Embed Size (px)
Citation preview

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Chemistry and Physics of Lipids 163 (2010) 94–101
Contents lists available at ScienceDirect
Chemistry and Physics of Lipids
journa l homepage: www.e lsev ier .com/ locate /chemphys l ip
Binding of a cationic phenazinium dye in anionic liposomal membrane:a spectacular modification in the photophysics
Debosreeta Bose, Debanjana Ghosh, Priyanka Das, Agnishwar Girigoswami,Deboleena Sarkar ∗, Nitin Chattopadhyay ∗
Department of Chemistry, Jadavpur University, Raja S. C. Mullick Road, Kolkata 700032, India
a r t i c l e i n f o
Article history:Received 8 June 2009Received in revised form 4 September 2009Accepted 16 October 2009Available online 27 October 2009
Keywords:LiposomeDimyristoyl-l-�-phosphatidylglycerol(DMPG)Phenazinium dyeFluorescencePartition coefficient
a b s t r a c t
Interaction of a cationic phenazinium dye, phenosafranin (PSF), with the anionic liposomal vesicle/bilayerof dimyristoyl-l-�-phosphatidylglycerol (DMPG) has been demonstrated using steady state and timeresolved fluorescence and fluorescence anisotropy techniques. The charge transfer emission spectrumof PSF shows a dramatic modification in terms of fluorescence yield together with an appreciable hyp-sochromic shift in the lipid environment. The blue shift indicates a lowering in polarity inside the vesicleas compared to that in bulk water. The fluorescence and fluorescence quenching studies and microp-olarity determination reveal that the cationic fluorophore has a profound binding interaction with theanionic DMPG membrane. Anisotropy study indicates the imposition of a motional restriction on theprobe inside the bilayer. The electrostatic interaction between the cationic dye and the anionic lipidmembrane has been argued to be the reason behind all these observations. The results could be usefulin analyzing membrane organization and heterogeneity in natural membranes exploiting PSF or alikecompounds as fluorescent probes.
© 2009 Elsevier Ireland Ltd. All rights reserved.
1. Introduction
Organized molecular assemblies such as vesicles or liposomescan be considered as large cooperative units being very dif-ferent from the individual structural units constituting them(Chattopadhyay and Mukherjee, 1993). Lipid vesicles are closedsystems consisting of one or more lamellae containing theamphiphiles. In vesicles, the hydrophobic part of the unit moleculesforms the interior of the bilayer and the hydrophilic portionremains in contact with the aqueous phase. Liposomes can beemployed as simple models for biological membranes (Gregoriadisand Allison, 1986). Phospholipid vesicles which have alternatinghydrophilic and hydrophobic parts provide opportunities for theformulation of controlled release and site specific delivery systems(Moyano et al., 2006; Teissie and Tsong, 1981; Correa and Schelly,1998a). Biphasic nature of the vesicles helps both lipophilic andhydrophilic ingredients to accommodate in it (Correa and Schelly,1998a,b; Asgharian and Schelly, 1999; Sujatha and Mishra, 1997,1998). The dye–liposome interaction is associated with many bio-logical processes in bio-membranes. However, the complicationscoming out of the wide diversity in composition and structures of
∗ Corresponding authors. Tel.: +91 94 3394 8648; fax: +91 33 2414 6266.E-mail addresses: [email protected] (D. Sarkar),
[email protected] (N. Chattopadhyay).
biological membranes can be avoided if we use synthetic liposomesor vesicles which mimic the geometry, topology and skeletal struc-ture of cell membranes (Moyano et al., 2006; Teissie and Tsong,1981; Correa and Schelly, 1998a,b). The ability of liposome to mimicthe behavior of natural membranes makes it safe and efficaciousvehicle for medicinal applications (Correa and Schelly, 1998a,b;Asgharian and Schelly, 1999; Sujatha and Mishra, 1997, 1998).Insights into many aspects of cell physiology, permeability, fusion,etc. can be explored exploiting these artificial membranes. Phos-pholipids are the fundamental matrix of natural membranes andrepresent the environment in which many proteins and enzymescan display their activities (Gregoriadis and Allison, 1986). Lipo-somes have been effectively utilized in diverse fields like drugloading, immunology, diagnostics, and food industry (Lasic, 1993).Lipid vesicles are also used as selective barriers, nano-capsulesand protective agents for the entrapped molecules (Needham andDewhirst, 2001; Chen and Regen, 2005).
Because of its sensitivity, selectivity and noninvasive nature,fluorescence spectroscopy has been widely used to characterizebiosystems. The micromolar concentration of a fluorescent proberequired for the investigation does not noticeably perturb the prop-erties of membranes. Penetration of the probe into the bilayermembrane from the bulk water modifies the photoprocesses sincethe polarity and viscosity in the immediate environments aroundthe probe are quite different from that of the bulk water (Das etal., 2008). It is thus important to study the liposomal microenvi-
0009-3084/$ – see front matter © 2009 Elsevier Ireland Ltd. All rights reserved.doi:10.1016/j.chemphyslip.2009.10.003

Author's personal copy
D. Bose et al. / Chemistry and Physics of Lipids 163 (2010) 94–101 95
Scheme 1. Structures of DMPG and PSF.
ronments and the modifications of the photophysics of the probein the liposome membranes. In the present study we have useda cationic phenazinium dye, namely, phenosafranin (PSF, 3,7-diamino-5-phenyl phenazinium chloride) as the fluorescent probe(Scheme 1). The dye has extensively been used in semiconductors,as energy sensitizer, and as a probe for studying various micro-heterogeneous environments including micelles, reverse micellesand polymeric matrices (Broglia et al., 2005; Jockush et al., 1997;Jayanthi and Ramamurthy, 1998; Gopidas and Kamat, 1990). It hasalso been established as a DNA intercalator (Sarkar et al., 2008b)and is found to form dimeric aggregates at higher concentrations(Sarkar et al., 2008a). It is red in color with a planar tricyclicphenazinium moiety bearing a positive charge. PSF is a water-soluble dye with characteristic broad absorption peak in the visibleregion (Broglia et al., 2005). Trapping of such biologically potentmolecules in different biomimetic environments attracts interestof the researchers because of their ability to achieve specific chem-ical efficiency as a result of organization in the reaction media. Here,we have studied the photophysics and binding interaction of PSFin anionic dimyristoyl-l-�-phosphatidylglycerol (DMPG) environ-ment (Scheme 1). The results demonstrate the effect of the lipidenvironment on the steady state fluorescence, fluorescence lifetimeand the steady state and time resolved anisotropy of the entrappedfluorophore.
2. Experimental procedures
2.1. Materials and instrumentation
Phenosafranin (PSF) and DMPG were purchased fromSigma–Aldrich (USA) and they were used as received. All thesolvents used were of UV spectroscopic grade (Spectrochem,India). AR grade potassium bromide and Tris–HCl buffer werepurchased from SRL (India). Triply distilled water was used formaking the experimental solutions. pH of the solutions weremaintained at 7.4 using Tris-HCl buffer. Concentration of PSF waskept at ca. 7.0 × 10−6 mol dm−3.
Absorption and steady state fluorescence measurements wereperformed using a Shimadzu UV-2450 spectrophotometer anda Horiba Jobin Yvon Fluoromax-4P spectrofluorimeter, respec-tively. The steady state fluorescence anisotropy was also measuredusing the same spectrofluorimeter. Steady state anisotropy, r, wasdefined by:
r = (IVV − G · IVH)(IVV + 2G · IVH)
(1)
where IVV and IVH are the intensities obtained with the excitationpolarizer oriented vertically and the emission polarizer orientedvertically and horizontally, respectively. The G factor was defined
as:
G = IHV
IHH(2)
I terms refer to parameters similar to those mentioned above for thehorizontal position of the excitation polarizer. Fluorescence life-times were determined from time resolved intensity decays by themethod of time correlated single photon counting (TCSPC) usinga picosecond diode laser at 403 nm (IBH, UK, nanoLED-07) as thelight source and TBX-04 as the detector. The decays were analyzedusing IBH DAS-6 decay analysis software. Goodness of fits was eval-uated by �2 criterion and the randomness of the residuals. Meanfluorescence lifetimes (�f) for bi-exponential iterative fitting werecalculated from the decay times (�i) and the relative amplitudes(ai) using the following relation
〈�f〉 = a1�1 + a2�2 (3)
For anisotropy decay measurements, the samples were excitedat 403 nm using a picosecond diode laser (IBH) and a Hamamatsumicrochannel plate photomultiplier tube (3809U). The instru-ment response function (FWHM) of the setup was 90 ps. For theanisotropy decays, we used a motorized polarizer in the emissionside. The emission intensities at parallel (IVV) and perpendicu-lar (IVH) polarizations were collected alternatively until a presetpeak difference was reached. The preset value, however, dependedon the tail matching of the parallel and perpendicular decays. Toimprove the signal-to-noise ratio, the data sets were averaged.The analysis of the data was done using IBH DAS, version 6, decayanalysis software. The quality of the fits was determined from thereduced �2 and the randomness of the residuals of the fitted func-tion to the data. The time resolved anisotropy [r(t)] was calculatedusing the following relation
r(t) = [IVV(t) − GIVH(t)][IVV(t) + 2GIVH(t)]
(4)
where G is the correction factor for the detector sensitivity to thepolarization detection of the emission.
All the experiments were performed at 25 ◦C temperature withair-equilibrated solutions.
2.2. Liposome preparation
Liposome of DMPG was prepared by the method of sonication(Huang, 1969). The phopholipid was first dissolved in 2:1 (v/v) chlo-roform:methanol solution and the solvent was evaporated understream of nitrogen. The resulted lipid film was then dried in vac-uum desiccator. The dry film was hydrated and swelled in Tris–HClbuffer at pH 7.4 containing 20 mM NaCl. The mixture was then vor-texed to disperse the lipids. The dispersion was then sonicated for30 min using Vibronics Ultrasonic P1 sonicator. The resulted sam-ple was centrifuged at 6000 rpm for 10 min to get rid of the foreignparticles, if any. All the spectroscopic experiments were performedwith freshly prepared vesicles.
3. Results and discussion
3.1. Steady state absorption and emission
PSF shows a broad unstructured absorption band with a max-imum at around 520 nm in aqueous buffer solution (Sarkar et al.,2008b). On addition of DMPG to the aqueous buffer solution of PSFthe band maximum shifts to ∼530 nm with a slight decrease inthe absorbance of PSF at higher lipid concentrations indicating thatthe environment around the probe gets modified in the lipid solu-tions from the pure aqueous phase. Discussions in the forthcomingsections reveal that a lowering in the polarity in the immediate
Author's personal copy
96 D. Bose et al. / Chemistry and Physics of Lipids 163 (2010) 94–101
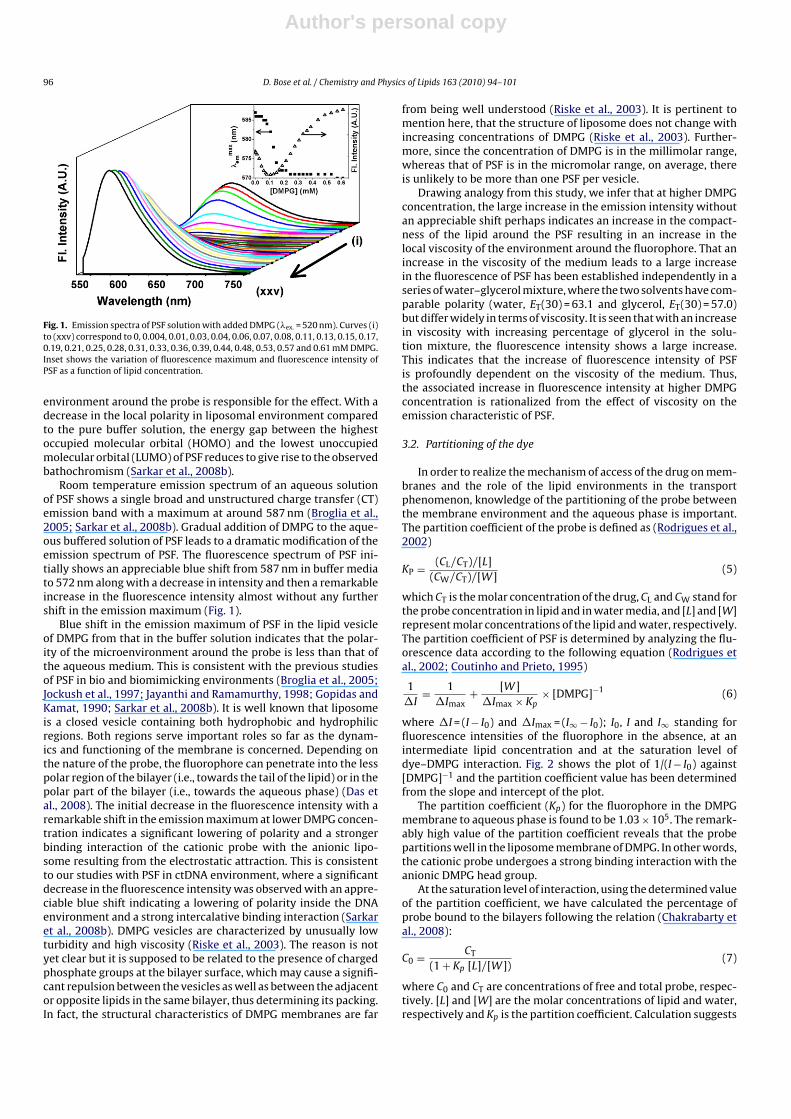
Fig. 1. Emission spectra of PSF solution with added DMPG (�ex. = 520 nm). Curves (i)to (xxv) correspond to 0, 0.004, 0.01, 0.03, 0.04, 0.06, 0.07, 0.08, 0.11, 0.13, 0.15, 0.17,0.19, 0.21, 0.25, 0.28, 0.31, 0.33, 0.36, 0.39, 0.44, 0.48, 0.53, 0.57 and 0.61 mM DMPG.Inset shows the variation of fluorescence maximum and fluorescence intensity ofPSF as a function of lipid concentration.
environment around the probe is responsible for the effect. With adecrease in the local polarity in liposomal environment comparedto the pure buffer solution, the energy gap between the highestoccupied molecular orbital (HOMO) and the lowest unoccupiedmolecular orbital (LUMO) of PSF reduces to give rise to the observedbathochromism (Sarkar et al., 2008b).
Room temperature emission spectrum of an aqueous solutionof PSF shows a single broad and unstructured charge transfer (CT)emission band with a maximum at around 587 nm (Broglia et al.,2005; Sarkar et al., 2008b). Gradual addition of DMPG to the aque-ous buffered solution of PSF leads to a dramatic modification of theemission spectrum of PSF. The fluorescence spectrum of PSF ini-tially shows an appreciable blue shift from 587 nm in buffer mediato 572 nm along with a decrease in intensity and then a remarkableincrease in the fluorescence intensity almost without any furthershift in the emission maximum (Fig. 1).
Blue shift in the emission maximum of PSF in the lipid vesicleof DMPG from that in the buffer solution indicates that the polar-ity of the microenvironment around the probe is less than that ofthe aqueous medium. This is consistent with the previous studiesof PSF in bio and biomimicking environments (Broglia et al., 2005;Jockush et al., 1997; Jayanthi and Ramamurthy, 1998; Gopidas andKamat, 1990; Sarkar et al., 2008b). It is well known that liposomeis a closed vesicle containing both hydrophobic and hydrophilicregions. Both regions serve important roles so far as the dynam-ics and functioning of the membrane is concerned. Depending onthe nature of the probe, the fluorophore can penetrate into the lesspolar region of the bilayer (i.e., towards the tail of the lipid) or in thepolar part of the bilayer (i.e., towards the aqueous phase) (Das etal., 2008). The initial decrease in the fluorescence intensity with aremarkable shift in the emission maximum at lower DMPG concen-tration indicates a significant lowering of polarity and a strongerbinding interaction of the cationic probe with the anionic lipo-some resulting from the electrostatic attraction. This is consistentto our studies with PSF in ctDNA environment, where a significantdecrease in the fluorescence intensity was observed with an appre-ciable blue shift indicating a lowering of polarity inside the DNAenvironment and a strong intercalative binding interaction (Sarkaret al., 2008b). DMPG vesicles are characterized by unusually lowturbidity and high viscosity (Riske et al., 2003). The reason is notyet clear but it is supposed to be related to the presence of chargedphosphate groups at the bilayer surface, which may cause a signifi-cant repulsion between the vesicles as well as between the adjacentor opposite lipids in the same bilayer, thus determining its packing.In fact, the structural characteristics of DMPG membranes are far
from being well understood (Riske et al., 2003). It is pertinent tomention here, that the structure of liposome does not change withincreasing concentrations of DMPG (Riske et al., 2003). Further-more, since the concentration of DMPG is in the millimolar range,whereas that of PSF is in the micromolar range, on average, thereis unlikely to be more than one PSF per vesicle.
Drawing analogy from this study, we infer that at higher DMPGconcentration, the large increase in the emission intensity withoutan appreciable shift perhaps indicates an increase in the compact-ness of the lipid around the PSF resulting in an increase in thelocal viscosity of the environment around the fluorophore. That anincrease in the viscosity of the medium leads to a large increasein the fluorescence of PSF has been established independently in aseries of water–glycerol mixture, where the two solvents have com-parable polarity (water, ET(30) = 63.1 and glycerol, ET(30) = 57.0)but differ widely in terms of viscosity. It is seen that with an increasein viscosity with increasing percentage of glycerol in the solu-tion mixture, the fluorescence intensity shows a large increase.This indicates that the increase of fluorescence intensity of PSFis profoundly dependent on the viscosity of the medium. Thus,the associated increase in fluorescence intensity at higher DMPGconcentration is rationalized from the effect of viscosity on theemission characteristic of PSF.
3.2. Partitioning of the dye
In order to realize the mechanism of access of the drug on mem-branes and the role of the lipid environments in the transportphenomenon, knowledge of the partitioning of the probe betweenthe membrane environment and the aqueous phase is important.The partition coefficient of the probe is defined as (Rodrigues et al.,2002)
KP = (CL/CT)/[L](CW/CT)/[W]
(5)
which CT is the molar concentration of the drug, CL and CW stand forthe probe concentration in lipid and in water media, and [L] and [W]represent molar concentrations of the lipid and water, respectively.The partition coefficient of PSF is determined by analyzing the flu-orescence data according to the following equation (Rodrigues etal., 2002; Coutinho and Prieto, 1995)
1�I
= 1�Imax
+ [W]�Imax × Kp
× [DMPG]−1 (6)
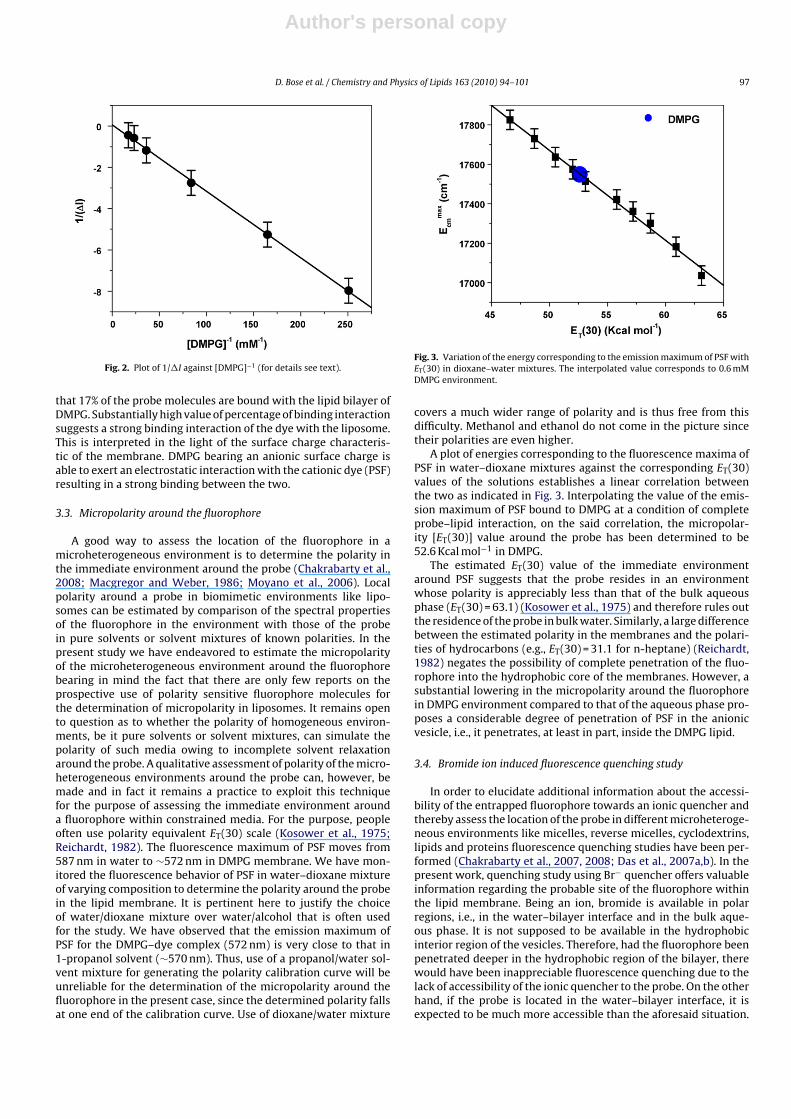
where �I = (I − I0) and �Imax = (I∞ − I0); I0, I and I∞ standing forfluorescence intensities of the fluorophore in the absence, at anintermediate lipid concentration and at the saturation level ofdye–DMPG interaction. Fig. 2 shows the plot of 1/(I − I0) against[DMPG]−1 and the partition coefficient value has been determinedfrom the slope and intercept of the plot.
The partition coefficient (Kp) for the fluorophore in the DMPGmembrane to aqueous phase is found to be 1.03 × 105. The remark-ably high value of the partition coefficient reveals that the probepartitions well in the liposome membrane of DMPG. In other words,the cationic probe undergoes a strong binding interaction with theanionic DMPG head group.
At the saturation level of interaction, using the determined valueof the partition coefficient, we have calculated the percentage ofprobe bound to the bilayers following the relation (Chakrabarty etal., 2008):
C0 = CT
(1 + Kp [L]/[W])(7)
where C0 and CT are concentrations of free and total probe, respec-tively. [L] and [W] are the molar concentrations of lipid and water,respectively and Kp is the partition coefficient. Calculation suggests
Author's personal copy
D. Bose et al. / Chemistry and Physics of Lipids 163 (2010) 94–101 97
Fig. 2. Plot of 1/�I against [DMPG]−1 (for details see text).
that 17% of the probe molecules are bound with the lipid bilayer ofDMPG. Substantially high value of percentage of binding interactionsuggests a strong binding interaction of the dye with the liposome.This is interpreted in the light of the surface charge characteris-tic of the membrane. DMPG bearing an anionic surface charge isable to exert an electrostatic interaction with the cationic dye (PSF)resulting in a strong binding between the two.
3.3. Micropolarity around the fluorophore
A good way to assess the location of the fluorophore in amicroheterogeneous environment is to determine the polarity inthe immediate environment around the probe (Chakrabarty et al.,2008; Macgregor and Weber, 1986; Moyano et al., 2006). Localpolarity around a probe in biomimetic environments like lipo-somes can be estimated by comparison of the spectral propertiesof the fluorophore in the environment with those of the probein pure solvents or solvent mixtures of known polarities. In thepresent study we have endeavored to estimate the micropolarityof the microheterogeneous environment around the fluorophorebearing in mind the fact that there are only few reports on theprospective use of polarity sensitive fluorophore molecules forthe determination of micropolarity in liposomes. It remains opento question as to whether the polarity of homogeneous environ-ments, be it pure solvents or solvent mixtures, can simulate thepolarity of such media owing to incomplete solvent relaxationaround the probe. A qualitative assessment of polarity of the micro-heterogeneous environments around the probe can, however, bemade and in fact it remains a practice to exploit this techniquefor the purpose of assessing the immediate environment arounda fluorophore within constrained media. For the purpose, peopleoften use polarity equivalent ET(30) scale (Kosower et al., 1975;Reichardt, 1982). The fluorescence maximum of PSF moves from587 nm in water to ∼572 nm in DMPG membrane. We have mon-itored the fluorescence behavior of PSF in water–dioxane mixtureof varying composition to determine the polarity around the probein the lipid membrane. It is pertinent here to justify the choiceof water/dioxane mixture over water/alcohol that is often usedfor the study. We have observed that the emission maximum ofPSF for the DMPG–dye complex (572 nm) is very close to that in1-propanol solvent (∼570 nm). Thus, use of a propanol/water sol-vent mixture for generating the polarity calibration curve will beunreliable for the determination of the micropolarity around thefluorophore in the present case, since the determined polarity fallsat one end of the calibration curve. Use of dioxane/water mixture
Fig. 3. Variation of the energy corresponding to the emission maximum of PSF withET(30) in dioxane–water mixtures. The interpolated value corresponds to 0.6 mMDMPG environment.
covers a much wider range of polarity and is thus free from thisdifficulty. Methanol and ethanol do not come in the picture sincetheir polarities are even higher.
A plot of energies corresponding to the fluorescence maxima ofPSF in water–dioxane mixtures against the corresponding ET(30)values of the solutions establishes a linear correlation betweenthe two as indicated in Fig. 3. Interpolating the value of the emis-sion maximum of PSF bound to DMPG at a condition of completeprobe–lipid interaction, on the said correlation, the micropolar-ity [ET(30)] value around the probe has been determined to be52.6 Kcal mol−1 in DMPG.
The estimated ET(30) value of the immediate environmentaround PSF suggests that the probe resides in an environmentwhose polarity is appreciably less than that of the bulk aqueousphase (ET(30) = 63.1) (Kosower et al., 1975) and therefore rules outthe residence of the probe in bulk water. Similarly, a large differencebetween the estimated polarity in the membranes and the polari-ties of hydrocarbons (e.g., ET(30) = 31.1 for n-heptane) (Reichardt,1982) negates the possibility of complete penetration of the fluo-rophore into the hydrophobic core of the membranes. However, asubstantial lowering in the micropolarity around the fluorophorein DMPG environment compared to that of the aqueous phase pro-poses a considerable degree of penetration of PSF in the anionicvesicle, i.e., it penetrates, at least in part, inside the DMPG lipid.
3.4. Bromide ion induced fluorescence quenching study
In order to elucidate additional information about the accessi-bility of the entrapped fluorophore towards an ionic quencher andthereby assess the location of the probe in different microheteroge-neous environments like micelles, reverse micelles, cyclodextrins,lipids and proteins fluorescence quenching studies have been per-formed (Chakrabarty et al., 2007, 2008; Das et al., 2007a,b). In thepresent work, quenching study using Br− quencher offers valuableinformation regarding the probable site of the fluorophore withinthe lipid membrane. Being an ion, bromide is available in polarregions, i.e., in the water–bilayer interface and in the bulk aque-ous phase. It is not supposed to be available in the hydrophobicinterior region of the vesicles. Therefore, had the fluorophore beenpenetrated deeper in the hydrophobic region of the bilayer, therewould have been inappreciable fluorescence quenching due to thelack of accessibility of the ionic quencher to the probe. On the otherhand, if the probe is located in the water–bilayer interface, it isexpected to be much more accessible than the aforesaid situation.
Author's personal copy
98 D. Bose et al. / Chemistry and Physics of Lipids 163 (2010) 94–101
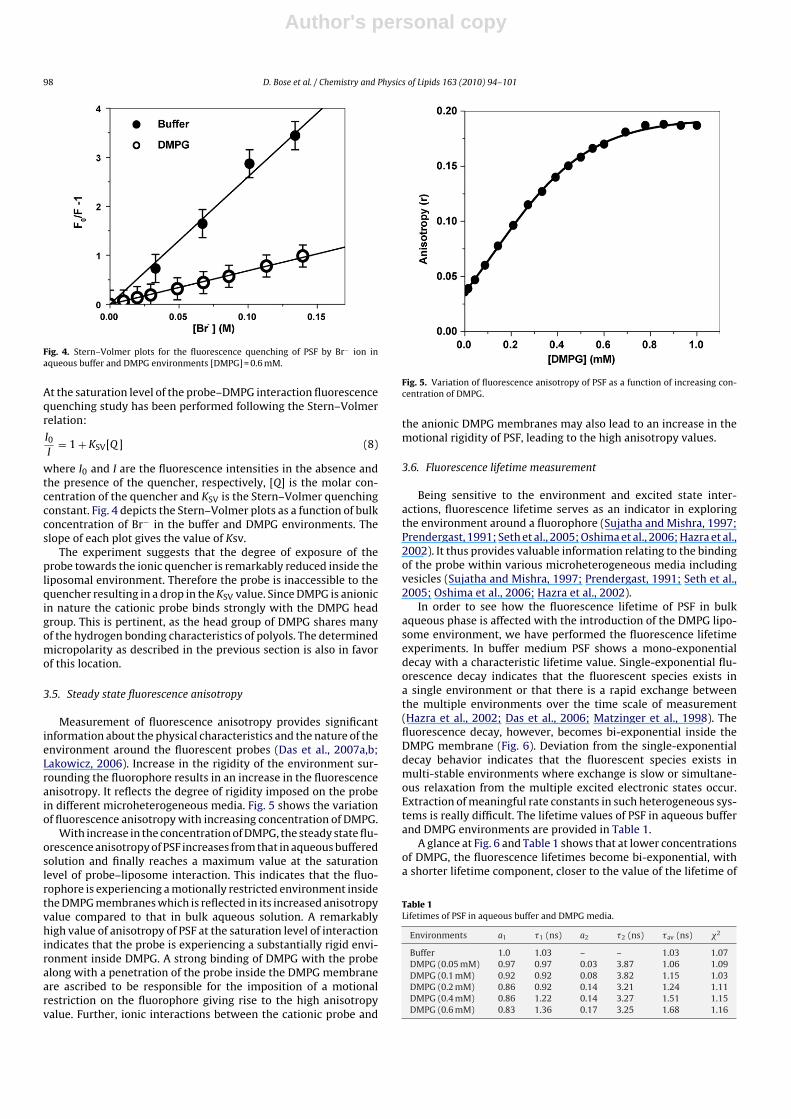
Fig. 4. Stern–Volmer plots for the fluorescence quenching of PSF by Br− ion inaqueous buffer and DMPG environments [DMPG] = 0.6 mM.
At the saturation level of the probe–DMPG interaction fluorescencequenching study has been performed following the Stern–Volmerrelation:
I0I
= 1 + KSV[Q ] (8)
where I0 and I are the fluorescence intensities in the absence andthe presence of the quencher, respectively, [Q] is the molar con-centration of the quencher and KSV is the Stern–Volmer quenchingconstant. Fig. 4 depicts the Stern–Volmer plots as a function of bulkconcentration of Br− in the buffer and DMPG environments. Theslope of each plot gives the value of Ksv.
The experiment suggests that the degree of exposure of theprobe towards the ionic quencher is remarkably reduced inside theliposomal environment. Therefore the probe is inaccessible to thequencher resulting in a drop in the KSV value. Since DMPG is anionicin nature the cationic probe binds strongly with the DMPG headgroup. This is pertinent, as the head group of DMPG shares manyof the hydrogen bonding characteristics of polyols. The determinedmicropolarity as described in the previous section is also in favorof this location.
3.5. Steady state fluorescence anisotropy
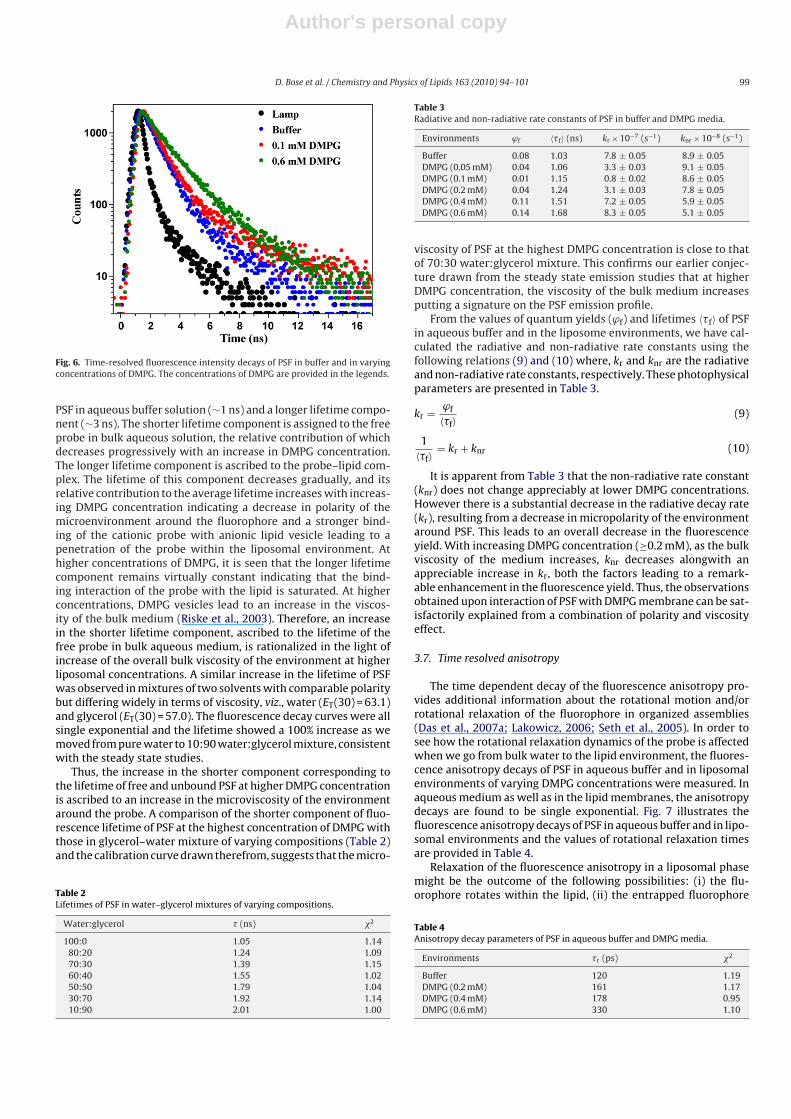
Measurement of fluorescence anisotropy provides significantinformation about the physical characteristics and the nature of theenvironment around the fluorescent probes (Das et al., 2007a,b;Lakowicz, 2006). Increase in the rigidity of the environment sur-rounding the fluorophore results in an increase in the fluorescenceanisotropy. It reflects the degree of rigidity imposed on the probein different microheterogeneous media. Fig. 5 shows the variationof fluorescence anisotropy with increasing concentration of DMPG.
With increase in the concentration of DMPG, the steady state flu-orescence anisotropy of PSF increases from that in aqueous bufferedsolution and finally reaches a maximum value at the saturationlevel of probe–liposome interaction. This indicates that the fluo-rophore is experiencing a motionally restricted environment insidethe DMPG membranes which is reflected in its increased anisotropyvalue compared to that in bulk aqueous solution. A remarkablyhigh value of anisotropy of PSF at the saturation level of interactionindicates that the probe is experiencing a substantially rigid envi-ronment inside DMPG. A strong binding of DMPG with the probealong with a penetration of the probe inside the DMPG membraneare ascribed to be responsible for the imposition of a motionalrestriction on the fluorophore giving rise to the high anisotropyvalue. Further, ionic interactions between the cationic probe and
Fig. 5. Variation of fluorescence anisotropy of PSF as a function of increasing con-centration of DMPG.
the anionic DMPG membranes may also lead to an increase in themotional rigidity of PSF, leading to the high anisotropy values.
3.6. Fluorescence lifetime measurement
Being sensitive to the environment and excited state inter-actions, fluorescence lifetime serves as an indicator in exploringthe environment around a fluorophore (Sujatha and Mishra, 1997;Prendergast, 1991; Seth et al., 2005; Oshima et al., 2006; Hazra et al.,2002). It thus provides valuable information relating to the bindingof the probe within various microheterogeneous media includingvesicles (Sujatha and Mishra, 1997; Prendergast, 1991; Seth et al.,2005; Oshima et al., 2006; Hazra et al., 2002).
In order to see how the fluorescence lifetime of PSF in bulkaqueous phase is affected with the introduction of the DMPG lipo-some environment, we have performed the fluorescence lifetimeexperiments. In buffer medium PSF shows a mono-exponentialdecay with a characteristic lifetime value. Single-exponential flu-orescence decay indicates that the fluorescent species exists ina single environment or that there is a rapid exchange betweenthe multiple environments over the time scale of measurement(Hazra et al., 2002; Das et al., 2006; Matzinger et al., 1998). Thefluorescence decay, however, becomes bi-exponential inside theDMPG membrane (Fig. 6). Deviation from the single-exponentialdecay behavior indicates that the fluorescent species exists inmulti-stable environments where exchange is slow or simultane-ous relaxation from the multiple excited electronic states occur.Extraction of meaningful rate constants in such heterogeneous sys-tems is really difficult. The lifetime values of PSF in aqueous bufferand DMPG environments are provided in Table 1.
A glance at Fig. 6 and Table 1 shows that at lower concentrationsof DMPG, the fluorescence lifetimes become bi-exponential, witha shorter lifetime component, closer to the value of the lifetime of
Table 1Lifetimes of PSF in aqueous buffer and DMPG media.
Environments a1 �1 (ns) a2 �2 (ns) �av (ns) �2
Buffer 1.0 1.03 – – 1.03 1.07DMPG (0.05 mM) 0.97 0.97 0.03 3.87 1.06 1.09DMPG (0.1 mM) 0.92 0.92 0.08 3.82 1.15 1.03DMPG (0.2 mM) 0.86 0.92 0.14 3.21 1.24 1.11DMPG (0.4 mM) 0.86 1.22 0.14 3.27 1.51 1.15DMPG (0.6 mM) 0.83 1.36 0.17 3.25 1.68 1.16
Author's personal copy
D. Bose et al. / Chemistry and Physics of Lipids 163 (2010) 94–101 99
Fig. 6. Time-resolved fluorescence intensity decays of PSF in buffer and in varyingconcentrations of DMPG. The concentrations of DMPG are provided in the legends.
PSF in aqueous buffer solution (∼1 ns) and a longer lifetime compo-nent (∼3 ns). The shorter lifetime component is assigned to the freeprobe in bulk aqueous solution, the relative contribution of whichdecreases progressively with an increase in DMPG concentration.The longer lifetime component is ascribed to the probe–lipid com-plex. The lifetime of this component decreases gradually, and itsrelative contribution to the average lifetime increases with increas-ing DMPG concentration indicating a decrease in polarity of themicroenvironment around the fluorophore and a stronger bind-ing of the cationic probe with anionic lipid vesicle leading to apenetration of the probe within the liposomal environment. Athigher concentrations of DMPG, it is seen that the longer lifetimecomponent remains virtually constant indicating that the bind-ing interaction of the probe with the lipid is saturated. At higherconcentrations, DMPG vesicles lead to an increase in the viscos-ity of the bulk medium (Riske et al., 2003). Therefore, an increasein the shorter lifetime component, ascribed to the lifetime of thefree probe in bulk aqueous medium, is rationalized in the light ofincrease of the overall bulk viscosity of the environment at higherliposomal concentrations. A similar increase in the lifetime of PSFwas observed in mixtures of two solvents with comparable polaritybut differing widely in terms of viscosity, viz., water (ET(30) = 63.1)and glycerol (ET(30) = 57.0). The fluorescence decay curves were allsingle exponential and the lifetime showed a 100% increase as wemoved from pure water to 10:90 water:glycerol mixture, consistentwith the steady state studies.
Thus, the increase in the shorter component corresponding tothe lifetime of free and unbound PSF at higher DMPG concentrationis ascribed to an increase in the microviscosity of the environmentaround the probe. A comparison of the shorter component of fluo-rescence lifetime of PSF at the highest concentration of DMPG withthose in glycerol–water mixture of varying compositions (Table 2)and the calibration curve drawn therefrom, suggests that the micro-
Table 2Lifetimes of PSF in water–glycerol mixtures of varying compositions.
Water:glycerol � (ns) �2
100:0 1.05 1.1480:20 1.24 1.0970:30 1.39 1.1560:40 1.55 1.0250:50 1.79 1.0430:70 1.92 1.1410:90 2.01 1.00
Table 3Radiative and non-radiative rate constants of PSF in buffer and DMPG media.
Environments ϕf 〈�f〉 (ns) kr × 10−7 (s−1) knr × 10−8 (s−1)
Buffer 0.08 1.03 7.8 ± 0.05 8.9 ± 0.05DMPG (0.05 mM) 0.04 1.06 3.3 ± 0.03 9.1 ± 0.05DMPG (0.1 mM) 0.01 1.15 0.8 ± 0.02 8.6 ± 0.05DMPG (0.2 mM) 0.04 1.24 3.1 ± 0.03 7.8 ± 0.05DMPG (0.4 mM) 0.11 1.51 7.2 ± 0.05 5.9 ± 0.05DMPG (0.6 mM) 0.14 1.68 8.3 ± 0.05 5.1 ± 0.05
viscosity of PSF at the highest DMPG concentration is close to thatof 70:30 water:glycerol mixture. This confirms our earlier conjec-ture drawn from the steady state emission studies that at higherDMPG concentration, the viscosity of the bulk medium increasesputting a signature on the PSF emission profile.
From the values of quantum yields (ϕf) and lifetimes 〈�f〉 of PSFin aqueous buffer and in the liposome environments, we have cal-culated the radiative and non-radiative rate constants using thefollowing relations (9) and (10) where, kr and knr are the radiativeand non-radiative rate constants, respectively. These photophysicalparameters are presented in Table 3.
kr = ϕf
〈�f〉(9)
1〈�f〉
= kr + knr (10)
It is apparent from Table 3 that the non-radiative rate constant(knr) does not change appreciably at lower DMPG concentrations.However there is a substantial decrease in the radiative decay rate(kr), resulting from a decrease in micropolarity of the environmentaround PSF. This leads to an overall decrease in the fluorescenceyield. With increasing DMPG concentration (≥0.2 mM), as the bulkviscosity of the medium increases, knr decreases alongwith anappreciable increase in kr, both the factors leading to a remark-able enhancement in the fluorescence yield. Thus, the observationsobtained upon interaction of PSF with DMPG membrane can be sat-isfactorily explained from a combination of polarity and viscosityeffect.
3.7. Time resolved anisotropy
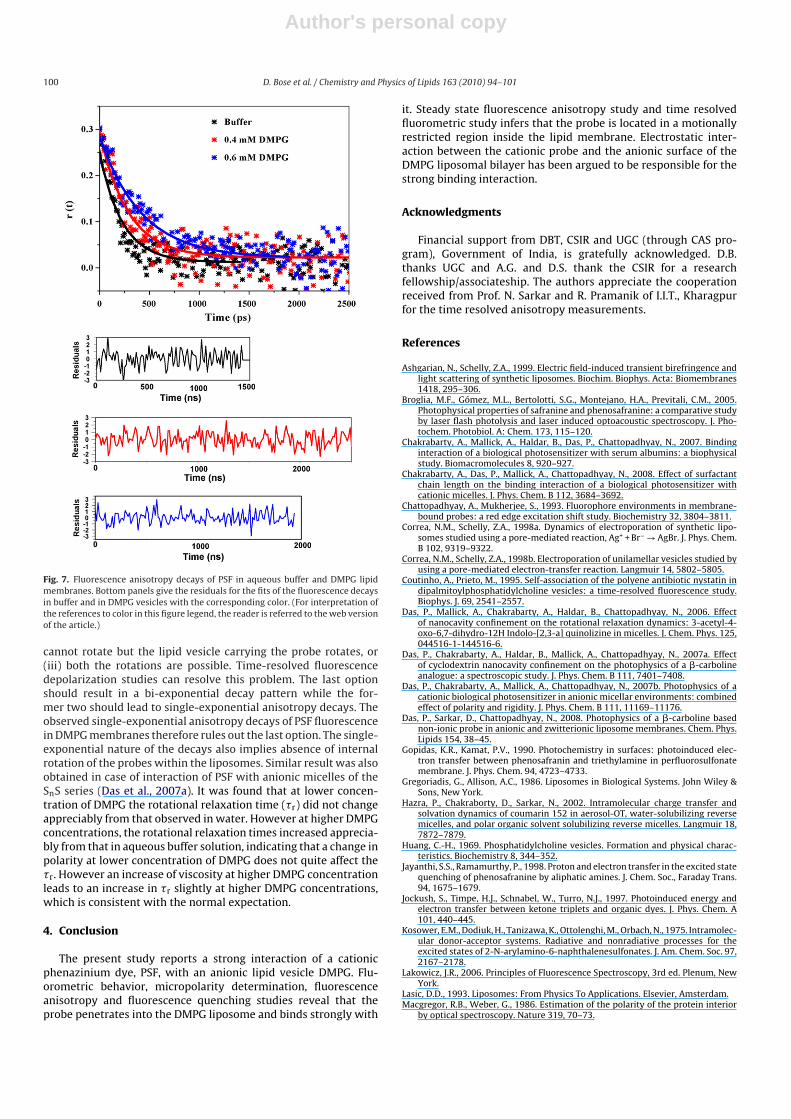
The time dependent decay of the fluorescence anisotropy pro-vides additional information about the rotational motion and/orrotational relaxation of the fluorophore in organized assemblies(Das et al., 2007a; Lakowicz, 2006; Seth et al., 2005). In order tosee how the rotational relaxation dynamics of the probe is affectedwhen we go from bulk water to the lipid environment, the fluores-cence anisotropy decays of PSF in aqueous buffer and in liposomalenvironments of varying DMPG concentrations were measured. Inaqueous medium as well as in the lipid membranes, the anisotropydecays are found to be single exponential. Fig. 7 illustrates thefluorescence anisotropy decays of PSF in aqueous buffer and in lipo-somal environments and the values of rotational relaxation timesare provided in Table 4.
Relaxation of the fluorescence anisotropy in a liposomal phasemight be the outcome of the following possibilities: (i) the flu-orophore rotates within the lipid, (ii) the entrapped fluorophore
Table 4Anisotropy decay parameters of PSF in aqueous buffer and DMPG media.
Environments �r (ps) �2
Buffer 120 1.19DMPG (0.2 mM) 161 1.17DMPG (0.4 mM) 178 0.95DMPG (0.6 mM) 330 1.10
Author's personal copy
100 D. Bose et al. / Chemistry and Physics of Lipids 163 (2010) 94–101
Fig. 7. Fluorescence anisotropy decays of PSF in aqueous buffer and DMPG lipidmembranes. Bottom panels give the residuals for the fits of the fluorescence decaysin buffer and in DMPG vesicles with the corresponding color. (For interpretation ofthe references to color in this figure legend, the reader is referred to the web versionof the article.)
cannot rotate but the lipid vesicle carrying the probe rotates, or(iii) both the rotations are possible. Time-resolved fluorescencedepolarization studies can resolve this problem. The last optionshould result in a bi-exponential decay pattern while the for-mer two should lead to single-exponential anisotropy decays. Theobserved single-exponential anisotropy decays of PSF fluorescencein DMPG membranes therefore rules out the last option. The single-exponential nature of the decays also implies absence of internalrotation of the probes within the liposomes. Similar result was alsoobtained in case of interaction of PSF with anionic micelles of theSnS series (Das et al., 2007a). It was found that at lower concen-tration of DMPG the rotational relaxation time (�r) did not changeappreciably from that observed in water. However at higher DMPGconcentrations, the rotational relaxation times increased apprecia-bly from that in aqueous buffer solution, indicating that a change inpolarity at lower concentration of DMPG does not quite affect the�r. However an increase of viscosity at higher DMPG concentrationleads to an increase in �r slightly at higher DMPG concentrations,which is consistent with the normal expectation.
4. Conclusion
The present study reports a strong interaction of a cationicphenazinium dye, PSF, with an anionic lipid vesicle DMPG. Flu-orometric behavior, micropolarity determination, fluorescenceanisotropy and fluorescence quenching studies reveal that theprobe penetrates into the DMPG liposome and binds strongly with
it. Steady state fluorescence anisotropy study and time resolvedfluorometric study infers that the probe is located in a motionallyrestricted region inside the lipid membrane. Electrostatic inter-action between the cationic probe and the anionic surface of theDMPG liposomal bilayer has been argued to be responsible for thestrong binding interaction.
Acknowledgments
Financial support from DBT, CSIR and UGC (through CAS pro-gram), Government of India, is gratefully acknowledged. D.B.thanks UGC and A.G. and D.S. thank the CSIR for a researchfellowship/associateship. The authors appreciate the cooperationreceived from Prof. N. Sarkar and R. Pramanik of I.I.T., Kharagpurfor the time resolved anisotropy measurements.
References
Ashgarian, N., Schelly, Z.A., 1999. Electric field-induced transient birefringence andlight scattering of synthetic liposomes. Biochim. Biophys. Acta: Biomembranes1418, 295–306.
Broglia, M.F., Gómez, M.L., Bertolotti, S.G., Montejano, H.A., Previtali, C.M., 2005.Photophysical properties of safranine and phenosafranine: a comparative studyby laser flash photolysis and laser induced optoacoustic spectroscopy. J. Pho-tochem. Photobiol. A: Chem. 173, 115–120.
Chakrabarty, A., Mallick, A., Haldar, B., Das, P., Chattopadhyay, N., 2007. Bindinginteraction of a biological photosensitizer with serum albumins: a biophysicalstudy. Biomacromolecules 8, 920–927.
Chakrabarty, A., Das, P., Mallick, A., Chattopadhyay, N., 2008. Effect of surfactantchain length on the binding interaction of a biological photosensitizer withcationic micelles. J. Phys. Chem. B 112, 3684–3692.
Chattopadhyay, A., Mukherjee, S., 1993. Fluorophore environments in membrane-bound probes: a red edge excitation shift study. Biochemistry 32, 3804–3811.
Correa, N.M., Schelly, Z.A., 1998a. Dynamics of electroporation of synthetic lipo-somes studied using a pore-mediated reaction, Ag+ + Br− → AgBr. J. Phys. Chem.B 102, 9319–9322.
Correa, N.M., Schelly, Z.A., 1998b. Electroporation of unilamellar vesicles studied byusing a pore-mediated electron-transfer reaction. Langmuir 14, 5802–5805.
Coutinho, A., Prieto, M., 1995. Self-association of the polyene antibiotic nystatin indipalmitoylphosphatidylcholine vesicles: a time-resolved fluorescence study.Biophys. J. 69, 2541–2557.
Das, P., Mallick, A., Chakrabarty, A., Haldar, B., Chattopadhyay, N., 2006. Effectof nanocavity confinement on the rotational relaxation dynamics: 3-acetyl-4-oxo-6,7-dihydro-12H Indolo-[2,3-a] quinolizine in micelles. J. Chem. Phys. 125,044516-1-144516-6.
Das, P., Chakrabarty, A., Haldar, B., Mallick, A., Chattopadhyay, N., 2007a. Effectof cyclodextrin nanocavity confinement on the photophysics of a �-carbolineanalogue: a spectroscopic study. J. Phys. Chem. B 111, 7401–7408.
Das, P., Chakrabarty, A., Mallick, A., Chattopadhyay, N., 2007b. Photophysics of acationic biological photosensitizer in anionic micellar environments: combinedeffect of polarity and rigidity. J. Phys. Chem. B 111, 11169–11176.
Das, P., Sarkar, D., Chattopadhyay, N., 2008. Photophysics of a �-carboline basednon-ionic probe in anionic and zwitterionic liposome membranes. Chem. Phys.Lipids 154, 38–45.
Gopidas, K.R., Kamat, P.V., 1990. Photochemistry in surfaces: photoinduced elec-tron transfer between phenosafranin and triethylamine in perfluorosulfonatemembrane. J. Phys. Chem. 94, 4723–4733.
Gregoriadis, G., Allison, A.C., 1986. Liposomes in Biological Systems. John Wiley &Sons, New York.
Hazra, P., Chakraborty, D., Sarkar, N., 2002. Intramolecular charge transfer andsolvation dynamics of coumarin 152 in aerosol-OT, water-solubilizing reversemicelles, and polar organic solvent solubilizing reverse micelles. Langmuir 18,7872–7879.
Huang, C.-H., 1969. Phosphatidylcholine vesicles. Formation and physical charac-teristics. Biochemistry 8, 344–352.
Jayanthi, S.S., Ramamurthy, P., 1998. Proton and electron transfer in the excited statequenching of phenosafranine by aliphatic amines. J. Chem. Soc., Faraday Trans.94, 1675–1679.
Jockush, S., Timpe, H.J., Schnabel, W., Turro, N.J., 1997. Photoinduced energy andelectron transfer between ketone triplets and organic dyes. J. Phys. Chem. A101, 440–445.
Kosower, E.M., Dodiuk, H., Tanizawa, K., Ottolenghi, M., Orbach, N., 1975. Intramolec-ular donor-acceptor systems. Radiative and nonradiative processes for theexcited states of 2-N-arylamino-6-naphthalenesulfonates. J. Am. Chem. Soc. 97,2167–2178.
Lakowicz, J.R., 2006. Principles of Fluorescence Spectroscopy, 3rd ed. Plenum, NewYork.
Lasic, D.D., 1993. Liposomes: From Physics To Applications. Elsevier, Amsterdam.Macgregor, R.B., Weber, G., 1986. Estimation of the polarity of the protein interior
by optical spectroscopy. Nature 319, 70–73.

Author's personal copy
D. Bose et al. / Chemistry and Physics of Lipids 163 (2010) 94–101 101
Matzinger, S., Hussey, D.M., Fayer, M.D., 1998. Fluorescent probe solubilization in theheadgroup and core regions of micelles: fluorescence lifetime and orientationalrelaxation measurements. J. Phys. Chem. B 102, 7216–7224.
Moyano, F., Biasutti, M.A., Silber, J.J., Correa, N.M., 2006. New insights on the behaviorof PRODAN in homogeneous media and in large unilamellar vesicles. J. Phys.Chem. B 110, 11838–11846.
Needham, D., Dewhirst, M.W., 2001. The development and testing of a newtemperature-sensitive drug delivery system for the treatment of solid tumors.Adv. Drug Deliv. Rev. 53, 285–305.
Oshima, J., Shiobara, S., Naoumi, H., Kaneko, S., Yoshihara, T., Mishra, A.K., Tobita,S., 2006. Extreme fluorescence sensitivity of some aniline derivatives to aque-ous and nonaqueous environments: mechanistic study and its implication as afluorescent probe. J. Phys. Chem. A 110, 4629–4637.
Prendergast, F.G., 1991. Time-resolved fluorescence techniques: methods and appli-cations in biology. Curr. Opin. Struct. Biol. 1, 1054–1059.
Reichardt, C., 1982. In: Ratajazak, H., Oriville-Thomas, W.J. (Eds.), Molecular Inter-actions, vol. 3. Wiley, New York.
Riske, K.A., Fernandez, R.M., Nascimento, O.R., Bales, B.L., Lamy-Freund, M.T., 2003.DMPG gel-fluid thermal transition monitored by a phospholipid spin labeled atthe acyl chain end. Chem. Phys. Lipids 124, 69–80.
Rodrigues, C., Gameiro, P., Reis, S., Lima, J.L.F.C., de Castro, B., 2002. Interactionof Grepafloxacin with large unilamellar liposomes:partition and fluores-
cence studies reveal the importance of charge interactions. Langmuir 18,10231–10236.
Sarkar, D., Das, P., Basak, S., Chattopadhyay, N., 2008a. Binding interaction of cationicphenazinium dyes with calf thymus DNA: a comparative study. J. Phys. Chem. B112, 9243–9249.
Sarkar, D., Das, P., Girigoswami, A., Chattopadhyay, N., 2008b. Spectroscopic char-acterization of phenazinium dye aggregates in water and acetonitrile media:effect of methyl substitution on the aggregation phenomenon. J. Phys. Chem. A112, 9684–9691.
Seth, D., Chakraborty, A., Setua, P., Chakraborty, D., Sarkar, N., 2005. Study of energytransfer from 7-amino coumarin donors to the rhodamine 6G acceptor in lecithinvesicles and sodium taurocholate-lecithin mixed aggregates. J. Phys. Chem. B109, 12080–12085.
Sujatha, J., Mishra, A.K., 1997. Effect of ionic and neutral surfactants on the propertiesof phospholipid vesicles: investigation using fluorescent probes. J. Photochem.Photobiol. A: Chem. 104, 173–178.
Sujatha, J., Mishra, A.K., 1998. Phase transition in phospholipid vesicles: excited stateprototropism of 1-naphthol as a novel probe concept. Langmuir 14, 2256–2262.
Teissie, J., Tsong, T.Y., 1981. Electric field induced transient pores in phospholipidbilayer vesicles. Biochemistry 20, 1548–1554.
Chen, W.-H., Regen, S.L., 2005. Thermally gated liposomes. J. Am. Chem. Soc. 127,6538–6539.





























