Embed Size (px)
Citation preview

149
Biogeographic Breaks in Vanuatu, a Nascent Oceanic Archipelago1
Alison M. Hamilton,2,3,5 Elaine R. Klein,4 and Christopher C. Austin2
Abstract: The study of distinct biogeographic demarcations has played a pivotalrole in our understanding processes responsible for patterns of species distribu-tions and, importantly, the role of geologic processes in promoting biotic diver-sification. Biogeographic barriers such as Wallace’s line have been shown to bethe result of old geologic processes shaping ancient faunal or floral diversi-fication events. Based on distributions of birds, bats, reptiles, plants, andinvertebrates we identify a distinct biogeographic disjunction in Vanuatu, a geo-logically nascent oceanic archipelago. We discuss mechanisms contributing tothis concordant pattern across these disparate taxonomic groups in light of geo-logic history, ocean currents, vegetation, soil, and bioclimatic data, and proposethe name Cheesman’s line to indicate the faunal and floral discontinuity be-tween the northern and southern islands of Vanuatu.
The distribution of plants and animalsamong the isolated, oceanic islands of thetropical Pacific has been of interest to evolu-tionary biologists for more than 150 yr(Sclater 1858, Wallace 1860, 1876). Disconti-nuity between distinct Asian and Australian
biotas has motivated a large body of research,resulting in designations of the western(Huxley’s line or Wallace’s line) and eastern(Lyddeker’s line) limits of a region of bioticoverlap between Asian and Australian faunas,a region generally referred to as Wallacea(Wallace 1860, Huxley 1868, Lydekker1896, Weber 1902, Mayr 1944, Hollowayand Jardine 1968, Simpson 1977). Efforts toidentify the line of faunal balance withinWallacea, west of which greater than 50%of the biota is derived from Asia and east ofwhich greater than 50% of the biota is Aus-tralian (Wallace 1876, Scrivenor et al. 1943,Mayr 1944, Lincoln 1975), led to detailedanalysis of the biogeography of the region,enabling an appreciation of the roles of geol-ogy, ecology, and evolution in shaping thesebiotas.
Within the Pacific Basin itself, the floraand fauna of the islands of the western Pacificare distinct from the biota of the islands oc-curring farther east and those in the northernor southern Pacific (Hedley 1899). Stoddart(1992:291) suggested that the most remark-able biotic discontinuity within the PacificBasin occurs at the Tonga Trench (Hedley’sline) and argued that this discontinuity wasbiologically significant because islands oneither side of Hedley’s line differed with re-spect to ‘‘altitude, climate, and other envi-ronmental constraints,’’ rather than simplygeographic coordinates.
Pacific Science (2010), vol. 64, no. 2:149–159doi: 10.2984/64.2.149: 2010 by University of Hawai‘i PressAll rights reserved
1 Funding for this project was provided by the Na-tional Science Foundation (DEB 0408010, DEB0445213, and DBI 0400797), an EPSCoR Fellowshipand grants from Graduate Women in Science, the Amer-ican Society of Ichthyologists and Herpetologists, the So-ciety for the Study of Amphibians and Reptiles, LSUchapter of Sigma Xi, the University of North Dakota(Graduate School, Office of Research and Program De-velopment and the Department of Biology), the LSUMuseum of Natural Science, and LSU BioGrads. Re-search was conducted under LSU IACUC Protocol no.03-121. Manuscript accepted 9 July 2009.
2 Department of Biological Sciences and Museum ofNatural Sciences, 119 Foster Hall, Louisiana State Uni-versity, Baton Rouge, Louisiana 70803.
3 Current address: Department of Ecology and Evo-lutionary Biology, 621 Charles E. Young Drive South,University of California Los Angeles, Los Angeles, Cali-fornia 90095.
4 Section of Integrative Biology and Texas MemorialMuseum, University of Texas, Austin, Texas 78712.
5 Corresponding author (e-mail: [email protected]).
(AutoPDF V7 25/11/09 11:51) UHP (7�10") Janson J-2210 Pacific Science, 64:2 (idp) PMU:(KN/W )18/11/2009 pp. 149–160 2210_64-2_01 (p. 149)

In addition to the broad distributionalpatterns in the Pacific Basin, it is importantto understand biogeographic patterns on asmaller scale, because the evolutionary his-tory of the lineages within the Pacific canbe accurately evaluated only when distri-butional patterns—and disjunctions withinthem—are recognized. The Outer Melane-sian Arc (OMA), stretching from New Guineathrough the Solomon Islands, Vanuatu Archi-pelago, Fijian Islands, the islands of Samoa,and southeast to Tonga (Kroenke and Rodda1984), is a major geological feature of thePacific Basin, west of the Tonga Trench.Components of the OMA are the result oftectonic activity over the last 11.2 to 2 myr(Kroenke and Rodda 1984), and the biota ofthis region results from speciation within theOMA (Filardi and Moyle 2005, Hamilton2008) as well as dispersal from SoutheastAsia, New Guinea, Australia, and New Cale-donia (Mayr 1965, Diamond and Mayr 1976,Bauer 1988, Brown 1991, Zug 1991, Flannery1995, 2000, Allison 1996, Austin 2000, Mayrand Diamond 2001, Carvajal and Adler 2005,Helgen 2007, Hamilton et al. 2009).
Historically, sharp biogeographic demar-cations that we recognize and name as lineshave been shown to be the result of old geo-logic processes shaped by ancient faunal andfloral diversification events. Wallace’s line,for example, identifies the suture betweenthe ancient Austral Papuan fauna of Gond-wanan origin, with the ancient northernAsian fauna derived from Laurasia. Similarly,Huxley’s line is concordant with the edge ofthe continental Sunda and Sahul shelves andthe deep Makassa and Lombok trenches(Hahn and Sytsma 1999), and Lyddeker’sline occurs along the edge of the continentalshelf to the west of West Irian and Australia(Simpson 1977). In contrast, our work in thegeologically young Vanuatu Archipelago sug-gests that biogeographic breaks can be foundin geologically young (likely less than 2 myrold) faunas.
The Vanuatu Archipelago comprises 82 is-lands. The oldest Vanuatu rocks are dated to14 mya; however, the entire archipelago wentthrough periods of emergence and submer-gence, with the last period of emergence
only in the last 2 myr (Carney et al. 1985,Greene et al. 1988a, Macfarlane et al. 1988).In addition to being biologically and geologi-cally nascent, the islands of Vanuatu are rela-tively isolated from other components of theOMA, surrounded by the Vanuatu Trench,the Vitiaz Trench, and the Johnson Trough(Coleman 1970). The current position ofVanuatu is a result of counterclockwise rota-tional movement since the Miocene (Gibbons1985, Greene et al. 1988a); before that rota-tion the islands of Vanuatu were located tothe north of Fiji and Tonga, adjacent to theVitiaz Arc (Chase and Seekins 1988) andmore isolated than now.
The geologically recent emergence of theVanuatu Archipelago (2 mya) explains someof the reptile and amphibian faunal discon-tinuities previously identified in Vanuatusuch as the absence of frogs (Platymantis)and elapid (Ogmodon) snakes (Schmidt 1930,Zweifel and Tyler 1982). In addition, theabsence of Ogmodon and Platymantis fromVanuatu—as well as the presence of Perochi-rus lizards in Vanuatu—has been suggestedto result from the north to south counter-clockwise rotational geologic movement ofthis archipelago (Gibbons 1985). The historicposition of Vanuatu was to the north of thecurrent location (Chase 1971); a location thatwould explain how groups such as Ogmodonand Platymantis could have colonized Fijifrom the Solomon Islands yet not be presentin Vanuatu. The distribution of Perochirus isrestricted to Micronesia, with a single species(P. guentheri) occurring in Vanuatu (Bauer1988). An older Vanuatu located north ofthe current position of the archipelago wouldhave permitted Perochirus to colonize Va-nuatu from Micronesia (Gibbons 1985). Theabsence of groups of vertebrates such as frogsand elapid snakes led to the suggestion thatthe fauna of Vanuatu may be depauperate(Baker 1928, 1929, Darlington 1948, Bauer1988). Recent examination of the terrestrialreptile diversity in Vanuatu in light of islandarea, isolation, and emergence history indi-cated that, in general, Vanuatu meets theexpectations of diversity generated by predic-tions of island biogeography theory (Hamil-ton et al. 2009).
(AutoPDF V7 25/11/09 11:51) UHP (7�10") Janson J-2210 Pacific Science, 64:2 (idp) PMU:(KN/W )18/11/2009 pp. 149–160 2210_64-2_01 (p. 150)
150 PACIFIC SCIENCE . April 2010

The origin and distribution of the biota ofVanuatu have not received much attentionbut when discussed have generally been con-sidered to result primarily from dispersal outof the Solomon Islands (Medway and Mar-shall 1975, Gibbons 1985, Bauer 1988, Otaet al. 1998). In addition, a distinct faunal andfloral break between Vanuatu and neighbor-ing New Caledonia has been noted (Bauer1988, Bauer and Vindum 1990, Bauer and Sa-dlier 1993, 2000, Hamilton et al. 2009). Here,we present data from our ongoing researchon the reptile fauna of the Vanuatu Archipel-ago on biogeographic patterns for squamatereptile species within this island group andnote a distinct biogeographic barrier withinthe Vanuatu Archipelago. We focus on thescincid lizard species Caledoniscincus atropunc-tatus (Roux, 1913) as an example, report newdistributional records for C. atropunctatus, anduse this species to illustrate and discuss gen-eral biogeographic patterns within the 82islands of Vanuatu. The biogeographic pat-terns highlighted by C. atropunctatus, particu-larly a biotic break between the islands ofsouthern Vanuatu (Erromango, Tanna, Fu-tuna, Aniwa, and Aneityum) and the islandsof central and northern Vanuatu, are congru-ent with patterns recovered from a review ofthe literature for a broad range of diverse tax-onomic groups including vertebrates, inverte-brates, and plants.
materials and methods
Between 2001 and 2005 we conducted fourcomprehensive field expeditions that surveyed53 sites on 19 islands throughout the VanuatuArchipelago and encompassed all major is-lands (Figure 1). At each site the lizard faunawas surveyed using a variety of techniques(Hamilton et al. 2007, 2008, Hamilton2008). Voucher specimens were collectedand deposited in the herpetology collectionat the Louisiana State University Museum ofNatural Science (lsumz).
We obtained data for seven bioclimaticvariables from WorldClim (Hijmans et al.2005), gridded to a 1 km resolution: annualmean temperature, mean diurnal range, max-imum temperature of the warmest month,
minimum temperature of the coldest month,annual precipitation, precipitation of the wet-test month, and precipitation of the driestmonth. These variables were chosen to cap-ture major variation among islands in annualmeans and seasonality of precipitation andtemperature because these environmentalconditions are likely to influence the distri-bution of a small, ectothermic reptile. Todetermine whether environments were differ-entiated across the species range limit of C.atropunctatus, we performed a principal com-ponents analysis (PCA) on environmentalvariables collected for the approximate centerof each island.
results and discussion
Relationship between Vanuatu and NewCaledonian Squamate Faunas
Despite the geographic proximity of the NewCaledonian Loyalty Islands and the southern-most Vanuatu island of Aneityum (only 150km between these island groups), the rep-tilian faunas of New Caledonia and theVanuatu Archipelago are dissimilar (Bauer1988, Hamilton et al. 2009). New Caledoniahas a highly endemic reptile fauna, with 86%of the terrestrial reptile species of New Cale-donia restricted to those islands (Bauer andSadlier 2000). Only nine of the 71 New Cale-donian squamate species are shared with Va-nuatu (Hamilton et al. 2009), and three ofthose nine species (Ramphotyphlops braminus,Lepidodactylus lugubris, and Hemidactylus frena-tus) are the result of human-mediated intro-ductions into both island groups (Bauer andSadlier 2000, Hamilton et al. 2009). Previousauthors have provided evidence for a distinctfaunal break between New Caledonia and theVanuatu Archipelago (Baker 1928, 1929,Solem 1976, Bauer 1988), and noted that thetwo regions are separated by the deep (6,400m) Vanuatu Trench (Greene et al. 1988b). Incontrast to the dissimilarity with Vanuatufauna, the New Caledonian squamate faunahas affinities with Australia and New Zealand(Bauer and Sadlier 1993, 2000), reflecting theGondwanan origin of New Caledonia. In ad-dition, the long history of island emergence
(AutoPDF V7 25/11/09 11:51) UHP (7�10") Janson J-2210 Pacific Science, 64:2 (idp) PMU:(KN/W )18/11/2009 pp. 149–160 2210_64-2_01 (p. 151)
Vanuatu Biogeography . Hamilton et al. 151

Figure 1. Distribution of Caledoniscincus atropunctatus in the Vanuatu Archipelago based on fieldwork conducted at 53sites on 19 islands over a 5 yr period is indicated by circles: survey sites at which C. atropunctatus was not observed areindicated with open circles and sites at which C. atropunctatus was collected are indicated by closed circles. The locationof Cheesman’s line, a biotic break within the Vanuatu Archipelago congruent with data from several taxonomic groupsincluding invertebrates, vertebrates, and plants is shown along with the summarized data and sources supporting thisbarrier. Distribution of C. atropunctatus in Vanuatu is congruent with this suggested biotic break because the species isrestricted to the islands south of Cheesman’s line.
(AutoPDF V7 25/11/09 11:51) UHP (7�10") Janson J-2210 Pacific Science, 64:2 (idp) PMU:(KN/W )18/11/2009 pp. 149–160 2210_64-2_01 (p. 152)

and isolation is likely responsible for the highrate of endemism in New Caledonia.
The oceanic islands of the Vanuatu Archi-pelago are considerably younger than NewCaledonia (Greene et al. 1988b, Macfarlaneet al. 1988, Bauer and Sadlier 2000), and, ingeneral, the squamate fauna is derived fromoverwater dispersal from New Guinea viathe Solomon Islands (Bauer 1988, Allison1996). Due to its Papuan origin, the Vanuatusquamate fauna is generally more similar tothose of other components of the Outer Mel-anesian Arc, such as the Fiji Archipelago andthe Solomon Islands (Hamilton et al. 2009).Of the six reptile species that are componentsof the native reptile fauna and have distribu-tions that encompass both New Caledoniaand the Vanuatu Archipelago, five are Papuanin origin, with Caledoniscincus atropunctatusrepresenting the sole species originating inNew Caledonia. This disparity may be dueto the ocean current between Vanuatu andthe New Caledonian Loyalty Islands, becausethe southerly direction of this flow system hasbeen suggested to promote colonization ofNew Caledonia from Vanuatu or Fiji (Gib-bons 1985, Treml et al. 2008).
Results of Herpetofaunal Surveys in Vanuatu
Fieldwork documented that Caledoniscincusatropunctatus was common and widely distrib-uted on Erromango Island, Aneityum Island,and Tanna Island (Figure 1). Collectionsmade during our fieldwork expanded theknown range of the species in Vanuatu to in-clude the southern islands of Aniwa and Fu-tuna (Figure 1). On all islands from which itwas collected, C. atropunctatus was numeri-cally common in both natural and semi-disturbed habitats. This species was foundin the leaf litter and was observed baskingin patches of sunlight on the forest floor.Although this species was collected frompatches of the forest edge adjacent to gar-dens, villages, and overgrown coconut planta-tions, it was more abundant in larger patchesof coastal forest or interior mixed hardwoodforests.
Burt and Burt (1932) also recorded C. atro-punctatus from the central island of Efate.
A.M.H. examined that specimen (amnh42002) and determined that it is not C. atro-punctatus but a species of Emoia based on thepresence of supranasals and fused frontopar-ietals (Brown 1991). We surveyed eight dif-ferent localities on and around Efate Island(Figure 1), and C. atropunctatus was never ob-served or collected despite extensive searchesof habitats typically occupied by this specieselsewhere in Vanuatu. Based on our extensivefield surveys and the erroneous single speci-men record from Efate we conclude that C.atropunctatus does not occur on Efate Islandor any of the northern or central islands.
Biogeographic Patterns within the VanuatuArchipelago
Caledoniscincus atropunctatus is distributedthroughout the southern islands of the Va-nuatu Archipelago but does not occur in thecentral and northern islands of this group.This distribution within Vanuatu mirrorsthe distribution of other lizard species. Thegecko Nactus pelagicus occurs on the samefive southern islands, with its congener, Nac-tus multicarinatus, being found on the north-ern and central islands. The distributions ofsome scincid lizards in the genus Emoia alsoprovide support for the presence of a faunalbreak between northern and central islands(the islands of the Torres Group south toEfate Island) and southern islands (Erro-mango Island south to Aneityum Island)(Figure 1). Emoia sanfordi is distributedthroughout the northern and central islandsof the Vanuatu Archipelago but does notoccur south of Efate Island (Hamilton 2008,Hamilton et al. 2008). Other species of Emoia(including E. aneityumensis and E. erronan)that are similar in body size, habitat use, andecology to E. sanfordi are endemic to southernVanuatu (Hamilton 2008) and further cor-roborate the presence of a distinct lizardfauna in the southern islands of the archipel-ago. In addition, other lizard species such asGekko vittatus (Gekkonidae) and Emoia nigra(Scincidae) have a distribution that encom-passes the Solomon Islands and the islandsof northern Vanuatu but no farther south(Medway and Marshall 1975, Cranbrook
(AutoPDF V7 25/11/09 11:51) UHP (7�10") Janson J-2210 Pacific Science, 64:2 (idp) PMU:(KN/W )18/11/2009 pp. 149–160 2210_64-2_01 (p. 153)
Vanuatu Biogeography . Hamilton et al. 153

1981, 1985a,b, Whitaker and Whitaker 1994,Hamilton Jennings and Austin 2002).
This biogeographic pattern is not re-stricted to squamate reptiles. Distinctlysimilar distributional breaks have been docu-mented in plants (Braithwaite 1975, Chew1975, Gillison 1975), bats (Medway and Mar-shall 1975, Flannery 1995), birds (Doughtyet al. 1999), freshwater gastropods (Haynes2000), ground beetles (Liebherr 2005), but-terflies (Gross 1975, Ackery et al. 1989), anda number of other invertebrates (Gross1975). Two general distributional patternscan be seen across a broad range of unrelatedtaxa: (1) a genus is distributed throughoutthe Vanuatu Archipelago, with the biota ofnorthern and central islands represented bya different assemblage of species than thesouthern Vanuatu islands; (2) each biotic re-gion in Vanuatu (northern and central versussouthern) is represented by a suite of genera,none of which bridges the biotic break be-tween Efate and Erromango islands.
The concordance between the Vanuatudistribution of C. atropunctatus and other bio-ta that dispersed from New Caledonia sug-gests that the occurrence of this species inVanuatu results from a natural dispersal eventfrom New Caledonia. The widespread distri-bution throughout the range and the highabundance and frequent occurrence of thisspecies in native, forested habitats providefurther support for the inclusion of C. atro-punctatus as a component of the native reptilefauna of Vanuatu.
The distributional disjunction between theislands of southern Vanuatu and the northernand central islands of Vanuatu was previouslynoted by Cheesman (1957), who suggestedthat this break represented a zoogeographicline analogous to Wallace’s line but not assharply defined. Cheesman (1957), in a pre–plate tectonics era, suggested a vicariancemodel for the pattern she observed. She ar-gued that the floral and faunal similarity ofthe southern islands of Erromango, Tanna,and Aneityum is due to a land-bridge connec-tion between these islands and presumed thatErromango, Tanna, and Aneityum were pre-viously a single landmass with a connectionto New Caledonia. Subsequent hypothesized
connection between this southern landmassand the northern islands of Vanuatu duringthe Miocene-Pliocene allowed the invasionfrom the north of the Papuan (Melanesian)biota that is present in southern Vanuatu(Cheesman 1957). The contemporary geo-logical literature, however, has provided abetter understanding of the tectonic and geo-logic history of the Vanuatu Archipelago andindicates that the islands of Vanuatu areyoung and of oceanic origin (Lee 1975). Fur-ther, there is no evidence for a previous con-nection between the islands of Erromango,Tanna, and Aneityum, nor for a connectionbetween these islands and the Gondwananlandmass of New Caledonia (Lee 1975,Greene et al. 1988b). The distributional pat-tern of the flora and fauna of Vanuatu there-fore has been the result of dispersal ratherthan vicariance.
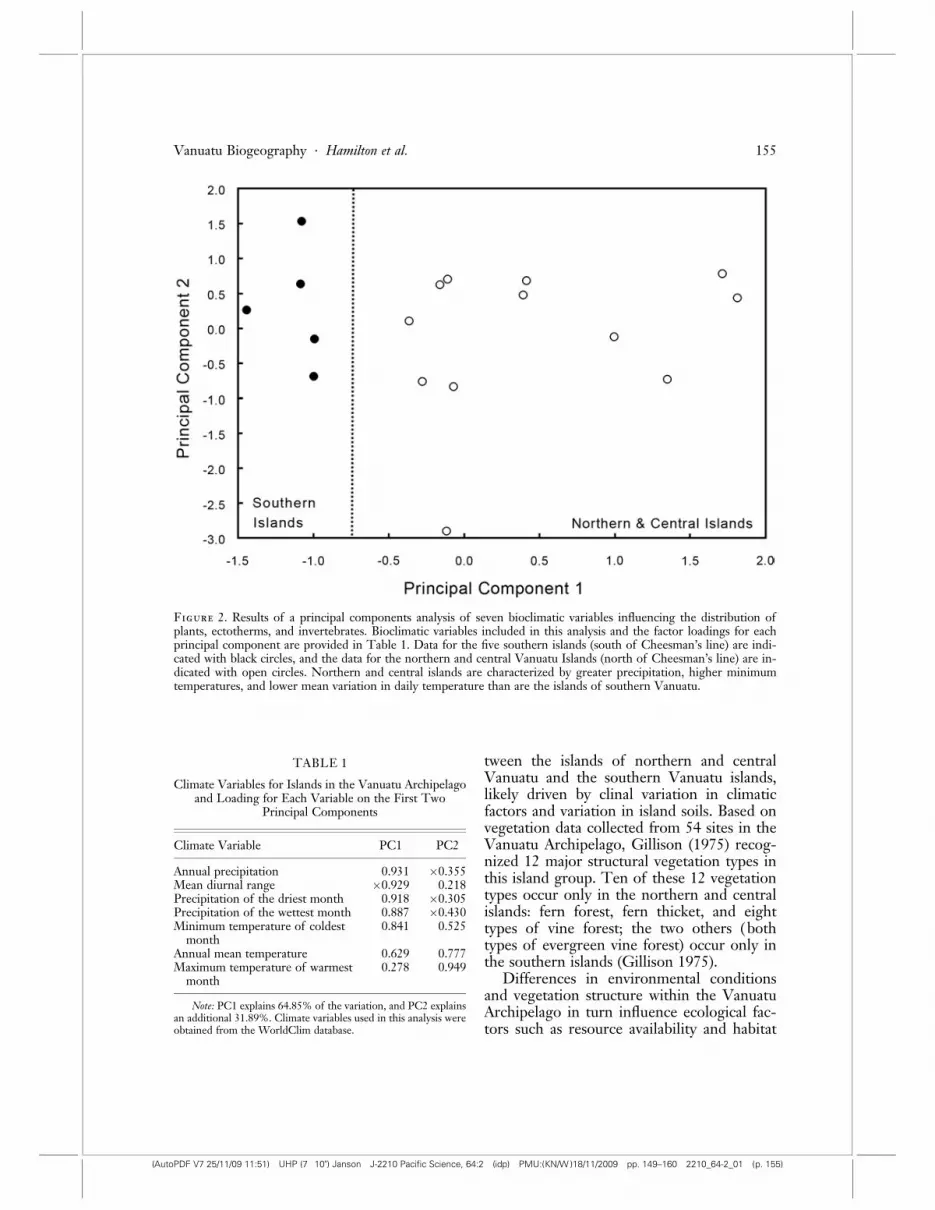
The biotic disjunction between the islandsof northern and central Vanuatu and those ofsouthern Vanuatu may be influenced by cli-mate as well as soil and vegetation type (Fig-ure 2). A PCA of seven climate variableslikely to influence the distribution of organ-isms (such as ectotherms and plants) sensi-tive to environmental conditions shows cleardifferences between the two biotic regions(Table 1, Figure 2). The climate data suggestthat the environmental conditions of the is-lands of northern and central Vanuatu arewetter and less thermally variable than onsouthern Vanuatu islands, as indicated by theincreases in mean annual precipitation andminimum temperature of the coldest monthin the northern islands, and the greater diur-nal temperature variation in the southern is-lands (Figure 2). Although the environmentsof the islands clearly vary with latitude, thePCA of the WorldClim bioclimatic data illus-trates the environmental differences amongislands within the Vanuatu Archipelago.
In addition to the latitudinal cline in envi-ronmental conditions, soil type varies amongthese islands. The islands of Erromango andTanna have different soils, and fewer soiltypes, than the central islands of Efate, Epi,Espiritu Santo, Maewo, Malakula, Pentecost,and the Shepherd Islands (Quantin 1975).Habitats and habitat diversity also vary be-
(AutoPDF V7 25/11/09 11:51) UHP (7�10") Janson J-2210 Pacific Science, 64:2 (idp) PMU:(KN/W )18/11/2009 pp. 149–160 2210_64-2_01 (p. 154)
154 PACIFIC SCIENCE . April 2010
tween the islands of northern and centralVanuatu and the southern Vanuatu islands,likely driven by clinal variation in climaticfactors and variation in island soils. Based onvegetation data collected from 54 sites in theVanuatu Archipelago, Gillison (1975) recog-nized 12 major structural vegetation types inthis island group. Ten of these 12 vegetationtypes occur only in the northern and centralislands: fern forest, fern thicket, and eighttypes of vine forest; the two others (bothtypes of evergreen vine forest) occur only inthe southern islands (Gillison 1975).
Differences in environmental conditionsand vegetation structure within the VanuatuArchipelago in turn influence ecological fac-tors such as resource availability and habitat
Figure 2. Results of a principal components analysis of seven bioclimatic variables influencing the distribution ofplants, ectotherms, and invertebrates. Bioclimatic variables included in this analysis and the factor loadings for eachprincipal component are provided in Table 1. Data for the five southern islands (south of Cheesman’s line) are indi-cated with black circles, and the data for the northern and central Vanuatu Islands (north of Cheesman’s line) are in-dicated with open circles. Northern and central islands are characterized by greater precipitation, higher minimumtemperatures, and lower mean variation in daily temperature than are the islands of southern Vanuatu.
TABLE 1
Climate Variables for Islands in the Vanuatu Archipelagoand Loading for Each Variable on the First Two
Principal Components
Climate Variable PC1 PC2
Annual precipitation 0.931 �0.355Mean diurnal range �0.929 0.218Precipitation of the driest month 0.918 �0.305Precipitation of the wettest month 0.887 �0.430Minimum temperature of coldest
month0.841 0.525
Annual mean temperature 0.629 0.777Maximum temperature of warmest
month0.278 0.949
Note: PC1 explains 64.85% of the variation, and PC2 explainsan additional 31.89%. Climate variables used in this analysis wereobtained from the WorldClim database.
(AutoPDF V7 25/11/09 11:51) UHP (7�10") Janson J-2210 Pacific Science, 64:2 (idp) PMU:(KN/W )18/11/2009 pp. 149–160 2210_64-2_01 (p. 155)
Vanuatu Biogeography . Hamilton et al. 155

suitability (such as the thermal preferences ofectothermic species) and contribute to theobserved biotic disjunction. We concludethat dispersal of the flora and fauna from thedisparate Papuan and New Caledonian re-gions combined with intra-archipelago differ-ences in climate, soil, and vegetation haveproduced one of the most distinct biogeo-graphic disjunctions in the Pacific. We sug-gest that this discontinuity between thenorthern (and central islands) of Vanuatuand the southern islands of the archipelagobe referred to as Cheesman’s line in honorof the contributions of Evelyn Cheesman tothe study of Pacific herpetology and biogeog-raphy.
acknowledgments
We thank Mr. Ernest Bani and Ms. DonnaKalfatak of the Environment Unit of the Re-public of Vanuatu for permits to conduct re-search on the reptiles of Vanuatu and forexport permits for tissues and voucher speci-mens collected in Vanuatu; numerous ni-Vanuatu chiefs and villagers for assistancethroughout Vanuatu; A. Freedman for assis-tance with analysis of the WorldClim data;and E. Hartfield, K. Blaha, M. Eckstut, andK. Grazyck for assistance with fieldwork inVanuatu. D. Kizirian provided access to theVanuatu Caledoniscincus specimens in theamnh. The manuscript was improved bycomments from A. Freedman, M. Eckstut,and two anonymous reviewers.
Literature Cited
Ackery, P. R., H. Taylor, and A. Renevier.1989. The milkweed butterfly Paranticapumila (Boisduval): Its biology and distri-bution in relation to the biogeography ofVanuatu. J. Nat. Hist. 23:713–723.
Allison, A. 1996. Zoogeography of amphib-ians and reptiles of New Guinea and thePacific region. SPB Academic Publishing,Amsterdam.
Austin, C. C. 2000. Molecular phylogeny andhistorical biogeography of Pacific islandboas (Candoia). Copeia 2000:341–352.
Baker, J. R. 1928. Non-marine vertebrate
fauna of the New Hebrides. Ann. Mag.Nat. Hist. 10:294–302.
———. 1929. Man and animals in the NewHebrides. George Routledge and Sons,London.
Bauer, A. M. 1988. Hypothesis: A geologicalbasis for some herpetofaunal disjunctionsin the Southwest Pacific, with special ref-erence to Vanuatu. Herpetol. J. 1:259–263.
Bauer, A. M., and R. A. Sadlier. 1993. Sys-tematics, biogeography and conservationof the lizards of New Caledonia. Bio-divers. Lett. 1:107–122.
———. 2000. The herpetofauna of New Ca-ledonia. Society for the Study of Amphib-ians and Reptiles, Ithaca, New York.
Bauer, A. M., and J. V. Vindum. 1990. Achecklist and key to the herpetofauna ofNew Caledonia, with remarks on biogeog-raphy. Proc. Calif. Acad. Sci. 47:17–45.
Braithwaite, A. F. 1975. The phytogeograph-ical relationships and origin of the NewHebrides fern flora. Philos. Trans. R. Soc.Lond. B Biol. Sci. 272:293–313.
Brown, W. C. 1991. Lizards of the genusEmoia (Scincidae) with observations ontheir evolution and biogeography. Mem.Calif. Acad. Sci. 15.
Burt, C. E., and M. D. Burt. 1932. Herpeto-logical results of the Whitney South SeaExpedition. VI. Bull. Am. Mus. Nat. Hist.63:461–597.
Carney, J. N., A. Macfarlane, and D. I. J.Mallick. 1985. The Vanuatu Island Arc:An outline of stratigraphy, structure andpetrology. Pages 683–718 in A. E. M.Nairn, F. G. Stehli, and S. Uyeda, eds.The ocean basins and margins. Vol. 7A:The Pacific Ocean. Plenum Press, NewYork.
Carvajal, A., and G. H. Adler. 2005. Biogeog-raphy of mammals on tropical Pacific is-lands. J. Biogeogr. 32:1561–1569.
Chase, C. G. 1971. Tectonic history of theFiji Plateau. Geol. Soc. Am. Bull. 82:3087–3110.
Chase, T. E., and B. A. Seekins. 1988. Sub-marine topography of the Vanuatu andsoutheastern Solomon Islands region.Pages 35–36 in H. G. Greene and F. L.
(AutoPDF V7 25/11/09 11:51) UHP (7�10") Janson J-2210 Pacific Science, 64:2 (idp) PMU:(KN/W )18/11/2009 pp. 149–160 2210_64-2_01 (p. 156)
156 PACIFIC SCIENCE . April 2010

Wong, eds. Geology and offshore re-sources of Pacific island arcs: Vanuatu re-gion. Circum-Pacific Council for Energyand Mineral Resources, Houston, Texas.
Cheesman, L. E. 1957. Biogeographical sig-nificance of Aneityum Island, New Hebri-des. Nature (Lond.) 180:903–904.
Chew, W.-L. 1975. The phanerogamic floraof the New Hebrides and its relationships.Philos. Trans. R. Soc. Lond. B Biol. Sci.272:315–328.
Coleman, P. J. 1970. Geology of the Solo-mon and New Hebrides islands, as part ofthe Melanesian re-entrant, Southwest Pa-cific. Pac. Sci. 24:289–314.
Cranbrook, E. O. 1981. A checklist and pre-liminary key to the lizards of Vanuatu.Naika 4:3–5.
———. 1985a. The herpetofauna of Va-nuatu. Br. Herpetol. Soc. Bull. 13.
———. 1985b. The lizards of Vanuatu.ASRA J. 2:47–71.
Darlington, P. J. 1948. The geographical dis-tribution of cold-blooded vertebrates. Q.Rev. Biol. 23:1–26, 105–123.
Diamond, J. M., and E. Mayr. 1976. Species-area relation for birds of the Solomon Ar-chipelago. Proc. Natl. Acad. Sci. U.S.A.73:262–266.
Doughty, C., N. Day, and A. Plant. 1999.Birds of the Solomons, Vanuatu, andNew Caledonia. A&C Black, London.
Filardi, C. E., and R. G. Moyle. 2005. Singleorigin of a pan-Pacific bird group and up-stream colonization of Australasia. Nature(Lond.) 438:216–219.
Flannery, T. 1995. Mammals of the South-West Pacific and Moluccan Islands. Cor-nell University Press, Ithaca, New York.
———. 2000. Mammalian zoogeography ofNew Guinea and the southwestern Pacific.Pages 399–406 in A. Keast and S. Miller,eds. The origin and evolution of Pacificisland biotas. New Guinea to eastern Poly-nesia: Patterns and processes. SPB Aca-demic Publishing, Amsterdam.
Gibbons, J. R. H. 1985. The biogeographyand evolution of Pacific island reptiles andamphibians. Pages 125–142 in G. Grigg,R. Shine, and H. Ehmann, eds. Biologyof Australasian frogs and reptiles. Surrey
Beatty & Sons, Chipping Norton, NewSouth Wales.
Gillison, A. N. 1975. Phytogeographical rela-tionships of the northern islands of theNew Hebrides. Philos. Trans. R. Soc.Lond. B Biol. Sci. 272:385–390.
Greene, H. G., A. Macfarlane, D. P. Johnson,and A. J. Crawford. 1988a. Structure andtectonics of the central New Hebridesarc. Pages 377–412 in H. G. Greene andF. L. Wong, eds. Geology and offshore re-sources of Pacific island arcs: Vanuatu re-gion. Circum-Pacific Council for Energyand Mineral Resources, Houston,Texas.
Greene, H. G., A. Macfarlane, and F. L.Wong. 1988b. Geology and offshore re-sources of Vanuatu: Introduction andsummary. Pages 1–25 in H. G. Greeneand F. L. Wong, eds. Geology and off-shore resources of Pacific island arcs: Va-nuatu region. Circum-Pacific Council forEnergy and Mineral Resources, Houston,Texas.
Gross, G. F. 1975. The land invertebrates ofthe New Hebrides and their relationships.Philos. Trans. R. Soc. Lond. B Biol. Sci.272:391–421.
Hahn, W. J., and K. J. Sytsma. 1999. Molec-ular systematics and biogeography of theSoutheast Asian genus Caryota (Palmae).Syst. Bot. 24:558–580.
Hamilton, A. M. 2008. Species boundaries,biogeography, and intra-archipelago ge-netic variation within the Emoia samoensisspecies group in the Vanuatu Archipelagoand Oceania. Ph.D. diss., Louisiana StateUniversity, Baton Rouge.
Hamilton, A. M., M. E. Eckstut, E. R. Klein,and C. C. Austin. 2008. Clutch size in thetropical scincid lizard Emoia sanfordi, a spe-cies endemic to the Vanuatu Archipelago.Zool. Sci. 25:843–848.
Hamilton, A. M., J. H. Hartman, and C. C.Austin. 2009. Island area and species diver-sity in the Southwest Pacific Ocean: Is thefauna of Vanuatu depauperate? Ecography32:247–258.
Hamilton, A. M., E. R. Klein, M. E. Eckstut,and E. E. Hartfield. 2007. A simple, inex-pensive method to capture arboreal lizards.Herpetol. Conserv. Biol. 2:164–167.
(AutoPDF V7 25/11/09 11:51) UHP (7�10") Janson J-2210 Pacific Science, 64:2 (idp) PMU:(KN/W )18/11/2009 pp. 149–160 2210_64-2_01 (p. 157)
Vanuatu Biogeography . Hamilton et al. 157

Hamilton Jennings, A. M., and C. C. Austin.2002. Report for research conducted inVanuatu during 2001 and 2002. Report tothe Vanuatu Environment Unit, Ministryof Natural Resources, Port Vila. Availablefrom the author.
Haynes, A. 2000. The distribution of fresh-water gastropods on four Vanuatu islands:Espiritu Santo, Pentecost, Efate andTanna (South Pacific). Ann. Limnol.-Int.J. Limnol. 36:101–111.
Hedley, C. 1899. A zoogeographic schemefor the mid-Pacific. Proc. Linn. Soc. N. S.W. 24:391–423.
Helgen, K. M. 2007. A reassessment of taxo-nomic diversity and geographic patterningin the Melanesian mammal fauna. Ph.D.thesis, University of Adelaide, Adelaide.
Hijmans, R. J., S. E. Cameron, J. L. Parra,P. G. Jones, and A. Jarvis. 2005. Veryhigh resolution interpolated climate sur-faces for global land areas. Int. J. Climatol.25:1965–1978.
Holloway, J. D., and N. Jardine. 1968. Twoapproaches to zoogeography: A studybased on the distribution of butterflies,birds, and bats in the Indo-Australianarea. Proc. Linn. Soc. Lond. 179:153–188.
Huxley, T. H. 1868. On the classification anddistribution of the Alectoromorphae andHeteromorphae. Proc. Zool. Soc. Lond.1868:294–319.
Kroenke, L. W., and P. Rodda. 1984. Ceno-zoic tectonic development of the South-west Pacific. United Nations Economicand Social Commission, Committee forCo-ordination of Joint Prospecting forMineral Resources in South Pacific Off-shore Areas (CCOP/SOPAC), New Zea-land.
Lee, K. E. 1975. Conclusions. Philos. Trans.R. Soc. Lond. B Biol. Sci. 272:477–486.
Liebherr, J. K. 2005. Platynini (Coleoptera:Carabidae) of Vanuatu: Miocene diversifi-cation on the Melanesian Arc. Invertebr.Syst. 19:263–295.
Lincoln, G. A. 1975. Bird counts on eitherside of Wallace’s line. Proc. Zool. Soc.Lond. 177:349–361.
Lydekker, R. 1896. A geographical history ofmammals. University Press, Cambridge.
Macfarlane, A., J. N. Carney, A. J. Crawford,and H. G. Greene. 1988. Vanuatu: A re-view of the onshore geology. Pages 45–91in H. G. Greene and F. L. Wong, eds. Ge-ology and offshore resources of Pacific is-land arcs: Vanuatu region. Circum-PacificCouncil for Energy and Mineral Re-sources, Houston, Texas.
Mayr, E. 1944. Wallace’s line in the lightof recent zoogeographic studies. Q. Rev.Biol. 19:1–14.
———. 1965. Avifauna: Turnover on islands.Science (Washington, D.C.) 150:1287–1588.
Mayr, E., and J. M. Diamond. 2001. Thebirds of northern Melanesia: Speciation,ecology, and biogeography. Oxford Uni-versity Press, New York.
Medway, L., and A. G. Marshall. 1975. Ter-restrial vertebrates of the New Hebrides:Origin and distribution. Philos. Trans. R.Soc. Lond. B Biol. Sci. 272:423–465.
Ota, H., R. N. Fisher, I. Ineich, T. J. Case,R. R. Radtkey, and G. R. Zug. 1998. Anew Lepidodactylus (Squamata: Gekkoni-dae) from Vanuatu. Herpetologica 54:325–332.
Quantin, P. 1975. Soils of the New Hebridesislands. Philos. Trans. R. Soc. Lond. BBiol. Sci. 272:287–292.
Roux, J. 1913. Les reptiles de la NouvelleCaledonie et des iles Loyalty. Appendice:Note sur quelques reptiles des Nouvelle-Hebrides, des ıles Banks et Santa Cruz.Pages 79–160 in F. Sarasin and J. Roux,eds. Nova Caledonia, Recherches scientifi-ques en Nouvelle Caledonie et aux IlesLoyalty. Zoologie. C. W. Kreidel’s Verlag,Wiesbaden.
Schmidt, K. P. 1930. Essay on the zoogeogra-phy of the Pacific islands. Pages 275–292in G. N. Shurcliff, ed. Jungle isles. Put-man, New York.
Sclater, P. L. 1858. On the geographical dis-tribution of the members of the class Aves.J. Linn. Soc. Lond. Zool. 2:130–145.
Scrivenor, J. B., T. H. Burkill, M. A. Smith,A. St. Corbet, H. K. A. Shaw, P. W. Ri-chards, and F. E. Zeuner. 1943. A discus-sion on the biogeographic division of theIndo-Australian Archipelago, with criti-
(AutoPDF V7 25/11/09 11:51) UHP (7�10") Janson J-2210 Pacific Science, 64:2 (idp) PMU:(KN/W )18/11/2009 pp. 149–160 2210_64-2_01 (p. 158)
158 PACIFIC SCIENCE . April 2010

cism of the Wallace and Weber lines andof any other dividing lines and with an at-tempt to obtain uniformity in the namesused for the divisions. Proc. Linn. Soc.Lond. 154:120–165.
Simpson, G. G. 1977. Too many lines: Thelimits of the Oriental and Australian zoo-geographic regions. Proc. Am. Philos.Soc. 121:107–120.
Solem, A. 1976. Endodontoid land snailsfrom Pacific islands (Mollusca: Pulmonata:Sigmurethra), Part I: Family Endodonti-dae. Field Musuem of Natural History,Chicago.
Stoddart, D. R. 1992. Biogeography of thetropical Pacific. Pac. Sci. 46:276–293.
Treml, E. A., P. N. Halpin, D. L. Urban, andL. F. Pratson. 2008. Modeling populationconnectivity by ocean currents, a graph-theoretic approach for marine conserva-tion. Landscape Ecol. 23:19–36.
Wallace, A. R. 1860. On the zoological geog-
raphy of the Malay Archipelago. J. Linn.Soc. Lond. 4:172–184.
———. 1876. The geographical distributionof animals. Macmillan, London.
Weber, M. 1902. Der Indo-Australische Ar-chipel und die Geschichte Seiner Tierwelt.Gustav Fischer, Jena.
Whitaker, A. H., and V. Whitaker. 1994. Asurvey of the herpetofauna near Matantas,Espiritu Santo; with notes on reptileselsewhere on Espiritu Santo and Efate,Vanuatu. Report to the Vanuatu Environ-ment Unit, Ministry of Natural Resources,Port Vila. Available from the Vanuatu En-vironment Unit.
Zug, G. R. 1991. Lizards of Fiji: Natural his-tory and systematics. Bishop MuseumPress, Honolulu.
Zweifel, R. G., and M. J. Tyler. 1982. Am-phibia of New Guinea. Pages 759–801 inJ. L. Gressitt, ed. Biogeography and ecol-ogy of New Guinea. W. Junk, The Hague.
(AutoPDF V7 25/11/09 11:51) UHP (7�10") Janson J-2210 Pacific Science, 64:2 (idp) PMU:(KN/W )18/11/2009 pp. 149–160 2210_64-2_01 (p. 159)
Vanuatu Biogeography . Hamilton et al. 159

(AutoPDF V7 25/11/09 11:51) UHP (7�10") Janson J-2210 Pacific Science, 64:2 (idp) PMU:(KN/W )18/11/2009 pp. 149–160 2210_64-2_01 (p. 160)





























