Embed Size (px)
Citation preview

ORIGINAL PAPER
Altered fast- and slow-twitch muscle fibre characteristicsin female mice with a (S248F) knock-in mutation of the brainneuronal nicotinic acetylcholine receptor
David J. Cannata Æ David I. Finkelstein ÆIlse Gantois Æ Yaroslav Teper ÆJohn Drago Æ Jan M. West
Received: 5 January 2009 / Accepted: 15 April 2009 / Published online: 29 April 2009
� Springer Science+Business Media B.V. 2009
Abstract We generated a mouse line with a missense
mutation (S248F) in the gene (CHRNA4) encoding the a4
subunit of neuronal nicotinic acetylcholine receptor
(nAChR). Mutant mice demonstrate brief nicotine induced
dystonia that resembles the clinical events seen in patients
with the same mutation. Drug-induced dystonia is more
pronounced in female mice, thus our aim was to determine
if the S248F mutation changed the properties of fast- and
slow-twitch muscle fibres from female mutant mice.
Reverse transcriptase-PCR confirmed CHRNA4 gene
expression in the brain but not skeletal muscles in normal
and mutant mice. Ca2? and Sr2? force activation curves
were obtained using skinned muscle fibres prepared from
slow-twitch (soleus) and fast-twitch (EDL) muscles. Two
significant results were found: (1) the (pCa50 - pSr50)
value from EDL fibres was smaller in mutant mice than in
wild type (1.01 vs. 1.30), (2) the percentage force produced
at pSr 5.5 was larger in mutants than in wild type (5.76 vs.
0.24%). Both results indicate a shift to slow-twitch char-
acteristics in the mutant. This conclusion is supported by
the identification of the myosin heavy chain (MHC) iso-
forms. Mutant EDL fibres expressed MHC I (usually only
found in slow-twitch fibres) as well as MHC IIa. Despite
the lack of spontaneous dystonic events, our findings sug-
gest that mutant mice may be having subclinical events or
the mutation results in a chronic alteration to muscle neural
input.
Keywords Ca2?/Sr2?-activation � Dystonia �Nicotinic acetylcholine receptor � S248F mutation �Skinned muscle fibre
Introduction
We have generated a mouse model of autosomal dominant
nocturnal frontal lobe epilepsy [ADNFLE] (Scheffer et al.
1995) originally described to be due to a missense mutation
(S248F) in the gene (CHRNA4) encoding the a4 subunit of
neuronal nicotinic acetylcholine receptor [nAChR] (Stein-
lein et al. 1995). Although spontaneous seizures are not
observed in these knock-in mice, they do exhibit nicotine-
induced behaviours that mimic the brief dystonic events
reported in ADNFLE (Teper et al. 2007).
Brain nAChRs are ligand-gated ion channels which can
be divided into muscle and neuronal receptor types. These
receptors are involved in fast synaptic transmission and are
located in both the peripheral and central nervous system
(Hogg et al. 2003). The receptors are composed of five
subunits surrounding a central pore and are made up of
different combinations of the acetylcholine binding sub-
units (a2–a10 and b2–b4; Hogg et al. 2003). The nAChRs
Electronic supplementary material The online version of thisarticle (doi:10.1007/s10974-009-9177-x) contains supplementarymaterial, which is available to authorized users.
D. J. Cannata (&) � J. M. West
School of Life and Environmental Sciences, Deakin University,
221 Burwood Highway, Melbourne, VIC 3125, Australia
e-mail: [email protected]
D. I. Finkelstein
The Mental Health Research Institute of Victoria, Melbourne,
VIC 3052, Australia
I. Gantois
Laboratory of Biological Psychology, Katholieke Universiteit
Leuven, Tiensestraat 102, 3000 Leuven, Belgium
Y. Teper � J. Drago
Howard Florey Institute, The University of Melbourne,
Melbourne, VIC 3010, Australia
123
J Muscle Res Cell Motil (2009) 30:73–83
DOI 10.1007/s10974-009-9177-x

with a ratio of (three) a4:(two) b2 subunit stoichiometry
are only found in the brain (Hogg et al. 2003); nAChRs
found in the peripheral nervous system including skeletal
muscles of adult animals lack a4 or b2 subunits, thus a
mutation in the a4 or b2 subunit of the nAChRs should
only directly involve receptors expressed in the brain
(Phillips et al. 1995; Wong et al. 2002).
In ADNFLE, the functional changes in the receptor
complex results from mutations in either the gene encoding
the a4 (CHRNA4) or b2 subunit (CHRNB2) of the nAChR
(Phillips et al. 1995; Wong et al. 2002). Five mutations
have been identified associated with an ADNFLE disease
phenotype, three in the a4 and two in the b2 subunit (Hogg
et al. 2003).
The S248F mutation of CHRNA4 causes the nAChR to
display an abnormal desensitization profile. The wild type
receptor does not desensitize whereas the S248F receptor
does (Teper et al. 2007). Synaptosome function was inves-
tigated in wild type, heterozygous S248F and homozygous
S248F mutant mice by assessing the release of 86Rb?,
GABA and dopamine from synaptosomes. The EC50 was
reduced in a gene dose dependent manner in mutant mice
suggesting that the mutation confers an increased sensitivity
to neuronal nicotine receptor agonists. The net biological
effect is difficult to predict and depends on the location of
the receptor on the neuron (i.e., pre-synaptic or post-syn-
aptic), the neurotransmitter species of the neuron and the
location of the neuron within a neural circuit (i.e., inhibitory
or excitatory). The S248F mutant mouse was generated to
address these complex questions in an in vivo context.
S248F mutant mice were shown to exhibit a stereotyp-
ical nicotine-induced dystonic arousal complex [DAC]
(Teper et al. 2007). The DAC is brief in duration and
involves exploratory-type side-to-side head and body
darting, dystonic forelimb posturing and an accentuated
Straub tail. In some cases, the dystonic forelimb posturing
is associated with digital clasping (Teper et al. 2007).
Although not highlighted in the original description of the
model, the DAC also involved bilateral hind limb midline
abduction (see supplementary video) resulting in the
hindquarters coming into close contact with the ventral
surface and at times rolling to the side. Despite the robust
nature of the DAC, S248F mice were not observed to have
spontaneous behavioural events. The DAC was not asso-
ciated with a change in the electroencephalogram and there
were no electroencephalographic changes in S248F mice
monitored over an extended period (Teper et al. 2007). The
DAC is considered a valid model of the dystonic episodes
seen in ADNFLE because of the phenotypic similarities
and the observation that both the DAC and the human
attacks are ameliorated by the prototypical anticonvulsant
carbamazepine and by the administration of low dose
nicotine.
We had shown in a Parkinson’s disease model that
changes are readily detected in contralateral muscles fol-
lowing a partial lesion of the substantia nigra using the
neurotoxin 6-hydroxydopamine. These changes were
identified in muscles despite the lack of abnormality in
spontaneous motor function (Sliwinski et al. 2005). As
there were no spontaneous dystonic events in S248F
knock-in mice we therefore sought to use a range of well
established assays of hind limb muscle physiology to
determine if the primary S248F brain mutation was asso-
ciated with changes in parameters indicative of sustained
alteration in neural output. This conclusion could be drawn
as CHRNA4 is not expressed in muscle and any changes
identified could be linked to changes in brain activity.
In this study we used female S248F mice as these ani-
mals have a higher penetrance of DAC (Teper et al. 2007).
In a recent study, 100% of the female animals displayed all
three previously described component behaviours of DAC
(saccadic behaviour, Straub tail and fore-limb dystonia),
compared to only 75% of male animals (Teper et al. 2007).
Females also obtained significantly higher scores than
males in all three DAC behaviours (Teper et al. 2007).
The pattern of motor neuron activation of skeletal
muscles exerts a powerful influence on muscle contractile
speed (Syrovy 1987). In vivo, rat slow-twitch muscles
(e.g., the soleus) are stimulated by a tonic frequency in the
range of 20–25 Hz and contract and relax slowly (Gun-
dersen et al. 1988). In contrast, fast-twitch muscles such as
the extensor digitorum longus (EDL) have considerably
faster twitch contraction and relaxation times and are
subject to phasic stimulation over the range of 60–90 Hz
(Hennig and Lomo 1985). These differences in neural
activation influence the distinction observed in the func-
tional properties of the muscle fibres.
The effect of changing the neural input on the phenotype
of the muscle has been extensively studied using various
experimental techniques. Cross-reinnervation studies ele-
gantly showed that when the slow-twitch soleus muscle
was reinnervated with a motor neuron previously supplying
a fast-twitch muscle the contractile speed of the slow-
twitch muscle became faster (Buller et al. 1960; Close
1969). Similarly, when the fast-twitch muscle was inner-
vated with a nerve previously supplying the soleus muscle
the speed of contraction slowed (Buller et al. 1960; Close
1969). Cross-reinnervation also resulted in changes in the
expression of the myofibrillar protein isoforms (Hoh 1975)
and their distribution within a particular fibre type (Pette
and Staron 1990).
Chronic low frequency stimulation mimics the tonic
nerve impulse patterns that a slow-twitch muscle receives
in the rabbit (10 Hz) and converts fast-twitch fatigable
muscles into slower contracting muscles that are more
fatigue resistant (Salmons and Vrbova 1969; Salmons and
74 J Muscle Res Cell Motil (2009) 30:73–83
123

Sreter 1976). Conversely, a slow-twitch muscle that has
undergone a transition to fast via cross-reinnervation can
revert back to its slow-twitch properties when stimulation
is used to restore tonic activity (Salmons and Sreter 1976).
The presence of changes in muscle tone in extrapyra-
midal diseases such as Parkinson’s disease and its modu-
lation by basal ganglia surgery is well recognised (Hayashi
et al. 2001). A number of other studies have investigated
the complex interaction between volitional movement
originating in the motor cortex and inputs from the basal
ganglia and brain stem on neural tone (Takakusaki et al.
2003, 2004b, 2004c) whereby descending pathways ulti-
mately influence muscle tone by altering excitability of
motor neurons (Takakusaki et al. 2004a). There are only a
few studies that directly investigated the secondary effects
in muscles at the physiological, histological or biochemical
level in humans with central nervous system diseases such
as Parkinson’s disease or rarer basal ganglia diseases
associated with limb dystonia such as dentatorubral pal-
lidoluysian atrophy and primary dystonia (Edstrom et al.
1979; Rossi et al. 1996; Cox et al. 2000; Sharott et al.
2008). Results from these studies suggest that chronic
changes in centrally mediated neural tone can impact on
muscle phenotype.
The aim of this study was to assess changes in the
properties of individual fast- and slow-twitch fibres from
the hind limb muscles of wild type and S248F knock-in
mice as an indirect measure of altered peripheral neural
tone. This is a unique study as it observes changes in
muscle fibre properties by possibly altering neural impulse
activity without electrical stimulation or physical detach-
ment/reattachment of motor neurons. Any changes
observed in muscle fibre properties are the result of the
S248F mutation of the nAChR in the brain and not from
any mechanical or surgical intervention.
Materials and methods
Ethical approval
All procedures involving transgenic and wild type mice con-
formed to the Australian National Health and Medical Research
Council (NHMRC) code of practice and were approved by the
Howard Florey Institute Animal Ethics Committee. All pre-
cautions were taken to minimise pain or discomfort.
Animals and tissue collection
The mouse model used in this study was developed and
housed at the Howard Florey Institute, Melbourne Uni-
versity (Teper et al. 2007). Mice had access to food and
water ad libitum. The development and genetic background
of these mice has been previously described (Teper et al.
2007). Briefly, heterozygous mice from the knock-in strain
(S248F mutated CHRNA4 gene expressed) and their cor-
responding wild type counterparts were on a 50:50 (129:
CD1) genetic background (Teper et al. 2007). PCR analysis
was used to genotype the knock-in mice. A total of eigh-
teen female animals (9, S248F knock-in; 9, wild type) were
used in this study. Mice were eight to twelve months old
and were drug naıve for this study.
On the day of tissue collection, mice were killed by
cervical dislocation, the entire hind limb was skinned,
dissected and placed directly into storage solution con-
taining 50% glycerol and 50% of relaxing solution con-
taining (mmol l-1): HEPES, 20; ATP, 2; MgCl2, 3; EGTA,
10; Propionic acid, 150; and stored at -20�C until use
(West et al. 1999).
RT-PCR
A qualitative reverse transcriptase-polymerase chain reac-
tion (RT-PCR) was performed to confirm the tissue loca-
tion of CHRNA4 containing nAChRs. Total RNA isolation
from tissue samples of the brain, soleus and EDL muscles
were obtained using a combined RNeasy Micro Kit (Qia-
gen) with TRIzolTM Reagent (Life technologies-BRL)
method. Complementary DNA synthesis was obtained
using the SuperScriptTM First-Strand Synthesis System for
RT-PCR (Invitrogen). Actin primers were used to confirm
the integrity of RNA in the samples (data not shown).
A buffer only negative control was run with all samples
to confirm that samples were not contaminated (data
not shown). The forward primer had the sequence
ATGTCACCTCCATCCGCATCC and the reverse primer
had the sequence AGATCATGGTGAAGAGCAG. These
primers are located at position 430 and 1,056 bp on the
cDNA sequence of NCBI BC053013.
Preparation of skinned fibres
Hind limbs taken from the storage solution were washed
with relaxing solution (solution A, Table 1) and the soleus
and EDL muscles were dissected. The muscles were thor-
oughly blotted and pinned under slight tension on a Sylgard
plate and submerged in paraffin oil. The plasma membrane
of the muscle fibres had been rendered permeable by pro-
longed exposure to glycerol in the storage solution. Single
fibres were isolated from the muscles, by teasing them from
the muscle using fine forceps (No. 5). Individual fibres
were then mounted between two pins of a force recording
apparatus using surgical silk (Deknatel, 9.0, USA) as pre-
viously described (West et al. 1999). One pin was attached
to a micromanipulator and the other to a force transducer
(SensorNor AME-802, Norway).
J Muscle Res Cell Motil (2009) 30:73–83 75
123

The dimensions (length and diameter) of individual
fibres were measured under paraffin oil. The fibre was then
removed from the paraffin oil, blotted with filter paper and
placed in a spectrophotometric vial containing solution A
(Table 1). Sarcomere length was determined by passing
the beam of a HeNe laser (JDS Uniphase, USA) through
the fibre as previously described (West et al. 1999). The
average sarcomere length for the fibres in each experi-
mental group has been summarised in Table 2.
Solutions to activate skinned fibres
Solutions used to activate and relax the muscle fibres have
been previously described (Ashley and Moisescu 1977;
West and Stephenson 1993) and are summarised in Table 1.
Previously determined apparent affinity constants (Kapp)
were used (Ashley and Moisescu 1977). The amount of free
EGTA (i.e., not complexed with Ca2? or Sr2?) in each
solution was determined by titration (Ashley and Moisescu
1977). Solutions were made containing Ca2? at concen-
trations between 3.88 9 10-8 and 2.55 9 10-5 M L-1
(pCa = - log10[Ca2?] between 7.41 and 4.59) and Sr2? at
concentrations between 2.35 9 10-8 and 4.65 9 10-4
M L-1 (pSr = -log10[Sr2?] between 7.62 and 3.33).
Experimental protocol
Single muscle fibres from adult fast- and slow-twitch
muscle can be identified physiologically based on their
sensitivity to Ca2? and Sr2? when the fibres are activated
(Fink et al. 1986, 1990; West and Stephenson 1993; Lynch
et al. 1995; West et al. 1999). Fast-twitch muscles typically
have a greatly different sensitivity to Ca2? and Sr2?,
whereas the sensitivity of the contractile apparatus to these
two divalent cations in slow-twitch muscles is similar.
Thus, Sr2? is a physiological tool used to characterise
individual muscle fibres as either fast- or slow-twitch.
Fibres were activated using the previously described
‘‘Ca2?/Sr2?-jump-technique’’ (Stephenson and Williams
1982). Each fibre was activated in a series of activating
solutions, containing increasing concentrations of Ca2? or
Sr2? (Moisescu 1976; Stephenson and Williams 1982) and
the force output recorded on a chart recorder (WR-3701,
Graphtec, Japan). Once a steady state force level was
achieved in one solution the fibre was then rapidly trans-
ferred to the next activating solution until a maximum
activation force response was obtained. To accommodate
any decrease in the ability of the fibres to develop force
during a staircase activation, a maximum force response
with either Ca2? or Sr2? was determined at the beginning
and end of a series of contractions by submerging the fibre
in solution containing sufficient Ca2? or Sr2? to maximally
activate (Po) the fibre (pCa, 4.59; pSr, 3.33). If the initial
and final forces differed by more than 10% the results were
discarded. If the decrease in force from the initial to the
final activation was \10%, force was assumed to have
declined linearly with time and the force measured at each
sub-maximal Ca2? and Sr2? concentration was normalised
to the estimated maximum force response at that time
(Rees and Stephenson 1987). Each fibre was activated in
both sets of solutions containing either Ca2? or Sr2?. The
order in which fibres were activated (i.e., Ca2? then Sr2? or
vice versa) was divided equally among preparations to
avoid any systematic effects due to the order of presenta-
tion. Maximum Ca2?- or Sr2?-activated force was
determined from the first maximum activation with the
respective divalent cation in each experimental run.
All experiments were conducted at room temperature
(23–25�C).
Data analysis for skinned fibre experiments
Steady-state isometric force production was measured over
a range of Ca2? and Sr2? concentrations. Force developed
when activated by either Ca2? or Sr2? has been expressed
relative to the maximum force (Po) measured using that
ion. This relationship between the force output (Pr = P/Po)
and the concentration of Ca2? and Sr2? can be described
by the Hill equation (Hill 1910); Pr = [x]n/([x]n50 ? [x]n),
where [x] = concentration of either Ca2? or Sr2?; n =
the Hill coefficient which is proportional to the slope of the
curves relating to Pr to pCa or pSr and [x]50 = the
Table 1 Composition of solutions for skinned fibres
Solution [K?] [EGTA] [HDTA] [Mg2?]total [Ca]total [Ca2?]free [Sr]total [Sr2?]free
A 117 50 – 10.3 – \10-6 – –
B 117 50 – 8.12 49.5 0.02 – –
H 117 0.2 49.8 8.51 – \1.7 9 10-6 – –
S 114 50 – 5.95 – – 40 0.283
Concentrations are in (mmol l-l). All solutions contained (mmol l-l): Na?, 36; HEPES, 60; total ATP, 8; creatine phosphate (CP), 10; sodium
azide (NaN3), 1; Mg2?, 1. pH was 7.10 ± 0.01 at 23–25�C. A = relaxing, B = activating (containing Ca2?), H = pre-activating, S = activating
(containing Sr2?). 1-6-diaminohexane-N-N-N0,N0-tetraacetic acid (HDTA). Solutions were titrated using a pH metric method (Ashley and
Moisescu 1977)
76 J Muscle Res Cell Motil (2009) 30:73–83
123

concentration that produces 50% of the maximum force
response when activated by either Ca2? or Sr2?.
All the data in this study have been presented with
relative force expressed as a function of pCa or pSr. Curves
were fitted to the experimental data using a Marquardt non-
linear regression algorithm using SigmaPlot 9.0. The
quantitative characteristics obtained when activating the
fibres with the physiological cations Ca2? and Sr2? are:
pCa10 (pSr10); the concentration of Ca2? (Sr2?) that
produces 10% of the maximum Ca2?- (Sr2?)-activated
force, giving an indication of the concentration of these
divalent cations that produces threshold levels of force.
pCa50 (pSr50); the concentration of Ca2? (Sr2?) that
produces 50% of the maximum Ca2?- (Sr2?)-force, which
is indicative of the sensitivity of the contractile apparatus
to these divalent cations.
nCa (nSr); a parameter reflecting the maximum slope of
the force–pCa (force–pSr) curve.
Parameters used for characterising muscle fibres based
on their activation profiles
The difference in Sr2? sensitivity of fast- and slow-twitch
fibres is directly related to the different Troponin C (TnC)
isoforms that are expressed in fast- and slow-twitch fibres
(O’Connell et al. 2004b). In this study the following
quantitative characteristics were used to classify the fibres
as fast- or slow twitch (1) pCa50 - pSr50; the difference
between the midpoints of the force–pCa (pCa50) and force–
pSr curves (pSr50). This quantifies the relative sensitivity of
the fibre to Ca2? and Sr2?. To accurately determine this
parameter, it was necessary to obtain a complete activation
profile for Ca2? and Sr2?. (2) The level of activation when
the concentration of Sr2? in the solution is 3.0 9
10-6 M L-1 (pSr 5.5). At this level of activation with Sr2?
slow-twitch fibres produce a force response at least 70% of
the maximum Sr2?-activated force whereas the fast-twitch
fibres produce less than 1% of the maximum Sr2?-activated
force.
Thus using our classification scheme a fibre dissected
from the EDL muscle was classified as fast-twitch, if
the pCa50 - pSr50 value was greater than 1.25 log units
and the force produced at pSr 5.5 was less than 1% of
the maximum Sr2?-activated force. A fibre dissected
from the soleus muscle was classified as slow-twitch, if
the pCa50 - pSr50 value was less than 0.75 log units
and the pSr 5.5 value was greater than 70% of the
maximum Sr2?-activated force. If fibres did not meet
the classification criteria for both parameters they were
considered to be neither fast- or slow-twitch. Fibres
dissected from the soleus muscle that met the criteria of
being classified as a fast-twitch fibre were not used in
the final analysis.
Myosin heavy chain separation
Myosin heavy chain (MHC) isoforms were separated via
SDS–PAGE, using a previously described method (Tal-
madge and Roy 1993). Briefly, single muscle fibres were
dissected from the EDL and soleus that were stored in 50%
glycerol and 50% relaxing solution, and were placed in a
SDS-reducing buffer (62.5 mM Tris–HCl (pH 6.8), 12.5%
Glycerol (v/v), and 2.3% SDS (w/v)) overnight at room
temperature (23–25�C). Single fibres were pooled from
three animals for analysis. Samples were then heated to
94�C for 5 min and stored at -20�C until required. MHC
separation was carried out using a Bio-Rad mini-Protean
III cell apparatus at 110 V (constant) at 4�C for 24 h.
Bands were visualised using a silver-staining kit (161-
0443, Bio-Rad).
Results
RT-PCR
Figure 1 shows RT-PCR analysis of the CHRNA4 gene.
This clearly shows that the nAChRs with the a4 subunit are
only located in the brain of the animal models. Expression
of the gene is evident by the strong band displayed at
626 bp. None of the other tissues (knock-in—EDL, soleus;
and wild type—EDL, soleus) expressed the CHRNA4
WT EDL
WT SOL
KIEDL
KISOL
KIBrain
Lane 1
600bp
100bp
Fig. 1 RT-PCR analysis of the CHRNA4 gene. WT wild-type, KI
knock-in, EDL extensor digitorum longus, SOL soleus. Lane 1 is
loaded with a 100 bp protein ladder. The only expression of the
CHRNA4 gene is evident in the lane loaded with brain tissue from the
knock-in mouse (626 bp)
J Muscle Res Cell Motil (2009) 30:73–83 77
123

gene. We used an intron spanning molecular amplification
paradigm to determine that mRNA rather than genomic
DNA was being amplified. The forward primer is com-
plementary to sequence located in exon 4 and the reverse
primer is complementary to sequence in exon 5.
Ca2?- and Sr2?- activation profiles of soleus and EDL
fibres from wild type animals
Typical activation profiles of a slow-twitch and a fast-
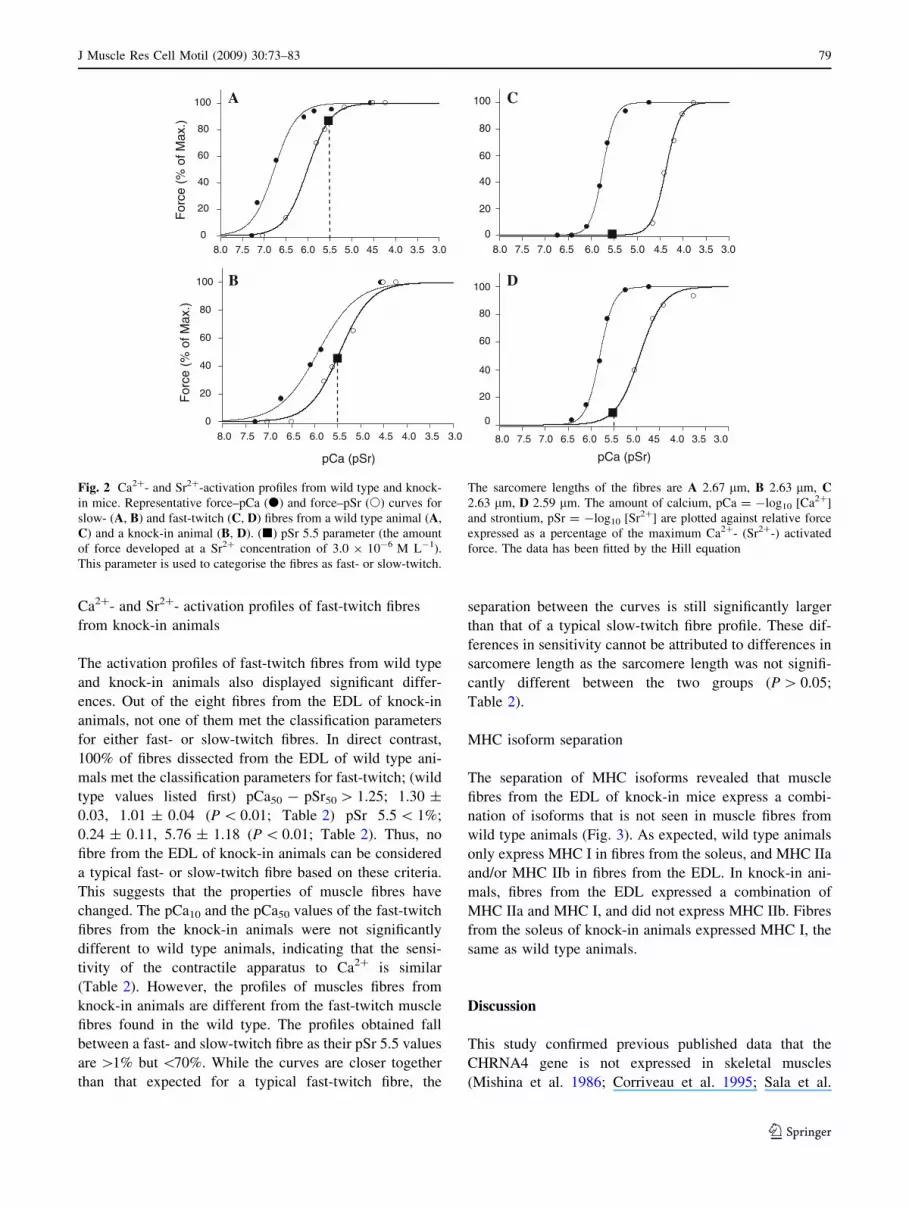
twitch fibre from a wild type animal are shown in Fig. 2A,
C. A summary of the parameters measured from wild type
and knock-in animals are shown in Table 2. In wild type
animals, all fast-twitch fibres from the EDL (n = 5) and all
slow-twitch fibres from the soleus (n = 5) met the classi-
fication criteria described in the method; fast-twitch fibres;
pCa50 - pSr50 [ 1.25 (1.30 ± 0.03) and pSr 5.5 \ 1%
(0.24 ± 0.11); slow-twitch; pCa50 - pSr50 \ 0.75 (0.55 ±
0.15) and pSr 5.5[75% (84.25 ± 2.53). In addition acti-
vation profiles of both fibre-types were consistent with other
well established parameters used to help identify fibre types
from other mammalian skeletal muscles (Lynch et al. 1995;
West et al. 1999; Bortolotto et al. 2000); the force–pCa curves
were steeper in fast-twitch fibres (nCa, 2.87 ± 0.21) than in
slow-twitch fibres (nCa, 1.51 ± 0.30; P\0.01; Table 2); and
slow-twitch fibres activated at lower concentrations of Ca2?
(pCa10) i.e., higher pCa values (7.36 ± 0.06) than fast-twitch
fibres (6.12 ± 0.06; P\0.01; Table 2).
Ca2?- and Sr2?- activation profiles of slow-twitch fibres
from knock-in animals
Activation profiles of the slow-twitch fibres from the wild
type and knock-in animals are significantly different
(Fig. 2A, B). According to the classification criteria, five of
the six slow-twitch fibres from knock-in animals had a
pCa50 - pSr50 value\0.75, but none of them had a pSr 5.5
value greater than 70%. Thus based on our scheme, none of
the fibres from the soleus of knock-in animals can be
considered typical slow-twitch nor can they be considered
fast-twitch as they do not meet the typical fast-twitch
classification either. In contrast, all fibres dissected from
the soleus of wild type animals can be considered typical
slow-twitch fibres based on our classification. Both force–
pCa and force–pSr curves of the fibres from knock-in
animals are shifted to the right (Fig. 2B) of the curves
obtained from the wild type animals (Fig. 2A). This indi-
cates that the contractile apparatus of the fibres from
knock-in animals is less sensitive to both Ca2? and Sr2?.
The pCa50 - pSr50 values of the slow-twitch fibres from
the knock-in and wild type animals were not significantly
different (P [ 0.05; Table 2). As the entire curves have
shifted to the right, the pCa10 value from fibres in the
knock-in animals was significantly less (7.18 ± 0.08;
P \ 0.01; Table 2) than wild type animals (7.36 ± 0.06;
Table 2). This means a higher concentration of Ca2? (pCa
7.18 = 6.6 9 10-8 M L-1) was needed to produce 10% of
the maximum force in the muscles from knock-in animals
than wild type animals (pCa 7.36 = 4.3 9 10-8 M L-1).
The pCa50 values for the slow-twitch fibres from the
knock-in animals were also significantly lower than those
from wild animals (P \ 0.05). A similar trend was shown
for Sr2?-activation properties (where pSr 5.5 decreased
from 84.25 to 54.16%; P \ 0.05; Table 2). The difference
in sensitivity of the fibres to Ca2? and Sr2? observed in this
study, cannot be due to different sarcomere lengths of the
fibres between groups (not significantly different P [ 0.05;
Table 2), which is known to have a significant effect on the
sensitivity of the fibres to Ca2? and Sr2? (Stephenson and
Williams 1982).
Table 2 Activation parameters of individual fibres from knock-in and wild type animals
Parameters Slow-twitch Fast-twitch
Wild-type Knock-in Wild-type Knock-in
pCa10 7.36 ± 0.06 7.18 ± 0.08* 6.12 ± 0.06 6.08 ± 0.03
pCa50 6.59 ± 0.18 6.00 ± 0.12** 5.78 ± 0.04 5.80 ± 0.01
nCa 1.51 ± 0.30 0.85 ± 0.09* 2.87 ± 0.21 3.62 ± 0.32*
pSr 5.5 (% force) 84.25 ± 2.53 54.16 ± 5.47* 0.24 ± 0.11 5.76 ± 1.18**
pCa50 - pSr50 0.55 ± 0.15 0.44 ± 0.09 1.30 ± 0.03 1.01 ± 0.04**
Max force (Ca/Sr) 0.96 ± 0.06 1.03 ± 0.05 0.99 ± 0.03 1.01 ± 0.05
SL (lm) 2.69 ± 0.30 2.56 ± 0.11 2.67 ± 0.05 2.56 ± 0.05
Values represented are the mean ± SE. Number of fibres from knock-in animals (soleus n = 6, EDL n = 8); number of fibres from wild type
animals (soleus n = 5, EDL n = 5). * P \ 0.05, ** P \ 0.01 indicates significant differences between wild type and knock-in groups for the
same muscle type. Max force (Ca/Sr) = maximum Ca2?-activated force/maximum Sr2?-activated force. This shows there is no significant
difference between the amount of force produced when maximally activated by Ca2? or Sr2?. SL sarcomere length. All slow-twitch fibres were
dissected from the soleus. All fast-twitch fibres were dissected from the EDL
78 J Muscle Res Cell Motil (2009) 30:73–83
123
Ca2?- and Sr2?- activation profiles of fast-twitch fibres
from knock-in animals
The activation profiles of fast-twitch fibres from wild type
and knock-in animals also displayed significant differ-
ences. Out of the eight fibres from the EDL of knock-in
animals, not one of them met the classification parameters
for either fast- or slow-twitch fibres. In direct contrast,
100% of fibres dissected from the EDL of wild type ani-
mals met the classification parameters for fast-twitch; (wild
type values listed first) pCa50 - pSr50 [ 1.25; 1.30 ±
0.03, 1.01 ± 0.04 (P \ 0.01; Table 2) pSr 5.5 \ 1%;
0.24 ± 0.11, 5.76 ± 1.18 (P \ 0.01; Table 2). Thus, no
fibre from the EDL of knock-in animals can be considered
a typical fast- or slow-twitch fibre based on these criteria.
This suggests that the properties of muscle fibres have
changed. The pCa10 and the pCa50 values of the fast-twitch
fibres from the knock-in animals were not significantly
different to wild type animals, indicating that the sensi-
tivity of the contractile apparatus to Ca2? is similar
(Table 2). However, the profiles of muscles fibres from
knock-in animals are different from the fast-twitch muscle
fibres found in the wild type. The profiles obtained fall
between a fast- and slow-twitch fibre as their pSr 5.5 values
are [1% but \70%. While the curves are closer together
than that expected for a typical fast-twitch fibre, the
separation between the curves is still significantly larger
than that of a typical slow-twitch fibre profile. These dif-
ferences in sensitivity cannot be attributed to differences in
sarcomere length as the sarcomere length was not signifi-
cantly different between the two groups (P [ 0.05;
Table 2).
MHC isoform separation
The separation of MHC isoforms revealed that muscle
fibres from the EDL of knock-in mice express a combi-
nation of isoforms that is not seen in muscle fibres from
wild type animals (Fig. 3). As expected, wild type animals
only express MHC I in fibres from the soleus, and MHC IIa
and/or MHC IIb in fibres from the EDL. In knock-in ani-
mals, fibres from the EDL expressed a combination of
MHC IIa and MHC I, and did not express MHC IIb. Fibres
from the soleus of knock-in animals expressed MHC I, the
same as wild type animals.
Discussion
This study confirmed previous published data that the
CHRNA4 gene is not expressed in skeletal muscles
(Mishina et al. 1986; Corriveau et al. 1995; Sala et al.
3.03.54.04.55.05.56.06.57.07.58.0
For
ce (
% o
f Max
.)
0
20
40
60
80
100
pCa (pSr) pCa (pSr)
3.03.54.04.55.05.56.06.57.07.58.0 3.03.54.04.55.05.56.06.57.07.58.0
3.03.54.04.55.05.56.06.57.07.58.0
For
ce (
% o
f Max
.)
0
20
40
60
80
100
0
20
40
60
80
100
0
20
40
60
80
100 DB
A C
Fig. 2 Ca2?- and Sr2?-activation profiles from wild type and knock-
in mice. Representative force–pCa (d) and force–pSr (s) curves for
slow- (A, B) and fast-twitch (C, D) fibres from a wild type animal (A,
C) and a knock-in animal (B, D). (j) pSr 5.5 parameter (the amount
of force developed at a Sr2? concentration of 3.0 9 10-6 M L-1).
This parameter is used to categorise the fibres as fast- or slow-twitch.
The sarcomere lengths of the fibres are A 2.67 lm, B 2.63 lm, C2.63 lm, D 2.59 lm. The amount of calcium, pCa = -log10 [Ca2?]
and strontium, pSr = -log10 [Sr2?] are plotted against relative force
expressed as a percentage of the maximum Ca2?- (Sr2?-) activated
force. The data has been fitted by the Hill equation
J Muscle Res Cell Motil (2009) 30:73–83 79
123

1996; Hogg et al. 2003). The assay used was designed to
detect both the mutant and wild type allele and we there-
fore also conclude that there is no aberrant expression of
the knock-in mutation in muscle of adult mice. Skeletal
muscles express a4 subunits transiently during very early
development (Corriveau et al. 1995). In this study, muscle
samples were taken from adult animals; thus the changes
observed in the properties of the skeletal muscles are
probably the result of the altered function of brain
expressed nAChRs. The skeletal muscles are not directly
affected by the S248F mutation as they do not express the
CHRNA4 gene. Transient expression of the mutant allele
during early development is unlikely to have a sustained
effect evident on analysis of adult muscle tissue.
The major finding in this study was that a mutation in
the nAChRs expressed in the brain induced changes in the
properties of the fast- and slow-twitch skeletal muscle
fibres in knock-in animals. These changes suggest the
neural impulse activity has been altered, which has been
clearly established to play a role in determining muscle
phenotype (Salmons and Sreter 1976; Salmons 1980;
Salmons and Henriksson 1981). This finding is highly
significant in terms of our model of ADNFLE. Mice with
the S248F mutation do not display spontaneous clinical
events suggestive of seizures or episodes of dystonia. A
very large number of mice were assessed over a long
period both by video recording of behaviour and epidural
and/or depth electrode electroencephalography. Dystonic
events resembling the brief nocturnal events seen in attacks
of ADNFLE in humans were only seen in the setting of
nicotine injection (Teper et al. 2007). The finding of the
current study suggest that mutant mice may either be
experiencing brief dystonic events that are not apparent by
simple behavioural observation or that the mutation is
associated with chronic modulation of neural input to the
muscles.
Fast-twitch fibres from the EDL and slow-twitch fibres
from the soleus of wild type animals showed the charac-
teristic activation properties observed for mammalian
muscles, exhibiting a greatly different sensitivity to Ca2?
and Sr2? in fast-twitch fibres, and a similar sensitivity in
Ca2? and Sr2? in slow-twitch fibres (Fink et al. 1986;
Bortolotto et al. 1999; West et al. 1999). The activation
profiles from the muscle fibres of the EDL from knock-in
animals were clearly different and not typical of fast-twitch
fibre profiles seen in wild type animals. The fibres showed
an increase in sensitivity to Sr2? which resulted in a
decreased pCa50 - pSr50 value and a higher pSr 5.5 value;
hence no fibre was classified as a typical fast- or slow-
twitch. A smaller separation between the force–pCa and
force–pSr curve is indicative of a greater contribution of
the slow-twitch component to the activation profile, as it is
known single fibres can express both fast- and slow-twitch
isoforms (Pette and Schnez 1977; Rubinstein et al. 1978;
Salmons and Henriksson 1981). MHC separation con-
firmed fibres from the EDL of knock-in animals
co-expressed MHC slow (MHC I) and fast (MHC IIa)
isoforms. MHC isoforms have been shown to have a strong
influence on the force–pSr curves (Bortolotto et al. 2000).
The expression of MHC I and possibly the emergence of
other slow isoforms being expressed in these fibres may be
responsible for changes in the activation properties. Due to
these changes, fibres from the EDL of knock-in animals
could be considered intermediate fibres. An increase in the
number of intermediate fibres in a muscle is considered an
indicator that a muscle is undergoing remodelling even
if the function of the muscle is not greatly affected
(Stephenson 2006).
In contrast, a different effect was seen in the properties
of slow-twitch fibres. The activation profiles of fibres from
the soleus muscle of knock-in animals showed a dramatic
change in sensitivity to both Ca2? and Sr2?. At pSr 5.5 the
force expressed as a percentage of the maximum signifi-
cantly decreased by over 30%. This, along with no sig-
nificant difference in the distance between the curves
(pCa50 - pSr50) of knock-in and wild type animals indi-
cates a decreased sensitivity to both Ca2? and Sr2?. This
may be due to a change in the TnC isoforms expressed,
changing the sensitivity of the contractile apparatus to
Ca2? and Sr2? (Grabarek et al. 1992; O’Connell et al.
2004a). There was no difference in the MHC isoforms
expressed (both groups expressed MHC I only).
Changes in the activation characteristics of muscle fibres
are due to the expression of the contractile and regulatory
proteins. The contribution of the different proteins
expressed in a fibre to the relationship between force and
Ca2? and Sr2? has been studied using single fibres from an
SOLEUS EDL WT KI WT KI
* * ******
Fig. 3 Myosin heavy chain isoform expression of single fibres.
Myosin heavy chain (MHC) isoforms were separated via SDS–PAGE.
Wild type (WT); knock-in (KI); MHC I (*); MHC IIa (**); MHC IIb
(***).10 fibres from three animals were pooled for analysis of WT
and KI soleus. 14 fibres from three animals were pooled for analysis
of WT and KI EDL. Fibres from the soleus of both groups only
expressed MHC I. In the EDL, fibres from knock-in animals
expressed MHC I and MHC IIa only. Fibres from the EDL of wild
type animals expressed varying amounts of MHC IIa and/or MHC IIb
but never expressed MHC I
80 J Muscle Res Cell Motil (2009) 30:73–83
123

EDL and a soleus from the hind limbs of rats which had
been tied in either series or parallel (Lynch et al. 1995).
When two individual fast-twitch fibres were tied in parallel
the Ca2?- and Sr2?-activation curves were typical of the
profile for a single fast-twitch muscle fibre (Lynch et al.
1995). However, different profiles were obtained when a
single fast-twitch and a single slow-twitch fibre were tied
together in parallel. Theoretical curves were used to predict
the contribution of the fast and slow twitch components of
the curve. When the prediction was a 30% contribution
from fast-twitch and 70% from slow-twitch, the separation
between the force–pCa and force–pSr curves (pCa50 -
pSr50) was 0.38 log units compared to when the prediction
was 90% from a fast-twitch and 10% from a slow-twitch
fibre, where the separation between the force–pCa and
force–pSr curves was 1.18 log units. Therefore, a separa-
tion of 1.30 log units between the force–pCa and force–pSr
curves (observed in fast-twitch fibres of wild type animals)
is indicative of a fast-twitch component greater than 90%
and a slow-twitch component smaller than 10%. Fibres
from the EDL of knock-in animals had a pCa50 - pSr50
value of 1.01 ± 0.04; therefore it could be predicted that
this may result from a 75% contribution from fast-twitch
and a 25% contribution from slow-twitch.
Differences observed in the activation profiles are
probably the result from changes to a combination of iso-
forms and their interaction, rather than just a change to one
specific protein. For example, the change in sensitivity to
Ca2? and Sr2? is probably due to alterations of TnC iso-
forms being expressed and their interactions with the dif-
ferent MHC isoforms. TnC is the subunit to which Ca2?
and Sr2? bind to activate muscle fibre contraction. Tro-
ponin together with tropomyosin forms the Ca2? regulatory
complex of the thin filament. Troponin consists of three
subunits, the tropomyosin binding subunit, troponin T
(TnT), the inhibitory subunit, troponin I (TnI) and the
calcium binding subunit, troponin C (TnC). Isoforms for all
subunits exist for fast- and slow-twitch muscle fibres (Pette
and Staron 1990). TnC expressed with different TnT iso-
form(s) have been correlated with changes in the force–
pCa profiles (O’Connell et al. 2004a; Kischel et al. 2005)
as well as with the different combination of myosin heavy
chain isoforms (Piquet et al. 1997; Geiger et al. 1999;
O’Connell et al. 2004a). It has been established that the
isoforms expressed can be influenced by neural impulse
activity (Sreter et al. 1973; Brown et al. 1983).
The slope of the force–pCa curve was also significantly
different in the muscles from knock-in animals which
further suggests changes in the isoforms and subunits
expressed (Pette and Staron 1990; Bicer and Reiser 2004).
Different combinations of TnT isoforms in combination
with tropomyosin subunits are thought to determine the
slope of the force–pCa curve (Schachat et al. 1987).
Multiple isoforms for all the subunits of tropomyosin and
troponin exist for both slow- and fast-twitch muscles (Pette
and Staron 1990; West et al. 1999).
Our results provide evidence for a change in neural
impulse activity received by skeletal muscles associated
with a knock-in mutation in the a4 subunit of nAChRs
expressed in the brain. How the nAChRs induce a change
in neural impulse activity is not fully understood. It has
been shown that the S248F mutation results in a signifi-
cantly higher affinity of the nAChR to ACh as observed by
shifts in the EC50 values (Teper et al. 2007). This suggests
that the mutated receptors are more sensitive to ACh and
would be activated at lower concentrations of ACh.
These results are in line with the proposal that centrally
mediated neural tone is a major determinant of biochemical
and ultimately the biophysical properties of muscles.
Despite the lack of spontaneous dystonic events, our find-
ings suggest that mutant mice may be having subclinical
events or the mutation may chronically alter neural input to
the muscles.
Acknowledgments This study was supported by a project grant
from the National Health and Medical Research Council (NHMRC).
J.D. is a NHMRC practitioner fellow. Authors would like to thank Jim
Massalas (Howard Florey Institute) for his assistance with the S248F
mouse strain and Prof. George Stephenson (La Trobe University) for
helpful suggestions and critically reading the manuscript.
References
Ashley CC, Moisescu DG (1977) Effect of changing the composition
of the bathing solutions upon the isometric tension–pCa
relationship in bundles of crustacean myofibrils. J Physiol 270:
627–652
Bicer S, Reiser PJ (2004) Myosin light chain isoform expression among
single mammalian skeletal muscle fibers: species variations.
J Muscle Res Cell Motil 25:623–633. doi:10.1007/s10974-004-
5070-9
Bortolotto SK, Stephenson DG, Stephenson GM (1999) Fiber type
populations and Ca2?-activation properties of single fibers in
soleus muscles from SHR and WKY rats. Am J Physiol 276:
C628–C637
Bortolotto SK, Cellini M, Stephenson DG, Stephenson GM (2000)
MHC isoform composition and Ca(2?)- or Sr(2?)-activation
properties of rat skeletal muscle fibers. Am J Physiol Cell Physiol
279:C1564–C1577
Brown WE, Salmons S, Whalen RG (1983) The sequential replace-
ment of myosin subunit isoforms during muscle type transfor-
mation induced by long term electrical stimulation. J Biol Chem
258:14686–14692
Buller AJ, Eccles JC, Eccles RM (1960) Interactions between
motoneurones and muscles in respect of the characteristic speeds
of their responses. J Physiol 150:417–439
Close R (1969) Dynamic properties of fast and slow skeletal muscles
of the rat after nerve cross-union. J Physiol 204:331–346
Corriveau RA, Romano SJ, Conroy WG, Oliva L, Berg DK (1995)
Expression of neuronal acetylcholine receptor genes in verte-
brate skeletal muscle during development. J Neurosci 15:
1372–1383
J Muscle Res Cell Motil (2009) 30:73–83 81
123

Cox H, Costin-Kelly NM, Ramani P, Whitehouse WP (2000) An
established case of dentatorubral pallidoluysian atrophy
(DRPLA) with unusual features on muscle biopsy. Eur J Paediatr
Neurol 4:119–123. doi:10.1053/ejpn.2000.0279
Edstrom L, Gremski W, Wroblewski R (1979) Sulphur and
phosphorus content in relation to fibre composition and atrophy
of skeletal muscle in patients with Parkinson’s disease. J Neurol
Sci 41:311–323. doi:10.1016/0022-510X(79)90092-3
Fink RH, Stephenson DG, Williams DA (1986) Calcium and
strontium activation of single skinned muscle fibres of normal
and dystrophic mice. J Physiol 373:513–525
Fink RH, Stephenson DG, Williams DA (1990) Physiological proper-
ties of skinned fibres from normal and dystrophic (Duchenne)
human muscle activated by Ca2? and Sr2?. J Physiol 420:337–353
Geiger PC, Cody MJ, Sieck GC (1999) Force-calcium relationship
depends on myosin heavy chain and troponin isoforms in rat
diaphragm muscle fibers. J Appl Physiol 87:1894–1900
Grabarek Z, Tao T, Gergely J (1992) Molecular mechanism of
troponin-C function. J Muscle Res Cell Motil 13:383–393. doi:
10.1007/BF01738034
Gundersen K, Leberer E, Lomo T, Pette D, Staron RS (1988) Fibre
types, calcium-sequestering proteins and metabolic enzymes in
denervated and chronically stimulated muscles of the rat. J Physiol
398:177–189
Hayashi R, Hashimoto T, Tada T, Ikeda S (2001) Relation between
changes in long-latency stretch reflexes and muscle stiffness in
Parkinson’s disease—comparison before and after unilateral palli-
dotomy. Clin Neurophysiol 112:1814–1821. doi:10.1016/S1388-
2457(01)00642-3
Hennig R, Lomo T (1985) Firing patterns of motor units in normal
rats. Nature 314:164–166. doi:10.1038/314164a0
Hill AV (1910) A new mathematical treatment of changes of ionic
concentration in muscle and nerve under the action of electric
currents, with a theory as to their mode of excitation. J Physiol
40:190–224
Hogg RC, Raggenbass M, Bertrand D (2003) Nicotinic acetylcholine
receptors: from structure to brain function. Rev Physiol Biochem
Pharmacol 147:1–46. doi:10.1007/s10254-003-0005-1
Hoh JF (1975) Selective and non-selective reinnervation of fast-twitch
and slow-twitch rat skeletal muscle. J Physiol 251:791–801
Kischel P, Bastide B, Muller M, Dubail F, Offredi F, Jin JP, Mounier
Y, Martial J (2005) Expression and functional properties of four
slow skeletal troponin T isoforms in rat muscles. Am J Physiol
Cell Physiol 289(2):C437–C443
Lynch GS, Stephenson DG, Williams DA (1995) Analysis of Ca2?
and Sr2? activation characteristics in skinned muscle fibre
preparations with different proportions of myofibrillar isoforms.
J Muscle Res Cell Motil 16:65–78. doi:10.1007/BF00125311
Mishina M, Takai T, Imoto K, Noda M, Takahashi T, Numa S,
Methfessel C, Sakmann B (1986) Molecular distinction between
fetal and adult forms of muscle acetylcholine receptor. Nature 321:
406–411. doi:10.1038/321406a0
Moisescu DG (1976) Kinetics of reaction in calcium-activated skinned
muscle fibres. Nature 262:610–613. doi:10.1038/262610a0
O’Connell B, Nguyen LT, Stephenson GM (2004a) A single-fibre
study of the relationship between MHC and TnC isoform
composition in rat skeletal muscle. Biochem J 378:269–274. doi:
10.1042/BJ20031170
O’Connell B, Stephenson DG, Blazev R, Stephenson GM (2004b)
Troponin C isoform composition determines differences in
Sr(2?)-activation characteristics between rat diaphragm fibers.
Am J Physiol Cell Physiol 287:C79–C87. doi:10.1152/ajpcell.
00555.2003
Pette D, Schnez U (1977) Coexistence of fast and slow type myosin
light chains in single muscle fibres during transformation as
induced by long term stimulation. FEBS Lett 83:128–130. doi:
10.1016/0014-5793(77)80656-X
Pette D, Staron RS (1990) Cellular and molecular diversities of
mammalian skeletal muscle fibers. Rev Physiol Biochem
Pharmacol 116:1–76
Phillips HA, Scheffer IE, Berkovic SF, Hollway GE, Sutherland GR,
Mulley JC (1995) Localization of a gene for autosomal dominant
nocturnal frontal lobe epilepsy to chromosome 20q 13.2. Nat
Genet 10:117–118. doi:10.1038/ng0595-117
Piquet F, Stevens L, Butler-Browne GS, Mounier Y (1997) Contrac-
tile properties and myosin heavy chain composition of newborn
rat soleus muscles at different stages of postnatal development.
J Muscle Res Cell Motil 18:71–79. doi:10.1023/A:1018633017
143
Rees BB, Stephenson DG (1987) Thermal dependence of maximum
Ca2?-activated force in skinned muscle fibres of the toad Bufomarinus acclimated at different temperatures. J Exp Biol 129:
309–327
Rossi B, Siciliano G, Carboncini MC, Manca ML, Massetani R,
Viacava P, Muratorio A (1996) Muscle modifications in
Parkinson’s disease: myoelectric manifestations. Electroencep-
halogr Clin Neurophysiol 101:211–218. doi:10.1016/0924-980X
(96)94672-X
Rubinstein N, Mabuchi K, Pepe F, Salmons S, Gergely J, Sreter F
(1978) Use of type-specific antimyosins to demonstrate the
transformation of individual fibers in chronically stimulated
rabbit fast muscles. J Cell Biol 79:252–261. doi:10.1083/jcb.
79.1.252
Sala C, Kimura I, Santoro G, Kimura M, Fumagalli G (1996)
Expression of two neuronal nicotinic receptor subunits in
innervated and denervated adult rat muscle. Neurosci Lett 215:
71–74. doi:10.1016/0304-3940(96)12923-2
Salmons S (1980) Functional adaptation in skeletal muscle. Trends
Neurosci 3:134–137. doi:10.1016/0166-2236(80)90050-8
Salmons S, Henriksson J (1981) The adaptive response of skeletal muscle
to increased use. Muscle Nerve 4:94–105. doi:10.1002/mus.8800
40204
Salmons S, Sreter FA (1976) Significance of impulse activity in the
transformation of skeletal muscle type. Nature 263:30–34. doi:
10.1038/263030a0
Salmons S, Vrbova G (1969) The influence of activity on some
contractile characteristics of mammalian fast and slow muscles.
J Physiol 201:535–549
Schachat FH, Diamond MS, Brandt PW (1987) Effect of different
troponin T-tropomyosin combinations on thin filament activa-
tion. J Mol Biol 198:551–554. doi:10.1016/0022-2836(87)
90300-7
Scheffer IE, Bhatia KP, Lopes-Cendes I, Fish, Marsden CD, Ander-
mann E, Andermann F, Desbiens R, Keene D, Cendes F et al
(1995) Autosomal dominant nocturnal frontal lobe epilepsy. A
distinctive clinical disorder. Brain 118(Pt 1):61–73. doi:10.1093/
brain/118.1.61
Sharott A, Grosse P, Kuhn AA, Salih F, Engel AK, Kupsch A,
Schneider GH, Krauss JK, Brown P (2008) Is the synchroniza-
tion between pallidal and muscle activity in primary dystonia
due to peripheral afferance or a motor drive? Brain 131:473–484.
doi:10.1093/brain/awm324
Sliwinski A, Stanic D, Finkelstein DI, Ilic M, West JM, Dooley PC
(2005) Alterations in the proportions of skeletal muscle proteins
following a unilateral lesion to the substantia nigra pars compacta
of rats. J Muscle Res Cell Motil 26:149–155. doi:10.1007/
s10974-005-6833-7
Sreter FA, Gergely J, Salmons S, Romanul F (1973) Synthesis by fast
muscle of myosin light chains characteristic of slow muscle in
response to long-term stimulation. Nat New Biol 241:17–19
82 J Muscle Res Cell Motil (2009) 30:73–83
123

Steinlein OK, Mulley JC, Propping P, Wallace RH, Phillips HA,
Sutherland GR, Scheffer IE, Berkovic SF (1995) A missense
mutation in the neuronal nicotinic acetylcholine receptor alpha 4
subunit is associated with autosomal dominant nocturnal frontal
lobe epilepsy. Nat Genet 11:201–203. doi:10.1038/ng1095-201
Stephenson DG (2006) Tubular system excitability: an essential
component of excitation-contraction coupling in fast-twitch
fibres of vertebrate skeletal muscle. J Muscle Res Cell Motil
27:259–274. doi:10.1007/s10974-006-9073-6
Stephenson DG, Williams DA (1982) Effects of sarcomere length on
the force–pCa relation in fast- and slow-twitch skinned muscle
fibres from the rat. J Physiol 333:637–653
Syrovy I (1987) Isoforms of contractile proteins. Prog Biophys Mol
Biol 49:1–27. doi:10.1016/0079-6107(87)90007-1
Takakusaki K, Habaguchi T, Ohtinata-Sugimoto J, Saitoh K,
Sakamoto T (2003) Basal ganglia efferents to the brainstem
centers controlling postural muscle tone and locomotion: a new
concept for understanding motor disorders in basal ganglia
dysfunction. Neuroscience 119:293–308. doi:10.1016/S0306-
4522(03)00095-2
Takakusaki K, Habaguchi T, Saitoh K, Kohyama J (2004a) Changes in
the excitability of hindlimb motoneurons during muscular atonia
induced by stimulating the pedunculopontine tegmental nucleus
in cats. Neuroscience 124:467–480. doi:10.1016/j.neuroscience.
2003.12.016
Takakusaki K, Oohinata-Sugimoto J, Saitoh K, Habaguchi T (2004b)
Role of basal ganglia-brainstem systems in the control of postural
muscle tone and locomotion. Prog Brain Res 143:231–237. doi:
10.1016/S0079-6123(03)43023-9
Takakusaki K, Saitoh K, Harada H, Kashiwayanagi M (2004c) Role
of basal ganglia-brainstem pathways in the control of motor
behaviors. Neurosci Res 50:137–151. doi:10.1016/j.neures.
2004.06.015
Talmadge RJ, Roy RR (1993) Electrophoretic separation of rat
skeletal muscle myosin heavy-chain isoforms. J Appl Physiol
75:2337–2340
Teper Y, Whyte D, Cahir E, Lester HA, Grady SR, Marks MJ, Cohen
BN, Fonck C, McClure-Begley T, McIntosh JM, Labarca C,
Lawrence A, Chen F, Gantois I, Davies PJ, Petrou S, Murphy M,
Waddington J, Horne MK, Berkovic SF, Drago J (2007)
Nicotine-induced dystonic arousal complex in a mouse line
harboring a human autosomal-dominant nocturnal frontal lobe
epilepsy mutation. J Neurosci 27:10128–10142. doi:10.1523/
JNEUROSCI.3042-07.2007
West JM, Stephenson DG (1993) Ca2? and Sr2? activation properties
of skinned muscle fibres with different regulatory systems from
crustacea and rat. J Physiol 462:579–596
West JM, Barclay CJ, Luff AR, Walker DW (1999) Developmental
changes in the activation properties and ultrastructure of fast-
and slow-twitch muscles from fetal sheep. J Muscle Res Cell
Motil 20:249–264. doi:10.1023/A:1005433809414
Wong JY, Ross SA, McColl C, Massalas JS, Powney E, Finkelstein
DI, Clark M, Horne MK, Berkovic SF, Drago J (2002)
Proconvulsant-induced seizures in alpha(4) nicotinic acetylcho-
line receptor subunit knockout mice. Neuropharmacology 43:
55–64. doi:10.1016/S0028-3908(02)00067-9
J Muscle Res Cell Motil (2009) 30:73–83 83
123

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.





























