Embed Size (px)
Citation preview

Chaos–order transition in foraging behavior of antsLixiang Lia,b, Haipeng Penga,1, Jürgen Kurthsb,1, Yixian Yanga,c, and Hans Joachim Schellnhuberb,d,1
aInformation Security Center, State Key Laboratory of Networking and Switching Technology, and cNational Engineering Laboratory for Disaster Backup andRecovery, Beijing University of Posts and Telecommunications, Beijing 100876, China; bPotsdam Institute for Climate Impact Research, D14473 Potsdam,Germany; and dSanta Fe Institute, Santa Fe, NM 87501
Contributed by Hans Joachim Schellnhuber, April 24, 2014 (sent for review February 5, 2014)
The study of the foraging behavior of group animals (especiallyants) is of practical ecological importance, but it also contributes tothe development of widely applicable optimization problem-solvingtechniques. Biologists have discovered that single ants exhibit low-dimensional deterministic-chaotic activities. However, the influencesof the nest, ants’ physical abilities, and ants’ knowledge (or experi-ence) on foraging behavior have received relatively little attention instudies of the collective behavior of ants. This paper provides newinsights into basic mechanisms of effective foraging for social insectsor group animals that have a home. We propose that the wholeforaging process of ants is controlled by three successive strategies:hunting, homing, and path building. A mathematical model is de-veloped to study this complex scheme. We show that the transitionfrom chaotic to periodic regimes observed in our model results froman optimization scheme for group animals with a home. Accordingto our investigation, the behavior of such insects is not representedby random but rather deterministic walks (as generated by deter-ministic dynamical systems, e.g., by maps) in a random environment:the animals use their intelligence and experience to guide them. Themore knowledge an ant has, the higher its foraging efficiency is. Whenyoung insects join the collective to forage with old and middle-agedants, it benefits the whole colony in the long run. The resulting strat-egy can even be optimal.
foraging dynamics | learning process | low-dimensional chaos |mathematical modeling | synchronization
Both experimental data analysis and mathematical modelingon the foraging behavior of group animals (especially ant
colonies) have recently captured much attention due to the highlevel of self-organizing structures that emerge at the collectivelevel (1–5). Random walking is a widely discussed strategy in theresearch literature on the foraging behavior of group animals(2, 6–8). Some ecologists maintain that especially Lévy flightschemes can appropriately be used to describe the foraging be-havior (6, 7). However, some recent studies have raised doubtswhether this is a valid conjecture (2, 8, 9). It is even argued thatthe rules of locomotion for a walker are always consistent with apurely deterministic model, rather than with a stochastic scheme(9, 10).On the other hand, in the studies on the foraging behavior of
animals, the existence of homes has so far received relativelylittle attention. Here we argue that the existence of a home ornest influences the foraging process to a large extent. Animalsare due to return to their homes because of increasing exhaus-tion of energy. Moreover each foraging process of an animal isalso a learning process. With foraging repetition, long-term mem-ory continues to accumulate, an animal’s knowledge about theenvironment of its nest gets richer, and the region that the animal isfamiliar with continues to enlarge. Moreover, animals’ physicalability and knowledge as determined by their age directly influencetheir foraging strategy. All these factors deserve close attention.There is already a rich history of research on the foraging
behavior of ant colonies (see, e.g., ref. 11). In particular foragingstrategies of ants were discussed in the context of solving dis-tributed control and optimization problems. Already 30 y ago, itwas proposed that Lévy flights might characterize the behav-ior of foraging ants (12). In 1990, Deneubourg et al. designed
a well-known wide binary bridge experiment which showed thatants could mark the path followed by a trail of pheromone andfind an optimal path between the nest and the food source (13).Based on similar experiments, Dorigo and coworkers (3) de-veloped ant colony optimization algorithms which have beenused for solving various difficult problems, including combina-torial optimization, object clustering, and routing selection incommunication networks. A limited binary bridge experimentwas presented to show that ants could even form two lanes tosolve traffic flow problems on crowded branches (4).However, all these experiments were conducted in special
man-designed environments, which were not identical to naturalones, so the ants’ free crawling was restricted. It was arguedthat unrestricted foraging ants might not perform Lévy flights.Moreover, through an experimental study on the dynamicalbehaviors of an isolated ant and a whole ant colony, Cole (14)discovered that the activity of an ant colony exhibited periodicbehavior, whereas the behavior of a single ant showed a low-dimensional deterministic chaotic pattern. In 1993, Solé et al.(15) constructed a 1D chaotic map following Cole to describe theforaging process of an isolated ant. Nemes and Roska (16)designed a cellular neural network model to describe the syn-chronized oscillating pattern of activity as a result of an array ofchaotic dynamic elements placed in a regular 2D grid. In 2006, Liand coworkers (17, 18) developed a chaotic ant swarm modelbuilding on Cole’s research to describe the phenomenon that thechaotic behavior of a single ant contributes to the self-organi-zation behavior of a whole ant colony. These models haveexplained some relationships between the chaotic (or random)strategy, individual dynamics, and group dynamics. However,these studies ignored the possibility that the ants also use theirown experience and intelligence to guide their foraging. Hence,further studies on the influences of physical ability, age, andknowledge on foraging behavior are needed to explain the bi-ological behavior of ants in nature.
Significance
We have studied the foraging behavior of group animals thatlive in fixed colonies (especially ants) as an important problemin ecology. Building on former findings on deterministic chaoticactivities of single ants, we uncovered that the transition fromchaotic to periodic regimes results from an optimization schemeof the self-organization of such an animal colony. We foundthat an effective foraging of ants mainly depends on their nestas well as their physical abilities and knowledge due to expe-rience. As an important outcome, the foraging behavior of antsis not represented by random, but rather by deterministicwalks, in a random environment: Ants use their intelligence andexperience to navigate.
Author contributions: L.L., H.P., and J.K. designed research; L.L., H.P., and Y.Y. performedresearch; H.P. analyzed data; and L.L., J.K., and H.J.S. wrote the paper.
The authors declare no conflict of interest.1To whom correspondence may be addressed. E-mail: [email protected],[email protected], or [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1407083111/-/DCSupplemental.
8392–8397 | PNAS | June 10, 2014 | vol. 111 | no. 23 www.pnas.org/cgi/doi/10.1073/pnas.1407083111

The aim of this paper is to provide a novel perspective onmechanisms of effective foraging of ant colonies. We assume thatthe foraging process of these animals is entirely controlled bythree successive strategies: hunting, homing, and path building. Amathematical model is developed to understand the whole for-aging process. We discuss the influences of the special regionaround the nest, the size of the food source, the search range, thelimitation of ants’ physical ability, and ants’ learning process withrespect to foraging behavior. Our analysis suggests that groupanimals that have a home do not perform random walks, but ratherdeterministic walks in a random environment. They use theirknowledge to guide them and their behavior is also influenced bytheir physical abilities, their age, and the existence of homes.
Foraging Strategies of AntsTo survive, ants need to leave their nest and forage for food. Thesurvival-of-the-fittest mechanism entails that ants do not onlyfind food, but also an optimal path between their nest and thefood source (19, 20). This reflects the collective intelligence of theinsects. Nest and food source indeed play important roles in ants’foraging behavior. The whole foraging process of ants may be de-scribed by three strategies: hunting, homing, and path building.
Hunting Strategy. During the first phase in foraging, some ants ofthe colony leave the nest to search for food; we call these “scoutants” (21). We assume that there is a food source located insome random environment. We consider the search for food asthe search for an unknown point (or region). Thus, initially, noant knows where the food is or in which direction it should go. Inparticular, there is no pheromone on the path before the foodsource is found. From Cole’s observational results on the be-havior of individual ants, we know that ants search chaoticallyaround their nest and initially have no impacts on their neigh-bors. For the details of chaotic and self-organizing ant behavior,see SI Text.Suppose that initially n ants go out to look for food. These ants
forage in a continuous search space S, often denoted Rl, the l-dimensional continuous space of real numbers, i.e., S = Rl. Eachant with a position vector ~si =~Zi =
�Zi1; . . . ;Zil
�attempts to
search for the location ~Pfood =�Pfood1; . . . ;Pfoodl
�of a food source
by minimizing an object function (cost function) f : S→R,where S = Rl and f ≥ 0, is a map from l dimensions real numberto real number, and when ~Zi =~Pfood; f
�~Zi
�= f
�~Pfood
�= 0. In
the search space S = Rl, the food possesses a small region (i.e., theneighborhood of the point ~Pfood). The bigger the food is, the largerthe region is. The values of the function f corresponding to differentpoints in the neighborhood of the food source have an upperbound △, where △ is a relatively small positive number. Here,larger △ means larger region around the point ~Pfood. So the valueof △ represents the size of the food source. f
�~Zi
�< △ means the
point ~Zi locates in a small neighborhood of the food source, i.e., theant finds the food.Fig. 1A provides a schematic diagram of ants crawling in the
search space where the black solid point represents the nest andthe green triangle the nearest food source. When food is scarce,ants need to go out to forage many times because they mustreturn to the nest to access the stored energy supply. We havedeveloped a map of the nest–food source–nest motion againsttime. This is sketched in Fig. 1B where the solid dot again rep-resents the nest and the green triangle the food source. Thelength of the optimal path from the nest to the food source isdenoted by L. In this map, ants forage chaotically, setting out atthe origin. If they do not find food, then the curve does not passthrough the green triangle. Because n ants conduct concurrentsearches, there is likely some ant i that quickly finds the foodsource (Fig. 1B). This hunting strategy, whereby ants leave thenest and return to it, is used throughout the foraging process.
Homing Strategy. The existence of a nest has an important in-fluence on the behavior of the ants during the whole search forfood. The homing strategy is triggered when a foraging antmakes a decision to return to the nest. The aim of that strategy isto find the home most efficiently. The nest of the ants is denotedby ~Pnest =
�Pnest1; . . . ;Pnestl
�. Different ants have different knowl-
edge about the environment of the nest, i.e., each ant considersdifferent domains Mi as nest neighborhoods (22). If ant i movesinto its nest neighborhood Mi =f~Zi :
��~Zi −~Pnest
��< cig, then weassume that it has found the nest (Fig. 1A), where the neigh-borhood range is determined by the positive constant ci, and alarger ci indicates that the ant has more knowledge about thenest environment. Obviously, older ants have larger ci becausethey have more knowledge about their nest. Foraging can beconsidered as searching for an unknown point (or small region),whereas searching for the nest is considered as searching fora special region with which an ant is very familiar. In the homingprocess, the ants use chaos and pheromone in combination withtheir own knowledge to search for the nest.Different ants have different search ranges, different continuing
search times, and different search paths. The starting time of thehoming strategy is very important. If it is triggered very early, thenthe probability of ants finding food will be minute; if it happensvery late, however, then the ants may not be able to return to thenest because their energy is exhausted. That is, each ant has its ownsearching time threshold, i.e., the tiring time ttired. When ttired isreached, the ant should start its homing strategy, independent ofhaving found food or not.
Fig. 1. Stylized presentation of the ant foraging process. The optimizedpath between the nest (black dot) and the food source (green triangle) haslength L. (A) Characteristic search behavior of scout ants distinguishedby different path colors. S is the overall foraging space and M the nest’sneighborhood, which the ants are quite familiar with. (B) The foragingmotion in S can be mapped onto a simplified trajectory, i.e., a relationship ofposition vs. time. As detailed in SI Text, time t is normalized in a way thathighlights periodicity and is represented by the abscissa in B–D. In the samecharts, the ordinate d represents the nearest distance of the respective antposition from the set L. Thus, each foraging excursion appears as a curvesegment between two consecutive black dots (home-to-home tour). Bdescribes the situation where n scouts search chaotically in parallel. The ithant happens to find the food source during its kth tour (as indicated by thepassing of the trajectory through the green triangle). (C and D) m-recruitedinsects are guided by the pheromone traces left by the successful ant i. (C)Initially, the pheromone intensity is rather low, thus its guiding influence onthe chaotic ant motion is very small. To find the optimal path, the recruitedinsects pursue various chaotically selected paths around the route marked bythe pheromone from the scout ant i. However, all those paths hit the greentriangle because the followers “know” the position of the food source fromthe pioneer. (D) With increasing pheromone concentration around the op-timal path, the chaotic foraging regime is gradually overcome. Eventuallythe new regime emerges where all ants pursue L to convey food from sourceto nest. Thus, all trajectories become straight lines.
Li et al. PNAS | June 10, 2014 | vol. 111 | no. 23 | 8393
PHYS
ICS
ECOLO
GY

Path-Building Strategy. When a scout ant finds a food source, itbegins returning to the nest and leaving pheromone on its returnpath as a means of communicating to fellow ants the location ofthe food and appropriate paths (3, 4). Once the scout ant hasreturned to the nest, it will gather recruited ants to follow its pathmarked with pheromone. We assume there are m recruits. Inaddition, there are still some freely walking scout ants looking forfood in the search space.Both the chaotic walking and the pheromone have important
impacts on the foraging process of recruited ants. Pheromoneinfluences their choice of path making. Ants encountering a pathpreviously laid with heavy amounts of pheromone are more likelyto decide to follow it. Pheromone is an evaporative substance,though. An autocatalytic collective behavior of recruited antsemerges, where the shorter the path, the higher its pheromonedensity becomes (23). During the initial construction phase ofthe optimal path, the orchestration between ants is weak, andchaotic movement dominates their behavior because of the verylow density of pheromone. Fig. 1C depicts the initial foragingbehavior of m recruited ants. This figure illustrates how the antsinitially foraged chaotically around the route marked by thepheromone of the scout ant, while leaving more pheromone ontheir wandering paths. This means that chaotic walking andpheromone interact in the optimization process.We use the pheromone field concept to help us understand
the behavior of the recruited ants. The pheromone field is thecause of self-organization among ants and is used to direct theirmovements. As time passes, the pheromone left by the recruitedants on their paths increases gradually, and a distinct pheromonefield is constructed between the food source and the nest. Thereinforcement of pheromone continues to weaken the chaoticbehavior of the ants. The evaporation of the pheromone causesthe density of pheromone on the shorter path to increase morequickly than on the longer one, which in turn causes more ants tochoose the shorter path. The increment of pheromone density onthe paths is equivalent to a decrement in the chaotic crawling ofinsects (SI Text). During the formation of the pheromone fieldand the finding of the optimal route, the ants still possess somechaotic crawling, which is eventually superseded by the phero-mone signals.Fig. 1D shows the final foraging phase of the m recruited ants.
The ants gradually enter a state of ordered periodic motionthrough self-organization in the colony. Obviously, the chaoticbehavior causes the ants to conduct a global search, whereas thepheromone field causes the ants to conduct a local search untilthey finally converge to periodic movements. In this process, thetransformation of dynamical behavior of an ant colony causes theemergence of something that may be called “swarm intelligence.”Thus, we consider the process by which ants begin their for-
aging and form their optimal path to be an intelligent process,whereby the status of ants is transformed from an asynchronouschaotic regime into a collective synchronous periodic act. Antsuse their intelligence and experience to guide their foragingprocess. We regard a foraging cycle of ants as the process inwhich ants leave their nest to find a food source until an optimalpath is found and then convey all of the food to their nest. Afterone foraging cycle is completed, the ants initiate another one tosurvive, searching for a new food source.
ResultsNow we show how the basic principles sketched above areapplied. For simplicity, we assume that there is only one foodsource in the search space and that there is only one optimalroute between the food source and the nest. The chaotic modelZ′ðt+ 1Þ=Z′ðtÞeμð1−Z′ðtÞÞ, constructed in ref. 15, is introduced tomimic the random activity of a single ant at the beginning of theforaging process, where Z′∈R+ is a continuous variable and μ
a positive constant. When μ= 3, the system is in a chaotic state(15, 18).Next, we take into account the influence of the nest and the
food source on the ants’ motion. Here, the organization of antssets in under the competing influences of the pheromone andthe ants’ chaotic crawling. The characteristic variable of chaoticcrawling represented by yi(t) is introduced, where 0≤ yiðtÞ< 1,and the value indicates the degree of chaotic crawling. A larger ymeans a higher degree of chaotic crawling. Based on the mecha-nism of chaotic annealing (24), the continual decrement dynamicsof yi(t) is represented by yiðtÞ= yiðt− 1Þð1+riÞ, which depends onthe self-organization factor ri (see Methods for details on ri). Themovement adjustment of each ant is executed as follows:
Zikðt+ 1Þ=�ZikðtÞ+Vk
�eð1−e−ayiðtÞÞ
�3−ψkðZikðtÞ+VkÞ
�
−Vk + e−2ayiðtÞ+b�jsinðωtÞj�Pfoodk −Pnestk
�− ðZikðtÞ−PnestkÞ
�;
[1]
where ψk can adjust the search range, V determines the searchregion of ant i and accounts for the option that ants can roamdiverse realms, ω is used to adjust the frequency of ants’ periodicoscillation between the nest and the food source, a is a suffi-ciently large positive constant such that the variable yi(t) couldhave a large enough impact on the position vector Zik, and b isthe local search factor which controls the local optimal pathstrategy. When yi approaches 0, b begins to work, where 0 ≤b < ln(2). If b ≥ ln(2), the system is unstable. When b = 0, thesystem is in a periodic oscillatory regime between the nest andthe food source without undergoing the process of a local search.When 0 < b < ln(2), the system starts from a transient chaos stateand finally converges to a periodic behavior.Now we analyze and examine the nonlinear dynamics of the
proposed chaotic ant foraging model. We use it to solve a con-crete optimization problem whose objective function is definedby f ðx1; x2Þ= ðx− 0:7Þ2 × ð0:1+ ð0:6+ x2Þ2Þ+ ðx2 − 0:5Þ2 × ð0:15+ð0:4+ x1Þ2Þ, where (0.7,0.5) is the global minimum of the energyfunction f. To simulate the foraging cycle, we assume that thepoint (0.7,0.5) is the position of the food source, and (0.4,0.4) isthe position of the nest. All of the figures in this section are theresults of numerical simulations for Eq. 1.
Food Hunting and Homing Processes. Ants of different ages havedifferent physical abilities and different knowledge about theirnest. The age of the ants thus has a significant impact on theirforaging behavior. Fig. S1 displays the foraging probability curvesof ants with different ages as △ changes, where △ is the size offood source. We find that when △ is fixed, old and middle-agedants hit the food source much more easily than the young ones.From Fig. S1, we also see that on the whole, the probability offinding food for an ant increases as △ increases, which is notvery surprising.Obviously, the tiring time ttired and the nest neighborhood
range constant c play crucial roles in the search process, whichshows that the physical ability and the knowledge the ants havehave an important impact on their foraging behavior. Fig. 2 Aand B shows the influence of ttired on the probability an antfinding food for different △. We see first that for a given groupsize, the foraging success increases as ttired grows. The greaterthe physical ability of an ant, the larger its ttired, i.e., the greater thephysical ability, the easier the foraging. Second, the higher thenumber of foraging ants, the larger the probability of findingfood. This agrees with traditional views reported in existingstudies on optimization (3, 17). Third, when the value of ttired isfixed, the larger the size of the food source, the greater theprobability of finding food.
8394 | www.pnas.org/cgi/doi/10.1073/pnas.1407083111 Li et al.
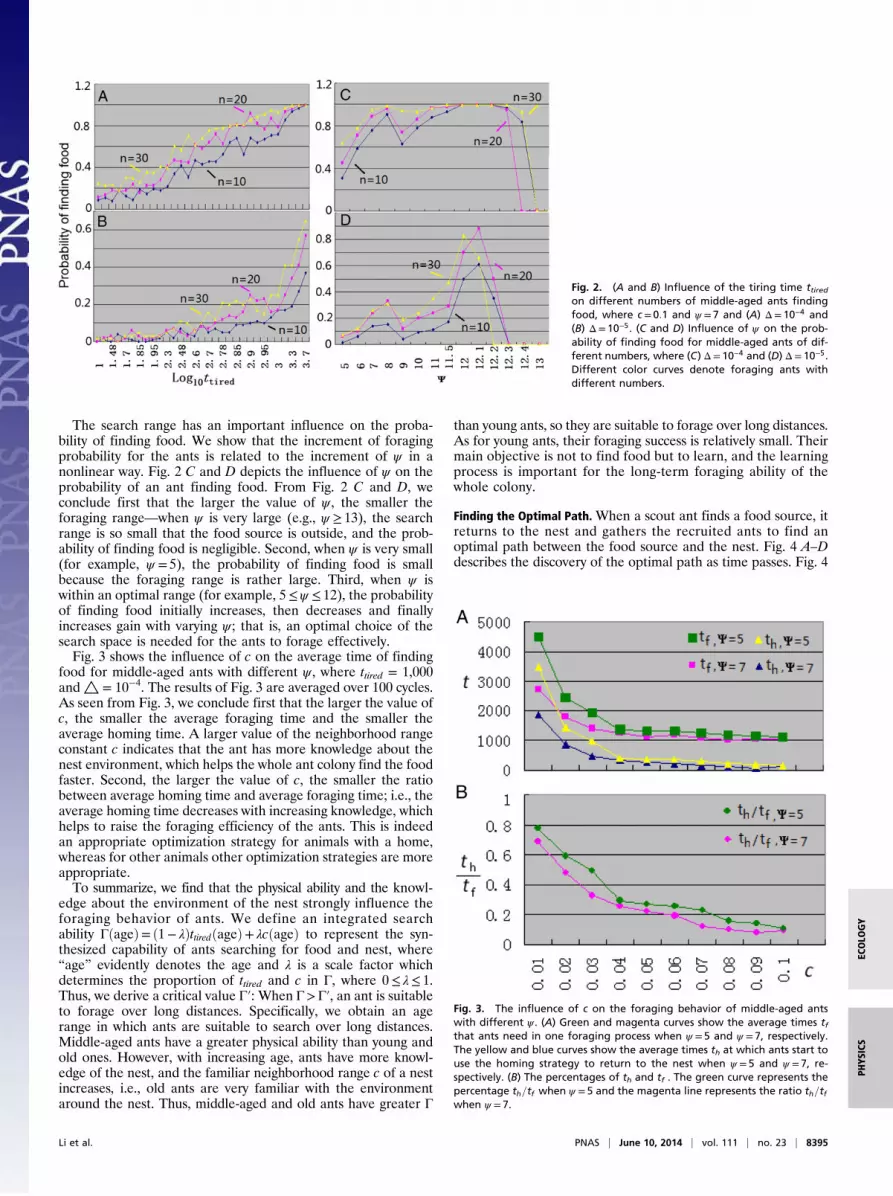
The search range has an important influence on the proba-bility of finding food. We show that the increment of foragingprobability for the ants is related to the increment of ψ in anonlinear way. Fig. 2 C and D depicts the influence of ψ on theprobability of an ant finding food. From Fig. 2 C and D, weconclude first that the larger the value of ψ , the smaller theforaging range—when ψ is very large (e.g., ψ ≥ 13), the searchrange is so small that the food source is outside, and the prob-ability of finding food is negligible. Second, when ψ is very small(for example, ψ = 5), the probability of finding food is smallbecause the foraging range is rather large. Third, when ψ iswithin an optimal range (for example, 5≤ψ ≤ 12), the probabilityof finding food initially increases, then decreases and finallyincreases gain with varying ψ ; that is, an optimal choice of thesearch space is needed for the ants to forage effectively.Fig. 3 shows the influence of c on the average time of finding
food for middle-aged ants with different ψ , where ttired = 1,000and△ = 10−4. The results of Fig. 3 are averaged over 100 cycles.As seen from Fig. 3, we conclude first that the larger the value ofc, the smaller the average foraging time and the smaller theaverage homing time. A larger value of the neighborhood rangeconstant c indicates that the ant has more knowledge about thenest environment, which helps the whole ant colony find the foodfaster. Second, the larger the value of c, the smaller the ratiobetween average homing time and average foraging time; i.e., theaverage homing time decreases with increasing knowledge, whichhelps to raise the foraging efficiency of the ants. This is indeedan appropriate optimization strategy for animals with a home,whereas for other animals other optimization strategies are moreappropriate.To summarize, we find that the physical ability and the knowl-
edge about the environment of the nest strongly influence theforaging behavior of ants. We define an integrated searchability ΓðageÞ= ð1− λÞttiredðageÞ+ λcðageÞ to represent the syn-thesized capability of ants searching for food and nest, where“age” evidently denotes the age and λ is a scale factor whichdetermines the proportion of ttired and c in Γ, where 0≤ λ≤ 1.Thus, we derive a critical value Γ′: When Γ>Γ′, an ant is suitableto forage over long distances. Specifically, we obtain an agerange in which ants are suitable to search over long distances.Middle-aged ants have a greater physical ability than young andold ones. However, with increasing age, ants have more knowl-edge of the nest, and the familiar neighborhood range c of a nestincreases, i.e., old ants are very familiar with the environmentaround the nest. Thus, middle-aged and old ants have greater Γ
than young ants, so they are suitable to forage over long distances.As for young ants, their foraging success is relatively small. Theirmain objective is not to find food but to learn, and the learningprocess is important for the long-term foraging ability of thewhole colony.
Finding the Optimal Path. When a scout ant finds a food source, itreturns to the nest and gathers the recruited ants to find anoptimal path between the food source and the nest. Fig. 4 A–Ddescribes the discovery of the optimal path as time passes. Fig. 4
Fig. 2. (A and B) Influence of the tiring time ttiredon different numbers of middle-aged ants findingfood, where c= 0:1 and ψ = 7 and (A) Δ= 10−4 and(B) Δ= 10−5. (C and D) Influence of ψ on the prob-ability of finding food for middle-aged ants of dif-ferent numbers, where (C) Δ= 10−4 and (D) Δ= 10−5.Different color curves denote foraging ants withdifferent numbers.
Fig. 3. The influence of c on the foraging behavior of middle-aged antswith different ψ . (A) Green and magenta curves show the average times tfthat ants need in one foraging process when ψ = 5 and ψ = 7, respectively.The yellow and blue curves show the average times th at which ants start touse the homing strategy to return to the nest when ψ = 5 and ψ =7, re-spectively. (B) The percentages of th and tf . The green curve represents thepercentage th=tf when ψ = 5 and the magenta line represents the ratio th=tfwhen ψ = 7.
Li et al. PNAS | June 10, 2014 | vol. 111 | no. 23 | 8395
PHYS
ICS
ECOLO
GY
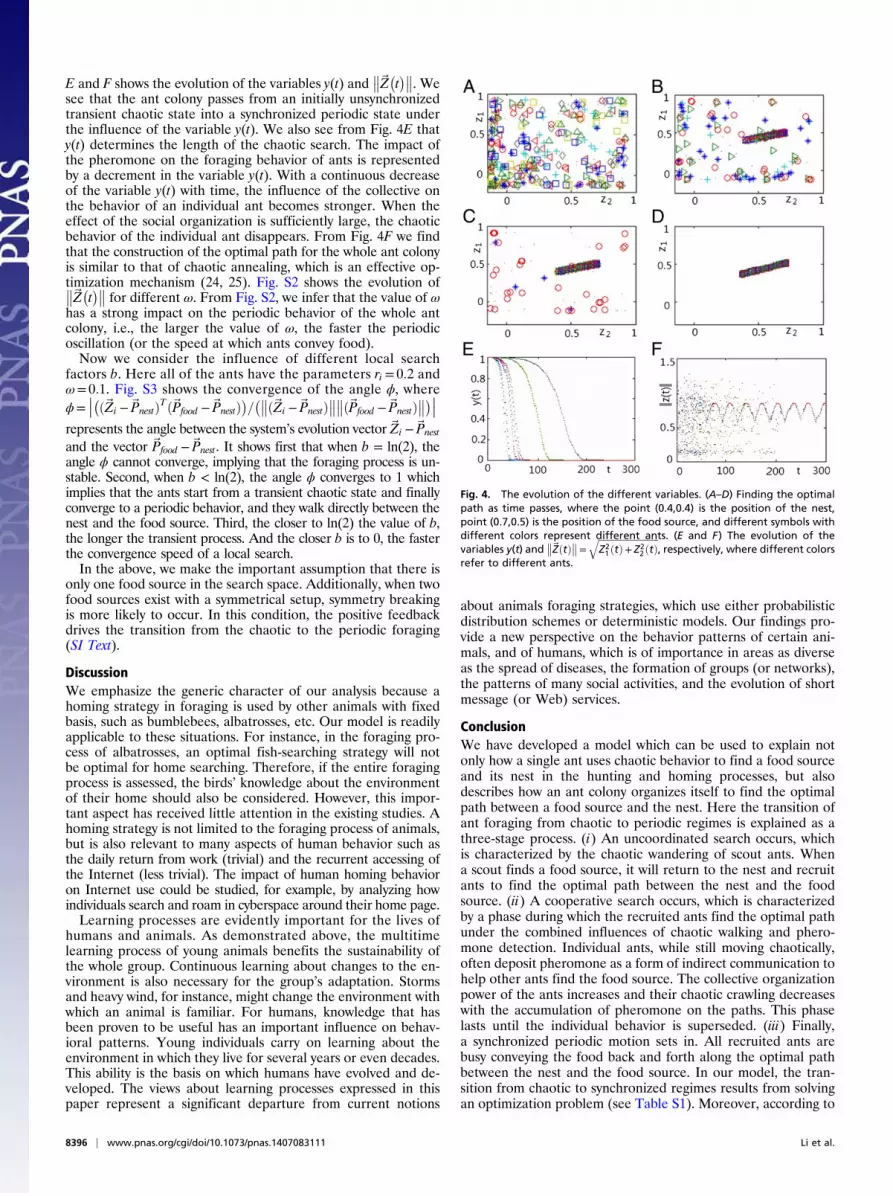
E and F shows the evolution of the variables y(t) and��~Z�t���. We
see that the ant colony passes from an initially unsynchronizedtransient chaotic state into a synchronized periodic state underthe influence of the variable y(t). We also see from Fig. 4E thaty(t) determines the length of the chaotic search. The impact ofthe pheromone on the foraging behavior of ants is representedby a decrement in the variable y(t). With a continuous decreaseof the variable y(t) with time, the influence of the collective onthe behavior of an individual ant becomes stronger. When theeffect of the social organization is sufficiently large, the chaoticbehavior of the individual ant disappears. From Fig. 4F we findthat the construction of the optimal path for the whole ant colonyis similar to that of chaotic annealing, which is an effective op-timization mechanism (24, 25). Fig. S2 shows the evolution of��~Z�t��� for different ω. From Fig. S2, we infer that the value of ωhas a strong impact on the periodic behavior of the whole antcolony, i.e., the larger the value of ω, the faster the periodicoscillation (or the speed at which ants convey food).Now we consider the influence of different local search
factors b. Here all of the ants have the parameters ri = 0:2 andω= 0:1. Fig. S3 shows the convergence of the angle ϕ, whereϕ=
���ð~Zi −~PnestÞTð~Pfood −~PnestÞ�=���ð~Zi −~PnestÞ
����ð~Pfood −~PnestÞ�����
represents the angle between the system’s evolution vector~Zi −~Pnest
and the vector ~Pfood −~Pnest. It shows first that when b = ln(2), theangle ϕ cannot converge, implying that the foraging process is un-stable. Second, when b < ln(2), the angle ϕ converges to 1 whichimplies that the ants start from a transient chaotic state and finallyconverge to a periodic behavior, and they walk directly between thenest and the food source. Third, the closer to ln(2) the value of b,the longer the transient process. And the closer b is to 0, the fasterthe convergence speed of a local search.In the above, we make the important assumption that there is
only one food source in the search space. Additionally, when twofood sources exist with a symmetrical setup, symmetry breakingis more likely to occur. In this condition, the positive feedbackdrives the transition from the chaotic to the periodic foraging(SI Text).
DiscussionWe emphasize the generic character of our analysis because ahoming strategy in foraging is used by other animals with fixedbasis, such as bumblebees, albatrosses, etc. Our model is readilyapplicable to these situations. For instance, in the foraging pro-cess of albatrosses, an optimal fish-searching strategy will notbe optimal for home searching. Therefore, if the entire foragingprocess is assessed, the birds’ knowledge about the environmentof their home should also be considered. However, this impor-tant aspect has received little attention in the existing studies. Ahoming strategy is not limited to the foraging process of animals,but is also relevant to many aspects of human behavior such asthe daily return from work (trivial) and the recurrent accessing ofthe Internet (less trivial). The impact of human homing behavioron Internet use could be studied, for example, by analyzing howindividuals search and roam in cyberspace around their home page.Learning processes are evidently important for the lives of
humans and animals. As demonstrated above, the multitimelearning process of young animals benefits the sustainability ofthe whole group. Continuous learning about changes to the en-vironment is also necessary for the group’s adaptation. Stormsand heavy wind, for instance, might change the environment withwhich an animal is familiar. For humans, knowledge that hasbeen proven to be useful has an important influence on behav-ioral patterns. Young individuals carry on learning about theenvironment in which they live for several years or even decades.This ability is the basis on which humans have evolved and de-veloped. The views about learning processes expressed in thispaper represent a significant departure from current notions
about animals foraging strategies, which use either probabilisticdistribution schemes or deterministic models. Our findings pro-vide a new perspective on the behavior patterns of certain ani-mals, and of humans, which is of importance in areas as diverseas the spread of diseases, the formation of groups (or networks),the patterns of many social activities, and the evolution of shortmessage (or Web) services.
ConclusionWe have developed a model which can be used to explain notonly how a single ant uses chaotic behavior to find a food sourceand its nest in the hunting and homing processes, but alsodescribes how an ant colony organizes itself to find the optimalpath between a food source and the nest. Here the transition ofant foraging from chaotic to periodic regimes is explained as athree-stage process. (i) An uncoordinated search occurs, whichis characterized by the chaotic wandering of scout ants. Whena scout finds a food source, it will return to the nest and recruitants to find the optimal path between the nest and the foodsource. (ii) A cooperative search occurs, which is characterizedby a phase during which the recruited ants find the optimal pathunder the combined influences of chaotic walking and phero-mone detection. Individual ants, while still moving chaotically,often deposit pheromone as a form of indirect communication tohelp other ants find the food source. The collective organizationpower of the ants increases and their chaotic crawling decreaseswith the accumulation of pheromone on the paths. This phaselasts until the individual behavior is superseded. (iii) Finally,a synchronized periodic motion sets in. All recruited ants arebusy conveying the food back and forth along the optimal pathbetween the nest and the food source. In our model, the tran-sition from chaotic to synchronized regimes results from solvingan optimization problem (see Table S1). Moreover, according to
Fig. 4. The evolution of the different variables. (A–D) Finding the optimalpath as time passes, where the point (0.4,0.4) is the position of the nest,point (0.7,0.5) is the position of the food source, and different symbols withdifferent colors represent different ants. (E and F) The evolution of thevariables y(t) and
��~ZðtÞ��= ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiZ21 ðtÞ+ Z2
2 ðtÞq
, respectively, where different colorsrefer to different ants.
8396 | www.pnas.org/cgi/doi/10.1073/pnas.1407083111 Li et al.

our analysis, physical ability, experience, and the existence ofa nest have important impacts on the foraging behavior ofant colonies.Through numerical experiments, we reach the following main
conclusions. (i) The age of the ants is crucial. Old and middle-aged ants find a food source much more easily than the youngones. However, pursuing a strategy whereby young ants foragetogether with old and middle-aged ants can be optimal because itbenefits the long-term foraging prospects of the whole colony.(ii) The physical ability of a single ant is also crucial. The greaterthe physical performance, the better the foraging. Therefore, it iseasier for a middle-aged ant to find a food source than youngerand older ants. (iii) The search range has an important influenceon the probability of finding food. In order for ants to forageeffectively, the range of the search space should lie within anoptimal realm. (iv) The foraging efficiency of group animals withhomes is clearly different from those without. For group animals,more knowledge about the neighborhood of the nest increasesforaging efficiency, i.e., the more knowledge of its home an an-imal has, the shorter its homing time. Based on these insights, wesuggest that for group animals that have a home, their foragingbehavior should not be characterized by random walking butrather by deterministic walking in random environments.
MethodsIn the foraging process, the movement strategy of a single ant i depends onthe current position of the ant ~ZiðtÞ, the best position found by itself or anyone of its neighbors, the position of the nest ~Pnest , the position of the foodsource~Pfood , the characteristic variable of chaotic crawling yiðtÞ, and the self-organization factor ri . Generally, the following function is used to describethe whole foraging process of ants:
~Zi
�t + 1
�=g
~Zi�t�,~Pfood ,~Pnest ,yi
�t�,ri
, [2]
where t means the current time step and g is a nonlinear function.To mimic an initially chaotic search, we introduce the chaotic model
Z′ðt + 1Þ= Z′ðtÞeμð1−Z′ðtÞÞ described by Solé et al. in ref. 15. Let Z′= ð1=μÞψZ,then we get Zðt + 1Þ= ZðtÞeμ−ψ · ZðtÞ, and the search center is approximately7:5=ð2ψÞ.
Here, the organization of ants sets in under the influences of the pher-omone and the chaotic crawling of ants. As time evolves, the pheromoneintensity increases and the chaotic crawling of ants is gradually reduced.Based on the annealing mechanism (24, 25), the adjustment of the chaoticbehavior of individual ant i is achieved by introducing a successively decreasingdynamical equation represented by yiðtÞ= yiðt −1Þð1+ri Þ. The self-organizationfactor ri is used to control the time of the chaotic search. If ri is very large, the
chaotic search is short, and vice versa. Because small changes are desired as timeevolves, ri is chosen typically in the range [0,0.5].
Moreover, the term jsinðωtÞjðPfoodk − PnestkÞ− ðZikðtÞ− PnestkÞ is introducedto achieve periodic oscillating behavior of an individual ant between thenest and the food source, where k is the kth dimension of the position vector.
The length of the optimal path is L=ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiPl
k=1
�Pfoodk − Pnestk
�2q. Adjustment of
the position of each ant obeys Eq. 1.In the food searching process, because there is no organization initially, the
position of the food source could be found by setting ri = 0. The foraging strategyof ants is to search ~Zi such that fð~ZiÞ<Δ. When ri = 0, e−ayi ðtÞ, and e−2ayi ðtÞ+bi
approximate to 0, and Eq. 1 applies, we get the following chaotic model:
Zik�t + 1
�=�ZikðtÞ+Vk
�e�3−ψk
�Zik ðtÞ+Vk
��−Vk : [3]
That is, the ants walk chaotically throughout the foraging process. Here,Vk = 7:5=ð2ψkÞ− Pnestk . The ants center around the nest and search for food.The search diameter ϕk depends on ψk and ϕk ≈ 7:5=ψk . Because all the nants conduct a parallel search in the search space, the ant colony quicklyfinds the food source ~Pfood .
In searching for the nest, the main aim of ant i is to use its homing strategyto find the neighborhood of its nest such that Mi =f~Zi :
��~Zi −~Pnest��< cig.
Here, older ants have larger ci because they have more knowledge abouttheir nest.
During the optimal path finding process, self-organization in the antcolony gradually occurs, where ri > 0. Under the influence of self-organiza-tion, the variable yiðtÞ of ants is gradually attenuated to 0. Eq. 1 is then atransient chaotic convergence process. The larger ri , the faster the alterationof yiðtÞ, and the faster the self-organization process of the system is formed.When yiðtÞ approaches 0, both e−ayi ðtÞ and e−2ayi ðtÞ approach 1, and param-eter b begins to work. Here, the search model becomes
Zik�t + 1
�= ZikðtÞ+ eb
�jsinðωtÞjðPfoodk − PnestkÞ− ðZikðtÞ− PnestkÞ�:
When 0 < b < ln(2), the system starts from a transient chaos state and finallyconverges to a periodic behavior, and the ants walk between the nest andthe food source to convey food. That is, the angle ϕ converges from ϕ< 1 toϕ= 1, where i = 1,. . .,n.
Because the chaotic search belongs to a global search and the searchcaused by pheromone belongs to a local one, the self-organization process isthe one that transfers from the global search to the local one. In this process,the ants finally find the optimal path along which they carry the foodperiodically.
ACKNOWLEDGMENTS. This work was partially supported by InternationalResearch Training Group 1740 (Deutsche Forschungsgemeinschaft andFundação de Amparo à Pesquisa do Estado de São Paulo), the Govern-ment of the Russian Federation (Grant 14.Z50.31.0033), the Beijing Cen-ter for Mathematics and Information Interdisciplinary Sciences, and theNational Natural Science Foundation of China (Grants 61100204 and61121061).
1. Couzin ID, Krause J, Franks NR, Levin SA (2005) Effective leadership and decision-making in animal groups on the move. Nature 433(7025):513–516.
2. Edwards AM, et al. (2007) Revisiting Lévy flight search patterns of wandering alba-trosses, bumblebees and deer. Nature 449(7165):1044–1048.
3. Bonabeau E, Dorigo M, Theraulaz G (2000) Inspiration for optimization from socialinsect behaviour. Nature 406(6791):39–42.
4. Dussutour A, Fourcassié V, Helbing D, Deneubourg JL (2004) Optimal traffic organi-zation in ants under crowded conditions. Nature 428(6978):70–73.
5. Wu Y, Zhou CS, Xiao JH, Kurths J, Schellnhuber HJ (2010) Evidence for a bimodaldistribution in human communication. Proc Natl Acad Sci USA 107(44):18803–18808.
6. Viswanathan GM, et al. (1996) Levy flight search patterns of wandering albatrosses.Nature 381(6581):413–415.
7. Reynolds AM (2011) On the origin of bursts and heavy tails in animal dynamics.Physica A 390(2):245–249.
8. Travis J (2007) Ecology. Do wandering albatrosses care about math? Science 318(5851):742–743.
9. Viswanathan GM, Raposo EP, da Luz MGE (2008) Lévy flights and superdiffusion inthe context of biological encounters and random searches. Phys Life Rev 5(3):133–150.
10. Bunimovich L (2004) Deterministic walks in random environments. Physica D 187(1-4):20–29.
11. Mailleux AC, Detrain C, Deneubourg JL (2005) Triggering and persistence of trail-laying in foragers of the ant Lasius niger. J Insect Physiol 51(3):297–304.
12. Shlesinger MF (1986) On Growth and Form, eds Stanley HF, Ostrowsky N (Nijhoff,Dordrecht, The Netherlands), pp 279–283.
13. Deneubourg JL, Aron S, Goss S, Pasteels JM (1990) The selforganizing Exploratorypattern of the Argentine ant. J Insect Behav 3(2):159–168.
14. Cole BJ (1991) Is animal behavior chaotic? Evidence from the activity of ants. Proc Biol
Sci 244(1311):253–259.15. Solé RV, Miramontes O, Goodwin BC (1993) Oscillations and chaos in ant societies.
J Theoret Biol 161(3):343–357.16. Nemes L, Roska T (1995) A CNN model of oscillation and chaos in ant colonies: A case
study. IEEE Trans Circuit Syst I 42(10):741–745.17. Li LX, Yang YX, Peng HP, Wang XD (2006) An optimization method inspired by
‘chaotic’ ant behavior. Int J Bifurcat Chaos 16(8):2351–2364.18. Peng HP, Li LX, Yang YX, Liu F (2010) Parameter estimation of dynamical systems
via a chaotic ant swarm. Phys Rev E Stat Nonlin Soft Matter Phys 81(1 Pt 2):
016207.19. Sudd JH (1967) An Introduction to the Behavior of Ants (Univ of Hull St Martin’s Press,
New York).20. Herbers JM (1983) Social organization in Leptothorax ants: Within- and between-
species patterns. Psyche (Stuttg) 90(4):361–386.21. Liang ZS, et al. (2012) Molecular determinants of scouting behavior in honey bees.
Science 335(6073):1225–1228.22. Sommer S, von Beeren C, Wehner R (2008) Multiroute memories in desert ants. Proc
Natl Acad Sci USA 105(1):317–322.23. Bonabeau E, Dorigo M, Theraulaz G (1999) Swarm Intelligence: From Natural to
Artificial System (Oxford Univ Press, Oxford).24. Chen L, Aihara K (1995) Chaotic simulated annealing by a neural network model with
transient chaos. Neural Netw 8(6):915–930.25. Aihara K (2002) Chaos engineering and its application to parallel distributed pro-
cessing with chaotic neural networks. Proc IEEE 90(5):919–930.
Li et al. PNAS | June 10, 2014 | vol. 111 | no. 23 | 8397
PHYS
ICS
ECOLO
GY

Supporting InformationLi et al. 10.1073/pnas.1407083111SI Text
Chaotic and Self-Organizing Ant BehaviorsThe first indication of deterministic chaos in insect behavior wasreported from experiments done on activity cycles in ants of thespecies Leptothax allardycei. Refuting popular fable, ants do notwork untiringly all day long. They may actually spend much timein inactivity (1). The study by Herbers on acts performed by antsof the Leptothorax longispinosus and Leptothorax ambiguus specieshas revealed that “ants spent two-thirds of their time apparentlydoing nothing at all” (2). The investigations on L. longispinosusexhibited well-defined rhythmic changes in movement activity lev-els. Cole used a solid-state automatic digitizing camera to performa careful experimental study on the dynamical behaviors of in-dividual isolated ants and an ant colony (3). The activity of numbersof individual workers was recorded and several interesting resultswere obtained:
i) The activity of the ant colony as a whole was shown to beperiodic with an activity cycle ranging from 15 to 37 min;
ii) The activity of single ants, recorded by using isolated indi-viduals, however, is not periodic. Individual ants have activitypatterns characterized by spontaneous bursts of activity fol-lowed by comparatively long periods of inactivity. As furtherindividuals are added, some synchronization begins to ap-pear, but periodic activity seems to take place only for a num-ber of workers similar to the normal colony size;
iii) Careful analysis of the experimental data revealed evidenceof low-dimensional deterministic chaos in the dynamics ofsingle ants and a periodic activity in the whole colony. Theattractor of single ant’s activity was reconstructed from theobtained time series and was shown to have a mean non-integer dimension of about 2.4, which leads itself to the con-struction of 3D phase space diagrams. The activity of the entirecolonies yielded an integer dimension.
Cole speculated that, “The existence of chaos in animal be-havior can have several important implications. Variation in thetemporal component of individual behavior may not be duesimply to chance variations in the stochastic world, but to de-terministic processes that depend on initial conditions” (3).
More Details on the Sketch Map of Fig. 1The process of each foraging is divided into two phases: One is thehunting (or finding) process and the other is the homing one. Forthe hunting (or finding) process, if the ant does not find the foodsource, then it begins the homing process at its tiring time ttired andreturns to the nest at time tnn; if it finds the food source at timetmmðtmm < ttiredÞ, then it begins the homing one at time tmm. Forthe searching process of an ant, let ZðtÞ be a point in Fig. 1A thatan ant passes in its crawling path at time t, and point Z in Fig. 1Bis mapped from point ZðtÞ in Fig. 1A. The line segment L is thedistance of the optimal path between the nest and the food source.Firstly, we consider the situation that an ant does not find the
food source in a foraging process and it begins the homing processat its tiring time. For the first foraging process of an ant, the pointin Fig. 1B is obtained as follows:
� At the hunting (or finding) time 0≤ t≤ ttired, the abscissa ofpoint Z is ðt=ttiredÞL, and the ordinate of point Z is dðtÞ, whichequals the distance from the point ZðtÞ to the line segment Lin Fig. 1A.
� At the homing time ttired ≤ t≤ tnn, the abscissa of point Z isL+ ðt− ttired=tnn − ttiredÞL, and the ordinate of point Z is dðtÞ,which equals the distance from the point ZðtÞ to the line seg-ment L in Fig. 1A.
At the n th foraging process of an ant, the ant has returned tothe nest for n− 1 times. Hence, for the n th foraging process ofthe ant, the point Z in Fig. 1B is mapped from the point ZðtÞ inFig. 1A as follows:
� At the time interval 0≤ t≤ ttired, the abscissa of point Z isð2ðn− 1Þ+ ðt=ttiredÞÞL and the ordinate of point Z is dðtÞ, whichequals the distance from the point ZðtÞ to the line segment Lin Fig. 1A;
� At the time interval ttired ≤ t≤ tnn, the abscissa of point Z isðð2n− 1Þ+ ððt− ttiredÞ=ðtnn − ttiredÞÞÞL and the ordinate of pointZ is dðtÞ, which equals the distance from the point ZðtÞ to theline segment L in Fig. 1A.
Secondly, we consider the situation in which an ant finds thefood source at time tmmðtmm < ttiredÞ in the foraging process (seethe k th path of ant i in Fig. 1B). Similarly, the point Z in Fig. 1Bis mapped from ZðtÞ in Fig. 1A as follows:
� At the time interval 0≤ t≤ tmm, the abscissa of point Z isð2ðk− 1Þ+ ðt=tmmÞÞL and its ordinate is dðtÞ, which equalsthe distance from the point ZðtÞ to the line segment L inFig. 1A;
� At the time interval tmm ≤ t≤ tnn, the abscissa of point Z isð2k− 1+ ððt− tmmÞ=ðtnn − tmmÞÞÞL and the ordinate of point Zis dðtÞ, which equals the distance from the point ZðtÞ to theline segment L in Fig. 1A.
Now, we consider the situation for the first foraging process inFig. 1 C and D, and the ant finds the food source at time tmm.The point Z in Fig. 1 C and D is mapped from ZðtÞ in Fig. 1A asfollows:
� At the time interval 0≤ t≤ tmm, the abscissa of point Z isðt=tmmÞL and its ordinate is dðtÞ, which equals the distancefrom the point ZðtÞ to the line segment L in Fig. 1A;
� At the time interval tmm ≤ t≤ tnn, the abscissa of point Z isL+ ððt− tmmÞ=ðtnn − tmmÞÞL and the ordinate of point Z isdðtÞ, which equals the distance from the point ZðtÞ to the linesegment L in Fig. 1A.
At the n th foraging process of an ant, the ant has returned tothe nest for n− 1 times. Hence, for the n th foraging process ofan ant (Fig. 1 C and D), the point Z is mapped from the pointZðtÞ in Fig. 1A as follows:
� At the time interval 0≤ t≤ tmm, the abscissa of point Z isð2ðn− 1Þ+ ðt=tmmÞÞL and the ordinate of point Z is dðtÞ, whichequals the distance from the point ZðtÞ to the line segment Lin Fig. 1A;
� At the time interval tmm ≤ t≤ tnn, the abscissa of point Z isð2n− 1+ ððt− tmmÞ=ðtnn − tmmÞÞÞL and the ordinate of pointZ is dðtÞ, which equals the distance from the point ZðtÞ tothe line segment L in Fig. 1A.
In short, the ordinate of Fig. 1 B–D is the distance from thepoint ZðtÞ to the line segment L and the abscissa of Fig. 1 B–D isthe multiple of L, where the path between two adjacent blackpoints in Fig. 1 B–D represents one foraging process for an ant.Note that different ants have different ttired, tnn, and tmm.
Li et al. www.pnas.org/cgi/content/short/1407083111 1 of 4

More Details on the Chaotic Ant Foraging ModelHere we present more detailed analyses of our model proposedin this paper. Fig. S1 shows the influences of the food sourcesize Δ on the probability of finding food for ants with differentages. Fig. S2 shows the evolution process of
��~ZðtÞ�� with dif-ferent ω. Fig. S3 gives the convergence process of the angleϕ=
����~Zi −~Pnest�T�~Pfood −~Pnest
�������~Zi −~Pnest������~Pfood −~Pnest
������,where b= logð1Þ, b= logð1:5Þ, b= logð1:8Þ, b= logð1:95Þ, b=logð1:98Þ, and b= logð2Þ, respectively. Here all of the ants havethe parameters ri = 0:2 and ω= 0:1.Ants change their foraging pattern from an initially chaotic
search, via a transient chaotic search, before finally arriving at anordered periodic movement during the whole foraging process inwhich they find the food, the nest, and the optimal path betweenthe food source and the nest. A chaotic attractor has a fractalstructure, i.e., the chaotic search of an ant meets a fractal searchpattern. Because many animals use fractal walking patterns, theforaging behavior of ants has some similarity with that of otheranimals. Through our analysis, we suggest that for the groupanimals that have a home, their behavior should not be char-acterized as random walks but as deterministic walks in randomenvironments because the animals use their intelligence and ex-perience to guide them and their behavior should be influencedby their physical abilities, their experience, and the existence ofa home.Let τðtÞ be the intensity of pheromone on the optimal path at
time t. Based on ref. 4, we have
τðt+ 1Þ= ρτðtÞ+ΔτðtÞΔτðtÞ= nnðtÞδ; [S1]
where nnðtÞ is the number of ants on the optimal path at time t, δis the quantity of pheromone that each ant laid on the optimalpath, and ρ is the evaporation coefficient.Because the variable yiðtÞ of an ant i is limited on the optimal
path, we define a threshold yp for the variable yiðtÞ. When yiðtÞ< yp,this ant i is on the optimal path. Thus, we can compute thenumber of ants whose value of yiðtÞ is less than the threshold yp.Then we can observe the evolution process of the intensity ofpheromone on the optimal path. In the finding process of theoptimal path, the intensity of the pheromone τðtÞ on the optimalpath becomes stronger and stronger as time evolves. This re-inforcement of pheromone makes the chaotic behavior of antsbecome weaker. The evaporation property of pheromone causesthe quantity of pheromone on the optimal path to increase morerapidly than on an alternative longer path. Subsequently more andmore ants to choose the optimal path and variable yiðtÞ will be-come smaller and smaller; that is, the increment process of thepheromone density on the paths is equivalent to the decrement ofthe chaotic crawling of ants.
The Impact of Two Symmetric Food SourcesWhen two sources of food exist and are relatively close, the op-timal paths from the nest would be very close to each other, whichwould lead to a symmetrical setup. Portha et al. (5) have examinedthe emergence of self-organized asymmetries in the spatial dis-tribution of ants within a symmetrical experimental setup, wheretwo food sources with a strictly symmetrical set up are considered.The relative amount of the pheromone at each branching point
of the trails determines the distribution of foragers between theforaging sectors. Due to the snowball effect of trail communication
(a positive feedback process), symmetry breakings (in which onesource is more exploited than the other) are more likely to occurfor stronger chemical trails. In this condition, positive feedbacksdrive the transition from the chaotic to the periodic foraging.For a more detailed experimental and analysis about self-
organized asymmetries, see ref. 5.
Computation of the Damage for the Lorenz SystemThe computation process of the damage extent for the continuouschaotic systems is shown in the following process. Firstly, werandomly generate the initial positions of all of the ants in the
search space. Then the initial position of ant i is ~̂θ0i =�θ̂0i1; . . . ; θ̂
0il
�and i= 1; 2; . . . ; n, where ~̂θ is the estimation of ~θ′. The searchingranges of unknown parameters are selected by Ψk, where k denotesthe search range of the k th element of a variable in search space.Based on the measurable state vector~x= ðx1; . . . ; xNÞT , we definethe following Euclidean norm objective function
f�~e�=
XWt=0
��x1ðtÞ− xKi1ðtÞ
�2+ . . . +
�xNðtÞ− xKiNðtÞ
�2�;
where t= 0; 1; . . . ;W . Thus, the problem of the computationprocess of damage extent of the dynamical system is transformedto that of using the proposed ant foraging model to search for
the suitable value of ~̂θKi =�θ̂Ki1; . . . ; θ̂
Kil
�, such that the objective
function f is globally minimized, where K is the iterative step of thechaotic ant foraging model. For each iteration step, we substitute ~̂θKiinto _~x=G
�~x;~θ′
�. Running the nonlinear _~x=G
�~x;~θ′
�, we attain the
corresponding state vector~xKi ðtÞ=�xKi1ðtÞ; . . . ; xKil ðtÞ
�T , and at eachiterative step, we need to calculate the corresponding objectivefunction f .Our example is an illustration of the well-known Lorenz system,
described by _x1 = θ1ðx2 − x1Þ, _x2 = ðθ2 − x3Þx1 − x2, and _x3 = x1x2 −θ3x3, where x1, x2, and x3 are the state variables, and θ1, θ2, andθ3 are positive constant parameters. The system is in a chaoticstate when θ1 = 10, θ2 = 28, and θ3 = 8=3. To simulate this, let theparameters of the Lorenz system after damage be θ′1 = 10:3,θ′2 = 27:8, and θ′3 = 3. We use a fourth order Runge–Kutta algo-rithm to solve the Lorenz system with a step length 0.01.Let us set the values of the proposed ant foraging model
parameters as yð0Þ= 0:999, Vd = 7:5=ð2ΨdÞ− θd, b= 2=3, H = 7,ϕ= 7:5=7, a= 2; 000, Ψ1 =Ψ2 =Ψ3 = 7:5=4, ri = 0:03+ 0:2× rand,where “rand” is a random number in the range ½0; 1�, W = 20 isthe parameter in objective function (6), and Δ= 0:005.Now we run the proposed model to estimate the parameters of
the Lorenz system after damage and compute the extent of theparameter damage. If many ants find the results, then the optimalresult is the output. If none of the ants find the food source in thesearch process, i.e., no results meet f ð~eÞ≤Δ= 0:005, then we usea star (★) to represent this case. The mean value for 10 iterationsrepresents the estimated value ~̂θ.Table S1 gives the corresponding estimation results of the
Lorenz system for 10 iterations. From Table S1, we can see thatwe achieved the estimation of the parameters after damage verywell. And the extent of damage is Γ̂=
���~θ−~̂θ���= 0:4969. Similarly,we could perform the above estimation process of a continuouschaotic system on a discrete nonlinear system.
1. Sudd JH (1967) An Introduction to the Behavior of Ants (Univ of Hull St Martin’s Press,New York).
2. Herbers JM (1983) Social organization in Leptothorax ants: Within- and between-species patterns. Psyche (Stuttg) 90(4):361–386.
3. Cole BJ (1991) Is animal behavior chaotic? Evidence from the activity of ants. Proc BiolSci 244(1131):253–259.
4. Mailleux AC, Detrain C, Deneubourg JL (2005) Triggering and persistence of trail-layingin foragers of the ant Lasius niger. J Insect Physiol 51(3):297–304.
5. Portha S, Deneubourg J-L, Detrain C (2002) Self-organized symmetries in ant foraging:A functional response to food type and colony needs. Behav Ecol 13(6):776–781.
Li et al. www.pnas.org/cgi/content/short/1407083111 2 of 4
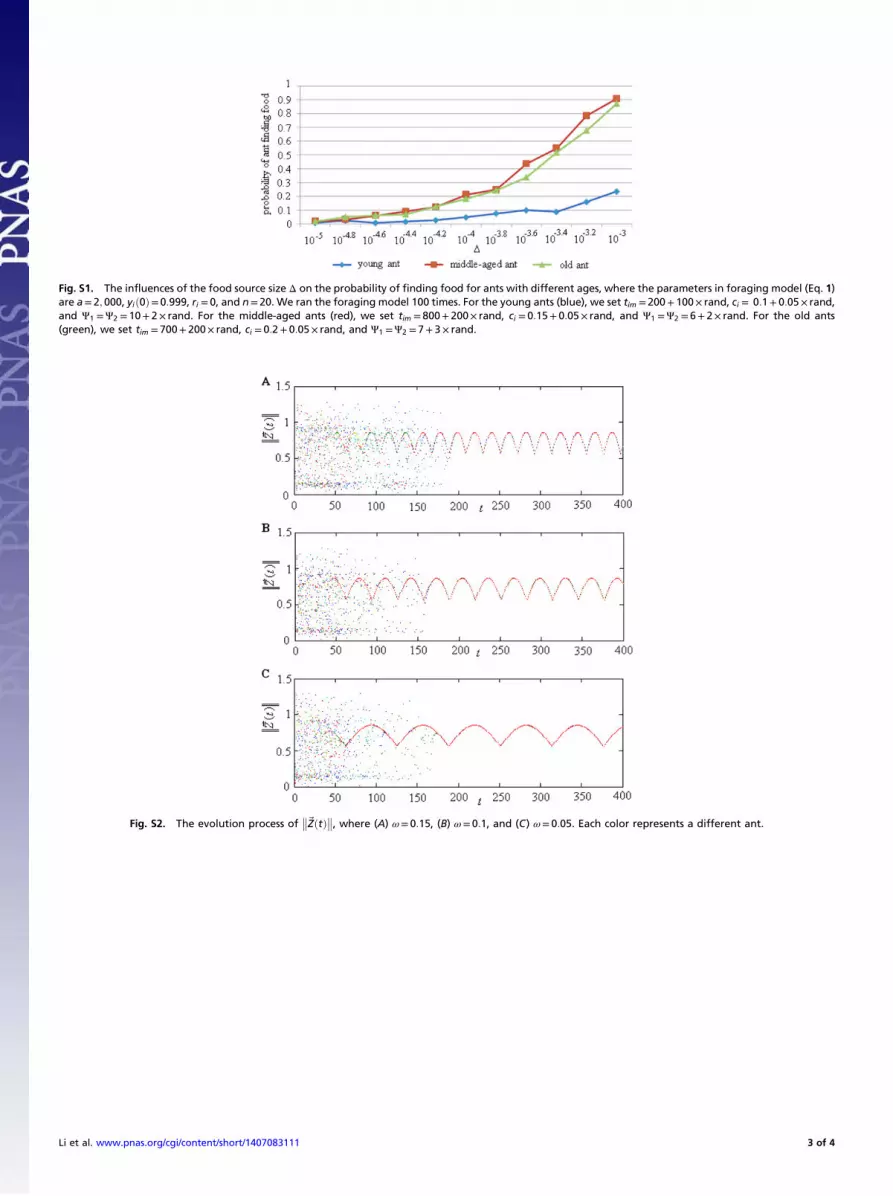
Fig. S1. The influences of the food source size Δ on the probability of finding food for ants with different ages, where the parameters in foraging model (Eq. 1)are a= 2; 000, yið0Þ=0:999, ri = 0, and n= 20. We ran the foraging model 100 times. For the young ants (blue), we set tim =200+ 100× rand, ci = 0:1+ 0:05× rand,and Ψ1 =Ψ2 = 10+ 2× rand. For the middle-aged ants (red), we set tim = 800+ 200× rand, ci = 0:15+ 0:05× rand, and Ψ1 =Ψ2 = 6+ 2× rand. For the old ants(green), we set tim = 700+ 200× rand, ci = 0:2+ 0:05× rand, and Ψ1 =Ψ2 = 7+ 3× rand.
Fig. S2. The evolution process of��~ZðtÞ��, where (A) ω= 0:15, (B) ω= 0:1, and (C) ω= 0:05. Each color represents a different ant.
Li et al. www.pnas.org/cgi/content/short/1407083111 3 of 4

Fig. S3. The convergence process of the angle ϕ=���ð~Zi −~PnestÞT ð~Pfood −~PnestÞ
�����ð~Zi −~PnestÞ����ð~Pfood −~PnestÞ
�����, where (A) b= logð1Þ, (B) b= logð1:5Þ, (C)b= logð1:8Þ, (D) b= logð1:95Þ, (E) b= logð1:98Þ, and (F) b= logð2Þ. Each color represents a different ant.
Table S1. The estimation results of parameters θ′1, θ′2, and θ′3Iterations θ̂1 θ̂2 θ̂3
1 10.2979 27.7964 3.00212 10.3076 27.7926 3.00693 10.3075 27.8021 3.00314 10.3012 27.8083 3.00345 10.3261 27.7890 3.00416 * * *7 10.3232 27.7996 2.99938 * * *9 * * *10 10.2842 27.8131 2.9995Average value 10.3068 27.8001 3.0036Average deviation 0.0068 0.0001 0.0036
* indicates that none of the ants find a food source in this foraging process.
Li et al. www.pnas.org/cgi/content/short/1407083111 4 of 4





























