Embed Size (px)
Citation preview

DEVELOPMENTAL BIOLOGY 96,23-31 (1983)
Cell Surface Proteins of Drosophila
I. Changes Induced by 20-Hydroxyecdysone
DANIEL F. WOODS AND CLI~ON A. POODRY
Department of Biology, Thimann Laboratories, lhiversity of Califmia, Santa Cmcz, California 95064
Received Februar?J 11, 1982; accepted in revised form October 5, 1982
Treatment of several ~~op~~~a cell lines with the molting hormone (20-hydroxyecdysone) resulted in biochemical
and cellular changes including the morphogeneti~ process of cell aggregation. Radiolabel~ng of the cell surface proteins
revealed 34 polypeptides that are modulated by the hormone’s action. This modulation included both expression of
“new” proteins and disappearance of preexisting polypeptides. Whereas most of the hormone-induced proteins were
lentil lectin-binding glycoproteins, only one group of disappearing proteins appears to bind lentil lectin. Labeling of
the cell surface prior to hormone addition revealed no specific modification of preexisting surface proteins which could
account for the protein changes observed with one possible exception. The potential relationship between the modulation
in surface proteins and the increase in cell-cell adhesion that occurs during hormone exposure is discussed.
INTRODUCTION
The cellular basis of morphogenesis, in particular the involvement of adhesive interactions between embryo cells, has been intensively studied in amphibian (Townes and Holtfreter, 1955) and chick (Steinberg, 1963; Mos- cona, 1974) systems. Models of cell interactions involv- ing tissue-specific cell adhesion molecules (Moscona, 1974) have been supported by reports of specific mole- cules which can either promote or inhibit aggregation of embryo cells, or can neutralize a F(ab)‘s effectiveness in inhibiting aggregation. In addition to the elegant studies on reaggregation in sponges (MeClay, 1974; for review see Miiller et al, 19’78) and aggregation in slime molds (Miiller and Gerisch, 1978; for review see Gerisch, 1977), studies of chick neural cells have revealed several specific adhesion molecules: “cognin,” a 50,000-dalton protein that promotes adhesion (Hausman and Mos- cona, 1976); cell adhesion molecule, a 140,000-dalton protein that can neutralize blocking antibodies (Rutis- hauser et a.[., 1976; Brackenbury et al., 1977; Thiery et al., 1977); and ligitin, a lO,OOO-dalton protein that can block aggregation (Jakoi and Marchase, 1979). A chick liver adhesion molecule, a glycoprotein of 65,000 to 68,000 daltons has been found which can neutralize anti-ag- gregation antibodies (Bertolotti et al., 1980; Nielson et al., 1981). Other studies have provided evidence for the involvement of specific molecules such as fibronectin (Yamada and Olden, 1978), glycosyl transferases (Rose- man, 1970), and lectins (Grabel et al., 1979) in vertebrate morphogenesis. However, the importance in vivo of the adhesion factors defined in. vitro remains an unknown and difficult problem to address in vertebrate systems.
The use of genetics, in particular temperature-sen- sitive mutations, in an organism such as Drosophila of- fers the promise of a test in viva of the developmental importance of any putative ligands or other molecules involved in intercellular interactions. However, the search for and analysis of mutants might be fraught with difficulties due to pleiotropy and the absence of a selection scheme other than for lethals. As an alter- native we have been exploring a system in which we identify a morphogenetic activity with Drosophila cells in ~~~~0, with the goal of identifying cell surface mol- ecules necessary for cellular interactions such as adhe- sion.
A number of cell lines have been established from Drosophila embryo cells (reviewed in Schneider and Blumenthal, 1978). They respond to exposure of the in- sect molting hormone, ZO-hydroxyecdysone, by under- going changes in protein synthesis (Best-Belpomme and Courgeon, 1976; Berger et aZ., 1980a), production of en- zymes (for reviews see Berger et al., 1980; Best-Bel- pomme et aE., 1980), and the formation of aggregates (Courgeon, 19’72). Savakis eC al. (1980) detected some changes in protein synthesis that occur within minutes of hormone application, but these authors found very little change in the overall pattern of synthesis. In con- trast, Silvert and Fristrom (1980) and Rick011 and Fris- trom (1983) reported that when imaginal discs were treated with ZO-hydrox~~ecdysone, over 15% of the mem- brane proteins underwent some type of change in syn- thesis. Thus, at metamorphosis, there are changes in membrane proteins, possibly cell surface proteins, con- current with morphogenetic movements involved in the evagination of imaginal discs (Fristrom and Fristrom,
OOlZ-1606/83/030023-09$03.00/O Copyright 0 1983 by Academic Press, Inc.
All rights of reproduction in any form reserved.

24 DEVELOPMENTAL BIOLOGY VOL~~~~ 96, 1983
1975). In this report we document the changes in the pattern of cell surface proteins during the aggregation response to hormone by several cell lines.
MATERIALS AND MET~~ODS
Cell Lines
The majority of the work shown was done using the Schneider L2D cell line originally isolated from late Oregon R embryos (Schneider, 1972) and kindly pro- vided by A. Blumenthal. Other lines used in this study were obtained from late embryos carrying the shibire’s’ mutation (Poodry, unpublished data) and several clones of this line, e.g., EH34A3 (Huala and Poodry, unpub- lished). All cells were maintained in Schneider’s tissue culture medium supplemented with 5% fetal bovine serum (FBS), bactopeptone, and insulin (6 mg/ml and 0.05 units/ml, respectively).
Cells were exposed to 1 X lo-” M 20-hydroxye~dysone (Sigma) for various lengths of time. In all initial ex- periments, cells were maintained in hormone for 96 hr, since this is near the exposure reported for maximal /3- galactosidase induction (Best-Belpomme et al., 1978). Nontreated cells were handled identically, except that no hormone was added.
Layering of the Cell Surface
For radiolabeling, the cells were removed from the plastic substrate by a gentle stream of medium, pel- leted, and washed once with phosphate-buffered saline (PBS). In general, lo7 cells were labeled with 200 PCi of Na-iz51 using Iodo-Gen (Pierce) as a catalyst accord- ing to the instruction of the manufacturer. The radio- labeled cells were washed in PBS, lysed in 10 mM Tris/ HCl, pH 7.6, containing 2% Nonidet-P40 (NP-40, Par- ticle Data Laboratories), and the protease inhibitors 0.1 mM phenylmethylsulfonyl fluoride (Sigma), 0.1 mg pep- statin (Sigma), and 20 KIU aprotinin (Sigma) (Hall et aZ., 1978). The cells were left in lysis buffer overnight at 4°C (approximately 12 hr), then centrifuged for 10 min at 12,000~. The pellet was extracted further with 2% sodium dodecyl sulfate (SDS) and 1% dithiothreitol (DTT). The radioactivities of the NP-40 and SDS-sol- uble fractions were determined using a Beckman 8000 gamma counter.
Approximately 4 million cpm of iodinated cell extract in 1% NP-40 were added to 100 ~1 of packed Lens cu- Zinaris agarose (3 mg/ml, Vector Laboratories) and al- lowed to incubate for 30 min. After incubation the aga- rose beads were washed several times with lectin buffer
(10 mM Tris/HCl, pH 7.8,0.14 M NaCl, 0.2% NP-40, 0.1 m&f CaC12, and 0.1 mM MgC12) to remove unbound pro- teins. The bound glycoproteins were released in lectin buffer containing 10% a-methyl-mannoside.
Gel Electrophcwesis
The radioiodinated proteins were separated using ei- ther the two-dimensional O’Farrell method (1975) or with nonequilibrium gel electrophoresis (NEPHGE) (O’Farrell et ul., 1977). Approximately 400,O~ cpm were loaded onto each gel. If the extract contained SDS, NP- 40 was added until the final ratio was 1:20 SDS/NP-40. The second-dimension gels were 10% (w/v) acrylamide (30% acrylamide and 0.8% bisacrylamide) and poly- merized using N,N,hl’~-tetramethylethylenediamine (TEMED) and 10% ammonium persulfate. All gel re- agents used were obtained from Bio-Rad with the ex- ception of the SDS (Sigma} a.nd ampholines (LKB). The subsequent gels were dried and autoradiographed using enhancing screens (DuPont) and Kodak film. At least five independent labelings and separations were carried out on hormone-treated and nontreated cells.
RESULTS
Mwphological Response of Cell Lines to 2?0- ~~dro~~ecd~s~,e
A typical response of the L2D cell line to hormone is shown in Fig. 1. The response is essentially the same as that described by Courgeon (1972) and that of our other cell lines (data not shown). The untreated cells were initially smaI1 and round (Fig. 1A). After approx- imately 12 hr of hormone stimulation, the cells became very active but showed little actual cell motility (un- published time-lapse observations). By 20 hr the cells assumed a flattened, monolayered appearance with a larger amount of cell surface in contact with the sub- strate. Twenty-four hours after addition of hormone small clumps of cells began to appear in the cell mono- layer. These cell aggregates continued to enlarge (Fig. 1B) with the addition of more cells from the monolayer (unpublished time-lapse observations) (Rosset, 1978). By 96 hr the aggregates were large enough to be easily dislodged from the substrate. These large aggregates could fuse with other cells and aggregates, producing some cell masses which exceeded 0.5 mm in diameter. Cells in the aggregates became closely apposed to one another (Fig. 1C). In addition to large areas of surface in close apposition, specialized junctions, including ad- hering zones and gap junctions, were plentiful (Figs. 1D and E). Septate junctions, typical of insect epithelia, were not seen.
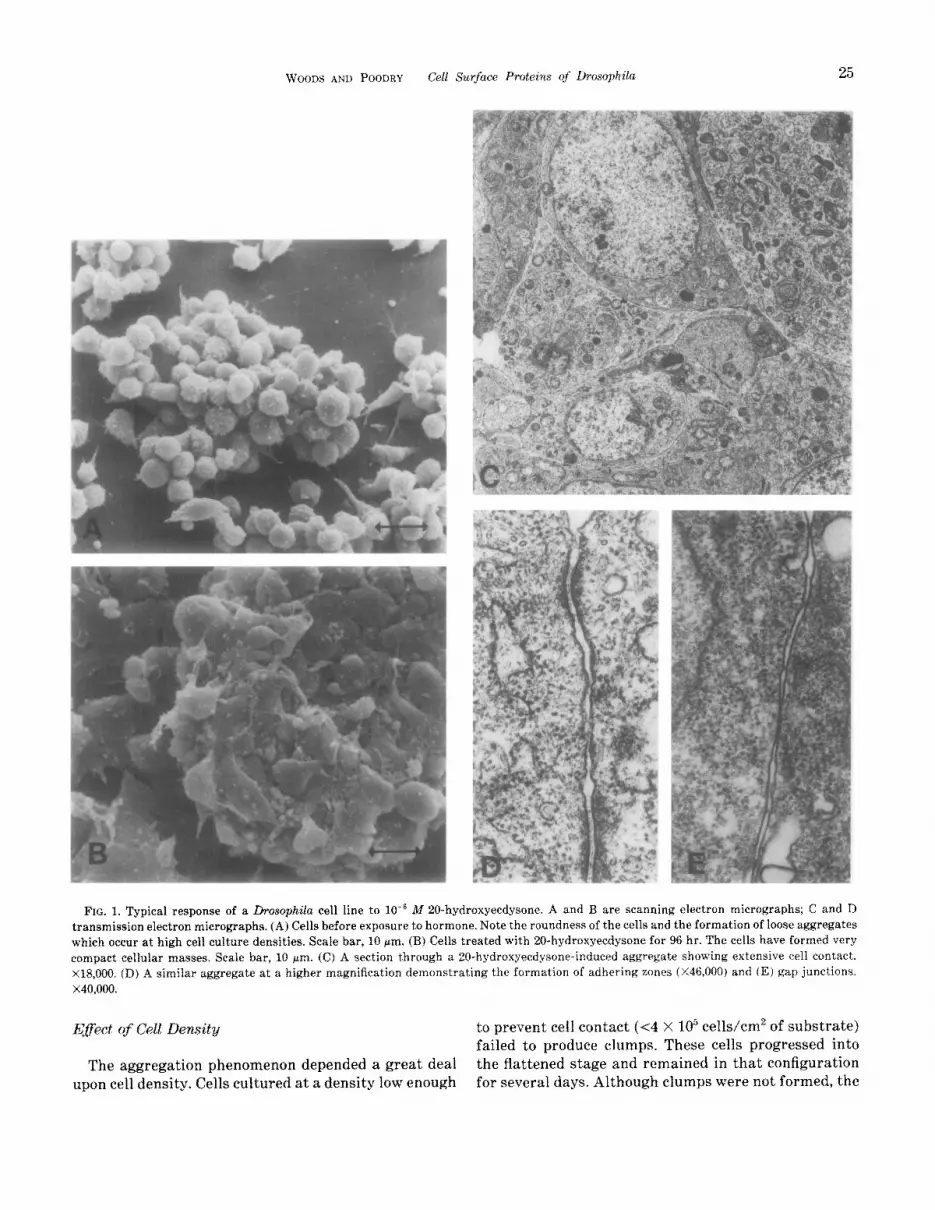
WOODS AND POODRY Cell Surfuce Proteins qf Drosophila 25
FIG. 1. Typical response of a Drosophila cell line to 10e6 M 20-hydroxyecdysone. A and B are scanning electron micrographs; C and D transmission electron micrographs. (A) Cells before exposure t,o hormone. Note the roundness of the cells and the formation of loose aggregates which occur at high cell culture densities. Scale bar, 10 pm. (B) Cells treated with 20-hydroxyecdysone for 96 hr. The cells have formed very compact cellular masses, Scale bar, 10 pm. (C) A section through a 20-h~droxye~d~sone-induced aggregate showing extensive cell contact. X18,000. (D) A similar aggregate at a higher magnification demonstrating the formation of adhering zones (X46.000) and (E) gap junctions, x40,000.
Eflect of Cell Density to prevent cell contact (~4 X lo5 cells/cm2 of substrate) failed to produce clumps. These cells progressed into
The aggregation phenomenon depended a great deal the flattened stage and remained in that configuration upon cell density. Cells cultured at a density low enough for several days. Although clumps were not formed, the

26 DEVELOF~ENT~L BIOLOGY VOLUME 96, 1933
cells still maintained the ability to aggregate. Clumps that were formed after concentrating hormone-treated low-density cultures appeared identical to clumps formed by cells maintained at a high-density through- out hormone stimulation.
Cdl Surface LabeZing of Cell Lines
The iodinated, NP-40-soluble cell surface proteins of L2D cells are shown in Fig. 2. Standard two-dimen- sional and nonequilibrium gel electrophoresis revealed over 150 labeled cell surface polypeptides. Extraction of labeled cells with 2% NP-40 solubilized over 60% of
; 30
12
*' - . II *tst92O 30 2 h ‘*
e
2 l I4 1 s t 15
16 1 $7 12
‘%V-- -- FIG. 2. Radiolabeled cell surface proteins of cells cultured without
(A) and with (B) lo-‘M ZO-hydroxyecdysone for 96 hr. Cells were iodinated and lysed overnight with 2% NP-40. Approximately 400,000 cpm from the soluble fraction were loaded onto an equilibrium iso- electric focusing gel, Separation in the second dimension was done using a 10% SDS-acrylamide gel. Proteins labeled with numbers were induced by the hormone, those with letters disappeared after hormone treatment. Starred spots represent proteins that showed no change and thus were used as reference markers to align the gels. Spot L (see Figs. 3 and 5) failed to resolve on these gels. The molecular weight standards used were myosin, phosphorylase B, bovine serum albumin, ovalbumin, carbonic anhydrase, and eytochrome c.
the bound radioactivity. Further extraction of the NP- 40 insoluble pellet with 2% SDS and 1% DTT solubilized approximately 90% of the remaining counts. While the SDS fraction revealed several proteins not observed in the NP-40”soluble fraction, no differences were seen be- tween hormone-treated and nontreated cells in this sub- set (data not shown). The starred spots on Fig. 2 point out spots which showed no change during hormone stimulation and serve as reference markers to align the gels. The majority of proteins present on the cell surface had an apparent pI of between 6.5 and 3.6 and a mo- lecular weight above 45K. This pattern of spots has been observed with only slight differences on every cell line we have studied thus far and on embryonic cells isolated from late gastrula.
Labeling with 125 I using Iodo-Gen as a catalyst ap- peared to be specific for cell surface components. Actin (pl 5.77-5.84, MW 44K), a major cytoplasmic protein, was not detected on our autoradiographs (cf. Fig. 2). Trypsinization (2.5 mg trypsin/ml) for 5 min at 22°C removed greater than 86% of the bound lz51 and essen- tially all of the radioactive proteins, as judged by au- toradiography of electrophoresed cell extracts. Labeling of the culture medium revealed no overlap between Dro- sophila cell surface proteins and proteins in the culture medium. Furthermore, cells freshly prepared from dis- persed embryos had a similar pattern to that of iodin- ated proteins (Sater et aE., in preparation). Therefore, the peptides we resolved were cell surface proteins, as opposed to internal proteins or proteins absorbed from culture medium.
Stimulaticm
Several differences were noted by comparing hor- mone-treated cells with untreated cells. At least 34 spots underwent reproducible changes after incubation in hormone. Twenty-two spots showed a quantitative in- crease in the amount of label (Fig. 2). The majority of these spots appeared in groups with similar molecular weight and PI. Concurrent with the increase in label of several proteins, the label in other proteins decreased (Fig. 2). This group included a minimum of 11 distinct spots. In contrast to the spots that increased in label, decreasing labeled proteins consisted of single unre- lated spots with the exception of a group labeled A-D (see Fig. 2A). NEPHGE gels revealed 1 additional spot that increased in label after hormone exposure (data not shown).
Sequence of Appearance of Cell Surface Proteins
A comparison of cells after various durations of hor- mone exposure revealed sequential changes of proteins

WOODS AND POODRY Cell Surface Proteins of Drosophila 27
on the cell surface. By 8 hr (before any observable mor- phological changes) a noticeable change had occurred on the cell surface. Several proteins, and in particular one labeled E on Fig. 2, had already started to decline in the relative amount of label, whereas no new or in- creased relative amounts of proteins were observed in these samples. After 20 hr of 20-hydroxyecdysone stim- ulation, i.e., just prior to the first observable aggrega- tion, proteins labeled in letters (with the exception of I) have decreased in the relative intensity of label. Sam- ples from longer exposures of hormone, i.e., up to 96 hr, showed essentially the same decrease in intensity as well as the reduction in the spot labeled I. A few spots (l-3, 12-17, 21-22; cf. Fig. 2B) showed a moderate in- crease by 20 hr. Forty-eight hours after hormone treat- ment the pattern of proteins from the cell surface was essentially identical to that seen on 96 hr gels, with the exception of the spot labeled 5 in Fig. 2B. At that time, a spot labeled 24 was present which had a very similar molecular weight, but a more basic ~1. In summary, most of the proteins that decreased in response to 20- hydroxyecdysone did so by 20 hr of exposure, whereas those that appear in response to hormone were present by 48 hr.
Effect of Cell Contact cm the Cell Surface
Cells cultured in the presence of 20-hydroxyeedysone for 96 hr at low density (4 X lo5 cells/em2 of substrate) lacked several of the proteins we have associated with hormone stimulation. Spots 7 through 14 (Fig. 3) all failed to show the typical quantitative increase after hormone addition. Since aggregation normally occurs
FIG. 3. Radiolabeied cell surface proteins from cells cultured at low cell density (see text) with lo-” M ZO-hydroxyecdysone for 96 hr. The labeled cells were handled as described in Fig. 2. Closed triangles point to the spots that are induced by hormone in both high- and low- density cells. Open triangles point to regions of the gel where spots are present in gels from high-density hormone-treated cells but are absent in low-density cells (cf. Fig. 2). The open circle indicates spot L.
before these proteins increase (see previous section), cell contact may be necessary to induce an accumulation or expression of these proteins. All of the other mod- ulated proteins, except a spot labeled L that normally disappeared, underwent changes comparable to those occurring in cells treated at high density.
Glycoproteins on the Cell Surface
Studies of the cell surface of other organisms have revealed that a signi~cant fraction of the external mem- brane proteins are glycoslyated (Bretscher, 1974; Tweto et al., 1976). Examination of the cell surface of Dro- sophila using lectin affinity chromatography has shown that most of the glycoproteins bound to Lens cu~i~a~.~ agglutin (LCA) (Woods and Poodry, in preparation). We used agarose-coupled LCA to affinity purify 12”i-labeled cell surface proteins. All but one of the hormone-in- duced proteins bound the LCA (see Fig. 4B). In contrast, only four (A-D) of the disappearing spots seemed to bind LCA (Fig. 4A). Other evidence based on both met-
8. s
-IEF-
FIG. 4. Lectin affinity chromatography of radiolabeled cell surface proteins from cells cultured without (A) and with (B) 20-hydroxyec- dysone. The NP-40 soluble fraction of the labeled cells was incubated with agarose bound Lens CT&W% and the bound proteins released with ~-methyl-mannoside. Open arrows indicate disappearing spots and closed arrows indicate increasing spots after hormone exposure.

28 DEVELOPMENTALBIOLOGY
abolic labeling and iectin binding (Woods and Poodry, in preparation) indicated that the modulated proteins that do not bind LCA were only slightly, if at all, gly- eosylated.
Several lines of evidence (Weller, 1979; Shur and Roth, 1973) point to modifications of preexisting proteins (i.e., glycosylation or phosphorylation~ during morphogen- esis. These modifications could cause changes in either isoelectric point or molecular weight leading to the type of changes observed after hormone treatment. To test this hypothesis, cells were labeled with lz51 prior to hormone addition. Prelabeled cells were then incubated for approximately 20 hr or until a noticeable morpho- logical change, i.e., flattening, had occurred in the hor- mone-treated cells. Three polypeptides were found that underwent specific changes (Fig. 5). One protein in- creased in molecular weight by approximately 4 kilo- daltons (spot 25). Although this change in molecular weight is slight, it was reproducible in gels analyzed from three independent prelabeling experiments. Spot 25 also appeared to be cell line specific since it was only present on EH34A3 cells. While the remaining proteins failed to reveal any shifts in molecular weight or PI, two proteins, E and L, showed a change in the rate of loss from the cell after hormone addition (Figs. 5C and D). Proteins E and L would normally show a decrease after hormone stimulation (Fig. 5B).
IXSCUSSION
Treatment of several ~~osoph,~Z~ cell lines with 20- hydroxyecdysone results in a dramatic sequence of mor- phological changes in the cells involved in the apparent differentiation of the cells (Cherbas et al., 197’7). One of the overt responses is an increase of cell to cell adhesion, as is demonstrated by the formation of aggregates (Courgeon, 1972). A likely correlate in Guo, considering the hormonal conditions to which the cells are exposed, might occur at pupation. Assuming at least some sim- ilarity between pupation and response of the cell line to molting hormone, the cellular adhesion we observe may be related to the vast amounts of cell rearrange- ment and morphogenesis occurring in the initial stages of metamorphosis (Fristrom and Fristrom, 1975). At the ultrastructural level, aggregates have some simi- larities to epithelial tissue such as a close apposition of cell membranes and the junctions formed. Imaginal an- lagen, which include the imaginal discs and precursors of the gut, are examples of epithelial tissues which irk vivo undergo rapid changes in function, shape, and cell
VOLUME 96.1983
c FIG. 5. A portion of an autoradiograph illustrating cell surface pro-
teins radiolabeled prior to 20-hydroxyecdysone addition. Cells were labeled with ‘%I then divided into 3 samples. One sample was lysed immediately while the other two samples were cultured with or with- out hormone for 20-24 hr before lysis. A fourth sample was labeled after exposure to 20-hydroxyecdysone for 72 hr. (A) Control cells (lysed immediately). (B) Hormone-treated cells labeled after exposure to hormone. (C) Cells cultured for 24 hr without hormone. (D) Proteins from cells that were cultured with hormone for 24 hr. The arrows point to three proteins that underwent some type of change after hormone exposure.
contact in response to ecdysone in preparation for pu- pation (Fristrom and Fristrom, 1975).
Labeling of the cell surface by radioiodination re- vealed a large number of polypeptides. A sizable frac- tion (34 of the 150 repeatably labeled proteins) under- went some type of modulation in response to hormone, such as a quantitative increase or decrease. In com- parison with earlier studies which detected new protein synthesis (Savakis et al., 1980), surface iodination re- vealed a lo-fold increase in the number of proteins that underwent change after the addition of 20-hydroxyec- dysone. Synthesis of cell surface proteins undoubtedly involved only a small fraction of the total metabolic activity in a cell. This factor may explain the failure of previous studies on cultured cells to detect the same dramatic changes. In addition, we have used metabolic

WOODS AND POODKY Cell Surfuce Proteins qf Drosophila 29
labeling with tritiated glucosamine or tritiated man- nose to focus on glycoproteins and have detected similar modulations in proteins occupying corresponding pos- tions in our two-dimensional gels (data not shown).
The difference in the pattern of cell surface proteins observed between hormone stimulated and nonstimu- lated cells may be explained in several ways. First, some proteins may cease synthesis and be slowly lost from the cell surface due to membrane turnover, while others increase in synthesis; therefore we detect a quantitative decrease or increase in relative labeling. Metabolic la- beling with [3H]glucosamine has verified that at least some glycoproteins undergo changes in incorporation and/or turnover (unpublished observations). Second, the changes in the amount of label may reflect accessibility of the protein to iodination. Proteins may change in their ability to be expressed on the cell surface due to final processing and yet show no change in the rate of synthesis. It is also possible that a conformational change could reveal new iodinatable sites or cover up old ones. Third, hormone stimulation may allow only specific populations of cells to divide while stopping cell division or causing cell death in other cell types. This last possibility seems rather unlikely since a clonal cell line gives the expected protein response after hormone treatment. We currently favor the interpretation that most of the change in patterns of labeled proteins re- flects a quantitative change in the amount of those pro- teins at the cell surface.
The large amount of change in the cell surface pro- teins of hormone-treated cells parallels the changes in membrane proteins reported by Silvert and Fristrom (1980) and Rick011 and Fristrom (1983). These authors described modulations in the various components (cy- toplasmic, ribosomal, chromatin, and membrane pro- teins) of imaginal discs and found that with the excep- tion of the membrane fraction, very little change is ob- served after stimulation of the disc with molting hormone. When the membrane fraction was examined, approximately 15% of the proteins showed some type of change, including both disappearance and appear- ance, a finding consistent with our data.
The vast majority of the proteins induced by ZO-hy- droxyecdysone bind lentil lectin and therefore may have a high mannose-type glycosylation. However, those pro- teins that disappear, with the exception of spots A-D, (Fig. 4) do not bind to lentil lectin. Thus the changes in cell surface lectin binding sites reported by Meta- kovski et nl. (1975) to be associated with hormone stim- ulation have been confirmed and are now recognized as changes in specific glycoproteins. The significance of this difference in glycoslyation is unclear but it should fa- cilitate purification of hormone-induced proteins using
lentil lectin and thus lead to the further characteriza- tion of these proteins.
Examination of the cell membrane during hormone exposure revealed a changing pattern of proteins on the cell surface. Eight hours after hormone addition, before any observable morphological change, the level of sev- eral proteins had already begun to decrease from the cell surface but new ones had not yet appeared. The first change in cell behavior and morphology that occurs soon after 8 hr could conceivably require a loss of pro- teins rather than the appearance of new proteins on the cell surface. Proteins lost after exposure to 20-hy- droxyecdysone could be involved in maintaining the cy- toskeletal network which undergoes modification dur- ing this period (Berger et al. 1980b; Locke and Huie, 1981). Just prior to the initial aggregation process (20 hr) all but one of the proteins destined to reduce in intensity had disappeared while several of the hor- mone-induced proteins showed a slight increase. If some
of the new proteins are invotved in the aggregation pro- cess then they should already be present. The new pro- teins that are absent at this stage are most likely not related to aggregation per se. In fact, these proteins overlap with the group of proteins that are absent from low-density cells. Thus, the proteins appear to result from secondary events, including cell contact and not from hormone stimulation. Several other systems have demonstrated a similar contact dependence for expres- sion of certain proteins (e.g., Alton and Lodish, 1977).
We examined the possibility that the hormone-in- duced changes we observe are actually the modification of preexisting proteins by labeling cells prior to hor- mone application. Only one indication of protein mod- ification was found. That is, there was generally no shift in pl or molecular weight of prelabeled proteins in re- sponse to hormone. A single protein (Fig. 5, protein 25) exhibited an apparent increase in molecular weight of about 4 kilodaltons.
With the exception of the elegant work done on re- ceptor proteins (Bretscher et al., 1980), prevailing the- ories of membrane turnover postulate that the mem- brane is degraded with no selective loss of any partic- ular component (e.g., Tweto and Doyle, 1977). Our results point to some type of selective control over the rate of loss of two proteins from the cell surface which nor- mally disappear after hormone exposure. However, sev- eral independent prelabelings reveal that not only does 20-hydroxyecdysone induce preferential loss of a com- ponent that normally disappears (Fig. 5, protein E) but also seems to increase the lifetime of a normally dis- appearing protein, L (Fig. 5). One way to explain this seemingly contradictory behavior of protein L is to pos- tulate that hormone stimulation results in the forma-

30 DEVELOPMENTAL BIOLOGY VOLUME 96, 1983
co 93 P co 66
a 45
6.5
20 0 cl 12
FIG, 6. Summary diagram of radiolabeled cell surface proteins and their modulation by 20-hydroxyecdysone (0) disappearing proteins; (@) proteins induced by hormone; (0) reference proteins; (0) glycoproteins; (4) proteins which failed to appear at low cell densities; (a) proteins which failed to disappear at low cell densities. Proteins are grouped and labeled based on the earliest time at which some type of change was observed (in hours after hormone addition). The molecular weight standards used were myosin, phosphorylase B, bovine serum albumin, ovalbumin, carbonic anhydrase, and cytochrome c.
tion of some type of stable complex involving L in such a way as to make it less accessible to iodination. This could also explain why L failed to disappear on low- density cells treated with hormone (Fig. 3). Therefore the “disappearance” of L may be due to conformational changes and not through actual loss from the cell sur- face. Further studies using a metabolic label should re- solve this problem.
Our main focus has been in identifying the protein(s) involved in the aggregation process. To this end we have demonstrated that a large number of proteins on the cell surface are modulated by the molting hormone (Fig. 6). Other data (Poodry et al., in preparation) would in- dicate that at least some of the aggregation-mediating components are proteins. However, several possible ex- planations exist for the aggregation event which need not rely on adhesion mediated through protein changes on the cell surface. One of these is a modification in the lateral mobility of certain proteins in the membrane. We already know that nontreated cells show some at- traction for each other but fail to form stable contacts. A change in the mobility of an adhesion factor could
conceivably be necessary for the formation of stable contacts over large areas of the cell membrane, without any concomitant change in surface proteins. To circum- vent this problem, we are now testing monoclonal an- tibodies that block the adhesion process. Using these antibodies we hope to identify precisely which mole- cules are involved in the aggregation event. Such in- formation will be a major step toward enabling us to use genetics to analyze the function of these cell surface proteins i?l sitti.
The authors would like to acknowledge the excellent technical as- sistance of Beverly L. Quon. We would also like to thank Dr. Patricia Calarco for her comments. We are especially grateful to Amy K. Sater for her help in both the experiments and writing of the manuscript. This research was supported by Grants GM 20401 and RR 08132 from the NIH.
REFERENCES
ALTON, T., and J&DISH, H. (1977). Synthesis of developmentally reg- mated proteins in D, dise~d~~ which are dependent on continued cell-cell interaction. Den Bid. 60, 207-216.
BERGER, E., FRANK, M., and ARELL, M. C. (1980). Ecdysone-induced changes in protein synthesis in embryonic Drosophila cells in cul-

WOODS AND POoDRy Cell 6 lurfuce Proteins of Drosophila
ture. In “Invertebrate Systems In Vitro” (E. Kurstak, K. Mara- morosch, and A. D~bendorfer, eds.) pp. 195-208. Else~~ier/North- Holland, Amsterdam.
BERGER, E., IRELAND, R., and WYSS, L. (1980a). Pattern of peptide synthesis in Drosophila cell lines and their hybrids. Static Cell Gen. 6, 719-729.
BERGER, E. M., SLOBODA, R. D., and IRELAND, R. C. (1980b). Tubulin content and synthesis in differentiating ~os~~h~~a cells in culture. CelE Motil 1, 113-129.
BERTOLOTTI, R., RUTISHAUSER, U., and EDELMAN, G. M. (1980). A cell surface molecule involved in aggregation of embryonic liver cells. Proc. Nut. Acad &i USA 77, 4831-4835.
BEST-BELPOMME, M., and COURGE~N, A.-M. (1976). Inductions prot- eiques par l’ecdysterone dans des clones cellulaires de Drosophila
melano.gaster cultives i?& vitro. C. R. Acud Sci. 282 Ser. D, 469-471. BEST-BELPOMME, M., CO’(IRCIEON, A.-M., and ECHALIER, G. (1980). De-
velopment of a model for the study of ecdysteroid action: Drosophila ~e~u~o~u~~e~ cells established in vitro. In “Progress in Ecdysone Research” (J. Hoffman, ed.), pp. 379-392. Elsevier/Nor~h-Holland, Amsterdam.
BEST-BELPOMME, M., CO~JR~EON, A.-M., and RAMBACH, A. (1978). fl- Galactosidase is induced by hormone in Drosophila lnelanogaster cell cultures. Proc. Nat. Rcud Sci. USA 75, 6102-6106.
BRACKE~~URY, R., THIERY, J.-P., RUTISHAUSER, U., and EDELMAN, G. M. (1977). Adhesion among neural cells of the chick embryo. I. An immunological assay for molecules involved in cell-cell binding. J. Biol. Chem. 252, 6835-6840.
BRETSCHER, M. S. (1974). Some aspects of membrane structure. 17~ “Perspectives in Membrane Biology” (0. S. Estrada and C. Gitler, eds.), pp. 3-24. Arademic Press, New York.
BRETSCHER, M. S., THOMSON, J. N., and PEARSE, B. M. F. (1980). Coated pits act as molecular filters. Proc. Nat. Acud. Sci. USA 77, 4156- 4159.
CHERBAS, P., CHERBAS, L., and WILLIAMS, C. M. (1977). Induction of acetylcholinesterase activity by ,%ecdysone in a Dr~~hila cell line. Scieme 197, 275277.
COURGEON, A.-M. (1972). Action of insect hormones at the cellular level. Morphological changes of a diploid cell line of Drosophila melmogaster treated with ecdysone and several analogues in vitro. Exp. Cell Rex 74, 327436.
FRISTROM, D., and FRISTROM, J. W. (1975). The mechanism of evagi- nation of imaginal discs of Drosophiln melanogaster. General con- siderations. Dew. Biol. 43, l-23.
GERISCH, G. (1977). Membrane sites implicated in cell adhesion: their deveiopmental control in ~~t~oste~~~rn discoideum. In “Interna- tional Ceil Biology 1976-1977” (B. Brinkley and K. Porter, eds.), pp. 36-42. Rockefeller Univ. Press, New York.
GRABEL, L., ROSEN, S., and MARTIN, G. (1979). Teratocarcinoma stem cells have a cell surface carbohydrate-binding component impli- cated in cell-cell adhesion. Cell 17, 477-484.
HALL, L. M., VON BORSTEL, R. W., OSMOND, B. C., HOELTZLI, S. D., and HUDSC)N, T. H. (19781. Genetic variants in an acetylcholine receptor from Drosophila melanogaster. FEBS Lett. 95, 243-246.
HAUSMAN, R. E., and MOSCONA, A. A. (1976). Isolation of retina-spe- cific cell-aggregating factor from membranes of embryonic neural retina tissue. Proc. &t. Acad. Sci GSA 73, 3594-3598.
JABOI, E., and MARCHASE, R. (1979). Ligatin from embryonic chick neural retina. J. Cell Biol. 80, 642-650.
LOCKE, M., and HUIE, P. (1981). Epidermal feet in insect, morphogen- esis. Nature (Londrm) 293, ‘733-735.
MCCLAY, D. (1974). Cell aggregation: Properties of cell surface factors from five species of sponge. J. Exp. Zod. 188, 89-102.
METAKOVSKII, E. V., CHERDANTSEVA, E. M., and GVOZDEV, V. A. (1977). Action of ecdysterone on surface membrane glycoproteins of Dro- sophila melanogaster cells in culture. lMoL Biol. 11, 158-170.
31
METAKOVSKII, E. V., KAKPAROV, V. T., and GVOZDEV, V. A. (1975). Effect of ecdysterone on subcultured cells from ~oso~~~~~ mela- nogaster. Stimulation of high molecular weight polypeptide syn- thesis and change in cell surface properties. Dokl Akad Nauk SSR 221, 960-963.
MOSCONA, A. A. (1974). Surface specification of embryonic cell:lectin receptors, cell recognition and specific cell ligands. In “The Cell
Surface in Development” (A. A. Moscona, ed.), pp. 67-99. Wiley, New York.
MILLER, K., and GERISCH, G. (1978). Specific glycoprotein as target site of adhesion-blocking fab in aggregating Dictgosteliwm cells. Nature (LcMzdoni 274, 445-449.
M~LLEK, W. E. G., MYIOLLEK, I., and ZAHN, R. K. 11978). Aggregation in sponges. Res. Mot. Biol. 8, 7-87.
NIELSON, L., PITTS, M., GRADY, S., and M~G~JIRE, E. (1981). Cell-cell adhesion in the embryonic chick: partial purification of liver adhe- sion molecules from liver membranes. Dev. Bid 86, 315-326.
O’FARRELL, P. H. (1975). High resolution two-dimensional gel elec- trophoresis of proteins. J. Viol. Chem. 250, 4007-4021.
O’FARRELL, P. Z., GOODMAN, H. M., and O’FARRELL, P. H. (1977). High resolution two-dimensional electrophoresis of basic as well as acidic proteins. Cell 12, 1133-1142.
RICKOLL, W. L., and FRISTROM, J. W. (1983). The effects of 20.hy- droxyeedgsone on the synthesis of membrane proteins in ~os~hiZu imaginal discs. De?:. Biol, 95, 275-287.
ROSEMAN, S. (19’70). The synthesis of complex carbohydrates by mul- tiglycosgltransferase systems and potential function in intercel- lular adhesion. Chem. Phys. Lipilzs 5, 270-299.
ROSSET, R. (1978). Effects of ecdysone on a ~os~ph~Lu cell line. Exp. Cell Ray. 111, 31-36.
RUTISHAUSER, U., THIERY, J.-P., BRACKENBURY, R., SELA, B.-A., and EDELMAN, G. M. (1976). Mechanisms of adhesion among cells from neural tissues of the chick embryo. Proc. Nut. Acad. Sci. USA 73, 577-581.
Sxvhers, C., DEMETRI, G., and CHERBAS, I’. (1980). Ecd~steroid-in- dueible polypeptides in a Drosophila cell line. Cell 22, 665-674.
SCHNEIDER, I. (1972). Cell lines derived from late embryonic stages of Drosophila melanogaster. J. Embryol. Exp. Morph01 27, 353-365.
SCHNEIDER, I., and BLUMENTHAL, A. B. (1978). Drosophila. cell and tissue culture. In “The Genetics and Biology of ~~soph~Za” (M. Ashburner and T. R. F. Wright, eds.) Vol. Za, pp. 266-305. Academic Press, New York.
SHUR, B., and ROTH, S. (1973). Localization and potential function of glyeosyltransferase in chick embryos. Amer. ZooE. 13, 1129-1135.
SILVERT, D., and FRISTROM, J. (1980). Biochemistry of imaginal discs: Retrospect and prospect. §. Biodem. 10,341-355.
STEINBERG, M. S. (1963). Reconstruction of tissues by dissociated cells. Science 141, 401-408.
THIERY, J.-P., BRACKENBURY, R., RUTISHATJSF,R, U., and EDELMAN, G. M. (1977). Purification of a cell-adhesion molecule for neural retina. J. Biol. Chem. 252,6841-6845.
TOWNES, P. L., and HOLTFRETER, J. (1955). Directed movements and selective adhesion of embryonic amphibian cells. J. Exp. Zool. 128, 53-120.
TU’ETO, J., and DOYI~E, D. (1977). Turnover of proteins of the eukary- otic cell surface. 1~ “Cell Surface Reviews” “The Synthesis, Assem- bly, and Turnover of Cell Surface Components” (G. Poste and G. Nieolson, eds.), vol. 4, pp. 137-163. Elsevier/North-Holland, Am- sterdam.
TWETO, J., FRIEDMAN, E., and DOYLE, D. (1976). Proteins of the hep- atoma tissue culture cell plasma membrane. J. Suprastmcct. 4,141- 149.
WELLER, E. M. (1979). “Protein Phosphorylation.” Pion, Ltd., London. YAMADA, K. M., and OLDEN, K. (1978). Fibronectins-Adhesive gly-
coproteins of cell surface and blood. Nature (London) 275, 1’79-184.





























