Embed Size (px)
Citation preview

Biochemical and Molecular Roles of nutrients
ß-Caroteneand Lutein Protect HepG2 Human Liver Cellsagainst Oxidant-Induced Damage1'2'3
KEITH R. MARTIN,4 MARK L. FAILLA5 AND J. CECIL SMITH, JR.*
Department of Food, Nutrition and Food Service Management, The university of North Carolinaat Greensboro,Greensboro, North Carolina 27412-500J, and *Carotenoids Research Unit,Beltsuille Human Nutrition Research Center, USDA, Beltsuiüe,MD 20705
ABSTRACT Numerous epidemiological studies support a strong inverse relationship between consumption of carotenoid-rich fruits and vegetables and theincidence of some degenerative diseases. One proposed mechanism of protection by carotenoids centerson their putative antioxidant activity, although directevidence in support of this contention is limited at thecellular level. The antioxidant potential of /J-carotene(BC) and lutein (LUT), carotenoids with or without provitamin A activity, respectively, was evaluated usingthe human liver cell line HepG2. Pilot studies showedthat a 90-min exposure of confluent cultures to 500/xmol/L iert-butylhydroperoxide (TBHP) at 37°Csignificantly (P < 0.05) increased lipid peroxidation andcellular leakage of lactate dehydrogenase (LDH), anddecreased the uptake of 3H-a-aminoisobutyric acidand 3H-2-deoxyglucose. Protein synthesis, mitochon
dria! activity and glucose oxidation were not affectedby TBHP treatment, suggesting that the plasma membrane was the primary site of TBHP-induced damage.Overnight incubation of cultures with •1 /xmol/L dl-a-tocopherol protected cells against oxidant-inducedchanges. In parallel studies, overnight incubation ofHepG2 in medium containing micelles with either BCor LUT (final concentrations of 1.1 and 10.9 /¿mol/L, respectively), the cell content of the carotenoidsincreased from <0.04 to 0.32 and 3.39 nmol/mg protein, respectively. Carotenoid-loaded cells were partially or completely protected against oxidant-inducedchanges in lipid peroxidation, LDH release and aminoacid and deoxyglucose transport. These data demonstrate that BC and LUT or their metabolites protectHepG2 cells against oxidant-induced damage and thatthe protective effect is independent of provitamin Aactivity. J. Nutr. 126: 2098-2106, 1996.
INDEXING KEY WORDS:
•carotenoids •antioxidant •amino acid transport•glucose transport •HepG2 human cell line
Carotenoids are naturally occurring pigments inplants that are involved in light-harvesting reactions
and protection of plant organdÃes against singlet oxygen-induced damage. Because free radical-induceddamage to mammalian tissues is believed to contributeto the aging process and to the development of somedegenerative diseases (Canfield et al. 1992), it has beenproposed that dietary carotenoids serve as antioxidantsin tissues (Thurnham 1994). This possibility is supported by numerous epidemiologie studies that indicate an inverse association between the increased intake of carotenoid-rich fruits and vegetables and theincidence of diseases such as cancer (Van Poppel andGoldbohm 1995). Additionally, in vitro studies havedemonstrated that carotenoids can inhibit chemicallyinduced neoplastic transformation (Bertram and Bort-kiewicz 1995), induce remission of oral leukoplakia(Garewal 1995), quench free radicals such as singletoxygen (Sies and Stahl 1995), and modulate immuneactivity (Meydani et al. 1995).
Possible mechanisms for carotenoid protection oftissues against oxidative damage include their antioxi-
1Preliminary reports of this research were presented at Experimental Biology 95, April 9-13, 1995, Atlanta, GA [Martin, K. R.,Failla, M. L. & Smith, J. C, Jr. |1995| /9-carotene and lutein protectHepG2 human liver cells against tert-butylhydroperoxide-induceddamage. FASEBJ.9: A1000 (abs.) and Experimental Biology 96, April14-17, 1996, Washington, DC [Martin, K. R., Failla, M. L. & Smith,J.C., Jr.11996)/0-Carotene and lutein attenuate oxidant-induced damage to plasma membrane in HepG2 human liver cells. FASEB J. 10:A732 (abs.|] and in the AIN Graduate Student Research Award Abstract Competition.
2Supported in part by the North Carolina Agricultural Experiment Station and USDA NRI 94-37207-1035.
3The costs of publication of this article were defrayed in part by
the payment of page charges. This article must therefore be herebymarked "advertisement" in accordance with 18 USC section 1734
solely to indicate this fact."Current address: Nutritional Immunology Laboratory, USDA
Human Nutrition Research Center on Aging at Tufts University,Boston, MA 02111.
5To whom correspondence should be addressed.
0022-3166/96 $3.00 ©1996 American Institute of Nutrition.Manuscript received 18 March 1996. Initial review completed 5 April 1996. Revision accepted 4 June 1996.
2098
by guest on July 21, 2011jn.nutrition.org
Dow
nloaded from

CAROTENOIDS AS ANTIOXIDANTS IN HUMAN LIVER CELLS 2099
dant capacity, interactions with other antioxidants andantioxidant enzymes, and conversion to retinoids andapocarotenoids (Canfìeldet al. 1990). Critical evaluation of the antioxidant potential of carotenoids in humans has been hampered in part by the relative lackof appropriate animal models. Human cell lines providea relevant model for investigating carotenoid function.The availability of highly differentiated human celllines with phenotypic properties similar to mature he-patocytes is particularly pertinent because the liver actively metabolizes drugs and other xenobiotics and generates high levels of free radicals (Moshage and Yap1992). We selected HepG2 human liver cells as ourmodel because these cells secrete plasma proteins andlipoproteins, express an inducible cytochrome P450system, and respond to hormones and cytokines in amanner similar to normal human liver (Javitt 1990).The objective of the present study was to determinewhether pretreatment of HepG2 cells with /3-carotene(BC)6and lutein (LUT), i.e., carotenoids with or withoutpro-vitamin A activity, respectively, confers cytopro-tection when cultures are subjected to oxidative stress.
MATERIALS AND METHODS
Supplies. All reagents were obtained from SigmaChemical (St. Louis, MO) unless otherwise noted. D-[U-14C]-glucose (sp. act. 89 MBq/mmol) and D-[6-14C]-glucose (sp. act. 181 MBq/mmol) and a-[methyl-3H]-
aminoisobutyric acid (sp. act. 252 GBq/mmol) werepurchased from DuPont/New England Nuclear (Wilmington, DE). 2-Deoxy-D-[2,6-3H] glucose (sp. act. 999GBq/mmol) and [4, 5-3H]-leucine (sp. act. 6 TBq/mmol)
were obtained from Amersham (Arlington Heights, IL).Tissue culture flasks (75 and 25 cm2) and multiwell
dishes were obtained from Costar (Cambridge, MA).Purified BC, LUT and echinenone were gifts from Dr.Fred Khachik, USDA, Beltsville Human Nutrition Research Center.
Cell culture. HepG2 cells were purchased fromAmerican Type Tissue Collection (ATCC, Rockville,MD) and stock cultures were maintained in T75 flasks.For experiments, cells were seeded in multiwell plasticdishes (15- or 35-mm diameter) or T25 flasks and incubated in a humidified atmosphere of air/CO2 (95:5 v/v)at 37°C.HepG2 cultures (passages 80-90) were grown
in minimal essential medium eagle (MEM) supplemented with 100 mL/L heat-inactivated fetal bovineserum (FBS), 1 mmol/L pyruvate, 2 mmol/L glutamine,
6Abbreviations used: AIB, a-aminoisobutyric acid; a-TC, a-to-copherol; BC, ,9-carotene; BHT, butylated hydroxytoluene; DOG, D-2-deoxyglucose; FBS, fetal bovine serum; HBSS, Hank's balanced salt
solution; LDH, láclate dehydrogenase; LUT, lutein; MEM, minimalessential medium eagle; MTT, 3[4,5-dimethylthiazol-2-yl]-2,5-diphe-nyl tetrazolium bromide,- TEARS, thiobarbituric acid reactive substances; TBHP, fert-butylhydroperoxide; TCA, trichloroacetic acid.
89 ¿zmol/Lgentamicin, 0.54 ¿zmol/Lfungizone and 15mmol/L N-2-hy droxyethylpiperazine-N' -2-ethanesul-
fonic acid (HEPES), pH 7.0 (complete medium). Medium was replaced every 2 d and experiments wereconducted with cultures at 1-3 d post-confluency.
Experiments were initiated by washing confluentcultures twice with Hank's balanced salt solution(HBSS) without phenol red at 37°C.Serum-free com
plete MEM alone (control) or containing 500 /^mol/Ltert-butylhydroperoxide (TBHP) was added and cultures were incubated at 37°Cfor 90 min. The indicated
conditions for the oxidative stress were selected as appropriate after performing pilot studies that examinedthe effects of varying the concentration of TBHP andlength of exposure on cell viability, levels of thiobarbituric acid reactive substances (TBARS) and lactate dehydrogenase (LDH) release from cell monolayers (seebelow).
Micellar preparation and cellular uptake of carotenoids and a-tocopherol. Carotenoids and dl-a-to-copherol (a-TC) were delivered to cells as micelles thathad been prepared by modification of the procedureof El-Gorab and Underwood (1973). All manipulationswith carotenoids were performed under yellow fluorescent lights to minimize photodecomposition. Appropriate volumes of stock solutions of the followingcompounds in either ethanol, hexane or chloroform/methanol were transferred to glass bottles to yield thefollowing final concentrations after dilution with complete medium: 1-a-phosphatidylcholine (16.7 ¿miol/L),monoolein (100 /umol/L), taurocholic acid (1.7 mmol/L), oleic acid (33.3 ^mol/L) and either BC, LUT or a-TC (15 /miol/L). Pilot studies were performed to examine the cytotoxic potential of the micellar preparationat the indicated and higher concentrations in which theratio of the individual components was kept constant.Morphological appearance of the monolayer and releaseof cytoplasmic LDH (see below) were the same in cultures incubated for at least 24 h in complete MEMalone or containing micelles with the components atthe final concentrations listed above. However, increasing the quantity of the micellar components by>50% had an adverse impact on the integrity of themonolayer.
After solvents were evaporated with a gentle streamof nitrogen, complete cell culture medium was addedto the bottles. Samples were transferred to a water bathsonicator (Sonic Systems, Newtown, PA) at room temperature and sonicated for three cycles of 5 min eachwith exchange of bath water between cycles. Sampleswere filter sterilized (SFCA syringe filter with 0.22-//mpore; Nalgene, Rochester, NY) to remove microcrystal-
line particles.The concentration of micellar a-TC and carotenoids
in filtered medium was measured by HPLC (Bieri et al.1979 and 1985). As indicated in Table 1, the filteredmicellar preparation contained 73% of the LUT and94% of the a-TC originally added. In contrast, filtered
by guest on July 21, 2011jn.nutrition.org
Dow
nloaded from

2100 MARTIN ET AL.
TABLE 1
Medium concentrations and accumulation of a-tocopheroland carotenoids by HepG2 cells1
/3-caroteneLuteina-TocopherolPost-filtered
medium^funol/L1.1
±0.31»10.9 ±0.17b14.1 ±1.49CCells3nmol/mg
protein0.32
±0.07»3.39 ±0.34C2.58 ±0.35b
1Values are means ±SEM,n = 4 for post-filtered medium and n= 6 for cells. Within a column, means with different superscriptsdiffer significantly, P < 0.05.
2 Medium contained 15 ¿imol/Lof the carotenoids or a-tocopherolbefore sterilization using a syringe filter with 0.22-/xm pores.
3 Mean levels of carotenoids and a-tocopherol in cells after incubation in medium containing micelles for only 20 h were <0.04 and0.26 nmol/mg protein, respectively.
samples contained only 7% of the BC added to theoriginal sample, i.e., 1.1 /zmol/LBC. Relative solubilityof BC in the micellar preparation, i.e., 7%, was similarto that reported by Canfield et al. (1990), viz., 4-13%.
To examine cellular accumulation of the lipophilictest compounds from medium with micelles, confluentmonolayers of HepG2 were exposed overnight (20 h) toMEM containing the micellar preparation alone or withone of BC, LUT or a-TC. After removal of medium,monolayers were washed two times with HBSS, collected in 1 mL of 0.1 mmol/L phosphate buffer containing 100 mL/L ethanol with 0.45 mmol/L butylatedhydroxytoluene (BHT) and sonicated 3-5 s using aSonic Dismembranator at 60% output (Fisher Scientific, Pittsburgh, PA). Aliquots (500 p.L]of the sonicatewere diluted with ethanol (500 /¿L)containing echinen-one (0.36 /xmol/L) and tocol (64 ¿imol/L)as internalstandards for analysis of carotenoids and a-tocopherol,respectively. Samples were extracted into hexane containing BHT (0.45 mmol/L), dried under nitrogen, reconstituted in the appropriate mobile phase and analyzed by HPLC as described below. The concentrationof LUT in cell homogenates was ~ 10-fold that of BC,whereas the concentration of a-TC was intermediatebetween LUT and BC (Table 1).
HPLC analysis. Carotenoids and a-TC were analyzed independently using HPLC (Hewlett-Packard,Model HP-1090, Avondale, PA) with a UV/visible dualbeam detector with wavelengths set at 450 or 292 nmfor carotenoids and a-TC, respectively, as described byBieri et al. (1979 and 1985). The column was a Micro-sorb-MV 5-Aim25 cm X 0.46 cm diameter C-18 ODS(Rainin, Woburn, MA) and was protected by a CIS Ad-sorbosphere, 5-/xm particle size, 0.75 X 0.46 cm guardcolumn (Alltech, Deerfield, IL). The mobile phase forthe analysis of carotenoids was 70:20:10 (v/v/v) aceto-nitrile (with 1.3mL/L triethylaminej/methylene chlo-ride/methanol containing 0.13 mmol/L ammonium acetate. Methanol was used as the mobile phase for the
analysis of a-TC. The flow rates throughout the iso-cratic analyses were 1.7 and 2.5 mL/min for analysisof carotenoids and a-TC, respectively. The columntemperature was maintained at 20"C by using a 40-cmHPLC column water jacket (Alltech) and a constant-temperature circulator (Fisher model 910; Fisher Scientific, McGaw Park, IL). Data were calculated based onthe analysis of a series of external standards that were>94% pure as verified by HPLC.
Evaluation of cytoprotectiue activity of carotenoidsand a-TC against TBHP-induced damage
Thioborbituric acid reactive substances (TEARS).Peroxidation of membrane lipids was estimated bymeasuring the levels of TEARS in cultures (mediumplus monolayer) immediately after a 90-min exposure±500 /zmol/L TBHP by a modification of the method ofBuege and Aust (1978).Briefly, 100 /¿Labsolute ethanolcontaining 4.5 mmol/L BHT was added to wells (9.6cm2) containing 2 mL MEM alone or with 500 ¿¿mol/LTBHP to quench further oxidation. An aliquot (1 mL)of the test medium was transferred to a 5-mL glassscrew cap tube and residual medium was discarded.The monolayer was washed three times with HBSS atroom temperature. Cellular material was collected in1 mL harvest solution containing 100 mL/L ethanolwith 4.5 mmol/L BHT, 3.5 mmol/L sodium dodecylsulfate, and 1 mmol/L disodium ethylenediaminetetra-acetic acid. After sonication for 10-20 s, one half ofthe sample (500 /xL)was combined with the aliquot ofmedium that had been transferred to the glass tubeto determine the total TBARS in cultures. Remainingsonicate was stored at -20°C for protein analysis.Tubes were heated at 95°Cfor 30 min after addition of
2 mL TBA solution [26 mmol/L thiobarbituric acid and920 mmol/L trichloroacetic acid (TCA) in 250 mmol/LHC1].Samples were cooled in ice, mixed, and aliquotswere removed and centrifuged at 16,000 X g for 10 minat 25°C.Supernate was analyzed spectrophotometri-cally at 532 nm. Tetramethoxypropane was used as astandard and various concentrations were processedidentically to samples. Data are expressed as pmolTBARS/mg cell protein.
Lactate dehydrogenase (LDH) release. The releaseof the cytoplasmic enzyme LDH (EC 1.1.1.27) into themedium by HepG2 monolayers during exposure toTBHP was used to assess the integrity of the cell membrane. Aliquots of medium were centrifuged for 5 minat 700 x g at 4°Cto remove debris. Monolayers werewashed with HBSS (25°C)and collected after incubation for 15 min in 1 mL diluted Triton X-100 (10 mL/L). Cellular material was sonicated for 5-10 s on ice,and the samples were centrifuged at 16,000 x g for 8min at 4°C.LDH activity was determined according to
Glascott et al. (1992). The reaction mixture contained2.8 mL of 0.1 mol/L phosphate buffer (pH 7.5) with 85
by guest on July 21, 2011jn.nutrition.org
Dow
nloaded from

CAROTENOIDS AS ANTIOXIDANTS IN HUMAN LIVER CELLS 2101
/¿mol/LNADH, 100 //L 23 mmol/L pyruvic acid, and500 /¿Lsupernate from either undiluted medium or cellhomogenate that had been diluted 1:50 with phosphatebuffer. The rate of change in absorption at 340 nm wasmonitored for 3 min (Beckman DU-640 UV/vis spectro-photometer, Fullerton, CA). The percentage of cellularLDH released to the medium was calculated as follows:% LDH release = [LDH activity in medium / (LDHactivity in medium + LDH activity in cells)] x 100.Pilot studies using either various dilutions of supernatefrom sonicated HepG2 cells that were not exposed toTBHP or purified rabbit muscle LDH showed that enzyme activity was not altered after exposure to 500/¿mol/LTBHP for 90 min at 37°C.
14C-Glucose oxidation. After incubating cultures
with or without TBHP for 90 min, monolayers (T25flasks) were washed twice with HBSS at 37°C.MEM
(5.5 mmol/L glucose) containing 100 mL/L FBS and 37kBq of either D-[U-14C]- or D-[6-14C]-glucose was addedto each flask and cultures incubated at 37°Cfor 2 h.Production of 14CO2was measured as described by Ell-
wood et al. (1993). Data are expressed as nmol glucoseoxidized/(h •mg cell protein).
Uptake and incorporation of 3H-leucine. After re
moval of medium ±TBHP, monolayers were washedtwice with HBSS before adding MEM containing 100mL/L FBS and 9.25 kBq [4,5-3H]-leucine. Cultures wereincubated for l h at 37°C.Medium was collected and
the cell monolayer washed twice with ice-cold HBSSand harvested as described previously. Cell homoge-nates were sonicated for 3-5 s, and an aliquot (200 /¿L)added to cold TCA (306 mmol/L) to precipitate 3H-
protein, vortexed and stored in ice for 15 min. Precipitated material was collected by centrifugation for 10min at 16,000 x g, washed twice with 306 mmol/LTCA and resuspended in a small volume of 1 mol/Lsodium hydroxide. Aliquots were transferred to vialscontaining 4 mL Ultima Gold scintillation cocktail(Packard, Meriden, CT) and analyzed by liquid scintillation spectrophotometry (Beckman model 6000SE LScounter,- Fullerton, CA) to quantify 3H. To determinethe amount of 3H-protein secreted into the medium, a
200-//L sample was added to a tube containing an equalvolume of water with bovine serum albumin (1 g/L) ascarrier and processed as described for the cell homoge-nates. The level of acid-precipitable 3H in cells and
medium represents protein synthesized during the labeling period. An additional aliquot of cell sonicate alsowas analyzed to determine total cellular radioactivity.3H-Leucine uptake was estimated by adding total cellular 3H and secretory protein 3H.
Transport of 3H-aminoisobutyric acid (*H-AIB)and 3H-deoxyglucose (3H-DOG). Control and TBHP-
treated monolayers were washed twice with 2 mL HBSSbefore adding an amino acid-free formulation of MEMwithout FBS and containing 3H-AIB (9.25 MBq/L) and
100 /¿mol/LAIB. Monolayers were incubated for 3 minat 37°C,washed twice with 2 mL amino acid-free MEM
at 0°Cand collected in 1 mL harvest solution. To measure 3H-DOG uptake, control and TBHP-treated cul
tures were incubated with a glucose-free formulation ofMEM without FBS and containing 3H-DOG (9.25 MBq/
L) and 100 //mol/L DOG for 10 min. Radiolabeled medium was removed and monolayers were washed twicewith 2 mL ice-cold, glucose-free MEM containing 200/¿mol/Lphloretin to inhibit DOG efflux. Pilot studiesshowed that uptakes of 3H-AIB and 3H-DOG were linear
for 4 and 20 min, respectively, using the described conditions. Cellular 3H was measured as described aboveand data are expressed as nmol/(min •mg protein).
Mitochondria! activity. To examine the impact oftreatment of cells with TBHP on mitochondrial activity, control and treated cells were washed twice withHBSS before the addition of serum-free MEM containing 60 /¿mol/L 3[4,5-dimethylthiazol-2-yl]-2,5-di-phenyltetrazolium bromide (MTT) for 30, 60 or 90 min.At indicated times, MEM was removed from replicatewells and dimethyl sulfoxide (1 mL) was added to lysecells and solubilize reduced crystals of MTT. After mixing for 5 min, the increase in optical density of reducedMTT was determined at 560 nm by UV/visible spectrophotometry (Scudiero et al. 1988).
Protein. Cellular protein was quantified as previously described (Ellwood et al. 1993).
Statistics. Data are expressed as means ±SEM.Allexperiments were repeated at least twice using triplicate cultures for each variable. When necessary, datawere transformed (Iog10) to stabilize variance amongtreatment groups. Either Student's i test or one-wayANOVA with post-hoc analysis using Tukey's multi
ple range procedure was used to determine statisticallysignificant differences (P < 0.05) between means(Snedecor and Cochrane 1989).
RESULTS
Prior to evaluating the cytoprotective influence ofcarotenoids against prooxidant injury, it was necessaryto establish a positive control for the protection ofHepG2 cultures against TBHP-induced damage. Mono-layers were incubated overnight in complete mediumcontaining micelles with increasing concentrations ofa-TC to examine the relationship between cellular content of a-TC and susceptibility to TBHP-induced injury. Cellular a-TC was increased significantly whenthe medium was supplemented with >3 /¿mol/La-TC(Fig. 1, upper panel] and was correlated positively (r =0.84) with medium a-TC between 1 and 30 /¿mol/L.The level of TEARS was measured to assess peroxida-tion of membrane polyunsaturated fatty acids in control and TBHP-treated samples. TBHP-induced elevation of TEARS in cultures was attenuated by pretreatment of monolayers with medium containing 1-30/¿mol/La-TC (Fig. 1, lower panel}. TBARS levels in
by guest on July 21, 2011jn.nutrition.org
Dow
nloaded from
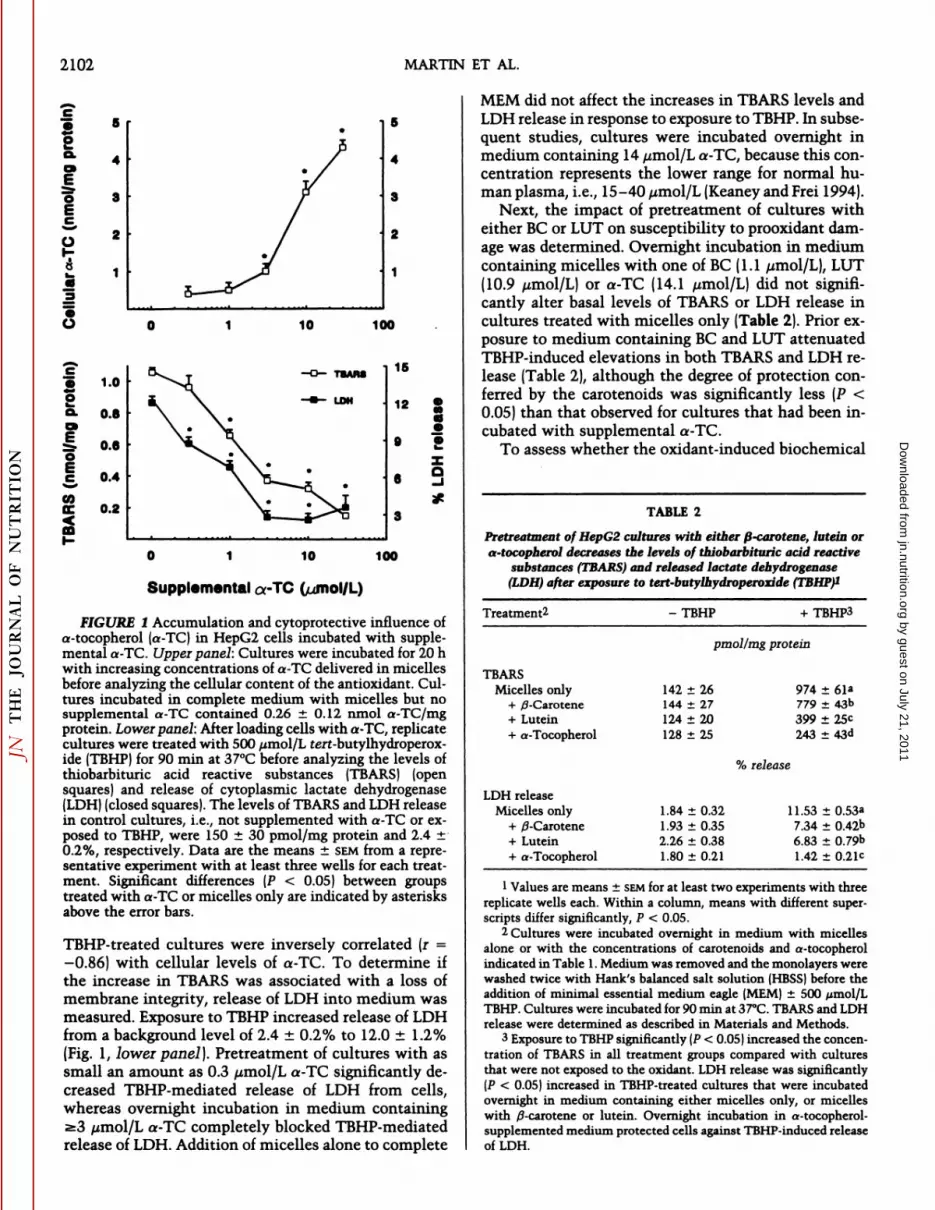
2102 MARTIN ET AL.
10 100
eE
coe
i
1.0
0.8
0.6
0.4
0.2
16
12 gOS
8xo
10 100
Supplemental orTC
FIGURE I Accumulation and cytoprotective influence ofa-tocopherol (a-TC) in HepG2 cells incubated with supplemental a-TC. Upper panel: Cultures were incubated for 20 hwith increasing concentrations of a-TC delivered in micellesbefore analyzing the cellular content of the antioxidant. Cultures incubated in complete medium with micelles but nosupplemental a-TC contained 0.26 ±0.12 nmol a-TC/mgprotein. Lower panel: After loading cells with a-TC, replicatecultures were treated with 500 ¿¿mol/Lferf-butylhydroperox-ide (TBHP) for 90 min at 37°Cbefore analyzing the levels of
thiobarbituric acid reactive substances (TEARS) (opensquares) and release of cytoplasmic lactate dehydrogenase(LDH) (closed squares). The levels of TEARS and LDH releasein control cultures, i.e., not supplemented with a-TC or exposed to TBHP, were 150 ±30 pmol/mg protein and 2.4 ±0.2%, respectively. Data are the means ±SEMfrom a representative experiment with at least three wells for each treatment. Significant differences (P < 0.05) between groupstreated with a-TC or micelles only are indicated by asterisksabove the error bars.
TBHP-treated cultures were inversely correlated (r =-0.86) with cellular levels of a-TC. To determine if
the increase in TBARS was associated with a loss ofmembrane integrity, release of LDH into medium wasmeasured. Exposure to TBHP increased release of LDHfrom a background level of 2.4 ±0.2% to 12.0 ±1.2%(Fig. 1, lower panel). Pretreatment of cultures with assmall an amount as 0.3 //mol/L a-TC significantly decreased TBHP-mediated release of LDH from cells,whereas overnight incubation in medium containinga3 /¿mol/La-TC completely blocked TBHP-mediatedrelease of LDH. Addition of micelles alone to complete
MEM did not affect the increases in TBARS levels andLDH release in response to exposure to TBHP. In subsequent studies, cultures were incubated overnight inmedium containing 14 //mol/L a-TC, because this concentration represents the lower range for normal human plasma, i.e., 15-40 ¿tmol/L(Keaney and Frei 1994).
Next, the impact of pretreatment of cultures witheither BC or LUT on susceptibility to prooxidant damage was determined. Overnight incubation in mediumcontaining micelles with one of BC (1.1 //mol/L), LUT(10.9 //mol/L) or a-TC (14.1 /zmol/L) did not significantly alter basal levels of TBARS or LDH release incultures treated with micelles only (Table 2). Prior exposure to medium containing BC and LUT attenuatedTBHP-induced elevations in both TBARS and LDH release (Table 2), although the degree of protection conferred by the carotenoids was significantly less (P <0.05) than that observed for cultures that had been incubated with supplemental a-TC.
To assess whether the oxidant-induced biochemical
TABLE 2
Pretreatment of HepG2 cultures with either ß-carotene, lutein ora-tocopherol decreases the levels of thiobarbituric acid reactive
substances (TBARS} and released lactate dehydrogenase(LDH) after exposure to tert-butylhydroperoxide (TBHP)1
Treatment2 -TBHP + TBHP3
pmol/mg protein
TBARSMicelles only
+ ^-Carotene+ Lutein+ a-Tocopherol
LDH releaseMicelles only
+ /3-Carotene+ Lutein+ a-Tocopherol
142 ±26144 ±27124 ±20128±25
974 ±61*779 ±43b399 ±25C243 ±43d
% release
1.84 ±0.321.93 ±0.352.26 ±0.381.80 ±0.21
11.53 ±0.53»7.34 ±0.42b6.83 ±0.79b1.42 ±0.21C
1Values are means ±SEMfor at least two experiments with threereplicate wells each. Within a column, means with different superscripts differ significantly, P < 0.05.
2 Cultures were incubated overnight in medium with micellesalone or with the concentrations of carotenoids and a-tocopherolindicated in Table 1. Medium was removed and the monolayers werewashed twice with Hank's balanced salt solution (HBSS) before the
addition of minimal essential medium eagle (MEM| ±500 ¿/mol/LTBHP. Cultures were incubated for 90 min at 37°C.TBARS and LDH
release were determined as described in Materials and Methods.3 Exposure to TBHP significantly (P < 0.05] increased the concen
tration of TBARS in all treatment groups compared with culturesthat were not exposed to the oxidant. LDH release was significantly(P < 0.05) increased in TBHP-treated cultures that were incubatedovernight in medium containing either micelles only, or micelleswith /9-carotene or lutein. Overnight incubation in a-tocopherol-supplemented medium protected cells against TBHP-induced releaseof LDH.
by guest on July 21, 2011jn.nutrition.org
Dow
nloaded from

CAROTENOIDS AS ANTIOXIDANTS IN HUMAN LIVER CELLS 2103
TABLE 3
Effect of exposure to tert-butylhydroperoxide (TBHP) onaccumulation and incorporation of 3H-leucine in cultures of
HepG2 pretreated overnight with either ß-carotene,lutein or a-tocopherol1
Treatment - TBHP + TBHP
nmol/mg protein
Cellular accumulation of3H-leucine
Micelles only+ /3-Carotene+ Lutein+ a-Tocopherol
202 ±3195 ±17190 ±21217 ±19
110 ±5'
199 ±12172 ±7219 ±12
Acid-precipitated(% total 3H)
3H-Leucineincorporationinto
proteinMicellesonly+/3-Carotene+Lutein+
a-Tocopherol36.0
±1.644.6±8.341.3±2.734.9±1.435.5
±1.341.2±3.437.9±2.231.1±3.5
1Values are means ±SEMfor at least two experiments with threereplicate wells per experiment. The presence of an asterisk as superscript indicates that the mean value for cultures treated with TBHPdiffers significantly (P < 0.05) from cultures that were not exposedto the oxidant.
2 Cultures were treated as described in the footnote for Table 2.After removal of medium ±TBHP, monolayers were washed. Complete minimal essential medium eagle (MEM) containing [4,5-3Hj-leucine was added to each well, and cultures were incubated at 37°C
for 1 h. Total cellular 3H and the percentage of 3H incorporated intoprotein were determined as described in Materials and Methods.
changes outlined above were associated with alterations in cellular activities, the uptake and incorporation of 3H-leucine into acid-precipitable protein wascompared in control and TBHP-treated cultures. Exposure of cultures to TBHP decreased cellular accumulation of 3H-leucine in 60 min by 52% (15.8 ±0.2 vs. 7.6±0.4 nmol/mg protein for control and TBHP-treatedcells, respectively; P < 0.05). Overnight incubation ofHepG2 with micelles containing BC, LUT and a-TCcompletely protected cells against TBHP-induced reduction of 3H-leucine accumulation (Table 3). The percentage of cellular 3H present in the acid insoluble fraction was similar in both control and TBHP-treated cultures suggesting that protein synthesis was notsignificantly altered by exposure to the prooxidant.These data suggested that TBHP impaired amino aciduptake. To address this possibility, the unidirectionaltransport of the non-metabolizable amino acid analogue a-aminoisobutyric acid (3H-AIB) was measured
in control and TBHP-treated cultures. As shown in Figure 2, treatment of cultures with TBHP decreased 3H-
AIB uptake to 16% of that of control [0.71 ±0.04vs.4.42 ±0.13 nmoljmin-mg protein), respectively; P <
0.05}. Overnight incubation of cultures in medium con
taining BC and LUT attenuated the impact of TBHPtreatment on 3H-AIB transport; cellular uptake of 3H-
AIB by BC- and LUT-treated cultures exposed to TBHPwas 33 and 60%, respectively,of that of the control (P< 0.05). Prior supplementation of cultures with a-TCwas more effective than the carotenoids at blockingTBHP-induced damage to the 3H-AIB transport process.
In a parallel series of studies, the impact of oxidantexposure on glucose utilization by HepG2 cells wasexamined. Cultures incubated with uniformly labeled(U)-14C-glucose after exposure to TBHP generated only56% as much 14CO2 as controls (P < 0.05). Because
mammalian cells respond to many toxicants by increasing the activity of the hexose monophosphatepathway, the apparent reduction in glucose oxidationcaused by TBHP may have reflected altered utilizationof the substrate rather than impaired glucose oxidation.Therefore, we next compared 6-14C-glucose oxidationby control and TBHP-treated cultures (Fig. 3), becausethe production of 14CO2 from this compound is due
primarily to glycolytic activity (Katz and Wood 1960).Overnight incubation of cultures in medium containing BC, LUT and a-TC completely protected cellsagainst the TBHP-mediated reduction in 6-14C-glucoseoxidation. Because TBHP decreased 3H-AIB uptake (see
above), it was necessary to consider whether the apparent decrease in 14C-glucose oxidation in TBHP-treated
cells might reflect impaired transport of glucose acrossthe plasma membrane, i.e., decreased cellular acquisition of the radiolabel. Similar to results for the oxidation of 6-14C-glucose, 3H-DOG uptake by TBHP-treated
+TBHP
6.
EB
it
l
CTL CTL a-TC BC LUT
FIGURE 2 Pretreatment with a-tocopherol (a-TC), /3-car-otene (BC)and lutein (LUT) attenuates oxidant-induced reduction of 3H-aminoisobutyric acid (3H-AIB)transport byHepG2 cells. Cultures were incubated overnight as describedin Table 1. After a 90-min exposure to tert-butylhydroperoxide (TBHP),cultures were pulsed with minimal essential medium eagle (MEM) containing 3H-AIB(18.5 kBq/well) for 3min and uptake of the radiolabel was determined. Data represent means ±SEMfrom at least three separate experimentseach performed in triplicate. Different letters above the errorbars indicate significant differences (P < 0.05) between themeans of the treatment groups.
by guest on July 21, 2011jn.nutrition.org
Dow
nloaded from
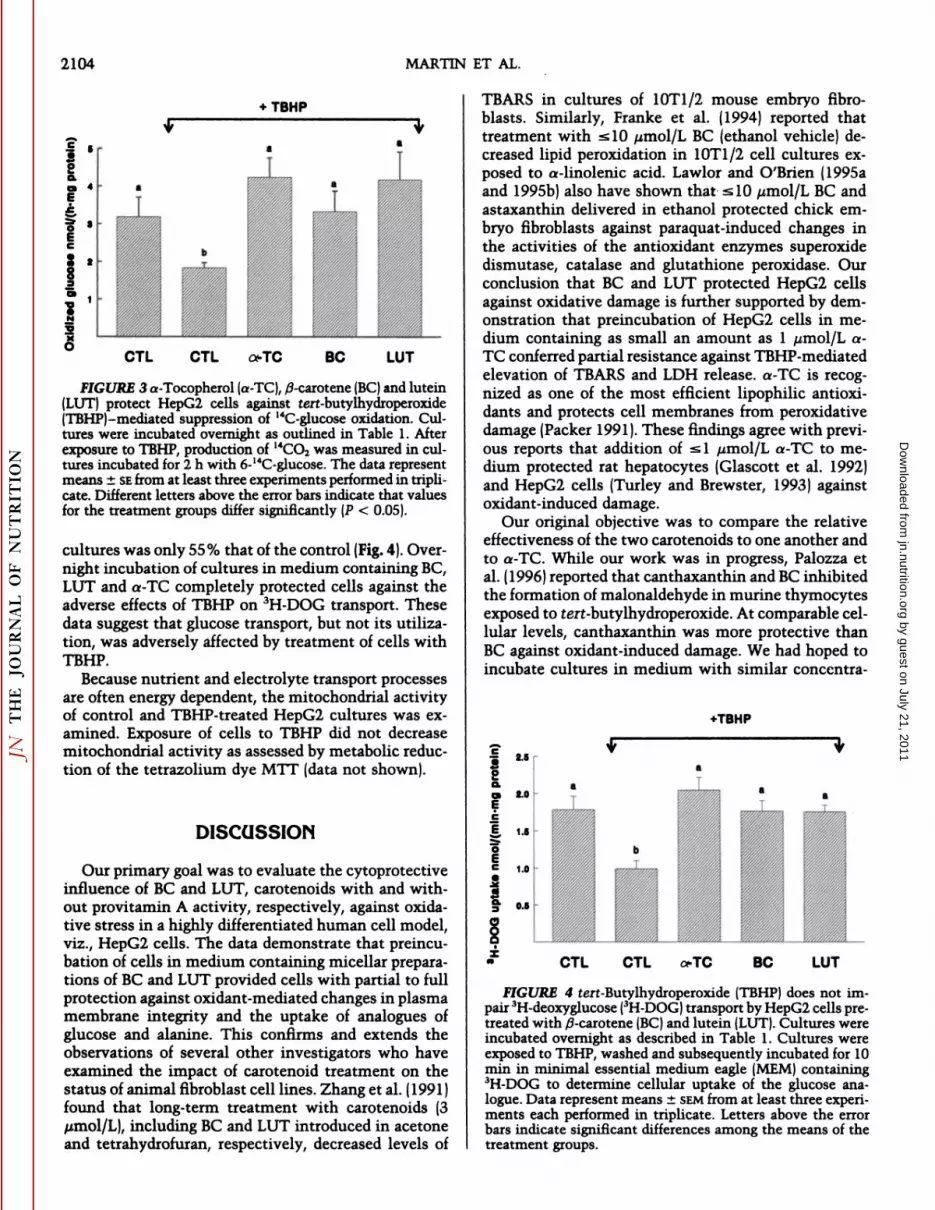
2104 MARTIN ET AL.
TBHP
•=»r
CTL CTL oTC LUT
FIGURE 3 a-Tocopherol (a-TC), /3-carotene (BC) and lutein(LUT) protect HepG2 cells against tert-butylhydroperoxide¡TEMP)-mediated suppression of 14C-glucose oxidation. Cul
tures were incubated overnight as outlined in Table 1. Afterexposure to TBHP, production of 14CO2was measured in cultures incubated for 2 h with 6-14C-glucose. The data represent
means ±SEfrom at least three experiments performed in triplicate. Different letters above the error bars indicate that valuesfor the treatment groups differ significantly (P < 0.05).
cultures was only 55% that of the control (Fig. 4). Overnight incubation of cultures in medium containing BC,LUT and a-TC completely protected cells against theadverse effects of TBHP on 3H-DOG transport. These
data suggest that glucose transport, but not its utilization, was adversely affected by treatment of cells withTBHP.
Because nutrient and electrolyte transport processesare often energy dependent, the mitochondrial activityof control and TBHP-treated HepG2 cultures was examined. Exposure of cells to TBHP did not decreasemitochondrial activity as assessed by metabolic reduction of the tetrazolium dye MTT (data not shown).
DISCUSSION
Our primary goal was to evaluate the cytoprotectiveinfluence of BC and LUT, carotenoids with and without provitamin A activity, respectively, against oxida-tive stress in a highly differentiated human cell model,viz., HepG2 cells. The data demonstrate that preincu-bation of cells in medium containing micellar preparations of BC and LUT provided cells with partial to fullprotection against oxidant-mediated changes in plasmamembrane integrity and the uptake of analogues ofglucose and alanine. This confirms and extends theobservations of several other investigators who haveexamined the impact of carotenoid treatment on thestatus of animal fibroblast cell lines. Zha.ng et al. (1991 )found that long-term treatment with carotenoids (3/xmol/L), including BC and LUT introduced in acetoneand tetrahydrofuran, respectively, decreased levels of
TEARS in cultures of 10T1/2 mouse embryo fibro-blasts. Similarly, Franke et al. (1994) reported thattreatment with <10 ¿¿mol/LBC (ethanol vehicle) decreased lipid peroxidation in 10T1/2 cell cultures exposed to a-linolenic acid. Lawlor and O'Brien (1995a
and 1995b) also have shown that s 10 ¿¿mol/LBC andastaxanthin delivered in ethanol protected chick embryo fibroblasts against paraquat-induced changes inthe activities of the antioxidant enzymes Superoxidedismutase, catalase and glutathione peroxidase. Ourconclusion that BC and LUT protected HepG2 cellsagainst oxidative damage is further supported by demonstration that preincubation of HepG2 cells in medium containing as small an amount as 1 ¿anol/La-TC conferred partial resistance against TBHP-mediatedelevation of TEARS and LDH release. a-TC is recognized as one of the most efficient lipophilic antioxi-dants and protects cell membranes from peroxidativedamage (Packer 1991). These findings agree with previous reports that addition of si /¿mol/La-TC to medium protected rat hepatocytes (Glascott et al. 1992)and HepG2 cells (Turley and Brewster, 1993) againstoxidant-induced damage.
Our original objective was to compare the relativeeffectiveness of the two carotenoids to one another andto a-TC. While our work was in progress, Palozza etal. (1996) reported that canthaxanthin and BC inhibitedthe formation of malonaldehyde in murine thymocytesexposed to terf-butylhydroperoxide. At comparable cellular levels, canthaxanthin was more protective thanBC against oxidant-induced damage. We had hoped toincubate cultures in medium with similar concentra-
t-TBHP
io.a
è
iee
||
ifi
2.0
1.0
CTL CTL a-TC BC LUT
FIGURE 4 tert-Butylhydroperoxide (TBHP) does not impair ''H-deoxyglucose (3H-DOG) transport by HepG2 cells pre-treated with /3-carotene (BC) and lutein (LUT). Cultures wereincubated overnight as described in Table 1. Cultures wereexposed to TBHP, washed and subsequently incubated for 10min in minimal essential medium eagle (MEM) containing•*H-DOGto determine cellular uptake of the glucose ana
logue. Data represent means ±SEMfrom at least three experiments each performed in triplicate. Letters above the errorbars indicate significant differences among the means of thetreatment groups.
by guest on July 21, 2011jn.nutrition.org
Dow
nloaded from

CAROTENOIDS AS ANTIOXIDANTS IN HUMAN LIVER CELLS 2105
tions of carotenoids and a-TC, i.e., 15 /¿mol/L.However, HPLC analysis of the concentration of the testcompounds in filtered medium revealed that BC waspoorly solubilized by micelles. As shown in Table 1,the final concentration of BC in filtered medium wasonly 1.1 ¿¿mol/L,whereas the concentrations of LUTand a-TC were 10- and 14-fold higher, respectively.The cellular concentration of BC was about 10% thatof LUT, suggesting that accumulation of these carotenoids was proportional to their levels in medium.Canfield et al. (1990) reported that the efficiency of BCincorporation into micelles was maximum at 15 yumol/L, and that the relative amount of BC incorporated intomicelles varied between 4 and 13% of the initial concentration. Similarly, Levin and Mokady (1995) reported that the maximum solubility of BC in micelleswas 14 //mol/L and that the isomer composition of BCmixtures affected the total amount of BC incorporated.It is likely that the low solubility of BC in our systemwas due in part to the need to decrease the concentrations of the components used for the preparation ofmicelles to prevent cytotoxicity. Also, taurocholatewas the only bile acid in our preparation, whereas theformulation used by Canfield et al. (1990) containedtaurocholate and five additional bile salts. Exclusion ofthe other bile acids may have affected the efficiency ofBC incorporation, because the types and relative quantities of lipids will influence the structure and size ofmicelles (El-Gorab and Underwood 1973). Additionally, we used a relatively short time for solubilizationof these compounds into micellar preparations and filter sterilized (0.22-//m pore) the material before use,thereby removing microcrystalline particles.
Both BC and LUT attenuated, but did not prevent,TBHP-induced increases in TEARS and LDH release.Pretreatment with a-TC prevented the enhanced leakage of cytoplasmic LDH and significantly reduced (P< 0.05) the rise in TEARS after exposure to TBHP.However, various factors preclude direct comparisonof the antioxidant potency of the carotenoids and a-TC in our model. First, as discussed above, the finalconcentrations of BC, LUT and a-TC in medium andcells differed. Second, cellular concentration by itselfdoes not provide insights about subcellular distributionand spatial orientation of the carotenoid within membranes (Britton 1995). Third, we did not consider thetypes or concentrations of carotenoid metabolites present in the cells. Khachik et al. (1995) have discussed thepotential role of carotenoid metabolites as mediators ofthe biological properties that are normally associatedwith their parent compounds. Finally, sites of toxicant-induced damage are likely to be influenced by both thechemical properties of the free radical produced andthe phenotypic characteristics of the cell. Therefore,specific conditions will influence the effectiveness ofantioxidants as cytoprotective agents against free radical-induced damage.
One proposed mechanism of damage by the prooxi-
dant TBHP is the initiation and propagation of lipidperoxidation of polyunsaturated fatty acids in theplasma membrane. However, TBHP or its metabolitesmay also interact with and damage various intracellu-lar processes (Aw and Rhoads 1994). Olson (1994) hasargued that evaluation of regulated cellular activitiesprovides greater sensitivity for detecting oxidative insult than simply monitoring the loss of barrier integrityor lethality. Therefore, we monitored the impact ofTBHP exposure on the ability of cultures to incorporate3H-leucine into protein. Exposure to the prooxidant de
creased cellular accumulation of the amino acid, butdid not alter its relative incorporation into protein.This suggested that TBHP-induced damage was localized to the plasma membrane in our model. In supportof this possibility, 3H-AIB uptake was suppressed by
84% after TBHP treatment. It is noteworthy that theuptake of 3H-leucine and 3H-AIB is mediated by differ
ent amino acid transporters, viz., systems L and A, respectively (Goenner et al. 1992), suggesting that TBHPadversely affected amino acid transporters. TBHP alsohas been reported to alter the transport of ions in several cell types. Srivastava et al. (1989) found that TBHPtreatment increased sodium entry into myocytes byaltering Na~ channels through increased lipid peroxida
tion. Elliott and Koliwad (1995) found that exposure ofendothelial cells to TBHP markedly inhibited agonist-stimulated uptake of Ca2+ across the plasma mem
brane, and that this impairment appeared to be due toan alteration in the membrane transport protein ratherthan to lipid peroxidation. TBHP-mediated reductionin 3H-AIB transport may result from altered membrane
fluidity, conformational changes of the transporter dueto peroxidation of membrane lipids in the local environment, or oxidative modification of the transporterprotein itself. Because BC, LUT and a-TC all significantly (P < 0.05) attenuated the impact of TBHP exposure on 3H-AIB uptake by HepG2 cells, it appears that
these compounds or their metabolites conferred partialor complete protection against oxidant-induced alterations in the plasma membrane. This protective influence was not limited to amino acid transporters, because pretreatment of the cultures with the carotenoidsalso prevented TBHP-induced impairment of 3H-DOG
transport.Transport activities are generally energy-dependent
processes requiring ATP for function of the Na+/K+
pump, maintenance of electrochemical gradients andproper function of the transporters. Exposure of HepG2cells to TBHP did not affect mitochondria as assessedby the ability of cells to reduce MTT. Similarly, exposure to eumene hydroperoxide did not adversely affectmitochondrial function in neural hybridoma cells(Vroegop et al. 1995). Thus, the impaired transport of3H-AIB and 3H-DOG by TBHP-treated HepG2 cells ap
pears to be due to direct damage of the cell surfacerather than a limitation in energy required to maintainnutrient transport.
by guest on July 21, 2011jn.nutrition.org
Dow
nloaded from

2106 MARTIN ET AL.
In summary, both BC and LUT partially or completely protected HepG2 human liver cells against freeradical-mediated damage as assessed by a panel of biochemical and functional indicators. The protective influence of LUT indicates that the carotenoids do nothave to be converted to retinoids for occurrence of thisphenomenon. It is unknown if the parent compoundsand/or their metabolites are the species that act as anti-oxidants within cells. The implications of these findings are that increased consumption of carotenoid-richfruits and vegetables may contribute to the protectionof human cells against the adverse effects of free radicals on cellular integrity.
ACKNOWLEDGMENTS
We thank Virginia Morris for technical assistancewith the initial HPLC analysis of a-tocopherol, GeorgeLoo for his helpful comments and Vivian Bullard forassistance in preparing the manuscript.
LITERATURE CITED
Aw, T. Y. & Rhoads, C. A. (1994) Glucose regulation of hydroper-oxide metabolism in rat intestinal cells: stimulation of reducednicotinamide adenine dinucleotide phosphate supply. J. Clin. Invest. 94: 2426-2434.
Bertram, J. S. and Bortkiewicz, H. J1995] Dietary carotenoids inhibit neoplastic transformation and modulate gene expression inmouse and human cells. Am. J. Clin. Nutr. 62: 1327S-1336S.
Bieri, J. G., Brown, E. D. & Smith, J. C., Jr. (1985) Determinationof individual carotenoids in human plasma by high performanceliquid chromatography. J. Liq. Chromatogr. 8: 473-484.
Bieri, J. G., Tolliver, T. J. & Catignani, G. L. (1979) Simultaneousdetermination of alpha-tocopherol and retinol in plasma or redcells by high pressure liquid chromatography. Am. J. Clin. Nutr.32: 2143-2146.
Britton, G. (1995) Structure and properties of carotenoids in relation to function. FASEB]. 9: 1551-1558.
Buege, J. A. & Aust, S. D. (1978) Microsomal lipid peroxidation.Methods Enzymol. 52: 302-310.
Canfield, L. M., Forage, J. W. Sa Valenzuela, J. G. (1992) Carotenoids as cellular antioxidants. Proc. Soc. Exp. Biol. Med. 200:260-265.
Canfleld, L. M., Fritz, T. A. & Tarara, T. E. (1990) Incorporationof beta carotene into mixed micelles. In: Methods in Enzymology(Packer, L., ed.), vol. 214, pp. 55-74. Academic Press, New York,NY.
El-Gorab, M. & Underwood, B.A. (1973) Solubilization of /î-caro-tene and retinol into aqueous solution of mixed micelles. Bio-chim. Biophys. Acta 306: 58-66.
Elliott, S. J. &Koliwad, S. K. (1995) Oxidant stress and endothelialmembrane transport. Free Radical Biol. & Med. 19: 649-658.
Ellwood, K. C., Chatzidakis, C. & Failla, M. L. (1993) Fructoseutilization by the human intestinal epithelial cell line Caco-2.Proc. Soc. Exp. Bio. Med. 202: 440-446.
Franke, A. A., Harwood, P. J., Shimamoto, T., Lumeng, S., Zhang,L.-X., Bertram, J. S., Wilkens, L. R., Marchand, L. L. &.Cooney,R. V. (1994) Effects of micronutrients and antioxidants on lipidperoxidation in human plasma and in cell culture. Cancer Lett.79: 17-26.
Garewal, H. (1995) Antioxidants in oral cancer prevention. Am.I. Clin. Nutr. 62: 1410S-1416S.
Glascott, P. A., Gilfor, E. a Farber, J. L. (1992) Effects of vitamin Eon the killing of cultured hepatocytes by tert-butylhydroperoxide.Mol. Pharmacol. 41: 1155-1162.
Goenner, S., Boutron, A., Soni, T., Lemonnier, A. & Moatti, N.(1992) Amino acid transport systems in the human hepatomacell line HepG2. Biochem. Biophys. Res. Commun. 189:472-479.
Javitt, N. B. (1990) HepG2 cells as a resource for metabolic studies: lipoprotein, cholesterol and bile acids. FASEBJ. 4: 161-168.
Katz, J. &Wood, H. G. (1960) The use of glucose-14Cfor the evalu
ation of the pathways of glucose metabolism. J. Biol. Chem. 235:2165-2177.
Keaney, J. F., Jr. & Frei, B. (1994) Antioxidant protection of low-density lipoprotein and its role in the protection of atheroscleroticvascular disease. In: Natural Antioxidants in Human Health andDisease (Frei,B.,éd.),pp. 303-351. Academic Press,New York,NY.
Khachik, F., Beecher, G. R. & Smith, J. C., Jr. (1995) Lutein, lyco-pene and their oxidative metabolites in chemoprevention of cancer. I. Cell. Biochem. 22: 236-246.
Lawlor, S. M. & O'Brien, N. M. (1995a) Astaxanthin: antioxidanteffects in chicken embryo fibroblasts. Nutr. Res. 15: 1695-1704.
Lawlor, S. M. & O'Brien, N. M. (1995b) Modulation of oxidativestress by ^-carotene in chicken embryo flbroblasts. Br. J. Nutr.73: 841-857.
Levin, G. & Mokady, S. (1995) Incorporation of all-trans- or 9-cis-j8-carotene into mixed micelles. Lipids 30: 177-179.
Meydani,S.N.,Wu,D., Santos, M.S.&.Hayek, M.G. (1995) Antioxidant and immune response in aged persons: overview of presentevidence. Am. J. Clin. Nutr. 62: 1462S-1476S.
Moshage,H. & Yap,S. H. (1992) Primary cultures of human hepatocytes: a unique system for studies in toxicology,virology,parasitol-ogy and liver pathophysiology in man. J. Hepatol. 15:404-413.
Olson, M. J. (1994) Evaluation of glucose metabolism in vitro:altered competence of intermediate metabolism as an index ofhepatotoxicity. Methods Toxicol. IB: 152-163.
Packer, L. (1991) Protective role of vitamin E in biological systems. Am. J. Clin. Nutr. 62: 1050S-1055S.
Palozza, P., Luberto, C., Ricci, P., Sgarlata, E., Calviello, G. &.Bartoli,G. M. (1996) Effect of /3-carotene and canthaxanthin on ten-butylhydroperoxide-induced lipid peroxidation in murine normaland tumor thymocytes. Arch. Biochem. Biophys. 325: 145-151.
Scudiero, D. A., Shoemaker, R. H., Paul, K. D., Monks, A., Tierney,S., Nofziger, T. H., Currens, M. J.,Seniff,D. & Boyd,M. R. (1988)Evaluation of a soluble tetrazolium/formazan assay for cellgrowth and drug sensitivity in culture using human and othertumor cell lines. Cancer Res. 48: 4827-4833.
Sies, H. & Stahl, W. (1995) Vitamins E and C, /3-carotene, andother carotenoids as antioxidants. Am. J. Clin. Nutr. 62: 1322S-1326S.
Snedecor, G. W. & Cochrane, W. G. (1989) Statistical Methods,8th ed. Iowa State University, Ames, IA.
Srivastava, S. K., Ansari, N. H., Liu, S., Izban, A., Das, B. & Szabo,G. (1989) The effect of oxidants on biomembranes and cellularmetabolism. Mol. Cell. Biochem. 91: 149-157.
Thurnham, D. I. (1994) Carotenoids: function and fallacies. Proc.Nutr. Soc. 53: 77-87.
Turley, C. P. & Brewster, M. A. (1993) a-Tocopherol protectsagainst a reduction of adenosylcobalamin in oxidatively stressedhuman cells. J. Nutr. 123: 1305-1312.
Van Poppel, G. & Goldbohn, R. A. (1995) Epidemiologie evidencefor /3-carotene and cancer prevention. Am. J. Clin. Nutr. 62:1393S-1402S.
Vroegop, S. M., Decker, D. E. & Buxser, S. E. (1995) Localizationof damage induced by reactive oxygen species in cultured cells.Free Radical Biol. & Med. 18: 141-151.
Zhang, L.-X., Cooney, R. V. & Bertram, }. S. (1991) Carotenoidsenhance gap junctional communication and inhibit lipid peroxidation in C3H/10T1/2 cells: relationship to their cancer chemo-preventive action. Carcinogenesis 12: 2109-2114.
by guest on July 21, 2011jn.nutrition.org
Dow
nloaded from





























