Embed Size (px)
Citation preview

Enzyme and Microbial Technology 36 (2005) 399–408
Cell cultures ofAjuga reptansL. to bioconvert emodin andaloe–emodin: an HPLC/ESI/MS investigation
Mulinacci Nadiaa,∗, la Marca Giancarloa, Innocenti Marziaa, Vincieri FrancoFa,Crespi-Perellino Nicolettab, Minghetti Anacletob
a Dipartimento di Scienze Farmaceutiche, Universita degli Studi de Firenze, via Ugo Schiff 6,50019 Sesto Fiorentino, Firenze, Italy
b Istituto di Ricerche Biotecnologiche, via Lago di Tovel 7, Altavilla Vicentina VI, Italy
Received 21 October 2003; accepted 18 March 2004
Abstract
In this paper, we describe the bioconversion of anthraquinones, emodin and aloe–emodin, carried out by cell cultures ofAjuga reptansL. Theo PLC/DADa ides wereo n with1 ides, werea s, resultedu©
K
1
ptcahptst
cge
les.ulesjuga
ge-ty ofolicwere
activeinggt com-
valu-that
lines
on-
ion
0d
ptimal biotransformation times ranged between 48 and 72 h of contact, and the % yields of bioconversion were near 35–38%. The Hnd ESI/MS analyses were mainly applied to perform a preliminary characterisation of the bioconverted molecules. Five glycosbtained from emodin, the main one being 6-O-�-malonyl galactoside; eight different compounds were obtained from aloe–emodi1-O-�-malonyl galactoside as predominant derivative. Among the other minor derivatives, diglycosides as such as acyl glycoslso detected. Finally, the total content of the phenyl propanoidic compound, the main natural metabolites of these cell culturenmodified by the addition of both the anthraquinones.2004 Elsevier Inc. All rights reserved.
eywords: Bioconversion; Cell culture; Emodin; Aloemodin;Ajuga reptans
. Introduction
In vitro plant cell cultures are employed for secondaryroduct biosynthesis for pharmaceutical and food applica-
ions and to biotransform appropriate precursors by spe-ific enzymatic reactions such as glycosilation, oxidationnd acylation[1]. The selective glycosilation of phenolicydroxyl groups by chemical means involves complicatedrotection–deprotection procedures and it is rather difficult
o obtain good yields from this procedure. Therefore, the pos-ibility of performing this reaction by means of biologicalools could be of interest for several applications.
Among the secondary metabolites of theAjuga reptansL.ell cultures, monoterpenoids, diterpenoids and extensivelylycosilated phenylpropanoids were found[2–4]. The pres-nce of these structures suggested these plant cell cultures as
∗ Corresponding author. Tel.: +39 055 2757288; fax: +39 055 240776.E-mail address:[email protected] (M. Nadia).
a potential tool for the glycosilation of exogenous molecuPrevious investigations carried out on flavonoidic molec[5,6] and on vanillin added as exogenous substances to Acell cultures, confirmed their ability to glycosidate exonous substrates. With the aim of evaluating the capabilithese cultures to carry out specific glycosilation on phensubstrates, also 1,4 and 1,8-dihydroxy-anthraquinonesconsidered. The 1,8-anthraquinone glycosides are theprinciples of herbal drugs with cathartic activity, belongto the Rhamnus, Aloe and Rheum genera[7–9], and amonthese compounds emodin and aloe–emodin are the mosmon aglycones.
Moreover, several recent studies were focused on eating the anticancer activity of emodin and aloe–emodinhave shown interesting results with human cancer celle.g. from liver and lung[10–12].
In this paper, we describe for the first time the biocversion of emodin and aloe–emodin by cell cultures ofAjugareptansL. with data relating to the optimal biotransformat
141-0229/$ – see front matter © 2004 Elsevier Inc. All rights reserved.
oi:10.1016/j.enzmictec.2004.03.029
400 M. Nadia et al. / Enzyme and Microbial Technology 36 (2005) 399–408
time and bioconversion yields. The chemical structure of theprincipal compounds is also described mainly by HPLC/DADanalysis and ESI/MS investigations.
2. Experimental
2.1. Materials
All solvents used were HPLC grade; CH3CN and MeOHfor HPLC were from E. Merck (Darmstadt, Germany), whilewater was from Baker (J.T. Baker, Italy).
For the NMR analysis the solvents used were CD3ODand DMSO from Merck (Darmstadt, Germany). All the an-thraquinone molecules were purchased from Extrasynthese(Lion-Geney, France).
2.2. Cell culture preparation
Callus cultures ofAjuga reptansL. were started from seedsof plants grown in the Botanical Garden of Casola Valsenio(Ravenna, Italy). Cells were grown on a modified Gamborgmedium containing naphthalene acetic acid 1 mg/L, kinetin1 mg/L and 2,4-dichloro-phenoxy acetic acid 0.2 mg/L (G5medium). Solid medium contained 7 g/L of agar. Suspendedc ells,g ing5 akera s-t resw solu-t
di
2
ons,s every2 askst ef-f plesw holics mi-n weret
2
ea-s w-i
%
The area values, expressed in mAu, both for emodin andaloe–emodin, were calculated at 430 nm corresponding to arelative maximum in the range 240–550 nm. Each datum isthe average of two determinations performed in a parallel wayduring the same experiment; % CV within 5% were obtained.
2.5. Extraction procedures
To concentrate the bioconverted products both for emodinand aloe–emodin, all supernatants from different times werecombined and filtered obtaining about 1.2 L of hydro alco-holic solution. These were evaporated up to water (about500 mL) and extracted with 4× 50 mL of CHCl3 to re-cover the residual free aglycone. The next step (developedto concentrate the anthraquinone molecules) was the ex-traction with 7× 50 mL of ethyl acetate. The total intenselyyellow-coloured extracts were then concentrated under vac-uum (28–30◦C) and collected with H2O/EtOH 1:9 (2 and4 mL) and used for semi-preparative HPLC.
2.6. HPLC analysis
The analyses were carried out using a HP 1090Lliquid chromatograph equipped with a DAD detector(Hewlett & Packard, Palo Alto, USA). The column was a4 -s a4 ntsw )C ed:4 ,( (A)a ally4 ysis5
dsw witha lk,
cules.
ultures were prepared by transferring about 2 g of crown on solid G5 medium, to 300 mL flasks contain0 mL of G5 liquid medium and shaken on a rotator sht 120 rpm at 28◦C. After 1 week, 5 mL portions were di
ributed into other identical flasks. Five days later cultuere supplemented with 2 mL of a sterile 50% sucrose
ion.All the anthraquinones (100�g/mL), previously dissolve
n 0.5 mL of MeOH, were added to 11-day-old cultures.
.3. Evaluation of the optimal biotransformation times
For time course determination of the biotransformatiamples for analysis were taken in the range 0–120 h,4 h. Then an equal volume of MeOH was added to the fl
o block enzymatic activities. In order to avoid browningects, 1 mg/mL of ascorbic acid was also added. The samere then homogenised and filtered and the hydro alcoolutions were directly analysed by HPLC/DAD. Deterations were carried out in duplicate and mean values
aken obtaining % CV values not over 5%.
.4. Evaluation of the biotransformation yields
The optimal biotransformation yields were obtained muring the area values from HPLC/DAD applying the follong formula:
yields=∑
Area of total bioconverted products
Area of aglycone at time 0× 100
.6 mm× 250 mm, 5�m, LiChrosorb RP18 (Merck, Darmtadt, Germany) maintained at 26◦C equipped withmm× 10 mm LiChrosorb RP18 pre column. The elueere (A) Acid H2O at pH 3.2 by H3PO4; (B) MeOH and (CH3CN. The following linear solvent gradient was applimin from (A) 92% and (C) 8%, within 8 min to (A) 72%
B) 20% and (C) 8%, with a plateau of 12 min, 16 min tond (B) 40% and (C) 20%, with a plateau of 8 min, finmin to reach (B) 60% and (C) 40%. Total time of anal2 min, flow 1 mL min−1.
Semipreparative HPLC: Separation of single compounas obtained by using a pump Series 250 LC equippedn LC 95 UV–vis detector (all from Perkin-Elmer, Norwa
Fig. 1. Reference structures for the considered anthraquinone mole

M. Nadia et al. / Enzyme and Microbial Technology 36 (2005) 399–408 401
Fig. 2. Chromatographic profiles at 430 and 330 nm of culture supernatants for samples collected at 72 h after feeding (a) emodin and (b) aloe–emodin.

402 M. Nadia et al. / Enzyme and Microbial Technology 36 (2005) 399–408
Fig. 3. Histograms to show the relative amounts (mAu) evaluated at 254 nmfor all the bioconverted molecules obtained at 48 and 72 h of contact: (a)emodin and (b) aloe–emodin.
CT, USA). The eluents were (A) Acid H2O at pH 3.2 byHCOOH; (B) MeOH and (C) CH3CN
The following linear solvent gradient was applied: from(A) 80%, (B) and (C) 10% to (A) 72%, (B) and (C) 14%,within 5 min, 7 min to reach (A) 70%, (B) 16% and (C)14%, 10 min to (A) 50%, (B) 20% and (C) 30% witha plateau of 14 min. Total time of analysis 36 min, flow5 mL min−1. The column was a 250 mm× 10 mm, 10�m,LiChrosphere RP18 (Merck, Darmstadt, Germany) equippedwith a 10 mm× 10 mm pre-column of the same phase.
For the analytical HPLC profiles the UV–vis spectra wererecorded in the range of 190–550 nm, and chromatogramswere acquired at 230, 254, 294, 330 and 430 nm. The ab-solute maximum of UV–vis spectra, both for emodin andaloe–emodin, was 254 nm; a characteristic maximum was at294 nm, while the chromatographic profiles at 430 nm wereused to identify aloe–emodin, emodin and their derivatives.The HPLC profiles at 330 nm were representative of thephenyl propanoidic derivatives, the main metabolites of theseplant cell cultures.
2.7. HPLC/MS analysis
The HPLC/MS analyses were performed using a HP1090L liquid chromatograph equipped with a DAD detector.T erat-i thefr polet n-
tors operated in the range 80–150 V. Due to the orthogonalposition of the nebulizer with respect to the capillary inlet,the chromatographic conditions were the same applied forthe HPLC/DAD analysis except for the acid modifier of thesolvent (A) that was substituted with HCOOH.
MS/MS analyses were carried out by an API 365 Sciextriple quadrupole instrument (Applied Biosystems, MSDSCIEX, Toronto, Canada), operating in negative ion mode.The mass spectrometric data were recorded in scan mode inthe mass range from 200 to 800 Th. The mass analyser wasoperated at unit mass resolution.
2.8. NMR analysis
1H NMR and 13C NMR spectra were recorded on Var-ian Gemini 200 spectrometers operating in FT. For1H and13C the following solvent signals were applied as internalstandard: 3.31 and 49.00 ppm from TMS, respectively, withCD3OD as solvent and 2.48 and 39.7 ppm in the case ofDMSO.
3. Results and discussion
3.1. Screening on anthraquinone molecules
ertt sev-e odin,1 car-r
uslye wnp ctionw (5-d were
TL olec-u
F
9
F
e
he interface was a HP 1100 MSD API-electrospray opng in negative ionisation mode (Hewlett & Packard) atollowing operating conditions: gas temperature 350◦C, flowate 10.0 L/min, nebulizer pressure 30–40 psi, quadruemperature 30◦C and capillary voltage 3500 V. Fragme
To check the ability of the Ajuga cell cultures to bio convhe anthraquinone nucleus, a preliminary screening onral dihydroxy anthraquinones (rhein, emodin, aloe–em,4 and 1,8-dihydroxy-anthraquinones, and aloin) wasied out at different times of contact.
The optimal time to add the precursors was previostablished corresponding to the initial logarithmic grohase (11-day-old cultures) and the glycosilation reaas improved supplementing the culture with sucroseay-old cells). Every 24 h of contact, the supernatants
able 1ist of the bioconverted compounds obtained for the two precursors, mlar weights in brackets.
Negative ion species (m/z)
rom emodin (Mw)1e Acetyl-diglycoside (636) 635, 2692e Unknown 457, 2533e Dimalonyl-diglycoside (766) 765, 677, 2694e 6-O-�-Galactoside (432) 431, 2695e 6-O-�-Malonyl galactoside (518) 517, 473, 311, 282, 26
rom aloe–emodin (Mw)1a Unknown 269, 623, 7992a Acetyl-glycoside (474) 473, 2693a Acetyl-glycoside (474) 473, 2694a Diglycoside (594) 593, 2685a Acetyl-diglycoside (636) 635, 593, 2686a 11-O-�-Galactoside (432) 431, 2687a Dimalonyl-diglycoside (766) 765, 677, 635, 2688a 11-O-�-Malonyl-galactoside (518) 517, 473, 268
, emodin; a, aloe–emodin.

M. Nadia et al. / Enzyme and Microbial Technology 36 (2005) 399–408 403
Fig. 4. Mass spectra of compound5e(a) ESI in negative ion mode and (b) MS/MS spectrum of [M−H]− ion atm/z 517.
drawn, filtered and directly analysed by HPLC/DAD to checkthe bioconversion process.
It was observed that 1,4-dihydroxy-anthraquinone andaloin were not bioconverted at all remaining unmodified upto 48 h of contact, rhein intoxicated the cells, while tracesof bioconverted products were only revealed for the 1,8-dihydroxy-anthraquinone. Finally, the chromatographic pro-files from emodin and aloe–emodin (chemical structure inFig. 1) show the presence of more polar molecules with char-acteristic UV–vis spectra after only 24 h of contact, therefore,these molecules were chosen to perform an in-depth investi-gation.
The best results in terms of bioconversion yields for boththese aglycones were found after 48–72 h of contact. Observ-ing the HPLC/DAD profiles of the supernatants of these sam-ples collected at 48 and 72 h of contact (Fig. 2), four and sevenderivatives were identified for emodin and aloe–emodin, re-
spectively. The presence of glycosilated anthraquinones wasconfirmed both by UV–vis and mass spectra.
Observing the distribution of the bioconverted moleculesat 48 and 72 h, irrelevant differences were observed for the
Table 2Proton signals (CD3OD) of the aromatic nucleus of the main glycosides fromemodin, aloe–emodin compared with frangulina (emodin 11-O-�-glucoside)
Emodin (ppm) Aloe–emodin (ppm)
Frangulina Compound5e
Compound4e
Compound6a
Compound8a
H2 7.64 7.65 7.64 7.46 7.46H4 7.46 7.48 7.48 7.87 7.86H5 7.15 7.15 7.17 7.76 7.76H6 – – – 7.81 7.81H7 6.96 6.95 6.92 7.34 7.35Hanom 5.64 5.13 5.14 4.44 4.45
404 M. Nadia et al. / Enzyme and Microbial Technology 36 (2005) 399–408
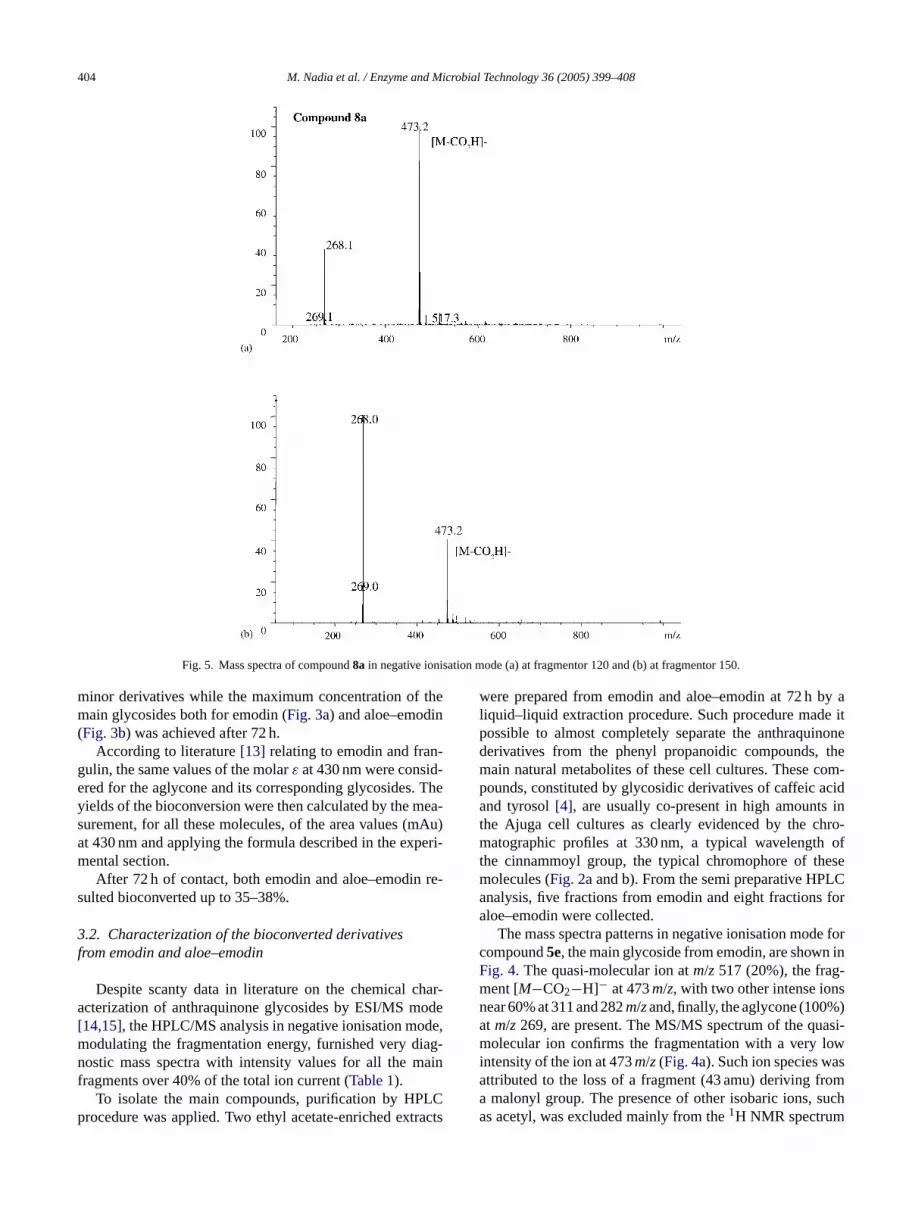
Fig. 5. Mass spectra of compound8a in negative ionisation mode (a) at fragmentor 120 and (b) at fragmentor 150.
minor derivatives while the maximum concentration of themain glycosides both for emodin (Fig. 3a) and aloe–emodin(Fig. 3b) was achieved after 72 h.
According to literature[13] relating to emodin and fran-gulin, the same values of the molarε at 430 nm were consid-ered for the aglycone and its corresponding glycosides. Theyields of the bioconversion were then calculated by the mea-surement, for all these molecules, of the area values (mAu)at 430 nm and applying the formula described in the experi-mental section.
After 72 h of contact, both emodin and aloe–emodin re-sulted bioconverted up to 35–38%.
3.2. Characterization of the bioconverted derivativesfrom emodin and aloe–emodin
Despite scanty data in literature on the chemical char-acterization of anthraquinone glycosides by ESI/MS mode[14,15], the HPLC/MS analysis in negative ionisation mode,modulating the fragmentation energy, furnished very diag-nostic mass spectra with intensity values for all the mainfragments over 40% of the total ion current (Table 1).
To isolate the main compounds, purification by HPLCprocedure was applied. Two ethyl acetate-enriched extracts
were prepared from emodin and aloe–emodin at 72 h by aliquid–liquid extraction procedure. Such procedure made itpossible to almost completely separate the anthraquinonederivatives from the phenyl propanoidic compounds, themain natural metabolites of these cell cultures. These com-pounds, constituted by glycosidic derivatives of caffeic acidand tyrosol[4], are usually co-present in high amounts inthe Ajuga cell cultures as clearly evidenced by the chro-matographic profiles at 330 nm, a typical wavelength ofthe cinnammoyl group, the typical chromophore of thesemolecules (Fig. 2a and b). From the semi preparative HPLCanalysis, five fractions from emodin and eight fractions foraloe–emodin were collected.
The mass spectra patterns in negative ionisation mode forcompound5e, the main glycoside from emodin, are shown inFig. 4. The quasi-molecular ion atm/z 517 (20%), the frag-ment [M−CO2−H]− at 473m/z, with two other intense ionsnear 60% at 311 and 282m/zand, finally, the aglycone (100%)at m/z 269, are present. The MS/MS spectrum of the quasi-molecular ion confirms the fragmentation with a very lowintensity of the ion at 473m/z(Fig. 4a). Such ion species wasattributed to the loss of a fragment (43 amu) deriving froma malonyl group. The presence of other isobaric ions, suchas acetyl, was excluded mainly from the1H NMR spectrum

M. Nadia et al. / Enzyme and Microbial Technology 36 (2005) 399–408 405
Fig. 6. Overlapping of the UV–vis spectra of free aglycones and their glycosides (a) emodin and derivatives4eand5eand (b) aloe–emodin and compounds6aand8a.
where the typical singlet corresponding to a methyl grouplinked to a carbonyl was not revealed. On the other hand, theformation of an adduct with HCOOH (the acidic modifier ofthe HPLC mobile phase) during the mass ionisation processwas excluded mainly because the same behaviour for analo-gous molecules, such as other bioconverted compounds andthe standard emodin 6-O-�-rhamnoside, was not evidenced.In addition compound5ewas submitted to an acidic hydroly-sis (HCl 1N in hydro alcoholic solution at room temperaturefor 1 week) to verify the loss of the acylic group in pres-ence of inorganic acids. By HPLC/DAD analysis the com-pound5e resulted almost completely converted in the cor-responding monoglycoside (4e). Moreover, comparing theNMR proton signals of compound5e and emodin-6-O-�-rhamnoside as pure standard, a� linkage, to the hydroxyl inC6, was revealed for both these molecules (Table 2). At thesame time, the presence in DMSO of the signals at 11.9 and12.1 ppm confirmed the existence of free hydroxyl groupsat C1 and C8, respectively. Finally, the presence of a galac-tose was ascertained by the13C NMR spectrum in CD3ODwith all the glycosidic carbons below 76 ppm; the glucosemoiety, linked to different aglycones, usually shows signalsover 76 ppm attributable to C3 and C5 of the sugar[15]. Fi-
nally, product5ewas identified as emodin 6-O-�-malonylgalactoside.
An analogous structure was observed for compound8a.In Fig. 5the mass spectra show very simple patterns; a quasi-molecular ion atm/z 517 and two intense fragments atm/z473 corresponding to [M−CO2−H]−, and atm/z268 relatingto the aglycone. The MS/MS spectrum ofm/z 517 confirmsthe fragmentation pattern reported inFig. 5. From 1H and13C NMR experiments, the hydroxyl groups in C1 and C8resulted free from linkage, as well as for compound5e. Alsothe sugar was again identified as a galactoside bound to thealcoholic hydroxyl in C11 of the aloe–emodin. From the cou-pling constant value of the anomeric proton, a� linkage wasagain confirmed. Finally, this compound was identified asaloe–emodin 11-O-�-malonyl galactoside.
Observing the mass spectra of compounds5eand8a, themain difference consists in the presence of the ion species atm/z 281 and 311 only for5e. Until now the origin of thesetwo intense fragments, certainly related to the structure ofcompound5e, as confirmed by the MS/MS spectra of bothions atm/z 517 and 473 (Fig. 5a and b) is not clear. Fur-thermore compound8a did not show, as expected, the ionspecies atm/z269, attributable to the aglycone, but rather an

406 M. Nadia et al. / Enzyme and Microbial Technology 36 (2005) 399–408
Fig. 7. MS spectra in negative mode of compound4e(a) at fragmentor 80 and (b) at fragmentor 150.
intense fragment atm/z 268. The same behaviour was alsoobserved for the other aloe–emodin glycosides7a, 6a, 5aand4a, all showing UV–vis spectra overlapped with that ofaloe–emodin.
The other identified bioconverted molecules, are sum-marised inTable 1. The two mono glycosides,4e and6a,were characterised by UV–vis, MS and NMR data. InFig. 6athe UV–vis spectra of emodin and their derivatives5eand4eare compared, evidencing an identical shape. The same com-parison concerning the pure aloe–emodin and its derivatives8aand6a is illustrated inFig. 6b: a complete overlapping ofthe UV–vis spectra is highlighted confirming that the chro-mophore remained unmodified after the bioconversion. Anoptimal ionisation in negative mode was again observed forall these minor compounds as illustrated inFig. 7where sig-nificant mass spectra of glycoside4eare reported as example.Both the mass spectra present intense molecular ions atm/z431 and the aglycone species atm/z 269. The difference of162 amu between these two fragments confirms the pres-ence of only one hexose. Finally, from the1H NMR signals(Table 2) the following structures were confirmed: emodin 6-
O-�-galactoside for4eand aloe–emodin 11-O-�-galactosidefor 6a.
Among the other minor compounds simple and acylatedmono- and diglycosides both for emodin and aloe–emodinwere detected (Table 1). The formation of acylated diglyco-sides of emodin (1eand3e) and aloe–emodin (4a, 5aand7a)was confirmed by their mass spectra at different fragmenta-tion energies showing the quasi-molecular ions, at apprecia-ble intensity, together with the loss of typical fragments. InFig. 8, the typical ion pattern of compound3eis reported withthe [M−H]− atm/z765, the [M−88−H]− atm/z677 and theaglycone ion at higher intensity.
For almost all the identified compounds the UV–vis spec-trum of the aglycone overlapped with their correspondingglycosides (e.g.Fig. 6), confirming that the hydroxyl groupsin C1 and C8 always remain free from linkages. The more po-lar compounds1a, 2aand3a, showing, bathochromic shiftsaround 20 nm for theλmax near 400 nm, represent the uniqueexceptions to this behaviour. These results confirm a modifi-cation of the chromophore, presumably involving a linkage ofthe sugar to the hydroxyls in C1 or C8. For the 1,8-dihydroxy-

M. Nadia et al. / Enzyme and Microbial Technology 36 (2005) 399–408 407
Fig. 8. MS spectra of compound3e, dimalonyl diglycoside, (a) at fragmentor 120 and (b) at fragmentor 150.
anthraquinone derivatives the typical intra molecular hydro-gen bonds, between the oxygen in C9 and the adjacent freehydroxyls, increase the hydrophobic characteristic of thesemolecules. The absence of such interaction can be correlatedto the higher polarity in reverse phase, of compounds2aand3awith respect to the other glycosides such as compounds4aand5a.
Work is in progress to collect more of these bioconvertedglycosides to definitively characterise their chemical struc-tures.
3.3. Secondary metabolites
Finally, the effect was evaluated of these anthraquinoneson the production of the main secondary metabolites ofthese cell cultures, constituted by phenyl propanoidic gly-cosides with the general chemical structure illustrated inFig. 9 [4]. In literature many interesting biological prop-erties are cited for analogous compounds: they can reducethe incidence of cataract, act as potent antioxidant and neu-
rosedative[16], antiinflammatory[17] and antiviral [18]agents.
The area values of the three main glycosides (seeFig. 2)were measured at 330 nm for samples at both 48 and 72 h ofcontact. No significant decrease of these phenyl propanoidiccompounds was observed with respect to the references sam-ples. Therefore, aloe–emodin and emodin do not interfere
Fig. 9. Chemical structure of the main phenyl propanoidic metabolites.

408 M. Nadia et al. / Enzyme and Microbial Technology 36 (2005) 399–408
with the metabolic pathway responsible of the biosynthesisof these interesting active principles of the Ajuga cell cul-tures.
4. Conclusions
Scanty data are available in literature on the chemicalcharacterisation of anthraquinone derivatives by ESI/MSmodes[14]. In this paper, we demonstrated that this an-alytical approach is a useful tool to characterize theseacyl anthraquinone glycosides and, to our knowledge, thesemolecules are described for the first time.
Moreover, increased attention has been recently registeredfor emodin and aloe–emodin for their potentiality as anti-cancer agents. Despite the increased scientific interest, natu-ral derivatives of these molecules such as glycosides, are noteasily found on the market as pure compounds and therefore,are not available to carry out biological studies. Our findingscan contribute to identify a new source of acyl glycosilatedanthraquinones and/or to furnish a standardised glycosidicpool of anthraquinones together with a group of biologicallyactive phenyl propanoids.
A
ntalC ccinif hert fi-n
R
L.zyme
Adv
[2] Kubo I, Kloke JA, Ganjian I, Ichikawa N, Matzumoto T. Efficientisolation of phytoecdysones from Ajuga plants by high-performanceliquid chromatography and droplet counter–current chromatography.J Chromatogr A 1983;257:157–61.
[3] Shimomura H, Sashida Y, Ogawa K. Iridoid glucosides and phenyl-propanoid glycosides in Ajuga species of Japan. Phytochemistry1987;26(7):1981–3.
[4] Garzotto C. Identificazione di composti prodotti da colture cellu-lari di Ajuga reptans. Degree Thesis, Faculty of Pharmacy, BolognaUniversity, 1994–95.
[5] Minghetti A, Crespi Perellino N, Mulinacci N, Speroni E. Biotrasfor-mazione di flavoni con colture di cellule diAjuga reptansL. In: Pro-ceedings of the VII National Congress of the Pharmacognosy ItalianSociety. Ed Patron; 1996. p. 103–12.
[6] Mulinacci N, Sansoni C, Romani A, Giaccherini C, Vincieri FF,Crespi Perellino N, et al. Glycosylation of some flavonoids byAjugareptansL. cell culture. TUH editions, Polyphenols Communications2000. Munchen-Frising 2000; 1:39-40.
[7] Della Loggia R. Piante officinali per infusi e tisane. Milano: OEMF;1993.
[8] European Pharmacopoea. IV ed. 2001.[9] Italian Pharmacopoea. XI ed. 2002.
[10] Kuo PL, Lin TC, Lin CC. The antiproliferative activity ofaloe–emodin is trough p53-dependent and p21-dependent apoptoticpathway in human hepatoma cell lines. Life Sci 2002;71:1879–92.
[11] Su HY, Cherng SH, Chen CC, Lee H. Emodin inhibits the mu-tagenicity and DNA adducts induced by 1-nitropyrene. Mutat Res1995;329:205–12.
[12] Sarno S, Moro S, Meggio F, Zagotto G, Dal ben D, Ghisellini P,et al. Toward the rational design of protein kinase casein kinase-2
[[ n
omet-
[ MR
[ in F,nyl-
[ icalRes
[ nsoninal
cknowledgements
We thank C.I.S.M (Mass Spectrometry Interdepartimeenter) of Florence and its director Dr. Giuseppe Piera
or his stimulating discussion and Dr. Sandra Gallori forechnical support. This work was developed with MIURancial support.
eferences
[1] Giri A, Dhingra V, Giri CC, Singh A, Ward PO, Narasu MBiotransformations using plant cells, organ cultures and ensystems: current trends and future prospects. Biotechnol2001;19:175–99.
inhibitors. Pharmacol Ther 2002;93:159–68.13] Merk Index. XIII ed. 2001.14] Goverdina C, Derksen H, Harm A, Niederlander G, Teris A, va
Beek. Analysis of anthraquinones inRubia tinctorumL. by liquidchromatography coupled with diode-array UV and mass spectrric detection. J Chromatogr A 2002;978(1–2):119–27.
15] Agraval PK, Bansal CM. Flavonoid glycosides by Carbon 13 Nof Flavonoids. Elsevier; 1989.
16] Daels-Rakotoarison DA, Seidel V, Gressier B, Brunet C, TillequBailleul F, et al. Neurosedative and antioxidant activities of phepropanoids fromBallota nigra. Drug Res 2000;50(1):16–23.
17] Deepak M, Handa SS. Antiinflammatory activity and chemcomposition of extracts of Verbena officinalis. Phytother2000;14(6):463–5.
18] Kernan MR, Amarquaye A, Chen JL, Chan J, Sesin DF, ParkiN, et al. Antiviral phenylpropanoid glycosides from the medicplant Markhamia lutea. J Nat Prod 1998;61(5):564–70.





























