Embed Size (px)
Citation preview

Int J Clin Lab Res 22:73-80, 1992
�9 Springer-Verlag 1992
CD38: A multi-lineage cell activation molecule with a split personality Fabio Malavasi 1, Ada Funaro 1, Massimo Alessio 1, Lucia B. DeMonte t, Clara M. Ausiello 2, Umberto Dianzani 3 Francesco Lanza 4, Elisabetta Magrini 1, Monica Momo i and Stefano Roggero 1
t Laboratorio di Biologia Ceitulare, Dipartimento di Genetica, Biologia e Chimica Medica, Universitfi di Torino e Centro CII/CIOS, CNR, Turin, Italy z Istituto CNR Tipizzazione Tissutale e Problemi della Dialisi, L'Aquila, Italy 3 Dipartimento di Medicina ed Oncologia Sperimentale, Universitfi di Torino, Turin, Italy 4 Cattedra di Ematologia, Universitfi di Ferrara, Ferrara, Italy
Summary. This review reports the characteristics of the human surface molecule CD38, a structure not linked to a definite line and predominantly expressed in early and activated phenotypes. The CD38 molecule consists of a single chain of 46 kDa, spanning the membrane and with the carboxyl terminus located in the extracellular com- partment. The CD38 molecule is also involved in the transduction of activation and proliferation signals, which are line unrestricted. The gene coding for the CD38 antigen has been cloned and used for the construction of simian and mouse transfectants expressing the human molecule. These cell models are used for the analysis of several unanswered issues, mainly concerning the in vivo function of CD38, the existence of a natural ligand and of polymorphism in the population.
Key words: Cluster designation - CD38/T10 - Agonistic antibody - Cell activation and proliferation
The aim of this review is twofold: first, to summarize the wide body of information available thus far on CD38, and then to provide recent additional evidence on the structure, function, expression and genetic control of the molecule whose conflicting characteristics are still waiting to be delineated in full.
Pattern of tissue distribution
The distribution of CD38 on normal cells is shown in Table 1. The CD38 structure is expressed by the majority of thymocytes, mainly of medullary origin. Resting and circulating T cells are CD38, but express the molecule upon activation, irrespective of the activation source [i.e., phytohemagglutinin (PHA), antigen or mixed lymphocye culture]. The same findings hold true for B cells. Lymph node germinal center lymphoblasts, plasma
Introduction
One of the most exciting aspects of current research on lymphocyte differentiation antigens is that an increasing number of molecules widely used as phenotype markers of several cell types have been reported to play crucial roles in cell regulation. The CD38 structure belongs to a group of cell surface antigens originally described by Reinherz et al. [15], who in the late 1970s extensively applied monoclonal antibodies (mAbs) in the detection of cell surface antigens for the definition of discrete stages of differentiation. T10 [the original definition now more precisely substituted by the Cluster Designation (CD) 38 (2nd International Workshop of White Cell Differentia- tion Antigens)] has been widely used as a marker in the study o fT and B cell activation and differentiation. It has also played an ancillary role in the categorization of lymphocyte malignancies of T and B cell origin.
Table 1. Distribution of the CD38 molecule on normal cells
Percentage of CD38 + ceils
Bone m a r r o w Mononuclear cells 10 T cells 6 TdT + cells 95 Pre-B cells 80 sIgM § cells 70 Plasma cells 80
Thymocytes 90
Peripheral blood T cells 8 B cells 5 NK cells 70 Monocytes 80 Granulocytes T blasts 90 B blasts 80
Fetal leukocytes 90
Offprint requests to: E Malavasi slg, Surface immunoglobulin; NK, natural killer

74 E Malavasi et al.: CD38 and cell activation
Table 2. Distribution of the CD38 molecule on tumor cell lines
CEM (T-ALL) + JURKAT (T-ALL) + HPB-ALL (T-ALL) + U937 (myelomonocytic) + HL60 (myeloid) + REH (pre-B) + RAJI (Burkitt's lymphoma) + DAUDI (Burkitt's lymphoma) + LG-14 (lymphoblastoid) + LP1 (myeloma) + MT3 (myeloma) + U266 (myeloma) + K562 (erythroblastoid) COLO 38 (melanoma)
cells and a variable number of intrafollicular cells express a significant amount of surface CD38. A significant pro- portion of normal bone marrow cells are CD38 § with plasma cells and hemopoietic TdT § precursors being stained by the specific mAb [13]. Among circulating leukocytes, CD38 is expressed by a significant (-~ 80%) proportion of large granular lymphocytes and mono- cytes, even under resting conditions. CD38 is expressed by 50%-80% of mononuclear cells from umbilical cord blood, and this level of expression is maintained for approximately the first 2-3 years of life [R. Maccario (University of Pavia), personal communication].
The distribution on tumor counterparts of selected leukocyte populations indicates that leukemias immor- talizing early differentiation stages maintain CD38 expression, while more differentiated phenotypes are CD38-[14]. However, there are several exceptions. A similar distribution to that observed on normal and dis- eased cells is seen among continuous cell lines. CD38 expression by the lines of hemopoietic origin used in the laboratory are reported in Table 2. Again it appears that the CD38 molecule marks the more undifferentiated phe- notypes, with the relevant exception of cells, such as plas- macytomas, believed to immortalize a mature phenotype.
CD38 structure
The CD38 molecule was originally described as a human cell surface molecule defined by mAb T10 [15]. This mol- ecule was reported to be present on thymocytes and on activated T cells. Cotner et al. [6] described a major sur- face glycoprotein of 46 kDa, with a 12-kDa chain [indis- tinguishable from fl2-microglobulin (flz m)], co-precipi- tating with variable intensity in all the lines analyzed. Terhorst et al. [18] presented evidence that T10 was an integral membrane protein, ruling out any correlation between T10 and the murine Qa2 products, due to the lack of a constant association with flz m.
A relationship between selected T cell surface molec- ules and HLA antigens was lately pursued by Bushkin et al. [2] who developed a mAb reacting with a monomor- phic p46,12 complex, similar to the T10 antigen. This reactivity was restricted to the Molt-4 line, whereas other cell types failed to express the p12 light chain. A relation-
ship between T10 and the Qa2 antigens was further ex- cluded by a lack of structural homology between the p46 and the murine class I antigens [6]. A relationship be- tween CD38 and MHC or the C D l a product was exclud- ed either by somatic cell genetics or by more detailed biochemical analysis of the structure. Katz et al. [12] mapped the gene encoding the CD38 structure to the human chromosome 4 by means of hetero-hybrids. MHC is located on chromosome 6, while CD1 is on chromo- some 1 [3]. The availability of the A10 antibody [with a good K d (10-8 M)] made a comprehensive evaluation of the CD38 molecule possible. As can be seen in Fig. 1 A, after surface labelling with 125iodine (I) and sodium dodecyl sulfate polyacrylamide gel electrophoresis, the immunoprecipitated CD38 molecule appears as a single chain of approximately 46 kDa. There are no apparent differences when the molecule is immunoprecipitated from cells belonging to different lineages, nor is there evidence of association with 12-kDa chains. Two-dimen- sional isoelectric focusing of CD38 and structures of a similar molecular weight indicates that the isoelectric point of CD38 ranges from 6.5 to 6.9, with five discrete spots, which is markedly different from either HLA class I (Fig. 1 B) or actin (not shown). An extensive bio- chemical analysis concluded that the CD38 molecule is a glycoprotein spanning the cell membrane and the carbo- hydrate component accounts for approximately 40% of the observed molecular weight. A thorough survey of the biochemical features of CD38 has been reported by Alessio et al. [1].
Functional implications
The addition of anti-CD38 mAb does not interfere with specific recognition by T or B cells, with the T-monocyte interactions, with allotypic response and with cytotoxic lymphocyte natural killer cell (NK) killing. However, a limited number of mAbs have the ability to transduce activation signals upon target binding; these antibodies are defined as agonistic, as opposed to the majority of antibodies which are antagonistic [5]. The effects induced by antibody binding may be studied at a phenotypic level (i.e., by evaluating the expression of molecules produced by activated or proliferating cells) or in functional terms (e.g., by studying the effector functions elicited by trans- membrane signalling or the production and release of relevant lymphokines). Following this approach, Funaro et al. [9] showed that the addition of A10 and IB4 (two specific anti-CD38 mAbs) to preparations of purified T cells, thymocytes and NK cells (CD16 § CD56 § caused activation. Forty-eight hours after the addition of the mAb (10 ~tg/0.5 x 1 0 6 cells per 100 Ixl), there was a signif- icant increase in 3H-thymidine incorporation (which peaked on the 3rd-4th day) indicating cell proliferation. The uptake of 3H-thymidine by T cells was lower than that obtained with CD3 activation but approximately similar to that obtained with CD2. Activation kinetics were faster with CD3 activation. A constant feature was the dependence on activation and proliferation of acces- sory cells. Similar results were also obtained with prepa-

F. Malavasi et al.: CD38 and cell activation 75
-200
o i i
-30
-14" A B
J - - Ib
|
"11[~
!
0"am -,r
Fig. 1. A SDS-PAGE analysis of the CD38 mole- cule. CD38 was immunoprecipitated from lysates of surface ~zsI-labeled M LC blasts and thymocytes. B Two-dimensional IEF gel electrophoresis of puri- fied ~zsI-CD38 molecule, as compared with human HLA-class I molecules
rations of purified NK cells. The general conclusions are that CD38 is involved in cell activation, with no restric- tion to a definite cell line.
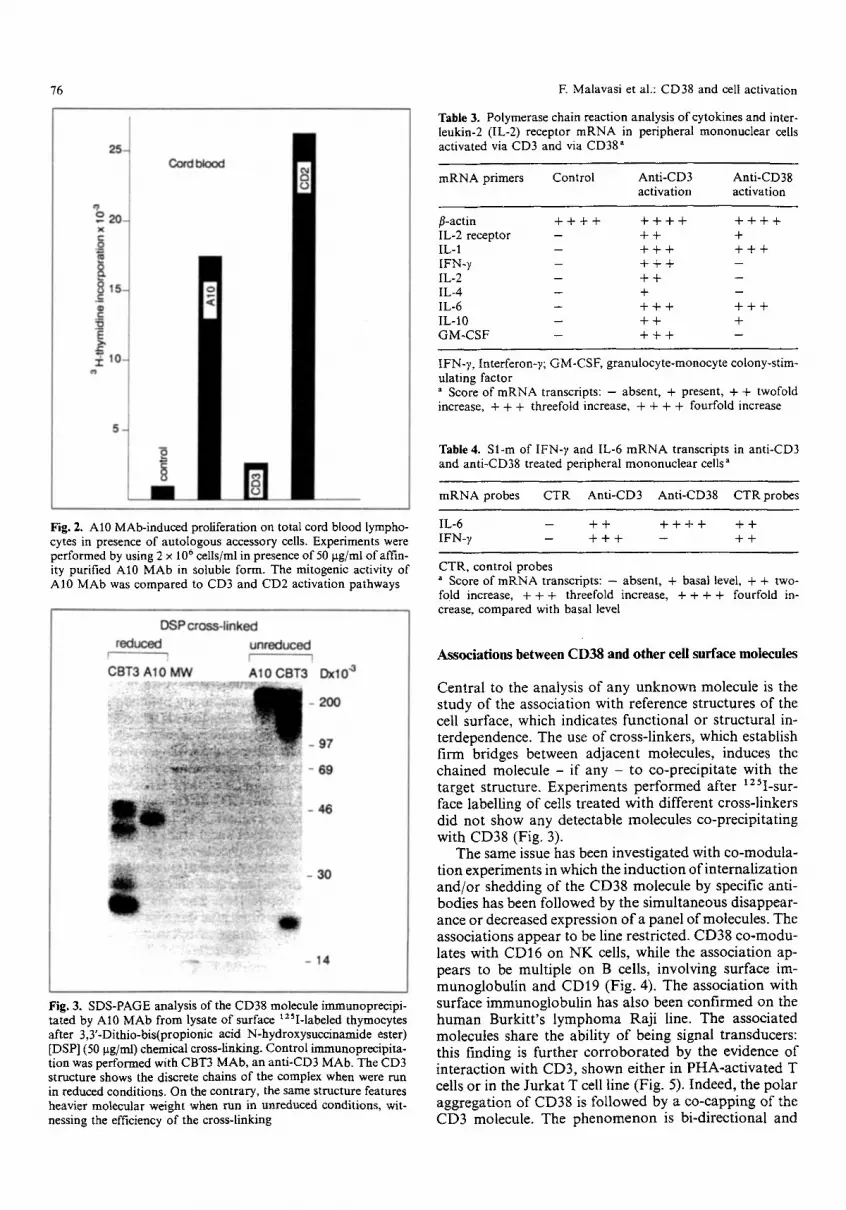
Finally, CD38 activation was investigated in relation to events induced via CD3 and CD2 in order to ascertain whether events were synergistic or antagonistic. The ad- dition of anti-CD38 mAb to cells treated with optimal amounts of anti-CD3 or, ant i-Tllz+3 was followed by a significantly enhanced activation and proliferation. CD38 did not appear to enhance or block the effects of a number of accessory molecules, i.e., CD45, CD44 and CD5. CD38 activation has been also investigated in um- bilical cord blood cells, in order to evaluate premature functions. The activation of such cells via CD3 is ex- tremely reduced [9], while the activation via CD2 is as in the adult controls and is accessory cell dependent (Fig. 2). CD38 activation appears to work efficiently in such cells.
Cytokine expression by CD38 activation
The mechanisms underlying activation via CD38 have been studied by assessing lymphokine release into the culture medium after CD38-driven activation. Surpris- ingly, it was observed that the mean level of interleukin-2 (IL-2) was in the same range as the control; however, this appeared to be related to a secondary high consumption of IL-2 linked to the increased metabolic rate of activated cells. This was further substantiated by an increased ex- pression of the CD25 molecule during activation. The involvement of IL-2 was indirectly confirmed by inhibi- tion of activation and proliferation by CB-TAC, an anti- CD25 blocking mAb. Significant amounts of IL-6 were
released upon activation via CD38 of cell line models (A. Mantovani, personal communication, 1990).
The CD38 activation process was compared with the classical CD3 activation pathways by analyzing the pro- duction by monocytes and T cells of messenger RNA (mRNA) of IL-1, interferon-7 (IFN-,/), IL-2, IL-4, IL-6, IL-10 and granulocyte-monocyte colony-stimulating fac- tor (GM-CSF). m R N A levels of the IL-2 receptor were used as parameters of lymphocyte activation. The cy- tokine m R N A was extracted from peripheral blood mononuclear cells which were cultured for 15 h in the presence of selected stimuli, i.e., the CBT3G mAb (anti- CD3) and IB4 mAb (anti-CD38). The polymerase chain reaction was used to evaluate the transcript levels of m R N A for all the cytokines under study [8]. Owing to the larger amounts of RNA needed to perform the test ( ~ 10 gg RNA for each experimental condition), the SI mapping method (Sl-m) was used only to measure IFN- 7 and IL-6 m R N A levels [10]. The relative density of m R N A bands was measured after densitometric analysis. Table 3 shows the percentage of relevant m R N A tran- scripts compared with the reference /3-actin m R N A transcripts. The patterns of IL-1 and GM-CSF tran- scripts induced in mononuclear cells activated via CD38 or CD3 were almost identical. Higher mRNA levels of IL-2, IFN-7, IL-10 and IL-2 receptor resulted after CD3 activation than CD38 activation. IL-6 mRNA transcripts observed during CD38-mediated activation were present at significantly higher levels than during CD3 activation (Tables 3 and 4). No information is available at the mo- ment concerning the stability of the messengers of the individual lymphokines.
76
2 5 -
Cord blood
2 0 - x e -
.g
O
815- ._=
E e-
~-10-
5 -
Fig. 2, A10 MAb-induced proliferation on total cord blood lympho- cytes in presence of autologous accessory cells. Experiments were performed by using 2 x 106 cells/ml in presence of 50 p.g/ml of affin- ity purified A10 MAb in soluble form. The mitogenic activity of A10 MAb was compared to CD3 and CD2 activation pathways
DSP cross-linked
reduced unreduced f ~ ] [ I
CBT3 A10 MW A10 CBT3 Oxl0 ~
- 2 0 0
- 9 7
- 6 9
- 4 6
3 0
~ ; :~ i �84 , ? ! i l
- 1 4
Fig. 3. SDS-PAGE analysis of the CD38 molecule immunoprecipi- tated by A10 MAb from lysate of surface t2SI-labeled thymocytes after 3,3'-Dithio-bis(propionic acid N-hydroxysuccinamide ester) [DSP] (50 gg/ml) chemical cross-linking. Control immunoprecipita- tion was performed with CBT3 MAb, an anti-CD3 MAb. The CD3 structure shows the discrete chains of the complex when were run in reduced conditions. On the contrary, the same structure features heavier molecular weight when run in unreduced conditions, wit- nessing the efficiency of the cross-linking
F. Malavasi et al.: CD38 and cell activation
Table 3. Polymerase chain reaction analysis of cytokines and inter- leukin-2 (IL-2) receptor mRNA in peripheral mononuclear cells activated via CD3 and via CD38 =
mRNA primers Control Anti-CD3 Anti-CD38 activation activation
fl-actin + + + + + + + + + + + + IL-2 receptor -- + + + IL-1 - + + + + + + IFN-y - + + + -- IL-2 - + + - IL-4 -- + -- IL-6 - + + + + + + IL-10 -- + + + GM-CSF - + + + -
IFN-y, Interferon-y; GM-CSF, granulocyte-monocyte colony-stim- ulating factor a Score of mRNA transcripts: - absent, + present, + + twofold increase, + + + threefold increase, + + + + fourfold increase
Table 4. Sl-m of IFN-y and IL-6 mRNA transcripts in anti-CD3 and anti-CD38 treated peripheral mononuclear cells a
mRNA probes CTR Anti-CD3 Anti-CD38 CTRprobes
IL-6 -- + + + + + + + + IFN- v - + + + - + +
CTR, control probes = Score of mRNA transcripts: -- absent, + basal level, + + two- fold increase, + + + threefold increase, + + + + fourfold in- crease, compared with basal level
Associations between CD38 and other cell surface molecules
Cen t r a l to the analysis o f any u n k n o w n m o l e c u l e is the s tudy o f the assoc ia t ion wi th re fe rence s t ruc tures o f the cell surface, which indica tes f unc t i ona l o r s t ruc tu ra l in- t e rdependence . T h e use o f c ross - l inkers , wh ich es tabl ish f i rm br idges be tween a d j a c e n t molecu les , induces the cha ined molecu le - i f any - to co -p rec ip i t a t e wi th the t a rge t s t ruc ture . E x p e r i m e n t s p e r f o r m e d af ter xzSI-sur- face label l ing o f cells t r ea ted wi th d i f fe ren t c ross- l inkers d id n o t s h o w any de tec tab le m o l e c u l e s co -p rec ip i t a t i ng wi th C D 3 8 (Fig. 3).
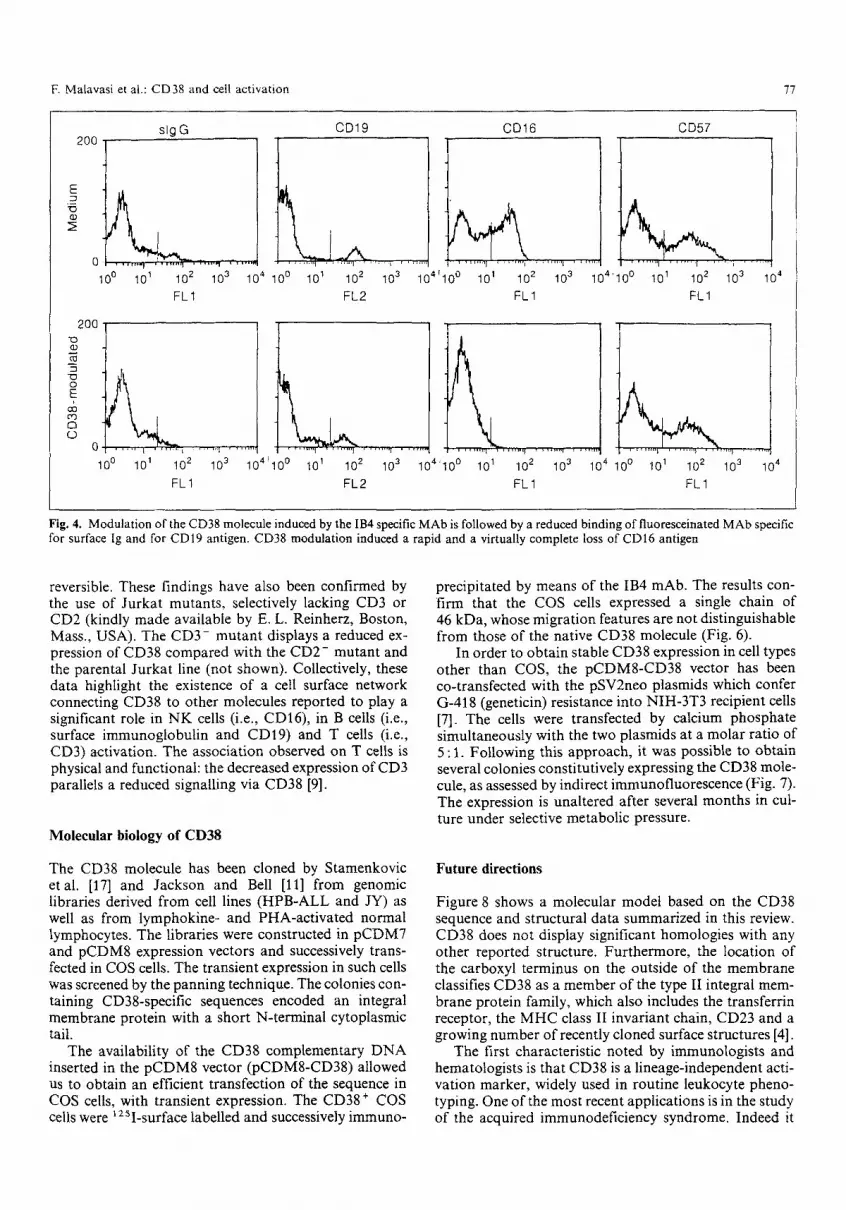
T h e same issue has been inves t iga t ed wi th c o - m o d u l a - t ion expe r imen t s in which the i n d u c t i o n o f i n t e rna l i za t i on a n d / o r shedd ing o f the C D 3 8 m o l e c u l e by specif ic ant i - bod ies has been fo l lowed by the s imu l t aneous d i sappea r - ance o r decreased express ion o f a pane l o f molecu les . T h e assoc ia t ions a p p e a r to be line res t r ic ted . C D 3 8 c o - m o d u - lates wi th C D 1 6 on N K cells, whi le the a s soc ia t ion ap- pears to be mul t ip l e on B cells, i nvo lv ing sur face im- m u n o g l o b u l i n and C D 1 9 (Fig . 4). T h e a s soc ia t ion wi th sur face i m m u n o g l o b u l i n has a l so been c o n f i r m e d o n the h u m a n Burk i t t ' s l y m p h o m a R a j i line. T h e assoc ia ted molecu le s share the abi l i ty o f be ing s ignal t ransducers : this f inding is fu r the r c o r r o b o r a t e d by the ev idence o f i n t e r ac t i on wi th C D 3 , s h o w n e i the r in P H A - a c t i v a t e d T cells o r in the J u r k a t T cell l ine (Fig. 5). Indeed , the po l a r a g g r e g a t i o n o f C D 3 8 is f o l l o w e d by a c o - c a p p i n g o f the C D 3 molecule . T h e p h e n o m e n o n is b i -d i r ec t iona l and
E Malavasi et al.: CD38 and cell activation 77
200
0 10 ~
2O0 0 O3
q[9 0 E
o
slgG CD19 CD16 CD57
101 102 103 104 100 101 102 103 10410 ~ 101 102 103 104 FL1 FL2 FL1
10 0
i
10 ~ 102 103 104100 101 102 103 104'10 o 101 102 103 104 FL1 FL2 FL1
t
100 10 ~ 102 103 104 FL1
10 o 101 10 2 10 3 10 4
FLI
Fig. 4. Modulation ofthe CD38 molecule induced by the IB4 specific MAb is followed by a reduced binding of fluoresceinated MAb specific for surface lg and for CDI9 antigen. CD38 modulation induced a rapid and a virtually complete loss of CD16 antigen
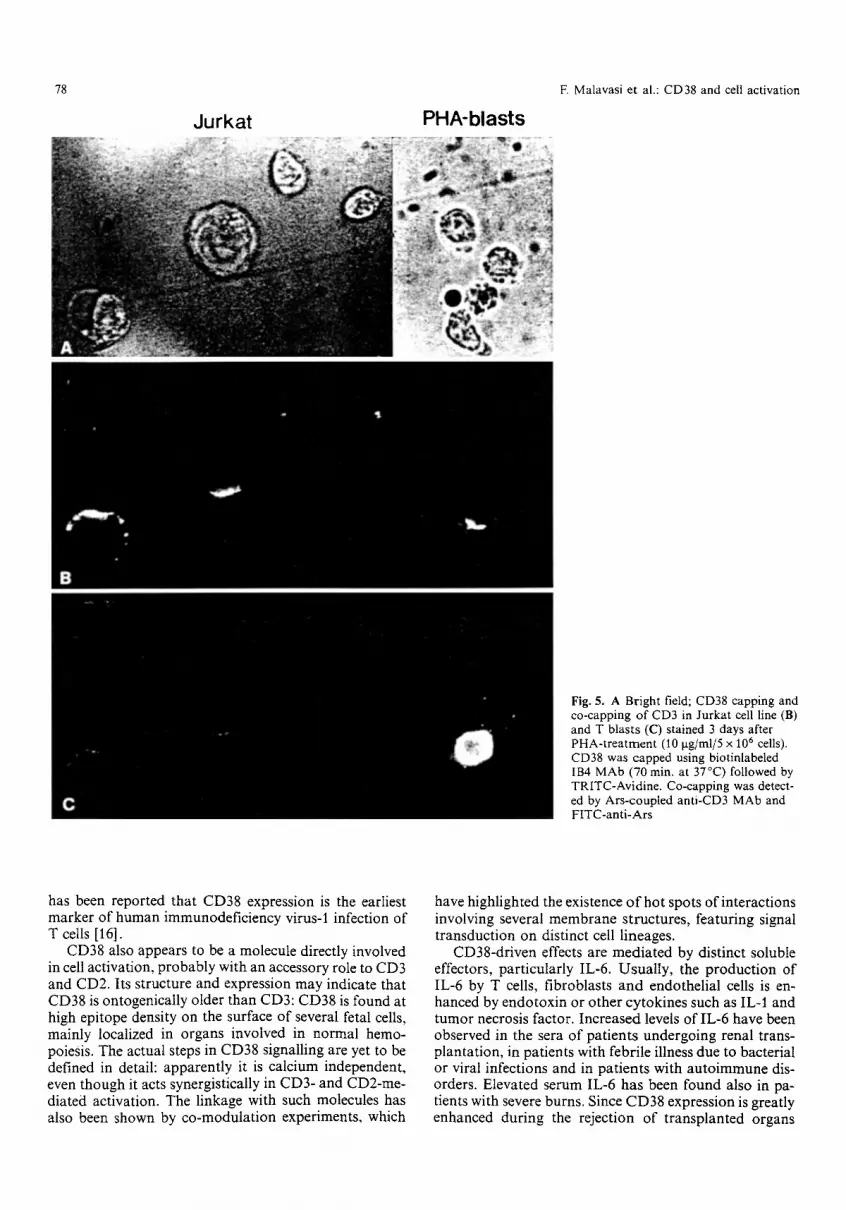
reversible. These findings have also been confirmed by the use of Jurkat mutants, selectively lacking CD3 or CD2 (kindly made available by E. L. Reinherz, Boston, Mass., USA). The CD3- mutant displays a reduced ex- pression of CD38 compared with the CD2- mutant and the parental Jurkat line (not shown). Collectively, these data highlight the existence of a cell surface network connecting CD38 to other molecules reported to play a significant role in NK cells (i.e., CD16), in B cells (i.e., surface immunoglobulin and CD19) and T cells (i.e., CD3) activation. The association observed on T cells is physical and functional: the decreased expression of CD3 parallels a reduced signalling via CD38 [9].
Molecular biology of CD38
The CD38 molecule has been cloned by Stamenkovic etal. [17] and Jackson and Bell [11] from genomic libraries derived from cell lines (HPB-ALL and JY) as well as from lymphokine- and PHA-activated normal lymphocytes. The libraries were constructed in pCDM7 and pCDM8 expression vectors and successively trans- fected in COS cells. The transient expression in such cells was screened by the panning technique. The colonies con- taining CD38-specific sequences encoded an integral membrane protein with a short N-terminal cytoplasmic tail.
The availability of the CD38 complementary DNA inserted in the pCDM8 vector (pCDM8-CD38) allowed us to obtain an efficient transfection of the sequence in COS cells, with transient expression. The CD38+ COS cells were l zSI-surface labelled and successively immuno-
precipitated by means of the IB4 mAb. The results con- firm that the COS cells expressed a single chain of 46 kDa, whose migration features are not distinguishable from those of the native CD38 molecule (Fig. 6).
In order to obtain stable CD38 expression in cell types other than COS, the pCDMS-CD38 vector has been co-transfected with the pSV2neo plasmids which confer G-418 (geneticin) resistance into NIH-3T3 recipient cells [7]. The cells were transfected by calcium phosphate simultaneously with the two plasmids at a molar ratio of 5:1. Following this approach, it was possible to obtain several colonies constitutively expressing the CD38 mole- cule, as assessed by indirect immunofluorescence (Fig. 7). The expression is unaltered after several months in cul- ture under selective metabolic pressure.
Future directions
Figure 8 shows a molecular model based on the CD38 sequence and structural data summarized in this review. CD38 does not display significant homologies with any other reported structure. Furthermore, the location of the carboxyl terminus on the outside of the membrane classifies CD38 as a member of the type lI integral mem- brane protein family, which also includes the transferrin receptor, the MHC class II invariant chain, CD23 and a growing number of recently cloned surface structures [4].
The first characteristic noted by immunologists and hematologists is that CD38 is a lineage-independent acti- vation marker, widely used in routine leukocyte pheno- typing. One of the most recent applications is in the study of the acquired immunodeficiency syndrome. Indeed it
78 F. Malavasi et al.: CD38 and cell activation
Jurkat PHA-blasts
Fig. 5. A Bright field; CD38 capping and co-capping of CD3 in Jurkat cell line (B) and T blasts (C) stained 3 days after PHA-treatment (10 I~g/ml/5 • 10 6 cells). CD38 was capped using biotinlabeled IB4 MAb (70 min. at 37~ followed by TRITC-Avidine. Co-capping was detect- ed by Ars-coupled anti-CD3 MAb and FITC-anti-Ars
has been reported that CD38 expression is the earliest marker of human immunodeficiency virus-1 infection of T cells [16].
CD38 also appears to be a molecule directly involved in cell activation, probably with an accessory role to CD3 and CD2. Its structure and expression may indicate that CD38 is ontogenically older than CD3:CD38 is found at high epitope density on the surface of several fetal cells, mainly localized in organs involved in normal hemo- poiesis. The actual steps in CD38 signalling are yet to be defined in detail: apparently it is calcium independent, even though it acts synergistically in CD3- and CD2-me- diated activation. The linkage with such molecules has also been shown by co-modulation experiments, which
have highlighted the existence o f hot spots of interactions involving several membrane structures, featuring signal transduction on distinct cell lineages.
CD38-driven effects are mediated by distinct soluble effectors, particularly IL-6. Usually, the production of IL-6 by T cells, fibroblasts and endothelial cells is en- hanced by endotoxin or other cytokines such as IL-1 and tumor necrosis factor. Increased levels of IL-6 have been observed in the sera of patients undergoing renal trans- plantation, in patients with febrile illness due to bacterial or viral infections and in patients with autoimmune dis- orders. Elevated serum IL-6 has been found also in pa- tients with severe burns. Since CD38 expression is greatly enhanced during the rejection of transplanted organs

E Malavasi et al.: CD38 and cell activation
Mrx lO "a MW IB4
200- --
92-
69- t=,
4 6 - " ~
3 0 - w
21- q.n ~ (
1 4 - "
Fig. 6. SDS-PAGE analysis of the CD38 molecule immunoprecipi- tated from pCDM8-CD38 transfected COS cells
79
200
100 101 10 2
FL 1
10 3 10 4
100
0 , , , , , , , ,
10 o 104 l ,,i,,,, I �9 ,i,,i,, r i , ,,,,i, I
101 1 0 2 10 3
FL I
Fig. 7. Immunofluorescence analysis performed on NIH-3T3 cells co-transfected with the pCDM8-CD38 vector and the pSV2-neo plasmid, x axis=fluorescence intensity; y axis=number of cells analyzed
0
NH 2
o %o Oo
Fig. 8. Hypothetical 3-dimensional structure of the CD38 molecule as inferred by com- puter analysis following Garnier's method

80 F. Malavasi et al.: CD38 and cell activation
and, in general, dur ing cell-mediated immune responses, it is likely that in these condit ions CD38 triggering is linked with increased IL-6 production.
The regulation of CD38 expression and the existence of a natural l igand specific for this structure are both elusive issues. There are some data suggesting that solu- ble CD38 molecules are present in biological fluids (e.g., amniotic fluids or sera of patients with myeloma). If con- firmed, these data would suggest for CD38 a model of immunomodu la t ion c o m m o n to other surface receptors (e.g., CD25, CD23, CD8, H L A class I,/~2 m, CD35, etc.). The more relevant quest ion concerns the natural ligand for CD38. Since CD38 is expressed in cells during "acti- vat ion," one would immediately see it as a receptor for a growth factor. Other data suggest that it is a receptor for a hetero- or homotypic adhesion molecule. In this case, the mouse cell expressing the human CD38 product may prove extremely useful in studying the optimal condit ions for adhesion.
The following issues require further investigation: (1) the efficiency of the permanent insertion of the gene in recipients other than NIH-3T3 and mainly in non-adher- ing cells, (2) the stability of the randomly inserted se- quence in the new genomic environment , (3), the quanti- tative expression in recipient cells (is there any linkage to cell cycle?) and (4) the analysis of the polymorphism of the CD38 gene, as hypothesized by Jackson and Bell [11]. The data favoring the existence of a polymorphism of CD38 are also supported by the findings of Stamenkovic et al. [17] who found differences in the m R N A of plasma cells (with the highest CD38 expression) and the majority of other cells. This is further substantiated by quanti ta- tive variations in the phenotypic expression of CD38 among normal individuals. Some findings suggest that normal healthy donors may be grouped as "high" and "low" responders in terms of cell activation via CD38.
Acknowledgements. This work was supported by the Project "AIDS" (Istituto Superiore di Sanitfi, Rome 1992), by the Associazione Ital- iana Ricerca Cancro, by the Special Projects "BBS" and "ACRO" (CNR, Rome). A. Funaro, M. Alessio, L. B. DeMonte and S. Rog- gero are recipients of a fellowship from the AIDS project, while M. Momo is a Tecnogen Research Fellow. E. Magrini, on leave of absence from Clinica Pediatrica III, Universitfi di Bologna, is sup- ported by the Fondazione AGEOP. The authors are greatly in- debted to Dr. D. Jackson, who generously made available the CD38 cDNA, and Dr. R. Maccario for helpful suggestions and data on age distribution of CD38 in newborn cells.
Note added in proof
The original title of this paper was "CD38: ...... with a schizophrenic personality". However our linguistic revisor found the term schizophrenic to specialistic and addressed to a limited audience. In turn she suggested split as an alternative. After sending this invited review, we found in the literature the paper by J. Gordon (Clin Exp Immunol 86: 356, 1991), who used a similar title, which was in turn borrowed by Professor H. Gould. And the end of this long chain which we have inadvertently entered, we are extremely grateful to Dr. J. Gordon for giving the permission to use the present title.
References
1. Alessio M, Roggero S, Funaro A, DeMonte LB, Peruzzi L, Geuna M, Malavasi F, CD38 molecule: structural and biochem- ical analysis on human T lymphocytes, thymocytes and plasma cells. J Immunol 145:878, 1990
2. Bushkin Y, Chorney M J, Diamante E, Lane C, Fu SM, Wang CY, Biochemical characterization of a P43,12 complex: com- parison with human and murine Class I molecules. Mol Im- munol 22:695, 1985
3. Calabi F, Bradbury A, The CD1 system. A review. Tissue Anti- gens 37: 1, 1991
4. Claesson L, Larhammar D, Rask L, Peterson PA, A cDNA clone for human invariant j chain of Class II histocompatibility antigens and its implication for the protein structure. Proc Natl Acad Sci USA 80: 7395, 1983
5. Clark EA, Ledbetter JA, Amplification of the immune response by agonistic antibodies. Immunol Today 7:267, 1986
6. Cotner T, Mashimo H, Kung PC, Goldstein G, Strominger JL, Human T cell surface antigens bearing a structural relationship to HLA antigens. Proc Natl Acad Sci USA 78: 3858, 1981
7. DeMonte LB, Nistic6 P, Tecce R, Dellabona P, Momo M, Anichini A, Mariani M, Natali PG, Malavasi F, Gene transfer by retrovirus-derived shuttle vectors in the generation of mu- rine bispecific monoclonal antibodies. Proc Natl Acad Sci USA 87: 2941, 1990
8. Ehler S, Smith KA, Differentiation ofT cell lymphokines gene expression: the in vitro acquisition ofT cell memory. J Exp Med 173:25, 1991
9. Funaro A, Spagnoli GC, Ausiello CM, Alessio M, Roggero S, Delia D, Zaccolo M, Malavasi F, Involvement of the multi- lineage CD38 molecule in a unique pathway of cell activation and proliferation. J Immunol 145:2390, 1990
10. Gessani S, McCandless P, Baglioni C, The glucocorticoid dexa- methasone inhibits synthesis of interferon by decreasing the level of its mRNA. J Biol Chem 283: 7454, 1988
11. Jackson DG, Bell JI, Isolation of a cDNA encoding the human CD38 (T10) molecule a cell surface glycoprotein with an un- usual discontinuous pattern of expression during lymphocyte differentiation. J Immunol 144:2811, 1990
12. Katz F, Povey S, Parkar M, Schneider C, Sutherland R, Stanley K, Solomon E, Greaves M, Chromosome assignment of mono- clonal antibody-defined determinants on human leukemic ceils. Eur J Immunol 13: 1008, 1983
13. Ma DDF, Sylwestrowicz, Granger S, Massaia M, Franks R, Janossy G, Hoffbrand AV, Distribution of terminal deoxy- nucleotidyl transferase and purine degradative and synthetic enzymes in subpopulations of human thymocytes. J Immunol 129:1430, 1982
14. Malavasi F, Caligaris-Cappio F, Dellabona P, Ricchiardi P, Carbonara AO, Characterization of a murine monoclonal anti- body specific for human early lymphohemopoietic cells. Hum Immunol 9:9, 1984
15. Reinherz EL, Kung PC, Goldstein G, Levey RH, Schlossman SF, Discrete stages of human intrathymic differentiation: anal- ysis of normal thymocytes and leukemic lymphoblasts on T-cell lineage. Proc Natl Acad Sci USA 77:1588, 1980
16. Schnizlein-Bick CT, Magier MR, Jones RB, Fife KH, Katz BP, Walker EB, Differences among mononuclear cell subpopula- tions in HIV seropositive or seronegative homosexual and heterosexual men as determined by four-color flow cytometry. J AIDS 3: 747, 1990
17. Stamenkovic I, Staunton J, Seed B, Molecular cloning of CD38. In: Knapp W, D6rken B, Gilks WR, Rieber EP, Schrnidt RE, Stein H, Kr von dem Borne AEG (eds) Leukocyte typing. IV. Oxford University Press, Oxford, p 87, 1989
18. Terhorst C, Agthoven A van, Le Clair K, Snow P, Reinherz EL, Schlossman SF, Biochemical studies of the human thymocyte cell-surface antigens T6, T9 and T10. Cell 23: 771, 1981





























