Embed Size (px)
Citation preview

J. Moll. Stud. (1998), 64,447-460 © The Malacological Society of London 1998
COMPARATIVE SPERM MORPHOLOGY OF THEPULMONATE LIMPETS TRIMUSCULUS COSTATUS,
T. RETICULATUS (TRIMUSCULIDAE) AND BURNUPIASTENOCHORIAS AND ANCYLUS FLUVIATILIS
(ANCYLIDAE)
A.N. HODGSON1 and J.M. HEALY2
'Department of Zoology and Entomology, Rhodes University, Grahamstown 6140, South Africae-mail: [email protected]
'Department of Zoology, University of Queensland, Brisbane, Queensland 4072, Australiae-mail [email protected]
(Received 20 November 1997; accepted 23 January 1998)
ABSTRACT
Sperm infrastructure is described for the first time inrepresentatives of the pulmonate 'limpet' familiesTrimusculidae (Trunusculus costatus, T. reticulatus:marine) and Ancylidae (Bumupia stenochorias,Ancylus fluviatilis: freshwater). All show character-istic heterobranch sperm features (a spheroidalacrosomal vesicle supported by an acrosomalpedestal; a helically keeled nucleus and a complex,very elongate midpiece featuring paracrystalline andmatrix layers sheathing the axoneme, coarse fibersand one or more glycogen helices). Posterior to themidpiece, a glycogen piece (axoneme sheathed byglycogen granules) and annulus are also present in allspecies. Taxonomically useful differences in theshape and dimensions of the acrosome, nucleus andmidpiece occur between the species. Results supportthe decision of recent workers to transfer the Tri-musculidae from the Siphonarioidea to a separatesuperfamily Trimusculoidea (characteristic spermfeatures including: narrow acrosomal pedestal over-lapping with nuclear apex; heavily keeled nucleus;midpiece with strongly projecting secondary andglycogen helices). The relationship of the Trimuscu-loidea to other pulmonates, as indicated by spermultrastructure, remains uncertain largely becausecomparative data for several important groups areunavailable. Spermatozoa of the two ancylids mostclosely resemble those of other investigated planor-boideans and to a lesser extent, those of the Lym-naeoidea. However, differences between Bumupiastenochorias (unique(?) accessory structure on theacrosomal pedestal; glycogen wedges within thenuclear fossa; other features similar to planorbids)and Ancylus fluviatilis (all sperm features verysimilar to planorbids) suggests that these patelliformancylids are not closely related.
AU conapondcDce to. Professor A.N. Hodgson, DepL Zoology AEntomology, Rhodes University, Grahamstown 6140, South Africa
INTRODUCTION
In recent years the phylogeny and classificationof pulmonate gastropods have been the subjectof several re-appraisals (see Bieler, 1992; Nord-sieck, 1992; Tillier & Ponder, 1992; Tillier et ai,1996 for reviews). Largely the stimulation forthese studies has been provided by the avail-ability of new sources of information (ultra-structural characters and molecular data) and amore refined knowledge of general anatomicalcharacters including character state homology.Traditionally the Pulmonata have comprisedtwo principal orders, the Basommatophora andStylommatophora although several authorshave suggested that the Basommatophora mightnot be a monophyletic group (Morton, 1955;Hubendick, 1978; Healy, 1983). A third orderknown under various names (for example,Systellommatophora, Gymnomorpha, Soleo-lifera) is typically recognized for variousamphibious-terrestrial slug groups such as theOnchidoidea and Rathousoidea (see Tillier,1984 for discussion).
Within the Basommatophora sensu latoa number of taxa have patelliform shellsincluding the marine Siphonariidae and Tri-musculidae and the freshwater Latiidae, Lan-cidae, Ancylidae, Acroloxidae, Protancylidaeand Patelloplanorbidae (Hubendick, 1945,1978; Boss, 1982). The Siphonariidae andTrimusculidae have usually been placed withinthe Siphonarioidea (Hubendick, 1978; Boss,1982; also Patelliformia of Thiele, 1931). Tillier(1984) proposed removing both the Siphonari-idae and Trimusculidae from the Basom-matophora and including them with the
Dow
nloaded from https://academ
ic.oup.com/m
ollus/article/64/4/447/981297 by guest on 13 July 2022

448 A.N. HODGSON & J.M. HEALY
Amphibolidae in a redefined and expandedAmphiboloidea (in Order Archeopulmonata,with Ellobioidea, Onchidioidea and Rath-ouisioidea; Tillier reserving 'Basommatophora'for the limnic groups only). More recentlyHaszpmnar & Huber (1990) have removed theTrimusculidae from the Basommatophorasensu lato, and placed them together with theEllobiidae and Stylommatophora in a higher-rank taxon, the Eupulmonata (an Orderoriginally introduced by Morton, (1955), forthe Stylommatophora, but expanded andredefined by later authors). Nordsieck (1992)proposed a classification of the Pulmonata inwhich his Suborders Siphonarioidei andAmphiboloidei were included in the Super-order Basommatophora whereas the OrderTrimusculiformes was removed from theBasommatophora and placed in the Super-order Eupulmonata. All proposals howeveragreed on retaining the ancylid limpets withinthe Basommatophora, specifically within thePlanorboidea (Hubendick, 1978), but preciserelationships of the Ancylidae remain uncer-tain.
Numerous studies have now demonstratedthat sperm morphology, particularly whenexamined at the ultrastructural level, can pro-vide extremely useful insights into taxonomicand phylogenetic relationships of gastropods(Thompson, 1973; Healy & Willan, 1984,1991;Kohnert & Storch, 1984; Koike, 1985; Healy,1988, 1993, 19%; Hodgson & Bernard, 1988).Whereas there have been several studies onthe sperm ultrastructure of siphonariid limpets(Healy, 1983; Sumikawa & Funakoshi, 1984;Azevedo & Corral, 1985; Hodgson et al., 1991),and of a number of other basommatophoran(sensu lato) groups including the Ellobioidea(Ellobiidae—Healy, 1983), Amphiboloidea(Amphibolidae—Healy, 1983), Lymnaeoidea(Lymnaeidae—Ohsako, 1971; Maxwell, 1975,1980; Rigby, 1982; Healy, 1983), Physoidea(Physidae—Ackerson & Koehler, 1977; Brack -enbury & Appleton, 1991a) and Planorboidea(Planorbidae—Bogitsh, 1974a,b, 1975; Jong-Brink et al., 1977; Kitajima & Paraense, 1976;Ackerson & Koehler, 1977; Brakenbury &Appleton, 1991b; Appleton & Brackenbury,1997), there have been no descriptions of thesperm of the marine Trimusculidae or thefreshwater Ancylidae. Clearly if sperm morph-ology is to be of assistance in resolving the tax-onomic questions such as the position ofthe trimusculids, and precise relationships ofthe Ancylidae, descriptions of these unstudiedtaxa are required.
In this account we describe the structure ofthe sperm of two species of Trimusculusand two ancylid limpets, then compare theseresults with those published for other Basom-matophora sensu lato and the Eupulmonatasensu Nordsieck (1992). In addition we com-ment on the potential value of sperm morph-ology in assisting the systematics of thesegastropods.
MATERIALS AND METHODS
Specimens of Trimusculus costatus (Krauss, 1948)and T. reticulatus (Sowerby, 1835) were collectedfrom the intertidal regions of the eastern Cape coastof South Africa (near East London), and California(Long Marine Labs, Santa Cruz) respectively.Bumupia stenochorias (Melvill and Ponsonby, 1903)was collected from the Blaukranz River close toGrahamstown in the eastern Cape of South Africa.Specimens of Ancylus fluviatilis MUller were col-lected from freshwater ponds near Reading, southernEngland.
For transmission electron microscopy, smallportions of the gonad and hermaphrodite duct weredissected from animals and fixed in 2.5% gluta-raldehyde in sea water (Trimusculus), 2.5% gluta-raldehyde in 0.1 M phosphate buffer (pH 7.2)(Bumupia & Ancytlus). All fixation was for 2 hoursat room temperature. After primary fixation, tissueswere washed in either 0.1M sodium cacodylatebuffer (marine tissues) or 0.1 M phosphate buffer(Bumupia, Ancylus) and postfixed in 1% osmiumtetroxide in the same buffer media for 90 minutes,dehydrated in a graded ethanol series and eitherembedded in an Araldite/Taab resin mixture viapropylene oxide (Trimusculus, Bumupia), or inSpurrs resin (Ancylus). Thin sections (silver/goldinterface), cut either on a RMC7 or LKB 2128 ultra-microtome, were stained in aqueous uranyl acetate(20 minutes) and lead citrate (3 minutes) beforebeing viewed either on a JEOL 1210 or Hitachi 300transmission electron microscope at 80 kv.
RESULTS
All four species examined produce complex,uniflagellate introsperm which exhibit, inanterio-posterior sequence, a short acrosomeand nucleus (collectively, the 'head' region), anelongate midpiece and a terminal glycogenpiece. The descriptions that follow are ofmature spermatozoa from the seminal vesicleregion of the hermaphrodite duct, except in thecase of Ancylus fluviatilis, where only ovo-testicular spermatozoa were available forstudy.
Dow
nloaded from https://academ
ic.oup.com/m
ollus/article/64/4/447/981297 by guest on 13 July 2022

SPERM STRUCTURE OF PULMONATE LIMPETS 449
Trimusculus reticulalus (Trimusculidae, Figure 1)
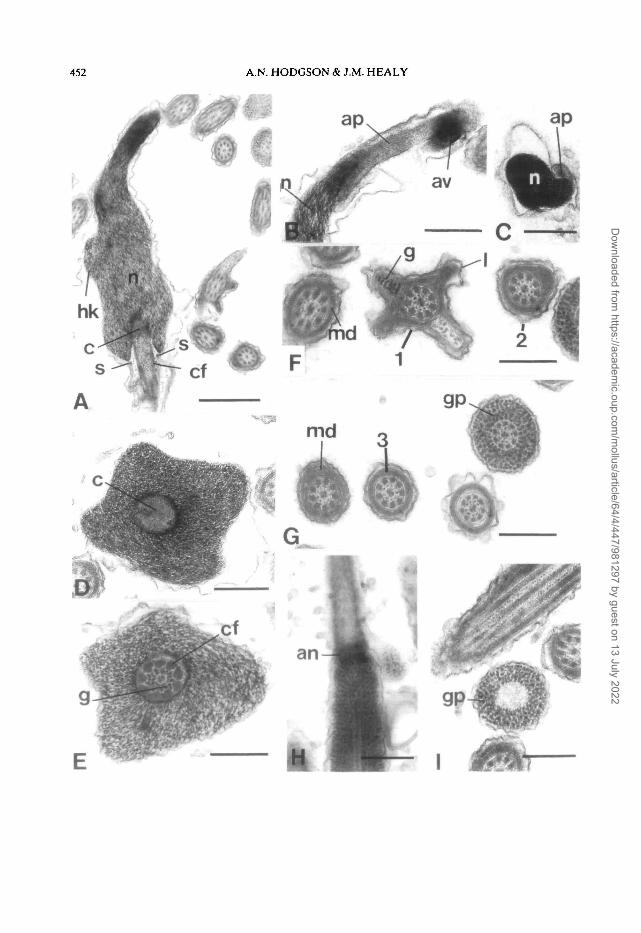
Sperm of T. reticulatus measure approximately 500u,m in total length. The head of the sperm is falciformwith a length of about 3.4 p.m and a basal diameter ofabout 1 u.m. The nucleus, which is uniformly elec-tron-dense, has a rounded, bilobed helical keel andtapers anteriorly. The anteriorly positioned acro-some consists of a cylindrical acTosomal pedestal(about 0.4 p.m long) at the apex of which is a smallvesicle about 0.4jj.m in diameter (Fig. 1B,C,D). Thebasal third of the acrosomal pedestal overlaps thenuclear apex (Fig. 1D,E). Posteriorly the nucleus hasa deep (about 1 n.m) V-shaped fossa which housesthe electron-dense centriolar complex from whichperiodically banded course fibers emerge (Fig. IF).Beneath the nucleus is a small sub-nuclear ring (Fig.
)The midpiece consists of an axoneme and coarse
fibre complex, two glycogen-filled helices, and anextensive, elongate mitochondrial derivative. Thenine periodically-striated coarse fibers are closelyassociated with the nine axonemal doublets (onecoarse fibre per doublet) and are thickest (150 nm)immediately posterior to the nucleus. Coarse fibrediameter decreases further posteriorly until in themiddle and distal regions of the midpiece the fibersare not observed. The mitochondrial derivative,which is composed of matrix and paracrystallinelayers, has a narrow lateral extension which graduallydecreases in size as it spirals posteriorly along themidpiece (Fig. 1G,H). The two glycogen-filledhelices, proximally have a wavelength of about1.6 jj.m (Fig. 1A) and decrease in size and numberalong the axoneme, eventually becoming reduced toone helix (Fig. 1H) before ultimately ending. Thedistal portion of the midpiece consists of theaxoneme and mitochondrial derivative only.
An annulus, in the form of a collar (Fig. II), marksthe junction between the midpiece and glycogenpiece. The anterior section of the glycogen piece isabout 0.7 jj.m in diameter, with a centrally placedaxoneme. Posteriorly the glycogen piece graduallydecreases in diameter and distally lacks the axoneme(Fig. U).
Trimusculus costatus (Trimusculidae, Figure 2)
The sperm of T. costatus is about 500 \im long. Thesperm head is about 4.6 jj.m long and contains anucleus (about 4.3 p.m long) in which the chromatinhas a fibrous appearance (Fig. 2A,D,E), the fibersbeing orientated along the long axis of the sperm.The nucleus has two or three broad helical keels (Fig.2A,D,E). Anteriorly the nucleus narrows and iscapped by an acrosomal complex. This complex con-sists of an acrosomal pedestal (about 0.6 u.m long)and small, spherical acrosomal vesicle (0.2 UJTI diam-eter) (Fig. 2B). The basal third of the pedestal over-laps the nuclear apex (Fig. 2B,C).
Posteriorly the nucleus has a deep fossa (about0.8 u,m) which houses the electron-dense centriolarcomplex and at the base of the nucleus a sub-nuclear
ring is present (Fig. 2A,D). The nine periodicallybanded coarse fibres (Fig. 2A,E) which have aninitial diameter of 100-150 nm, emerge from thecentriolar complex and surround the axoneme.Glycogen granules are present between the centralpair of microtubules of the axoneme and thedoublets/coarse fibres (Fig. 2E).
At the proximal end of the midpiece the mito-chondrial derivative surrounds two glycogen helices(helical wavelength anteriorly = 1.4 nm) which arediametrically opposed. In addition, the mitochon-drial derivative has two lateral arms (Fig. 2F) whichare also diametrically opposed. Posteriorly the glyco-gen helices are reduced to a single unit before endingand the mitochondrial arms are reduced in size (Fig.2F,G).
An annulus, similar in appearance to thatdescribed for T. reticulatus, marks the junction of themidpiece and glycogen piece which is about 0.8 u.m indiameter (Fig. 2H). The axoneme terminates beforethe end of the glycogen piece, which posteriorly has acentral canal filled with flocculent material (Fig. 21).
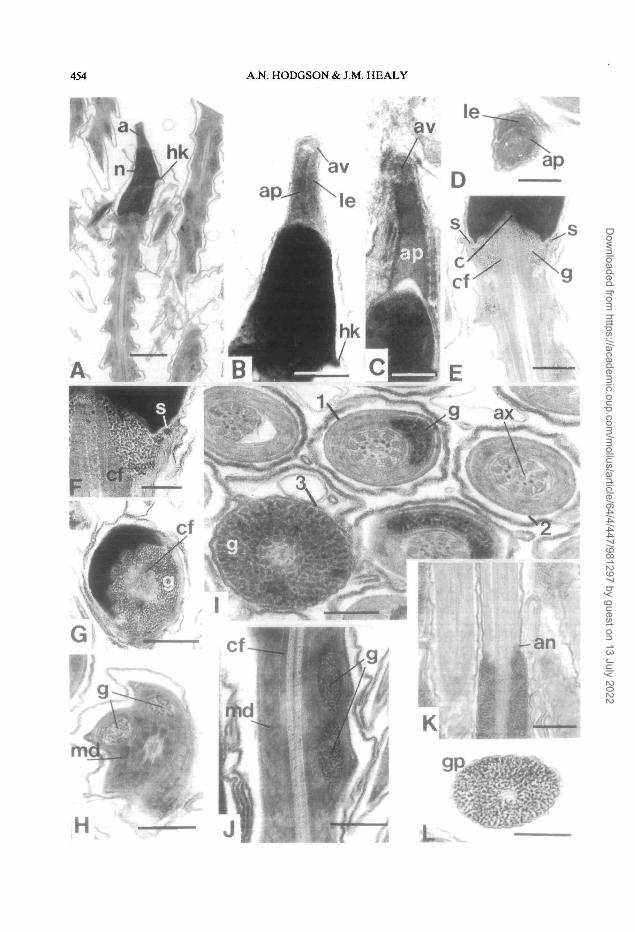
Bumupia stenochorias (Ancylidae, Figure 3)
The sperm from the seminal vesicle region of thehermaphrodite duct are about 465 u,m long. Thehead (about 3.0 u.m long) contains an electron-densenucleus (about 2.1 nm long X 1.0 jj.m basal diameter)which has a small lateral projection, the helical keel(Fig. 3A,B). Anteriorly the nucleus narrows and isrounded, whereas posteriorly there is a wide nuclearfossa which houses the centriolar complex (Fig.3E,F). At the base of the nucleus are small electron-dense structures which possibly form a continuoussub-nuclear ring (Fig. 3E,F). The acrosomal complex(about 0.8 u.m long X 0.4 u.m basal diameter) issituated anterior to the nucleus. It consists of a cylin-drical, hollow acrosomal pedestal (length 0.7 jim,basal diameter 0.3 p.m) flanked by a lateral element,and a small (0.1 jim diameter) spherical acrosomalvesicle (Fig. 3B,D) which is most clearly seen/observed in late spermatids (Fig. 3C).
The neck region of the midpiece consists of acentriolar complex implanted into a broad V-shapedfossa in the base of the nucleus. Nine very shallowdepressions within the nuclear fossa are filled by ninetriangular deposits of glycogen granules (Fig.3E,F,G). Anteriorly, the axoneme, which has a 9 + 2arrangement of microtubules, is surrounded by ninecoarse fibres which are periodically banded and havetheir origin in the centriolar complex (Fig. 3E,F,G).The fibres initially have a diameter of 150 nm but thisdecreases along the midpiece. They are absent fromthe mid- and posterior regions of the midpiece andthe glycogen piece. The coarse fibres are in turnsurrounded by the mitochondrial derivative (Fig.3H,J). At the proximal end of the midpiece the mito-chondrial derivative surrounds two glycogen helices(Fig. 3HJ). In B. stenochorias the helices each have aperiodicity of about 0.8 u,m. Posteriorly along themidpiece, the two glycogen helices are reduced(possibly by coalescence) to form a single helix which
Dow
nloaded from https://academ
ic.oup.com/m
ollus/article/64/4/447/981297 by guest on 13 July 2022

450 A.N. HODGSON & J.M. HEALY
Dow
nloaded from https://academ
ic.oup.com/m
ollus/article/64/4/447/981297 by guest on 13 July 2022

SPERM STRUCTURE OF PULMONATE LIMPETS 451
Figure 1. Longitudinal sections (L.S.) and transverse sections (T.S.) through hermaphrodite duct spermatozoaof Trimusculus reliculalus.A. L.S. of the head and midpiece showing glycogen helix (g) and lateral arm (1) of the mitochondrial derivative.B,C4X L.S. of the head showing falciform shape of the nucleus (n) and the apical acrosomal vesicle (av) andacrosomal pedestal (ap). E. T.S. of the anterior of the nucelus (n) and posterior section of the acrosomalpedestal (ap). F. L.S. of the posterior nucleus, neck region and anterior midpiece. G,H. T.S. through regions ofthe midpiece (anterior to posterior, 1-4 respectively) and glycogen piece (gp). I. L.S. of the annulus (an) at thejunction of the midpiece and glycogen piece. J. T.S. of the glycogen piece (gp) in which the axoneme is absentfrom the centre.Abbreviations: af, anterior nuclear fossa; ap, acrosomal pedestal; av, acrosomal vesicle; c, centriolar complex;cf, coarse fibres associated with axonemal doublets; g, glycogen; gp, glycogen piece; hk, helical keel of nucleus;1, lateral arm of mitochondrial derivative; md, mitochondrial derivative; n, nucleus; s, sub-nuclear ring.Scale bars: A = 1.0 u.m; B-D and F-J = 0.5 nm; E = 0.25 u,m
in turn gradually diminishes in extent and ultimatelyends, leaving the axoneme and coarse fibres sur-rounded only by the paracrystalline and matrix com-ponents of the mitochondrial derivative (Fig. 31).
Posterior to the midpiece the axoneme is sur-rounded by glycogen granules. At the junction of themidpiece and glycogen piece there is a distinct annu-lus (Fig. 3K). The anterior section of the glycogenpiece is about 1 |xm in diameter, the diameterdecreasing posteriorly. The posterior section of theglycogen piece lacks an axoneme and is filled withglycogen only (Fig. 3L).
Ancylusfluviatilis (Ancylidae, Figure 4)
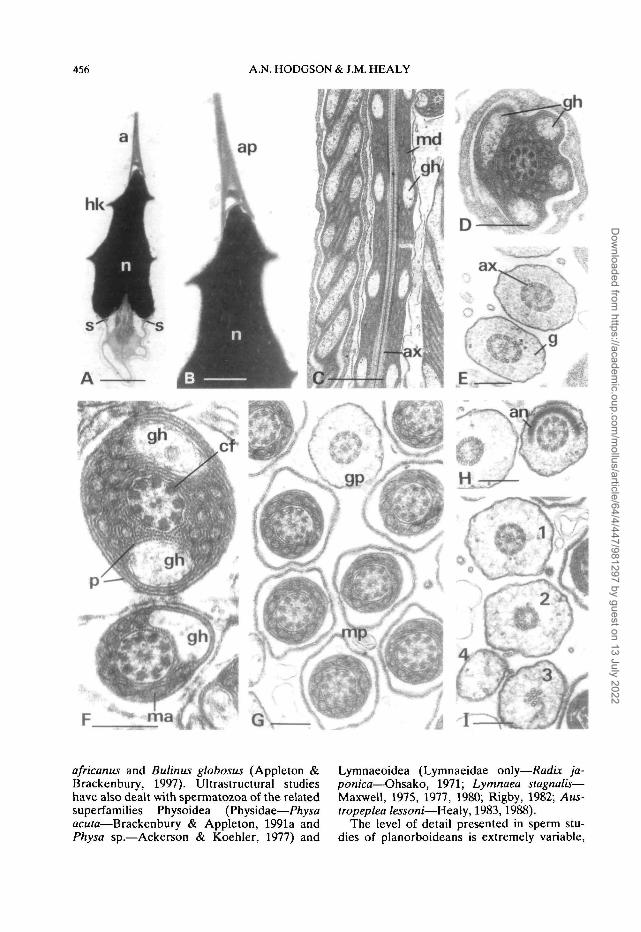
Spermatozoa from the ovotestis are about 330-350nm long. The head (about 3.0 p.m long) consists ofan elongate-conical, electron-dense nucleus (about2.1 n,m long X 1.0 \ym basal diameter) cappedapically by a finely tapered acrosomal complex (Fig.4A,B). One or two sharp-edged keels run helicallyalong the length of the nucleus (Fig. 4A). Thenuclear apex is sheathed by the base of the acrosomalpedestal. Basally the nucleus exhibits a broad,shallow, V-shaped fossa housing the centriolar com-plex and initial portion of the axoneme and coarsefibers (Fig. 4A). The acrosomal complex exhibits atall, conical, finely tapered, acrosomal pedestal(about 13-1.6 ji,m long X 0.3 (im basal diameter)seated on the nuclear apex (Fig. 4B). A central,slightly irregular cavity is present within the pedestal.Too few mature acrosomes were observed to estab-lish the presence and shape of the acrosomal vesicle.
The neck region of the midpiece consists of acentriolar complex implanted into the V-shaped pos-terior nuclear fossa. A sub-nuclear ring is present(Fig. 4A). Anteriorly, the axoneme, which has a 9+ 2 arrangement of microtubules, is surroundedby nine thick, coarse fibers emanating from thecentriolar complex (Fig. 4A). They are reduced indiameter in the middle and posterior regions of themidpiece. The coarse fibres are in turn surrounded bythe mitochondrial derivative which is composed ofhelically coiled matrix material enveloped in exten-sive layers of paracrystalline layers (also helicallycoiled) (Fig. 4C,D,F,G). At the proximal end of themidpiece the mitochondrial derivative surroundsfour glycogen helices (Fig. 4D), the walls of which
are formed from the paracrystalline layers. Thehelices each have a periodicity of about 13 u.m.Moving posteriorly along the midpiece, the glycogenhelices are reduced from four to one in number(Fig. 4F) until only the mitochondrial derivative andits associated membranes surround the axoneme andcoarse fibers (Fig. 4G).
Posterior to the midpiece the axoneme issurrounded by glycogen granules to form the glyco-gen piece (Fig. 4G-I). The axonemal doublets in thisregion are not accompanied by coarse fibers, indi-cating that the fibers terminate either at or imme-diately prior to the annulus. The annulus, at the junc-tion of the midpiece and glycogen piece, is evidentlywell developed but was only partly observed in asingle transverse section (Fig. 4H). The glycogenpiece has a diameter of approximately 0.5 |j.mthroughout most of its length. Terminally the 9 + 2axoneme undergoes the degenerative transition in itsstructure: 1) loss of central pair of microtubules (Fig.4H, left); 2) angling of doublets and marked reduc-tion of axonemal diameter (Fig. 41, 2); 3) loss andrandom organization of doublets (Fig. 41, 3); 4) totalloss of axonemal microtubules (posterior tip com-posed of glycogen surrounded by plasma membrane(Fig. 41,4).
DISCUSSION
Spermatozoa of Trimusculus reticulatus, T.costatus, Burnupia stenochorias and Ancylusfluviatilis exhibit morphological features whichare characteristic of the vast majority of hetero-branch gastropods. The more significant ofthese features include the following: 1) anacrosome composed of an acrosomal pedestaland membrane-bound acrosomal vesicle; 2) anucleus which typically exhibits one or morehelical keels and is posteriorly invaginated; 3)a midpiece composed of a complex mitochon-drial derivative (paracrystalline and matrixcomponents) which surrounds the axonemalcomplex and one or more glycogen-filledhelices and 4) and a 9 + 2 pattern axoneme, the
Dow
nloaded from https://academ
ic.oup.com/m
ollus/article/64/4/447/981297 by guest on 13 July 2022
452 A.N. HODGSON & J.M. HEALY
ap
Dow
nloaded from https://academ
ic.oup.com/m
ollus/article/64/4/447/981297 by guest on 13 July 2022

SPERM STRUCTURE OF PULMONATE LIMPETS 453
Figure 2. Longitudinal sections (L.S.) and transverse sections (T.S.) through hermaphrodite duct spermatozoaof Trimusculus costatus.A. L.S. through the head showing the fibrous nature of the chromatin of the nucleus (n), centriolar complex (c)and sub-nuclear ring (s). B. L.S. though the apical region of the nucleus (n), the acrosomal pedestal (ap) andacrosomal vesicle (av). C T.S. through the anterior nucleus (n) and base of the acrosomal pedestal (ap). D.T.S. through the centriolar complex (c) in the posterior fossa of the nucleus. E. T.S. through the base of thenucleus showing the coarse fibres (cf) surrounding the axoneme. F,G. T.S. through different regions of the mid-piece (anterior to posterior region 1-3 respectively), and glycogen piece (gp). H. L.S. through the annulus (an)at the junction of the midpiece and glycogen piece. L T.S. through the posterior glycogen piece.Abbreviations: c, centriolar complex; cf, coarse fibres associated with axonemal doublets; g. glycogen; gp,glycogen piece; hk, helical keel of nucleus; 1, lateral arm of mitochondrial derivative; md, mitochondrial deriva-tive; n, nucleus; s, sub-nuclear ring.Scale bars: A = 1.0 um; B-I = 0.5 u.m.
nine doublets of which are closely associatedwith nine periodically banded coarse fibres(Healy, 1983; see also Healy, 1993 for furtherdiscussion of major variants from the standardheterobranch sperm pattern). Significantly thespermatozoa of Trimusculus reticulatus, T.costatus, Burnupia stenochorias and Ancylusfluviatilis also show features which differentiateeach from other investigated pulmonates,thereby allowing some commentary to be givenin relation to the possible taxonomic implica-tions of our results.
The Siphonariidae (Siphonarioidea) andAmphibolidae (Amphiboloidea) are usuallyregarded as being closely related (e.g. seeThiele, 1931; Hubendick, 1945, 1978; Boss,1982; Tillier, 1984; Haszprunar & Huber, 1990;Nordsieck, 1992). Characteristic features ofSiphonaria spermatozoa include: 1) a usuallyfibrous nucleus; 2) a relatively long acrosomalpedestal (>0.8 u,m long) slightly draped overthe nuclear apex; 3) a midpiece featuring only asingle glycogen helix; 4) a sleeve-like annulus;5) a well developed glycogen piece (with anintact axoneme for most of its length) and 6)possibly the absence of a sub-nuclear ring(Healy, 1983; Sumikawa & Funakoshi, 1984;Azevedo & Corral, 1985; Hodgson et al., 1990).Results of Healy (1983, 1988) show thatspermatozoa of Salinator (Amphibolidae, twostudied species) differed substantially fromthose of Siphonaria (Siphonariidae, sevenstudied species) and Trimusculus (Trimusculi-dae, two studied species), specifically in thesubstructure of the acrosomal pedestal (hollow-cylindrical and associated with a helical bodywhich also coils around the nucleus), annulus(a simple ring and not sleeve-like) and theglycogen piece (totally lacking axoneme). Suchdifferences support the continued recognitionof a separate superfamily for the Amphiboli-dae (Amphiboloidea) but cast considerabledoubt over the validity of the higher taxon
Thalassophila (embracing marine Amphiboli-dae and Siphonariidae—see Nordsieck, 1992).Significantly, results of rRNA sequence studiesled Tillier et al., (1996) to conclude that theThalassophila were at best a paraphyletic andnot monophyletic grouping, and that theAmphibolidae may even belong outside thePulmonata. Further study of amphibolid sper-matozoa seems warranted.
The Trimusculidae have traditionally beenincluded within the Siphonarioidea, princi-pally because of the radially-ridged, patelliformshell, wide pallial cavity and amphibious-marine lifestyle (for example, see Hubendick,1945,1978; Boss, 1982; Siphonarioidea = StirpsPatelliformia of Thiele, 1931). However, inrecent years the anatomy of the Trimusculidaeand other groups of amphibious pulmonateshas been re-assessed, particularly in relationto details of the central nervous system. Usingthis information, Haszprunar & Huber (1990)removed the Trimusculidae and Ellobiidaefrom the Basommatophora sensu lato andincluded both, together with the Stylom-matophora, in a 'new' ordinal rank group, theEupulmonata (a pulmonate ordinal namesomewhat tentatively introduced by Morton,1955 for Stylommatophora). The ordersBasommatophora and Systellommatophorawere retained by Haszprunar and Huber andthe Basommatophora redefined to reflect theremoval of the Trimusculidae and Ellobiidae.Nordsieck (1992) not only accepted thesechanges but also incorporated the Systel-lommatophora (= Onchidioidea + Rathou-isioidea) within the 'superorder' Eupulmonata.The present study has shown that trimusculidspermatozoa differ from those of siphonariidsin a number of features, the most importantbeing: 1) presence of a smaller acrosomalpedestal which substantially overlaps with thenuclear apex; 2) pronounced keels on thenucleus and 3) presence of prominent
Dow
nloaded from https://academ
ic.oup.com/m
ollus/article/64/4/447/981297 by guest on 13 July 2022
454 A.N. HODGSON & J.M. HEALY
le
Dow
nloaded from https://academ
ic.oup.com/m
ollus/article/64/4/447/981297 by guest on 13 July 2022

SPERM STRUCTURE OF PULMONATE LIMPETS 455
Figure 3. Longitudinal sections (L.S.) and transverse sections (T.S.) through hermaphrodite duct spermatozoaof Bumupia stenochorias.A. L.S. of the head and anterior region of the mid-piece. B. L.S. of the nucleus and acrosome. C. L.S. of theacrosome of a late spermatid. D. T.S. section of the acrosomal pedestal (ap) and lateral element (le). E. L.S. ofthe neck region. F. Higher magnification of the posterior nucleus showing sub-nuclear ring (s) and periodicallybanded coarse fibres (cf).G. T.S. of the posterior nucleus and posterior fossae containing glycogen (g). H. T.S.midpiece showing two glycogen helices (g). I. T.S. of regions of the midpiece with a single glycogen helix (1),area with mitochondrial derivative only (2) and glycogen piece (3). J. L.S. midpiece showing two glycogenhelices (g). K. L.S. showing annulus (an) at the junction of the midpiece and glycogen piece. L. T.S. glycogenpiece with no axoneme.Abbreviations: a, acrosome; ap, acrosomal pedestal; av, acrosomal vesicle; c, centriolar complex; cf, coarsefibres associated with axonemal doublets; g, glycogen compartment; gp, glycogen piece; hk, helical keel ofnucleus; le, lateral element of acrosomal pedestal; md, mitochondrial derivative; n, nucleus; s, sub-nuclear ring.Scale bars: A = 1.0 (j.m; B, E-G, I-K = 0.5 p.m; C,D,H = 0.25 jim
secondary helices (lateral projections) and twoglycogen helices in the midpiece. Such differ-ences make it impossible to identify any spermsynapomorphy for the Siphonarioidea sensulato, but do support the view (e.g. Nordsieck,1992) that trimusculids are worthy of their ownhigher-level status within the Pulmonata(Trimusculoidea, Trimusculiformes). Presentlythere are no overwhelming spermatologicalreasons to closely associate the Trimusculidaewith the Ellobioidea (see Healy, 1983 formicrographs) or with any other group ofpulmonates, although it should be added thatthis situation may be remedied once detailedcomparative sperm data become available forunstudied genera, families and superfamilies.Unfortunately no published DNA and RNAstudies have yet included trimusculids amongtaxa being investigated.
Although spermatozoa of T. reticulatus andT. costatus differ from each other in the sub-structure of the nucleus (fibrous in T. costatus,homogeneous in T. reticulatus) as well as thenumber of lateral projections of the mito-chondrial derivative (one in T. reticulatus, twoin T. costatus), the general form of the acro-somal complex (overlapping nuclear apex),nucleus (short, with strong keels), midpieceand glycogen piece are consistent and presum-ably are characteristic of the Trimusculidae.The fact that two species of the same genus(Trimusculus) have different forms of chro-matin would suggest that this morphologicalfeature is probably of little systematic value.Chromatin with a fibrous appearance has beenreported for six species of Siphonaria(Sumikawa & Funakoshi, 1984; Azevedo &Corral, 1985; Hodgson et al., 1991) and theonchidoideans Onchidium damellii and Onchi-della celtica (Healy, 1986; Selmi et al., 1988),whereas in most other pulmonates the chro-matin has a homogeneous appearance (e.g.
Takaichi & Sawada, 1973; Takaichi, 1975;Atkinson, 1982; Maxwell, 1983; Healy, 1983;Healy & Jamieson, 1989; Brackenbury &Appleton, 1991a,b; Cuezzo, 1995a,b). Variousfactors may be at work in determining whethersperm nuclei of heterobranch gastropods havefibrous or homogeneous substructure, includ-ing fixation and osmotic effects, pH, structuralchanges to membranes, nuclear protein tran-sitions or other physiological alterations (seeSelmi et al., 1988; Healy & Willan, 1991, fordiscussion and further literature). However,almost nothing is known of the extracellularbiochemical influences on heterobranch sper-matozoa either within the hermaphrodite ductor the receptaculum prior to fertilization.
All recent proposed classifications of thePulmonata place the ancylids within the limnicBasommatophora (the 'Branchiopulmonata' ofMorton, 1955), usually within the Planorboidea(Harry, 1964; Hubendick, 1978; Boss, 1982;Tillier, 1984; Nordsieck, 1992). Hubendick(1978) fused the Planorbidae and Ancylidaeinto a single family the Ancyloplanorbidaebut in most classifications the Ancylidae areretained as a separate family for the bulk ofthe patelliform planorboideans (Thiele, 1931;Boettger, 1954; Boss, 1982). Although anumber of important limnic families have yetto be investigated in relation to sperm ultra-structure (e.g. Chilinidae, Latiidae), enough isknown to make comparison with the resultspresented herein for the two patelliformancylids, Bumupia stenochorias and Ancylusfluviatilis.
Five species of Planorboidea (Planorbidae)have previously been examined for sperm/spermatid ultrastructure—Planorbarius corneus(Starke & Nolte, 1970), Biomphalaria glabrata(Bogitsh, 1974a,b, 1975; Kitajima & Paraense,1976; Jong-Brink el al., 1977), Bulinus tropicus(Brackenbury & Appleton, 1991b), Bulinus
Dow
nloaded from https://academ
ic.oup.com/m
ollus/article/64/4/447/981297 by guest on 13 July 2022
456 A.N. HODGSON & J.M. HEALY
africanus and Bulinus globosus (Appleton &Brackenbury, 1997). Ultrastructural studieshave also dealt with spermatozoa of the relatedsuperfamilies Physoidea (Physidae—Physaacuia—Brackenbury & Appleton, 1991a andPhysa sp.—Ackerson & Koehler, 1977) and
Lymnaeoidea (Lymnaeidae only—Radix ja-ponica—Ohsako, 1971; Lymnaea siagnalis—Maxwell, 1975, 1977, 1980; Rigby, 1982; Aus-iropeplea /essom—Healy, 1983,1988).
The level of detail presented in sperm stu-dies of planorboideans is extremely variable,
Dow
nloaded from https://academ
ic.oup.com/m
ollus/article/64/4/447/981297 by guest on 13 July 2022

SPERM STRUCTURE OF PULMONATE LIMPETS 457
Figure 4. Longitudinal sections (L. S.) and transverse sections (T.S.) through ovotesticular spermatozoa ofAncylus flu viatilis.A. L.S. of the head. B. L.S. detail of previous figure showing internal structure of acrosomal pedestal (ap).C L.S. and oblique L.S. of middle region of midpiece showing two spiraling glycogen helices (sperm slightlyimmature). D. T.S. through midpiece in immediate post-nuclear region, showing four glycogen helices (spermslightly immature, still associated with cytoplasmic sheath). E. T.S. glycogen piece. F. T.S. middle and posteriorregions of midpiece showing details of internal substructure of the mitochondrial derivative andaxonemal/coarse fibre complex (cf). G. T.S. of posterior region of midpiece (after termination of glycogenhelix) and glycogen piece (gp). H. T.S. (oblique) portion of annulus (an). I. Sequence of structural changesoccurring in terminal region of glycogen piece (1, axoneme intact; 2, loss of central axonemal singlets, doubletsbeginning to degenerate; 3, 'axoneme' reduced to four disorganized doublets; 4, terminal portion without anymicrotubular elements).Abbreviations: a, acrosome; an, annulus; ap, acrosomal pedestal; ax, axoneme; cf, coarse fibres associated withaxonemal doublets; g, glycogen granules; gh, glycogen helix; gp, glycogen piece; hk, helical keel of nucleus;ma, matrix component of mitochondrial derivative; md, mitochondrial derivative; mp, midpiece; n, nucleus;p, paracrystalline component of mitochondrial derivative; s, sub-nuclear ring.Scale bars: A, C = 1.0 u,m; B,D = 0.5 (i.m; E-I = 0.25 u.m.
and unfortunately acrosomal features havebeen treated rather cursorily in all the availableliterature, thereby limiting the scope of thepresent discussion. Our results for Ancylusfluviatilis (Ancylidae), although incompleteand based on ovotesticular sperm, show severalsimilarities to other planorboideans, especiallyplanorbids. Acrosomal pedestal and nuclearmorphology are very similar to Biomphalariaglabrata (see Kitajima & Paraense, 1976); mid-piece morphology is similar to all investigatedplanorbids, but closest to Bulinus tropicuswhich, like Ancylus fluviatilis also shows amaximum of four glycogen helices (Bracken-bury & Appleton, 1991b; a condition elsewhererecorded within the Heterobranchia only in the'cephalaspid' opisthobranch Acteon tornatilis—see Thompson, 1973). Spermatozoa of Bur-nupia stenochorias (currently placed in theAncylidae) exhibit the same angulate second-ary helices on the mitochondrial derivativeseen in Biomphalaria glabrata (Planorbidae)(compare Figure 3H of this study with Figure5h of Kitajima & Paraense, 1976). However,there are also features which distinguish Bur-nupia stenochorias from other planorboideansincluding Ancylus fluviatilis (like B. stenocho-rias, an ancylid) notably the morphology of theacrosomal pedestal (columnar pedestal withlateral accessory structure which overlaps withnuclear apex), and the presence of glycogengranules in nine shallow recesses of the nuclearfossa. The close resemblance of ancylidspermatozoa to those of various planorbidscould be interpreted as further support forHubendick's (1978) amalgamation of theAncylidae and Planorbidae into a single family,the Ancyloplanorbidae, or suggest a commonorigin for ancylids and planorbids (Burch,
1962, however points out chromosomal num-ber differences between the two groups).
Spermatozoa of the patelliform freshwaterpulmonate Acroloxus lacustris show numerousdifferences from the ancylid limpets, includinga highly elongate nucleus, an elongate acro-somal pedestal and typically a single glycogenhelix in the midpiece, suggesting possibleconnections with the Glacidorboidea and/orAmphiboloidea (Healy in preparation). Burch(1962) using light microscopy, cited theextreme elongation of the sperm 'head' ( =acrosome + nucleus) of A. lacustris and chro-mosomal differences in this species as evidencefor establishing a family Acroloxidae (andremoving the genus Acroloxus from the Ancyl-idae)—a decision generally accepted in mostclassifications (usually the group is alsoaccorded superfamily status—see Boss, 1982).
From the spermatozoal perspective the Plan-orboidea appear to be closest to the Physoideaand Lymnaeoidea among the Basommato-phora sensu lato (very small, almost obsoleteacrosomal vesicle (always present in Physidae);very short, strongly-keeled nucleus, presenceof a sub-nuclear ring; mitochondrial derivativewith multiple glycogen helices and secondaryhelices; well developed glycogen piece withintact axoneme). Lymnaeid, physid and planor-bid spermatozoa are however each character-ized by a series of distinguishing features tojudge from the available data. For lymnaeidsthese include: 1) a bilaterally symmetrical,broadly lobulate, acrosomal pedestal (withinternal cylinder axially); 2) nucleus with mul-tiple, square-profile keels; 3) a very shallowcentriolar fossa; 4) a midpiece featuring 6-7,square-profile, secondary keels (anteriorly)and 5) a consistently sleeve-like annulus
Dow
nloaded from https://academ
ic.oup.com/m
ollus/article/64/4/447/981297 by guest on 13 July 2022

458 A.N. HODGSON & J.M. HEALY
(Ohsako, 1971; Rigby, 1982; Healy, 1983).Spermatozoa of Physa acuta (Physidae) show anumber of differences from those of planorbidsand lymnaeids, principally in relation to thenucleus (with perinuclear material) and themidpiece (triangular in transverse section an-teriorly, with three glycogen helices, decliningto two and one helices posteriorly) (Bracken-bury & Appleton, 1991a). UnfortunatelyBrackenbury & Appleton do not providedetailed micrographs of the acrosomal com-plex, the structure of which could have shedlight onto relationships of physids to otherlimnic basommatophorans. The only otherstudy of physid spermatozoa is a brief note byAckerson & Koehler (1977) detailing the formof the annulus and midpiece/glycogen piecejunction (and associated membranes). Wewould strongly recommend that further, ideallycomparative, ultrastmctural work on physidspermatozoa be carried out to test relation-ships of this family.
ACKNOWLEDGEMENTS
It is a pleasure to thank: Mary Bursey (East LondonMuseum, South Africa) for locating the populationsof Trimusculus costatus. Professor John and Dr VickiPearse for their warm hospitality to ANH whilst inSanta Cruz, Mrs Lil Haigh and Mr Heather Davies-Coleman for providing specimens of Bumupia steno-chorias and Dr. John Taylor and Ms Emily Glover(Natural History Museum, London) for providingglutaraldehyde-fixed specimens of AncylusfluviatUis.Mrs Val Hodgson (Dept. of Zoology, Rhodes Uni-versity) and Mrs Lina Daddow (Dept. of Zoology,University of Queensland) are thanked for assistancewith sectioning of the material. This work was sup-ported by a grant from the Foundation for ResearchDevelopment (to ANH) and by a Senior ResearchFellowship from the Australian Research Council,the Keith Sutherland Award of the AustralianMuseum and a University of Queensland EnablingGrant (to JMH).
REFERENCES
ACKERSON, A. & KOEHLER, L. 1977. Vesiculationand loss of the plasma membrane along the mid-piece of Physa sp. spermatozoon. 35th AnnualProceedings of the Electron Microscopical Societyof America (Boston): 618-619.
APPLETON, C.C. & BRACKENBURY, T.D. 1997. Acomparative study of the mature spermatozoa ofBulinus africanus (Krauss, 1848) and B. globosus(Morelet, 1866) (Gastropoda: Planorbidae). Jour-nal of Molluscan Studies, 63: 401-406.
ATKINSON, J.W. 1982. An ultrastmctural analysis ofthe mature spermatozoon of Anguispira altemata(Say) (Pulmonata, Stylommatophora). Journal ofMorphology, 173: 249-257.
AZEVEDO, C. & CORRAL, L. 1985. The fine structureof the spermatozoa of Siphonaria algestrae (Gas-tropoda, Pulmonata). Journal of Morphology, 186:107-117.
BIELER, R. 1992. Gastropod phylogeny and systemat-ics. Annual Review of Ecology and Systematics, 23:311-338.
BOETTGER, C.R. 1954. Die Systematik der euthy-neuren Schnecken. Deutsche Zoologischer Gesell-schaft Verhandlungen (Zoologischer Anzeiger,Supplement), 18: 253-280.
BOGITSH, BJ. 1974a. Cytochemical observations onthe midpiece of the spermatozoa of the gastropodBiomphalaria glabrata. Transactions of the Ameri-can Microscopical Society, 93: 211-219.
BOGITSH, BJ. 1974b. Acid phosphatase activity inspermatids of the snail Biomphalaria glabrata.Transactions of the American MicroscopicalSociety, 93: 363-373.
BOGITSH, BJ. 1975. The cytochemical localization ofsuccinic dehydrogenase in the sperm mitochondriaof the gastropod Biomphalaria glabrata. A com-parison of two methods. Journal of Histochemistryand Cytochemistry, 23: 75-79.
Boss, K.J. 1982. Mollusca. In: Synopsis and Classifi-cation of Living Organisms. (S.P. Parker, ed. inchief), 2: 945-1166. McGraw-Hill, New York.
BRACKENBURY, T.D. & APPLETON, C.C. 1991a. Mor-phology of the mature spermatozoon of Physaacuta (Draparnaud, 1801) (Gastropoda: Physidae).Journal of Molluscan Studies, 57: 211-218.
BRACKENBURY, T.D. AND APPLETON, C.C. 1991b.Morphology of the mature spermatozoon ofBulinus tropicus (Krauss, 1848) (Gastropoda:Planorbidae). Malacologia, 33: 273-280.
BURCH, J.B. 1962. Cytotaxonomic studies of fresh-water limpets (Gastropoda: Basommatophora). I.The European lake limpet, Acroloxus lacustris.Malacologia, 1: 55-72.
CUEZZO, M.G. 1995a. Spermatogenesis and spermstructure in the Neotropical pulmonate snailScutalus tupacii (d'Orbigny). Veliger, 38: 212-222.
CUEZZO, M.G. 1995b. Ultrastructure of the maturespermatozoa of the land snail Epiphragmophoratucumanensis (Doering, 1874) (Gastropoda:Helicoidea). Journal of Molluscan Studies, 61:1-7.
HARRY, H.W. 1964. The anatomy of Chilina fluctu-osa Gray reexamined, with prolegomena on thephylogeny of the higher limnic Basommatophora(Gastropoda: Pulmonata). Malacologia, 1: 355-385.
HASZPRUNAR, G. & HUBER, G. 1990. On the centralnervous system of Smeagolidae and Rhodopidae,two families questionably allied with the Gymno-morpha (Gastropoda: Euthyneura). Journal ofZoology, London, 220:185-199.
HEALY, J.M. 1983. An ultrastmctural study ofbasommatophoran spermatozoa (Mollusca, Gas-tropoda). Zoologica Scripta, 12: 57-66.
Dow
nloaded from https://academ
ic.oup.com/m
ollus/article/64/4/447/981297 by guest on 13 July 2022

SPERM STRUCTURE OF PULMONATE LIMPETS 459
HEALY, J.M. 1986. Electron microscopic observa-tions on the spermatozoa of a marine pulmonateslug Onchidium damellii (Gastropoda, Onchidi-acea). Journal of Submicroscopic Cytology, 18:587-594.
HEALY, J.M. 1988. Sperm morphology and itssystematic importance in the Gastropoda. Malaco-logical Review. Supplement, 4: 251-266.
HEALY, J.M. 1993. Comparative sperm ultrastructureand spermiogenesis in basal heterobranch gastro-pods (Valvatoidea, Architectonicoidea, Rissoel-loidea, Omalogyroidea and Pyramidelloidea)(Mollusca). Zoologica Scripta, 22: 263-276.
HEALY, J.M. 1996. Molluscan sperm ultrastructure:correlation with taxonomic units within the Gastro-poda, Cephalopoda and Bivalvia. In: Origin andRadiation of the Mollusca. (J. Taylor, ed.), 99-113.Oxford University Press, Oxford.
HEALY, J.M. & JAMIESON, B.G.M. 1989. An ultra-structural study of spermatozoa of Helix aspersaand Helix pomatia (Gastropoda, Pulmonata).Journal of Molluscan Studies, 55: 389-404.
HEALY, J.M. & WILLAN, R.C. 1984. Ultrastructureand phylogenetic significance of notaspideanspermatozoa (Mollusca, Gastropoda, Opistho-branchia). Zoologica Scripta, 13:107-120.
HEALY, J.M. & WILLAN, R.C. 1991. Nudibranchspermatozoa: comparative ultrastructure andsystematic importance. Veliger, 34: 134-165.
HODGSON, A.N. & BERNARD, R.T.F. 1988. A com-parison of the structure of the spermatozoa andspermatogenesis of 16 species of patellid limpet(Mollusca: Gastropoda: Archaeogastropoda).Journal of Morphology, 195: 205-223.
HODGSON, A.N., BERNARD, R.T.F. & LINDLEY, D.S.1991. Comparative spermatology of four sympatricspecies of Siphonaria (Pulmonata: Basom-matophora). Journal of Molluscan Studies, 57:309-322.
HUBENDICK, B. 1945. Phylogenie und Tiergeographieder Siphonariidae. Zur Kentniss der Phylogenie inder Ordnung Basommatophora und der Urspringsder Pulmonatengruppe. Zoologiska BidrUgen franUppsala, 24:1-216.
HUBENDICK, B. 1978. Systematics and comparativemorphology of the Basommatophora. In: Pul-monates (V. Fretter & J.F. Peake, eds), 2A: 1-47.Academic Press, London.
JONG-BRINK, DE, M., BOER, H.H., HOMMES, T.G. &KODDE, A. 1977. Spermatogenesis and the role ofSertoli cells in the freshwater snail Biomphalariaglabrata. Cell and Tissue Research, 181: 37-58.
KITAJIMA, E.W. & PARAENSE, W.L. 1976. The ultra-structure of the mature sperm of the freshwatersnail Biomphalaria glabrata (Mollusca: Gastro-poda). Transactions of the American MicroscopicalSociety, 95: 1-10.
KOHNERT, R. & STORCH, V. 1984. Vergleichend-ultra-strukturelle Untersuchungen zur Morphologieeupyrener Spermien der Monotocardia (Proso-branchia). Zoologischer Jahrbucher, 111: 51-93.
KOIKE, K. 1985. Comparative ultrastructural studieson the spermatozoa of the Prosobranchia (Mol-
lusca: Gastropoda). Science Report of the Facultyof Education, Gunma University, 34: 33-153.
MAXWELL, W.L. 1975. Scanning electron microscopestudies of pulmonate spermatozoa, Veliger, 18:31-33.
MAXWELL, W.L. 1977. Freeze-etching studies ofpulmonate spermatozoa. Veliger, 20: 71-74.
MAXWELL, W.L. 1980. Distribution of glycogendeposits in two euthyneuran sperm tails, Inter-national Journal of Invertebrate Reproduction, 2:245-249.
MAXWELL, W.L. 1983. Mollusca. In: Reproductivebiology of invertebrates II: Spermatogenesis andsperm function (K.G. Adiyodi & R.G. Adiyodi,eds), 275-319. John Wiley and Sons, Chichester.
MORTON, J.E. 1955. The evolution of the Ellobiidaewith a discussion on the origin of the Pulmonata.Proceedings of the Zoological Society of London,125: 127-168.
NORDSIECK, H. (1990) 1992. Phylogeny and systemof the Pulmonata (Gastropoda). Archiv fiirMolluskenkunde, 121: 31-52.
OHSAKO, N. 1971. Electron microscopic studies onthe sperm of fresh-water snail, Radix japonica Jay.I. Structural characterization. Reports of theFaculty of Science, Kagoshima University (EarthScience and Biology), 4: 63-70.
RIGBY, J.E. 1982. The fine structure of differentiatingspermatozoa and Sertoli cells in the gonad of thepond snail Lymnaea stagnalis. Journal of Mollus-can Studies, 48: 111-123.
SELMI, M.G., Giusn, F. & MANGANELLI, G. 1988.The fine structure of the mature spermatozoon ofOnchidella celtica (Cuvier) (Mollusca: Gastro-poda) and its phylogenetic implications. Journal ofUltrastructure and Molecular Structure Research,99:107-123.
STARKE, JJ. AND NOLTE, A. 1970. Tubulikorper imZytoplasma de spermatiden von Planorbariuscomeus L. (Basommatophora). Zeitschrift fiir Zell-forschung und Mikroskopische Anatomie, 105:210-224.
SUMIKAWA, S. & FUNAKOSHI, C. 1984. The fine struc-ture of mature spermatozoa in two species of theSiphonariidae (Pulmonata, Basommatophora).Venus, 43:143-155.
TAKAICHI, S. 1975. The fine structure of spermatozoaof Euhadra hickonis. Zoological Magazine, Tokyo,84: 48-57.
TAKAICHI, S. & SAWADA, N. 1973. An electronmicroscope study on sperm formation in Euhadrahickonis. Memoirs of the Ehime University, seriesB, 7:17-58.
THIELE, J. 1931. Handbuch der systematischenWeichtterkunde, P. Fischer, Jena.
THOMPSON, T.E. 1973. Euthyneuran and othermolluscan spermatozoa. Malacologia, 14: 167-206(plus addendum, 443-444).
TILLIER, S. 1984. Relationships of gymnomorphgastropods (Mollusca, Gastropoda). ZoologicalJournal of the Linnean Society, 82: 345-362.
TILLIER, S. AND PONDER, W.F. 1992. New species ofSmeagol from Australia and New Zealand, with a
Dow
nloaded from https://academ
ic.oup.com/m
ollus/article/64/4/447/981297 by guest on 13 July 2022

460 A.N. HODGSON & J.M. HEALY
discussion of the affinities of the genus (Gastro- pods from rRNA sequences, and tempo and age ofpoda: Pulmonata). Journal of Molluscan Studies, the stylomatophoran radiation. In: Origin and58: 135-155. Evolutionary Radiation of the Mollusca (J. Taylor,
TILLIER, S., MASSELOT, M. & TILLIER, A. 19%. ed.), 267-284. Oxford University Press, Oxford.Phylogenetic relationships of the pulmonate gastro-
Dow
nloaded from https://academ
ic.oup.com/m
ollus/article/64/4/447/981297 by guest on 13 July 2022





























