Embed Size (px)
Citation preview

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Genetically switched D-lactate production in Escherichia coli
Li Zhou a,b, Dan-Dan Niu c, Kang-Ming Tian a,b, Xian-Zhong Chen a,b, Bernard A. Prior c,Wei Shen a,b, Gui-Yang Shi a,d, Suren Singh e, Zheng-Xiang Wang a,b,n
a Center for Bioresource & Bioenergy, School of Biotechnology, Jiangnan University, 1800 Lihu Avenue, Wuxi 214122, People’s Republic of Chinab The Key Laboratory of Industrial Biotechnology of Ministry of Education, Jiangnan University, 1800 Lihu Avenue, Wuxi 214122, People’s Republic of Chinac Department of Microbiology, University of Stellenbosch, Private Bag X1, Matieland 7602, South Africad National Engineering Laboratory for Cereal Fermentation Technology, Jiangnan University, 1800 Lihu Avenue, Wuxi 214122, People’s Republic of Chinae Department of Biotechnology & Food Technology, Faculty of Applied Sciences, Durban University of Technology, P.O. Box 1334, Durban 4001, South Africa
a r t i c l e i n f o
Article history:
Received 6 August 2011
Received in revised form
3 May 2012
Accepted 11 May 2012Available online 8 June 2012
Keywords:
Genetic switch
Thermo-regulation
D-Lactate production
Lambda pL and pR tandem promoters
LdhA
a b s t r a c t
During a fermentation process, the formation of the desired product during the cell growth phase
competes with the biomass for substrates or inhibits cell growth directly, which results in a decrease in
production efficiency. A genetic switch is required to precisely separate growth from production and to
simplify the fermentation process. The ldhA promoter, which encodes the fermentative D-lactate
dehydrogenase (LDH) in the lactate producer Escherichia coli CICIM B0013-070 (ack-pta pps pflB dld poxB
adhE frdA), was replaced with the l pR and pL promoters (as a genetic switch) using genomic recombi-
nation and the thermo-controllable strain B0013–070B (B0013-070, ldhAp::kan-cIts857-pR–pL), which
could produce two-fold higher LDH activity at 42 1C than the B0013-070 strain, was created. When the
genetic switch was turned off at 33 1C, strain B0013-070B produced 10% more biomass aerobically than
strain B0013-070 and produced only trace levels of lactate which could reduce the growth inhibition
caused by oxygen insufficiency in large scale fermentation. However, 42 1C is the most efficient
temperature for switching on lactate production. The volumetric productivity of B0013-070B improved
by 9% compared to that of strain B0013-070 when it was grown aerobically at 33 1C with a short
thermo-induction at 42 1C and then switched to the production phase at 42 1C. In a bioreactor
experiment using scaled-up conditions that were optimized in a shake flask experiment, strain
B0013-070B produced 122.8 g/l D-lactate with an increased oxygen-limited productivity of 0.89
g/g �h. The results revealed the effectiveness of using a genetic switch to regulate cell growth and
the production of a metabolic compound.
& 2012 Elsevier Inc. All rights reserved.
1. Introduction
Microorganisms have been used as cell factories for theproduction of many homologous and heterologous metabolicproducts, including fermentation products (organic acids, suchas lactate, acetate and butyrate as well as neutral products, suchas ethanol, butanol, acetone and butanediol), amino acids andnumerous secondary metabolites (Kern et al., 2007; Krarner,1996; Mijts and Schmidt-Dannert, 2003; Zhang et al., 2011).However, many of these end compounds are detrimental to cellsor metabolic fluxes are affected due to the over production of thedesired product. For instance, over expressing fermentative D-lac-tate dehydrogenase (LDH, encoded by the ldhA gene) to overproduce D-lactate in Escherichia coli resulted in very poor growth
because excess LDH converts the pyruvate pool to lactate, whichcreates a shortage of 3-carbon metabolic intermediates (Bunchet al., 1997). Therefore, it is desired that the intensified produc-tion of the desired product can be controlled and that it can beinitiated after a high cell density develops. A two-phase fermen-tation with separate growth and production phases has been usedin many studies to achieve medium density cultures (10�20 g/ldry cell weight [DCW]) and greater volumetric productivity(Vemuri et al., 2002; Zhou et al., 2011; Zhu et al., 2007), wherehigh-density cultures and improved volumetric productivity isalso a primary objective for many E. coli based processes (Shiloachand Fass, 2005). However, a certain amount of the end productcould still be produced during the growth phase, which mayinterfere with the ability to produce biomass. For example, E. coli
B0013–070, which contains eight mutations in the ackA, pta, pps,pflB, dld, poxB, adhE and frdA genes, was previously reported toproduce 125 g/l D-lactate and a low level of by-products whentwo-phase fermentation was used (aerobic growth and oxygen-limited production [without the need for strict anaerobic
Contents lists available at SciVerse ScienceDirect
journal homepage: www.elsevier.com/locate/ymben
Metabolic Engineering
1096-7176/$ - see front matter & 2012 Elsevier Inc. All rights reserved.
http://dx.doi.org/10.1016/j.ymben.2012.05.004
n Corresponding author at: Center for Bioresource & Bioenergy, School of
Biotechnology, Jiangnan University, 1800 Lihu Avenue, Wuxi 214122, People’s
Republic of China. Fax: þ86 510 85918121.
E-mail address: [email protected] (Z.-X. Wang).
Metabolic Engineering 14 (2012) 560–568

Author's personal copy
conditions]) (Zhou et al., 2011). However, as much as 7 g/l oflactate may accumulate during the cell growth phase, whichcompetes with cell synthesis for pyruvate. Moreover, the oxygensupply may be insufficient in full-scale fermentations due tolimitations on power, and lactate generation during the growthphase will increase to the detriment of cell mass accumulation,which will result in a low volumetric production of lactate duringthe later production phase. Therefore, in addition to optimizationof the cultivation conditions, genetic approaches also need to bedeveloped to accurately regulate the metabolic fluxes. Further-more, strain selection and breeding could be significant in devel-oping new growth conditions that are simpler and easier.
The leftward (pL) and rightward (pR) promoters of bacterioph-age l enable a simple temperature change to efficiently andrapidly switch on or off the expression of the recombinant protein(Elvin et al., 1990; Love et al., 1996). These promoters are apromising technology that can be used as a genetic switch(Ptashne, 1992) to regulate metabolism. Furthermore, productinhibition effect on the repressible operon will be eliminated ifthe native promoter is changed, which may improve productionof the end product. Recently, a single genetic switch and acomplicated switch composed of several interrelated geneticswitches (in the field of synthetic biology) have been used toprecisely control metabolic pathways and to explore networktopologies (Anesiadis et al., 2008; Atkinson et al., 2003; Gardneret al., 2000; Hasty et al., 2002; Weber et al., 2009). Yet, theseresearches have mostly focused on plasmid expression whereplasmid loss is a significant limitation if antibiotic selection isremoved. Moreover, as Yuan et al. (2006) indicated, over-expres-sion of genes on multi-copy vectors has the disadvantage ofsegregational instability, metabolic burden and the potential forgene rearrangements. Mass production of the catalyzing enzymecan also reduce carbon resource. Beyond Yuan et al., few studieshave considered improving the production of desired compoundsthrough the use of chromosomal gene regulation.
In the present investigation, the production of D-lactate, whichis an important chiral intermediate and substrate for polylacticacid (PLA) production, is used as an example to illustrate theeffectiveness of a genetic switch. The D-lactate producer B0013-070 was used as the host strain, and the pR–pL promoters wereexploited as a genetic switch to regulate the expression of LDHand the subsequent production of lactate. The promoter of thechromosomal ldhA gene, ldhAp, in strain B0013-070 was replacedby the pR–pL promoters, which were derived from the plasmidpPL451 (Love et al., 1996), to construct strain B0013-070B(B0013-070, ldhAp::kan-cIts857-pR–pL). Subsequently, the ability
of the genetic switch to control production was studied usingaerobic and oxygen-limited two-phase fermentation with glucoseas the sole carbon source. Consequently, lactate accumulationwas expected to be minimized during the cell growth phase toreduce the strict oxygen requirement for growth and intensifiedduring the oxygen-limited phase by over-expression of LDH.Moreover, the importance of the genetic switch during metabolicregulation may also be applicable to the production of otherhomologous and heterologous compounds.
2. Materials and methods
2.1. Strains
The genotypes of the microbial strains and plasmids used inthe present study are summarized in Table 1. The Escherichia coli
strain CICIM B0013-070 (Dack-pta::dif Dpps::dif DpflB::dif
Ddld::dif DpoxB::FRT DadhE::dif DfrdA::dif) was reported pre-viously (Zhou et al., 2011) and kept in the Culture and Informa-tion Center of Industrial Microorganisms of China Universities(http://cicim-cu.jiangnan.edu.cn/). Strain B0013-070B (B0013-070, ldhAp::Km-cIts857-pR–pL) was constructed during this study.
To construct the B0013-070B, Km-cIts857-pR–pL cassette whichencodes a kanamycin resistance selectable marker, the l repres-sor and the tandem l pR and pL promoters, was placed before thestart codon of the chromosomal ldhA gene in B0013-070. Detailsof the primers used in this study are provided in Table 2. The ldhA’
gene was cloned from the genomic DNA of B0013-070 using PCRamplification and the primers Ec-lA3-5 and YldhA3. The resulting
Table 1E. coli strains and plasmids used in this study.
Strains Relevant characteristics Source or reference
CICIM B0013-070 B0013, Dack-pta::dif Dpps::dif DpflB::dif Ddld::dif DpoxB::FRT DadhE::dif DfrdA::dif (Zhou et al., 2011)
CICIM B0013-070B B0013-070, ldhAp::kan-cIts857-pR–pL This study
Plasmids Relevant characteristics Source or reference
pMD18-T simple bla; TA cloning vector TaKaRa, Japan
pPL451 bla, lcIts857, pR pL (Love et al., 1996)
pSKsymKm bla kan CICIM-CUa; (Overhage et al., 1999)
T-ldhA0 bla, ldhA0; ldhA0 (PCR using Ec-lA3-5/YldhA3) from E. coli B0013-070 cloned into
pMD18-T simple vector
This study
pPL-kan bla kan, lcIts857, pR pL; kan SmaI fragment from pSKsymKm cloned into reverse
PCR fragment of pPL451 (using PPL1/PPL2 primers)
This study
T-ldhAp::kan-cIts857-pR–pL bla kan, ldhAp::kan-cIts857-pR–pL; kan-cIts857-pR–pL (PCR using PPL3/PPL4 primers)
from pPL-kan (EcoRI and EcoRV digested) cloned into reverse PCR fragment of T-ldhA0
(using Ec-RlA1/Ec-RlA2 primers and digested by EcoRI)
This study
pKD46 bla, g b exo (red recombinase), temperature-conditional pSC101 replicon CGSCb; (Datsenko and Wanner, 2000)
a The Culture and Information Center of Industrial Microorganisms of China Universities, Jiangnan University.b Genetic Stock Center, Yale University.
Table 2Primers used in this study.
Primers Sequence (50-30) restriction sites are italic/underlined Restriction
sites
Ec-lA3-5 TCCGGTACCaCAGCCCGAGCGTCATCAG KpnIa
YldhA3 GTCAAGGTCGACGTTATTGAAACCG
Ec-RlA1 TAAGAATTCaATGAAACTCGCCGTTTATAGCACAAAACAG EcoRIa
Ec-RlA2 AAGACTTTCTCCAGTGATGTTGAATCACAT
PPL1 AGCTTGGCTGCAGGTGATGATTATCAGC
PPL2 ATCGCCGGCAATTCGTAATCATGG
PPL3 TAAGATATCaCCATGATTACGAATTGCCGGC EcoRVa
PPL4 TAAGAATTCaAGTTAACCTCCTTAGGATCCCAATGCTT EcoRIa
YldhA4 TCATCTAATGCAATACGTGTCCCGAGC
a Restriction sites with corresponding restriction enzyme.
L. Zhou et al. / Metabolic Engineering 14 (2012) 560–568 561

Author's personal copy
550-bp PCR fragment, which included the promoter and part ofthe structural region of the ldhA gene, was inserted into thepMD18-T simple vector to create T-ldhA’. The reverse PCR frag-ment from plasmid pPL451 was amplified using the primers PPL1and PPL2. The amplified fragment was then ligated to thekanamycin resistance gene (966-bp), which was isolated fromthe SmaI–SmaI sites of pSKsymKm (Overhage et al., 1999) (CICIMP0121; http://cicim-cu.jiangnan.edu.cn/), to create pPL-kan. Sub-sequently, the kan-cIts857-pR–pL fragment was amplified frompPL-kan using the primers PPL3 and PPL4. After digestion by EcoRIand EcoRV, this fragment was cloned into the reverse PCRfragment of T-ldhA’, which was amplified using the primers Ec-RlA1 and Ec-RlA2 and then digested by EcoRI, to produce plasmidT-ldhAp::kan-cIts857-pR–pL. Afterwards, this plasmid was digestedwith KpnI and gel purified. The resulting fragment was used as thetemplate for PCR amplification using the primers Ec-lA3-5 andYldhA3 to create the ldhAp::kan-cIts857-pR–pL cassette. The result-ing DNA was electroporated into strain B0013-070, which con-tained the Red recombinase plasmid pKD46 (Baba et al., 2006;Datsenko and Wanner, 2000; Sambrook and Russell, 2001).Kanamycin resistant colonies were then selected. The presenceof the ldhAp::kan-cIts857-pR–pL mutation was confirmed by per-forming colony PCR with the primers YldhA3 and YldhA4. PCRamplification with these two primers yielded a 658-bp fragmentfrom the wild-type ldhA gene and a 3,033-bp fragment from theldhAp::kan-cIts857-pR–pL mutation, which were further confirmedby nucleotide sequencing. The resulting recombinant strain wasdesignated B0013-070B (B0013-070, ldhAp::kan-cIts857-pR–pL).
2.2. Measurement of LDH activity
Strains (stored as glycerol stocks at –80 1C) were first grownon Luria-Bertani (LB) plates for approximately 24 h at 30 1C andthen the colonies were transferred to 50 ml of LB medium in a250-ml flask. After growing at 30 1C while shaking at 200 rpm for11 h, the cells were collected and used to inoculate 50 ml offreshly modified M9 medium containing 5 g/l glucose at a con-centration of 0.02 g/l dry cell weight (DCW) as previouslyreported (Zhou et al., 2011). The flasks were incubated at 30 1Cwhile shaking at 200 rpm for 6 h and then at 30 1C and 42 1C, foranother 7 h. Crude cell extracts were prepared using the bacterialsoluble total protein preparation kit (GENMED Scientifics Inc.,Arlington, MA, USA). The extracts were assayed for LDH activityusing a kit to colorimetrically determine the total bacterial LDHactivity (GENMED Scientifics Inc., Arlington, MA, USA). One unit ofthe overall LDH activity was defined as the amount of enzymerequired to transform 1 mmol of pyruvate per minute. The proteinconcentration in the crude extracts was determined using theBradford method (Bradford, 1976), and bovine serum albuminwas used as the standard. The LDH activity was divided by thecorresponding protein concentration to calculate the specific LDHactivity. Modified M9 medium contained (per liter): 15.11 gNa2HPO4 �12H2O, 3 g KH2PO4, 1 g NH4Cl, 0.5 g NaCl. One ml offilter-sterilized 1 M MgSO4, and 1 ml of filter-sterilized traceelement solution containing (per liter) 2.4 g FeCl3 �6H2O, 0.3 gCoCl2 �6H2O, 0.15 g CuCl2 �2H2O, 0.3 g ZnCl2, 0.3 g Na2MO4 �2H2O,0.075 g H3BO3, 0.495 g MnCl2 �4H2O was added to a liter of thefinal medium.
2.3. Cultivation conditions for determining the optimal aerobic
growth temperature
Cells were pre-cultured in LB medium as described in the‘‘Measurement of LDH activity’’. After inoculating the bacteriainto modified M9 medium containing 5 g/l glucose, the flasks
were incubated at 30 1C, 33 1C, 35 1C, 37 1C and 40 1C whileshaking at 200 rpm for 13 h.
2.4. Cultivation conditions for determining induction temperature
The cells were pre-cultured and inoculated into modified M9medium as described above. For aerobic cultivation, the flaskswere initially kept at 30 1C while shaking at 200 rpm for 7 h, andthen the cultures were incubated at 30 1C, 35 1C, 37 1C, 40 1C,42 1C and 45 1C while shaking at 200 rpm for another 4 h toinduce the expression of LDH. Afterwards, 1.5 ml of 500 g/lglucose and 2 g CaCO3 (for the purpose of pH adjustment) wereadded to the broth, and the flasks were kept stationary at 30 1C toinitiate the oxygen-limited phase. The two-phase fermentations(with an aerobic and oxygen-limited phase) were stopped after82 h.
2.5. Cultivation conditions for determining the duration of thermal
induction
Cells were pre-cultured and inoculated in modified M9 med-ium as described above. A two-phase fermentation with anaerobic phase, followed by an oxygen-limited phase was initiated.The aerobic growth phase was conducted at 33 1C while shakingat 200 rpm for 9.5 h. Afterwards, the cultures were incubated at42 1C while shaking at 200 rpm for another 0.5 h to induceexpression of LDH. Then, 1.5 ml of 500 g/l glucose and 2 g CaCO3
were added to the broth, and the flasks were kept stationary at42 1C to start the oxygen-limited phase. The two-phase fermenta-tions were stopped when the glucose was depleted. As a control,another experiment was conducted at 33 1C while shaking at200 rpm for 10 h and then kept stationary at 42 1C.
2.6. Bioreactor experiments
For the bioreactor experiments, cells were pre-cultured andinoculated into a 7-liter bioreactor (Bioflow110; New BrunswickScientific Co., Edison, NJ, USA) as previously reported (Zhou et al.,2011), except for the temperatures used were 30 1C and 33 1C forthe pre-cultivation and bioreactor growth, respectively. Growthwas initiated by sparging air into the bioreactor at 1 to 7 l/min,and the dissolved oxygen concentration was maintained above40% saturation by agitation at 200 to 1000 rpm. The pH wasmaintained at 7 by automatically feeding concentrated NH4OHand 10% (vol/vol) H2SO4 (Zhou et al., 2011). The temperature wasset at 42 1C when the cell concentration reached an OD600 ofapproximately 20 (�7.6 g/l DCW). After 45 min, the oxygen-limited phase was initiated. During this second phase, the airsparging was stopped, the agitation was slowed to 100 rpm, thepH was maintained at 7 using 25% (wt/vol) Ca(OH)2 and thetemperature was maintained at 42 1C. The residual glucose con-centration was maintained above 10 g/l by adding glucose in fourbatches (649.5 g of glucose was added in total) as indicated inFig. 3. The fermentations were stopped when the glucose wasexhausted.
2.7. Analytical methods
The cell mass was estimated by measuring the optical densityat 600 nm (if CaCO3 or Ca(OH)2 had been added during fermenta-tion, samples were pretreated with 1 M HCl at 20-fold of thesample volume to remove the suspended substances), and the dryweight of the cells was calculated using a standard curve(1 OD600¼0.38 g/l DCW) (Zhou et al., 2011). The glucose concen-tration was estimated using a glucose biosensor (Zhou et al.,2011). Samples were pretreated with H2SO4 (at 5% of the sample
L. Zhou et al. / Metabolic Engineering 14 (2012) 560–568562

Author's personal copy
volume) to release organic acids that precipitated with CaCO3 orCa(OH)2 during fermentation. Organic acid concentrations weremeasured by HPLC as described previously (Zhou et al., 2011).
3. Results and discussion
3.1. Replacement of the ldhA promoter (ldhAp) with the pR and pL
promoters
Genomic DNA analysis indicated that the kan-cIts857-pR–pL
fragment was correctly inserted into the ldhAp locus of the B0013-070 chromosome (data not shown). The resulting mutant wasnamed B0013-070B (B0013-070, ldhAp::kan-cIts857-pR–pL), andthe structure of the upstream region of ldhA is shown in Fig. 1. Theresidual ldhAp gene remained in the B0013-070B strain but wasnot functional (Fig. 1). DNA sequencing indicated that the regionupstream of ldhAp in B0013-070B was not altered after recombi-nation (data not shown). Therefore, the function of the upstreamgene, ydbH, is not affected. Subsequently, B0013-070B and theparent strain, B0013-070, were cultured at 30 1C and 42 1C, andthe activity of lactate dehydrogenase (LDH) was investigated(Fig. 2). The specific LDH activity of B0013-070 at 301C wasstandardized to 100%. Comparatively, B0013-070B had low LDHactivity at 30 1C and high activity at 42 1C. Since the LDH activitywas lower at 42 1C than at 30 1C in the B0013-070 strain, thetemperature change alone does not fully account for the behaviorof strain B0013-070B. Therefore, the expression of ldhA wassuccessfully controlled by the temperature through the pR–pL
promoters. Villaverde et al. (1993) reported that no enzymaticactivity was detected at temperatures below 32 1C from the pR–pL
promoters, although a very slight band corresponding to themolecular mass of the recombinant protein was perceptible evenat 28 1C using Western blot analysis. In the present investigation,
the strain differences may account for the detectable LDH activityin B0013-070B at 30 1C because even a mutant strain of B0013-070 with the ldhA gene deleted can produce trace levels of lactate(data not shown). For the convenience of genetic manipulation, anEcoRI site was included before the start codon of ldhA gene instrain B0013-070B (Fig. 1). Consequently, the space between thestart codon and the recombinant ribosome-binding site (RBS) wasextended to 11-bp, which might reduce the initiation efficiency ofthe pR–pL promoters. Nevertheless, the resulting LDH activity inthe B0013-070B strain was two-fold higher than in the B0013-070 strain when it was cultured at 42 1C (Fig. 2), which issufficient for lactate production.
The temperature-inducible tandem pL and pR promoters havebeen used to switch on plasmids in synthetic biology (Basu et al.,2004; Hooshangi et al., 2005). However, as we realized previously,over-expression of the catalyzing enzyme on a multi-copy-num-ber plasmid decreases the yield of the desired product (data notshown), which is possibly due to a higher consumption of thesubstrate by the recombinant protein and a disruption of thecentral metabolic flux. In this study, we inserted a genetic switchupstream of the native chromosomal ldhA gene in the lactateproducing strain B0013-070 to regulate the production of lactate.The aim of this project was to decrease lactate generation duringthe cell growth phase to improve biomass accumulation and thento strengthen lactate production after a high cell density wasreached without reducing the overall yield of lactate.
3.2. The effect of temperature on aerobic growth
E. coli recombinant cultures carrying heat inducible expressionvectors were generally grown at 30 1C to achieve a high celldensity and to avoid the adverse effects of the recombinantprotein on cell growth (Norma et al., 2010). However, cells arenormally grown slowly at a low temperature (Ratkowsky et al.,1983). A higher growth rate and a shorter duration of the growthphase will increase productivity of the overall fermentationprocess. Meanwhile, the high cell level obtained during thegrowth phase may increase the volumetric productivity duringthe following production phase. Therefore, it is necessary tooptimize the growth temperature to improve the growth rateand biomass yield during the aerobic phase.
B0013-070B and its parent strain, B0013-070, were grownaerobically at 30 1C, 33 1C, 35 1C, 37 1C and 40 1C in shake flasksusing glucose as the sole carbon source. The fermentation data areshown in Table 3. The parent strain, B0013-070, accumulated ahigher biomass and produced less lactate (in terms of yield andconcentration) at 30 1C compared with other temperatures. How-ever, the lower growth rate was undesirable because ideally theunproductive growth phase should be as short as possible toachieve a higher overall level of productivity. Only trace levels oflactate were produced by B0013-070B when grown at 30 1C,which was in accordance with the low LDH activity that wasdetected at 30 1C. After increasing the cultivation temperature to33 1C and 35 1C, the specific growth rate of B0013-070B wasincreased by 12% compared to when it was grown at 30 1C. Thegenetic switch was slightly turned on and produced a minimalamount of lactate in the B0013-070B strain, which is consistentwith the report that indicated that denaturation of the lambdacI857 repressor is very inefficient at temperatures below 36 1C(Villaverde et al., 1993). Because of the significant decrease in theloss of substrate due to lactate production, the fluxes wereredistributed at the pyruvate node for biomass accumulation,and the biomass yields in the B0013-070B strain at 33 1C were10% higher than for the B0013-070 strain. However, a notablyhigher amount of pyruvate accumulated because the pathwaywas blocked in the B0013-070B strain. When grown at 37 1C,
Fig. 2. Comparison of specific LDH activities of E. coli CICIM B0013-070 and
B0013-070B.
Fig. 1. Chromosomal structure of the ldhA upstream region of E. coli CICIM B0013-
070B. ldhAp: promoter of ldhA gene. The ribosome binding site (RBS) is marked.
The start codon (ATG) is italic.
L. Zhou et al. / Metabolic Engineering 14 (2012) 560–568 563

Author's personal copy
lactate accumulation was much higher in the B0013-070B strainthan when it was grown at 35 1C but was still less than for theB0013-070 strain at 37 1C, which indicates that the LDH expres-sion level in the B0013-070B strain was not as high as in the wildtype strain even at 37 1C. However, when strain B0013-070B wasgrown at 40 1C, the lactate yield was considerably higher and theaccumulation of pyruvate was noticeably lower than for theB0013-070 strain, which indicates that expression of LDH wastriggered. The higher lactate accumulation in strain B0013-070Bresulted in lower biomass yields than for B0013-070. Acetate andsuccinate yields were not affected by the temperature. Thespecific growth rates of both strains increased as the temperatureincreased from 30 1C to 37 1C but significantly decreased at 40 1Cdue to the limitation of methionine synthesis in the minimalmedium (Ron and Davis, 1971). Although the growth rate at 33 1Cwas not higher than at 37 1C, it was still maintained at a highlevel. Based on the higher biomass yield and growth rate and thelower lactate yield, 33 1C was selected as the optimal temperaturefor aerobic growth.
In the present study, genetic methods were used to controlmetabolism, to improve biomass accumulation and to createsimpler cultivation conditions. The maximum specific growthrate and the biomass yield are adversely affected by fermentativeproducts, such as acetate, ethanol, propionate, isobutyrate andlactate (Pan et al., 1987). Therefore, lactate production should belimited. Lactate is the main fermentative product during thegrowth phase of strain B0013-070 (Zhou et al., 2011). However,the cell mass accumulation rate of strain B0013-070 was notaffected by down regulating ldhA expression (in the catabolicpathway, Table 3). This effect may be because the pronouncedeffect of acetic acid (Luli and Strohl, 1990; Pan et al., 1987) did notdecrease (except at 40 1C) or because the microbial growthkinetics were linked to upstream molecular events that controlthe production of rate-limiting enzymes (Koutinas et al., 2011).After optimization of the growth temperature, the biomass yieldof strain B0013-070B at 33 1C improved by 13% compared to thatof strain B0013-070 at 37 1C (strain B0013-070 was previouslyreported to be grown at 37 1C; (Zhou et al., 2011)) with a minimalamount of lactate. This improvement could be more significantwhen oxygen supply is deficient, and therefore enable large-scalefermentation with easier available fermentation process.
3.3. The effect of the LDH induction temperature on lactate
production
The thermo-induced expression system was generally inducedat 42 1C to initiate production of recombinant proteins. However,dramatic cell death and lower production levels were observedwhen cultures were induced at temperatures up to 42 1C
(Villaverde et al., 1993). Some other drawbacks, such as cell wallalterations or incorrect folding of recombinant proteins that arecaused by the high temperature, have also been reported whenthe bacteria are grown at 42 1C (Villaverde et al., 1993). Moreover,because strain B0013-070B has significantly higher LDH activityat 42 1C (Fig. 2), the amount of LDH produced at a lowertemperature might be adequate for lactate production. Therefore,this system may allow control of LDH expression at a lowertemperature than the current system.
To determine the induction temperature for LDH expression, atwo-phase fermentation (aerobic and oxygen-limited) with aserial temperature induction from 30 1C to 42 1C was conductedwith strain B0013-070B, and strain B0013-070 was used as acontrol strain (Table 4). Synthesis of LDH was initially maintainedin the off state by culturing at 30 1C until the late growth phase(aerobic heat induction for 4 h) and was switched off again duringthe oxygen-limited phase to eliminate LDH formation duringthese processes. Due to the low cultivation temperature of 30 1Cthat is used for the second phase, the lactate production byB0013-070 was much lower than was reported previously (Zhouet al., 2011), and the overall lactate yields were lower. In thecontrol experiment with constant cultivation at 30 1C, B0013-070B produced a low level of lactate and had a low glucoseconsumption rate, which indicates that lactate production wasmostly switched off. After increasing the induction temperature,lactate production and yields increased gradually, which suggeststhat lactate production can be progressively turned on by increas-ing the temperature. However, the overall lactate yield fromB0013-070B was still lower than that of B0013-070 when theywere induced at 40 1C. When induced at 42 1C, the lactate yield ofstrain B0013-070B reached same level as that of strain B0013-070, and the oxygen-limited lactate specific productivity was 33%higher than for strain B0013-070, which could be resulted fromthe higher LDH activity in B0013-070B at 42 1C. The low acetatelevel produced by strain B0013-070B was not significantlyaffected by changing the induction temperature. The accumula-tion of succinate did not change in a linear manner as theinduction temperature rose, which may have resulted from theredistribution of the carbon flow between lactate and otherproducts at different temperatures in strain B0013-070B. Thepyruvate yield in B0013-070B was generally decreased at lowerinduction temperatures, which is consistent with the previousreport that indicated that in the range of 30 1C to 42 1C, 42 1C isthe most efficient temperature for inducing the tandem pR and pL
promoters, which control protein expression (Villaverde et al.,1993). Additionally, 42 1C is also the most efficient temperaturefor increasing production of the metabolic product and wastherefore used as the induction temperature in the followingexperiments.
Table 3Comparison of fermentation data (mean7standard deviation of triplicate experiments) when the bacteria were grown at different temperatures.
Strain Growth
temperature
Specific growth
rate m (1/h)
Lactate
titer (g/l)
Yield (g/100 g glucose)a
Biomass Lactate Pyruvate Acetate Succinate
B0013-070 30 1C 0.4070.00 1.770.1 26.471.2 27.371.3 16.370.6 0.970.1 0.070.0
33 1C 0.4670.00 2.370.1 23.470.3 37.171.6 12.670.7 1.170.1 0.070.0
35 1C 0.4770.00 2.470.1 22.471.0 40.372.2 11.170.3 1.270.1 0.070.0
37 1C 0.4670.01 2.470.1 22.871.2 41.071.3 14.470.7 1.370.0 0.070.0
40 1C 0.4070.02 2.470.1 16.670.7 46.670.5 20.472.2 0.870.0 0.170.0
B0013-070B 30 1C 0.4070.01 0.070.0 28.770.8 0.670.1 51.373.1 1.370.1 0.070.0
33 1C 0.4570.01 0.170.0 25.871.4 1.770.1 56.173.8 1.270.1 0.170.0
35 1C 0.4570.01 0.270.0 24.470.6 3.070.2 57.870.8 1.370.1 0.170.0
37 1C 0.4670.00 0.970.0 22.970.8 16.770.9 44.172.2 1.270.1 0.170.0
40 1C 0.4370.01 3.270.1 14.070.5 65.773.2 3.570.5 0.670.1 0.070.0
a Average yield during the growth phase.
L. Zhou et al. / Metabolic Engineering 14 (2012) 560–568564
Author's personal copy
Several studies have reported on the regulation of cI857-controlled protein production at different induction temperatures(Soares et al., 2003; Villaverde et al., 1993). In contrast, theproduct of interest in the present investigation is the chemicalproduct of the recombinant protein. Although over-expression ofLDH was necessary to increase lactate productivity during theoxygen-limited phase, it was not expected to produce a masslevel to waste the cellular resource for lactate production. There-fore, a novel induction strategy was required. Because lactategeneration during the aerobic phase was reduced, the overallproduction of lactate in strain B0013-070B at induction tempera-tures lower than 42 1C were not as high as from strain B0013-070despite the fact that comparable amounts of oxygen-limitedlactate production was obtained when they were induced at40 1C. Irrespective of the considerable increase in specific pro-ductivity, the volumetric productivity by strain B0013-070B didnot improved when compared to strain B0013-070 because alower cell mass accumulated aerobically at 42 1C. Therefore, it isnecessary to further optimize this stage and the duration ofthermal induction.
3.4. Effect of the duration of LDH thermal induction on lactate
production
Cell yield is much lower when the bacteria are grown at 42 1Cthan when they are grown at 33 1C (data not shown). Therefore, ashort induction period or even no induction at 42 1C during theaerobic growth phase was preferred. Furthermore, concurrentformation of LDH at 42 1C during the oxygen-limited productionphase (growth-limited phase) was expected to reinforce lactateproduction. The experiments with different thermal inductionduration in the aerobic phase and oxygen-limited cultivation at42 1C were performed in shake flasks using strain B0013-070 asthe control strain, and the results were summarized in Table 5.
Without thermal induction under aerobic conditions, the over-all biomass yield from strain B0013-070B was higher than fromstrain B0013-070. Lactate productivity by strain B0013-070B wasslightly increased compared to that of strain B0013-070, whichindicates that strain B0013-070B could produce sufficient LDH at42 1C even under oxygen-limited conditions. The short cultivationtime at 42 1C resulted in only a negligible reduction in biomassformation during the aerobic phase (Table 5), and aerobic biomassyields from strain B0013-070B were still higher than from strainB0013-070 by 10%, which suggests that a short aerobic inductionis feasible. Lactate productivity by strain B0013-070B was furtherimproved by approximately 4% and was 4% higher than that ofstrain B0013-070 under the same conditions. Therefore, a shortinduction time for LDH expression is better for larger scalefermentations. Lactate production and oxygen-limited lactateyield by strain B0013-070B was greater than that of strainB0013-070, whereas the levels of the by-products succinate, inthe competing route, and pyruvate, which is the precursor oflactate, were lower than for strain B0013-070.
Several previous studies have substantially increased lactateproduction by removing competing pathways from E. coli strains(Zhou et al., 2003a, 2006, 2011, Zhu and Shimizu, 2004, 2005; Zhuet al., 2007) without over expressing LDH. Zhou et al. reportedthat increased lactate productivity can be achieved by signifi-cantly increasing LDH activity in strain SZ85 (focA-pflB frdBC adhE
ackA ldhA::ldhL) after selecting for improved growth (Zhou et al.,2003b), which demonstrates a promising capacity for increasedlactate production by over expressing LDH even in a metabolicallyengineered lactate producer. In the present investigation, over-expression of LDH was dynamically regulated during lactatefermentation. Consequently, high efficient cell accumulation (asa result of switching off lactate production) and lactateT
ab
le4
Co
mp
ari
son
of
ferm
en
tati
on
da
ta(m
ea
n7
sta
nd
ard
de
via
tio
no
ftr
ipli
cate
ex
pe
rim
en
ts)
wh
en
the
ex
pre
ssio
no
fLD
Hw
as
ind
uce
da
td
iffe
ren
tte
mp
era
ture
s.
Str
ain
Ind
uct
ion
tem
pe
ratu
re
Lact
ate
vo
lum
etr
ic
pro
du
ctiv
ity
(g/L
h)a
Ox
yg
en
-lim
ite
d
lact
ate
spe
cifi
c
pro
du
ctiv
ity
(g/g
h)b
Ox
yg
en
-lim
ite
d
spe
cifi
cg
luco
se
con
sum
pti
on
rate
(g/g
h)c
Ox
yg
en
-lim
ite
d
lact
ate
yie
ld
(g/1
00
gg
luco
se)d
Lact
ate
tite
r
(g/l
)
Yie
ld(g
/10
0g
glu
cose
)e
Bio
ma
ssLa
cta
teA
ceta
teS
ucc
ina
teP
yru
va
te
B0
01
3-0
70
301C
0.1
87
0.0
00
.107
0.0
10
.117
0.0
18
9.57
0.2
15
.27
0.4
8.77
0.4
69
.07
1.8
0.67
0.0
3.17
0.1
2.07
0.1
351C
0.1
97
0.0
10
.117
0.0
10
.137
0.0
18
3.37
4.2
16
.17
0.6
8.27
0.3
73
.57
3.2
0.47
0.1
2.37
0.1
1.67
0.1
371C
0.1
97
0.0
10
.127
0.0
00
.147
0.0
18
2.87
4.4
16
.17
0.7
7.77
0.1
75
.27
2.3
0.47
0.0
1.97
0.1
2.07
0.1
401C
0.1
97
0.0
10
.137
0.0
10
.137
0.0
29
8.37
4.6
16
.27
0.9
7.27
0.3
76
.57
2.3
0.47
0.0
1.97
0.1
2.17
0.0
421C
0.1
97
0.0
10
.127
0.0
00
.147
0.0
09
1.37
1.9
16
.07
0.9
7.37
0.2
76
.27
3.7
0.47
0.0
1.97
0.0
2.07
0.1
B0
01
3-0
70
B3
01C
f0
.027
0.0
00
.027
0.0
00
.087
0.0
12
2.37
1.8
1.87
0.0
9.57
0.5
12
.47
0.8
0.77
0.0
9.57
0.5
21
.27
1.0
351C
0.1
17
0.0
10
.087
0.0
10
.127
0.0
16
8.97
3.6
9.47
0.5
8.67
0.4
48
.67
2.3
0.57
0.0
0.97
0.0
23
.97
1.1
371C
0.1
77
0.0
00
.117
0.0
00
.137
0.0
08
7.07
0.6
14
.07
0.0
7.87
0.2
64
.57
1.5
0.47
0.0
1.87
0.1
9.87
0.3
401C
0.1
87
0.0
00
.147
0.0
10
.147
0.0
19
5.07
2.1
14
.87
0.0
7.17
0.3
68
.07
1.2
0.37
0.0
2.67
0.1
1.47
0.1
421C
0.1
97
0.0
10
.167
0.0
10
.167
0.0
29
7.47
4.7
16
.17
0.6
5.77
0.3
75
.37
1.7
0.27
0.0
1.47
0.1
1.87
0.1
aA
ve
rag
ev
olu
me
tric
pro
du
ctiv
ity
for
lact
ate
du
rin
gth
eo
ve
rall
ferm
en
tati
on
pro
cess
.b
Av
era
ge
spe
cifi
cla
cta
tep
rod
uct
ivit
yd
uri
ng
ox
yg
en
-lim
ite
dp
ha
se.
cA
ve
rag
esp
eci
fic
glu
cose
con
sum
pti
on
rate
du
rin
go
xy
ge
nli
mit
ed
ph
ase
.d
Av
era
ge
yie
ldd
uri
ng
the
ox
yg
en
-lim
ite
dp
ha
se.
eA
ve
rag
ey
ield
du
rin
gth
eo
ve
rall
ferm
en
tati
on
pro
cess
.f
Inco
mp
lete
uti
liza
tio
no
fg
luco
sesu
bst
rate
.
L. Zhou et al. / Metabolic Engineering 14 (2012) 560–568 565
Author's personal copy
production (as a result of the increased LDH activity) wereachieved separately using easier available fermentation processwith strain B0013-070B.
3.5. Bioreactor experiment
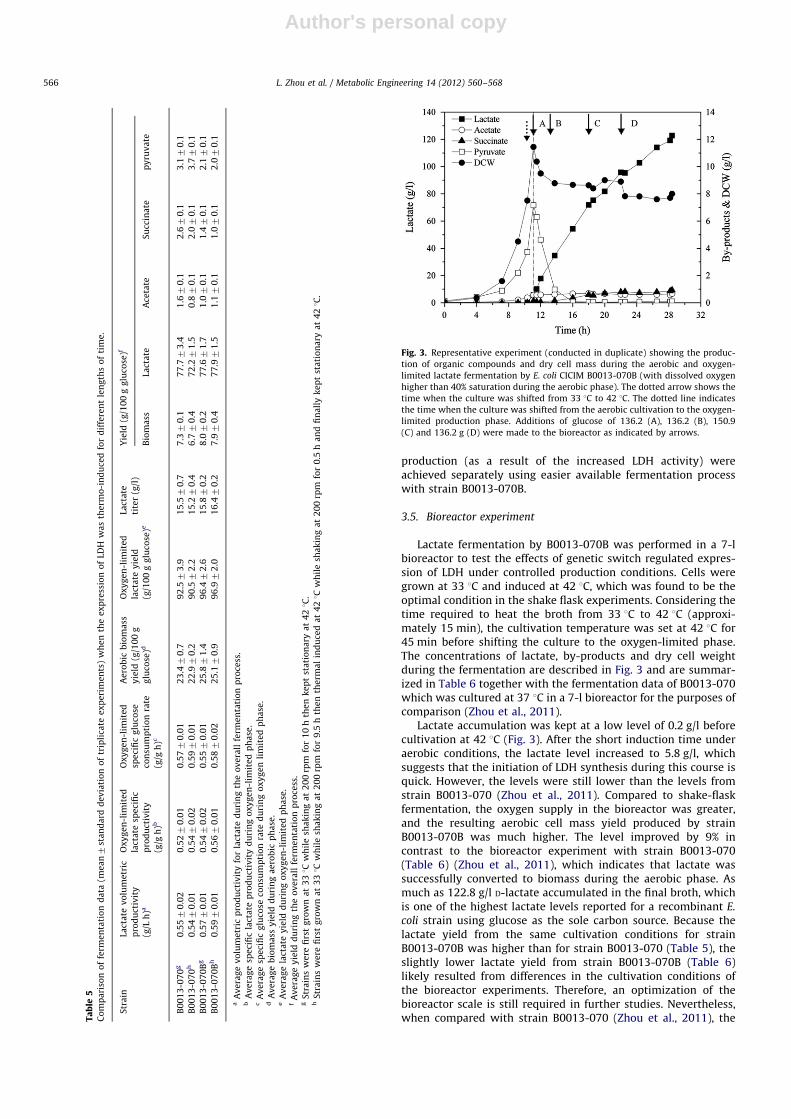
Lactate fermentation by B0013-070B was performed in a 7-lbioreactor to test the effects of genetic switch regulated expres-sion of LDH under controlled production conditions. Cells weregrown at 33 1C and induced at 42 1C, which was found to be theoptimal condition in the shake flask experiments. Considering thetime required to heat the broth from 33 1C to 42 1C (approxi-mately 15 min), the cultivation temperature was set at 42 1C for45 min before shifting the culture to the oxygen-limited phase.The concentrations of lactate, by-products and dry cell weightduring the fermentation are described in Fig. 3 and are summar-ized in Table 6 together with the fermentation data of B0013-070which was cultured at 37 1C in a 7-l bioreactor for the purposes ofcomparison (Zhou et al., 2011).
Lactate accumulation was kept at a low level of 0.2 g/l beforecultivation at 42 1C (Fig. 3). After the short induction time underaerobic conditions, the lactate level increased to 5.8 g/l, whichsuggests that the initiation of LDH synthesis during this course isquick. However, the levels were still lower than the levels fromstrain B0013-070 (Zhou et al., 2011). Compared to shake-flaskfermentation, the oxygen supply in the bioreactor was greater,and the resulting aerobic cell mass yield produced by strainB0013-070B was much higher. The level improved by 9% incontrast to the bioreactor experiment with strain B0013-070(Table 6) (Zhou et al., 2011), which indicates that lactate wassuccessfully converted to biomass during the aerobic phase. Asmuch as 122.8 g/l D-lactate accumulated in the final broth, whichis one of the highest lactate levels reported for a recombinant E.
coli strain using glucose as the sole carbon source. Because thelactate yield from the same cultivation conditions for strainB0013-070B was higher than for strain B0013-070 (Table 5), theslightly lower lactate yield from strain B0013-070B (Table 6)likely resulted from differences in the cultivation conditions ofthe bioreactor experiments. Therefore, an optimization of thebioreactor scale is still required in further studies. Nevertheless,when compared with strain B0013-070 (Zhou et al., 2011), theT
ab
le5
Co
mp
ari
son
of
ferm
en
tati
on
da
ta(m
ea
n7
sta
nd
ard
de
via
tio
no
ftr
ipli
cate
ex
pe
rim
en
ts)
wh
en
the
ex
pre
ssio
no
fLD
Hw
as
the
rmo
-in
du
ced
for
dif
fere
nt
len
gth
so
fti
me
.
Str
ain
Lact
ate
vo
lum
etr
ic
pro
du
ctiv
ity
(g/L
h)a
Ox
yg
en
-lim
ite
d
lact
ate
spe
cifi
c
pro
du
ctiv
ity
(g/g
h)b
Ox
yg
en
-lim
ite
d
spe
cifi
cg
luco
se
con
sum
pti
on
rate
(g/g
h)c
Ae
rob
icb
iom
ass
yie
ld(g
/10
0g
glu
cose
)d
Ox
yg
en
-lim
ite
d
lact
ate
yie
ld
(g/1
00
gg
luco
se)e
Lact
ate
tite
r(g
/l)
Yie
ld(g
/10
0g
glu
cose
)f
Bio
ma
ssLa
cta
teA
ceta
teS
ucc
ina
tep
yru
va
te
B0
01
3-0
70
g0
.557
0.0
20
.527
0.0
10
.577
0.0
12
3.47
0.7
92
.57
3.9
15
.57
0.7
7.37
0.1
77
.77
3.4
1.67
0.1
2.67
0.1
3.17
0.1
B0
01
3-0
70
h0
.547
0.0
10
.547
0.0
20
.597
0.0
12
2.97
0.2
90
.57
2.2
15
.27
0.4
6.77
0.4
72
.27
1.5
0.87
0.1
2.07
0.1
3.77
0.1
B0
01
3-0
70
Bg
0.5
77
0.0
10
.547
0.0
20
.557
0.0
12
5.87
1.4
96
.47
2.6
15
.87
0.2
8.07
0.2
77
.67
1.7
1.07
0.1
1.47
0.1
2.17
0.1
B0
01
3-0
70
Bh
0.5
97
0.0
10
.567
0.0
10
.587
0.0
22
5.17
0.9
96
.97
2.0
16
.47
0.2
7.97
0.4
77
.97
1.5
1.17
0.1
1.07
0.1
2.07
0.1
aA
ve
rag
ev
olu
me
tric
pro
du
ctiv
ity
for
lact
ate
du
rin
gth
eo
ve
rall
ferm
en
tati
on
pro
cess
.b
Av
era
ge
spe
cifi
cla
cta
tep
rod
uct
ivit
yd
uri
ng
ox
yg
en
-lim
ite
dp
ha
se.
cA
ve
rag
esp
eci
fic
glu
cose
con
sum
pti
on
rate
du
rin
go
xy
ge
nli
mit
ed
ph
ase
.d
Av
era
ge
bio
ma
ssy
ield
du
rin
ga
ero
bic
ph
ase
.e
Av
era
ge
lact
ate
yie
ldd
uri
ng
ox
yg
en
-lim
ite
dp
ha
se.
fA
ve
rag
ey
ield
du
rin
gth
eo
ve
rall
ferm
en
tati
on
pro
cess
.g
Str
ain
sw
ere
firs
tg
row
na
t3
31C
wh
ile
sha
kin
ga
t2
00
rpm
for
10
hth
en
ke
pt
sta
tio
na
rya
t4
21C
.h
Str
ain
sw
ere
firs
tg
row
na
t3
31C
wh
ile
sha
kin
ga
t2
00
rpm
for
9.5
hth
en
the
rma
lin
du
ced
at
421C
wh
ile
sha
kin
ga
t2
00
rpm
for
0.5
ha
nd
fin
all
yk
ep
tst
ati
on
ary
at
421C
.
Fig. 3. Representative experiment (conducted in duplicate) showing the produc-
tion of organic compounds and dry cell mass during the aerobic and oxygen-
limited lactate fermentation by E. coli CICIM B0013-070B (with dissolved oxygen
higher than 40% saturation during the aerobic phase). The dotted arrow shows the
time when the culture was shifted from 33 1C to 42 1C. The dotted line indicates
the time when the culture was shifted from the aerobic cultivation to the oxygen-
limited production phase. Additions of glucose of 136.2 (A), 136.2 (B), 150.9
(C) and 136.2 g (D) were made to the bioreactor as indicated by arrows.
L. Zhou et al. / Metabolic Engineering 14 (2012) 560–568566

Author's personal copy
volumetric production of lactate and the specific productivityduring the oxygen-limited phase increased by 51% and 46% instrain B0013-070B, respectively. Considering that the first phasewas not shortened, the overall volumetric productivity increasedby 22%, which illustrates the significance of using the geneticswitch. Low levels of less than 1 g/l acetate, succinate andpyruvate accumulated as by-products. In comparison with ourprevious report (Zhou et al., 2011), acetate accumulation duringthe aerobic phase was lower in strain B0013-070B and wasfollowed by a minimal acetate yield of 0.4 g/100 g glucose(Table 6), which may be because the oxygen supply was increasedduring the growth phase. These results were consistent with aprevious report that found that a succinate producer, E. coli
AFP111, accumulates a significant amount of acetate at a mediumoxygen transfer rate (volumetric oxygen mass transfer coeffi-cients [kLa] is 52), and never accumulates acetate while at a highoxygen transfer rate (kLa is 69) (Vemuri et al., 2002). Becauselactate production is blocked, the pyruvate level steadilyincreased as the cell mass accumulated during the aerobic phaseand then decreased swiftly after the initiation of the oxygen-limited phase; only trace amounts were detectable in the finalbroth, which indicates that the high lactate production rate occursduring the second phase.
Using two-phase fed batch fermentation, an engineered E. coli
strain, ALS974, produced 138 g/l lactate with a second-phaseproductivity of 6.3 g/l h when acetate was used as a carbonsource for cell growth in addition to glucose, and the productionphase was conducted under strict anaerobic conditions (Zhu et al.,2007). The overall productivity further improved to 4.2 g/l h withonly minimal creation of by-products when cell-recycle fermen-tation was used (Zhu et al., 2008). In the present investigation, theoverall volumetric productivity and the oxygen-limited produc-tivity was as high as 4.32 and 6.73 g/l h, respectively. These levelsare some of the highest levels ever produced by a recombinant E.
coli strain that did not require special substrates or strict aerobicand anaerobic conditions.
4. Conclusions
The l pR–pL promoters could be used as a genetic switch toregulate cell growth and the production of a chemical compound(lactate) by regulating chromosomal gene expression of thecatalyzing enzyme (LDH). Switching off lactate generation duringthe aerobic cell growth phase can improve biomass accumulation,causing a decrease in the strict oxygen requirement for cellgrowth, and intensifying LDH expression after the genetic switchis switched on could promote lactate formation during theproduction phase. High efficient cell accumulation and lactateproduction were achieved separately using easier available fer-mentation process with strain B0013-070B. The thermal induc-tion strategies could be useful for other research, particularlythose that are studying fermentation products.
Acknowledgments
This work was partly funded by the Sino-South Africa Coop-eration Program 2009DFA31300 and CS06-L10 and the NationalNatural Science Foundation of China no. 21006039.
References
Anesiadis, N., Cluett, W.R., Mahadevan, R., 2008. Dynamic metabolic engineeringfor increasing bioprocess productivity. Metab. Eng. 10, 255–266.T
ab
le6
Co
mp
ari
son
of
ferm
en
tati
on
da
ta(m
ea
n7
ran
ge
of
du
pli
cate
ex
pe
rim
en
ts)
fro
mth
eb
iore
act
or
ex
pe
rim
en
ts.
Str
ain
sO
ve
rall
vo
lum
etr
ic
lact
ate
pro
du
ctiv
ity
(g/l
h)a
Ox
yg
en
-
lim
ite
d
vo
lum
etr
ic
lact
ate
pro
du
ctiv
ity
(g/l
h)b
Ox
yg
en
-
lim
ite
dsp
eci
fic
lact
ate
pro
du
ctiv
ity
(g/g
h)c
Ox
yg
en
-
lim
ite
dsp
eci
fic
glu
cose
con
sum
pti
on
rate
(g/g
h)d
Ae
rob
icb
iom
ass
yie
ld
(g/1
00
gg
luco
se)
e
Ox
yg
en
-lim
ite
d
lact
ate
yie
ld
(g/1
00
gg
luco
se)f
Lact
ate
tite
r(g
/l)
Yie
ld(g
/10
0g
glu
cose
)gS
ou
rce
s
Bio
ma
ssLa
cta
teA
ceta
teS
ucc
ina
teP
yru
va
te
B0
01
3-0
70
B4
.327
0.1
36
.737
0.3
30
.897
0.0
40
.957
0.0
43
7.97
1.2
93
.57
4.3
12
2.87
3.7
5.37
0.2
84
.37
2.1
0.47
0.0
0.67
0.0
0.17
0.0
Th
isst
ud
y
B0
01
3-0
70
3.3
27
0.0
34
.467
0.0
60
.617
0.0
20
.637
0.0
13
4.87
0.8
96
.07
0.3
12
5.17
2.1
4.97
0.1
86
.67
1.3
4.37
1.4
0.97
0.3
0.27
0.0
(Zh
ou
et
al.
,
20
11
)
aA
ve
rag
ev
olu
me
tric
pro
du
ctiv
ity
for
lact
ate
du
rin
gth
eo
ve
rall
ferm
en
tati
on
pro
cess
.b
Av
era
ge
vo
lum
etr
icp
rod
uct
ivit
yfo
rla
cta
ted
uri
ng
the
ox
yg
en
-lim
ite
dp
ha
se.
cA
ve
rag
esp
eci
fic
lact
ate
pro
du
ctiv
ity
du
rin
gth
eo
xy
ge
n-l
imit
ed
ph
ase
.d
Av
era
ge
spe
cifi
cg
luco
seco
nsu
mp
tio
nra
ted
uri
ng
the
ox
yg
en
-lim
ite
dp
ha
se.
eA
ve
rag
eb
iom
ass
yie
ldd
uri
ng
ae
rob
icp
ha
se.
fA
ve
rag
ela
cta
tey
ield
du
rin
gth
eo
xy
ge
n-l
imit
ed
ph
ase
.g
Av
era
ge
yie
ldd
uri
ng
the
ov
era
llfe
rme
nta
tio
np
roce
ss.
L. Zhou et al. / Metabolic Engineering 14 (2012) 560–568 567

Author's personal copy
Atkinson, M.R., Savageau, M.A., Myers, J.T., Ninfa, A.J., 2003. Development ofgenetic circuitry exhibiting toggle switch or oscillatory behavior in Escherichiacoli. Cell. 113, 597–607.
Baba, T., Ara, T., Hasegawa, M., Takai, Y., Okumura, Y., Baba, M., Datsenko, K.A.,Tomita, M., Wanner, B.L., Mori, H., 2006. Construction of Escherichia coli K-12in-frame, single-gene knockout mutants: the Keio collection. Mol. Syst. Biol. 22006.0008.
Basu, S., Mehreja, R., Thiberge, S., Chen, M.T., Weiss, R., 2004. Spatiotemporalcontrol of gene expression with pulse-generating networks. Proc. Natl. Acad.Sci. U. S. A. 101, 6355–6360.
Bradford, M.M., 1976. A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dye binding.Anal. Biochem. 72, 248–254.
Bunch, P.K., Mat-Jan, F., Lee, N., Clark, D.P., 1997. The ldhA gene encoding thefermentative lactate dehydrogenase of Escherichia coli. Microbiology. 143,187–195.
Datsenko, K.A., Wanner, B.L., 2000. One-step inactivation of chromosomal genes inEscherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. U. S. A. 97,6640–6645.
Elvin, C.M., Thompson, P.R., Argall, M.E., Philip Hendr, N., Stamford, P.J., Lilley, P.E.,Dixon, N.E., 1990. Modified bacteriophage lambda promoter vectors for over-production of proteins in Escherichia coli. Gene. 87, 123–126.
Gardner, T.S., Cantor, C.R., Collins, J.J., 2000. Construction of a genetic toggle switchin Escherichia coli. Nature. 403, 339–342.
Hasty, J., McMillen, D., Collins, J.J., 2002. Engineered gene circuits. Nature. 420,224–230.
Hooshangi, S., Thiberge, S., Weiss, R., 2005. Ultrasensitivity and noise propagationin a synthetic transcriptional cascade. Proc. Natl. Acad. Sci. U. S. A. 102,3581–3586.
Kern, A., Tilley, E., Hunter, I.S., Legisa, M., Glieder, A., 2007. Engineering primarymetabolic pathways of industrial micro-organisms. J. Biotechnol. 129, 6–29.
Koutinas, M., Kiparissides, A., Silva-Rocha, R., Lam, M.C., Martins dos Santos, V.A.P.,de Lorenzo, V., Pistikopoulos, E.N., Mantalaris, A., 2011. Linking genes tomicrobial growth kinetics–An integrated biochemical systems engineeringapproach. Metab. Eng. 13, 401–413.
Krarner, R., 1996. Genetic and physiological approaches for the production ofamino acids. J. Biotechnol. 45, 1–21.
Love, C.A., Lilley, P.E., Dixon, N.E., 1996. Stable high-copy-number bacteriophage[lambda] promoter vectors for overproduction of proteins in Escherichia coli.Gene. 176, 49–53.
Luli, G.W., Strohl, W.R., 1990. Comparison of growth, acetate production, andacetate inhibition of Escherichia coli strains in batch and fed-batch fermenta-tions. Appl. Environ. Microbiol. 56, 1004–1011.
Mijts, B.N., Schmidt-Dannert, C., 2003. Engineering of secondary metabolite path-ways. Curr. Opin. Biotechnol. 14, 597–602.
Norma, V.C., Luis, C., Nestor, P., Octavio, R., Mauricio, T.R., 2010. Production ofrecombinant proteins in E. coli by the heat inducible expression system basedon the phage lambda pL and/or pR promoters. Microb. Cell Fact. 9, 18–24.
Overhage, J., Priefert, H., Rabenhorst, J., Steinbuchel, A., 1999. Biotransformation ofeugenol to vanillin by a mutant of Pseudomonas sp. strain HR199 constructedby disruption of the vanillin dehydrogenase (vdh) gene. Appl. Microbiol.Biotechnol. 52, 820–828.
Pan, J.G., Rhee, J.S., Lebeault, J.M., 1987. Physiological constraints in increasingbiomass concentration of Escherichia coli B in fed-batch culture. Biotechnol.Lett. 9, 89–94.
Ptashne, M., 1992. A genetic switch: phage [lambda] and higher organisms. CellPress, Cambridge.
Ratkowsky, D.A., Lowry, R.K., McMeekin, T.A., Stokes, A.N., Chandler, R.E., 1983.Model for bacterial culture growth rate throughout the entire biokinetictemperature range. J. Bacteriol. 154, 1222–1226.
Ron, E.Z., Davis, B.D., 1971. Growth rate of Escherichia coli at elevated tempera-tures: limitation by methionine. J. Bacteriol. 107, 391–396.
Sambrook, J., Russell, D.W., 2001. Molecular cloning: a laboratory manual. ColdSpring Harbor Laboratory Press, New York.
Shiloach, J., Fass, R., 2005. Growing E. coli to high cell density—a historicalperspective on method development. Biotechnol. Adv. 23, 345–357.
Soares, C.R.J., Gomide, F.I.C., Ueda, E.K.M., Bartolini, P., 2003. Periplasmic expres-sion of human growth hormone via plasmid vectors containing the lPL
promoter: use of HPLC for product quantification. Protein Eng. 16, 1131–1138.Vemuri, G.N., Eiteman, M.A., Altman, E., 2002. Succinate production in dual-phase
Escherichia coli fermentations depends on the time of transition from aerobicto anaerobic conditions. J. Ind. Microbiol. Biotechnol. 28, 325–332.
Villaverde, A., Benito, A., Viaplana, E., Cubarsi, R., 1993. Fine regulation of cI857-controlled gene expression in continuous culture of recombinant Escherichiacoli by temperature. Appl. Environ. Microbiol. 59, 3485–3487.
Weber, W., Lienhart, C., Daoud-El Baba, M., Fussenegger, M., 2009. A biotin-triggered genetic switch in mammalian cells and mice. Metab. Eng. 11,117–124.
Yuan, L.Z., Rouvi�ere, P.E., LaRossa, R.A., Suh, W., 2006. Chromosomal promoterreplacement of the isoprenoid pathway for enhancing carotenoid productionin E. coli. Metab. Eng. 8, 79–90.
Zhang, X.Z., Sathitsuksanoh, N., Zhu, Z., Percival Zhang, Y.H., 2011. One-stepproduction of lactate from cellulose as the sole carbon source without anyother organic nutrient by recombinant cellulolytic Bacillus subtilis. Metab.Eng. 13, 364–372.
Zhou, L., Zuo, Z.R., Chen, X.Z., Niu, D.D., Tian, K.M., Prior, B.A., Shen, W., Shi, G.Y.,Singh, S., Wang, Z.X., 2011. Evaluation of genetic manipulation strategies onD-lactate production by Escherichia coli. Curr. Microbiol. 62, 981–989.
Zhou, S., Causey, T.B., Hasona, A., Shanmugam, K.T., Ingram, L.O., 2003a. Productionof optically pure D-lactic acid in mineral salts medium by metabolicallyengineered Escherichia coli W3110. Appl. Environ. Microbiol. 69, 399–407.
Zhou, S., Shanmugam, K.T., Ingram, L.O., 2003b. Functional replacement of theEscherichia coli D-(�)-lactate dehydrogenase gene (ldhA) with the L-(þ)-lactatedehydrogenase gene (ldhL) from Pediococcus acidilactici. Appl. Environ. Micro-biol. 69, 2237–2244.
Zhou, S., Shanmugam, K.T., Yomano, L.P., Grabar, T.B., Ingram, L.O., 2006. Fermen-tation of 12% (w/v) glucose to 1.2 M lactate by Escherichia coli strain SZ194using mineral salts medium. Biotechnol. Lett. 28, 663–670.
Zhu, J., Shimizu, K., 2004. The effect of pfl gene knockout on the metabolism foroptically pure D-lactate production by Escherichia coli. Appl. Microbiol. Bio-technol. 64, 367–375.
Zhu, J., Shimizu, K., 2005. Effect of a single-gene knockout on the metabolicregulation in Escherichia coli for D-lactate production under microaerobiccondition. Metab. Eng. 7, 104–115.
Zhu, Y., Eiteman, M.A., Altman, E., 2008. Indirect monitoring of acetate exhaustionand cell recycle improve lactate production by non-growing Escherichia coli.Biotechnol. Lett. 30, 1943–1946.
Zhu, Y., Eiteman, M.A., DeWitt, K., Altman, E., 2007. Homolactate fermentation bymetabolically engineered Escherichia coli strains. Appl. Environ. Microbiol. 73,456–464.
L. Zhou et al. / Metabolic Engineering 14 (2012) 560–568568





























