Embed Size (px)
Citation preview

R E S E A R C H A R T I C L E
Characterizationofmicrobial communities in theaqueousphaseofa constructedmodelwetland treating1,2-dichloroethene-contaminatedgroundwaterGwenael Imfeld1, Cristian Estop Aragones1, Ingo Fetzer2, Eva Meszaros3, Simone Zeiger1, IvonneNijenhuis1, Marcell Nikolausz4, Sylvain Delerce1 & Hans H. Richnow1
1Department of Isotope Biogeochemistry, Helmholtz Centre for Environmental Research – UFZ, Leipzig, Germany; 2Department of Environmental
Microbiology, Helmholtz Centre for Environmental Research – UFZ, Leipzig, Germany; 3Department of Microbiology, Eotvos Lorand University of
Science, Budapest, Hungary; and 4Department of Environmental Biotechnology, Helmholtz Centre for Environmental Research – UFZ, Leipzig, Germany
Correspondence: Gwenael Imfeld,
Laboratory of Hydrology and Geochemistry
of Strasbourg (LHyGeS), University of
Strasbourg/ENGEES, CNRS 1, quai Koch BP
61039, F-67 070 Strasbourg, France. Tel.:
133 0 3 88 24 82 43; fax: 133 0 3 88 24 82
84; e-mail:
Received 26 May 2009; revised 16 November
2009; accepted 16 November 2009.
Final version published online 19 January 2010.
DOI:10.1111/j.1574-6941.2009.00825.x
Editor: Max Haggblom
Keywords
chlorinated ethenes; compound-specific
isotope analysis; DGGE; ordination methods;
Dehalococcoides; biodegradation.
Abstract
The dynamics and composition of microbial communities in the aqueous phase of
a model wetland supplied with cis- and trans-1,2-dichloroethenes (DCE)-con-
taminated groundwater was characterized. PCR-denaturing gradient gel electro-
phoresis analysis of water samples obtained from different parts of the wetland
revealed that changes of the bacterial community structure coincided with a
succession of the hydrochemical conditions in the wetland, from oxic towards
anoxic conditions. During this transition phase, the appearance of vinyl chloride
and ethene correlated with the presence of putative dechlorinating bacteria
(Dehalococcoides spp., Geobacter spp. and Dehalobacter spp.). Additionally, a shift
of the DCE isotopic composition indicated the progressive prevalence of reductive
dechlorination in the wetland. Although the DCE degradation processes varied
over time, biodegradation activity was maintained in the wetland system. 16S
rRNA gene libraries revealed that Proteobacteria accounted for 4 50% of 16S
rRNA genes clone libraries, whereas�17% of the sequences from the wetland were
related to sulphate reducers. Based on a multiple-method approach, this study
illustrates the linkage between microbial community dynamics and composition,
changes of hydrochemical conditions and processes of DCE degradation in a
wetland system.
Introduction
Dichloroethenes (DCE) and carcinogenic vinyl chloride
(VC) often originate from the reductive dechlorination of
tetrachloroethene (PCE) or trichloroethene (TCE) and tend
to accumulate within anoxic aquifers (McCarty & Semperini,
1994; Vogel, 1994; Maymo-Gatell et al., 1997). Over the
course of contaminant migration from ground to surface
water, the contaminant flux can be intercepted by wetlands
(Reddy & Dangelo, 1997; Stottmeister & Wiessner, 2003).
Biodegradation of DCE in wetlands is likely to be controlled
by a complex and dynamic assemblage of adjacent aerobic
and anaerobic zones that typically prevail in biogeochemical
heterogenic environments (Armenante et al., 1992; Master
et al., 2002; Meade & D’Angelo, 2005). Wetland systems
located in draining areas of groundwater may thus contri-
bute to natural attenuation of intermediate chlorinated
hydrocarbons, such as DCE and VC (McCarty & Semperini,
1994; Lorah & Olsen, 1999; Amon et al., 2007). In aerobic
zones, oxidation of DCE can occur metabolically (Bradley &
Chapelle, 2000; Coleman et al., 2002) or cometabolically
(Kim & Semprini, 2005). Alternatively, anaerobic oxidation
of DCE occurring under Mn(IV)-, Fe(III)- or SO4-reducing
conditions has been demonstrated in laboratory experiments
(Bradley & Chapelle, 1998; Hata et al., 2004). Under strictly
anaerobic conditions, DCE can additionally be subjected to
reductive dechlorination and finally transformed to ethene
(Maymo-Gatell et al., 1997). However, little is known about
microbial diversity and biogeochemical processes in wetlands
receiving water fluxes contaminated by organic chemicals.
Recently, constructed wetlands used for water remedia-
tion have been investigated with respect to their ability to
FEMS Microbiol Ecol 72 (2010) 74–88Journal compilation c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original German government works
MIC
ROBI
OLO
GY
EC
OLO
GY

remove chlorinated hydrocarbons. Most investigations so
far have consisted of contaminant concentrations and
isotopic composition analyses (Kassenga et al., 2003; Keefe
et al., 2004; Amon et al., 2007; Braeckevelt et al., 2007;
Imfeld et al., 2008a). However, these analyses can be
efficiently coupled with culture-independent molecular
techniques in order to provide additional information on
the phylogenetic diversity and structure of microbial com-
munities in contaminated wetlands (Nocker et al., 2007;
Weiss & Cozzarelli, 2008). DNA fingerprinting techniques
were applied for characterization of microbial diversity and
community structure dynamics in constructed wetlands
treating domestic and dairy wastewater (Truu et al., 2005;
Ibekwe et al., 2007), as well as assessing the microbial
community composition of wetland leachates (Walsh et al.,
2002). In addition to the general overview of the microbial
diversity, specific detection of bacteria affiliated to the
genera Dehalobacter, Dehalococcoides or Geobacter degrading
chlorinated ethenes can be used as a more specific indicator
for ongoing potential dechlorinating activity (Loffler et al.,
2000; Hendrickson et al., 2002; Duhamel et al., 2004; Imfeld
et al., 2008b; Nikolausz et al., 2008). However, microbial
community diversity, structures and degradation activities
in wetland systems often vary on both spatial and temporal
scales with ambient geochemical conditions. Therefore,
statistical tools relating microbial community structures to
hydrochemical processes across ecological scales allow to
gain a more consistent insight into the contaminant
attenuation potential of heterogeneous systems (Ramette,
2007).
This study examined the dynamics of the microbial
communities in the aqueous phase of a model system
representing wetland zones located at the interface be-
tween anoxic DCE-contaminated aquifers and oxic surface
water bodies. A previous study revealed the occurrence of
biodegradation activity and major hydrochemical changes
from oxic to strongly reducing conditions in the model
wetland system (Imfeld et al., 2008a). These changes were
associated with a parallel shift in the dominant degrada-
tion mechanisms over time: the sequence was initiated by
DCE degradation under oxic conditions and progressively
changed towards reductive dechlorination under anoxic
conditions. It is hypothesized that changes of the micro-
bial diversity affected the prevailing degradation mechan-
ism in the model wetland, which in turn may influence the
system functioning over time with respect to organic
contaminant removal. Therefore, the first objective of the
present study was to characterize the wetland microbial
community during the transition phase from prevailing
aerobic to anaerobic conditions. The second objective was
to identify the potential contribution of the microbial
community to the biogeochemical processes that influ-
ence DCE transformation in the wetland system, mainly
focusing on reductive dechlorinating bacteria and their
activity.
Materials and methods
System design and sampling procedure
The model wetland was a rectangular chamber
(201� 60� 5 cm), filled with quartz sand and planted with
common rush (Juncus effusus, L.) (Fig. 1). The wetland was
continuously supplied with groundwater from a stainless-
steel tank. The contaminated groundwater was collected
bimonthly from the Bitterfeld/Wolfen site and kept under
anoxic conditions (N2 atmosphere). Regular geochemical
analysis of the hydrogeochemistry revealed a stable hydro-
chemistry of the supplied groundwater over time, with cis-
and trans-1,2-DCE as the dominant contaminants (see
Supporting Information, Appendix S1). Additional quanti-
ties of DCE were spiked into the tanks during groundwater
sampling in order to increase DCE average concentrations to
6.5 and 1.5 mg L�1 for cis- and trans-DCE, respectively. The
system was operated in a flow-through mode with a rate of
1.8 L day�1 contaminated groundwater, corresponding to a
retention time of approximately 15 days (for more details
regarding the system design, refer to Imfeld et al., 2008a).
The system was equipped with a cooling system maintaining
the tank and the wetland temperature constantly at
11� 2 1C.
The biogeochemical development of the wetland was
followed for a period of 430 days, as described in Imfeld
et al. (2008a). However, pore water sampling for the micro-
bial investigations presented in this study started when
indications of the establishment of anaerobic conditions in
the wetland were observed. Samples were retrieved at days
199, 227, 255, 283 and 311. Water samples were collected
from the tank, the inflow chamber, the pond, and at four
vertical profiles across the sand compartment to conduct
hydrogeochemical and microbial analyses in parallel. Verti-
cal profiles in the sand compartment were taken at 6, 49, 94
and 139 cm from the inflow via the three sampling ports at
20, 32 and 44 cm depths (Fig. 1).
For the microbial analyses, pore water samples were
retrieved from the wetland at days 199, 227, 255, 283 and
311 from the wetland using sterile syringes. The samples
collected at each sampling day were then pooled in two
integrative samples. Each of these integrative pore water
samples consisted of six 20-mL subsamples, which were
separately pooled in sterile vials. The first series of six pore
water subsamples was retrieved from the two vertical
profiles at the inflow side of the sand compartment (6 and
49 cm from the inflow). The second series of six subsamples
was retrieved from the two vertical profiles at the pond side
of the sand compartment (94 and 139 cm). This integrative
FEMS Microbiol Ecol 72 (2010) 74–88 Journal compilation c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original German government works
75Microbial communities in a wetland treating dichloroethenes

sampling procedure accounted for the dominant spatial
development of the hydrogeochemical gradients within the
model wetland that occurred along the flow path across the
wetland rather than over depth (see Supporting Informa-
tion, Fig. S1). In parallel, 0.75-L water samples were col-
lected in cleaned and sterile Schott bottles from the tank, the
inflow and the outflow pond for microbial investigations.
The sampled material was immediately cooled to 4 1C to
slow down further transformation processes, and samples
for microbial analyses were filtered within o 3 h after
sampling.
Hydrogeochemical analysis and compound-specific carbon isotope analysis
Quantification of cis-, trans-1,2-dichloroethene, VC and
ethene was performed with a gas chromatograph equipped
with a flame ionization detector (Varian Chrompack CP-
3800, Middelburg, the Netherlands), with detection limits of
50, 30, 5 and 5 mg L�1, respectively, according to Nijenhuis
et al. (2007). Geochemical parameters (pH, redox potential,
Cl�, NH41, PO43�, Fe21, total Fe, O2, SO4
2� and total
sulphide) were determined according to DIN and DIN EN
ISO standards, following the laboratory procedures de-
scribed in Imfeld et al. (2008a). Stable carbon isotope
compositions of DCE were measured using a GC-combus-
tion-isotope ratio MS system (GC-C-IRMS) (see Nijenhuis
et al., 2007; Imfeld et al., 2008b). The carbon isotope ratio
for an individual compound is reported in d-notation (%)
relative to the Vienna Pee Dee Belemnite standard (V-PDB,
IAEA-Vienna) (Coplen et al., 2006). The analytical error
is � 0.5 d unit and incorporates both the accuracy and the
reproducibility on at least three replicate measurements of
the sample.
Molecular analysis
DNA extraction
In order to avoid perturbation of the system, only water
samples have been retrieved during the course of the
experiment. Water samples were filtered through a sterile
0.2 mm membrane (MoBio Water DNA kit, Carlsbad, CA).
The membranes were stored at � 20 1C until extraction.
DNA was extracted by disrupting microorganisms with a
bead beater (Fast Prep System, Qbiogene, Irvine, CA),
applying a FastDNA spin kit for DNA extraction (BIO101,
La Jolla, CA) and elution in 50 mL nuclease-free water.
16S rRNA gene-targeted PCR
PCR was used to amplify part of the 16S rRNA genes from
Bacteria and Archaea. The PCR mix per reaction contained
1� PCR buffer (with 1.5 mM MgCl2) (Qiagen, Hilden,
(a)
50
600
530
480
505
Inflow Outflow
Pond
(b)
590450 450490
1390940
1980
60
(mm)
Fig. 1. Scheme of the model-constructed
wetland (a) and location of the pore water
sampling devices (b). The model horizontal
subsurface flow wetland was filled with
quartz sand (average depth = 54 cm;
kfaverage = 2.27� 0.14�10�4 m s�1;
grain size = 0.40–0.63 mm) and planted
with common rush (Juncus effusus, L.). A
50-cm-long water pond remained in direct
contact with the atmosphere. Groundwater was
continuously pumped from the 50 L tank at a
constant flow rate (0.4 mL min�1). Supplied
groundwater (50 L tank) was maintained under
anaerobic conditions at a constant N2 pressure
(0.5 mbar). (b) Pore water sampling devices (�)
were located at 6, 49, 94 and 139 cm from the
inflow. At these distances, the sampling ports
were mounted at 20, 32 and 44 cm depths from
the surface.
FEMS Microbiol Ecol 72 (2010) 74–88Journal compilation c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original German government works
76 G. Imfeld et al.

Germany), 0.2 mM (each) dNTP (Qiagen), 0.5 mM (each)
forward and reverse primer (Invitrogen), 1.5 U of HotStar-
Taq DNA polymerase (Qiagen), 1 : 10 v : v of DNA template
and molecular-grade water (Promega, Madison, WI). Eu-
bacterial primers 27f (Lane, 1991) and 1378r (Heuer et al.,
1997) were used to amplify almost the complete 16S rRNA
gene using the following amplification program: 95 1C
(15 min), followed by 30 cycles of 95 1C (30 s), 51 1C (30 s)
and 72 1C (50 s), completed with an additional 30 min at
72 1C. The second-round PCR for denaturing gradient gel
electrophoresis (DGGE) analysis used universal primers
GC968f (Nubel et al., 1996) and 1378r. The conditions for
PCR amplifications were as follows: 95 1C (15 min), fol-
lowed by 30 cycles of 95 1C (30 s), 60 1C (1 min), 60–55 1C
(0.5 1C min�1), 72 1C (1 min) and a final extension for
30 min at 72 1C. In parallel, to test the presence of the genera
Dehalobacter (Schlotelburg et al., 2002) and Geobacter
(Duhamel & Edwards, 2006) in the model system, a second
round of PCR with specific primers was performed using the
PCR products from the universal 16S rRNA gene amplifica-
tion as a template (Imfeld et al., 2008b). A Dehalococcoides-
specific amplification protocol according to Imfeld et al.
(2008b) was used for detecting Dehalococcoides-affiliated
bacteria, using specific primers Fp DHC 1 and Rp DHC
1377 for the first round of PCR, and the second PCR was
carried out with a nested Dehalococcoides-specific primer set
(Fp DHC 774 and Rp DHC 1212) (Hendrickson et al.,
2002). The specificity of the Dehalobacter- and Dehalococ-
coides-specific primer sets was previously tested in a pre-
vious study using DNA samples from the same groundwater
that was used for this experiment (Nijenhuis et al., 2007).
PCR for amplification of the 16S rRNA genes of Archaea
used Archaea-specific primers ARC21f and ARC958r ac-
cording to Bano et al. (2004). The second-round PCR for
DGGE used primers GC-ARC344f and UNIV517r under the
conditions described previously (Bano et al., 2004).
Cloning sequencing
Two samples were selected for setting up 16S rRNA gene
libraries. The first sample consisted of groundwater supplied
to the wetland between days 200 and 315. The second
sample consisted of pore water retrieved at day 227 from
the sand compartment at the vertical profiles located at 6
and 49 cm from the inflow. The latter sample was selected in
order to assess the bacterial composition associated with the
developing front of iron sulphide mineral precipitates in the
wetland (Fig. S2). PCR products obtained with primers 27f
and 1378r were ligated into a pGEM-TEasyTM plasmid
(Promega), and were transformed into competent Escher-
ichia coli JM109 cells. Following plasmid extraction and
amplification using vector-specific M13f and M13r primers
(Stratagen, La Jolla, CA), the PCR products of clones were
separately digested with 1 U of the tetrameric site restriction
endonuclease Hin6I and BsuRI for amplified rDNA restric-
tion analysis (Fermentas, St. Leon-Rot, Germany). Clones
with the same pattern with both enzymes were grouped into
an operating taxonomic unit (OTU) (Massol-Deya et al.,
1995). Before sequencing, the 16S rRNA genes of each OTU
were purified using an ABI PRISM BigDye Terminator Cycle
Sequencing Kit V. 3.0 (Applied Biosystems, Foster City, CA)
and sequenced using an ABI PRISM 3100 DNA analyzer
(Applied Biosystems). Putative chimeric sequences (about
10% of the sequences) were discarded from the dataset after
detection analysis performed using the Chimera Detection
Program of the RDP-II (Cole et al., 2009). Examination of
phylogenetic relationships and taxonomic assignments at a
confidence level of 50% was performed using the naıve
Bayesian rRNA Classifier and the Sequence Match tools of
the RDP-II (release 10, http://rdp.cme.msu.edu/), respec-
tively (Wang et al., 2007; Cole et al., 2009). Bacterial
nucleotide sequence data are available in the European
Molecular Biology Laboratory database under accession
numbers FM205929–FM206115.
Molecular fingerprinting analysis ofmicrobial communities
Amplicons obtained from the amplification of bacterial or
archaeal 16S rRNA genes were separated using the DCode
Universal Mutation Detection System (BioRad, Munich,
Germany) DGGE system (for protocol details see Imfeld
et al., 2008b). The DGGE fingerprints were normalized
according to the reference patterns and compared with the
GELCOMPARII software (Applied Maths, Kortrijk, Belgium).
DGGE banding patterns were then converted into a binary
presence–absence matrix subsequently used for further
statistical analysis. Distinct bands of 16S rRNA gene-DGGE
gels for Archaea were excised from the gels and placed in
30-mL sterile nanopure water overnight to elute DNA. Direct
sequencing of the bands was subsequently performed as
described in Cloning sequencing. Archaeal sequence data of
DGGE bands are available under accession numbers
FM211566–FM211590.
Data analysis
Statistical data analyses were performed using the R software
ver. 2.7.2 (R Development Core Team, 2008) with additional
package ‘VEGAN’ (Oksanen et al., 2008) for multivariate
analyses. Principal component analysis (PCA) was applied
to assess the spatio-temporal dynamics of the hydrochemi-
cal variables (pH, Eh, Cl�, NH41, PO43�, Fe21, O2, SO4
2�,
total sulphide, cis- and trans-DCE concentrations and iso-
topic composition). The PCA was based on the Euclidian
correlation matrix of data obtained at days 227, 255, 283 and
FEMS Microbiol Ecol 72 (2010) 74–88 Journal compilation c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original German government works
77Microbial communities in a wetland treating dichloroethenes

311. Correspondingly, a nonmetric multidimensional scal-
ing (nMDS) was used to evaluate the changes in bacterial
community structures from day 227 to 311. Before the
analyses, the binary numerical matrix of DGGE bands was
converted into a Euclidean dissimilarity matrix. This con-
version was chosen as the most appropriate for the data
according to the highest rank-order dissimilarity test with
gradient separation after a comparison of indices (Oksanen
et al., 2008). The relationship between the community
profiles and the hydrogeochemical variables was interpreted
by fitting the environmental vectors a posteriori onto the
nMDS. Hydrochemical vectors were fitted onto the com-
munity matrix by maximizing their correlations with the
ordination configuration. In the final plot, the vectors, each
representing a hydrochemical variable, point towards the
direction that corresponds to the largest change of the
variable value. Hence, hydrochemical variables significantly
associated with the observed changes in the bacterial com-
munity structures can be revealed. The significance of the
fitted vectors was assessed by conducting a Monte-Carlo
permutation test with 1000 permutation steps. Only hydro-
chemical variables with a significance level of Po 0.05 were
considered significant.
Results
Hydrochemistry
Detailed hydrogeochemical data are provided separately for
each sampling date in Tables S1–S8. The main geochemical
changes in the wetland occurring from days 199 to 430 are
characterized by a progressive decrease of dissolved oxygen
concentrations (0–3.63 mg L�1) accompanied by an increase
of dissolved ferrous iron (from 0.01 to 4.18 mg L�1) and
sulphide (from o 5 to 1573 mM) concentrations (Fig. 2).
The concentration values of VC and ethene increased in the
wetland between days 225 and 430 from o 5 to 518� 13
and o 5 to 102� 5 mg L�1, respectively. This indicates the
occurrence of ongoing reductive dechlorination in the wet-
land. The concentrations of VC and ethene increased and
coincided with significant and increasing isotopic enrich-
ment of both trans- and cis-DCE (Dd13C4 0.5%) between
the inflow and the end of the sand compartment ongoing
from day 225 (Fig. 2c and d). This suggests that DCE
oxidation, associated with a lower isotope fractionation,
was progressively replaced over time by reductive dechlori-
nation, associated with larger isotope fractionation (Imfeld
et al., 2008a). Thus, the progressive development of anoxic
conditions in the wetland reflected a shift of the prevailing
DCE degradation process.
PCA of hydrogeochemical variables (pH, Eh, Cl�, NH41,
PO43�, Fe21, O2, SO4
2�, total sulphide, cis- and trans-DCE
concentrations and isotopic composition) underscored the
major hydrogeochemical trends in the wetland from days
227 to 311 (Fig. 3). This period corresponds to the period of
microbial community structure characterization. Hydroche-
mical profiles of the tank and the inflow clustered together
over time (Fig. 3a). This emphasizes that the quality of the
supplied water did not significantly change over the investi-
gation period. In contrast, hydrochemical profiles from the
0
2
4
6
8
10
12
0
500
1000
1500
2000
0
5
10
15
20
25
30
35
199 227 255 283 311 340 396 430 199 227 255 283 311 340 396 430
Δδ13
C (
‰)
Δδ13
C (
‰)
Fe(
II) (
mg
L–1)
Sul
phid
e (μ
M)
Time (day) Time (day)
(a)
(b)
(c)
(d)
Inflow 6 and 49 cm 94 and 139 cm Outflow
0
2
4
6
8
10
199 227 255 283 311 340 396 430199 227 255 283 311 340 396 430
Fig. 2. Iron(II) (a) and sulphide (b) concentrations
and the mean carbon isotopic signature of
trans- (c) and cis- (d) 1,2-dichloroethenes at the
inflow, the sand compartment (vertical profiles at
6, 49 and 94, 139 cm from the inflow) and the
outflow in the model wetland between days 199
and 430. Error bars for iron(II) and sulphide mean
concentration values indicate the SDs of the six
concentration values retrieved at the sampling
ports from the sand compartment (at 6, 49 and
94, 139 cm from the inflow, respectively). The
dashed lines indicate the time period of microbial
community structure analysis.
FEMS Microbiol Ecol 72 (2010) 74–88Journal compilation c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original German government works
78 G. Imfeld et al.
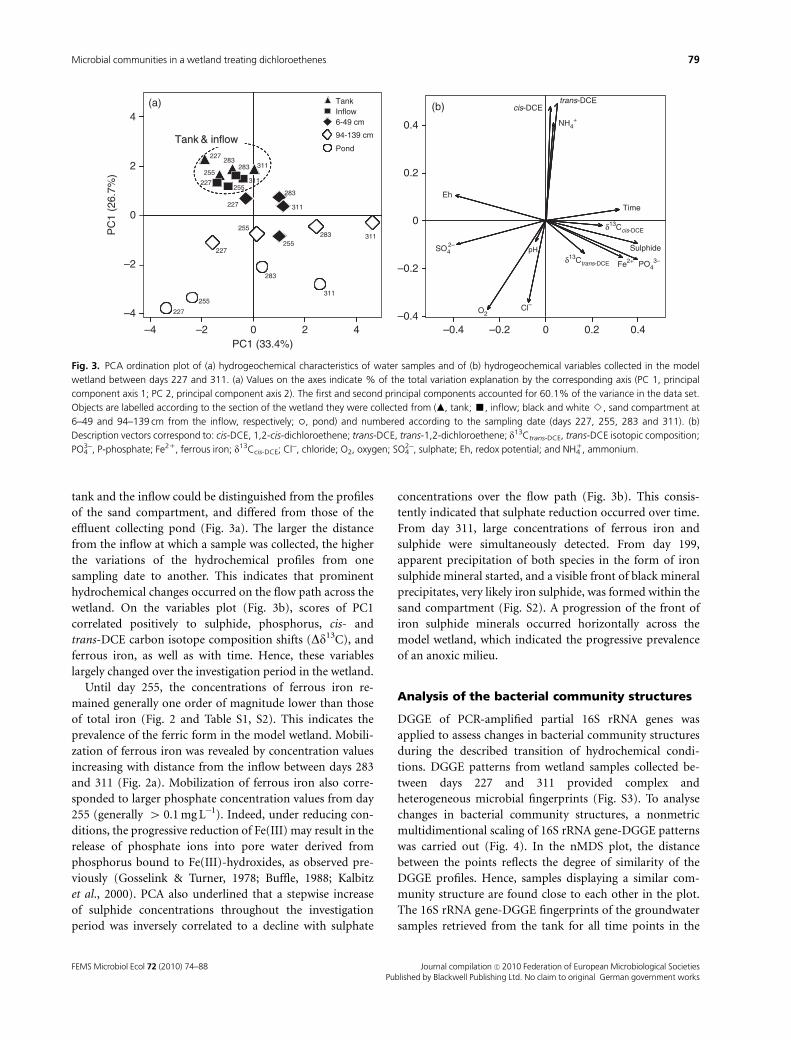
tank and the inflow could be distinguished from the profiles
of the sand compartment, and differed from those of the
effluent collecting pond (Fig. 3a). The larger the distance
from the inflow at which a sample was collected, the higher
the variations of the hydrochemical profiles from one
sampling date to another. This indicates that prominent
hydrochemical changes occurred on the flow path across the
wetland. On the variables plot (Fig. 3b), scores of PC1
correlated positively to sulphide, phosphorus, cis- and
trans-DCE carbon isotope composition shifts (Dd13C), and
ferrous iron, as well as with time. Hence, these variables
largely changed over the investigation period in the wetland.
Until day 255, the concentrations of ferrous iron re-
mained generally one order of magnitude lower than those
of total iron (Fig. 2 and Table S1, S2). This indicates the
prevalence of the ferric form in the model wetland. Mobili-
zation of ferrous iron was revealed by concentration values
increasing with distance from the inflow between days 283
and 311 (Fig. 2a). Mobilization of ferrous iron also corre-
sponded to larger phosphate concentration values from day
255 (generally 4 0.1 mg L�1). Indeed, under reducing con-
ditions, the progressive reduction of Fe(III) may result in the
release of phosphate ions into pore water derived from
phosphorus bound to Fe(III)-hydroxides, as observed pre-
viously (Gosselink & Turner, 1978; Buffle, 1988; Kalbitz
et al., 2000). PCA also underlined that a stepwise increase
of sulphide concentrations throughout the investigation
period was inversely correlated to a decline with sulphate
concentrations over the flow path (Fig. 3b). This consis-
tently indicated that sulphate reduction occurred over time.
From day 311, large concentrations of ferrous iron and
sulphide were simultaneously detected. From day 199,
apparent precipitation of both species in the form of iron
sulphide mineral started, and a visible front of black mineral
precipitates, very likely iron sulphide, was formed within the
sand compartment (Fig. S2). A progression of the front of
iron sulphide minerals occurred horizontally across the
model wetland, which indicated the progressive prevalence
of an anoxic milieu.
Analysis of the bacterial community structures
DGGE of PCR-amplified partial 16S rRNA genes was
applied to assess changes in bacterial community structures
during the described transition of hydrochemical condi-
tions. DGGE patterns from wetland samples collected be-
tween days 227 and 311 provided complex and
heterogeneous microbial fingerprints (Fig. S3). To analyse
changes in bacterial community structures, a nonmetric
multidimentional scaling of 16S rRNA gene-DGGE patterns
was carried out (Fig. 4). In the nMDS plot, the distance
between the points reflects the degree of similarity of the
DGGE profiles. Hence, samples displaying a similar com-
munity structure are found close to each other in the plot.
The 16S rRNA gene-DGGE fingerprints of the groundwater
samples retrieved from the tank for all time points in the
(a) (b)
–4 –2 0 2 4
–4
–2
0
2
4
PC1 (33.4%)
PC
1 (2
6.7%
)
–0.4 –0.2 0 0.2 0.4–0.4
–0.2
0
0.2
0.4
Fig. 3. PCA ordination plot of (a) hydrogeochemical characteristics of water samples and of (b) hydrogeochemical variables collected in the model
wetland between days 227 and 311. (a) Values on the axes indicate % of the total variation explanation by the corresponding axis (PC 1, principal
component axis 1; PC 2, principal component axis 2). The first and second principal components accounted for 60.1% of the variance in the data set.
Objects are labelled according to the section of the wetland they were collected from (m, tank; ’, inflow; black and white B, sand compartment at
6–49 and 94–139 cm from the inflow, respectively; �, pond) and numbered according to the sampling date (days 227, 255, 283 and 311). (b)
Description vectors correspond to: cis-DCE, 1,2-cis-dichloroethene; trans-DCE, trans-1,2-dichloroethene; d13Ctrans-DCE, trans-DCE isotopic composition;
PO43�, P-phosphate; Fe21, ferrous iron; d13Ccis-DCE; Cl�, chloride; O2, oxygen; SO4
2�, sulphate; Eh, redox potential; and NH41, ammonium.
FEMS Microbiol Ecol 72 (2010) 74–88 Journal compilation c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original German government works
79Microbial communities in a wetland treating dichloroethenes

observed period were found to be close together. This
emphasizes the similarity of community structures in the
supplied groundwater throughout the investigation period.
However, the analysis revealed that the bacterial community
structure changed over time at the inflow, the sand com-
partment and the pond. The sand compartment and pond
samples from day 227 to 255 were separated from samples
from days 283 to 311 (Fig. 4). This suggests that relatively
large changes in the bacterial community structure occurred
during the first investigation period (days 227–253),
whereas later only smaller changes were observed.
The relationship between the bacterial community dy-
namics and the observed hydrochemical development in the
wetland was inferred statistically. The resultant characteris-
tic variables such as time, phosphate and sulphide vectors
positively correlated with changes in the microbial commu-
nity structures, whereas the sulphate vector correlated
negatively (Po 0.05) (Fig. 4). Thus, the observed changes
in the microbial community structures over time statistically
correlated with sulphate reduction in the wetland. The
initial changes in the bacterial community structures corre-
sponded to the mobilization of both ferrous iron and
sulphide within the system, and thus reflected major varia-
tions of the prevailing redox conditions.
Detection of potential reductive dechlorinatingbacteria
An indication of the presence of putative reductive dechlori-
nating bacteria in the wetland was provided by taxon-
specific assays. The presence of Dehalobacter, Dehalococcoides
and Geobacter DNA at the inflow, the sand compartment
and the pond of the model wetland was tested from day 199
to 430 (Table 1). Geobacter spp. DNA was detected in all
samples retrieved from the model system throughout the
investigation period. The genus Geobacter sp. contains
metabolically versatile iron-reducers, including members
capable of dechlorination of chlorinated ethenes during
iron reduction (Sung et al., 2006). Dehalobacter spp. was
observed from day 311 and coincided with the presence of
Dehalococcoides spp. DNA. DNA of all the targeted genera
could be detected across the system at day 430, suggesting
the existence of a consortium of reductive dechlorinating
bacteria. The presence of Dehalococcoides-like bacteria was
detected from day 253 in the sand compartment; this
presence correlated with both the detection of VC and
ethene as well as a larger shift in the carbon isotope
composition of both cis- and trans-DCE (Table 1; Fig. 2c
and d).
Sequence analysis of the clone libraries
Two 16S rRNA gene clone libraries were constructed to gain
an insight into the composition of bacterial communities of
the supplied groundwater and the model wetland at day 227
(Table 2). Details on the phylogenetic affiliation and se-
quence similarities of retrieved 16S rRNA gene clones are
provided in Fig. 5. A total of 138 and 165 clones were
obtained and restriction pattern analysis further refined
these sequences, resulting in, respectively, 89 and 97 OTUs
for the groundwater and wetland samples. Because the
ΣS2–
–15 –10 –5 0 5 10 15
–10
–5
0
5
10
–15
NMDS 1
NM
DS
2TimeTime
ΣS2–
SO42–SO42–
PO34–PO34–
311
255
283
311
283
255
227
311
283
255
227
255
283
311
255
311
283
:6-49 cm :Pond:Tank
Sand compartment
:Inflow :94-139 cm
Fig. 4. nMDS plot (two dimensional) of the 16S rDNA gene DGGE
patterns of the bacterial community from water samples, showing the
community changes (stress value: 18.5%). Objects are labelled according
to location (m, tank; ’, inflow; black and white B, sand compartment
at 6–49 and 94–139 cm from the inflow, respectively; �, pond) and
numbered according to the sampling date (day 227, 255, 283 and 311).
Vector arrows of maximum correlation with the corresponding DGGE
patterns are superimposed depicting the direction, magnitude and
correlation of four most important characterizing components (SO42�,
PO34�, time and SS2�). The significance of fitted vectors is assessed using
permutation of all assessed variables a posteriori by permutation of
variables at Po 0.05.
Table 1. Detection of Dehalococcoides, Dehalobacter and Geobacter
DNA in the model wetland using taxon-specific assays
Taxon-specific assay
Time (days)
225 253 280 309 430
Geobacter spp. a, b, c, d a, b, c, d a, b, c, d a, b, c, d a, b, c, d
Dehalobacter spp. – – – a, b a, b, c, d
Dehalococcoides spp. – a, b a a, b, c a, b, c, d
� , absence of the targeted genera. Letters correspond to the part of the
wetland at which samples were retrieved: a, inflow; b, sand compart-
ment at 6 and 49 cm from the inflow; c, sand compartment at 94 and
139 cm from the inflow; d, pond.
FEMS Microbiol Ecol 72 (2010) 74–88Journal compilation c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original German government works
80 G. Imfeld et al.

rarefaction curves did not indicate a clear tendency towards
saturation, the discovery of additional sequences is expected
by increasing the number of investigated clones (Fig. S4). It
is therefore very likely that the current sampling strategy did
not allow detection of minor populations of the bacterial
communities, including those potentially involved in con-
taminant degradation. Consequently, accurate quantitative
estimates of the bacterial groups present in these samples
could not be provided in this study, and relative clone
frequencies mainly apply to dominant populations of the
communities. Clone sequences from the groundwater and
the wetland could be affiliated with, respectively, eight and
11 classes of the domain Bacteria (Table 2). Overall, riboso-
mal sequences affiliated to the Proteobacteria largely domi-
nated, and accounted for about half of both clone libraries.
About 25% of the clones could not be assigned to any
bacterial phylum at the defined confidence level of 50%.
Sequences affiliated to the Beta- and Deltaproteobacteria
were retrieved in higher numbers than the other groups in
both libraries. The majority of sequences within the b-
subgroup in the groundwater were similar to sequences
affiliated to the neutrophilic and chemolithotrophic iron-
oxidizing Gallionella ferruginea, in agreement with the
hydrogeochemistry of the supplied groundwater. Geobacter
sp. accounted for 7% of the wetland clone library. Sequences
affiliated to Geobacter sp. had a close relationship with
sequences retrieved from Fe(III)-reducing enrichment cul-
tures obtained from contaminated sediments (Holmes et al.,
2004; Scala et al., 2006; Kittelmann & Friedrich, 2008).
Sequences affiliated to Verrumicrobia, Chloroflexi and Spiro-
chaetes were exclusively detected in the constructed wetland.
Within the phylum Chloroflexi, one clone sequence was
affiliated to the Levilinea genus. The presence of non-
Dehalococcoides Chloroflexi populations also raises the even-
tuality of their involvement in reductive dechlorination
(Watts et al., 2005). Recently, the existence of putative
reductive dechlorinating phylotypes within Chloroflexi that
includes uncultured microorganisms was revealed (Fager-
vold et al., 2005). While Dehalococcoides is so far the only
genus containing only members dependent on reductive
dehalogenation as an energy-gaining process, the involve-
ment of Chloroflexi members in the degradation process
cannot be excluded. In contrast, no Dehaloccocoides-
affiliated sequences could be retrieved, which underscores
the higher sensitivities of the taxon-specific PCR assay
(Hendrickson et al., 2002) over the cloning-sequencing
procedure. Lack of Dehaloccoides-like sequences in the clone
library, despite the obvious reductive dehalogenation activ-
ity in the wetland, could be due to the relatively high
degradation capacity of Dehalococcoides compared with the
slow-growing nature of these organisms (with a doubling
time of approximately 1 day) (He et al., 2005; Tang et al.,
2009). An additional explanation is that the Dehalococcoides
strains described so far have only one copy of the ribosomal
genes, while the predominant Proteobacteria have many
more (up to 13, 4.17 in average), leading to lower sensitivity
for their detection (Lee et al., 2009). Noticeably, sequences
related to the TM7 candidate division accounted for about
9% of the groundwater clone library, but could not be
retrieved from the wetland. Some members of this division
were suggested to be associated with TCE and cis-DCE
cometabolic oxidation (Lowe et al., 2002; Connon et al.,
2005).
To gain insights into Archaea diversity, the community
composition of the Archaea was assessed at day 255, when
methane production in the wetland was observed (Imfeld
et al., 2008a). This assessment also aimed at identifying
methanogen candidates in the wetland. High archaeal
diversity (15–19 bands) with variation between the segments
of the wetland was observed, suggesting changes in the micro-
bial community composition over the flow path (Fig. S5).
Bands with a high intensity within the DGGE pattern were
excised from the gel to identify predominant Archaea. The
detailed outcome of sequence analysis of selected bands is
Table 2. Relative clone frequencies in major phylogenetic groups of the
clone libraries from the original groundwater (supplied to the wetland
between days 200 and 310) and the wetland pore water (sampling day 227)
Group
Relative frequency (%)
Groundwater Wetland
Proteobacteria 57 34
Alphaproteobacteria 2 7
Magnetospirillum spp. 0 2
Betaproteobacteria 35 16
Gallionella spp. 17 1
Deltaproteobacteria 10 7
Geobacter spp. 0 7
Desulfovibrio spp. 1 15
Epsilonproteobacteria 2 2
Gammaproteobacteria 7 2
Dokdonella 0 1
Thiothrix spp. 1 1
Unclassified Proteobacteria 1 0
Firmicute
‘Clostridia’ Clostridiales 2 4
TM7 Division 9 0
Bacteroidetes 2 4
Lentisphaerae 1 5
OD1 division 3 2
Actinobacteria 1 2
Verrucomicrobia 0 2
Cloroflexi 1 2
Acidobacteria 1 1
Spirochaetes 0 1
Unclassified Bacteria 24 18
Number of clones 138 165
Number of OTUs 89 97
FEMS Microbiol Ecol 72 (2010) 74–88 Journal compilation c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original German government works
81Microbial communities in a wetland treating dichloroethenes

(a) (b)
Candidatedivision OD1
Fig. 5. Neighbour-joining tree of 16S rRNA gene sequences depicting the relationships among community members of the (a) groundwater and (b)
constructed wetland as revealed by comparative analysis of 16S rRNA gene sequences and those stored in the ARB database and GenBank. 16S rRNA
gene bacterial sequences determined in this study are highlighted in grey and were deposited in GenBank under accession numbers
FM205929–FM206115. Bootstrap values of 4 50% are indicated at the node of the branch. Scale bar represents a 10% estimated change.
FEMS Microbiol Ecol 72 (2010) 74–88Journal compilation c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original German government works
82 G. Imfeld et al.

provided in Fig. 6. Comparative analyses of sequences
retrieved from GenBank revealed that all 11 examined
sequences proved to belong to Euryarchaeota, including
eight sequence types affiliated to hydrogenotrophic metha-
nogenic genera (Methanomicrobiales, Methanococcales and
Methanopyrales). Two sequences were related to the Archaeo-
globi class and one sequence was identified to be a member
of the Halobacteria class.
Discussion
In this study, we assessed the biogeochemical development
of a wetland treating DCE-contaminated groundwater over
time, with a particular emphasis on the microbial character-
ization. Microbial and hydrochemical investigations were
combined to characterize in parallel the wetland microbial
community and ambient hydrochemistry in the wetland.
Microbial analyses performed in this study were exclusively
based on liquid-phase samples, allowing a temporal and
spatial assessment of the microbial community with mini-
mal disturbance of the system during sampling. Owing to
the dimensions of the wetland systems, parallel sediment
collections would have resulted in a significant disturbance
of ongoing biogeochemical changes over space and time.
However, this liquid sampling design very likely results in an
incomplete view of the wetland microbiology as the liquid
phase is generally less heterogeneously distributed than the
solid material, and specific populations or activities may be
asymmetrically distributed between the pore water and the
sediment phases (Lehman & O’Connell, 2002; Lehman,
2007). Nevertheless, changes of the hydrochemistry in the
wetland could be identified and corresponded to a temporal
shift in the prevailing pathway of DCE degradation, from
degradation reactions under oxic conditions, to reductive
dechlorination under anoxic conditions. These changes
reflected variations of both the bacterial community struc-
tures and the composition of the guild of putative dechlori-
nating bacteria.
Microbial respiration coupled to degradation of organic
carbon and hydrogen consumption by various groups of
reducing bacteria is assumed to be a dominant degradation
mechanism in various anaerobic soils and sediments (Hol-
mer & Storkholm, 2001). The high availability of electron
donors due to decaying plant material was evidenced by
high DOC concentrations in the wetland (Imfeld et al.,
2008a), which sustained the coexistence of iron and sulphate
reduction, in agreement with previous observations in
freshwater wetlands (Alewell et al., 2004). In the wetland
Fig. 6. Neighbour-joining tree of 16S rRNA gene sequences depicting the phylogenic relationship of 16S rRNA gene archaeal sequences from excised
DGGE bands to other related organisms. 16S rRNA gene archaeal sequences determined in this study are highlighted in grey and are designated
according to their origin in the wetland (tank, inflow, sand compartment or outflow) (GenBank accession numbers in parentheses). Bootstrap values of
4 50% are indicated at the node of the branch. Scale bar represents 5% estimated change.
FEMS Microbiol Ecol 72 (2010) 74–88 Journal compilation c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original German government works
83Microbial communities in a wetland treating dichloroethenes

system, the spatial and temporal development of zones of
iron sulphide precipitation was likely sustained by (1) the
maintenance of reducing conditions created by continuous
supply of anoxic groundwater, (2) the constant supply of
nonreduced precursors [Fe(III) and sulphate] in the sup-
plied groundwater and (3) the high availability of electron
donors from decaying plant organic matter within the
wetland. These spatial and temporal changes in the wetland
hydrochemical properties likely shaped the ecological habi-
tats within the porous sand compartment. For instance, the
accumulation of decaying plant organic material and the
visible deposits of iron sulphide mineral in the wetland
likely resulted in the reduction of hydraulic conductivity and
void space of the porous medium, oxygen supply and
increased dispersion, as described previously (Tanner &
Sukias, 1995; Garcia et al., 2004). Changes in both the
hydrochemistry and the hydrodynamic properties of the
sand compartment can in turn affect both the structures and
the diversity of the microbial community by changing fluxes
and transport patterns of organic substrates and nutrients.
Although substantial variations in the bacterial community
structures were observed throughout the investigation per-
iod, the contaminant biodegradation capacity was main-
tained. Nevertheless, changes in the microbial community
structures coincided with a shift in the DCE carbon isotope
composition, which indicated changes in the DCE degrada-
tion pathway.
Although changes in the microbial community structures
coincided with a shift in the DCE degradation pathway as
suggested by compound-specific isotope analysis, degrading
bacterial populations were also expected to change. Detec-
tion of dechlorinating bacteria provided an additional line
of evidence that reductive dechlorination represented a key
mechanism for contaminant removal in the wetland. As
suggested by the isotopic composition analysis, reductive
dechlorinating activity likely has become more important
over time, until it completely overcame oxidative degrada-
tion. Decreasing availability of electron acceptors for aerobic
(i.e. O2) or anaerobic oxidation (i.e. Fe31) may have
progressively limited oxidative degradation of DCE or VC
during the course of the investigation period (Bradley &
Chapelle, 1998). Although no quantitative approach was
used in the study, Geobacter sp. likely dominated at the
beginning of the transition phases, when iron reduction
occurred. However, the involvement of this genus in reduc-
tive dechlorination in the wetland cannot be proven with the
presented data. The stepwise detection of members of the
reductive dechlorinating guild over time also reflected
hydrogeochemical changes in the wetland. Under anoxic
conditions, favourable conditions for microbial reductive
dechlorination of DCE were progressively achieved within
the wetland. Hence, the detection of ethene along with the
simultaneous presence of putative reductive dechlorinating
bacteria under strongly reducing conditions emphasized
that reductive dechlorination of DCE prevails at the end of
the investigation period. Although cultivated members of
the genera Geobacter and Dehalobacter are capable of partial
dechlorination of PCE and TCE to cis-DCE, Dehalococcoides
is the only group known so far to be capable of complete
dechlorination of PCE/TCE to ethene (Maymo-Gatell et al.,
1997; Seshadri et al., 2005). In the case of a hypothetical
change of the prevailing groundwater contaminants, the
simultaneous presence of these reductive dechlorinating
bacteria also suggests that the wetland system could sustain
reductive dechlorination of PCE or TCE up to ethene.
The 16S rRNA gene clone library indicated that the model
wetland sustained diverse microbial populations capable of
various terminal electron-accepting processes. A large pro-
portion of recovered microorganisms were putatively driv-
ing iron and sulphate reduction, in agreement with the
hydrochemical observations. Interactions among the var-
ious populations of the wetland system can result in changes
of the hydrochemical conditions and may affect members of
the reductive dechlorinating guild. This in turn can influ-
ence the wetland DCE-degrading capacity. For instance,
sulphate-reducing and reductive dechlorinating micro-
organisms are able to thrive at similar H2 levels (Heimann
et al., 2005). Sulphate-reducing bacteria were relevant in the
wetland when the transition from an oxic to a prevailing
anoxic regime started, and possibly compete for an electron
donor or its fermentation products, thus determining the
rate and extent of DCE dechlorination (Aulenta et al., 2002;
Heimann et al., 2007). However, reductive dechlorinating
activity was observed concurrently with sulphate reduction,
and even increased progressively throughout the investiga-
tion period. This suggests that dechlorinators could com-
pete for electron usage, in agreement with previous
observations (Aulenta et al., 2007). Although considered as
ubiquitous in anaerobic systems, cometabolic dechlorinat-
ing processes (presumably fortuitous) are generally incap-
able of mediating complete reduction to ethene (McCarty &
Semperini, 1994). Nevertheless, if the present reductive
dechlorinating bacteria do not display enzymatic capabil-
ities to use iron or sulphate, competitive exclusion may
influence organisms involved in the cometabolic dechlor-
ination of cis-DCE. These include methanogens, present in
the wetland, and acetogens, both of which are highly
influenced by H2 consumption, as well as Archaea. Although
no Archaea have been identified in dechlorination processes,
their direct or indirect role in dechorination is not well
known. Examples of the syntrophic association of dechlor-
inating bacteria within a methanogenic microbial consor-
tium have been reported. Species also capable of
cometabolic dechlorination include Methanosarcina thermo-
phila, Methanosarcina mazei, which can dechlorinate PCE to
TCE, and Methanobacterium thermoautotrophicum and
FEMS Microbiol Ecol 72 (2010) 74–88Journal compilation c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original German government works
84 G. Imfeld et al.

Methanothrix soehngenii, both capable of reductively
dechlorinating cis-DCE to chloroethane (Fantroussi et al.,
1998; Holliger et al., 2003).
Though the DCE degradation mechanism varied over
time, the biodegrading function of the model wetland was
maintained. Although coupling reduction and oxidation
processes in wetland systems to reach an efficient transfor-
mation of some chlorinated hydrocarbons and their trans-
formation products may be of interest, possible changes in
the degradation mechanism over the lifespan of a wetland
system need to be carefully considered. Further studies
integrating hydrochemical and microbial approaches are
required to understand how biodegradation mechanisms
and processes influence the transformation of chlorinated
hydrocarbons over the lifespan of wetland systems.
Acknowledgements
G.I. and E.M. were supported by a European Union Marie
Curie Early Stage Training Fellowship (AXIOM, contract no.
MEST-CT-2004-8332). C.E.A. was supported by a Leonardo
Da Vinci exchange grant. We thank the Analytical Chemistry
Department of the UFZ for measurements of the anorganic
parameters and anonymous reviewers for critical review and
helpful comments. This work was supported by the Helm-
holtz Centre for Environmental Research – UFZ in the scope
of the SAFIRA II Research Programme (Revitalization of
Contaminated Land and Groundwater at Megasites, project
‘Compartment Transfer’).
References
Alewell C, Lischeid G, Hell U & Manderscheid B (2004) High
temporal resolution of ion fluxes in semi-natural ecosystems –
gain of information or waste of resources? Biogeochemistry 69:
19–35.
Amon JP, Agrawal A, Shelley ML et al. (2007) Development of a
wetland constructed for the treatment of groundwater
contaminated by chlorinated ethenes. Ecol Eng 30: 51–66.
Armenante PM, Kafkewitz D, Lewandowski G & Kung CM
(1992) Integrated anaerobic–aerobic process for the
biodegradation of chlorinated aromatic-compounds. Environ
Prog 11: 113–122.
Aulenta F, Majone M, Verbo P & Tandoi V (2002) Complete
dechlorination of tetrachloroethene to ethene in presence of
methanogenesis and acetogenesis by an anaerobic sediment
microcosm. Biodegradation 13: 411–424.
Aulenta F, Pera A, Rossetti S, Papini MP & Majone M (2007)
Relevance of side reactions in anaerobic reductive
dechlorination microcosms amended with different electron
donors. Water Res 41: 27–38.
Bano N, Ruffin S, Ransom B & Hollibaugh JT (2004)
Phylogenetic composition of Arctic Ocean archaeal
assemblages and comparison with antarctic assemblages.
Appl Environ Microb 70: 781–789.
Bradley PM & Chapelle FH (1998) Microbial mineralization of
VC and DCE under different terminal electron accepting
conditions. Anaerobe 4: 81–87.
Bradley PM & Chapelle FH (2000) Aerobic microbial
mineralization of dichloroethene as sole carbon substrate.
Environ Sci Technol 34: 221–223.
Braeckevelt M, Rokadia H, Imfeld G et al. (2007) Assessment of
in situ biodegradation of monochlorobenzene in
contaminated groundwater treated in a constructed wetland.
Environmental Pollut 148: 428–437.
Buffle J (1988) Complexation properties of homologous
comlexants and choice of measuring methods. Complexation
Reactions in Aquatic Systems: An Analytical Approach
(Chalmers RA & Masson M, eds), pp. 304–383. Ellis Horwood
Series in Analytical Chemistry, Chichester, UK.
Cole JR, Wang Q, Cardenas E et al. (2009) The ribosomal
database project: improved alignments and new tools for
rRNA analysis. Nucleic Acids Res 37: D141–D145 DOI:
10.1093/nar/gkn879.
Coleman NV, Mattes TE, Gossett JM & Spain JC (2002)
Biodegradation of cis-dichloroethene as the sole carbon source
by a beta-proteobacterium. Appl Environ Microb 68: 2726–2730.
Connon SA, Tovanabootr A, Dolan M, Vergin K, Giovannoni SJ
& Semprini L (2005) Bacterial community composition
determined by culture-independent and -dependent methods
during propane-stimulated bioremediation in
trichloroethene-contaminated groundwater. Environ Microbiol
7: 165–178.
Coplen TB, Brand WA, Gehre M, Groning M, Meijer HAJ, Toman
B & Verkouteren RM (2006) New guidelines for delta C-13
measurements. Anal Chem 78: 2439–2441.
Duhamel M & Edwards EA (2006) Microbial composition of
chlorinated ethene-degrading cultures dominated by
Dehalococcoides. FEMS Microbiol Ecol 58: 538–549.
Duhamel M, Mo K & Edwards EA (2004) Characterization of a
highly enriched Dehalococcoides-containing culture that grows
on vinyl chloride and trichloroethene. Appl Environ Microb 70:
5538–5545.
Fagervold SK, Watts JEM, May HD & Sowers KR (2005)
Sequential reductive dechlorination of meta-chlorinated
polychlorinated biphenyl congeners in sediment microcosms
by two different Chloroflexi phylotypes. Appl Environ Microb
71: 8085–8090.
Fantroussi SE, Naveau H & Agathos SN (1998) Anaerobic
dechlorinating bacteria. Biotechnol Progr 14: 167–188.
Garcia J, Chiva J, Aguirre P, Alvarez E, Sierra JP & Mujeriego R
(2004) Hydraulic behaviour of horizontal subsurface flow
constructed wetlands with different aspect ratio and granular
medium size. Ecol Eng 23: 177–187.
Gosselink JG & Turner RE (1978) The role of hydrology in
freshwater wetland ecosystems. Freshwater Wetlands, Ecological
Processes and Management Potential (Good RE, Whigham DF
& Simpson RL, eds), pp. 63–78. Academic Press, New York.
FEMS Microbiol Ecol 72 (2010) 74–88 Journal compilation c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original German government works
85Microbial communities in a wetland treating dichloroethenes

Hata J, Miyata N, Kim ES, Takamizawa K & Iwahori K (2004)
Anaerobic degradation of cis-1,2-dichloroethylene and vinyl
chloride by Clostridium sp strain DC1 isolated from landfill
leachate sediment. J Biosci Bioeng 97: 196–201.
He J, Sung Y, Krajmalnik-Brown R, Ritalahti KM & Loffler FE
(2005) Isolation and characterization of Dehalococcoides sp.
strain FL2, a trichloroethene (TCE)- and 1,2-dichloroethene-
respiring anaerobe. Environ Microbiol 7: 1442–1450.
Heimann AC, Friis AK & Jakobsen R (2005) Effects of sulfate on
anaerobic chloroethene degradation by an enriched culture
under transient and steady-state hydrogen supply. Water Res
39: 3579–3586.
Heimann AC, Friis AK, Scheutz C & Jakobsen R (2007) Dynamics
of reductive TCE dechlorination in two distinct H-2 supply
scenarios and at various temperatures. Biodegradation 18:
167–179.
Hendrickson E, Payne J, Young R et al. (2002) Molecular analysis
of Dehalococcoides 16S ribosomal DNA from chloroethene-
contaminated sites throughout North America and Europe.
Appl Environ Microb 68: 485–495.
Heuer H, Krsek M, Baker P, Smalla K & Wellington EMH (1997)
Analysis of actinomycete communities by specific
amplification of genes encoding 16S rRNA and gel-
electrophoretic separation in denaturing gradients. Appl
Environ Microb 63: 3233–3241.
Holliger C, Regeard C & Dietert G (2003) Dehalogenation by
anaerobic bacteria. Dehalogenation: Microbial Processes and
Environmental Applications (Haggblom MM & Bossert ID,
eds), pp. 115–157. Kluwer Academic Publishers, Boston, MA.
Holmer M & Storkholm P (2001) Sulphate reduction and
sulphur cycling in lake sediments: a review. Freshwater Biol 46:
431–451.
Holmes DE, Nevin KP & Lovley DR (2004) In situ expression of
nifD in Geobacteraceae in subsurface sediments. Appl Environ
Microb 70: 7251–7259.
Ibekwe AM, Lyon SR, Leddy M & Jacobson-Meyers M (2007)
Impact of plant density and microbial composition on water
quality from a free water surface constructed wetland. J Appl
Microbiol 102: 921–936.
Imfeld G, Estop C, Zeiger S et al. (2008a) Tracking in situ
biodegradation of 1,2-dichloroethenes in a model wetland.
Environ Sci Technol 42: 7924–7930.
Imfeld G, Nijenhuis I, Nikolausz M et al. (2008b) Assessment of
in situ degradation of chlorinated ethenes and bacterial
community structure in a complex contaminated groundwater
system. Water Res 42: 871–882.
Kalbitz K, Solinger S, Park JH, Michalzik B & Matzner E (2000)
Controls on the dynamics of dissolved organic matter in soils:
a review. Soil Sci 165: 277–304.
Kassenga GR, Pardue JH, Blair S & Ferraro T (2003) Treatment of
chlorinated volatile organic compounds in upflow wetland
mesocosms. Ecol Eng 19: 305–323.
Keefe SH, Barber LB, Runkel RL & Ryan JN (2004) Fate of volatile
organic compounds in constructed wastewater treatment
wetlands. Environ Sci Technol 38: 2209–2216.
Kim Y & Semprini L (2005) Cometabolic transformation of cis-
1,2-dichloroethyleneand cis-1,2-dichloroethylene epoxide by a
butane-grown mixed culture. Water Sci Technol 52: 125–131.
Kittelmann S & Friedrich MW (2008) Identification of novel
perchloroethene-respiring microorganisms in anoxic river
sediment by RNA-based stable isotope probing. Environ
Microbiol 10: 31–46.
Lane DJ (1991) 16S/23S rRNA sequencing. Nucleic Acid
Techniques in Bacterial Systematics (Stackebrandt E &
Goodfellow M, eds), pp. 115–148. Wiley, New York.
Lee ZMP, Bussema C & Schmidt TM (2009) rrnDB: documenting
the number of rRNA and tRNA genes in bacteria and archaea.
Nucleic Acids Res 37: D489–D493.
Lehman RM (2007) Understanding of aquifer microbiology is
tightly linked to sampling approaches. Geomicrobiol J 24:
331–341.
Lehman RM & O’Connell SP (2002) Comparison of extracellular
enzyme activities and community composition of attached
and free-living bacteria in porous medium columns. Appl
Environ Microb 68: 1569–1575.
Loffler F, Sun Q, Li J & Tiedje J (2000) 16S rRNA gene-based
detection of tetrachloroethene-dechlorinating Desulfuromonas
and Dehalococcoides species. Appl Environ Microb 66:
1369–1374.
Lorah MM & Olsen LD (1999) Degradation of 1,1,2,2-
tetrachloroethane in a freshwater tidal wetland: field and
laboratory evidence. Environ Sci Technol 33: 227–234.
Lowe M, Madsen EL, Schindler K, Smith C, Emrich S, Robb F &
Halden RU (2002) Geochemistry and microbial diversity of a
trichloroethene-contaminated Superfund site undergoing
intrinsic in situ reductive dechlorination. FEMS Microbiol Ecol
40: 123–134.
Massol-Deya AA, Odelson DA, Hickey RF & Tiedje JM (1995)
Bacterial community fingerprinting of amplified 16S and
16–23S ribosomal DNA gene sequences and restriction
endonuclease analysis (ARDRA). Molecular Microbial Ecology
Manual, Vol. 3.3.2 (Akkermans ADL, van Elsas JD & de Ruijn
FJ, eds), pp. 1–8. Kluwer Academic Publishers, Dordrecht, The
Netherlands.
Master ER, Lai VWM, Kuipers B, Cullen WR & Mohn WW
(2002) Sequential anaerobic–aerobic treatment of soil
contaminated with weathered aroclor 1260. Environ Sci
Technol 36: 100–103.
Maymo-Gatell X, Chien YT, Gossett JM & Zinder SH (1997)
Isolation of a bacterium that reductively dechlorinates
tetrachloroethene to ethane. Science 276: 1568–1571.
McCarty P & Semperini L (1994) Groundwater treatment for
chlorinated solvents. Handbook of bioremediation (Norris RD,
Hinchee RE, Brown R, McCarty PL, Semprini L, Wilson DH,
Kampbell M, Reinhard EG, Bouwer R, Borden C, Vogel TM,
Thomas JM, Ward CH, eds), pp. 87–116. Lewis Publisher,
Boca Raton, FL.
Meade T & D’Angelo EM (2005) [C-14]Pentachlorophenol
mineralization in the rice rhizosphere with established
oxidized and reduced soil layers. Chemosphere 61: 48–55.
FEMS Microbiol Ecol 72 (2010) 74–88Journal compilation c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original German government works
86 G. Imfeld et al.

Nijenhuis I, Nikolausz M, Koth A et al. (2007) Assessment of the
natural attenuation of chlorinated ethenes in an anaerobic
contaminated aquifer in the Bitterfeld/Wolfen area using stable
isotope techniques, microcosm studies and molecular
biomarkers. Chemosphere 67: 300–311.
Nikolausz M, Chatzinotas A, Palatinszky M, Imfeld G, Martinez P
& Kastner M (2008) Single-nucleotide primer extension assay
for detection and sequence typing of ‘Dehalococcoides’ spp.
Appl Environ Microb 74: 300–304.
Nocker A, Burr M & Camper AK (2007) Genotypic microbial
community profiling: a critical technical review. Microb Ecol
54: 276–289.
Nubel U, Engelen B, Felske A et al. (1996) Sequence
heterogeneities of genes encoding 16S rRNAs in Paenibacillus
polymyxa detected by temperature gradient gel electrophoresis.
J Bacteriol 178: 5636–5643.
Oksanen J, Kindt R, Legendre P, O’Hara RB, Simpson GL,
Solymos P, Stevens MHH & Wagner H (2008) Vegan:
community ecology package version 1.15-1. Available at http://
cran.r-project.org/
Ramette A (2007) Multivariate analyses in microbial ecology.
FEMS Microbiol Ecol 62: 142–160.
R Development Core Team (2008) R: A Language and
Environment for Statistical Computing. R Foundation for
Statistical Computing, Vienna, Austria. http://www.R-project.
org.
Reddy KR & Dangelo EM (1997) Biogeochemical indicators to
evaluate pollutant removal efficiency in constructed wetlands.
Water Sci Technol 35: 1–10.
Scala DJ, Hacherl EL, Cowan R, Young LY & Kosson DS (2006)
Characterization of Fe(III)-reducing enrichment cultures and
isolation of Fe(III)-reducing bacteria from the Savannah River
site, South Carolina. Res Microbiol 157: 772–783.
Schlotelburg C, von Wintzingerode C, Hauck R, von
Wintzingerode F, Hegemann W & Gobel U (2002) Microbial
structure of an anaerobic bioreactor population that
continuously dechlorinates 1,2-dichloropropane. FEMS
Microbiol Ecol 39: 229–237.
Seshadri R, Adrian L, Fouts D et al. (2005) Genome sequence of
the PCE-dechlorinating bacterium Dehalococcoides
ethenogenes. Science 307: 105–108.
Stottmeister U & Wiessner A (2003) Effects of plants and
microorganisms in constructed wetlands for wastewater
treatment. Biotechnol Adv 22: 93–117.
Sung Y, Fletcher KF, Ritalaliti KM et al. (2006) Geobacter lovleyi sp
nov strain SZ, a novel metal-reducing and tetrachloroethene-
dechlorinating bacterium. Appl Environ Microb 72: 2775–2782.
Tang YJJ, Yi S, Zhuang WQ, Zinder SH, Keasling JD & Alvarez-
Cohen L (2009) Investigation of carbon metabolism in
‘Dehalococcoides ethenogenes’ strain 195 by use of isotopomer
and transcriptomic analyses. J Bacteriol 191: 5224–5231.
Tanner CC & Sukias JP (1995) Accumulation of organic solids in
gravel-bed constructed wetlands. Water Sci Technol 32:
229–239.
Truu J, Nurk K, Juhanson J & Mander U (2005) Variation of
microbiological parameters within planted soil filter for
domestic wastewater treatment. J Environ Sci Heal A 40:
1191–1200.
Vogel TM (1994) Natural bioremediation of chlorinated solvents.
Handbook of Bioremediation (Norris RD, Hinchee RE, Brown
R, et al., eds), pp. 201–225. Lewis Publisher, Boca Raton, FL.
Walsh KA, Hill TCJ, Moffett BF, Harris JA, Shaw PJ & Wallace JS
(2002) Molecular characterisation of bacteria in a wetland
used to remove ammoniacal-N from landfill leachate. Waste
Manage Res 20: 529–535.
Wang Q, Garrity GM, Tiedje JM & Cole JR (2007) Naive Bayesian
classifier for rapid assignment of rRNA sequences into the new
bacterial taxonomy. Appl Environ Microb 73: 5261–5267.
Watts JEM, Fagervold SK, May HD & Sowers KR (2005) A PCR-
based specific assay reveals a population of bacteria within the
Chloroflexi associated with the reductive dehalogenation of
polychlorinated biphenyls. Microbiol-SGM 151: 2039–2046.
Weiss JV & Cozzarelli IM (2008) Biodegradation in contaminated
aquifers: incorporating microbial/molecular methods. Ground
Water 46: 305–322.
Supporting Information
Additional Supporting Information may be found in the
online version of this article:
Appendix S1. Additional information and results.
Fig. S1. Prevailing spatial hydrogeochemical gradients de-
velopment in the sand compartment of the model wetland
at days 227, 283 and 311 of the investigation.
Fig. S2. Iron sulphide minerals precipitation in the model
subsurface horizontal-flow model wetland treating cis- and
trans-DCE-contaminated groundwater at day 227.
Fig. S3. Example of the DGGE patterns of Bacteria 16S rRNA
gene PCR products amplified from water-DNA extracts
obtained across the model wetland at the tank, the inflow,
the sand compartment (Sample AB: pooled samples retrieved
at 6 and 49 cm from the inflow; and CD: pooled samples
retrieved at 94 and 139 cm from the inflow) and the pond.
Fig. S4. Rarefaction curve of bacterial 16S rRNA gene clones
recovered from groundwater (m) and from the wetland (�).
Fig. S5. DGGE patterns of Archaea 16S rRNA gene PCR
products amplified from water-DNA extracts obtained
across the model wetland at the tank, the inflow, the sand
compartment (pooled samples retrieved at 6 and 49 cm
from the inflow) and the pond at day 255.
Table S1. Results of the geochemical and contaminant
analyses in the sand compartment of the model constructed
wetland at day 199 of the microbial investigation period.
Table S2. Results of the geochemical and contaminant
analyses in the sand compartment of the model constructed
wetland at day 227 of the microbial investigation period.
FEMS Microbiol Ecol 72 (2010) 74–88 Journal compilation c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original German government works
87Microbial communities in a wetland treating dichloroethenes

Table S3. Results of the geochemical and contaminant
analyses in the sand compartment of the model constructed
wetland at day 255 of the microbial investigation period.
Table S4. Results of the geochemical and contaminant
analyses in the sand compartment of the model constructed
wetland at day 283 of the microbial investigation period.
Table S5. Results of the geochemical and contaminant
analyses in the sand compartment of the model constructed
wetland at day 311 of the microbial investigation period.
Table S6. Results of the geochemical and contaminant
analyses in the sand compartment of the model constructed
wetland at day 340 of the microbial investigation period.
Table S7. Results of the geochemical and contaminant
analyses in the sand compartment of the model constructed
wetland at day 396 of the microbial investigation period.
Table S8. Results of the geochemical and contaminant
analyses in the sand compartment of the model constructed
wetland at day 430 of the microbial investigation period.
Please note: Wiley-Blackwell is not responsible for the
content or functionality of any supporting materials sup-
plied by the authors. Any queries (other than missing
material) should be directed to the corresponding author
for the article.
FEMS Microbiol Ecol 72 (2010) 74–88Journal compilation c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. No claim to original German government works
88 G. Imfeld et al.
![Synthesis, Antimalarial Activity, and Molecular Modeling of New Pyrrolo[1,2- a ]quinoxalines, Bispyrrolo[1,2- a ]quinoxalines, Bispyrido[3,2- e ]pyrrolo[1,2- a ]pyrazines, and Bispyrrolo[1,2-](https://img.dokumen.tips/doc/110x75/635500188ae64d6d7f0af0b9/synthesis-antimalarial-activity-and-molecular-modeling-of-new-pyrrolo12-a-quinoxalines.jpg)




























