Embed Size (px)
Citation preview

Abstract This study describes time course and ultra-structural changes during axonal degeneration of differ-ent neurones within the tympanal nerve of the locustSchistocerca gregaria. The tympanal nerve innervatesthe tergit and pleurit of the first abdominal segment andcontains the axons of both sensory and motor neurones.The majority of axons (approx. 97%) belong to severaltypes of sensory neurones: mechano- and chemosensitivehair sensilla, multipolar neurones, campaniform sensillaand sensory cells of a scolopidial organ, the auditory or-gan. Axons of campaniform sensilla, of auditory sensorycells and of motor neurones are wrapped by glial cellprocesses. In contrast, the very small and numerous ax-ons (diameter <1 µm) of multipolar neurones and hairsensilla are not separated individually by glia sheets.Distal parts of sensory and motor axons show differentreactions to axotomy: 1 week after separation from theirsomata, distal parts of motor axons are invaded by glialcell processes. This results in fascicles of small axonbundles. In contrast, distal parts of most sensory axonsdegenerate rapidly after being lesioned. The time to on-set of degeneration depends on distance from the lesionsite and on the type of sensory neurone. In axons of au-ditory sensory neurones, ultrastructural signs of degener-ation can be found as soon as 2 days after lesion. Aftercomplete lysis of distal parts of axons, glial cell process-es invade the space formerly occupied by sensory axons.The rapid degeneration of distal auditory axon parts al-lows it to be excluded that they provide a structure thatleads regenerating axons to their targets. Proximal partsof severed axons do not degenerate.
Key words Ear · Sensory cells · Motor neurones · Glia ·Insect · Regeneration · Schistocerca gregaria (Insecta)
Introduction
After lesion many axons undergo degenerative changesin their distal parts. This process has been used to revealthe axonal composition of peripheral nerves and the cen-tral projection areas of sensory neurones (Schürmann1980; Distler and Boeckh 1997). However, in inverte-brates a long-term survival of distal parts of the lesionedaxon has also been reported (Boulton 1969; Bittner andMann 1976; Ballinger and Bittner 1980; Bittner 1991).This long-term survival has been proposed to be impor-tant for guiding the regenerating proximal axon; the out-growing axons may orientate along the remaining distalaxon parts or fuse with them (Hoy et al. 1967; Birse andBittner 1981; Bittner 1991; Lyckman et al. 1992; vonBernhardi and Muller 1995).
Axons of auditory receptor cells of grasshoppers havethe capacity to regrow into their target area after experi-mental lesion (Lakes and Kalmring 1991; Lakes-Harlanand Pfahlert 1995; Jacobs 1997). Here, they form func-tional synapses onto auditory interneurones. To date,nothing is known about the fate of distal axon parts ofauditory neurones. Do they degenerate rapidly after le-sion, or do they persist until the proximal axons regener-ate, and so perhaps provide a guidance structure forthem? Can the possibility of a fusion of axonal processesduring regeneration be completely ruled out? The an-swers to these questions are important for our under-standing of pathfinding and target recognition of regen-erating neurones. Here, we describe the ultrastructuralchanges of auditory sensory axons of the locust Schis-tocerca gregaria after their separation from the somata.
Before entering the ganglia of the ventral nerve cord,the axons of auditory sensory cells run within the audito-ry nerve. This nerve innervates the tergit and pleurit ofthe first abdominal segment, including the ear. Next tothe axons of auditory cells, it contains motor axons and
K. Jacobs · R. Lakes-Harlan (✉)Georg-August-Universität Göttingen, Abteilung Neurobiologie,Institut für Zoologie und Anthropologie, Berliner Str. 28, D-37073 Göttingen, Germanye-mail: [email protected]: +49 551 395409Fax: +49 551 395438
Present address: K. Jacobs, Sussex Centre for Neuroscience, University of Sussex,Brighton BN1 9QE, UK
Cell Tissue Res (1999) 298:167–178 © Springer-Verlag 1999Digital Object Identifier (DOI) 10.1007/s004419900056
R E G U L A R A R T I C L E
Kirsten Jacobs · Reinhard Lakes-Harlan
Axonal degeneration within the tympanal nerve of Schistocerca gregaria
Received: 4 February 1999 / Accepted: 4 May 1999 / Published online: 1 July 1999

axons of different other types of sensory neurones,which we identified by performing different experimen-tal types of surgery. An analysis of ultrastructural chang-es within these sensory and motor axons revealed majordifferences in their reaction following lesion. As experi-mental conditions were exactly the same for all neuro-nes, this must be explained by intrinsic properties of thedifferent neurone types.
Materials and methods
Animals and surgery
Adult Schistocerca gregaria locusts were obtained from a labora-tory colony reared under crowded conditions in our institute. Inorder to identify different types of axons and to investigate chang-es in ultrastructure during degeneration of sensory fibres, we per-formed three different types of surgery (Fig. 1): cutting the tympa-nal nerve about 500 µm peripheral from the metathoracic ganglionto investigate degeneration of distal parts (=parts which are nolonger connected to the cell bodies) of all sensory axons (axotomy1), extirpation of the auditory organ to induce degeneration of au-ditory axons and that of other sensory cells at the tympanal mem-brane (axotomy 2); and removal of the tympanal membrane with-out destroying the auditory organ to cause degeneration of distalaxonal parts of non-auditory sensory cells at the tympanal mem-brane, leaving the auditory axons intact (axotomy 3). For the firsttype of surgery, adult animals, aged up to 5 days after the imaginalmoult, were cooled at 4°C and fixed ventral side up on a Plasticineplatform. The sternum was opened by cutting the cuticle with a ra-zor blade, and nerve 6 of the metathoracic ganglion was cut (Fig.1; axotomy 1). After the surgery, the cuticle was replaced andfixed with wax to avoid loss of haemolymph. The animals werekept at 25°C and killed after various postoperative periods (1–25days). For the second and third type of surgery the animals werefixed lying on one side. The wings were pinned up to expose theear. Thereafter, either the tympanal membrane was opened withfine forceps and the auditory organ was excised (axotomy 2) orthe tympanal membrane was removed, leaving the auditory organinside the animal (axotomy 3). The wound was covered with pe-troleum jelly and the animals were killed after the same postopera-tive periods as animals with type 1 surgery.
Light microscopy
Axonal morphology and the types of sensory neurones at the earwere revealed by axonal fillings of the tympanal nerve with neuro-biotin. The nerve was cut about 500 µm peripheral to the ganglionand was marked centrifugally by immersing the cut end in a neu-robiotin solution (5% neurobiotin in 1 M potassium acetate) for 12 h at 4°C. Afterwards, the ear was removed from the animal andfixed for 1 h in 4% paraformaldehyde in phosphate buffer (PB).After several rinses in PB containing 1% Triton-X 100, the prepa-rations were incubated overnight in a solution of avidin with bio-tinylated horseradish peroxidase (Vector Elite Kit) at room tem-perature. The preparations were washed in PB several times andincubated in diaminobenzidine and nickel chloride (Vector Kit) for30 min at room temperature before transferring the preparations toa solution of H2O2 in PB. When labelled neurones became visible,the reaction was stopped with PB. The preparations were dehy-drated, cleared in methyl salicylate and viewed using a LeicaDMRB microscope.
Centrally located arborizations and cell bodies of motorneuro-nes as well as sensory projections were analysed by axonal fillingsof the tympanal nerve connected with the metathoracic ganglion(“central part of the nerve” in experimental animals). The tympan-ic nerve was placed into a glass capillary filled with 5% NiCl2 so-lution. The preparation was transferred to a moist chamber and
stored at 20°C for 12 h. The nickel was precipitated with freshlyprepared 1% ammonium sulphide before fixation with Bouin’s fix-ative (30 min at room temperature). The preparations were silverintensified according to Bacon and Altman (1977). Preparationswere dehydrated through a series of ethanols and cleared in meth-ylsalicylate.
Electron microscopy
The metathoracic ganglion including the central stump of the tym-panal nerve was dissected from animals at a selected time aftersurgery. In cut preparations (axotomy 1) the peripheral region ofthe nerve was taken out of the animal as well. All preparationswere immediately fixed in 2.5% glutaraldehyde in freshly pre-pared 0.1 M phosphate-buffered saline (PBS; pH 7.4) for a periodof 2.5 h. Next the tissue was washed in 0.1 M PBS containing 5%sucrose for 15 min with several changes. Postfixation followed fora further hour in 2% osmium tetroxide dissolved in the buffer so-lution. The tissue was then again washed in PBS and dehydratedin a graded ethanol series. The 70% ethanol step was performedfor at least 30 min using a saturated solution of uranyl acetate in70% ethanol. Preparations were incubated twice for 10 min in pro-pylene oxide and embedded in Araldite. Thin (60–80 nm) sectionsof peripheral and central nerve regions (Fig. 1; EM1, EM2) wereobtained using an ultramicrotome (Reichert Jung Ultracut E). Thesections were placed on 50-mesh or single-slot grids coated withFormvar and stained with lead citrate before being viewed withthe electron microscope (Zeiss EM10). The entire cross section of
168
Fig. 1 Schematic drawing of the experimental preparation. Indi-cated are the sites of three different types of surgery (axotomy1–3) at the tympanal nerve (N6). Axotomy 1 divides axons of mo-tor neurones as well as all axons of peripheral sensory neuronesinto two parts, axotomy 2 and 3 only axons of different types ofsensory neurones at the ear. The first site of investigation (EM1)lies centrally to all sites of surgery (=central region of the tympa-nal nerve with distal parts of lesioned sensory axons). The secondsite of investigation lies peripherally to the nerve cut (EM2; axoto-my 1=peripheral region of tympanal nerve with distal parts of mo-tor axons) (cs campaniform sensilla, sc auditory sensory cells, tmtympanal membrane, TH3 metathoracic ganglion)

the nerve was photographed via an overlapping series of expo-sures; these were printed at a final magnification of ×2500. Countswere made of the total number of axons and the total number ofmitochondria. Statistical analysis of the changes in number of mi-tochondria was performed using Student’s t-test. Axon diameterwas calculated as the mean length of two perpendicular linesthrough the centre of the axonal profile. The percentage area thataxons and glial cells occupied within a nerve cross section was de-termined from measurements along two perpendicular linesthrough the nerve centre.
Results
Innervation area of the tympanal nerve
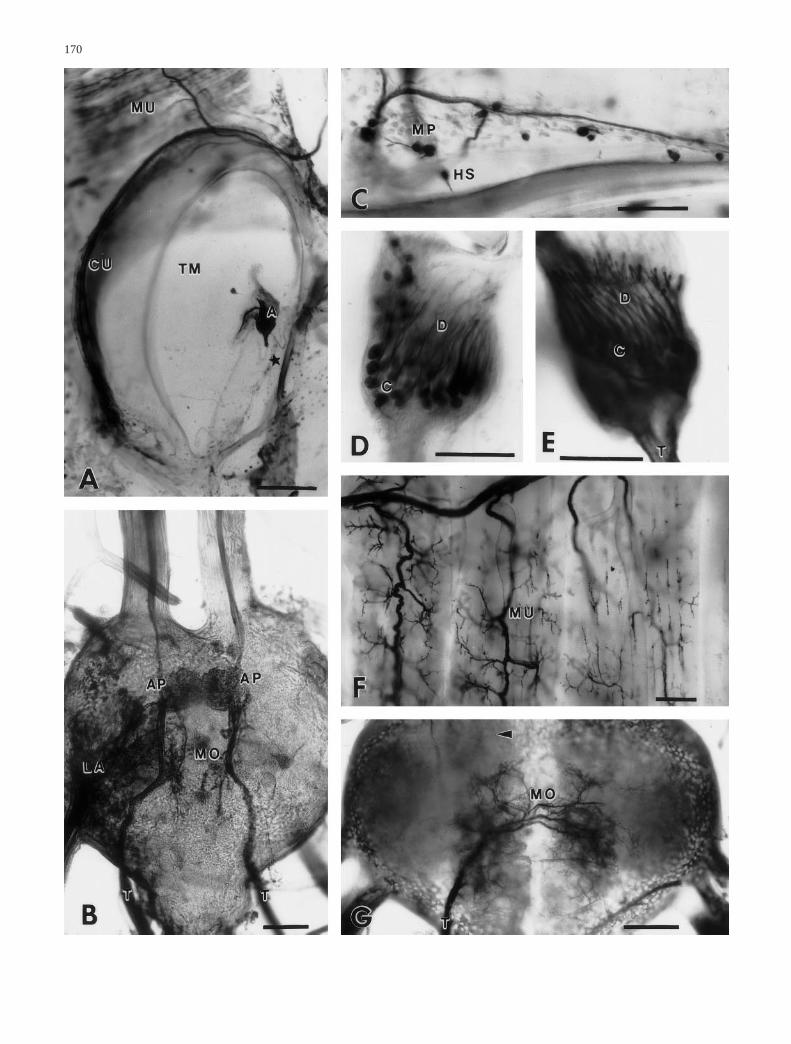
The tympanal nerve of the grasshopper Schistocercagregaria contains axons of sensory cells and of motor-neurones which innervate the tergit and pleurit of thefirst abdominal segment. Sensory cells at the tergal cuti-cle comprise mainly hair sensilla and multipolar neuro-nes. The pleurit contains the animal’s ear. The most ob-vious sensory organ of the ear is a scolopidial organ,which is attached to the tympanal membrane. This organbears the cell bodies and dendrites of about 80 auditorysensory cells (Fig. 2A,D; Gray 1960; Jacobs et al. 1999),and is therefore referred to as auditory organ. Next to it,the tympanal membrane and the cuticular rim of the earcontain a variety of smaller sensory organs, includingmultipolar neurones, campaniform sensilla and mech-ano- and chemosensory hairs (Fig. 2A,C). About 20campaniform sensilla as well as multipolar sensory cellslie directly underneath the auditory organ. Several multi-polar neurones and hair sensilla are situated at the borderof the tympanal membrane and the adjacent cuticle ofthe segment.
Ultrastructure of the tympanal nerve
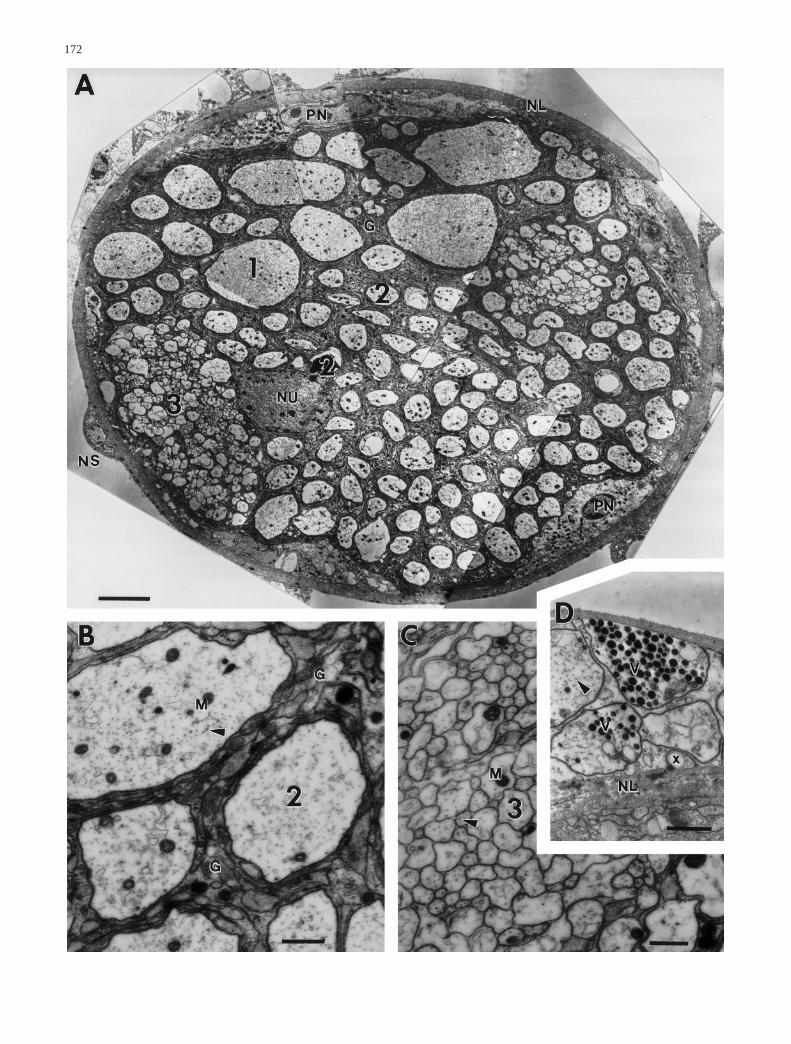
The tympanal nerve has a diameter of 50–60 µm andcontains between 835 and 865 axons of different sizes(Figs. 3A, 5; individuals differ slightly). The axons occu-py between 53% and 63% of the nerve’s area; the cellu-lar processes of glial cells, their nuclei and the extracel-lular matrix cover the remaining area (Fig. 6A). The nu-clei of perineural glial cells (one to two per nerve sec-tion) are located at the periphery of the tympanal nerve,whereas the nuclei of subperineural glia lie medially be-tween the axons (one nucleus per nerve section). A bun-dle of presumptive neurosecretory axons is attached atthe neurolemma of each tympanal nerve (Fig. 3D).
Within the nerve, three different types of axons can bedistinguished according to their morphology and size:The first type includes 4–6 large axons with a diameterof 8–12 µm; the second comprises between 123 and 143axons with a diameter between 1.5 and 6 µm; and thethird consists of 707–716 axons with a diameter smallerthan 1 µm (Figs. 3A, 5). The first two types of axons areindividually separated by subperineural glial cell pro-cesses which form layers around each axon (Fig. 3B).
Type 3 axons mostly occur in groups, forming one tothree fascicles within the tympanal nerve (Fig. 3C). Thenumber of axons of all types shows minor variations be-tween different individuals (Fig. 5).
The axoplasm contains neurotubules running parallelto the axon axis, randomly distributed mitochondria anda few vacuoles (Fig. 3B). No vesicles have been found,except in the presumptive neurosecretory axons attachedto the nerve (Fig. 3D). The number of mitochondria cor-relates with the type of axon: type 1 axons have a meanof 28±7 (n=12, N=2; n=number of axons, N=number ofpreparations) mitochondria, type 2 axons 5.7±3.4(n=264, N=2) mitochondria and type 3 axons approxi-mately one mitochondria per 25 axon profiles (Fig. 6B).In contrast to that of axons, the cytoplasm of glial cellsappears more electron-dense and contains mitochondria,microtubules and small vesicles, ribosomes, rough endo-plasmic reticulum and Golgi apparatus. The latter arefound predominantly next to the nuclei.
Correlation between neurone type and axon morphology
In order to correlate the sensory neurones at the abdomi-nal segment to axon types within the tympanal nerve, weperformed three different types of surgery (Fig. 1; see“Materials and methods”). All of these types of surgeryeventually lead to massive degeneration of distal parts ofsensory axons (see below for time course and ultrastruc-tural changes), but not necessarily to that of distal partsof motor axons.
Type 1 axons
In the central region of the tympanal nerve (Fig. 1; EM1)the number of type 1 axons is not affected by any type ofsurgery. Twenty-five days postcut, four to six type 1 ax-ons can still be found. In the region of the nerve (Fig. 1;EM2), these axons disappear and are replaced by bun-dles of smaller axons (Fig. 4C,D). The type 1 axons willtherefore be ascribed as motor axons (see “Discussion”).
Type 2 axons
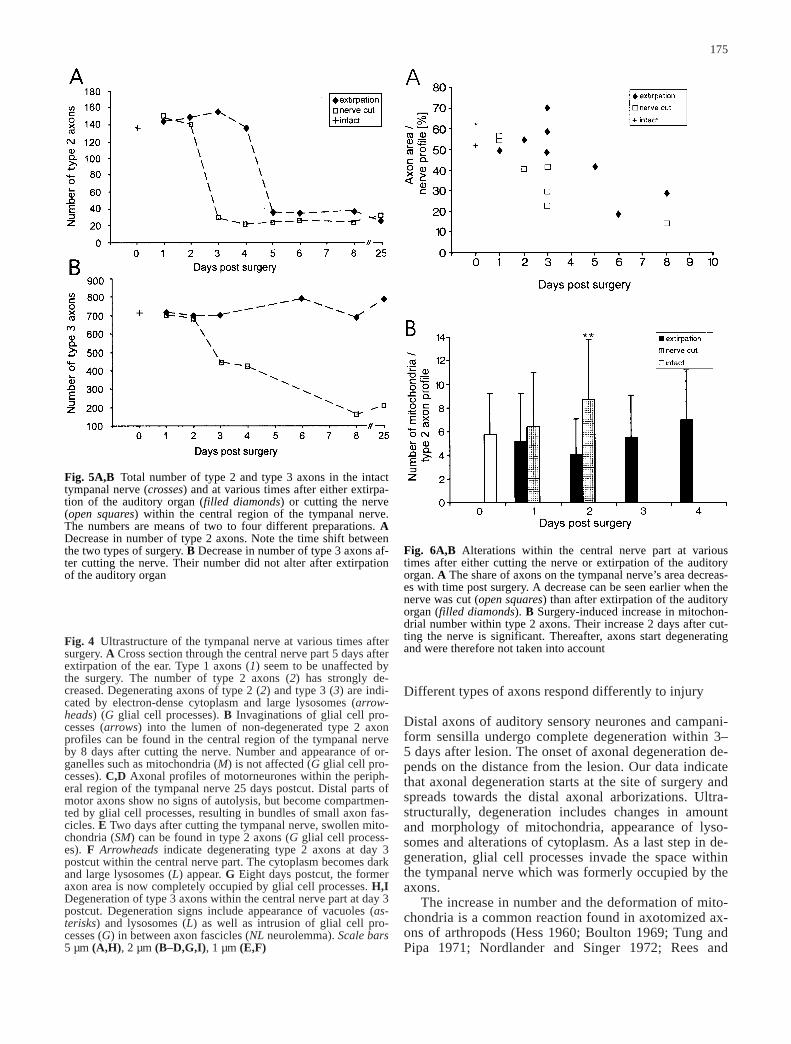
After cutting the tympanal nerve or after extirpation ofthe auditory organ, 26–32 axons can still be found in thecentral region of the tympanal nerve (Fig. 5A). Thismeans that 110–120 axons from the original approxi-mately 140 profiles have disappeared. After removal ofthe tympanal membrane, 118 axons remain intact, andabout 20 disappear. In the peripheral region of the nerveabout 100–110 axons remain intact after cutting. Wetherefore conclude that 26–32 axons belong to motor-neurones, and furthermore that the remaining 100–110axons of this type originate at the ear, comprising pre-sumably about 20 axons of campaniform sensilla andabout 80 axons of auditory receptor cells.
169
170

Type 3 axons
Neither extirpation of the auditory organ nor removalof the tympanal membrane has any significant effect onthe number of type 3 axons. However, cutting the tym-panal nerve results in a strong decrease in the numberof type 3 axons within the central region of the tympa-nal nerve; 25 days postsurgery only 204 axons re-mained (Fig. 5B). Within the peripheral region of thenerve the number in relation to the intact fasciclescould not be determined as the distal axon parts of mo-torneurones are replaced by bundles of smaller axons(Fig. 4C,D). In summary we conclude that type 3 axonsprobably belong mainly to hair sensilla and multipolarsensilla, located at the tergit and pleurit of the first ab-dominal segment.
Morphological and ultrastructural changes after lesion
Axonal marking of the lesioned nerve reveals light mi-croscopically that the central projections of sensory neu-rones disappear within a few days after lesion. Prepara-tions on the 6th day after lesion show normal arboriza-tions of dendrites of motorneurones, but none of sensorycells (Fig. 2A,G). These preparations also show 15–26(n=8) marked cell bodies of motorneurones. Similarmarkings into the periphery show that sensory cell bod-ies are also not degenerated after lesion (Fig. 2E). How-ever, in contrast to the sensory axons, distal parts of mo-tor axons can still be labelled on the 6th day after lesion(Fig. 2F).
Lesion-induced ultrastructural changes were analysedsystematically in the central region of the nerve (thenerve connected to the ganglion), as we were mainly in-terested in degeneration of auditory fibres and the rele-vance of this to their subsequent regeneration. Neverthe-less, for a comparison of ultrastructural changes withinaxons of motorneurones and sensory neurones, we alsoanalysed the peripheral nerve part at 3, 8 and 25 dayspostcut.
The diameter of the central nerve part does notchange after lesion: it ranges from 48 µm to 60 µm untilthe 25th day after surgery. A closer look reveals, howev-er, dramatic changes in the relation between axons andglial cell processes (Fig. 6A): the proportion of axons inthe central nerve area decreases to about 13% by day 8post surgery. By then, glial cell processes and extracellu-lar matrix covers the remaining area (Figs. 4G, 6A). Theinvading glial cell processes show no obvious differ-ences in organelle composition from those in intact ani-mals. The number of nuclei from perineural and subperi-neural glial cells does not differ from that in intact tym-panal nerves, indicating growth of glial cell processesrather than proliferation of the cells themselves. Regard-less of the time after the surgery, one to two nuclei fromperineural glia and one nucleus from subperineural gliaare present per axon profile.
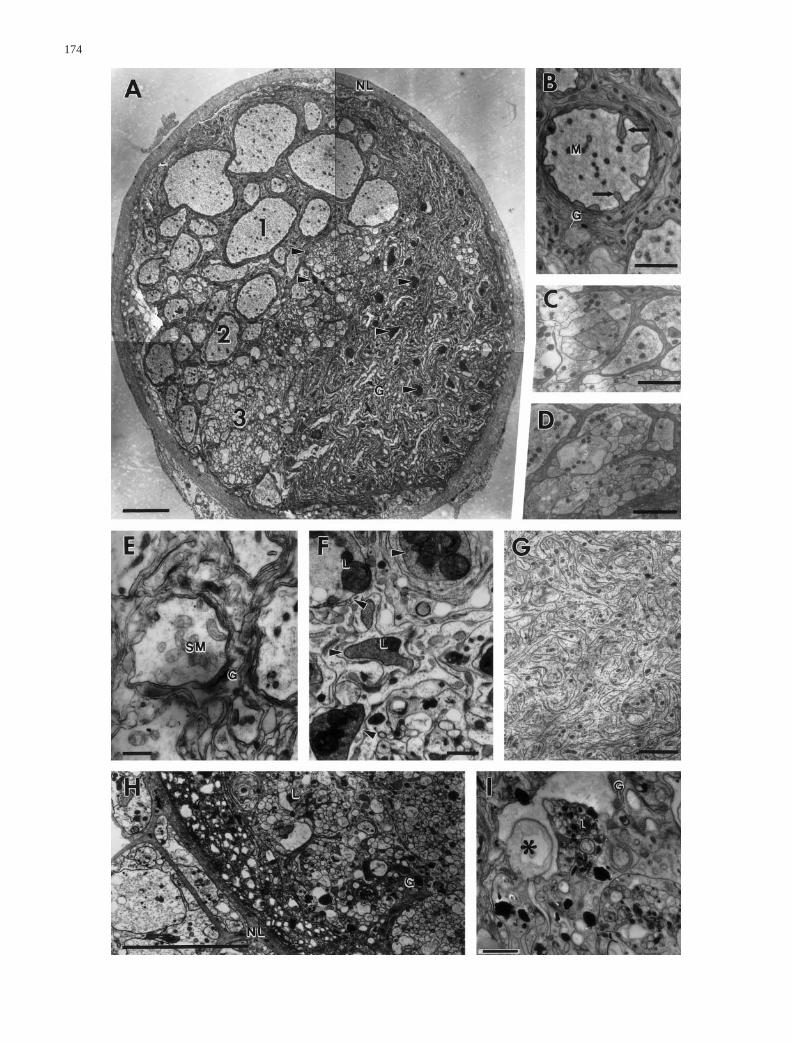
Alterations in ultrastructure of axons after surgery de-pend on the type of axon profile. Type 1 axons show nochanges in the type or number of organelles at any timeafter surgery. However, starting at the 3rd day postcut,glial cell processes invading the axons can be observed(Fig. 4A). The number and length of these glial cell pro-cesses seem to increase with time postcut, but they neverbecome predominant. By contrast, within the peripheralregion of the nerve, type 1 axons show remarkablechanges. The axonal profiles are invaded deeply by glialcell processes, which results in division of the large ax-ons into smaller profiles of axon-like structure (Fig. 4C).Thereafter, the former site of the type 1 axons within thenerve is occupied by bundles of small profiles (Fig. 4D).These bundles are present in the nerve at least until 25 days postcut without showing signs of degenerationsuch as the appearance of lysosomes.
The appearance of type 2 axons does not change dur-ing the 1st day after cutting the tympanal nerve. By day2 postcut the number of mitochondria per axon profileincreases significantly (P<0.001) to 8.7±5 (n=289, N=2,Fig. 6B) within the central region of the tympanal nerve.In some axon profiles the mitochondria appear some-what deformed and swollen (Fig. 4E). Three days aftersurgery the axoplasm becomes quite electron dense andalmost indistinguishable from that of glial cells. In mostaxon profiles, however, the cytoplasm is completely re-placed by lysosomes and disintegrating mitochondria bythis time (Fig. 4A,F). In the peripheral region of the tym-panal nerve, some type 2 axons are replaced by bundlesof axons, like those described for type 1 axons.
The earliest changes in the ultrastructure of type 3 ax-ons are visible at day 3 postcut (central region of thetympanal nerve). The ultrastructural changes include anincrease in the number of mitochondria and lysosomes,as in type 2 axons (Fig. 4H,I). Axon groups formed bytype 3 axons disintegrate due to invasion of glial cellprocesses. In contrast to type 2 axons, signs of degenera-tion in type 3 axons can still be detected at day 8 postcut,indicating that type 3 axons do not all degenerate at thesame time (Fig. 5B). In the peripheral region of thenerve no changes have been found after a cut.
171
Fig. 2 Photographs of neurobiotin- or NiCl2-marked neuronalstructures of the tympanal nerve before (A–D) and after lesion(E–G). A Overview of the ear of Schistocerca gregaria as re-vealed by centrifugal backfill. The auditory sensory cells (A) arelocated on the tympanal membrane (TM) adjacent to numeroussmall sensilla (asterisk) shown in C. B Central projection of theauditory receptor cells (AP) shown on both sides of the animal.The motoric arborizations (MO) are largely out of focus. C Hairsensilla (HS) and multipolar sensilla (MP) on the tympanal mem-brane (area marked in A). D Cell bodies (C) and dendrites of theauditory sensory cells. E Auditory sensory cells 6 days after nervelesion. The cells are not degenerated. F Arborizations of motor-neurones onto tergal muscles (MU) 6 days after lesion. No degen-eration can be detected. G Six days after lesion only motorneuronearborizations (MO) can be seen in the metathoracic ganglion. Au-ditory central arborizations cannot be marked (arrowhead), due tothe axonal degeneration (CU cuticle, LA projection of leg affer-ents, T tympanal nerve). Scales 1 mm (A), 100 µm (B,G), 200 µm(C,F)
172

Onset of degeneration of type 2 axons depends on distance to the lesion
Cutting the tympanal nerve results in an earlier degener-ation of type 2 axons in comparison to an extirpation ofthe auditory organ: In the former type of surgery, duringthe first 2 days post surgery between 131 and 153 axonswith intact morphology can still be found (Fig. 5A). Atday 3 postcut their number has decreased to a mean of28 axons, at day 8 to 24 axons. In contrast, after extirpa-tion of the auditory organ the number of intact type 2 ax-ons remains constant during the first 4 days after surgery,with a mean of 146. The decrease in axon number doesnot start until day 5, when the number drops to 35 (Fig.5A) and subsequently to 26 axons per nerve at the 25thday. This delay is also reflected by the ultrastructuralchanges within the axon. The increase in area occupiedby glial cells as well as ultrastructural changes within theaxon profiles occurs with 2 days delay; the density ofmitochondria increases at day 5 post surgery to 7±4.2(n=83, N=1; Fig. 6B). Lysosomes and dark cytoplasmare also not found until the 5th day after extirpation.
Discussion
This study describes changes in the ultrastructure of thetympanal nerve of Schistocerca gregaria following le-sion. Degeneration experiments were performed: (1) tocorrelate ultrastructurally distinguishable axons withinthe tympanal nerve to different types of sensory neuro-nes at the ear and (2) to investigate morphologicalchanges in sensory axons and motor axons that are corre-lated to degeneration.
All types of sensory neurones showed axonal degen-eration after experimental lesion, although the onset ofdegeneration depended on both kind of applied surgeryand neurone type. Ultrastructural changes during axonaldegeneration were similar in all types of sensory neuro-nes and will be discussed below. The results of this studyallow speculation about pathfinding of regenerating sen-sory fibres after lesion.
The tympanal nerve of the adult locust
The tympanal nerve contains axons of motorneuronesand of several types of sensory neurones, which inner-vate the tergit and pleurit of the first abdominal segmentincluding the ear. The sensory neurones include about700 multipolar neurones and mechano- and chemosensi-tive hair sensilla, about 20 campaniform sensilla and ascolopidial organ, which contains about 80 auditory sen-sory cells. Axon number varied slightly between differ-ent individuals. All different types of sensory neuronescan be found directly at the ears of the animals.
After cutting the tympanal nerve, 4–6 type 1 axonsand 26–32 type 2 axons remain intact in the centralnerve region, indicating that they are connected to cellbodies within the central nervous system. Thus, proba-bly all of these axons belong to motorneurones andneuromodulatory neurones (e.g. dorsal unpaired radianneurones). The number corresponds to that of neuronalcell bodies labelled by centripetal nerve backfilling, al-though the latter number is slightly lower, probably dueto incomplete marking. The identification as motorneu-rones is also supported by a dorsal location of their ax-ons within the nerve. Hence, motorneurones have thelargest diameters of all tympanal nerve axons and areindividually wrapped by glia processes. In contrast,most of the remaining approximately 800 type 2 andtype 3 axons degenerate in the central nerve part. Theyare therefore likely to have peripherally located cellbodies and thus to be axons of sensory cells. The largenumber seems somewhat surprising because the mainsensory organ of the first abdominal segment is the ear,with only about 80 auditory sensory cells and about 20campaniform sensilla. The different types of surgery re-vealed that both sensory neurone types belong to type 2axons. They are ultrastructurally indistinguishable. Fur-thermore the different physiological and morphologicaltypes of auditory cells (Römer 1976; Halex et al. 1988;Jacobs et al. 1999) do not show any ultrastructural dif-ferences. In summary, the ultrastructurally homogene-ous type 2 axons comprise axons of motorneurones,scolopidial cells and campaniform sensilla. The smallaxons (type 3) seem to belong to mechanosensory andmultineuronal chemosensory hair sensilla and multipo-lar neurones. Such sensilla occur in large numbers atthe cuticle as well as at the cuticular rim of the ear andeven at the tympanal membrane. The function of thesensilla at the tympanal membrane is so far completelyunknown. They may be involved in measuring low-fre-quency oscillations of the tympanal membrane whichare caused by respiration. These oscillations are knownto interfere with hearing and should therefore be takeninto account by the animals (Meyer and Elsner 1995).Some of these axons appear not to degenerate even af-ter 25 days. We do not know whether this reflects aprolonged survival, or whether these axons belong tounidentified cells which have their somata in the cen-tral nervous system.
173
Fig. 3A–D Ultrastructure of the intact tympanal nerve. A Crosssection through the tympanal nerve of a control animal shows ax-onal profiles of various sizes. According to their size, profileshave been classified into three types; the larger ones [type 1 (1)and type 2 axons (2), see text] occur individually surrounded byglial cell processes (G) and the smaller ones [type 3 axons (3)] infascicles. Nuclei of subperineural glia (NU) are found between ax-ons, that of perineural glia (PN) directly beneath the neurolemma(NL) (NS presumptive neurosecretory cells). B High-power viewof a representative type 2 axon (2) which contains mitochondria(M) and neurotubules (arrowheads) (G glial cell processes). CHigh-power view of type 3 axon fascicles. Axons (3) are not indi-vidually wrapped by glial cell processes. They contain mitochon-dria (M) and neurotubules (arrowhead). D Axons of presumptiveneurosecretory cells accompany each tympanal nerve. They con-tain a large number of dense core vesicles (V), mitochondria, neu-rotubules (arrowhead) and some vacuoles (x). Scale bars 5 µm(A), 0.5 µm (B,C), 1 µm (D)
174
Different types of axons respond differently to injury
Distal axons of auditory sensory neurones and campani-form sensilla undergo complete degeneration within 3–5 days after lesion. The onset of axonal degeneration de-pends on the distance from the lesion. Our data indicatethat axonal degeneration starts at the site of surgery andspreads towards the distal axonal arborizations. Ultra-structurally, degeneration includes changes in amountand morphology of mitochondria, appearance of lyso-somes and alterations of cytoplasm. As a last step in de-generation, glial cell processes invade the space withinthe tympanal nerve which was formerly occupied by theaxons.
The increase in number and the deformation of mito-chondria is a common reaction found in axotomized ax-ons of arthropods (Hess 1960; Boulton 1969; Tung andPipa 1971; Nordlander and Singer 1972; Rees and
175
Fig. 4 Ultrastructure of the tympanal nerve at various times aftersurgery. A Cross section through the central nerve part 5 days afterextirpation of the ear. Type 1 axons (1) seem to be unaffected bythe surgery. The number of type 2 axons (2) has strongly de-creased. Degenerating axons of type 2 (2) and type 3 (3) are indi-cated by electron-dense cytoplasm and large lysosomes (arrow-heads) (G glial cell processes). B Invaginations of glial cell pro-cesses (arrows) into the lumen of non-degenerated type 2 axonprofiles can be found in the central region of the tympanal nerveby 8 days after cutting the nerve. Number and appearance of or-ganelles such as mitochondria (M) is not affected (G glial cell pro-cesses). C,D Axonal profiles of motorneurones within the periph-eral region of the tympanal nerve 25 days postcut. Distal parts ofmotor axons show no signs of autolysis, but become compartmen-ted by glial cell processes, resulting in bundles of small axon fas-cicles. E Two days after cutting the tympanal nerve, swollen mito-chondria (SM) can be found in type 2 axons (G glial cell process-es). F Arrowheads indicate degenerating type 2 axons at day 3postcut within the central nerve part. The cytoplasm becomes darkand large lysosomes (L) appear. G Eight days postcut, the formeraxon area is now completely occupied by glial cell processes. H,IDegeneration of type 3 axons within the central nerve part at day 3postcut. Degeneration signs include appearance of vacuoles (as-terisks) and lysosomes (L) as well as intrusion of glial cell pro-cesses (G) in between axon fascicles (NL neurolemma). Scale bars5 µm (A,H), 2 µm (B–D,G,I), 1 µm (E,F)
Fig. 5A,B Total number of type 2 and type 3 axons in the intacttympanal nerve (crosses) and at various times after either extirpa-tion of the auditory organ (filled diamonds) or cutting the nerve(open squares) within the central region of the tympanal nerve.The numbers are means of two to four different preparations. ADecrease in number of type 2 axons. Note the time shift betweenthe two types of surgery. B Decrease in number of type 3 axons af-ter cutting the nerve. Their number did not alter after extirpationof the auditory organ
Fig. 6A,B Alterations within the central nerve part at varioustimes after either cutting the nerve or extirpation of the auditoryorgan. A The share of axons on the tympanal nerve’s area decreas-es with time post surgery. A decrease can be seen earlier when thenerve was cut (open squares) than after extirpation of the auditoryorgan (filled diamonds). B Surgery-induced increase in mitochon-drial number within type 2 axons. Their increase 2 days after cut-ting the nerve is significant. Thereafter, axons start degeneratingand were therefore not taken into account

Usherwood 1972; Bittner and Mann 1976; Meiri et al.1983; Brandstätter et al. 1991). Changes in mitochondri-al morphology are not restricted to distal axonal parts,but can also occur in proximal axonal parts. In our ex-periments the significant increase in mitochondrial num-ber is found only after a cut of the tympanal nerve, notafter extirpation of the auditory organ (Fig. 6B). This in-dicates that the increase is a local reaction at the site oflesion and not directly involved in the degeneration pro-cess. The most obvious sign of degeneration in the dis-tal parts of tympanal sensory axons is the occurrence ofdark cytoplasm and of large lysosomes within the axons,as is also known from other insect preparations (Farleyand Milburn 1969; Brandstätter et al. 1991). This sug-gests autolytic processes rather than digestion by gliacell processes. In contrast, in motor axons and interneu-rones of cockroaches and crayfishes, degeneration ofaxons is accompanied by invagination of glial cell pro-cesses filled with lysosomes (Nordlander and Singer1972; Bittner and Mann 1976; Wood and Usherwood1979), indicating digestion of distal axon parts by glialcells.
Interestingly, the morphologically homogeneous type2 axons of the tympanal nerve belonging to sensory ormotor neurones show very different reactions to a sepa-ration from their somata. In contrast to rapid degenera-tion of distal parts of sensory axons, distal parts of motoraxons do not undergo autolysis – at least not during thefirst 25 days after lesion. Previous studies have shownthat the time it takes distal axonal parts to degenerate canvary largely depending on animals and temperature (e.g.,Rees and Usherwood 1972; Nordlander and Singer 1973;Bittner and Mann 1976; Muller et al. 1987; Bittner 1988;Blundon et al. 1990). Our results indicate that – indepen-dent of extrinsic influences – intrinsic properties of neu-rones are also likely to determine method and duration ofthe degeneration process.
The most dramatic change in the appearance ofanucleate motor axons is their partitioning by invadingglial cell processes. A similar mechanism is well knownfrom crayfish motorneurones (Ballinger and Bittner1980; Govind et al. 1992; Parnas et al. 1998), where dis-tal axons are maintained physiologically functional forup to several months after separation from their somata(Hoy et al. 1967; Govind et al. 1992). It has been sug-gested that the invading glial cells produce the neces-sary proteins to maintain axonal function of survivingdistal axons (Bittner 1988; Bittner 1991; Parnas et al.1998). The amount of proteins transferred from glialcells to the axon would be extended by the invasion ofglial cell processes into the axon, as this increases thesurface area between the axon membrane and the glialelements. We have not investigated whether the distalmotor axons remain functional, but nevertheless our re-sults give a first hint of differential reactions to lesion offunctional different types of axons in the nervoussystem of insects.
Implications of axonal degeneration on regeneration of auditory afferents
After lesion, auditory sensory cells of grasshoppershave the capability to regrow axons into their target ar-ea and to form functional synapses onto auditory inter-neurones (Lakes and Kalmring 1991; Pfahlert andLakes-Harlan 1997). Our results proved that the regen-erating fibres cannot fuse with the distal axonal parts ofauditory cells, as they degenerate within a few days.Furthermore, regenerating auditory fibres cannot be di-rected to their target areas by distal parts of the cut ax-ons as regeneration can take up to 3 weeks, by whichtime degeneration is long complete. Orientation alongdegeneration products or glial cells is possible, howev-er, as these persist up to the time outgrowing axons en-ter the nerve. The role of glial cells on regeneration ofaxons has been intensively investigated in vertebratecentral and peripheral nervous system, leading to astrong differentiation between different types of glialcells: in the CNS white matter, oligodendrocytes areknown to inhibit regeneration by presenting and pro-ducing inhibitory factors, and microglia and especiallyastrocytes prevent regeneration by forming a glial scarthrough which axons cannot grow (for reviews, seeSchwab and Bertholdi 1996; Aldskogius and Kozlova1998; Bregman 1998). In mammalian peripheralnerves, where regeneration does take part, Schwanncells have been reported to be either permissive for re-generation or even promoting regeneration. Further-more, a rapid degeneration of distal axon parts has beenshown to influence rate and specificity of axonal regen-eration. Experimentally delayed Wallerian degenerationof sensory axons delays the onset of regeneration(Brown et al. 1992; Dailey et al. 1998; Bregman 1998),and experimentally induced predegeneration of sensorydistal axons accelerates their regeneration and increasestheir specificity (Hasan et al. 1996; Kerns et al. 1993;Brushart et al. 1998). Predegeneration is considered toeither stimulate trophic factor production, or to removeinhibitors of regeneration allowing axons earlier andmore extensive contact with specific pathway markers.To date, in insects no trophic factors have been identi-fied, but it is known from an in vitro assay that sensoryneurones respond to mammalian trophic factors by in-creased axonal outgrowth (Pfahlert and Lakes-Harlan1997). This might be taken as a hint that in vivo a fastdegeneration of distal auditory axons might also inducethe release of so far unknown stimulating factors andtherefore be a prerequisite for fast axonal regeneration.Nevertheless, quick degeneration or promotion of re-generation through glial cells seems not to be a decisivefactor in selection of specific pathways of regeneratingaxons in the tympanal nerve of the locust: studies onsingle regenerating auditory fibres revealed that withinthe central nervous system a large number of them fol-low the hair sensilla/multipolar neurone pathway (Ja-cobs 1997). At least some of the small axons (type 3) ofhair sensilla or multipolar neurones appear to degener-
176

ate at a much slower rate than auditory axons. Thus, itis possible that regenerating fibres orientate preferen-tially along not yet degenerated sensory axons whichoriginate in the same nerve rather than search for cellu-lar or molecular cues of their former specific pathway.This view is further supported by ultrastructural studiesin which regenerated fibres were mostly found in closeproximity to intact fibres and not to glial cells in theirspecific pathway (Jacobs 1997).
Acknowledgements We want to thank S. Becker and M. Winklerfor help and technical assistance, and Dr. P. Phelan for general dis-cussion and correcting the language. We are grateful to Prof. Dr.F.-W. Schürmann for providing the electron-microscopic facilitiesand Dr. N. Elsner for general support. Dr. A. Mücke provided thephotograph in Fig. 2B. This study was supported by the DeutscheForschungsgemeinschaft, SFB 406.
References
Aldskogius H, Kozlova EN (1998) Central neuron-glial and glial-glial interactions following axon injury. Prog Neurobiol 55:1–26
Bacon SP, Altman JS (1977) A silver intensification method forcobalt-filled neurons in wholemount preparations. Brain Res138:359–363
Ballinger ML, Bittner GD (1980) Ultrastructural studies of se-vered medial giant and other CNS axons in crayfish. Cell TissRes 208:123–133
Birse SC, Bittner GD (1981) Regeneration of earthworm giant ax-ons following transection or ablation. J Neurophysiol 45:724–742
Bittner GD (1988) Long term survival of severed distal axonalstumps in vertebrates and invertebrates. Am Zool 28:1165–1179
Bittner GD (1991) Long-term survival of anucleate axons and itsimplications for nerve regeneration. Trends Neurosci 14:188–193
Bittner D, Mann DW (1976) Differential survival of isolated por-tions of crayfish axons. Cell Tiss Res 169:301–311
Blundon JA, Sheller AR, Moehlenbruck JW, Bittner GD (1990)Effect of temperature on long term survival of anucleate giantaxons in crayfish and goldfish. J Comp Neurol 297:377–391
Boulton PS (1969) Degeneration and regeneration in the insectcentral nervous system. (I.). Z Zellforsch Mikros Anat 101:98–118
Brandstätter JH, Shaw SR, Meinertzhagen IA (1991) Terminal de-generation and synaptic disassembly following receptor photo-ablation in the retina of the fly’s compound eye. J Neurosci11:1930–1941
Bregman BS (1998) Regeneration in the spinal cord. Curr OpinNeurobiol 8:800–807
Brown MC, Perry VH, Hunt SP, Lapper SR (1992) Consequencesof slow Wallerian degeneration for regenerating motor andsensory axons. J Neurobiol 23:521–536
Brushart TM, Gerber J, Kessens P, Chen Y-G, Royall RM (1998)Contributions of pathway and neuron to preferential motor re-innervation. J Neurosci 18:8674–8681
Dailey AT, Avellino AM, Benthem L, Silver J, Kliot M (1998)Complement depletion reduces macrophage infiltration andactivation during wallerian degeneration and axonal regenera-tion. J Neurosci 18:6713–6722
Distler PG, Boeckh J (1997) Central projections of the maxillaryand antennal nerves in the mosquito Aedes aegypti. J Exp Biol200:1873–1879
Farley RD, Milburn NS (1969) Structure and function of the giantfibre system in the cockroach, Periplaneta americana. J InsectPhysiol 15:457–476
Govind CK, Blundon JA, Kirk MD (1992) Functional degenera-tion of isolated central stumps of crayfish sensory axons. JComp Neurol 322:111–120
Gray EG (1960) The fine structure of the insect ear. Philos TransR Soc Lond Biol 243:75–94
Halex H, Kaiser W, Kalmring K (1988) Projection areas andbranching patterns of the tympanal receptor cells in migratorylocusts, Locusta migratoria and Schistocerca gregaria. CellTiss Res 253:517–528
Hasan NA, Neumann MM, de Souky MA, So KF, Bedi KS (1996)The influence of predegenerated nerve grafts on axonal regen-eration from prelesioned peripheral nerves. J Anat 189:293–302
Hess A (1960) The fine structure of degenerating nerve fibers,their sheaths and their terminations in the central nerve cord ofthe cockroach (Periplaneta americana). J Biophys BiochemCytol 7:339–344
Hoy RR, Bittner GD, Kennedy D (1967) Regeneration in crusta-cean motoneurons: evidence for axonal fusion. Science 156:251–252
Jacobs K (1997) Axonale Degeneration und Regeneration tympan-aler Sinneszellen von Schistocerca gregaria. PhD Thesis. I.Zoologisches Institut, Georg-August-Universität Göttingen
Jacobs K, Lakes-Harlan R (1997) Lectin histochemistry of the me-tathoracic ganglion of the locust Schistocerca gregaria beforeand after axotomy of the tympanal nerve. J Comp Neurol 387:255–265
Jacobs K, Otte B, Lakes-Harlan R (1999) Tympanal receptor cellsof Schistocerca gregaria: correlation of soma positions anddendrite attachment sites, central projections and physiologies.J Exp Zool 283:210–285
Kerns JM, Danielsen N, Holmquist B, Kanje M, Lundborg G (1993)The influence of predegeneration on regeneration through pe-ripheral nerve grafts in the rat. Exp Neurol 122:28–36
Lakes R, Kalmring K (1991) Regeneration of the projection andsynaptic connections of tympanic receptor fibers of Locustamigratoria (Orthoptera) after axotomy. J Neurobiol 22:169–181
Lakes-Harlan R, Pfahlert C (1995) Regeneration of axotomizedtympanal nerve fibres in the adult grasshopper Chorthippusbiguttulus (L.) (Orthoptera: Acrididae) correlates with regain-ing the localization ability. J Comp Physiol A 176:797–807
Lyckman AW, Heidelbaugh SM, Bittner GD (1992) Analysis ofneuritic outgrowth from severed giant axons in Lumbricus ter-restris. J Comp Neurol 318:426–438
Meiri H, Dormann A, Spira ME (1983) Comparison of ultrastruc-tural changes in proximal and distal segments of transected gi-ant fibers of the cockroach Periplaneta americana. Brain Res263:1–14
Meyer J, Elsner N (1995) How respiration affects auditory sensi-tivity in the grasshopper Chorthippus biguttulus (L.). J CompPhysiol A 176:563–573
Muller KJ, Gu X, McClade-McCulloh E, Mason A (1987) Tinker-ing with successful synapse regeneration in the leech: addinginsult to injury. J Exp Biol 132:207–221
Nordlander R, Singer M (1972) Electron microscopy of severedmotor fibers in the crayfish. Z Zellforsch Mikros Anat 126:157–181
Nordlander RH, Singer M (1973) Effects of temperature on the ul-trastructure of severed crayfish motor axons. J Exp Zool 184:289–302
Parnas I, Shahrabany-Baranes N, Feinstein N, Grant P, Adelsberg-er H, Dudel J (1998) Changes in the ultrastructure of survivingdistal segments of severed axons of the rock lobster. J ExpBiol 201:779–791
Pfahlert C, Lakes-Harlan R (1997) Responses of insect neuronesto neurotrophic factors in vitro. Naturwissenschaften 84:163–165
Rees D, Usherwood PNR (1972) Fine structure of normal and de-generating motor axons and nerve-muscle synapses in the lo-cust, Schistocerca gregaria. Comp Biochem Physiol A 43:83–101
177

Römer H (1976) Die Informationsverarbeitung tympanaler Re-zeptorelemente von Locusta migratoria (Acrididae, Ortho-ptera). J Comp Physiol 109:101–122
Schürmann F-W (1980) Experimental anterograde degeneration ofnerve fibers: a tool for combined light- and electron-micro-scopic studies of the insect nervous system. In: Strausfeld NJ,Miller TA (eds) Neuroanatomical Techniques, Springer-Ver-lag, New York, Heidelberg, Berlin, pp 206–239
Schwab ME, Bertholdi D (1996) Degeneration and regeneration ofaxons in the lesioned spinal cord. Physiol Rev 76:319–370
Tung AS-C, Pipa RL (1971) Fine structure of transected intergan-glionic connectives and degenerating axons of wax moth lar-vae. J Ultrastruc Res 36:694–707
von Bernhardi R, Muller K (1995) Repair of the central nervoussystem: lessons from lesions in leech. J Neurobiol 27:353–366
Wood MR, Usherwood PNR (1979) Ultrastructural changes incockroach leg muscle following unilateral neurotomy. J Ultra-struct Res 68:265–280
178





















