Embed Size (px)
Citation preview

THE ANATOMICAL RECORD 292:1213–1228 (2009)
Comparative Analysis of Paw PadStructure in the Clouded Leopard
(Neofelis nebulosa) and Domestic Cat(Felis catus)
CHRIS HUBBARD,* VIRGINIA NAPLES, ERIN ROSS, AND BURCU CARLONDepartment of Biological Sciences, Northern Illinois University, Dekalb, Illinois
ABSTRACTThe Clouded leopard (Neofelis nebulosa) is a medium-sized highly ar-
boreal cat. This study compares the structure of the digital, metacarpaland metatarsal pads of the manus and pes in N. nebulosa to that of thedomestic cat (Felis catus). Covered by a stratified squamous cornified epi-thelium, the pads have a supple deposit of subepidermal fat that is parti-tioned by collagen fibers and extensively anchored to the muscle tendonsheaths. In both animals, a pes metatarsal pad suspensory ligament origi-nates from the Mm. flexores digitorum profundi tendon and forms 3–4small branches that project through the dermal fat layer and attach tothe pad epidermis. In the cat manus, four tendons of equal size extendfrom the M. flexor digitorum superficialis (FDS) to form the manica flexo-ria in digits 2–4 from which extends a metacarpal pad suspensory liga-ment (MPSL) on digits 2 and 5 that extends into the tela subcutanea andepidermis. On digits 3 and 4 MPSL extends directly from the FDS tendonitself. In contrast, manus FDS tendons 1 and 5 in N. nebulosa were thinand either project directly to the tela subcutanea (tendon 1) or connectwith the manica flexoria forming a metacarpal pad suspensory ligament(tendon 5). Tendons 2–4 connect with the manica flexoria from whichMPSL project into the tela subcutanea and epidermis. In both species, thesuspensory ligaments may serve to contract the pad to conform to theunder lying substrate, thus enhancing the animal’s ability to grip brancheswhile climbing. Anat Rec, 292:1213–1228, 2009. VVC 2009 Wiley-Liss, Inc.
Keywords: metacarpal pad; metatarsal pad; clouded leopard;Neofelis nebulosa
The Clouded leopard (Neofelis,nebulosa, N. nebulosa)is the smallest ‘‘big cat,’’ that has the body size andshape of a small cat but with pantherine cranial anddental structure. These cats have the longest caninetooth-to-body size ratio among extant felids (Nowell andJackson, 1996; Sunquist and Sunquist, 2002), and can-ines with sharp-edged posterior keels, similar to extinctsaber-tooth cats (Christiansen, 2006). Canine lengthaffects other cranial features, including muzzle elonga-tion, a lengthened diastema between the canine andreduced first premolar, and a greatly elongated skull(Guggisberg, 1975). These cats also have short legs, witheven shorter forelegs, robust bodies and greatly enlargedforefeet that can assume a spoon shape for more effec-tive gripping of supporting branches. N. nebulosa weigh
from 16 to 23 kg and are about 60–106-cm long (headand body). The extremely long tail, used for balancingwhile climbing, adds 55–91 cm. to their total length(Wozencraft, 2005). N. nebulosa are solitary except forfemales with young (Yamada and Durant, 1989).
*Correspondence to: Dr. Chris Hubbard, Department of Bio-logical Sciences, Northern Illinois University, Castle Dr. Dekalb,IL 60115 E-mail: [email protected]
Received 7 September 2007; Accepted 18 April 2009
DOI 10.1002/ar.20930Published online 15 June 2009 in Wiley InterScience (www.interscience.wiley.com).
VVC 2009 WILEY-LISS, INC.
The common name, Clouded leopard, derives from coatmarkings; large, irregular cloud-like pale spots outlinedin black or brown surrounded by a pale ring on pale yel-low to light brown background (Gunderson, 1976; Nowelland Jackson, 1996; Sunquist and Sunquist, 2002).Recent coat color, pattern variation and genetic analysesdivide N. nebulosa into two species (Grassman et al.,2005; Buckley-Beason et al., 2006; Kitchener et al., 2006).N. nebulosa occurs widely across mainland Asia (Loxton,1973), while the new N. diardi is restricted to Indonesia(Kitchener et al., 2006). Both species occur in mangroveswamps in Borneo (Davis, 1962; Davies and Payne,1982), grass and scrublands (Santiapillai and Ashby,1988; Johns, 1989; Dinerstein and Mehta, 1989; Rabino-witz and Walker, 1991) subtropical forests, primaryclosed evergreen tropical forests, (Rabinowitz, 1988; Aus-tin, 2002), tropical rainforests (Johns, 1989) and open,dry partially forested areas. N. nebulosa adapt to differ-ent habitats, and is not restricted by human perturba-tion, because they can occupy semidisturbed habitats(Rabinowitz et al., 1987) of up to 3,000 square meters(Jerdon, 1874; Biswas et al., 1985). Habitat destructionis a major contributor to their highly endangered status[(CITES I); Groombridge, 1994; Baillie and Groombridge,1996; Nowell and Jackson, 1996] as is poaching for Asianmedicines and their beautiful coats (Low, 1991).
N. nebulosa are rare, shy, mainly nocturnal, and arbo-real, hunting and resting in trees (Grassman, 2001).They are generalist, opportunistic predators, takingbirds, arboreal monkeys and large prey, including pigs,cattle, young buffalo, goats, deer, and even porcupines(Nowak, 1999). These cats differ from typical felids inbeing able to run down trees headfirst, and to movealong horizontal supports with the body suspendedbeneath the branches (Layhausen, 1979; Choudhury,1996; Conforti, 1996; Nowell and Jackson, 1996; Ghose,2002; Grassman and Tewes, 2002). These unusual,highly acrobatic abilities have been also reported in themuch smaller margay and marbled cats, but the largerN. nebulosa displays greater adaptations for extreme ar-boreal abilities (Leopold, 1959; Grzimek, 1975; Lekaguland McNeely, 1977; Nowell and Jackson, 1996). Themargay (Leopardus weidii) is from Central and SouthAmerica, and the marbled cat, (Pardofelis marmorata)shares the Asian range of N. nebulosa (Pocock, 1932;Sunquist and Sunquist, 2002). The margay and perhapsthe marbled cat, have increased intratarsal flexibility toallow movements that are impossible in other felids(Grzimek, 1975), but there are no studies of more super-ficial structures that also correlate with tarsal move-ments (Layhausen, 1979; Choudhury, 1996; Ghose,2002). Information on gross and histological anatomyand function of paw pads is sparse, and unsurprisingly,nothing has been published on these structures inN. nebulosa (Cutts and Krause, 1983; Alexander et al.,1986; Benz et al., 2005.).
MATERIALS AND METHODS
Two N. nebulosa were obtained through the Smithso-nian Institution Osteopreparatory laboratory and arehoused in the research collection of the biological sciencesdepartment of Northern Illinois University (NIU 2105-2106). Both animals died of natural causes. Following ne-cropsy, the animals were transported to Northern Illinois
University for further dissection and analysis of therecorded results. Preserved specimens of six, F. catus,were obtained from NASCO Biological Supply Companyand were studied as a comparison to N. nebulosa. Beforedissection, tissue sample cubes were removed for histolog-ical analysis from the metacarpal and metatarsal pads(�10 mm2 for N. nebulosa and 5 mm2 for the domestic cat)as well as the digital pads (�5 mm2 for N. nebulosa and2–3 mm2 for F. catus). Each sample of tissue included epi-dermis, dermis and subdermis, ending at the deep fasciacovering the muscle or tendons of the metacarpal or meta-tarsal pads. One sample block was taken from of the cen-ter and one from the medial and lateral side of themetacarpal or metatarsal pads of manus and pes for eachN. nebulosa. A single sample was removed from the centerof the fore and hind paw pads of two F. catus specimens.One sample was obtained from each of digits 2–4 of eachmanus and pes. Samples were placed in buffered formalinfor 1 week then dehydrated and embedded in paraffin.The tissue blocks were sectioned yielding 10–15 serialsections (depending on the size) which were placed on aglass slide for staining using standard histological meth-ods (Presnell and Schreibman, 1997). Three slidesfrom each sample block were chosen randomly. One eachwas stained with either hemotoxylin and eosin (H&E,Presnell and Schreibman, 1997), a gold nerve stain (Gar-vey et al., 1987) or Verhoeff ’s elastin stain (Presnell and
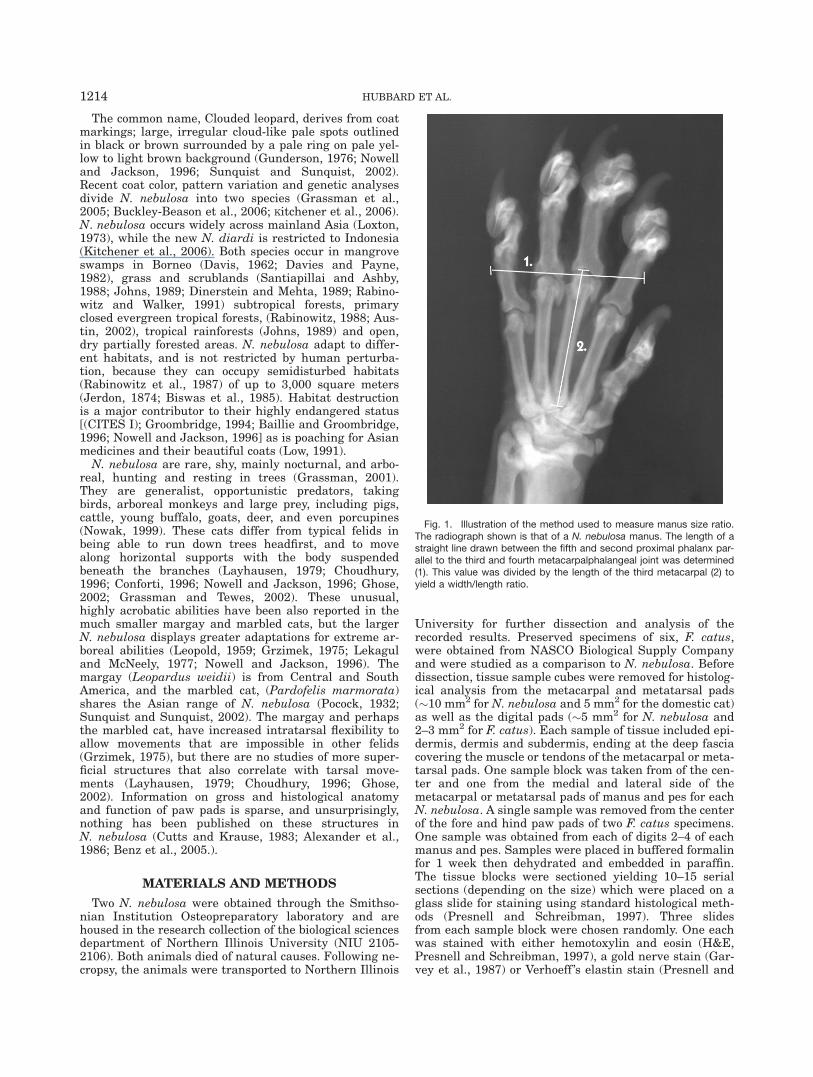
Fig. 1. Illustration of the method used to measure manus size ratio.The radiograph shown is that of a N. nebulosa manus. The length of astraight line drawn between the fifth and second proximal phalanx par-allel to the third and fourth metacarpalphalangeal joint was determined(1). This value was divided by the length of the third metacarpal (2) toyield a width/length ratio.
1214 HUBBARD ET AL.

Schreibman, 1997). A representative section was chosenat random in each H&E slide. The stratium corniumlayer and combined noncornified layers were then meas-ured separately at 40� for the N. nebulosa or at 100�for F. catus using an ocular micrometer (calibrated at 40�and 100� power using the 1/10 mm etched squares ina hemocytometer). The relative adipose content invarious layers of the sample, nerve fiber numbers andlocation, and elastic fiber density were noted for eachslide.
Radiographic images of N. nebulosa and F. catus wereused to compare the relative size of the manus and pes.Radiographs were obtained using a portable GeneralElectric Mobile 100-15 X-ray machine and AXB-90 AlphaImages Medical high speed film. For each specimen,each paw was X-rayed separately in an anterior/poste-rior position. Following film development the radio-graphs were analyzed by measuring the distancebetween the lateral boarders of digits 2 and 5 with aline that passed between the metacarpal/metatarsal pha-langeal joints of digits 3 and 4 (line 1, Fig. 1). A secondmeasurement was made of the length of the third meta-
carpal bone (line 2, Fig. 1). These two measurementswere then expressed as a ratio (width/length) and aver-aged for the manus and pes of each animal.
RESULTSAnatomy of Manus and Pes Metacarpal,Metatarsal and Digital Pads
F. catus
Manus. Although the anatomy of F. catus has beenmore completely documented than that of many domesticspecies, many aspects remain to be investigated (Reighartand Jennings, 1925; Crouch, 1969; Walker, 1986; Rose-nzweig, 1990). In particular, we were interested in study-ing the structural features of the metacarpal, metatarsaland digital pads in the manus and pes that may assist inlocomotor activities. The most notable of our findings wasthe presence of a group of small ligamentous structuresthat are collectively designated in this article as eithermetacarpal (manus) or metatarsal (pes) pad suspensoryligaments. These structures project into the connective
Fig. 2. This illustration is a composite diagram of gross and histo-logic structures observed in the manus of N. nebulosa. (1) Metacarpalsuspensory ligaments project from the M. flexor digitorum superficialistendons of digits 3, 4, and 5 into the metacarpal fat pad and appearto attach to regions adjacent to the epidermis. (2) Metacarpal padsuspensory ligaments extend from the manica flexoria of digit 2 into
the metacarpal fat pad. (3) Metacarpal pad suspensory ligaments fromthe distal end of manica flexoria of digits 2–4 into the metacarpal padconnective tissue. Bundles of collagen fibers running parallel to theepidermis and adipose tissue compartments are present in the subcu-taneous compartment.
ANALYSIS OF PAW PAD STRUCTURE 1215
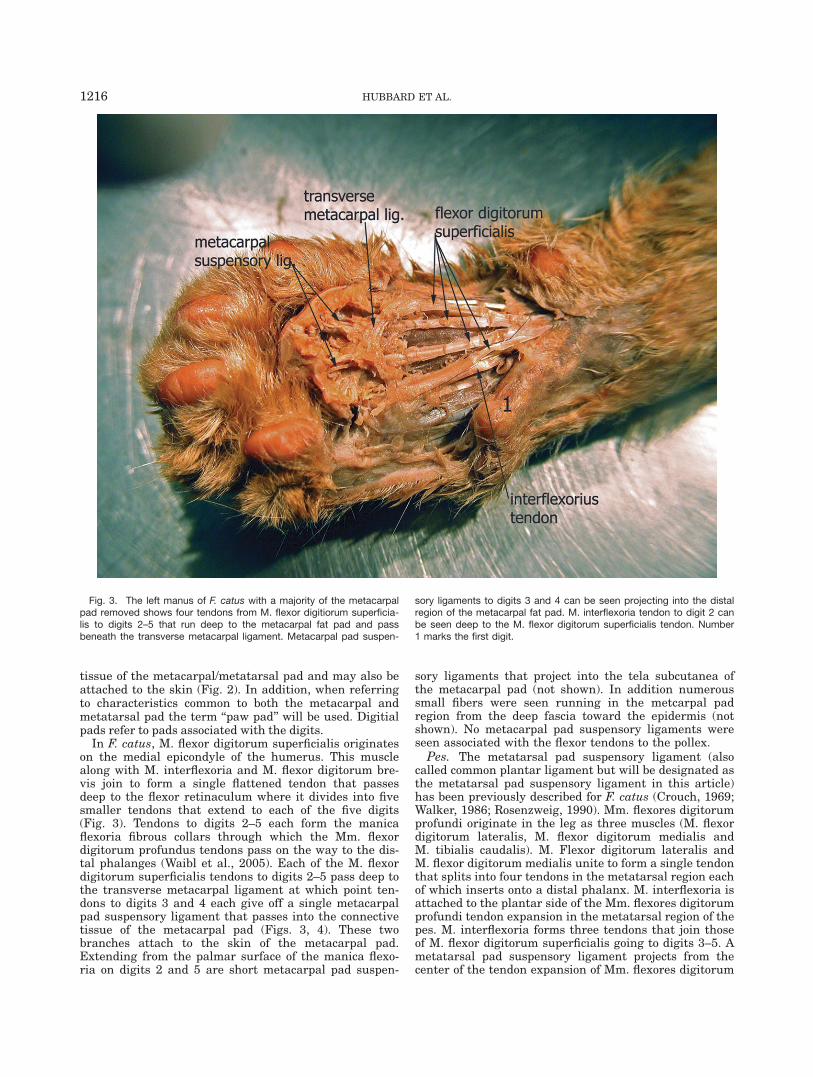
tissue of the metacarpal/metatarsal pad and may also beattached to the skin (Fig. 2). In addition, when referringto characteristics common to both the metacarpal andmetatarsal pad the term ‘‘paw pad’’ will be used. Digitialpads refer to pads associated with the digits.
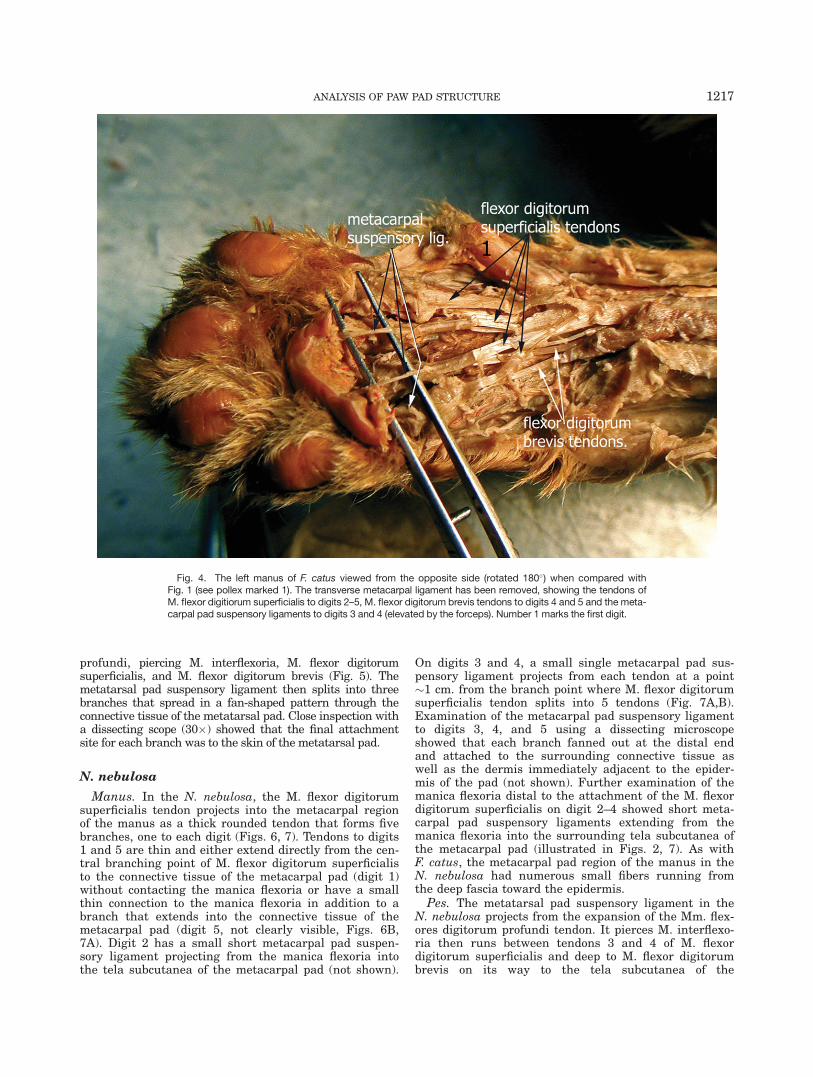
In F. catus, M. flexor digitorum superficialis originateson the medial epicondyle of the humerus. This musclealong with M. interflexoria and M. flexor digitorum bre-vis join to form a single flattened tendon that passesdeep to the flexor retinaculum where it divides into fivesmaller tendons that extend to each of the five digits(Fig. 3). Tendons to digits 2–5 each form the manicaflexoria fibrous collars through which the Mm. flexordigitorum profundus tendons pass on the way to the dis-tal phalanges (Waibl et al., 2005). Each of the M. flexordigitorum superficialis tendons to digits 2–5 pass deep tothe transverse metacarpal ligament at which point ten-dons to digits 3 and 4 each give off a single metacarpalpad suspensory ligament that passes into the connectivetissue of the metacarpal pad (Figs. 3, 4). These twobranches attach to the skin of the metacarpal pad.Extending from the palmar surface of the manica flexo-ria on digits 2 and 5 are short metacarpal pad suspen-
sory ligaments that project into the tela subcutanea ofthe metacarpal pad (not shown). In addition numeroussmall fibers were seen running in the metcarpal padregion from the deep fascia toward the epidermis (notshown). No metacarpal pad suspensory ligaments wereseen associated with the flexor tendons to the pollex.Pes. The metatarsal pad suspensory ligament (also
called common plantar ligament but will be designated asthe metatarsal pad suspensory ligament in this article)has been previously described for F. catus (Crouch, 1969;Walker, 1986; Rosenzweig, 1990). Mm. flexores digitorumprofundi originate in the leg as three muscles (M. flexordigitorum lateralis, M. flexor digitorum medialis andM. tibialis caudalis). M. Flexor digitorum lateralis andM. flexor digitorum medialis unite to form a single tendonthat splits into four tendons in the metatarsal region eachof which inserts onto a distal phalanx. M. interflexoria isattached to the plantar side of the Mm. flexores digitorumprofundi tendon expansion in the metatarsal region of thepes. M. interflexoria forms three tendons that join thoseof M. flexor digitorum superficialis going to digits 3–5. Ametatarsal pad suspensory ligament projects from thecenter of the tendon expansion of Mm. flexores digitorum
Fig. 3. The left manus of F. catus with a majority of the metacarpalpad removed shows four tendons from M. flexor digitiorum superficia-lis to digits 2–5 that run deep to the metacarpal fat pad and passbeneath the transverse metacarpal ligament. Metacarpal pad suspen-
sory ligaments to digits 3 and 4 can be seen projecting into the distalregion of the metacarpal fat pad. M. interflexoria tendon to digit 2 canbe seen deep to the M. flexor digitorum superficialis tendon. Number1 marks the first digit.
1216 HUBBARD ET AL.
profundi, piercing M. interflexoria, M. flexor digitorumsuperficialis, and M. flexor digitorum brevis (Fig. 5). Themetatarsal pad suspensory ligament then splits into threebranches that spread in a fan-shaped pattern through theconnective tissue of the metatarsal pad. Close inspection witha dissecting scope (30�) showed that the final attachmentsite for each branch was to the skin of the metatarsal pad.
N. nebulosa
Manus. In the N. nebulosa, the M. flexor digitorumsuperficialis tendon projects into the metacarpal regionof the manus as a thick rounded tendon that forms fivebranches, one to each digit (Figs. 6, 7). Tendons to digits1 and 5 are thin and either extend directly from the cen-tral branching point of M. flexor digitorum superficialisto the connective tissue of the metacarpal pad (digit 1)without contacting the manica flexoria or have a smallthin connection to the manica flexoria in addition to abranch that extends into the connective tissue of themetacarpal pad (digit 5, not clearly visible, Figs. 6B,7A). Digit 2 has a small short metacarpal pad suspen-sory ligament projecting from the manica flexoria intothe tela subcutanea of the metacarpal pad (not shown).
On digits 3 and 4, a small single metacarpal pad sus-pensory ligament projects from each tendon at a point�1 cm. from the branch point where M. flexor digitorumsuperficialis tendon splits into 5 tendons (Fig. 7A,B).Examination of the metacarpal pad suspensory ligamentto digits 3, 4, and 5 using a dissecting microscopeshowed that each branch fanned out at the distal endand attached to the surrounding connective tissue aswell as the dermis immediately adjacent to the epider-mis of the pad (not shown). Further examination of themanica flexoria distal to the attachment of the M. flexordigitorum superficialis on digit 2–4 showed short meta-carpal pad suspensory ligaments extending from themanica flexoria into the surrounding tela subcutanea ofthe metacarpal pad (illustrated in Figs. 2, 7). As withF. catus, the metacarpal pad region of the manus in theN. nebulosa had numerous small fibers running fromthe deep fascia toward the epidermis.Pes. The metatarsal pad suspensory ligament in the
N. nebulosa projects from the expansion of the Mm. flex-ores digitorum profundi tendon. It pierces M. interflexo-ria then runs between tendons 3 and 4 of M. flexordigitorum superficialis and deep to M. flexor digitorumbrevis on its way to the tela subcutanea of the
Fig. 4. The left manus of F. catus viewed from the opposite side (rotated 180�) when compared withFig. 1 (see pollex marked 1). The transverse metacarpal ligament has been removed, showing the tendons ofM. flexor digitiorum superficialis to digits 2–5, M. flexor digitorum brevis tendons to digits 4 and 5 and the meta-carpal pad suspensory ligaments to digits 3 and 4 (elevated by the forceps). Number 1 marks the first digit.
ANALYSIS OF PAW PAD STRUCTURE 1217
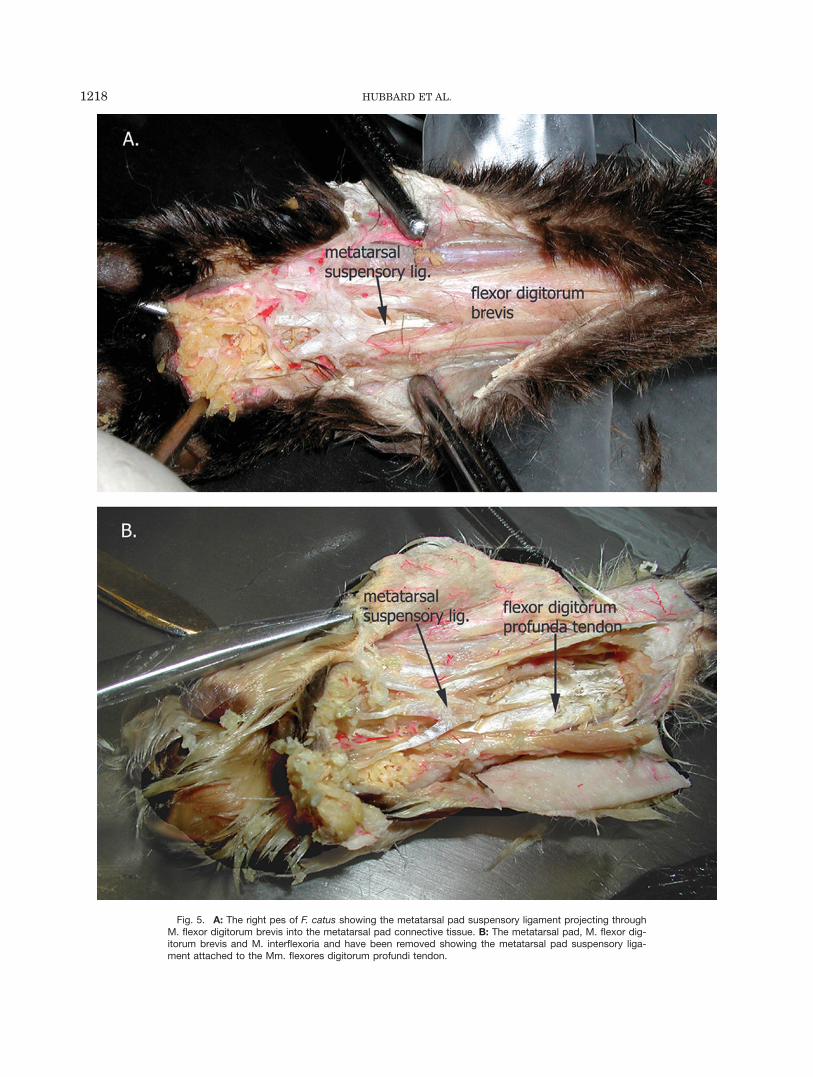
Fig. 5. A: The right pes of F. catus showing the metatarsal pad suspensory ligament projecting throughM. flexor digitorum brevis into the metatarsal pad connective tissue. B: The metatarsal pad, M. flexor dig-itorum brevis and M. interflexoria and have been removed showing the metatarsal pad suspensory liga-ment attached to the Mm. flexores digitorum profundi tendon.
1218 HUBBARD ET AL.
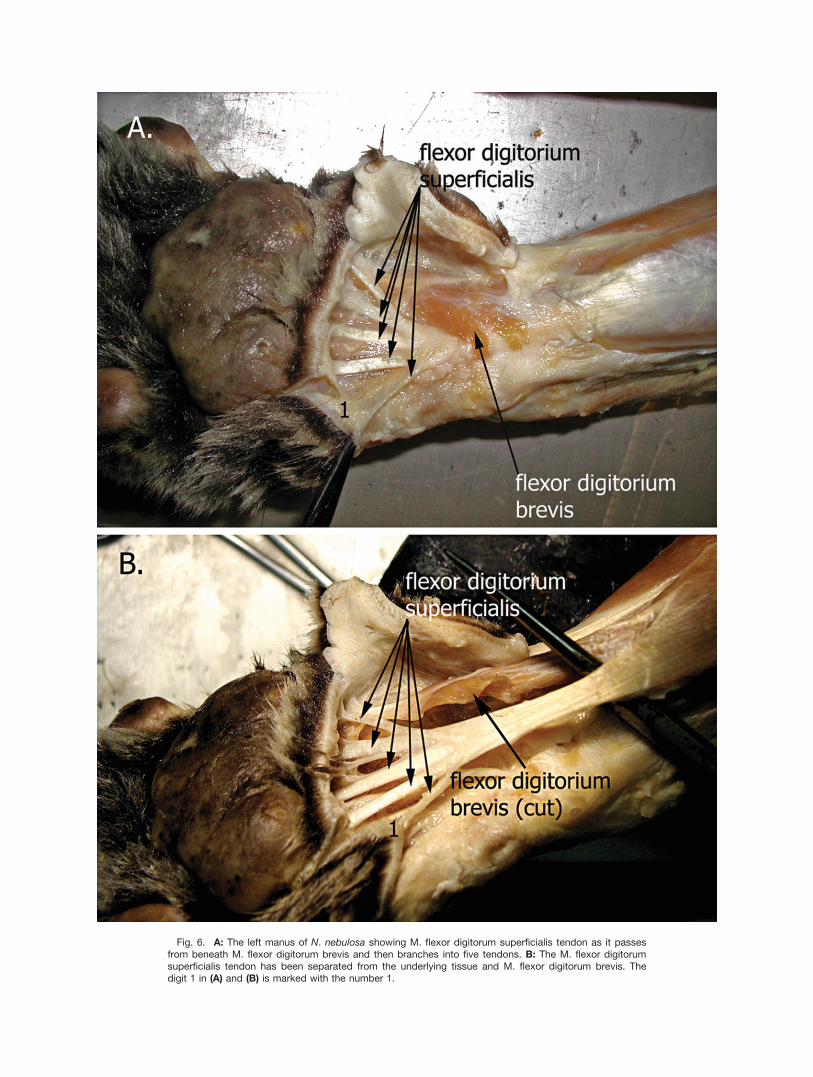
Fig. 6. A: The left manus of N. nebulosa showing M. flexor digitorum superficialis tendon as it passesfrom beneath M. flexor digitorum brevis and then branches into five tendons. B: The M. flexor digitorumsuperficialis tendon has been separated from the underlying tissue and M. flexor digitorum brevis. Thedigit 1 in (A) and (B) is marked with the number 1.
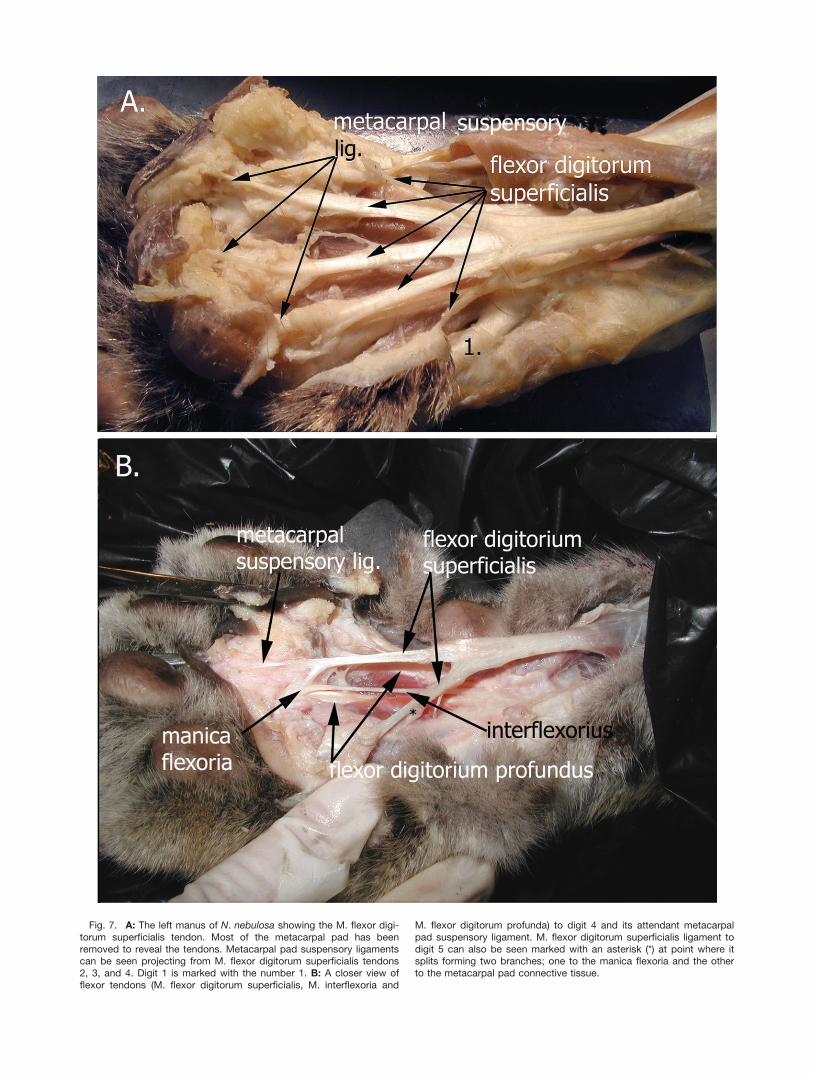
Fig. 7. A: The left manus of N. nebulosa showing the M. flexor digi-torum superficialis tendon. Most of the metacarpal pad has beenremoved to reveal the tendons. Metacarpal pad suspensory ligamentscan be seen projecting from M. flexor digitorum superficialis tendons2, 3, and 4. Digit 1 is marked with the number 1. B: A closer view offlexor tendons (M. flexor digitorum superficialis, M. interflexoria and
M. flexor digitorum profunda) to digit 4 and its attendant metacarpalpad suspensory ligament. M. flexor digitorum superficialis ligament todigit 5 can also be seen marked with an asterisk (*) at point where itsplits forming two branches; one to the manica flexoria and the otherto the metacarpal pad connective tissue.
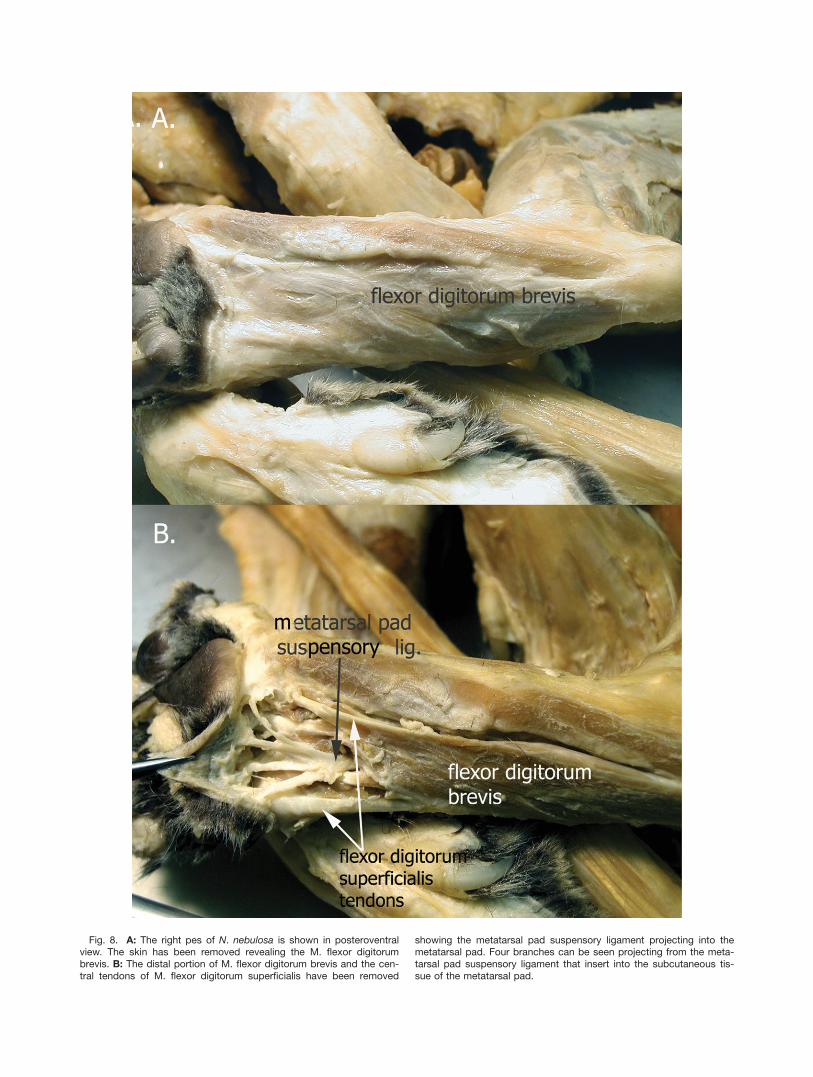
Fig. 8. A: The right pes of N. nebulosa is shown in posteroventralview. The skin has been removed revealing the M. flexor digitorumbrevis. B: The distal portion of M. flexor digitorum brevis and the cen-tral tendons of M. flexor digitorum superficialis have been removed
showing the metatarsal pad suspensory ligament projecting into themetatarsal pad. Four branches can be seen projecting from the meta-tarsal pad suspensory ligament that insert into the subcutaneous tis-sue of the metatarsal pad.

metatarsal pad (Figs. 8, 9). This ligament forms threebranches that pass through the connective tissue andattach to the skin of the medial, lateral and centralregions of the metatarsal pad (Fig. 8B).
In addition to the tendons described earlier, numeroussmall anchoring fibers were seen running through theadipose connective tissue of the metatarsal pad in boththe manus and pes (not shown). These fibers appear tobe attached to the tendon sheaths deep to the metatarsalpad. They pass through the connective tissue and attachto the skin of the metatarsal pad.
Histological Analysis
The central thickness of the metacarpal and metatar-sal pads extending from the muscle tendons to the epi-dermal surface in F. catus averaged 4 mm (3.4–4.5 mm).Central digital pad thickness averaged 2 mm (1.25–2.5 mm).
In the N. nebulosa, the metacarpal and metatarsalpad thickness were each �10 mm (8–12 mm) while thedigital pads were �4 mm (3.4–4.5 mm) at the thickestpoint.
Tissue samples stained with H&E from F. catus andN. nebulosa showed similar morphologic characteristicsexcept for the epithelial thickness. Initial analysis ofsample measurements taken from the medial, lateral,and central metacarpal and metatarsal pads in theN. nebulosa and F. catus were not significantly different
Fig. 9. A deep dissection of the right pes of N. nebulosa showingthe Mm. flexores digitorum profundi tendon and the attachment site ofthe metatarsal pad suspensory ligament. The M. flexor digitorum bre-vis and the M. flexor digitorum superficialis tendon have been
detached and folded toward the distal end of the pes. Mm. interflexoriihas been removed to show the attachment site of the metatarsal padsuspensory ligament. M. quadratus plantae can also be seen attachedto Mm. flexores digitorum profundi tendon.
TABLE 1. The thickness of the stratiumcornium compared to non-cornified layers of the
epidermis in the metatarsal, metacarpal and digitalpads of the N. nebulosa and domestic cat
N. nebulosa Metacarpal pad corn. 93.8 � 1.41
Metatarsal pad corn. 97.0 � 2.81
Metacarpal pad noncorn. 24.5 � 0.22
Metatarsal noncorn. 22.5 � 0.32
Manus digit pad corn. 76.5 � 1.23
Pes digit pad corn. 79.4 � 1.83
Manus digit pad noncorn. 17.8 � 0.64
Pes digit pad noncorn. 14.4 � 0.24
F. catus Metacarpal pad corn. 26.1 � 0.52
Metatarsal pad corn. 25.8 � 0.32
Metacarpal pad noncorn. 10.8 � 0.15
Metatarsal pad noncorn. 10.9 � 0.35
Manus digit pad corn. 17.1 � 0.34
Pes digit pad corn. 21.5 � 0.32
Manus digit pad noncorn. 8.5 � 0.15
Pes digit pad noncorn. 10.3 � 0.25
Values are expressed as mean lm � SEM. For both N. neb-ulosa and F. catus, metacarpal or metartarsal pads, N ¼ 8,digital pads N ¼ 24. Values with different numbers are sig-nificantly different from each other (P � 0.05).Abbreviations: corn, stratum corneum; noncorn, non-corni-fied layers.
1222 HUBBARD ET AL.
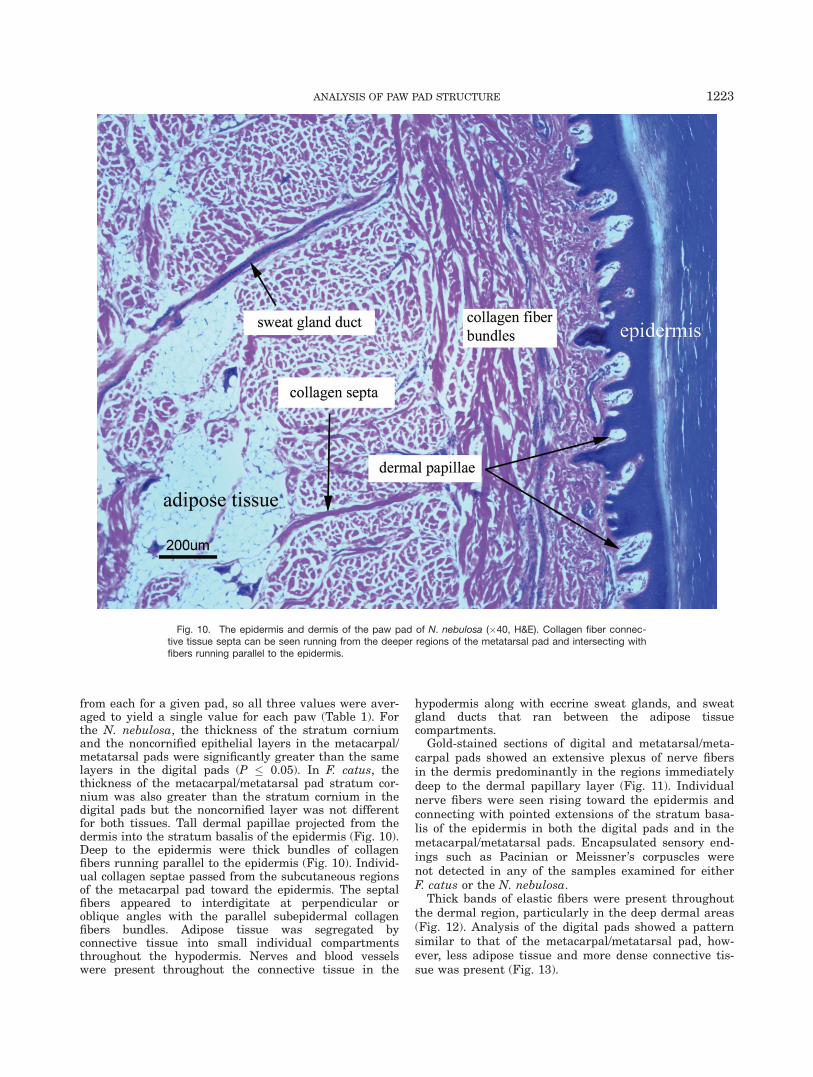
from each for a given pad, so all three values were aver-aged to yield a single value for each paw (Table 1). Forthe N. nebulosa, the thickness of the stratum corniumand the noncornified epithelial layers in the metacarpal/metatarsal pads were significantly greater than the samelayers in the digital pads (P � 0.05). In F. catus, thethickness of the metacarpal/metatarsal pad stratum cor-nium was also greater than the stratum cornium in thedigital pads but the noncornified layer was not differentfor both tissues. Tall dermal papillae projected from thedermis into the stratum basalis of the epidermis (Fig. 10).Deep to the epidermis were thick bundles of collagenfibers running parallel to the epidermis (Fig. 10). Individ-ual collagen septae passed from the subcutaneous regionsof the metacarpal pad toward the epidermis. The septalfibers appeared to interdigitate at perpendicular oroblique angles with the parallel subepidermal collagenfibers bundles. Adipose tissue was segregated byconnective tissue into small individual compartmentsthroughout the hypodermis. Nerves and blood vesselswere present throughout the connective tissue in the
hypodermis along with eccrine sweat glands, and sweatgland ducts that ran between the adipose tissuecompartments.
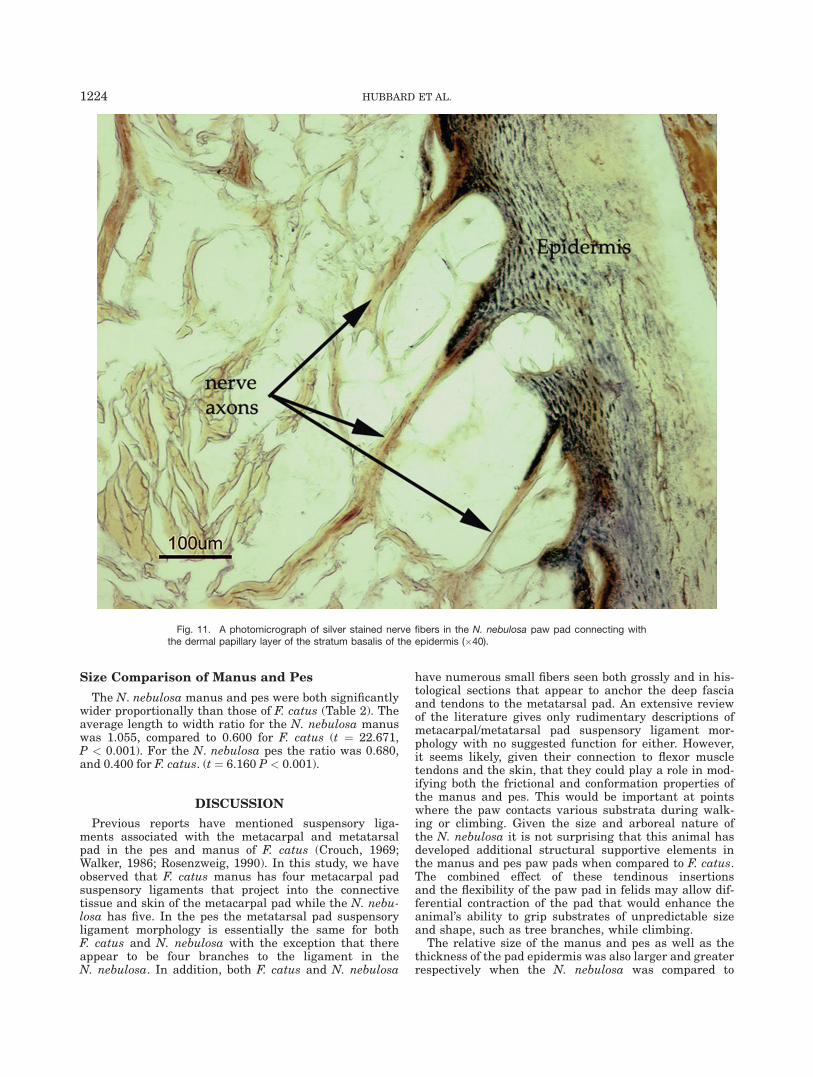
Gold-stained sections of digital and metatarsal/meta-carpal pads showed an extensive plexus of nerve fibersin the dermis predominantly in the regions immediatelydeep to the dermal papillary layer (Fig. 11). Individualnerve fibers were seen rising toward the epidermis andconnecting with pointed extensions of the stratum basa-lis of the epidermis in both the digital pads and in themetacarpal/metatarsal pads. Encapsulated sensory end-ings such as Pacinian or Meissner’s corpuscles werenot detected in any of the samples examined for eitherF. catus or the N. nebulosa.
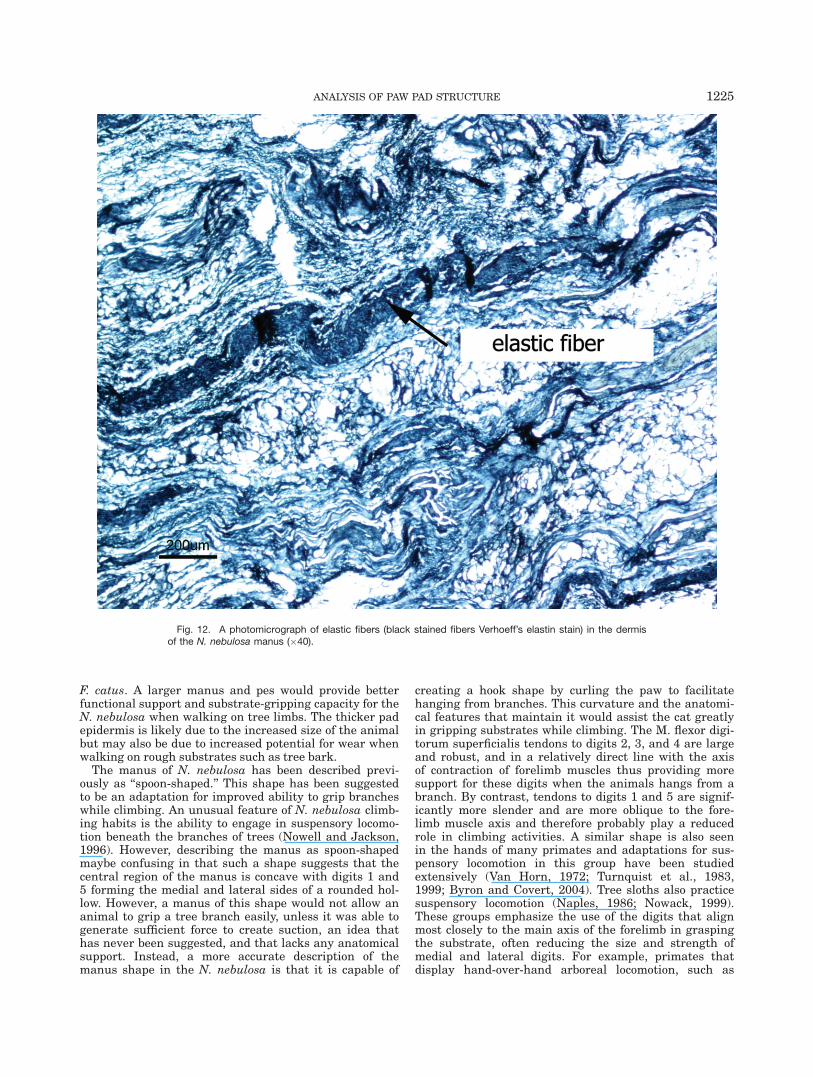
Thick bands of elastic fibers were present throughoutthe dermal region, particularly in the deep dermal areas(Fig. 12). Analysis of the digital pads showed a patternsimilar to that of the metacarpal/metatarsal pad, how-ever, less adipose tissue and more dense connective tis-sue was present (Fig. 13).
Fig. 10. The epidermis and dermis of the paw pad of N. nebulosa (�40, H&E). Collagen fiber connec-tive tissue septa can be seen running from the deeper regions of the metatarsal pad and intersecting withfibers running parallel to the epidermis.
ANALYSIS OF PAW PAD STRUCTURE 1223
Size Comparison of Manus and Pes
The N. nebulosa manus and pes were both significantlywider proportionally than those of F. catus (Table 2). Theaverage length to width ratio for the N. nebulosa manuswas 1.055, compared to 0.600 for F. catus (t ¼ 22.671,P < 0.001). For the N. nebulosa pes the ratio was 0.680,and 0.400 for F. catus. (t ¼ 6.160 P < 0.001).
DISCUSSION
Previous reports have mentioned suspensory liga-ments associated with the metacarpal and metatarsalpad in the pes and manus of F. catus (Crouch, 1969;Walker, 1986; Rosenzweig, 1990). In this study, we haveobserved that F. catus manus has four metacarpal padsuspensory ligaments that project into the connectivetissue and skin of the metacarpal pad while the N. nebu-losa has five. In the pes the metatarsal pad suspensoryligament morphology is essentially the same for bothF. catus and N. nebulosa with the exception that thereappear to be four branches to the ligament in theN. nebulosa. In addition, both F. catus and N. nebulosa
have numerous small fibers seen both grossly and in his-tological sections that appear to anchor the deep fasciaand tendons to the metatarsal pad. An extensive reviewof the literature gives only rudimentary descriptions ofmetacarpal/metatarsal pad suspensory ligament mor-phology with no suggested function for either. However,it seems likely, given their connection to flexor muscletendons and the skin, that they could play a role in mod-ifying both the frictional and conformation properties ofthe manus and pes. This would be important at pointswhere the paw contacts various substrata during walk-ing or climbing. Given the size and arboreal nature ofthe N. nebulosa it is not surprising that this animal hasdeveloped additional structural supportive elements inthe manus and pes paw pads when compared to F. catus.The combined effect of these tendinous insertionsand the flexibility of the paw pad in felids may allow dif-ferential contraction of the pad that would enhance theanimal’s ability to grip substrates of unpredictable sizeand shape, such as tree branches, while climbing.
The relative size of the manus and pes as well as thethickness of the pad epidermis was also larger and greaterrespectively when the N. nebulosa was compared to
Fig. 11. A photomicrograph of silver stained nerve fibers in the N. nebulosa paw pad connecting withthe dermal papillary layer of the stratum basalis of the epidermis (�40).
1224 HUBBARD ET AL.
F. catus. A larger manus and pes would provide betterfunctional support and substrate-gripping capacity for theN. nebulosa when walking on tree limbs. The thicker padepidermis is likely due to the increased size of the animalbut may also be due to increased potential for wear whenwalking on rough substrates such as tree bark.
The manus of N. nebulosa has been described previ-ously as ‘‘spoon-shaped.’’ This shape has been suggestedto be an adaptation for improved ability to grip brancheswhile climbing. An unusual feature of N. nebulosa climb-ing habits is the ability to engage in suspensory locomo-tion beneath the branches of trees (Nowell and Jackson,1996). However, describing the manus as spoon-shapedmaybe confusing in that such a shape suggests that thecentral region of the manus is concave with digits 1 and5 forming the medial and lateral sides of a rounded hol-low. However, a manus of this shape would not allow ananimal to grip a tree branch easily, unless it was able togenerate sufficient force to create suction, an idea thathas never been suggested, and that lacks any anatomicalsupport. Instead, a more accurate description of themanus shape in the N. nebulosa is that it is capable of
creating a hook shape by curling the paw to facilitatehanging from branches. This curvature and the anatomi-cal features that maintain it would assist the cat greatlyin gripping substrates while climbing. The M. flexor digi-torum superficialis tendons to digits 2, 3, and 4 are largeand robust, and in a relatively direct line with the axisof contraction of forelimb muscles thus providing moresupport for these digits when the animals hangs from abranch. By contrast, tendons to digits 1 and 5 are signif-icantly more slender and are more oblique to the fore-limb muscle axis and therefore probably play a reducedrole in climbing activities. A similar shape is also seenin the hands of many primates and adaptations for sus-pensory locomotion in this group have been studiedextensively (Van Horn, 1972; Turnquist et al., 1983,1999; Byron and Covert, 2004). Tree sloths also practicesuspensory locomotion (Naples, 1986; Nowack, 1999).These groups emphasize the use of the digits that alignmost closely to the main axis of the forelimb in graspingthe substrate, often reducing the size and strength ofmedial and lateral digits. For example, primates thatdisplay hand-over-hand arboreal locomotion, such as
Fig. 12. A photomicrograph of elastic fibers (black stained fibers Verhoeff’s elastin stain) in the dermisof the N. nebulosa manus (�40).
ANALYSIS OF PAW PAD STRUCTURE 1225

gibbons and spider monkeys, often have thumbs that arereduced in size and are not used as a part of this grip(Van Horn, 1972; Turnquist, et al., 1999). Sloths showhands that are even more restricted to grasping curvedsurfaces by reduction of the digits to three in the brady-podids or even two in choloepids (Nowak, 1999).
This difference in the relative size of the tendons con-trasts with the condition in F. catus, where all of theM. flexor digitorum superficialis tendons are of similarsize. The increase in the relative size of the tendons inthe N. nebulosa paw that are more in direct line withthe bellies of the forearm muscles would better sup-port increased tension by these muscles produced whilemaintaining a hook-shaped paw during suspensionbeneath a tree limb, and can be considered an adapta-tion for this specialized kind of locomotion.
Histological analysis of the metacarpal/metatarsalpads in the N. nebulosa and F. catus shows that they areorganized into adipose tissue compartments surroundedby dense collagen and elastic fibers. The collagen fibersappear to serve as anchors between the subdermalregions and the epidermis in addition to acting as parti-tioning agents. Previous studies in the elephant and
F. catus indicate that the adipose compartments, com-bined with the profusion of elastic fibers impart a cush-ioning effect to the metacarpal and metatarsal pad(Alexander et al., 1986; Ker, 1999; Liebich, 1999; Weis-sengruber et al., 2006). Adipose tissue compartmentshave been suggested to act as the equivalent of a fluid-filled cushion where large volume fluid flow is limitedduring compression by partitioning connective tissueseptae in a way that is similar to isolated springs in abed mattress (Ker, 1999; Weissengruber et al., 2006).This would appear to enhance conformational contouringof the paw pad to variable substrates during walkingand climbing as well as providing a cushioning effect.Tensing of the paw pad tendons when the flexor musclesare active in either the manus or pes would appear tostiffen the pad by pulling on the connective tissue ele-ments surrounding the adipose compartments. This mayact to stabilize the pad further, and enhance the gripduring climbing.
The silver stained slides of the subepidermal region ofthe paw pads showed a large numbers nerve fibers butthe pad was surprisingly devoid of mechanoreceptorssuch as the Pacinian and Meissner corpuscles that are
Fig. 13. The digital pad of the N. nebulosa manus (�40, H&E). Adipose tissue compartments can beseen interspersed between thick bundles of collagen fibers. A sweat gland is also present.
1226 HUBBARD ET AL.

prevalent in the feet, fingers and digital pads of ele-phants, humans, primates, certain rodents and marsu-pials (Weissengruber et al., 2006). This seems unusualgiven that these structures are useful for sensing vari-able surfaces to facilitate walking (Weissengruber et al.,2006). While it is possible that these were located inother regions of the pad that were not tested, it seemsunlikely because all major regions of the metacarpal/metatarsal pads and digital pads were examined. Ofnote, however, is the unusual way in which nerves fromthe dermal region attach to pointed projections of the ep-idermis (see Fig. 11). It is possible that these representsome undermined sensory modality.
Numerous eccrine sweat glands were present in allhistological samples for both the N. nebulosa andF. catus. Previous studies have indicated that sweatfrom these glands improves the frictional capacity of thepaw pad and provides scent markings (Meyer and Bar-tels, 1989). It is likely the function is similar in F. catusand N. nebulosa.
ACKNOWLEDGMENTS
The authors thank Charley Potter and John Ososky andthe Smithsonian Institution for facilitating this study.
LITERATURE CITED
Alexander RMcN, Bennett MB, Ker RF. 1986. Mechanical proper-ties and function of the metatarsal pads of some mammals. J ZoolLond 209:405–419.
Austin SC. 2002. Ecology of sympatric carnivores in Khao YaiNational Park, Thailand. Ph.D. Dissertation. Texas A& M Univer-sity–Kingsville and Texas A&M–College Station, Kingsville andCollege Station.
Baillie J, Groombridge B, editors. 1996. Red list of threatened ani-mals. Switzerland: IUCN, Gland.
Benz A, Zenker W, Hildebrandt TB, Weissengruber G, Geyer H. 2005.About the macroscopic and microscopic morphology of elephantshooves (Elephantidae) Verh Ber Erkrg Zootlere 42:167–170.
Biswas B, Ghose RK, Ghosel DK. 1985. Progress Report 2 on pilotsurvey of the WWF- India/Zoological survey of India collaborativeproject on the status of the lesser cats in eastern India. ZoologicalSurvey of India, Calcutta.
Buckley-Beason VA, Johnson W, Nash WG, Stanyonp R, MenningerJC, Driscoll CA, Howard J, Bush M, Page JE, Roelke ME, StoneG, Martinelli PP, Wen C, Ling L, Duraisingham RK, Lam PV,O’Brien S. 2006. Molecular evidence for species-level distinctionsin N. nebulosa. Curr Biol 16:2371–2376.
Byron CD, Covert HH. 2004. Unexpected locomotor behavior: bra-chiation by an old world monkey (Pygathrix nemaceus). J Zool263:101–106.
Choudhury A. 1996. The marbled cat, Felis marmorata in Assam—some recent records. J Bombay Nat Hist Soc 93:583–584.
Christiansen P. 2006. Sabertooth characters in the clouded leopard(Neofelis nebulosa Griffiths 1821). J Morphol 267:1186–1198.
Conforti K. 1996. The status and distribution of small carnivoresin the Huai Kaeng/Thung Yai Naresuan Wildlife Sanctuaries,west-central Thailand. M.S. Thesis, Univesrity of Minnesota,Minneapolis-St. Paul.
Crouch JE. 1969. Text-Atlas of cat anatomy. Philadelphia: Lea andFebinger.
Cutts JH, Krause WJ. 1983. Structure of the paws in Didelphis vir-giniana. Anat Anz 154:329–335.
Davies G, Payne J. 1982. A journal survey of Sabah. UnpublishedReport WWF Maylasia for the Game Branch, Forest Department,Sabah.
Davis DD. 1962. Mammals of the lowland rainforest of North Bor-neo. Bull Nat Hist Mus (Singapore) 31:1–129.
Dinerstein E, Mehta JN. 1989. The clouded leopard in Nepal. Oryx23:199–201.
Garvey W, Fathi A, Bigelow F, Jimenez C, Carpenter B. 1987 A newreliable silver impregnation technique for nervous system. J His-totechnol 10:245–247.
Ghose D. 2002. First sighting of clouded leopard Neofelis nebulosafrom the Blue Mountain National Park, Mizoram, India. Curr Sci83:20–21.
Grassman LI. 2001. Spatial ecology and conservation of the felidcommunity in Phu Khieo Wildlife Sanctuary, Thailand. Report toCat Action Treasury.
Grassman LI, Tewes ME. 2002. Observations of a marbled cat pairin northeastern Thailand. The World Conservation Union, CatSpecialist Group, Bougy, Switzerland. Cat News 36:9.
Grassman LI, Tewes ME, Silvy NJ, Kreetiyutanovet K. 2005. Ecol-ogy of three sympatric felids in a mixed evergreen forest inNorth-central Thailand. J Mammal 86:29–38.
Groombridge B, editor. 1994. IUCN Red list of threatened animals.Switzerland: IUCN, Gland.
Grzimek B, editor. 1975. Grzimek’s animal life encyclopedia: mam-mals, Vol. 1–13. New York: Van Nostrand, Reinhold.
Guggisberg CAW. 1975. Wild cats of the World. London: David andCharles.
Gunderson HL. 1976. Mammalogy. New York: McGraw-Hill. p 483.Jerdon TC. 1874. The Mammals of India: natural history. London:
John Wheldon Press.Johns AD. 1989. Timber, the environment and wildlife in Malaysian
rainforests. Final Report to Institute of South-East Asian Biology.Aberdeen, Scotland, U.K.: University of Aberdeen.
Ker RF. 1999. The design of soft collagenous load-bearing tissues.J Exp Biol 202:3315–3324.
Kitchener AC, Beaumont MA, Richardson D. 2006. Geographicalvariation in the clouded leopard, Neofelis nebulosa, reveals twospecies. Curr Biol 16:2377–2383.
Layhausen P. 1979. Cat behavior: the predatory and social behaviorof domestic and wild cats. New York: Garland Press.
Lekagul B, McNeely JA. 1977. Mammals of Thailand. Bankok:Sahakarnbhat. p 758.
Leopold AS. 1959. Wildlife of Mexico. Berkeley: Univ. CaliforniaPress. p xxxi þ 568
TABLE 2. Comparison of the manus and pessize ratio in F. catus and N. nebulosa
Width (cm) Length (cm) W/L Ratio
ManusN. nebulosa 1 4.40 4.15 1.06N. nebulosa 2 5.20 4.95 1.05
Mean 1.055F. catus 1 1.70 2.75 0.62F. catus 2 1.55 2.70 0.57F. catus 3 1.75 2.85 0.61
Mean 0.600
PesN. nebulosa 1 3.80 5.90 0.64N. nebulosa 2 4.50 6.20 0.73
Mean 0.680F. catus 1 1.45 4.15 0.35F. catus 2 1.85 4.40 0.42F. catus 3 1.90 4.45 0.43
Mean 0.400
Analysis using two-sample Student’s t test showed signifi-cant differences in the W/L ratios between the N. nebulosaand F. catus in both the manus (t ¼ 22.671, P < 0.001) andpes (t ¼ 6.160, P < 0.001).
ANALYSIS OF PAW PAD STRUCTURE 1227

Liebich HG. 1999. Funktionelle Histologie der Haussaugetiere. 3rded. Stuttgart: Schattauer.
Low J. 1991. The smuggling of endangered wildlife across the Tai-wan Strait. Cambridge: TRAFFIC International.
Loxton H. 1973. The beauty of big cats. London: Triune Books.Meyer W, Bartels T. 1989. Histochemical study on the eccrine
glands in the metatarsal pad of the cat. Basic Appl Histochem33:219–238.
Naples VL. 1986. The morphology and function of the hyoid regionin the tree sloths, Bradypus and Choloepus. J Mammal 67:712–724.
Nowak RM. 1999. Walker’s Mammals of the World. 6th ed.Baltimore, London: Johns Hopkins University Press, Vol. 1:p 1–836.
Nowell K, Jackson P. 1996. Wild cats: status survey and conserva-tion action plan. IUCN (World Conservation Union), Gland, Swit-zerland. p xxiv, 382.
Pocock RI. 1932. The marbled cat (Pardofelis marmorata) and someother oriental species, with a definition of new genus of the Feli-dae. Proc Zool Soc Lond 741–766.
Presnell JK, Schreibman MP. 1997. Humason’s anatomical tissuetechniques. 5th ed. Baltimore, London: Johns Hopkins UniversityPress.
Rabinowitz AR. 1988. The clouded leopard in Taiwan. Oryx 22:46–47.
Rabinowitz AR, Walker SR. 1991. The carnivore community in adry tropical forest mosaic in Huai Khe Khaeng Wildlife Sanctu-ary, Thailand. J Trop Ecol 7:37–47.
Rabinowitz AR, Andau P, Chai PPK. 1987. The clouded leopard inMalaysian Borneo. Oryx 22:107–111.
Reighart JE, Jennings HS. 1925. Anatomy of the cat. New York:Henry Holt and Company.
Rosenzweig LJ. 1990 Anatomy of the cat. Text and dissection guide.Dubuque, Iowa: Wm C Brown.
Santiapillai C, Ashby KR. 1988. The clouded leopard in Sumatra.Oryx 22:44–45.
Sunquist ME, Sunquist F. 2002. Wild cats of the world. UnitedKingdom: Cambridge University Press.
Turnquist JE, Schmidt D, Rose MD, Cant JG. 1983. Forelimb mus-culature and ligaments in Ateles, the spider monkey. Am J PhysAnthropol 62:209–226.
Turnquist JE, Schmidt D, Rose MD, Cant JG. 1999. Pendularmotion in the brachiation of captive Lagothrix and Ateles. Am JPrimatol 48:263–281.
Van Horn RN. 1972. Structural adaptations to climbing in the gib-bon hand. Am Anthropol 74:326–334.
Waibl H, Gasse H, Hashimoto Y, Budras KD, Constatinescu GM,Saber AS, Simoens P, Salazar I, Sotonyi P, Augsburger H, Bra-gulla A. 2005. Nomina anatomical Veterinaria. 5th ed. Hannover:International committee on Veterinary Gross Anatomical Nomen-clature. World Association of Veterinary Anatomists.
Walker WF. 1986. Vertebrate dissection. Sanders College PublicationsPhiladelphia, PA.
Weissengruber GE, Egger GF, Hutchinson JR, Groenewald HB,Elsasser L, Famini D, Forstenpointner G. 2006. The structure ofthe cushions in the feet of African elephants (Loxodonta afri-cana). J Anat 209:781–792.
Wozencraft WC. 2005. The clouded leopard. In: Wilson, DE, ReederDM, editors. Mammal Species of the World, 3rd ed. Baltimore,London: Johns Hopkins Press. p 545–546.
Yamada JK, Durant BS. 1989. Reproductive parameters of cloudedleopard (Neofelis nebulosa). Zoo Biol 8:223–231.
1228 HUBBARD ET AL.





























