Embed Size (px)
Citation preview

Environment International 75 (2015) 180–198
Contents lists available at ScienceDirect
Environment International
j ourna l homepage: www.e lsev ie r .com/ locate /env int
Review
Integrated phytobial remediation for sustainable management of arsenicin soil and water
Madhumita Roy b,⁎, Ashok K. Giri a, Sourav Dutta b, Pritam Mukherjee b
a Molecular and Human Genetics Division, CSIR-Indian Institute of Chemical Biology, 4Raja S.C. Mallick Road, Kolkata 700032, West Bengal, Indiab Techno India University, Salt Lake, Kolkata 700091, India
⁎ Corresponding author.
http://dx.doi.org/10.1016/j.envint.2014.11.0100160-4120/© 2014 Elsevier Ltd. All rights reserved.
a b s t r a c t
a r t i c l e i n f oArticle history:Received 14 June 2014Received in revised form 10 November 2014Accepted 15 November 2014Available online xxxx
Keywords:ArsenicBioremediationPhytoremediationRhizoremediationHyperaccumulationTransgenicConstructed wetland
Arsenic (As), cited as the most hazardous substance by the U.S. Agency for Toxic Substance and Disease Registry(ATSDR, 2005), is an ubiquitousmetalloidwhich when ingested for prolonged periods cause extensive health ef-fects leading to ultimate untimely death. Plants andmicrobes can helpmitigate soil and groundwater As problemsince they have evolved elaborate detoxification machineries against this toxic metalloid as a result of their co-existence with this since the origin of life on earth. Utilization of the phytoremediation and bioremediation po-tential of the plants and microbes, respectively, is now regarded as two innovative tools that encompassbiology, geology, biotechnology and allied sciences with cutting edge applications for sustainable mitigation ofAs epidemic. Discovery of As hyperaccumulating plants that uptake and concentrate large amounts of thistoxic metalloid in their shoots or roots offered new hope to As phytoremediation, solar power based nature'sown green remediation. This review focuses on how phytoremediation and bioremediation can be merged to-gether to form an integrated phytobial remediation which could synergistically achieve the goal of large scale re-moval of As from soil, sediment and groundwater and overcome the drawbacks of the either processes alone. Thereview also points to the feasibility of the introduction of transgenic plants andmicrobes that bring new hope formore efficient treatment of As. The review identifies one critical research gap on the importance of remediation ofAs contaminated groundwater not only for drinking purpose but also for irrigation purpose and stresses thatmore research should be conducted on the use of constructed wetland, one of themost suitable areas of applica-tion of phytobial remediation. Finally the review has narrowed down on different phytoinvestigation andphytodisposal methods, which constitute the most essential and the most difficult part of pilot scale and fieldscale applications of phytoremediation programs.
© 2014 Elsevier Ltd. All rights reserved.
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1812. Bioremediation of arsenic . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1823. Phytoremediation of arsenic . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 183
3.1. Mechanism of phytoremediation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1843.1.1. Arsenic uptake . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1843.1.2. Arsenic transport . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1843.1.3. Detoxification . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 185
3.2. Different types of arsenic phytoremediation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1853.2.1. Phytoextraction/phytoaccumulation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1853.2.2. Phytostabilization/phytoimmobilisation/phytorestoration . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1863.2.3. Phytovolatilization/rhizovolatilization . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1863.2.4. Phytofiltration/rhizofiltration . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1863.2.5. Phytostimulation/natural attenuation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 187
4. Phytobial remediation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1874.1. Phytobialremediation by free living microbes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 188
4.1.1. Mobilization . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 188

181M. Roy et al. / Environment International 75 (2015) 180–198
4.1.2. Immobilization and bioaccumulation/biosequestration . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1884.1.3. Biotransformation/volatilization . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 189
4.2. Phytobialremediation by rhizospheric microbes/phytorhizoremediation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1894.3. Phytobialremediation by endophytic microbes/endophytoremediation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1894.4. Phytobialremediation by mycorrizal fungi/phytomycoremediation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 189
5. Strategies for enhanced phytoremediation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1905.1. Transgenic technology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 190
5.1.1. Transgenic microbes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1905.1.2. Transgenic rhizomicrobes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1905.1.3. Transgenic plants . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 190
5.2. Other methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1915.2.1. Biostimulation/bioaugmentation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1915.2.2. Phytoremediation with electrokinetics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1915.2.3. Permeable reactive barrier . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 191
6. Areas of application of phytobial technology and some case studies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1927. Phytoinvestigation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1928. Phytodisposal . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1929. Conclusion and recommendations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 193Acknowledgment . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 194References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 194
1. Introduction
The heavy metalloid arsenic (As) ranking the group V element ofthe periodic table is the 20th most abundant element in the Earth'spresent-day crust. Evidences indicate that it was an abundant ele-ment on the primordial Earth surface and served as an energy sourceto some beginning life forms while posed biochemical challenge forothers (Olsen et al., 1999). As the Earth cooled, heavy elements likesulfur and As were engulfed within the core and mantle leaving a lit-tle on the surface of the Earth causing life to flourish. But occasionalvolcanic eruptions, leaching from geologic formations caused it tosurface again. Anthropomorphic sources like application of As con-taining fungicides, pesticides, insecticides and herbicides and irre-sponsible mining activities also contributed its unusual highpercentage in some agricultural fields. The survival of large varietyof life forms including microbes and plants in the presence of thistoxic metalloid is not a surprise but a result of million years of adap-tation, natural selection and evolution (Oremland et al., 2009). Alarge body of literature is present on biogeochemical, pharmacolog-ical and toxicological behavior of As (Oremland and Stolz, 2003; Pottet al., 2001; Smedley and Kinniburgh, 2002; Rosen, 2002).
Well-documented cases of As poisoning have been found inArgentina, Chile, China, Mexico, Taiwan, India and Bangladesh (seereview by Christopher and Haque, 2012). The world's largest As-hitpopulation would be found in the Bengal basin covering parts ofBangladesh and West Bengal, India where millions of people con-sume unsafe concentrations of As because of the presence of shallowtube wells (N100 m) that yield water containing N10 μg L−1 of Aswhich is the maximum contaminant level (MCL) of As in safe drink-ing water. Arsenicosis or As induced health defects due to prolongedexposure to sub-acute levels of As in drinking water include variousdermatological skin lesions, cardiovascular, peripheral vascular dis-eases, neurological disorders, respiratory diseases, limb deformations,eye problems and various forms of cancer and death (Paul et al., 2013;Naujokas et al., 2013). Due to chronic exposure to very low levels ofAs that always exist in both soil and water (Smedley and Kinniburgh,2002), As tolerance or detoxification systems has evolved in most, ifnot all, living organisms, including humans (Rosen, 2002). Health ef-fects, genetic damage and As susceptibility extensively studied by ourgroup indicates that genetic variants and poor nutritional status are re-sponsible for As induced toxicity, susceptibility and carcinogenicity (DeChaudhuri et al., 2008a, 2008b; Banerjee et al., 2011). Ground-waterused for cultivation of staple food crops such as rice, wheat
contaminated with As aggravates the As epidemic (Banerjee et al.,2013; Abedin et al., 2002; Brammer, 2009; Das et al., 1995). Long termuse of As contaminated irrigation water also leads to accumulation ofAs in the fields. Abandoned mines also cause As poisoning in the sur-rounding agricultural soils (Kwon et al., 2012; Susaya et al., 2010). Live-stock fed on As-containing vegetation are source of As food chaincontamination (Samal et al., 2011). The use of alternative source ofdrinking water (deep tube wells of N150 ft depth) may solve drinkingwater crisis locally but for irrigation purpose large scale groundwaterremediation is the only sustainable solution as the overuse of deeperpumping for irrigation or large scale use other than domestic purposes,would induce downwardmigration of As from upper shallow aquifer todeep aquifer, permanently destroying this deep resource (Michael andVoss, 2008; Ravenscroft et al., 2013). Several water treatment technolo-gies like adsorption, co-precipitation, ion exchange, reverse osmosisandmembrane processes can effectively remove As from contaminatedgroundwater (Cheremisinoff, 1998). But most of them are costly andvery large scale application including decontamination of the contami-nated groundwater reservoir is practically not possible. Bioremediationand phytoremediation are the alternative solutions that are cost-effective and environmentally friendly. In contrast to bioremediationwhich in general concept employs microbes, phytoremediationemploys plants. Microflora associated with plants i.e. endophyticbacteria, rhizosphere bacteria and mycorrhizae assist the plants inAs uptake, translocation and accumulation and this process is termedas rhizoremediation. Lynch and Moffat (2005) first used theterm phytobialremediation to redefine phytoremediation assistedby microbes. The discovery of As hyperaccumulator plant Pteris vittataby Ma et al. (2001), paved the way for discovery of other Ashyperaccumulator/accumulator plants and this made phytoremediationof As a reality from imagination.
The present review is a comprehensive complete review that hasdiscussed all aspects of As phytobialremediation (flowchart in thegraphical abstract shows the structure of the review). The main ob-jective of the review is to address the issue that bioremediation,phytoremediation and rhizoremediation are intricately related, com-plementary to each other's disadvantages (Table 1) and is synergistical-ly more effective than individual procedures alone. They should belaunched together to form a holistic approach to stop the aggressionof this world's greatest mass poisoning agent and save millions oflives. Along with showing the detailed mechanism of bioremediationand phytoremediation of As, the authors have discussed how plantsand microbes help each other in the whole process and how

Table 1Some gaps of phytoremediation that can be complemented by bioremediation.
Gaps of phytoremediation Bioremediation fills up the gap
Phytoremediation is limited toshallow soil and shallowgroundwater due to limitations ofplant root length.
Microbes are present at all depths of soiland groundwater. In places wherepumping of groundwater is not possiblefor phytoremediation, bioremediation cancarry out the cleaning job.
High contaminant concentration maylimit vegetation growth.
Microbes may be employed to reducecontaminant load and make the sitehabitable by plants engaged inphytoremediation.
Requirement of long time period forachieving the goal.
Microbes can speed up the process ofcontaminant removal by either indirectlyassisting the plant in several ways (e.g.nitrogen fixation, providing access to biounavailable mineral nutrients includingphosphorus, production of plant growthregulators such as auxins, gibberellinesand cytokinins, suppression of ethylenesynthesis by ACC deaminase, preventinggrowth of pathogenic bacteria) or bydirectly participating in removal of toxicarsenite.
Disposal of huge biomass containingtoxic As is a problem.
Volume of biomass can be reduced bymicrobially mediated pretreatments likecomposting and compaction that greatlywould reduce volume of biomass.
Table 2Speciation states of arsenic: arsenate (As+5), arsenite (As+3), elemental As (As0)and arsine (As−3). The toxicity of As species in vivo follows the order DMA(III),MMA(III) N As(III) N As(V) N DMA(V), MMA(V) N TMAO (Akter et al., 2005).
Species of arsenic Chemical formula and main features
Arsenate As(V) (dominant form in oxic water)Arsenate, an analog of phosphate, has moderate toxicity as itis a potent inhibitor of oxidative phosphorylation. Arsenate isless mobile and therefore less bio available as it more readilygets attached to inorganic surfaces, prominently Fe(III). It canbe present as various protonated forms depending on pH:H3AsO4, H2AsO4−, HAsO4
2−, and AsO43−.
Arsenite As(III) (dominant form in anoxic water)(below pH 9.2, H3AsO3). This form has the highest toxicity asit has high affinity to bind with sulfhydryl groups of proteinsdisrupting enzymes involved in cellular metabolism. It ismore mobile, and quickly comes into aqueous solvents and soare more bioavailable.
Arsenic trioxide As2O3
Many agricultural and medical products used for a variety ofpurposes comprises this form of As.
Arsine AsH3
This form is highly toxic, colorless, flammable, non-irritatinggas. Because of their diluted presence in the atmosphere verylittle information is present regarding natural cycles of thesesubstances.
Organoarseniccompounds
Naturally occurring substances like betaine andarsenobetaine are non toxic and benign. These compoundsare common in many shellfishes and elasmobranchs speciesinhabiting seas. However what benefits they confer to theirmarine host or why these marine forms accumulate thesecompounds is not known. Microbiological transformations ofthese compounds release arsenate.
182 M. Roy et al. / Environment International 75 (2015) 180–198
transgenic and other technology can be used to enhance the effect ofphytobialremediation. The authors have stressed that more researchshould be conducted for As removal in constructed wetland systemand inventing new disposal and recycling techniques for As scaveng-ing from disposed biomass, the two most neglected areas of Asphytobialremediation.
2. Bioremediation of arsenic
Microbes represent the largest number of life forms on earthwhich encompasses all three major domains of life: eukaryotes, bac-teria, archaea and viruses. Survival strategies of microbes in the pres-ence of As through oxidation, reduction, sequestration, extrusionand methylation reactions regulate global cycle of As. Intricate roleof microbes in geochemical cycle of As can be found in many reviews(Mukhopadhyay et al., 2002; Oremland and Stolz, 2003; Stolz et al.,2006; Tsai et al., 2009). Table 2 shows different species of As presentin the biosphere. Prior to undertake a phyto cum bioremediationplanning it is necessary to study the speciation status of As in thatsite in soil–water–plant–human continuum because based on thegeological features of the site and the variance of the microbial com-munity, distinct As species would predominate.
Broadly microbes can be divided into two groups depending onwhether they gain energy or drain energy for their association withAs (Fig. 1). Several aerobic/anaerobic chemoautotrophic bacteria(CAO) that gain energy by oxidizing arsenite and use oxygen or, ni-trate as their terminal electron acceptor have been isolated (Hoeftet al., 2004, 2007; Kulp et al., 2008; Oremland et al., 2002; Santiniand Vanden Hoven, 2004). Heterotrophic bacteria don't gain energybut drains energy either for detoxifying or respiring on As. Hetero-trophic bacteria can be categorized into two types, arsenate reducingbacteria and arsenite oxidizing bacteria. Heterotrophic arsenate re-ducing bacteria are again of two types, cytoplasmic As(V) reducingbacteria and respiratory As(V) reducing bacteria. Heterotrophic ar-senite oxidizing bacteria (HAO) use As(III) as an electron donor atthe beginning of the respiratory chain. Cytoplasmic arsenate reduc-ing bacteria widely present in nature possess As resistant operon(ars) as part of their As detoxification machinery. The As resistanceoperon is represented by either arsABCDR (e.g. plasmid R733 ofEscherichia coli) or arsBCR (e.g. plasmid pI258 of Staphylococcus
aureus). Arsenate enters bacterial cells through phosphate trans-porters namely Pit and Pst while arsenite enters through glycerolchannel GlpF (Rosen and Liu, 2009). Arsenate reductase ArsC geneof the Ars operon then reduces As(V) to As(III) and the As(III) iseffluxed from the cell through ATP dependent efflux pump ArsAB/ArsB. In some cases cells store As(III) either in free form (rare) and/or in conjugation with thiol ligands. Instead of As(V), cells storemore toxic As(III) because this form is able to form inert complexesby binding with many thiol ligands like GSH, PCs, and MTs (Tsaiet al., 2009). Saccharomyces cerevisiae Arr2p is the eukaryotic homo-logue of bacterial arsenate reductase (Mukhopadhyay and Rosen,1998, 2001). The respiratory arsenate reductase (arr) containing mi-crobes that use As(V) as a terminal electron acceptor in anaerobicrespiration is referred as dissimilatory arsenate-respiring prokary-otes (DARPs) (Hollibaugh et al., 2006; Perez-Jimenez et al., 2005).Many organic compounds like acetate, formate, lactate, aromaticsand inorganic compounds like hydrogen and sulfide serve as elec-tron donors during oxidation by these DARPs, releasing As(III). Asthese DARPs are constitutive or facultative anaerobes they aremore abundant in the anoxic region of the aquifers and contributeto the release of adsorbed and coprecipitated As (McArthur et al.,2001; Harvey et al., 2002; Nickson et al., 1998, 2000). Using arrgene as molecular marker (Malasarn et al., 2004), unculturedDARBs were detected from Mono and Searles lakes (Hollibaughet al., 2006; Kulp et al., 2007), As-contaminated groundwater aquiferof Cambodia (Lear et al., 2007; Pederick et al., 2007), and deep andshallow tube wells of Bangladesh (Sutton et al., 2009). Although amajor group of microbes increase environmental As(III) levelthrough arsmediated cytoplasmic reductase or arrmediated respira-tory reductase but another major group balances the As(III) level byoxidizing it again to As(V). The heterotrophic As(III) oxidizing bacte-ria (HAO) and chemoautotrophic As(III) oxidizing bacteria (CAO)both possess oxidase enzyme of slightly different variant (AioA/AioB). Vast numbers of bacteria have been isolated from As contam-inated soils, mines, river sediments and geothermal springs that ox-idize arsenite to arsenate and therefore has bioremediation potential

Fig. 1. Different detoxification strategies and interaction of microbes with arsenic under aerobic and anaerobic conditions. *MAsV and MAsIII are the methylated forms of arsenate andarsenite, respectively.
183M. Roy et al. / Environment International 75 (2015) 180–198
(Donahoe-Christiansen et al., 2004; Gihring et al., 2001; Macur et al.,2004; Oremland et al., 2002; Salmassi et al., 2006). The HAO, CAO andArr possessing bacteria may or may not possess ars operon. In addi-tion to oxidation/reduction reactions, methylation/demethylationreactions are the other major means of regulating global As cycle. Mi-crobial methylation has been reviewed by various authors (Bentley andChasteen, 2002; Frankenberger, 2001; Poser, 2005). The biomethylationpathway involves reduction of As(V) to As(III) and then oxidative addi-tion of methyl groups to the compounds. After formation thesemethyl-ated volatile compounds are released from the microbes and thisphenomenon is called as biovolatilization. Arsenic is also converted tovarious benign organoarsenic compounds found in higher abundancein some marine animals, algae and plants.
Fig. 1 shows the three types of detoxification strategies against As inmicrobes: i) active effluxing of As ii) sequestration within cell as freeAs(III) or as conjugates with thiols iii) biotransformation and methyla-tion. Among themvast group of As(III) oxidizers andAsmethylatingmi-crobes directly participates in As bioremediation. Others most notablyDARPs increase As(III) level in anoxic condition by mobilizing solidAs(V) to dissolved As(III). ArsC possessing microbes although widelydistributed don't increase environmental As(III) level as the reductaseis cytoplasmic in nature and only accepts aqueous As(V) that has en-tered the cell and doesn't mobilize it from environment. The ars operonacts as a mere detoxification strategy. The As(III) is either effluxed fromcell or sequestered inside cell by complex formation with metal chelat-ing proteins. In addition to those shown in Fig. 1 there are many bene-ficial microbes who actively bioremediate As by physical absorption ofAs in their cell wall or precipitating it. A detailed description of these
microbes can be found in Section 4.1while discussing the role of free liv-ing microbes in phytobialremediation of As.
3. Phytoremediation of arsenic
Plants can be classified into As resistant and nonresistant types de-pending on their response to As. Arsenic nonresistant plants cannotgrow or show symptoms of toxicity when grown in As containing soil.Arsenic resistant plants (tolerant, accumulator and hyperaccumulator)can tolerate and grow in moderate to high concentration of As withoutshowing toxic symptoms of arsenicosis. Arsenic tolerant plants resistuptake of As while growing in As containing soil. Arsenic accumulators(accumulate low to moderate concentration of As in root) andhyperaccumulator plants (accumulate very high amount of As inabove ground parts) uptake As but resist it from causing toxic effect.To reduce As species into the food chain and to create As resistant trans-genic crops thorough understanding of themolecularmechanisms of Asuptake and detoxification is of primary importance. Data from thesestudies can be harnessed to create model phytoremediator plant. Bio-chemistry of them has been discussed in many reviews (Xie et al.,2009; Zhao et al., 2010) and would be discussed here in brief. Fig. 2shows two modes of As phytoremediation by hyperaccumulators andaccumulators/tolerators and the underlying mechanism of As uptake,transport and detoxification. Hyperaccumulators (mainly Pteris group)remediate As bymaximum containmentmethod i.e. uptake and storingthem in different underground and aboveground plant parts withoutharm to itself. Others like accumulators and tolerators remediate sites
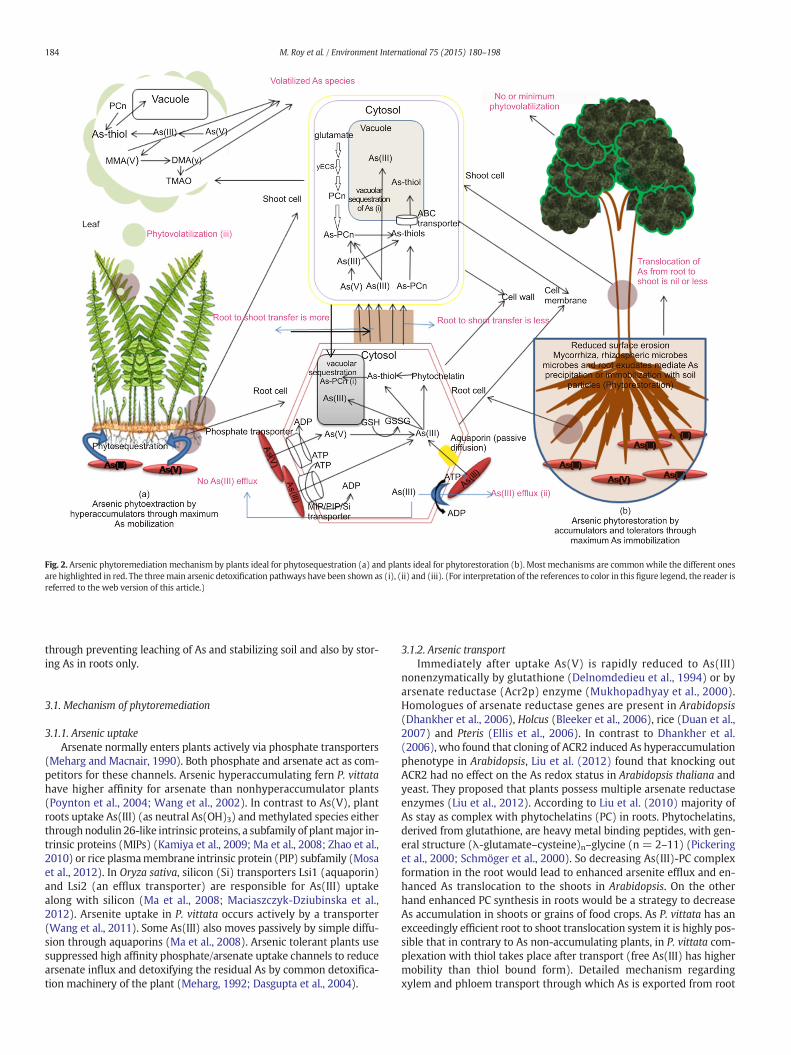
Fig. 2. Arsenic phytoremediation mechanism by plants ideal for phytosequestration (a) and plants ideal for phytorestoration (b). Most mechanisms are commonwhile the different onesare highlighted in red. The threemain arsenic detoxification pathways have been shown as (i), (ii) and (iii). (For interpretation of the references to color in this figure legend, the reader isreferred to the web version of this article.)
184 M. Roy et al. / Environment International 75 (2015) 180–198
through preventing leaching of As and stabilizing soil and also by stor-ing As in roots only.
3.1. Mechanism of phytoremediation
3.1.1. Arsenic uptakeArsenate normally enters plants actively via phosphate transporters
(Meharg and Macnair, 1990). Both phosphate and arsenate act as com-petitors for these channels. Arsenic hyperaccumulating fern P. vittatahave higher affinity for arsenate than nonhyperaccumulator plants(Poynton et al., 2004; Wang et al., 2002). In contrast to As(V), plantroots uptake As(III) (as neutral As(OH)3) andmethylated species eitherthrough nodulin 26-like intrinsic proteins, a subfamily of plantmajor in-trinsic proteins (MIPs) (Kamiya et al., 2009; Ma et al., 2008; Zhao et al.,2010) or rice plasmamembrane intrinsic protein (PIP) subfamily (Mosaet al., 2012). In Oryza sativa, silicon (Si) transporters Lsi1 (aquaporin)and Lsi2 (an efflux transporter) are responsible for As(III) uptakealong with silicon (Ma et al., 2008; Maciaszczyk-Dziubinska et al.,2012). Arsenite uptake in P. vittata occurs actively by a transporter(Wang et al., 2011). Some As(III) also moves passively by simple diffu-sion through aquaporins (Ma et al., 2008). Arsenic tolerant plants usesuppressed high affinity phosphate/arsenate uptake channels to reducearsenate influx and detoxifying the residual As by common detoxifica-tion machinery of the plant (Meharg, 1992; Dasgupta et al., 2004).
3.1.2. Arsenic transportImmediately after uptake As(V) is rapidly reduced to As(III)
nonenzymatically by glutathione (Delnomdedieu et al., 1994) or byarsenate reductase (Acr2p) enzyme (Mukhopadhyay et al., 2000).Homologues of arsenate reductase genes are present in Arabidopsis(Dhankher et al., 2006), Holcus (Bleeker et al., 2006), rice (Duan et al.,2007) and Pteris (Ellis et al., 2006). In contrast to Dhankher et al.(2006), who found that cloning of ACR2 induced As hyperaccumulationphenotype in Arabidopsis, Liu et al. (2012) found that knocking outACR2 had no effect on the As redox status in Arabidopsis thaliana andyeast. They proposed that plants possess multiple arsenate reductaseenzymes (Liu et al., 2012). According to Liu et al. (2010) majority ofAs stay as complex with phytochelatins (PC) in roots. Phytochelatins,derived from glutathione, are heavy metal binding peptides, with gen-eral structure (λ-glutamate–cysteine)n–glycine (n = 2–11) (Pickeringet al., 2000; Schmöger et al., 2000). So decreasing As(III)-PC complexformation in the root would lead to enhanced arsenite efflux and en-hanced As translocation to the shoots in Arabidopsis. On the otherhand enhanced PC synthesis in roots would be a strategy to decreaseAs accumulation in shoots or grains of food crops. As P. vittata has anexceedingly efficient root to shoot translocation system it is highly pos-sible that in contrary to As non-accumulating plants, in P. vittata com-plexation with thiol takes place after transport (free As(III) has highermobility than thiol bound form). Detailed mechanism regardingxylem and phloem transport through which As is exported from root

185M. Roy et al. / Environment International 75 (2015) 180–198
to shoot and then distributed/accumulated to various parts of the shootis not known. Controversy persists regarding the forms of As (freeAs(V)/free As(III)/ thiol bound complex) during transport.
3.1.3. DetoxificationThree types of detoxification pathways exist in As resistant plants
(Fig. 2). The major one is complextion of As(III) with thiol compoundsfollowed by sequestration of the compounds in vacuoles. Effluxing ofAs(III) and methylation cum volatilization are other two means of de-toxification. Arsenite either in root or shoot forms complex with thiols,particularly phytochelatins (PCs). Sequestration of the metal(loid)–PCcomplexes into vacuoles is a strategy by the plant to protect its cellularcomponents from the reactive metal(loid)s. The transporters responsi-ble for active transport of PC-conjugated As(III) have been identifiedas ABC transporters by Song et al. (2010). He also identified vacuolarPC transporters as ABCC-type phytochelatin transporters in Arabidopsis.The resultingAs(III)–PC2 complex isfinally detoxified by its safe storagewithin plant vacuoles. Arsenic nonresistant/sensitive plants cannot sur-vive in moderate to high As containing zones due to damage of rootgrowth or death due to generation of reactive oxygen species by theconversion of arsenite to different methylated species (Barrachinaet al., 1995; Meharg and Macnair, 1991). Aspergillus sp. P37 exhibits ahypertolerant phenotype for As(V) (up to 0.2 M) and reports suggestthat thiol groups play important role in hypertolerancewithmajor frac-tion of the cellular As sequestered in the vacuoles as glutathione conju-gate As(GS)3 (Canovas et al., 2004). Second type of detoxification thatoperates in parallel is active efflux of As(III) from root cells to the exter-nal medium (Xu et al., 2007; Zhang et al., 2009). In rice Lsi1 (NIP) thatoperates as As(III) gate also plays a role in As(III) efflux in rice rootsexposed to As(V) (Zhao et al., 2010). Third detoxification mode ofAs is phytovolatilization of As. Plant species with higher uptake ofAs(V) into roots tends to have higher uptake of MMA and DMA intoroots. Methylated forms of As taken directly from root or generatedthrough methylation of As(III) in plant tissue undergo a series of meth-ylations catalyzed by S-adenosylmethyltransferase, ultimately leadingto its volatilization.
Various antioxidant defence systems generated in response againstAs induced oxidative stress have been characterized in different plants.Recently Tripathi et al. (2012) coined the term “Arsenomics” in a reviewand discussed As induced transcriptomic, proteomic and metabolomechanges. Themechanisms behind hyperaccumulation are enhanced up-take of As(V), decreased arsenite efflux to the external medium withreduced arsenite–thiol complexation and, greatly enhanced xylemtranslocation and vacuolar sequestration of As(III) in fronds. WhenP. vittata grows in soils containing As(V), As is taken up by the rootand is concomitantly reduced to As(III) (Su et al., 2008). This is rapidlytransported to the vacuoles of the upper and lower epidermal cellsand trichomes of the fronds. Fronds accumulate 25 times more Asthan the roots (Tu and Ma, 2005). Arsenic is stored in vacuoles of thefronds in free forms and not conjugated with PCs (Chen et al., 2005;Lombi et al., 2002). The accumulation of free As(III) in the P. vittata vac-uoles contrasts with that observed in other nonhyperaccumulatingplants where As(V) taken up by the root is reduced to As(III) and re-mains sequestered in the root by binding to the thiolate ligands ofphytochelatins (Pickering et al., 2000; Dhankher et al., 2002). Howevera small portion of As(III) (b5%) in P. vittata is retained as a thiolate com-plex in a cylindrical sheath 40- to 50-mm thick, immediately surround-ing the veins of vascular tissue of the fronds (Pickering et al., 2006).Hyperaccumlation of As in P. vittata involves novel or additional Astransporters. One vacuolar arsenite transporter protein ACR3 was iden-tified in P. vittata that effluxes As(III) into vacuoles for sequestration(Indriolo et al., 2010). Biochemical investigation shows that when com-pared to an As-sensitive fern Pteris ensiformis, P. vittata has tolerance tooxidative stress, higher levels of antioxidant enzymes (Singh et al.,2006; Srivastava et al., 2005) and greater levels of reduced GSH. Aglutaredoxin (PvGrx5) was characterized in P. vittata that contributed
to As(III) resistance by playing a role in regulating intracellular As(III)levels, by modulating the aquaglyceroporins (Sundaram et al., 2008).Arsenic also induces enhanced expression of various antioxidant en-zymes like superoxide dismutase, ascorbate peroxidise, and catalase(Cao et al., 2004; Singh et al., 2006; Srivastava et al., 2005) in As-hyperaccumulating plants.
The phenomenon of As hyperaccumulation is not confined to terres-trial plants only but is found in many aquatic plants too (Favas et al.,2012). Some aquatic macrophytes have been found to contain 100–50,000 times As than the surrounding water As. Physicochemicaladsorption facilitated by co-deposition of Fe hydroxides to the plant'ssurface was suggested as probable mechanism (Robinson et al., 2006).The rootless duckweed Wolffia globosa can be an ideal agent forphytofiltration as it accumulates N1000mg kg−1 As in frond, and toler-ate As up to 400 mg kg−1. The passive absorption of the metal by theapoplast and the absence of root-to-shoot translocation barrier mayhave powered the aquatic macrophyte to absorb such a huge collectionof As and Cd (Xie et al., 2013b; Zhang et al., 2009). However a recentstudy has revealed an important role of PC in As detoxification inW. globosa (Zhang et al., 2012). Some photosynthetic microorganismsincludingmarine and freshwater algae, can accumulate high concentra-tion of As andmetabolize them into arsenosugar like organic As species(Phillips, 1990).
3.2. Different types of arsenic phytoremediation
Arsenic can be phytoremediated using five mechanisms ofphytoremediation.
3.2.1. Phytoextraction/phytoaccumulationIn this process, plant roots absorb contaminant alongwith other nu-
trients and water and store them in various parts of the plants (Fig. 2a).Plants can be divided into four groups according to their ability tophytoextract As. Plants storing metals upto 1% of their dry weight istermed as hyperaccumulator. Indicators and accumulators are thoseplantswhose uptake and translocation reflectmetal concentration in in-terstitialwaterwith andwithout showing toxic symptoms, respectively.Excluders are plants who restrict the uptake of toxic metals (Baker,1981). A breakthrough step for As phytoremediation program was thediscovery of the As hyperaccumulator fern P. vittata by scientists fromthe University of Florida (Ma et al., 2001). This common brake fern,known popularly as Edenfern, presents an extraordinary capacity tophytocollect 2.3% As in its biomass and stores 93% of it in the fronds(Ma et al., 2001; Tu andMa, 2002). It soaks up As 200 times concentrat-ed than the surrounding soil. Although hyperaccumulation is not re-quired for plant metabolism, natural selection may have selected thisfeature as a beneficiary role for the plants. Plants can use the ability asa defensive strategy to kill pathogens and keep away the herbivores ora survival strategy to avoid competition from metal nontolerant plantsand survival in metal polluted areas. A recent study by Ye et al. (2011)has shown that phytoremediation by P. vittata markedly reduces Asconcentration in rice from As contaminated paddy soil in a pot experi-ment. The study also observed that DMA (dimethylarsinic acid) contentbecame the minor species or undetectable after phytoremediationwhich was the dominant form in untreated sample.
Recently researchers are focussing on exploring the As accumulatingabilities of native plants and they have identified several As accumulat-ing/hyperaccumulating native plants (Castillo-Michel et al., 2011;Antosiewicz et al., 2008; García-Salgado et al., 2012; Tripathi et al.,2012). Although As accumulating plants accumulate less As but the ul-timate goal can be achieved in less time owing to their high biomas. Italso reduces invasion fear by exotic fern species and reduces cost.Brassicas, are very good candidate plants for hyperaccumulation of Asand other heavy metals (Pickering et al., 2000). Some recent studieshave found maize, the most widely planted crop, has good prospects inphytoremediation (Requejo and Tena, 2005; Ding et al., 2011). Through

186 M. Roy et al. / Environment International 75 (2015) 180–198
a long-term study, Niazi et al. (2012) calculated the phytoremediationefficiency and time needed for complete phytoremediation of As bytwo hyperaccumulating ferns P. calomelanos var. austroamericana andP. vittata. They calculated that both the above two fern species wouldtake 6–15 years time to reduce level of As in soil below the EIL (ecologicalinvestigation level) value (20 mg kg−1).
3.2.2. Phytostabilization/phytoimmobilisation/phytorestorationPhytostabilization or phytorestoration is the use of the extensive
root system of plants with its associated rhizobiome to hold the pollut-ant at the site, reducing it's leaching to groundwater andwindblown airexposure due to soil erosion (Fig. 2b). It is also useful in infiltration con-trol i.e. preventing significant amount of rainwater percolating into thegroundwater. The target of phytostabilization is not the removal of Asbut rather stabilization of As at the site and reduce the risk of its expo-sure. Ideal As tolerant plants for phytostabilization not only give goodvegetation cover of the polluted soil by possessing an extensive root sys-tem immobilizing the contaminant in the rhizosphere but also poorlytranslocate As from root to shoot reducing food chain transfer risk.Selected candidate plants should be easy to grow and establish densecanopies and root systems quickly. The vegetation ground coverprovides hydraulic control suppressing the lateral and vertical transferof pollutants into the groundwater (Berti and Cunningham, 2000;Cunningham et al., 1995; Cunningham and Ow, 1996; Schnoor, 2000).Phytosequestration also prevents migration of the contaminant to soil,water and air by phytochemical complexation of themetal contaminantwith different root exudates. Phytochemicals exuded into the rhizo-sphere causes precipitation of the contaminants in the root zone.Some transport proteins associate themselves with the exterior rootmembrane irreversibly and stabilize As by binding to it on the root sur-faces. Others facilitate transfer and safe storage of As into the vacuoles ofroot cells preventing translocation to the shoot and above ground parts.Phytostabilization is the interim strategy where immediate decontami-nation strategies cannot be launched to clean vast areas of As-pollutedsites due to the lack of remediation funds or other reasons. Before begin-ning plantation, the contaminated soil should be prepared with a seedbed with lime, fertilizer, or other chemical amendments to fix As. Themost appreciable soil amendments for fixing As are phosphate fertil-izers, organic matter, iron or manganese oxyhydroxides, clay particles,and mixtures of these amendments. Arsenic is generally adsorbed bysoil cations such as iron, aluminum and calcium, forming insolublesalts. This immobilization occurs more frequently in clay or organicsoils rather than sandy soils where bound As is prone to movement byerosion of soil particles.
3.2.3. Phytovolatilization/rhizovolatilizationAnother method of phytoremediation of metal involves the use of
wild type or genetically modified plants which alone or in associationwith rhizospheric microbes strip As from soil, biologically convertthem to volatile forms within the plant, and release them into the at-mosphere. Although volatilization of As from natural environments(Frankenberger and Arshad, 2002) has been reported, but percent-age of rhizomicrobes contribution towards the total fraction is notknown. Information available on volatilization of As for soil suggeststhat in the absence of plant roots, volatile compounds account onlyfor small proportions of total As (Turpeinen et al., 1999).
Phytovolatilization is the principle mechanism behind ecologicalrestoration of As contaminated land by bioenergy crop Arundo donaxL. (Mirza et al., 2011). Arsenic accumulation order in different plantparts of A. donax L. was: Leaf As(V) N Shoot As(V) N Root As(V) N Solu-tion As(V) solution As(III) Leaf As(III) N Shoot As(III) N Root As(III).A. donax L. volatilized 7.2–22% As at supplied As concentration of300–1000 μg L−1. Volatilization was theway to avoid toxic effects of As.
Several experiments have shown that P. vittatta also possesphytovolatilization activity towards As. The vapor released fromthe frond of P. vittata was found to contain both As(III) and As(V).
Sakakibara et al. (2010) found that the fern removed a maximumratio of 90% of the total As from As-contaminated soils in greenhouse,where the environment was similar to the subtropics. However, if alarge amount of arsenic had been released from the contaminated siteinto the atmosphere by the fern, the processmay have caused a second-ary As-contamination to the surrounding environment.
But in general although phytovolatilization is quite promising forsome metal remediation like Se and Hg it is not suitable for As becausesome gaseous forms of As are very toxic. Recent studies have shown thatsomemethylated volatile intermediates like monomethylarsonous acid(MMAIII) and dimethylarsinous acid (DMAIII) are more toxic than theinorganic arsenic species (Petrick et al., 2000). Dimethylarsine andtrimethylarsine are about 100 times more potent genotoxic than mostgenotoxic non-volatile arsenical dimethylarsinous acid (Andreweset al., 2003). So in light of the toxicity of MMA(III) and DMA(III) andtheir identification in human urine a rethinking is necessary on thismode of detoxification.
3.2.4. Phytofiltration/rhizofiltrationRhizofiltration is afiltrationmethod that uses plants to remove pollut-
ants fromwater. In rhizofiltration plants are raised in greenhouses hydro-ponically i.e. roots suspended in water containing essential minerals andthen transplanting them to As containing water where the plants absorband store As in their roots (Rahman and Hasegawa, 2011; Dushenkovet al., 1995; Favas et al., 2012). Arsenic hyperaccumulating moonlightfern and brake fern were found successful when tested hydroponicallyfor their phytofiltration abilities (Baldwin and Butcher, 2007). In compar-ison to conventional cultivation methods, hydroponics allows bettermonitoring and characterization of pollutant uptake as potential complexinteractions of soil componentswith target elements can be avoided. Rootexudates cause change in rhizosphere pH which in turn cause As to pre-cipitate out onto root surfaces. As the roots are saturated with As theyare harvested for disposal. Controversy persists regarding whetherrhizofiltration should select plants that would exclusively store As inroots and not in shoots or those plants that would store As in both rootand shoot compartments with equal efficiency. The first case wouldallow efficient disposal of As (Dushenkov et al., 1995)whereas the secondcategorywould allow increased rhizofiltration ability (Zhu et al., 1999). Inspite of this difference in opinions, it is apparent that ideal plants selectedfor rhizofiltration should have the ability to tolerate and accumulatesignificant amounts of As in conjunction with large root biomass or rootsurface area, lowmaintenance cost, easy handling and generation ofmin-imum secondary waste. Several aquatic plants have the ability to removeAs from water, including aquatic macrophyte Eleocharis macrostachya(Olmos-Márquez et al., 2012), Hydrilla verticillata (L.f.) Royle (Xueand Yan, 2011), Phragmities australis (Ghassemzadeha et al., 2008),W. globosa (Zhang et al., 2012), and water hyacinth (Eichhorniacrassipes) (Alvarado et al., 2008). Among many other aquatic macro-phytes (for complete list see review by Rahman and Hasegawa, 2011),water hyacinth (E. crassipes), duckweeds (Lemna gibba, Lemna minor,Spirodela polyrhiza and Wolffia), water spinach (Ipomoea aquatica),water ferns (Azolla caroliniana, Azolla pinnata and Azolla filiculoides),water cabbage (Pistia stratiotes), hydrilla (Hydrilla verticillata) and wa-tercress (Nasturtium officinale, Nasturtium microphyllum) have beenproposed to be ideal agents for aquaphytoremediation due to their Ashyperaccumulation ability and growth behavior. However, most aquat-ic plants have inherent drawbacks due to their small, slow-growingroots and high water content in their cells complicating the disposalprocess of drying, composting, or incineration. Terrestrial plants arecurrently under investigation as alternatives for rhizofiltration becausein contrast to small roots of aquatic plants, their longer, strong, fibrousroot systems give large surface areas for increased metal sorption. Inanother development, Rofkar and Dwyer (2013) took an initiative totreat As laden water through engineered wetland system. Here theyscreened several native wetland plants in hydroponic and identified

187M. Roy et al. / Environment International 75 (2015) 180–198
on two plants C. stricta and S. pectinata as suitable candidates for theengineered wetland system.
3.2.5. Phytostimulation/natural attenuationNatural attenuation (NA) or in situ clean up is nature's own way of
remediating a polluted site. The naturally occurring slowmoving intrin-sic processes can be accelerated for designing an environmentally safecost effective treatment strategy. Natural attenuation of As mainly in-volves immobilization method through sorption to solid phases. Hy-droxides of aluminum, clay, iron, manganese, sulfide minerals, andnatural organic matters are the significant As adsorbents. Arsenic speci-ation status and on site geochemical characteristics including pH, redoxpotential, and co-occurring ions effect the rate of sorption. Microbialtransformations and hyperaccumulation also constitute a part ofNA. Addition or removal of certain factors from soil would enhancephytoavailability of As. For example the addition of P increases theAs phytoremediation capacity of Anadenanthera peregrina plants byincreasing As accumulation and alleviating As-induced oxidativestress (Gomes et al., 2013). An elaborate discussion on different pro-cesses of natural attenuation of As would be found in the review byWang and Mulligan (2006).
4. Phytobial remediation
Phytobial remediation combines bioremediation andphytoremediation both to reduce the level of target contaminant.Fig. 3 shows howmicrobes and plants take part in natural geochemicalcycle of arsenic. Soil inhabits largest number of microbes on earth. Soilmicrobes are the key players of ecosystems; they drive major
Fig. 3. Role of microbes and plants in the flow of arsenic through the biosphere. Red colored arshow pathways of arsenic that are dominant in arsenic hyperaccumulating plants; violet arrowsplant roots (ideal phytoimmobilizer) and arsenic hyper accumulators. Blue arrows show reactierences to color in this figure legend, the reader is referred to the web version of this article.)
biogeochemical processes, regulate plant productivity, and recycle nu-trients (see review by Heijden Van Der et al., 2008). Soil microbes en-hance plant productivity in nutrient deficient ecosystems byproviding growth-limiting factors N and P to plants. They may alsodecrease plant productivity by acting as pathogens and competingwith plants for nutrients. They may also transform nutrients into inac-cessible forms decreasing productivity or into accessible forms increas-ing productivity to plants. In addition microbes play key roles in Asglobal cycle along with other elements of biosphere participating inrock weathering, mineral dissolution and biotransformation reactions.In view of the active role of soil microbiome on ecosystem, Asphytoremediation program cannot be successful without knowing theindigenous microbial flora predominating in that area. Particularly ifthe region has large deposits of As containingminerals, As dissimilatorybacteria would be actively engaged in obtaining energy from such com-pounds and doing so would release a huge portion of bioavailable Asagain (Ahmann et al., 1994; Malasarn et al., 2004; Islam et al., 2004).So a test site cannot be declared As free if that site is rich in As bearingminerals. The site shouldmaintain a perfect balance between its dissim-ilatory arsenate reducing microbial flora and a combined phyto-mi-crobial flora that fishes/scavenges out the bioavailable As as soonas it is released. The process of As release and its consumptionwould continue until all the As laden mineral resources continue tofeed the microbes. Once all the sequestered As comes in the solutionand invasion of new As species from xenobiotic sources can beprevented a site can be declared As free. So in a natural systemboth microbes and plants play intricate role in restoring its environ-ment against the toxic effect of pollutants including As. In a reviewRahman et al. (2014) has shown role of both prokaryotes and
rows display events favoring arsenic mobilization. In the plant cell, brown colored arrowsshow reactions that are common for both arsenic accumulator plants that accumulate it toons that are dominant in plants for phytosequestration only. (For interpretation of the ref-

188 M. Roy et al. / Environment International 75 (2015) 180–198
eukaryotes as bioorganisms in As removal process. In this review weare showing the individual role and mechanism of free living,rhizomicrobes and endomicrobes separately and their interactionwith plants in the As removal process. Among the soil microbes, rhi-zosphere associated microbes is termed as rhizobiome and the rootassociated mycorrizha forms a special subcategory termed asmycorhizobiome. Endophytobiome is another term that denotesthe entire microbiome that resides within plant. The rest of the mi-crobes in the bulk soil have been simply termed here as free livingsoil microbiome. Very recently relationship of microbial communi-ties and functional genes and their synecologic interactions withthe biogeochemical characters of an As contaminated vertical soilgradient were analyzed (Xiong et al., 2012). Significant changeswere observed with important genes contributing to As resistance,phosphorus utilization, carbon and nitrogen cycling. RbcL and nifHgenes were selected as biomarkers to show how anaerobic life tookthe position of aerobic life with increasing depth (Xiong et al.,2012). An integrated model of the metabolic interactions takingplace in an As rich acid mine drainage was built up and the role of in-dividual microorganisms residing in that complex community wasdeciphered by Bertin et al. (2011). These metagenomic studies notonly provided important insights into the heterogeneity of microbialcommunity metabolic potential but also enriched our understandingof natural attenuation mechanism and future biodiversity preserva-tion for As bioremediation. Culture dependent study is also impor-tant as it allows full characterization of the individual isolates andidentifies novel bacterial species/genes (Delavat et al., 2012).
4.1. Phytobialremediation by free living microbes
Free-livingmicrobes not only have immense role on the partitioningof different species of As in soil, water, and air but they regulate Asuptake by plants in differentways. Their assistance in phytoremediationof As by immobization/mobilization/transformation/sequestration/volatilization reactions are discussed here. In some nutrient poorcontaminated sites application of microbes may provide plants vitalnutrients.
4.1.1. MobilizationMicroorganisms mobilize metals through a variety of reactions in-
cluding redox transformations, volatilization, leaching and chelation.Bacteria respiring on As cause mobilization of As in anoxic sub-surfaceaquifer by oxidizing organic matter present in the aquifer and useAs(V) or Fe(III) as the terminal electron acceptors during the process(see Fig. 1). The DARPs mobilize As by either reductive release ofAs(III) and Fe(II) (e.g. iron-reducing DARP Sulfurospirillum barnesii orthrough release of adsorbed As from the surfaces of either Fe(III) orAl(OH)3minerals via reduction of As(V) to As(III)). Iron-reducing bacte-ria, such asGeobactermay cause the release of As(V). Iron respiring bac-teria reduces Fe(III) to Fe(II), that has a lower binding affinity to As(V).The released As(V) may be reabsorbed to other minerals. If this processtakes place in the presence of DARPs or if the iron respiring bacteria areDARPs (as many bacteria can use multiple electron acceptors), As(III)will be released instead of As(V). Organic matters present speed upthis mobilization process by supplying nutrients to these microbes(Dobran and Zagury, 2006). It is important to understand that amongthe arsenate reducing microbes only the DARPs can reduce both aque-ous and solid phase As(V) because the arr gene is located either freelyin periplasm or as membrane bound periplasmic protein with catalyticsite in the periplasm. But microbes possessing arsC as free cytoplasmicprotein reduce only aqueous As(V) that has entered the cell but cannotreduce solid phase As(V). Shewanella sp. strain ANA-3, possessing bothtypes of reductases (Saltikov and Newman, 2003; Saltikov et al.,2003), employ only Arr gene in reducing solid-phase As(V) (Malasarnet al., 2004). Yamamura et al. (2008) investigated bioremediation poten-tial of a DARB, Bacillus selenatarsenatis SF-1 recovering As-contaminated
soils by reductive dissolution of As. They also observed that removal ofAs from the contaminated soil can be enhanced by the co-presence ofanthraquinone-2,6-disulfonate (AQDS) along with B. selenatarsenatiswhich concomitantly releases Fe(II). This suggested that combined useof DARP and electron-shuttling compounds would be effective in biore-mediation of As-contaminated soils. Lee et al. (2009) further developeda noble hybrid method by combining microbes mediated anaerobicbioleaching and electrokinetics together. Previously laboratory and fieldbased studies has shown electrokinetics as a promisingmethod for simul-taneously recovering multiple metal contaminants. But in case of As, themobility and removal efficiency of Aswas slower than other heavymetals(Wang et al., 2005; Virkutyte et al., 2002). So he speeded up As removalperformance of electrokinetics by combining it with microbial metal re-duction in the anaerobic bioleaching. An integrated phytoremediationprogram should include addition of more metal reducing bacteria to theeffected site which would cause enhanced mobilization of solid phaseAs to more bioavailable As. Plants selected for phytoextraction would ac-cumulate this As into harvestable tissue which would be disposed later.All themicrobialmediatedmobilization reactions except volatilization in-creases the phytoavailable pool of As. So inoculation of As mobilizing mi-crobes in As contaminated sites would speed up the process ofphytoextraction and hence phytoremediation.
4.1.2. Immobilization and bioaccumulation/biosequestrationIn contrast to mobilization, immobilization technique is used to
decrease the mobility of contaminants by changing the physical orchemical attributes of the contaminant. Although mobilization and im-mobilization are antagonistic to each other, both contribute to Asphytobialremediation in their own ways. Microbes play an immenserole in As immobilization since the widespread microbial arsenite oxi-dase enzyme (possessed by HAO and CAOs) oxidizes more mobile andtoxic As(III) to less toxic and lessmobile As(V)which is readily absorbedby various metal oxides and removed by physico-chemical methods(Leist et al., 2000). Bioremediation potential of various As(III) oxidizingmicrobes has been tested either in bioreactor based system (Dastidarand Wang, 2012) or laboratory based assay (Bahar et al., 2012). Micro-bial biomass also acts as a metal sink, by intracellular sequestration, orprecipitation of metals. A high As(III) resistant bacterium Sporosarcinaginsengisoli CR5was investigated for microbially induced calcite precip-itation process for bioremediation which in the future may stand as apromising, environmental friendly technology for remediation of theAs contaminated sites (Achal et al., 2012). Arsenic immobilization bysulfate reduction is awell-knownphenomenon. Sulfate-reducing bacte-ria produce sulfide which reacts to precipitate As. Keimowitz et al.(2007), showed that this phenomenon could be utilized at landfill andother sites where As mobilization is a problem. While some eukaryoticmicrobes like Candida glabrata metabolizes extracellular sulfate tosulfide (Thomas and SurdinKerjan, 1997), that acts as an electrondonor for As(V) reduction (Rochette et al., 2000), other yeast likeSchizosaccharomyces pombe (Dameron and Winge, 1990) andC. glabrata (Krumov et al., 2007) incorporates sulfide to form a morestable, highmolecular weight PC–metal–sulfide complex in the vacuole(Kneer and Zenk, 1997; Mendoza-Cozatl et al., 2006). Some microbes(e.g. Herminiimonas asoxydans) produce extracellular polymeric sub-stances, which help in the immobilization of metals (Muller et al.,2007). Another strategy of As removal by immobilization is via bio sorp-tion reaction. As the name suggest it involves the use of biological ma-terial for the removal of contaminating substances from a solution.Both living and dead bacteria, microalgae, and fungi can take part inbiosorption reaction, as it is a physio-chemical process (Gadd, 2010).Living cells of Bacillus cereus (Giri et al., 2012), native and chemicallymodified biomass of Aspergillus niger and Neosartorya fischeri (Litteraet al., 2011) and dried biomass of Spirogyra hyalina (Kumar andOommen, 2012) were tested as bioadsorbents for treating As contami-nated water. Microbial mediated immobilization should be applied tosites selected for phytosequestration. These microbes could also be

189M. Roy et al. / Environment International 75 (2015) 180–198
used to develop biofilm-based reactors (Chang et al., 2006) useful in re-moving As from contaminated waters (Lieveremont et al., 2009). Sitesthat contain phytotoxic concentration of As, such As-sequestering mi-crobes would be helpful for reducing the As concentrations andmakingthe sites habitable for plants that can carry out phytoremediation.
4.1.3. Biotransformation/volatilizationA large number of bacteria, fungi and algae are capable of As bio-
methylation (Huysmans and Frankenberger, 1991; Frankenbergerand Arshad, 2002). Microbial biomethylation of As, a detoxificationpathway was previously sought to have bioremediation potential(Turpeinen et al., 1999, 2002; Bentley and Chasteen, 2002). Howeverrecently it gained controversy as the mono and dimethylated arsinesare very genotoxic and the DNA damaging potency follows the orderDMA(III) NMMA(III) N (AsV/AsIII) NMMA(V) NDMA(V) N TMA(V) (Masset al., 2001). The volatile arsines in air have short life span and their quickoxidation again contribute to soil andwater pool of As(III)/As(V). In addi-tion to biotransformation manymicrobes have been found in nature thatbioaccumulate large amount of As (Takeuchi et al., 2007; Joshi et al., 2008;Xie et al., 2013a, Tripathi et al., 2007).
4.2. Phytobialremediation by rhizosphericmicrobes/phytorhizoremediation
Rhizosphere is the microecological zone surrounding plant roots.The roots release a number of compounds (exudates, secretions, muci-lages, mucigel, and lysates) in the rhizosphere establishing a highly dy-namic and active bacterial community distinctly different from the bulksoil microbial community. While plant exudates are low molecularweight surplus sugars, amino acids, and aromatics, secretions are thelow/high molecular weight by-products of plant metabolism. Lysatesare mixtures of cellular enzymes from lysed root cells. Mucilages arecells sloughed off from the root cap andmucigel is a gelly like substance(slime) that coats the surface of a root. It increases contact betweenplant roots and the surrounding soil and prevents dehydration duringdry spells. The root system aerates the soil, provides a steady-stateredox environment and a starting material for colonization of plantgrowth promoting rhizobacteria (PGPR). PGPR directly enhance plantnutrition and growth by solubilization of essential minerals like phos-phorus (P). Indirectly it helps plant growth by preventing growth ofphytopathogenic organisms. Some phosphorus starving plants showan increased exudation of carboxylic acids, like citric and malic acidwhich make soil more acidic favoring chelation of metal cations andsubsequent formation of soluble metal-chelate conjugates with P. Thisresults in enhanced phytoavailability of P (Kirk et al., 1999). Someplantsalso excrete protons and/or reducing and chelating compounds causingco-dissolution of As from Fe-oxides/hydroxides and render Asmore sol-uble and bioavailable to plants. Some PGPR are active nitrogen fixingmicrobes while others synthesize siderophores that solubilize andsequester iron from soil. Some accelerate plant growth by producingdifferent phytohormones (e.g. auxins and cytokinins). Some microbesunder iron-starved conditions secrete siderophores, which are smallmolecular weight organic compounds to chelate and solubilize iron. Var-ious siderophores possess various ligand binding groups like catecholate,phenolate or hydroxamate and although they are specific ferric iron che-lator, but canbind to othermetals includingAs. Siderophores producedbyPseudomonas azotoformans, were shown to act as a washing agent likeEDTA and citric acid to mobilize and thus remove As (oxyanion) fromcontaminated soil (Nair et al., 2007). Some acidifuge plants secrete oxalicand citric acidwhich effectivelymobilize P and Fe from lime stone (Strömet al., 1994). Root vicinity normally exhibits aerobic condition and hencean increase of rhizosphere pH favour mobilization and uptake of As(V).Application of nitrogen fertilizers increases As uptake as N2 affectsrhizosphere pH, due to its participation in cation/anion uptake ratio.However legumes and actinorhizal plants play different roles in rhizo-sphere acidification. Legumes produce root nodules housing N2 fixingmi-crobes. Rhizosphere acidification by the N2-fixing symbionts favors
As(V) immobilization. So rhizoremediation of As is accomplished by ei-ther of the three ways, stimulation of plant growth by accelerating plantnutrition, making it bio-unavailable to living organisms by immobiliza-tion of the metal in the rhizospheric zone and/or enhancement of metalaccumulation in the plant. Sometimes special structures called ironplaques are encountered in the rhizosphere of wetland plants due to pre-cipitation of iron oxyhydroxides on the root surface (Hansel et al., 2002).The iron plaques have strong affinity for arsenate sequestration reducingit's bioavailability to root cells (Chen et al., 2005). In ageochip based anal-ysis, Xiong et al. (2010) found more abundant distribution of genes re-sponsible for denitrification, phosphorus utilization, sulfur reduction,and As resistant genes in the rhizosphere of P. vittata. Wang et al.(2012) observed microbial As oxidation and reduction in rhizosphere ofP. vittata L. and they isolated strains that exhibited dual ability for bothAs(V) reduction and As(III) oxidation. In another study by Yang et al.(2012), the application of As(V) reducers was shown to increase plantbiomass and total concentration of As in the fronds in comparison tocontrol. So As may be made more accessible to plants by design of aplant–microbial consortium having the capacity to mobilize metals byregulating rhizosphere pH or release of biosurfactants or ligand exuda-tion. An integrated approach of rhizosphere management combiningco-cropping, improved agronomic practices, and soilmanagement shouldbe undertaken to face the challenge of complex and heterogenous multi-ply polluted “real world” soils. Fitz and Wenzel (2002) and Wenzel(2009) reviewed the role of rhizobacteria in phytoremediation of Asand other heavy metals.
4.3. Phytobialremediation by endophytic microbes/endophytoremediation
An endophyte is a bacterium or fungus that lives endosymbioticallywithin a plant. It occasionally invades the plant from rhizosphere, orphyllosphere or seed and then colonizes vascular tissue of the plantfor at least a part of its life without harming the host (Ryan et al.,2008). Endophytes are ubiquitous on earth and almost all the 30,0000species of land plants existing on earth have been found to host one ormultiple endophytes (Strobel et al., 2004). They are important parts ofplant microbiome comprising epiphytes, mycorrhizal fungi, pathogens,and saprotrophs. All of them participate in plant growth and responsesto herbivores, pathogens, and environmental changes like droughts andflood in one or the other way. Some fungal endophytes produce usefulsecondary metabolites. This endophytobiology demands more researchso that the huge potential of these relatively unstudied endophytobiomecan be fully exploited (see review on fungal endophytes by Porras-Alfaroand Bayman, 2011). Although the natural ability of endophytes to de-grade various xenobiotics has been applied for phytoremediation ofmany mono- and dichlorinated benzoic acids (Siciliano et al., 2001), vol-atile organic compounds (Barac et al., 2004), andmono and polycyclic ar-omatic hydrocarbons (Germaine et al., 2006, Aken Van et al., 2004;Taghavi et al., 2005) but only scarce reports are present reporting endo-phytes in As tolerance. Dourado et al. (2012) first time reported the isola-tion and characterization of Methylobacterium strains from mangroveplants showing tolerance to various heavy metals including As. Isolationof an As-resistant bacterium, AsRB1 from the phyllosphere (fronds) ofP. vittata (Rathinasabapathi et al., 2006) is encouraging to concentratemore study in this relatively neglected endophytobiology of As.
4.4. Phytobialremediation by mycorrizal fungi/phytomycoremediation
Majority of land plants have live-in-together relationshipwithmanytypes of mycorrhizal fungi. In this mutualistic relationship, the mycor-rhizal fungal hyphae help the plant to get more water and nutrientsby increasing the surface area of root and giving access to the smallestpores of soil that root tips cannot access (Sylvia et al., 2005). In return,the fungi receive food from the plant. Ectomycorrhizae produces a netlike structure called Hartig net that interacts with the plant root byweaving between the root cortical cells. Mycorrhiza that interacts

190 M. Roy et al. / Environment International 75 (2015) 180–198
with the plant root by producing a branched arbuscule that growswithinthe root cortical cell and produces a direct connection between the plantand fungal cytoplasm is referred to as endomycorrhiza or arbuscularmy-corrhiza (AM) (Sylvia et al., 2005). The significance of AM fungi in soilbioremediation study recently got attention (reviewed by Gohre andPaszkowski, 2006; Khan, 2005) because of its potential to enhancephytoextraction by hyperaccumulating ferns and phytostabilizationby rooted vegetation. In the presence of As tolerant AM fungi, Ashyperaccumulation ability of P. vittata increases significantly as they as-sist the plant for enhanced growth and As uptake by increasing root sur-face area, chlorophyll content and As translocation factor (Trotta et al.,2006). Leung et al. (2006) noted increase in As concentration in thesoil caused an increase in mycorrhizal infection in P. vittata. Agely et al.(2005) observed that in the presence of AM fungi As accumulation infrond increased several folds. Increased P transporters in the hyphae en-hanced As(V) uptake. However the mycorrhizal association of crop spe-cies growing in As contaminated areas contributes to the enhancedaccumulation of As since the fungi mediated enhanced acquisition ofphosphate is accompanied by enhanced uptake of As. Some AM fungiisolated from Brake Fern are Glomus mosseae, Glomus geosporum andGlomus etunicatum (Wu et al., 2009). Orłowska et al. (2012) investigatedthe role of several indigenous and non-indigenous arbuscular mycorrhi-zal fungi (AMF) on Plantago lanceolata L. growing on As rich mine wasteand he observed higher shoot and root biomass in P. lanceolata inoculat-ed with AMF than the non-inoculated plants.
5. Strategies for enhanced phytoremediation
Phytoremediation efficiency of native plants/ferns can be enhancedby the combination of other strategies like the following.
5.1. Transgenic technology
Extensive technological development is needed to bring down Aslevel in drinking water below 10 ppb. Transgenic technology creates anew generation of genetically modified organisms that brings newhope to boost up natural bioremediation potentials of plants and mi-crobes and customizing the plant microbe mutual relation for betterbioremediation of As.
5.1.1. Transgenic microbesThe bioremediation potential of microbes can be harnessed by ge-
netically equipping them with weapons to fight against As better thanwild type cells. As discussed earlier the bacterial As resistance genesform ars operon confering resistance to arsenicals. In the upstream re-gion of the promoter and coding region of ars operon lies one arsRgene that codes for one regulatory protein that specifically senses onlyAs(III). Kostal et al. (2004) overexpressed ArsR in recombinant E. colicells which accumulated 13- to 60-fold higher levels of As(III) andAs(V) than control cells. They removed 98% of 50 ppb As(III) from con-taminated water. These cells were able to accumulate more As thanthose cells that accumulates As by producing PC (Sauge-Merke et al.,2003). Singh et al. (2008) constructed an engineered E. coli strain thatexpressed fMT from Fucus vesiculosus (Merrifield et al., 2004). MTs(metallothioproteins) are members of cysteine-rich protein familythat form stable bondswithmetals.MTs that bindwith As have been re-ported in the alga F. vesiculosus (Merrifield et al., 2004), but none havebeen isolated in bacteria. When Singh et al. (2008) overexpressedAs(III) specific transporter GlpF with fMT, the engineered E. coli concen-trated high levels of intracellular As. Recent advancement of genomeshuffling enabled the creation of a customized arsenate detoxificationpathway in E. coli (Crameri et al., 1997). Recombinants containing theoptimized operon displayed 40 fold higher resistance than control. Inanother study, S-adenosylmethionine methyltransferase gene (arsM)gene, responsible for biomethylation of As(III) to the comparativelyless toxic pentavalent species DMA(V) and TMAO, was identified in
Rhodopseudomonas palustris (Qin et al., 2006). This biomethylationgene arsMwas cloned in E. coli and verified for its ability to produce vol-atile trimethylarsine from inorganic As (Yuan et al., 2008). Overexpres-sion of arsM in Sphingomonas desiccabilis and Bacillus idriensis causedthe production of methylated As gas 10 times higher in comparison towild type (Liu et al., 2011). Chauhan et al. (2009) made a metagenomiclibrary froman industrial effluent treatment sludge and there they iden-tified a novel As(V) resistance gene (arsN) encoding a acetyltransferaselike protein. Overexpression of that ArsN caused higher resistance of Asin recombinant E. coli cell.
5.1.2. Transgenic rhizomicrobesThe transgenic technology can also be exploited to construct a better
micro phyto mutualistic association. Brazil et al. (1995) constructed anengineered PCB degrader Pseudomonas fluorescens F113rifPCB whichprotects the plant by serving as an effective rhizosphere colonizer. Thestable insertion of ars operon into the recombinant strain allowed it tosurvive in the soil having inhibitory concentration of As and carryout its bioremediation activity (Ryan et al., 2007). PGPR having ACC(1-aminocyclopropane-1-carboxylic acid) deaminase activity lowersstress-induced ethylene levels in plants and increases tolerance forheavy metals, fungal phytopathogens and flooding (Stearns and Glick,2003). Transgenic canola plants Enterobacter cloacae CAL2 expressingE. cloacae UW4 ACC deaminase accumulated approximately four timesmore As(V) in comparison to un-transformed canola (Nie et al., 2002).In the presence of As(V), ACC deaminase hydrolyzes excess ACC lower-ing the inhibitory level of ethylene in seeds and enhances the process ofgermination.
5.1.3. Transgenic plantsIntroduction of transgenic technology opened up new hope of
As phytoremediation through the introduction of new transgenicplants with enhanced As removal capacity from soil or transgeniccrops with enhanced ability to restrict uptake of As from soil.Genes have been identified that encodes As uptake channels andtransporters in As accumulator and hyperaccumulator plants.So through inactivation or deletion of the genes correspondingto different variants of phosphate transporters, NIP aquaporins,and Lsi2-like carrier proteins, it is possible to block the entry ofAs(V)/As(III) into roots. Complex formation with ligands such asphytochelatins (PCs), metallothioneins (MTs) or glutathione (GSH)and further sequestration or compartmentalization of the complexesin specialized tissues like trichomes or vacuoles is a detoxificationprocess. So genetic engineering for As phytoremediation triggerseither enhanced As sequestration in root (rhizofiltration) or en-hanced hyperaccumulation of As in shoot/frond (phytoextraction)or phytovolatilization pathways. Root surface area and root biomassfor enhanced rhizofiltration can be attained by induction of hairyroot phenotype (Eapen et al., 2003; Nedelkoska and Doran, 2000).Hyperaccumulation could be enhanced by targeting transportergenes and PC synthesis genes responsible for increased translocationto shoot and increased storage to vacuoles, respectively.Metal tolerancecan also be increased by causing hyper expression of oxidative stressrelated genes (Foyer and Noctor, 2005). Another target would be thegenes that cause As methylation. Transgenic rice plants have beencreated that methylate and volatilize As (Meng et al., 2011). Fear offood chain contamination can be addressed by novel biotechnologicalapproaches like creation of designer plants that would phytoextractand accumulate large amounts of As and at the same time would resistAs food chain transfer by expressing some toxin genes or conditionallethality genes that would distract the herbivores and resist pest attack.
Zhu and Rosen (2009) have recently reviewed progresses made intransgenic field. Here we have highlighted the major successes madein this field. Dhankher et al. (2002) overexpressed arsenate reductase(ArsC) and gamma-glutamyl cysteine synthase (λ-ECS) genes togetherinto the model plant Arabidopsis with the objective of creating

191M. Roy et al. / Environment International 75 (2015) 180–198
transgenic plants with increased As accumulation ability. While ArsCmediates reduction of As(V) to As(III) in the stem and leaves, γ-ECS cat-alyzes the first step in the phytochelatin synthesis pathway increasingthe pool of phytochelatin like thiol compounds throughout the plants.As a result, the transgenic plants accumulated higher amounts of As inthe leaves than the wild types. They expressed arsC under a light-inducible promoter, to ensure arsenate reduction occurs always in theshoot and never in the roots. Thus, As(V) in the transgenic plantsmoves only uni-directionally (from root to leaf) before being capturedby the arsC or λ-ECS. Singh et al. (2008) cloned and expressedA. thaliana phytochelatin synthase (AtPCS) gene into the yeastS. cerevisiae and the transgenic yeast showed enhanced As accumula-tion. When Guo et al. (2008) cloned and overexpressed a λ-glutamylcysteine synthetase (λ-ECS) gene of S. cerevisiae intoA. thaliana, it showed elevated tolerance and accumulation of As. As astep forward to construct transgenic rice resistant to As, Duan et al.(2012) cloned the As(III) extrusion pump (ScAcr3p) of Saccharomycescerevisiae into rice. As a result, roots of these transgenic plants effluxedAs many times higher than normal wild type. It is known that sulfidepresent in soil decreases the toxicity of As by creating metal precipitates.Some eukaryotes also have taken this strategy by incorporating sulfideand forming a high molecular weight PC–metal–sulfide precipitation inthe vacuole (Mendoza-Cozatl et al., 2006). However in the bacterial king-dom hydrogen sulfide production is only restricted to anaerobic sulfate-reducing bacteria that produce H2S only under strict anaerobic condition.To overcome the limitation, Wang et al. (2001) took cysteinedesulfhydrase (CysDes) gene from Treponema denticola and expressedit in E. coli for aerobic sulfide production using cysteine as the substrate.Cysteine desulfhydrase gene encodes for an aminotransferase enzymethat produces hydrogen sulfide from cysteine under aerobic condition.Recently Tsai et al. (2012) co-expressed cysteine desulfhydrase andAtPCS genes together in S. cerevisiae to increase the level of intracellularAs through the formation of stable PC–As–sulfide complexes. Yeast cellscontaining both AtPCS and CysDes showed elevated level of intracellu-lar As accumulation, confirming the coordinated action of H2S and PCsin the overall bioaccumulation of As. With the objective of aquaticphytoremediation, phytochelatin synthase (PCS) gene from an aquaticsubmerged rootless macrophyte, Ceratophyllum demersum cv. L.(CdPCS1)was expressed in tobacco plants and these transgenic tobaccoplants showed enhanced PC synthesis (Shukla et al., 2012).
However as the introduction of any GMO in field is under strongcontroversy due to the risk of horizontal gene transfer (HGT) towild type organisms, it is likely that the introduction of transgeniccandidates for phytobial remediation would be also under strictscrutiny. However the use of conditional lethality genes or insertionof transgenes into chloroplast DNA may prevent fear of HGT(Davison, 2005). However transgenic bacteria or transgenic endo-phytes would enjoy many relaxations as gene transfer among them-selves (in case of Agrobacterium between plant and microbe) alwayshappen in nature due to conjugation like events and should not beconsidered ‘genetically modified’. For a wider acceptance of trans-genic biotechnology more research should be carried out for under-standing underlying risk and solutions needs to be worked out forrisk mitigation (Linacre et al., 2003; Davison, 2005).
5.2. Other methods
5.2.1. Biostimulation/bioaugmentationNatural attenuation that occurs at a slow pace in nature can be
boosted up by bio stimulation i.e. addition of various biotic factors(microbes) and bioaugmentation i.e. addition of various abioticfactors (chelating agents and others). If phytoimmobilization is thetarget, As immobilizing microbes and abiotic factors that causesimmobilization of As in soils can be introduced. Preservation of anacidic and oxidizing atmosphere is the main requirement to ensuresolid phase sorption of As which is the main factor for long-lasting
immobilization of As. Proper mixture of H2O2 and Fe salts(e.g., FeCl2 and FeSO4) can precipitate As from groundwater. H2O2
helps in the formation of Fe hydroxides, oxidation of As(III) toAs(V), and creates an oxidizing environment to favor As(V) sorption.pH buffering agents should be used to maintain desired pH in therange of 4–7 to enhance As sorption and prevent As remobilization.Biosparging technology and different types of oxidants can be usedto ensure proper mixing of oxygen to the underground soil andwater. Biosurfactant (rhamnolipid, surfactin, and sophorolipid)foam technology can be used to deliver nutrients or exogenousmicrobes into the subsurface soil/water. The application of naturalorganic matter such as humic acids, which are sometimes abundantin many soils, increases Fe bioavailability as they can chelateFefunctioning as electron acceptors. Different types of inexpensive,nontoxic and efficient adsorbents (e.g. amorphous Fe hydroxidesor Fe hydroxides coated sands) are used to remove As from ground-water. This action can be further boosted by feeding Fe-oxidizingbacteria (e.g. Gallionella ferruginea and Leoptothrix ochracea,indigenous in most groundwaters) into the groundwater (bioaug-mentation). Effects of various soil amendments have been testedon P. vittata (Cao et al., 2003; Santos et al., 2008; Tu and Ma, 2003).A minimum P/As molar ratio of 1:2 in soil solution increasesefficiency of plants to remove of As. Addition of calcium (Ca2+)(Liao et al., 2007; Li et al., 2006), selenium (Se) (Srivastava et al.,2009) and nitrogen (N) (Liao et al., 2007) increase As accumulationof P. vittata. Huang et al. (2012) showed organic matters added tosoil significantly enhanced As methylation and volatilization fromthe paddy field. These recommendations should be followed for de-veloping cultivation approaches for efficient phytoremediation of As.
5.2.2. Phytoremediation with electrokineticsIn this technique a low intensity direct current is passed without
disturbing soil architecture between two electrodes placed verticallyin the soil. Current drives separation of organic and inorganicmolecules. Water and heavy metal cations move towards cathodeby electroosmosis and electromigration, respectively. Anions andsmall charged particles like colloids migrate towards the anode byelectromigration. Laboratory scale studies of electrokinetics combinedwith phytoremediation showed good prospect in heavy metal remedia-tion of Zn, Pb,Cu, Cd and As (O'Connor et al., 2003; Aboughalma et al.,2008; Cameselle et al., 2013). However this is an emerging area andmore large-scale validations are needed to further evaluate feasibility ofthis technique as a low cost remediation alternative. DuPont laboratorieshave designed a technology combining phytoextraction with electroki-netics and chelate based in situ soil washing. The combined action wasmore effective than the single techniques alone. Chelates brought moreions into solution, electrokinetics speeded up the migration of relativelyimmobile ions, and properly selected plants provided an enhanced sur-face area for absorption of the ions and their chelated forms.
5.2.3. Permeable reactive barrierPRB technique conceptualized in 1990 is an effective remediation
technology for the treatment of contaminated groundwater. It is asubsurface emplacement of reactive materials or a treatment zonethrough which a dissolved contaminant plume must move as itflows, typically under natural gradient. Treated water exits theother side of the PRB. This in situ method for remediatingdissolved-phase contaminants in groundwater combines a passivechemical or biological treatment zone with subsurface fluid flowmanagement. The contaminated groundwater flows through thetreatment zone of PRB installed in its flowpath. The treatment materialuses immobilized microbes or plants (Köber et al., 2005), zero valentiron (Richard et al., 2008) or nanoparticles to absorbAs fromgroundwa-ter or soil treatment/landfill leachate.

192 M. Roy et al. / Environment International 75 (2015) 180–198
6. Areas of application of phytobial technology and some casestudies
The following areas have been identified by the authors wherephytobial technology can be applied for As removal from soil andgroundwater.
1. Constructedwetland (CW) or Treatmentwetland for treatingAs con-taminated groundwater: Constructed wetlands are artificial waterbodies where matrix, vegetation and the associated invertebrateand microbial communities are designed to simulate and mimic theability of natural wetlands to remove pollutants from water includ-ing As, and are good example of ecological engineering (Kadlec andKnight, 1996). CWs are cost effective, provide ideal application ofphytobial remediation, and present a “green” image within theurban/industrial context. Two types of constructed wetlands are incommon use: the first type, the free water surface (FWS) wetlandis a basin like structurewherewater cannot seep to the undergroundsoil but is directly exposed to the atmosphere like any natural waterbody.The second type subsurface flow (SF)wetland also consists of a basinwith a barrier to prevent seepage to soil, but the bed contains a suit-able depth of porous media (rock or gravel) and the water is notexposed to atmosphere and the water level in the bed remainsbelow the top of the rock or gravel. The flow path of the contaminatedwater may be vertical or horizontal. If the water surface is maintainedbelow themedia surface there is little risk of odors, or insect vectors inthe SF type and the media provides greater available surface area fortreatment. However, the areawould be smaller in area than a FWS sys-temdesigned for the samewastewater conditions. Lizama et al. (2011)have reviewed on potential removal mechanisms that may operate inthe complex bioreactor system of CW.
2. Constructed wetland for treating acid mine drainage site: Acid minedrainage (AMD) has been considered one of the industries' toughestproblems to solve. Most water remediation techniques are timelyand costly, and AMDcan continue over a period ofmany years, there-fore requiring treatment of AMD to continue until well after the landhas been abandoned bymining activities. Constructed wetlands pro-vide a continuous, low-cost and effective solution to the growingproblem of AMD sites from mining industries.
3. Riparian buffer: Arsenic builds up in agricultural fields not only endup in the crops but agricultural run offs contaminate surface waterbodies and nearby streams also. If plants with ability to sequesterAs are planted as riparian buffer between agricultural fields and riv-ers, it may reduce the impact of this pollutant.
4. Dry covers/landfill cover/vegetated cap: The agricultural fields/abandon mine areas degraded beyond use can be recovered withthe use of native plants able to phytoextract As. This is called vegetatedcap or cover. It controls hydraulic flow, prevents soil erosion, preventsleachate formation, minimize infiltration of precipitation. A vegetatedcap can be constructed over landfill or groundwater. Here they alsoact as hydraulic barrier.
Phytoremediation technology has been successfully applied for fieldscale decontamination of lead (Blaylock and Huang, 2000), uranium(Entry et al., 1996) and many others metals. The various successfulpilot scale and bench scale applications of As phytoremediation are en-couraging enough to go for large scale field application which has al-ready proven successful in groundwater (Favas et al., 2012; Schnoor,2000) and soil As remediation (Ye et al., 2011). Jointly with EPA's SBIRProgram, Edenspace Systems Corporation has recently developed anAs phytoremediation technology for in situ treatment of soils containinglarge amounts of CCA (copper chromiumarsenate). Some large scale ap-plications of phytoremediation are underway by FMC Corporation(http,//www.fmc-middleport.com/cms/cmsdocumentlibrary/workplans/
phyto08.pdf) and NEWPCP (http,//dspace.library.drexel.edu/bitstream/1860/123/1/Project.pdf).
7. Phytoinvestigation
Pilot scale studies and field applications require regular monitoringto determine phytoremediation performance of the vegetation of eachharvesting batch. Repeated cropping is continued until As level comesdown to a negligible limit. Monitoring involves analysis of As speciationstatus of different plant parts and the soil. It also involves visual inspec-tion of the health and vigor of the plants including pest problem, regularcollection of fallen plant parts and others. Assessment of soil As bioavail-ability should be performed at regular intervals. Nabeel et al. (2011)showed how As speciation study can be performed using X-ray Absorp-tion Near Edge Structure (XANES). This device coupled with sequentialextraction device can be used for As phytoavailability measurementalso. Bradham et al. (2011) developed one rapid, reliable, and inexpen-sive method for As phytoavailability measurement through in vivo andin vitro assays. Biosensors can be engaged for rapid and reliable moni-toring of the rate of elimination of bioavailable fraction of the pollutants(Shin, 2011). So far many biosensors have been developed and tested fordetecting bioavailable fraction of As (Corbisiera et al., 1993; Tauriainenet al., 1997; Ramanathan et al., 1997; Petänen and Romantschuk, 2003).Whole cell biosensors use reporter gene luciferase or green fluorescentprotein (GFP). All As sensing biosensors are triggered by As(V)/As(III),the analyte which enters the biosensor and activates the well regu-lated transcription of the reporter gene (luc/lux or GFP gene) presentdownstream with regulator and promoter of ars operon. A quick re-sponse is generated, usually luminescence or fluorescencewhich is cap-tured and measured by sensitive fiber optic devices, resulting in real-time data. Like biosensors, phytosensors (indicator plants) could bedesigned that would monitor phytotoxicity and indicate the extent ofphytoremediation.
8. Phytodisposal
The main disadvantage of phytoremediation technology is safe dis-posal of huge quantity of toxic As enriched biomass (Sas-Nowosielskaet al., 2004). Composting and compaction have been suggested as postharvest pretreatments for volume reduction (Cao et al., 2010; Blaylockand Huang, 2000) before final disposal. The high As content of plantwaste doesn't pose detrimental to the microorganisms involved incomposting and compaction. For sustainable use of phytoremediation,large biomass generated as a byproduct of phytoremediation shouldbe reused in an integrated approach through thermochemical conver-sion process, which involves generation of thermal and electrical ener-gy. Combustion, gasification and pyrolysis are the principle methodsof thermochemical energy conversion used for generating electricaland thermal power. Thermochemical energy conversion is the bestuse of disposal of phytoextracted biomass because it cannot be utilizedas fodder and fertilizers due to the presence of toxic As. Combustionprocedure burns the biomass reducing volume to 2–5% and heat energyis generated. Heat should be consumed immediately or converted toelectricity and should not be used for secondary fuel. Air emissionsmust be controlled through gas cleaning devices. The ash producedfrom combustion should be disposed properly or recycled. Research isneeded to formulate a procedure through which metal can be recycledfrom the ash. Before landfilling the ash must be subjected to pretreat-ments like solidificationwith cement and concrete. Gasification is a pro-cess for generation of clean and combustive gas in a high temperaturemetallurgical furnace. Here the biomass are subjected to various chem-ical processes at high temperature (1100–1500 °C) producing amixtureof gases called as producer gas and/or pyro-gas or syn gas (Iyer et al.,2002). Although high temperature is needed but this can be compensat-ed from the resulting syngas that can be used as fuel. Pyrolysis is anoth-er thermochemical decomposition procedure that involves processing

Fig. 4. Advantages and disadvantages of phytoremediation and areas where more research is needed.
193M. Roy et al. / Environment International 75 (2015) 180–198
the biomass anaerobically at a lower temperature (~380 °C) thangastification and produces oil and coke (liquid/semiliquid) instead of asynthesis gas. Heavy metals stay in the coke, which could be fed laterin smelter (Koppolua et al., 2003). Co-incineration on short term andlow-temperature pyrolysis or high-temperature gasification on a long-term basis was suggested by Helsen and Bulck (2005) to treat CCA.Yan et al. (2008) gave some clues on As volatilization behavior duringincineration of P. vittata biomass. Flue gas cleaning equipment can beused to capture released As. Environmental exposure of As due to es-cape by volatilization should be kept to a minimum during the thermochemical processing. Pyrolysis process in comparison to incineration orgastification gives better recovery of As and less release of volatilized Asspecies. Nicks and Chambers (1994) described how thermochemical con-version can bring the biomass generated from As phytoaccumulatingplants into a source of revenue. More research is needed on the develop-ment of improved phytodisposal end management system as the futureof phytoremediation depends upon them.
9. Conclusion and recommendations
More than 30 and 20 years have passed since dissolved As was firstconfirmed in groundwater of West Bengal and Bangladesh in 1983 and1993, respectively (Smith et al., 2000). But still now sustainable meansof supplying safe drinking water has not been widely achieved and
millions of sufferers are still exposed to toxic concentration of As. Al-though conventional methods of As mitigation alternatives like solidifi-cation/vitrification/soil washing/acid extraction/flushing for soiltreatment and precipitation/co-precipitation/adsorption/ion exchangefor groundwater treatments exist (EPA, 2002), but these are difficult toimplement on a large scale due to their environmentally invasive nature,high cost involved in decontamination of the vast contaminated sites. Incontrast although phytoremediation requires a commitment of time butit suits well for use at very large field sites with low capital cost, low op-erational and low management cost; at sites where As concentrationsare below regulatory action levels and only “polishing treatment” is re-quired for eco restoration. Fig. 4 shows the advantages and disadvantagesof phytoremediation and the areas where more knowledge explorationsare required. With the vast amount of information available on As soiland groundwater remediation techniques, we identify the current re-search gaps and areas where more research is needed as mentionedbelow
1. In contrary to worldwide attention for solving the drinking watercrisis due to groundwater As problem, crisis for As free irrigationwater in As effected region has got almost no attention. Ifimmediation attention is not paid, As present in irrigation waterwould be the second most important cause of arsenicosis epidemicin the developing countries where majority of the As effected popu-lation is also suffering frommalnutrition. Arsenic accumulates in top

194 M. Roy et al. / Environment International 75 (2015) 180–198
soil due to irrigation with As contaminated water and results in Asfood chain contamination. For irrigation purpose we cannot use thedeep As free aquifer resource due the fear of downward migrationof As due to overuse of deep aquifer. So decontamination of ground-water from As would be the only option for supply to the farmers.The authors suggest that phytobialremediation can be applied inconstructed wetland system to solve this problem as large amountof groundwater can be remediated through this C.W. system.
2. At present, no official guidance is available on the design criteria forsetting up of a large wetland for arsenic removal. Extensive lab,pilot and field scale study and geological modeling experiments arerequired to develop a CW.More research should be conducted on un-derstanding of arsenic removal mechanisms operating in a CW.
3. More research is needed on development of phytodisposal technolo-gy and new technology may be invented to extract the heavy metal-loid As from the disposed biomass and reuse of the safe biomasswithheavy amount of sequestered carbon for biofuel or fodder.
In summary, we can say that phytobial remediation either alone orin companywith others offers the future low cost solution of As remedi-ation over vast region of As contaminated soil and groundwater and es-thetically would please the society.
Acknowledgment
The authors thank ICMR (ICMR GAP-296) and CSIR (NWP-0052) forproviding financial support for the preparation of this manuscript. Theauthors also sincerely apologize to the investigators whose publicationswere unintentionally omitted from this review due to the presence of avast number of investigations done on arsenic phyto and bioremediation.
References
Abedin, M.J., Feldmann, J., Meharg, A.A., 2002. Uptake kinetics of arsenic species in riceplants. Plant Physiol. 128, 1120–1128.
Aboughalma, H., Bi, R., Schlaak, M., 2008. Electrokinetic enhancement on phytoremediationin Zn, Pb, Cu and Cd contaminated soil using potato plants. J. Environ. Sci. Health A Tox.Hazard. Subst. Environ. Eng. 43, 926–933.
Achal, V., Pan, X., Fu, Q., Zhang, D., 2012. Biomineralization based remediation of As(III)contaminated soil by Sporosarcina ginsengisoli. J. Hazard. Mater. 201–202, 178–184.
Agely, A.A., Sylvia, D.M., Ma, L., 2005. Mycorrhiza increase arsenic uptake by thehyperaccumulator Chinese brake fern (Pteris vittata L.). J. Environ. Qual. 34, 2181–2186.
Ahmann, D., Roberts, A.L., Krumholz, L.R., Morel, F.M., 1994. Microbe grows by reducingarsenic. Nature 371, 750.
Aken Van, B., Peres, C., Doty, S., Yoon, J., Schnoor, J., 2004.Methylobacterium populi sp. nov.,a novel aerobic, pink pigmented, facultatively methylotrophic, methane-ultilisingbacterium isolated from poplar trees (Populus deltoides × nigra DN34). Evol. Microbiol.54, 1191–1196.
Akter, K.F., Owens, G., Davey, D.E., Naidu, R.K., 2005. Arsenic speciation and toxicity in bi-ological system. Rev. Environ. Contam. Toxicol. 184, 97–149.
Alvarado, S., Guédez, M., Lué-merú, M.P., Nelson, G., Alvaro, A., Jesús, A.C., Gyula, Z., 2008.Arsenic removal from waters by bioremediation with the aquatic plants water hya-cinth (Eichhornia crassipes) and lesser duckweed (Lemna minor). Bioresour. Technol.99, 8436–8440.
Andrewes, P., Kitchin, K.T., Wallace, K., 2003. Dimethylarsine and trimethylarsine are po-tent genotoxins in vitro. Chem. Res. Toxicol. 16, 994–1003.
Antosiewicz, D.M., Escudĕ-duran, C., Wierzbowska, E., Skłodowska, A., 2008. Indigenousplant species with the potential for the phytoremediation of arsenic and metals con-taminated soils. Water Air Soil Pollut. 193, 197–210.
Bahar, M.M., Megharaj, M., Naidu, R., 2012. Arsenic bioremediation potential of a newarsenite-oxidizing bacterium Stenotrophomonas sp. MM-7 isolated from soil. Biodeg-radation 23, 803–812.
Baker, A.J.M., 1981. Accumulators and excluders, strategies in the response of plants totrace metals. J. Plant Nutr. 3, 643–654.
Baldwin, P.R., Butcher, D.J., 2007. Phytoremediation of arsenic by two hyperaccumulatorsin a hydroponic environment. Microchem. J. 85, 297–300.
Banerjee, N., Nandy, S., Kearns, J.K., Bandyopadhyay, A.K., Das, J.K., Majumder, P., Basu, S.,Banerjee, S., Sau, T.J., States, J.C., Giri, A.K., 2011. Polymorphisms in the TNF-α and IL10gene promoters and risk of arsenic-induced skin lesions and other nondermatologicalhealth effects. Toxicol. Sci. 121, 132–139.
Banerjee, M., Banerjee, N., Bhattacharjee, P., Mondal, D., Lythgoe, P.R., Martincz, M., Pan, J.,Polya, D.A., Giri, A.K., 2013. High arsenic in rice is associated with elevated genotoxiceffects in humans. Sci. Rep. 3, 2195.
Barac, T., Taghavi, S., Borremans, B., Provoost, A., Oeyen, L., Colpaert, J.V., Vangronsveld, J.,Van Der Lelie, D., 2004. Engineered endophytic bacteria improve phyto-remediationof water soluble, volatile, organic pollutants. Nat. Biotechnol. 22, 583–588.
Barrachina, A.C., Carbonell, F.B., Beneyto, J.M., 1995. Arsenic uptake, distribution, and ac-cumulation in tomato plants — human health risk. Fresenius Environ. Bull. 4,395–400.
Bentley, R., Chasteen, T.G., 2002. Microbial methylation of metalloids, arsenic, antimony,and bismuth. Microbiol. Mol. Biol. Rev. 66, 250–271.
Berti, W.R., Cunningham, S.D., 2000. Phytostabilization of metals. In: Raskin, I., Ensley, B.D.(Eds.), Phytoremediation of Toxic Metals, Using Plants to Clean-up the Environment.John Wiley and Sons, Inc., New York, pp. 71–88.
Bertin, P.N., Heinrich-Salmeron, A., Pelletier, E., Goulhen-Chollet, F., Arsène-Ploetze, F.,Gallien, S., et al., 2011. Metabolic diversity among main microorganisms inside anarsenic-rich ecosystem revealed by meta- and proteo-genomics. ISME J. 5,1735–1747.
Blaylock, M.J., Huang, J.W., 2000. Phytoextraction of metals. In: Raskin, I., Ensley, B.D.(Eds.), Phytoremediation of Toxic Metals, Using Plants to Clean up the Environment.John Wiley and Sons, New York, pp. 53–70.
Bleeker, P.M., Hakvoort, H.W.J., Bliek, M., Souer, E., Schat, H., 2006. Enhanced arsenate re-duction by a CDC25-like tyrosine phosphatase explains increased phytochelatin accu-mulation in arsenate-tolerant Holcus lanatus. Plant J. 45, 917–929.
Bradham, K.D., Scheckel, K.G., Nelson, C.M., Seales, P.E., Lee, G.E., Hughes, M.F., Miller, B.W.,Yeow, A., Gilmore, T., Serda, S.M., Harper, S., Thomas, D.J., 2011. Relative bioavailabilityand bioaccessibility and speciation of arsenic in contaminated soils. Environ. HealthPerspect. 119, 1629–1634.
Brammer, H., 2009. Mitigation of arsenic contamination in irrigated paddy soils in southand south-east Asia. Environ. Int. 35, 856–863.
Brazil, G.M., Kenefick, L., Callanan, M., Haro, A., Lorenzo, V.D., Dowling, D.N., O'gara, F.,1995. Construction of a rhizosphere pseudomonad with potential to degradepolychlorinated biphenyls and detection of bph gene expression in the rhizosphere.Appl. Environ. Microbiol. 61, 1946–1952.
Cameselle, C., Chirakkara, R.A., Reddy, K.R., 2013. Electrokinetic-enhanced phytoremediationof soils, status and opportunities. Chemosphere http://dx.doi.org/10.1016/j.chemosphere.2013.06.029.
Ca'novas, D., Riet, V., Henk, S., Lorenzo, V.D., 2004. The role of thiol species in thehypertolerance of Aspergillus sp. P37 to arsenic. J. Biol. Chem. 279, 51234–51240.
Cao, X.D., Ma, L.Q., Shiralipour, A., 2003. Effects of compost and phosphate amendmentson arsenic mobility in soils and arsenic uptake by the hyperaccumulator Pteris vittataL. Environ. Pollut. 126, 157–167.
Cao, X.D., Ma, L.Q., Tu, C., 2004. Antioxidative responses to arsenic in the arsenichyperaccumulator Chinese brake fern (Pteris vittata L.). Environ. Pollut. 128,317–325.
Cao, X., Ma, L., Shiralipour, A., Harris, W., 2010. Biomass reduction and arsenic transforma-tion during composting of arsenic-rich hyperaccumulator Pteris vittata L. Environ. Sci.Pollut. Res. 17, 586–594.
Castillo-Michel, H., Hernandez-Viezcas, J., Dokken, K.M., Marcus, M.A., Peralta-Videa, J.R.,Gardea-Torresdey, J.L., 2011. Localization and speciation of arsenic in soil and desertplant parkinsonia florida using μXRF and μXANES. Environ. Sci. Technol. 45,7848–7854.
Chang, W.C., Hsu, G.S., Chiang, S.M., Su, M.C., 2006. Heavy metal removal from aqueoussolution by wasted biomass from a combined AS-biofilm process. Bioresour. Technol.97, 1503–1508.
Chauhan, N.S., Ranjan, R., Purohit, H.J., Kalia, V.C., Sharma, R., 2009. Identification of genesconferring arsenic resistance to Escherichia coli from an effluent treatment plantsludge metagenomic library. FEMS Microbiol. Ecol. 67, 130–139.
Chen, Z., Zhu, Y.G., Liu, W.J., Meharg, A.A., 2005. Direct evidence showing the effect of rootsurface iron plaque on arsenite and arsenate uptake into rice (Oryza sativa) roots.New Phytol. 165, 91–97.
Cheremisinoff, N.P., 1998. Groundwater Remediation and Treatment Technologies. NoyesPublications, p. 395.
Christopher, O.A., Haque, A.M.M., 2012. Arsenic contamination in irrigation water for riceproduction in Bangladesh, a review. Trends Appl. Sci. Res. 7, 331–349.
Corbisiera, P., Jib, G., Nuytsa, G., Mergeaya,M., Silver, S., 1993. LuxAB gene fusionswith thearsenic and cadmium resistance operons of Staphylococcus aureus plasmid pI258.FEMS Microbiol. Lett. 110, 231–238.
Crameri, A., Dawes, G., Rodriguez, E., Silver, S., Stemmer, W.P.C., 1997. Molecular evolu-tion of an arsenate detoxification pathway DNA shuffling. Nat. Biotechnol. 15,436–438.
Cunningham, S.D., Berti, W.R., Huang, J.W., 1995. Phytoremediation of contaminated soils.Trends Biotechnol. 13, 393–397.
Cunningham, S.D., Ow, D.W., 1996. Promises and prospects of phytoremediation. PlantPhysiol. 110, 715–719.
Dameron, C.T., Winge, D.R., 1990. Peptide-mediated formation of quantum semiconduc-tors. Trends Biotechnol. 8, 3–6.
Das, D., Chatterjee, A., Mandal, B.K., Samanta, G., Chakraborti, D., Chanda, B., 1995. Arsenicinground water in six districts of West Bengal, India, the biggest As calamity in theworld. Part 2. Arsenic concentration in drinking water, hair, nails, urine, skin-scaleand liver tissue (biopsy) of the affected people. Analyst 120, 917–925.
Dasgupta, T., Hossain, S.A., Meharg, A.A., Price, A.H., 2004. An arsenate tolerance gene onchromosome 6 of rice. New Phytol. 163, 45–49.
Dastidar, A., Wang, Y.T., 2012. Modeling arsenite oxidation by chemoautotrophicThiomonas arsenivorans strain b6 in a packed-bed bioreactor. Sci. Total Environ.432, 113–121.
Davison, J., 2005. Risk mitigation of genetically modified bacteria and plants designed forbioremediation. J. Ind. Microbiol. Biotechnol. 32, 639–650.
De Chaudhuri, S., Kundu, M., Banerjee, M., Das, J.K., Majumdar, P., Basu, S., Roychoudhury, S.,Singh, K.K., Giri, A.K., 2008a. Arsenic-induced health effects and genetic damage in ker-atotic individuals, involvement of p53 arginine variant and chromosomal aberrations inarsenic susceptibility. Mutat. Res. 659, 118–125.

195M. Roy et al. / Environment International 75 (2015) 180–198
De Chaudhuri, S., Ghosh, P., Sarma, N., Majumdar, P., Sau, T.J., Basu, S.,Roychoudhury, S., Ray, K., Giri, A., 2008b. Genetic variants associated with arse-nic susceptibility, study of purine nucleoside phosphorylase, arsenic (+3)methyltransferase, and glutathione S-transferase omega genes. Environ. HealthPerspect. 116, 501–505.
Delavat, F., Lett, M.C., Lièvremont, D., 2012. Novel and unexpected bacterial diversity in anarsenic-rich ecosystem revealed by culture-dependent approaches. Biol. Direct 7, 28.
Delnomdedieu, M., Basti, M.M., Otvos, J.D., Thomas, D.J., 1994. Reduction of binding of ar-senate and dimethylarsinate by glutathione, a magnetic resonances study. Chem.Biol. Interact. 90, 139–155.
Dhankher, O.P., Li, Y., Rosen, B.P., Shi, J., Salt, D., Senecoff, J.F., Sashti, N.A., Meagher, R.B.,2002. Engineering tolerance and hyperaccumulation of arsenic in plants by combin-ing arsenate reductase and γ-glutamylcysteine synthetase expression. Nat. Biotechnol.20, 1140–1145.
Dhankher, O.P., Rosen, B.P., Mckinney, E.C., Meagher, R.B., 2006. Hyperaccumulation of ar-senic in the shoots of Arabidopsis silenced for arsenate reductase (ACR2). Proc. Natl.Acad. Sci. U. S. A. 103, 5413–5418.
Ding, D., Li, W., Song, G., Qi, H., Liu, J., 2011. Identification of QTLs for arsenic accumulationin maize (Zea mays L.) using a RIL population. PLoS ONE 6, e25646.
Dobran, S., Zagury, G.J., 2006. Arsenic speciation and mobilization in CCAcontaminationsoils, influence of organic matter content. Sci. Total Environ. 364, 239–250.
Donahoe-Christiansen, J., D'imperio, S., Jackson, C.R., Inskeep,W.P., Mcdermott, T.R., 2004.Arsenite-oxidizing hydrogenobaculum strain isolated from an acid-sulfatechloridegeothermal spring in Yellowstone National Park. Appl. Environ. Microbiol. 70,1865–1868.
Dourado, M.N., Ferreira, A., Araújo, W.L., Azevedo, J.L., Lacav, P.T., 2012. The diversity ofendophytic methylotrophic bacteria in an oil-contaminated and an oil-freemangroveecosystem and their tolerance to heavy metals. Biotechnol. Res. Int. http://dx.doi.org/10.1155/2012/759865.
Duan, G.L., Zhou, Y., Tong, Y.P., Mukhopadhyay, R., Rosen, B.P., 2007. A CDC25 homologuefrom rice functions as an arsenate reductase. New Phytol. 174, 311–321.
Duan, G., Kamiya, T., Ishikawa, S., Arao, T., Fujiwara, T., 2012. Expressing ScACR3 in riceenhanced arsenite efflux and reduced as accumulation in rice grains. Plant Cell Phys-iol. 53, 154–163.
Dushenkov, V., Kumar, P.B.A.N., Motto, H., Raskin, I., 1995. Rhizofiltration, the use ofplants to remove heavy metals from aqueous streams. Environ. Sci. Technol. 29,1239–1245.
Eapen, S., Suseelan, K., Tivarekar, S., Kotwal, S., Mitra, R., 2003. Potential for rhizofiltrationof uranium using hairy root cultures of Brassica juncea and Chenopodium amaranticolor.Environ. Res. 91, 127–133.
Ellis, D.R., Gumaelius, L., Indriolo, E., Pickering, I.J., Banks, J.A., 2006. A novel arsenate re-ductase from the arsenic hyperaccumulating fern Pteris vittata. Plant Physiol. 141,1544–1554.
Entry, J.A., Vance, N.C., Hamilton, M.A., Zabowski, D., Watrud, L.S., Adriano, D.C., 1996.Phytoremediation of soil contaminated with low concentrations of radionuclides.Water Air Soil Pollut. 88, 167–176.
EPA—Solid Waste, Emergency Response (5102G), 2002. Arsenic Treatment Technologiesfor Soil, Waste and Water. www.epa.gov/tioclu-in.org/As.
Favas, P.J., Pratas, J., Prasad, M.N., 2012. Accumulation of arsenic by aquatic plants in large-scale field conditions, opportunities for phytoremediation and bioindication. Sci.Total Environ. 433, 390–397.
Fitz, W.J., Wenzel, W.W., 2002. Arsenic transformations in the soil/rhizosphere/plant sys-tem, fundamentals and potential application to phytoremediation. J. Biotechnol. 99,259–278.
Foyer, C.H., Noctor, G., 2005. Oxidant and antioxidant signalling in plants, a re-evaluationof the concept of oxidative stress in a physiological context. Plant Cell Environ. 28,1056–1071.
Frankenberger, W.T.J.R. (Ed.), 2001. Environmental Chemistry of Arsenic. Marcel Dekker,New York.
Frankenberger,W., Arshad, M., 2002. Volatilization of arsenic. In: Frankenberger, W. (Ed.),Environmental Chemistry of Arsenic. Marcel Dekker, pp. 363–380.
Gadd, G.M., 2010. Metals, minerals and microbes, geomicrobiology and bioremediation.Microbiology 156, 609–643.
García-Salgado, S., García-Casillas, D., Angeles Quijano-Nieto, M., Milagros Bonilla-Simón, M.,2012. Arsenic and heavy metal uptake and accumulation in native plant species fromsoils polluted by mining activities. Water Air Soil Pollut. 223, 559–572.
Germaine, K., Liu, X., Cabellos, G., Hogan, J., Ryan, D., Dowling, D.N., 2006. Bacterialendophyte-enhanced phyto-remediation of the organochlorine herbicide 2,4-dichlorophenoxyacetic acid. FEMS Microbiol. Ecol. 57, 302–310.
Ghassemzadeha, F., Yousefzadehb, H., Arbab Zavarc, M.H., 2008. Arsenic phytoremediationby Phragmites australis, green technology. Int. J. Environ. Stud. 65, 587–594.
Gihring, T.M., Druschel, G.K., Mccleskey, R.B., Hamers, R.J., Banfield, J.F., 2001. Rapid arse-nite oxidation by Thermus aquaticus and Thermus thermophilus, field and laboratoryinvestigations. Environ. Sci. Technol. 35, 3857–3862.
Giri, A.K., Patel, R.K., Mishra, P.C., 2012. Biosorption of As(V) from aqueous solutions byliving cells of Bacillus cereus. Water Sci. Technol. 66, 699–707.
Gohre, V., Paszkowski, U., 2006. Contribution of the arbuscular mycorrhizal symbiosis toheavy metal phytoremediation. Planta 223, 1115–1122.
Gomes, M.P., Carvalho, M., Carvalho, G.S., Marques, T.C.L.L.S.M., Garcia, Q.S., Guilherme, L.R.G.,Soares, A.M., 2013. Phosphorus improves arsenic phytoremediation by Anadenantheraperegrinaby alleviating induced oxidative stress. Int. J. Phytoremediation 15, 633–646.
Guo, J.B., Dai, X.J., Xu,W.Z., Ma, M., 2008. Overexpressing GSH1 and AsPCS1 simultaneous-ly increases the tolerance and accumulation of cadmium and As in Arabidopsisthaliana. Chemosphere 72, 1020–1026.
Hansel, C.M., La Force, M.J., Fendorf, S., Sutton, S., 2002. Spatial and temporal association ofAs and Fe species on aquatic plant roots. Environ. Sci. Technol. 36, 1988–1994.
Harvey, C.F., Swartz, C.H., Badruzzaman, A.B., Keon-Blute, N., Yu, W., Ali, M.A., Jay, J.,Beckie, R., Niedan, V., Brabander, D., Oates, P.M., Ashfaque, K.N., Islam, S., Hemond, H.F.,Ahmed,M.F., 2002. Arsenicmobility and groundwater extraction in Bangladesh. Science298, 1602–1606.
Heijden Van Der, M.G.A., Bardgett, R.D., Van Straalen, N.M., 2008. The unseen majority,soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems.Ecol. Lett. 11, 296–310.
Helsen, L., Bulck, E.V.D., 2005. Review of disposal technologies for chromated copper arse-nate (CCA) treated wood waste, with detailed analyses of thermochemical conver-sion processes. Environ. Pollut. 134, 301–314.
Hoeft, S.E., Kulp, T.R., Stolz, J.F., Hollibaugh, J.T., Oremland, R.S., 2004. Dissimilatory arse-nate reduction with sulfide as the electron donor, experiments with Mono Lakewater and isolation of strain MLMS-1, a chemoautotrophic arsenate-respirer. Appl.Environ. Microbiol. 70, 2741–2747.
Hoeft, S.E., Blum, J.S., Stolz, J.F., Tabita, F.R., Witte, B., King, G.M., 2007. Alkalilimnicolaehrlichii sp. nov., a novel, arsenite-oxidizing haloalkaliphilic gammaproteobacteriumcapable of chemoautotrophic or heterotrophic growth with nitrate or oxygen as theelectron acceptor. Int. J. Syst. Evol. Microbiol. 57, 504–512.
Hollibaugh, J.T., Budinoff, C., Hollibaugh, R.A., Ransom, B., Bano, N., 2006. Sulfide oxidationcoupled to arsenate reduction by a diverse microbial community in a Soda Lake. Appl.Environ. Microbiol. 72, 2043–2049.
Huang, H., Jia, Y., Sun, G.X., Zhu, Y.G., 2012. Arsenic speciation and volatilization fromflooded paddy soils amended with different organic matters. Environ. Sci. Technol.46, 2163–2168.
Huysmans, D.K., Frankenberger, W.T., 1991. Evolution of trimethylarsine by a Penicil-lium sp. isolated from agricultural evaporation pond water. Sci. Total Environ.105, 13–28.
Indriolo, E., Na, G., Ellis, D., Salt, D.E., Banks, J.A., 2010. Vacuolar arsenite transporter nec-essary for as tolerance in the As hyperaccumulating fern Pteris vittata is missing inflowering plants. Plant Cell 22, 2045–2057.
Islam, F.S., Gault, A.G., Boothman, C., Polya, D.A., Charnock, J.M., Chatterjee, D., Lloyd, J.R.,2004. Role of metal-reducing bacteria in As release from Bengal delta sediments. Na-ture 430, 68–71.
Iyer, P.V.R., Rao, T.R., Grover, P.D., 2002. Biomass Thermochemical Characterization, Thirded. , p. 38.
Joshi, D.N., Patel, J.S., Flora, S.J.S., Kalia, K., 2008. As accumulation by Pseudomonas stutzeriand its response to some thiol chelators. Environ. Health Prev. Med. 13, 257–263.
Kadlec, R.H., Knight, R.L., 1996. Treatment Wetlands. Lewis Publishers, Boca Raton, FL.USA.
Kamiya, T., Tanaka, M., Mitani, N., Ma, J.F., Maeshima, M., Fujiwara, T., 2009. NIP1;1, anaquaporin homolog, determines the arsenite sensitivity of Arabidopsis thaliana. J.Biol. Chem. 284, 2114–2120.
Keimowitz, A.R., Mailloux, B.J., Cole, P., Stute, M., Simpson, H.J., Chillrud, S.N., 2007. Labo-ratory investigations of enhanced sulfate reduction as a groundwater As remediationstrategy. Environ. Sci. Technol. 41, 6718–6724.
Khan, A.G., 2005. Role of soil microbes in the rhizospheres of plants growing on tracemetal contaminated soils in phytoremediation. J. Trace Elem. Med. Biol. 8,355–364.
Kirk, G.J.D., Santos, E.E., Findenegg, G.R., 1999. Phosphate solubilisation by organic anionexcretion from rice (Oryza sativa L.) growing in aerobic soil. Plant Soil 211, 11–18.
Kneer, R., Zenk, M.H., 1997. The formation of Cd–phytochelatin complexes in plant cellcultures. Phytochemistry 44, 69–74.
Köber, R., Daus, B., Ebert, M., Mattusch, J., Welter, E., Dahmke, A., 2005. Compost-basedpermeable reactive barriers for the source treatment of arsenic contaminations inaquifers: column studies and solid-phase investigations. Environ. Sci. Technol. 39(19), 7650–7655.
Koppolua, L., Agblover, F.A., Clements, L.D., 2003. Pyrolysis as a technique for separatingheavy metals from hyperaccumulators. Part II, lab-scale pyrolysis of synthetichyperaccumulator biomass. Biomass Bioenergy 25, 651–663.
Kostal, J., Yang, R., Wu, C.H., Mulchandani, A., Chen, W., 2004. Enhanced arsenic accumu-lation in engineered bacterial cells expressing ArsR. Appl. Environ. Microbiol. 70,4582–4587.
Krumov, N., Oder, S., Perner-Nochta, I., Angelov, A., Posten, C., 2007. Accumulation of CdSnanoparticles by yeasts in a fed-batch bioprocess. J. Biotechnol. 132, 481–486.
Kulp, T.R., Han, S., Saltikov, C., Lanoil, B., Zargr, K., Oremland, R.S., 2007. Effects of imposedsalinity gradients on dissimilatory arsenate-reduction, sulfate reduction, and othermicrobial processes in sediments from two California soda lakes. Appl. Environ.Microbiol. 73, 5130–5137.
Kulp, T.R., Hoeft, S.E., Asao, M., Madigan, M.T., Hollibaugh, J.T., 2008. As(III) fuelsanoxygenic photosynthesis in hot spring biofilms fromMono Lake, California. Science321, 967–970.
Kumar, J.I., Oommen, C., 2012. Removal of heavy metals by biosorption using freshwateralga Spirogyra hyalina. J. Environ. Biol. 33, 27–31.
Kwon, J.C., Lee, J.S., Jung, M.C., 2012. Arsenic contamination in agricultural soils surround-ing mining sites in relation to geology and mineralization types. Appl. Geochem. 27,1020–1026.
Lear, G., Polya, D.A., Song, B., Gault, A.G., Lloyd, J.R., 2007. Molecular analysis of arsenate-reducing bacteria within Cambodian sediments following amendment with acetate.Appl. Environ. Microbiol. 73, 1041–1048.
Lee, K.Y., Yoon, I.H., Lee, B.T., Kim, S.O., Kim, K.W., 2009. A novel combination of anaerobicbioleaching and electrokinetics for as removal from mine tailing soil. Environ. Sci.Technol. 43, 9354–9360.
Leist, M., Casey, R.J., Caridi, D., 2000. The management of As wastes, problems and pros-pects. J. Hazard. Mater. 76, 125–138.
Leung, H.M., Ye, Z.H., Wong, M.H., 2006. Interactions of mycorrhizal fungi with Pterisvittata (As hyperaccumulator) in As-contaminated soils. Environ. Pollut. 139, 1–8.

196 M. Roy et al. / Environment International 75 (2015) 180–198
Li, W.X., Chen, T.B., Huang, Z.C., Lei, M., Liao, X.Y., 2006. Effect of arsenic on chloroplastultrastructure and calcium distribution in As hyperaccumulator P. vittata L.Chemosphere 62, 803–809.
Liao, X.Y., Chen, T.B., Xiao, X.Y., Xie, H., Yan, X.L., Zhai, L.M., Wu, B., 2007. Selecting appro-priate forms of nitrogen fertilizer to enhance soil As removal by P. vittata, a new ap-proach in phytoremediation. Int. J. Phytoremediation 9, 269–280.
Lieveremont, D., Bertin, P.N., Lett, M.C., 2009. Arsenic in contaminated waters, biogeo-chemical cycle, microbial metabolism and biotreatment processes. Biochimie 91,1229–1237.
Linacre, N.A., Whiting, S.N., Baker, A.J.M., Angle, J.S., Ades, P.K., 2003. Transgenics andphytoremediation: the need for an integrated risk assessment, management, andcommunication strategy. Int. J. Phytoremediation 5, 181–185.
Littera, P., Urík, M., Sevc, J., Kolencík, M., Gardosová, K., Molnárová, M., 2011. Removal ofarsenic from aqueous environments by native and chemically modified biomass ofAspergillus niger and Neosartorya fischeri. Environ. Technol. 32, 1215–1222.
Liu, W.J., Wood, B.A., Raab, A., Mcgrath, S.P., Zhao, F.J., Feldmann, J., 2010. Complexation ofarsenite with phytochelatins reduces arsenite efflux and translocation from roots toshoots in Arabidopsis thaliana. Plant Physiol. 152, 2211–2221.
Liu, S., Zhang, F., Chen, J., Sun, G., 2011. Arsenic removal from contaminated soil viabiovolatilization by genetically engineered bacteria under laboratory conditions. J.Environ. Sci. 23, 1544–1550.
Liu, W., Schat, H., Bliek, M., Chen, Y., Mcgrath, S.P., 2012. Knocking out ACR2 does not af-fect arsenic redox status in Arabidopsis thaliana, implications for arsenic detoxifica-tion and accumulation in plants. PLoS ONE 7, e42408.
Lizama, A.K., Fletcher, T.D., Sun, G., 2011. Removal processes for arsenic in constructedwetlands. Chemosphere 84, 1032–1043.
Lombi, E., Zhao, F.J., Fuhrmann, M., Ma, L.Q., Mcgrath, S.P., 2002. Arsenic distribution andspeciation in the fronds of the hyperaccumulator Pteris vittata. New Phytol. 156,195–203.
Lynch, J.M., Moffat, A.J., 2005. Bioremediation— prospects for the future application of in-novative applied biological research. Ann. Appl. Biol. 146, 217–221.
Ma, L.Q., Komar, K.M., Tu, C., Zhang, W., Cai, Y., Kennelley, E.D., 2001. A fern thathyperaccumulates arsenic. Nature 409, 579.
Ma, J.F., Yamaji, N., Mitani, N., Xu, X.Y., Su, Y.H., Mcgrath, S.P., Zhao, F.J., 2008. Transportersof arsenite in rice and their role in arsenic accumulation in rice grain. Proc. Natl. Acad.Sci. U. S. A. 105, 9931–9935.
Maciaszczyk-Dziubinska, E., Wawrzycka, D., Wysocki, R., 2012. Arsenic and antimonytransporters in eukaryotes. Int. J. Mol. Sci. 13, 3527–3548.
Macur, R.E., Jackson, C.R., Botero, L.M., Mcdermott, T.R., Inskeep,W.P., 2004. Bacterial pop-ulations associatedwith the oxidation and reduction of arsenic in an unsaturated soil.Environ. Sci. Technol. 38, 104–111.
Malasarn, D., Saltikov, C., Campbell, K.M., Santini, J.M., Hering, J.G., Newman, D.K., 2004.arrA is a reliable marker for arsenic(V) respiration. Science 306, 455.
Mass, M.J., Tennant, A., Roop, B.C., Cullen, W.R., Styblo, M., Thomas, D.J., Kligerman, A.D.,2001. Methylated trivalent arsenic species are genotoxic. Chem. Res. Toxicol. 14,355–361.
Mcarthur, J.M., Ravenscroft, P., Safiullah, S., Thirlwall, M.F., 2001. Arsenic in groundwater,testing pollution mechanisms for sedimentary aquifers in Bangladesh. Water Resour.Res. 37, 109–117.
Meharg, A.A., Macnair, M.R., 1990. An altered phosphate uptake system in arsenate toler-ant Holcus lanatus. New Phytol. 116, 29–35.
Meharg, A.A., Macnair, M.R., 1991. The mechanisms of arsenate tolerance in Deschampsiacespitosa (L.) Beauv and Agrostis capillaris L. New Phytol. 119, 291–297.
Meharg, A.A., Macnair, M.R., 1992. Suppression of the high affinity phosphate uptake sys-tem, a mechanism of arsenate tolerance in Holcus lanatus L. J. Exp. Bot. 43, 519–524.
Mendoza-Cozatl, D.G., Rodriguez-Zavala, J.S., Rodriguez-Enriquez, S., Mendoza-Hernandez,G., Briones-Gallardo, R., Moreno-Sanchez, R., 2006. Phytochelatin-cadmium-sulfidehigh-molecular-mass complexes of Euglena gracilis. FEBS J. 273, 5703–5713.
Meng, X.Y., Qin, J., Wang, L.H., Duan, G.L., Sun, G.X., Wu, H.L., Chu, C.C., Ling, H.Q., Rosen, B.P.,Zhu, Y.G., 2011. Arsenic biotransformation and volatilization in transgenic rice. NewPhytol. 191, 49–56.
Merrifield, M.E., Ngu, T., Stillman, M.J., 2004. Arsenic binding to Fucus vesiculosusmetallo-thionein. Biochem. Biophys. Res. Commun. 324, 127–132.
Michael, H., Voss, C.I., 2008. Evaluation of the sustainability of deep groundwater as anarsenic-safe resource in the Bengal Basin. Proc. Natl. Acad. Sci. U. S. A. 105,8531–8536.
Mirza, N., Pervez, A., Mahmood, Q., Shah, M.M., Shafqat, M.N., 2011. Ecological restorationof arsenic contaminated soil by Arundo donax L. Ecol. Eng. 37, 1949–1956.
Mosa, K.A., Kumar, K., Chhikara, S., Mcdermott, J., Liu, Z., Musante, C., White, J.C.,Dhankher, O.P., 2012. Members of rice plasma membrane intrinsic proteins sub-family are involved in arsenite permeability and tolerance in plants. Transgenic Res.21, 1265–1277.
Mukhopadhyay, R., Rosen, B.P., 1998. Saccharomyces cerevisiae ACR2 gene encodes an ar-senate reductase. FEMS Microbiol. Lett. 168, 127–136.
Mukhopadhyay, R., Shi, J., Rosen, B.P., 2000. Purification and characterization of ACR2p,the Saccharomyces cerevisiae arsenate reductase. J. Biol. Chem. 275, 21149–21157.
Mukhopadhyay, R., Rosen, B.P., 2001. The phosphatase C(X)5R motif is required for cata-lytic activity of the Saccharomyces cerevisiae Acr2p arsenate reductase. J. Biol. Chem.276, 34738–34742.
Mukhopadhyay, R., Rosen, B.P., Phung, L.T., Silver, S., 2002. Microbial arsenic, fromgeocycles to genes to enzymes. FEMS Microbiol. Rev. 26, 311–325.
Muller, D., Médigue, C., Koechler, S., 2007. A tale of two oxidation states, bacterial coloni-zation of arsenic-rich environments. PLoS Genet. 3, 518–530.
Nabeel, K.N., Singh, B., Shah, P., 2011. Arsenic speciation and phytoavailability in contam-inated soils using a sequential extraction procedure and XANES spectroscopy. Envi-ron. Sci. Technol. 45, 7135–7142.
Nair, A., Juwarkar, A.A., Singh, S.K., 2007. Production and characterization of siderophoresand its application in arsenic removal from contaminated soil. Water Air Soil Pollut.180, 199–212.
Naujokas,M.F., Anderson, B., Ahsan, H., Aposhian, H.V., Graziano, J.H., Thompson, C., Suk,W.A.,2013. The broad scope of health effects from chronic arsenic exposure, update on aworldwide public health problem. Environ. Health Perspect. 121, 295–302.
Nedelkoska, T.V., Doran, P.M., 2000. Hyperaccumulation of cadmium by hairy roots ofThlaspi caerulescens. Biotechnol. Bioeng. 67, 607–615.
Niazi, N.K., Singh, B., Zwieten, L.V., Kachenko, A.G., 2012. Phytoremediation of anarsenic-contaminated site using Pteris vittata L. and Pityrogramma calomelanosvar. austroamericana, a long-term study. Environ. Sci. Pollut. Res. 19, 3506–3515.
Nicks, L., Chambers, M.F., 1994. Nickel farm. Discov. 19.Nickson, R., Mcarthur, J.M., Burgess, W.G., Ahmed, K.M., Ravenscroft, P., Rahman,M., 1998.
Arsenic poisoning in Bangladesh groundwater. Nature 395, 338.Nickson, R., Mcarthur, J.M., Ravenscroft, P., Burgess, W.G., Ahmed, K.M., 2000. Mechanism
of arsenic release to groundwater, Bangladesh and West Bengal. Appl. Geochem. 15,403–413.
Nie, L., Shah, S., Rashid, A., Burd, G.I., Dixon, D.G., Glick, B.R., 2002. Phytoremediation of ar-senate contaminated soil by transgenic canola and the plant growth-promoting bac-terium Enterobacter cloacae CAL2. Plant Physiol. Biochem. 40, 355–361.
O'connor, C.S., Lepp, N.W., Edwards, R., Sunderland, G., 2003. The combined use of electro-kinetic remediation and phytoremediation to decontaminate metal-polluted soils, alaboratory-scale feasibility study. Environ. Monit. Assess. 84, 141–158.
Olmos-Márquez, M.A., Alarcón-Herrera, M.T., Martín-Domínguez, I.R., 2012. Performanceof Eleocharis macrostachya and its importance for arsenic retention in constructedwetlands. Environ. Sci. Pollut. Res. Int. 19, 763–771.
Olsen, E.J., Kracher, A., Davis, A.M., Steele, I.M., Hutcheon, I.D., Bunch, T.E., 1999. The phos-phates of IIIAB iron meteorites. Meteorit. Planet. Sci. 34, 285–300.
Oremland, R.S., Hoeft, S.E., Bano, N., Hollibaugh, R.A., Hollibaugh, J.T., 2002. Anaerobic ox-idation of arsenite in Mono Lake water and by a facultative, arsenite-oxidizing che-moautotroph, strain MLHE-1. Appl. Environ. Microbiol. 68, 4795–4802.
Oremland, R.S., Stolz, J.F., 2003. The ecology of arsenic. Science 299, 939–944.Oremland, R.S., Saltkov, C.W., Wolfe-Simon, F., Stolz, J.F., 2009. Arsenic in the evolution of
earth and extraterrestrial ecosystems. Geomicrobiol. J. 26, 522–536.Orłowska, E., Godzik, B., Turnau, K., 2012. Effect of different arbuscular mycorrhizal fungal
isolates on growth and arsenic accumulation in Plantago lanceolata L. Environ. Pollut.168, 121–130.
Paul, S., Das, N., Bhattacharjee, P., Banerjee, M., Das, J.K., Sarma, N., Sarkar, A.,Bandyopadhyay, A.K., Sau, T.J., Basu, S., Banerjee, S., Majumder, P., Giri, A.K., 2013.Arsenic-induced toxicity and carcinogenicity, a two-wave cross-sectional study inarsenicosis individuals in West Bengal, India. J. Expo. Sci. Environ. Epidemiol. 23,156–162.
Pederick, R.L., Gault, A.G., Charnock, J.M., Polya, D.A., Lloyd, J.R., 2007. Probing thebiogeochemistry of arsenics, response of two contrasting aquifer sedimentsfrom Cambodia to stimulation by arsenate and ferric iron. J. Environ. Sci. HealthA42, 1763–1774.
Perez-Jim 'Enez, J.R., Defraia, C., Young, L.Y., 2005. Arsenate respiratory reductase gene (arrA)for Desulfosporosinus sp. strain Y5. Biochem. Biophys. Res. Commun. 338, 825–829.
Petänen, T., Romantschuk, M., 2003. Toxicity and bioavailability to bacteria of particle-associated arsenite and mercury. Chemosphere 50, 409–413.
Petrick, J.S., Ayala-Fierro, F., Cullen, W.R., Carter, D.E., Aposhian, H.V., 2000.Monomethylarsonous acid (MMA(III)) is more toxic than arsenite in Changhuman hepatocytes. Toxicol. Appl. Pharmacol. 163, 203–207.
Phillips, D.J.H., 1990. Arsenic in aquatic organisms: a review, emphasizing chemical speci-ation. Aquat. Toxicol. 16, 151–186.
Pickering, I.J., Prince, R.C., George, M.J., Smith, R.D., George, G.N., Salt, D.E., 2000. Reductionand coordination of arsenic in Indian mustard. Plant Physiol. 122, 1171–1177.
Pickering, I.J., Gumaelius, L., Harris, H.H., Prince, R.C., Hirsch, G., Banks, J.A., Salt, D.E.,George, G.N., 2006. Localizing the biochemical transformations of arsenate in ahyperaccumulating fern. Environ. Sci. Technol. 40, 5010–5014.
Porras-Alfaro, A., Bayman, P., 2011. Hidden fungi, emergent properties, endophytes andmicrobiomes. Annu. Rev. Phytopathol. 49, 291–315.
Poser, A., 2005. Microbiology of the arsenic, microbiology of the origin of volatile arseniccompounds. Geoecology 1–10.
Pott, W.A., Benjamin, S.A., Yang, R.S.H., 2001. Pharmacokinetics, metabolism, and carcino-genicity of arsenic. Rev. Environ. Contam. Toxicol. 169, 165–214.
Poynton, C.Y., Huang, J.W.W., Blaylock, M.J., Kochian, L.V., Elless, M.P., 2004. Mechanismsof arsenic hyperaccumulation in Pteris species, root As influx and translocation. Planta219, 1080–1088.
Qin, J., Rosen, B.P., Zhang, Y., Wang, G., Franke, S., Rensing, C., 2006. Arsenic detoxificationand evolution of trimethylarsine gas by a microbial arsenite S-adenosylmethioninemethyltransferase. Proc. Natl. Acad. Sci. U. S. A. 103, 2075–2080.
Rahman,M.A., Hasegawa, H., 2011. Aquatic arsenic, phytoremediation using floatingmac-rophytes. Chemosphere 83, 633–646.
Rahman, S., Kim, K.H., Saha, S.K., Swaraz, A.M., 2014. Review of remediation techniquesfor arsenic (As) contamination: a novel approach utilizing bio-organisms. J. Environ.Manag. 134, 175–185.
Ramanathan, S., Shi, W., Rosen, B.P., Daunert, S., 1997. Sensing antimonite and arsenite atthe subattomole level with genetically engineered bioluminescent bacteria. Anal.Chem. 69, 3380–3384.
Rathinasabapathi, B., Raman, S.B., Kertulis, G., Ma, L.Q., 2006. Arsenic resistantproteobacterium in the phyllosphere of the Chinese brake fern (Pteris vittata L.) re-duces arsenate to arsenite. Can. J. Microbiol. 52, 695–700.
Ravenscroft, P., Mcarthur, J.M., Hoque, M.A., 2013. Stable groundwater quality in deepaquifers of Southern Bangladesh, the case against sustainable abstraction. Sci. TotalEnviron. 454–455, 627–638.

197M. Roy et al. / Environment International 75 (2015) 180–198
Requejo, R., Tena, M., 2005. Proteome analysis of maize roots reveals that oxidative stressis a main contributing factor to plant arsenic toxicity. Phytochemistry 66, 1519–1528.
Richard, T.W., Steven, D.A., Douglas, G.B., Randall, R.R., Tony, R.L., Cindy, J.P., 2008. FieldApplication of a Permeable Reactive Barrier for Treatment of Arsenic in GroundWaterEPA 600/R-08/093 www.epa.gov/ada (September).
Robinson, B., Kim, N., Marchetti, M., Moni, C., Schroeter, L., 2006. Arsenic hyperaccumulationby aquaticmacrophytes in the Taupo Volcanic Zone, New Zealand. Environ. Exp. Bot. 58,206–215.
Rochette, E.A., Bostick, B.C., Li, G.C., Fendorf, S., 2000. Kinetics of arsenate reduction by dis-solved sulfide. Environ. Sci. Technol. 34, 4714–4720.
Rofkar, J.R., Dwyer, D.F., 2013. Irrigation of three wetland species and a hyperaccumlatingfern with arsenic-laden solutions, observations of growth, As uptake, nutrient status,and chlorophyll content. Int. J. Phytoremediation 15, 561–572.
Rosen, B.P., 2002. Transport and detoxification systems for transition metals, heavymetals andmetalloids in eukaryotic and prokaryotic microbes. Comp. Biochem. Phys-iol. A Mol. Integr. Physiol. 133, 689–693.
Rosen, B.R., Liu, Z.J., 2009. Transport pathways for arsenic and selenium, a minireview. En-viron. Int. 35, 512–515.
Ryan, R.P., Ryan, D., Dowling, D.N., 2007. Plant protection by the recombinant, root-colonizing Pseudomonas fluorescens F113rifPCB strain expressing arsenic resistance,improving rhizoremediation. Lett. Appl. Microbiol. 45, 668–674.
Ryan, R.P., Germaine, K., Franks, A., Ryan, D.J., Dowling, D.N., 2008. Bacterial endophytes,recent developments and applications. FEMS Microbiol. Lett. 278, 1–9.
Sakakibara, M., Watanabe, A., Sano, S., Inoue, M., Kaise, T., 2010. Phytoextraction andphytovolatilization of arsenic from As contaminated soils by Pteris. Proceedings ofthe Annual International Conference on Soils, Sediments, Water and Energy. vol. 12(Article 26).
Salmassi, T.M., Walker, J.J., Newman, D.K., Leadbetter, J.R., Pace, N.R., Hering, J.G., 2006.Community and cultivation analysis of arsenite oxidizing biofilms at Hot Creek. Envi-ron. Microbiol. 8, 50–59.
Saltikov, C.W., Cifuentes, A., Venkateswaran, K., Newman, D.K., 2003. The ars detoxifica-tion system is advantageous but not required for As(V) respiration by the geneticallytractable Shewanella species strain ANA-3. Appl. Environ. Microbiol. 69, 2800–2809.
Saltikov, C.W., Newman, D.K., 2003. Genetic identification of a respiratory arsenate reduc-tase. Proc. Natl. Acad. Sci. U. S. A. 100, 10983–10988.
Samal, A.C., Kar, S., Bhattacharya, P., Santra, S.C., 2011. Human exposure to As throughfoodstuffs cultivated using arsenic contaminated groundwater in areas of West Ben-gal, India. J. Environ. Sci. Health 46, 1259–1265.
Santini, J.M., Vanden Hoven, R.N., 2004. Molybdenum containing arsenite oxidase of thechemolithoautotrophic arsenite oxidizer NT-26. J. Bacteriol. 186, 1614–1619.
Santos, J.A.G., Gonzaga, M.I.S., Ma, L.Q., Srivastava, M., 2008. Timing of phosphate applica-tion affects arsenic phytoextraction by P. vittata L. of different ages. Environ. Pollut.154, 306–311.
Sas-Nowosielska, A., Kucharski, R., Małkowski, E., Pogrzeba, M., Kuperberg, J.M., Krynski, K.,2004. Phytoextraction crop disposal—an unsolved problem. Environ. Pollut. 128,373–379.
Sauge-Merke, S., Cuine, S., Carrier, P., Lecomte-Pradines, C., Luu, D.T., Peltier, G., 2003. En-hanced toxic metal accumulation in engineered bacterial cells expressing Arabidopsisthaliana phytochelatin synthase. Appl. Environ. Microbiol. 69, 490–494.
Schmöger, M.E.V., Oven, M., Grill, E., 2000. Detoxification of arsenic by phytochelatins inplants. Plant Physiol. 122, 793–801.
Schnoor, J.L., 2000. Phytostabilization of metals using hybrid poplar trees. In: Raskin, I.,Ensley, B.D. (Eds.), Phytoremediation of Toxic Metals, Using Plants to Clean-up theEnvironment. John Wiley and Sons, Inc., New York, pp. 133–150.
Shin, H.J., 2011. Genetically engineered microbial biosensors for in situ monitoring of en-vironmental pollution. Appl. Microbiol. Biotechnol. 89, 867–877.
Shukla, D., Kesari, R., Mishra, S., Dwivedi, S., Tripathi, R.D., Nath, P., Trivedi, P.K., 2012. Ex-pression of phytochelatin synthase from aquatic macrophyte Ceratophyllumdemersum L. enhances cadmium and arsenic accumulation in tobacco. Plant CellRep. 31, 1687–1699.
Siciliano, S., Fortin, N., Himoc, N., 2001. Selection of specific endophytic bacterial geno-types by plants in response to soil contamination. Appl. Environ. Microbiol. 67,2469–2475.
Singh, N., Ma, L.Q., Srivastava, M., Rathinasabapathi, B., 2006. Metabolic adaptations toarsenic-induced oxidative stress in Pteris vittata L. and Pteris ensiformis L. Plant Sci.170, 274–282.
Singh, S., Lee, W., Dasilva, N.A., Mulchandani, A., Chen, W., 2008. Enhanced arsenic accu-mulation by engineered yeast cells expressing Arabidopsis thaliana phytochelatinsynthase. Biotechnol. Bioeng. 99, 333–340.
Smedley, P.L., Kinniburgh, D.G., 2002. A review of the source, behaviour and distributionof arsenic in natural waters. Appl. Geochem. 17, 517–568.
Smith, A.H., Lingas, E.O., Rahman, M., 2000. Contamination of drinking-water by arsenic inBangladesh, a public health emergency. Bull. World Health Organ. 78, 9.
Song, W.Y., Park, J., Mendoza-Cózatl, D.G., Suter-Grotemeyer, M., Shim, D., Hörtensteiner, S.,Geisler, M., Weder, B., Rea, P.A., Rentsch, D., Schroeder, J.I., Lee, Y., Martinoia, E., 2010.Arsenic tolerance in Arabidopsis is mediated by two ABCC-type phytochelatin trans-porters. Proc. Natl. Acad. Sci. U. S. A. 107, 21187–21192.
Srivastava, M., Ma, L.Q., Singh, N., Singh, S., 2005. Antioxidant responses of hyper-accumulator and sensitive fern species to arsenic. J. Exp. Bot. 415, 1335–1342.
Srivastava, M., Ma, L.Q., Rathinasabapathi, B., Srivastava, P., 2009. Effects of selenium onarsenic uptake in arsenic hyperaccumulator P. vittata L. Bioresour. Technol. 100,1115–1121.
Stearns, J.C., Glick, B.R., 2003. Transgenic plants with altered ethylene biosynthesis or per-ception. Biotechnol. Adv. 21, 193–210.
Stolz, J.F., Basu, P., Santini, J.M., Oremland, R.S., 2006. Arsenic and selenium in microbialmetabolism. Annu. Rev. Microbiol. 60, 107–130.
Strobel, G., Daisy, B., Castillo, U., Harper, J., 2004. Natural products from endophyticmicro-organisms. J. Nat. Prod. 67, 257–268.
Ström, L., Olsson, T., Tyler, G., 1994. Differences between calcifuge and acidifuge plants inroot exudation of low molecular organic acids. Plant Soil 167, 239–245.
Su, Y.H., Mcgrath, S.P., Zhu, Y.G., Zhao, F.J., 2008. Highly efficient xylem transport of arse-nite in the arsenic hyperaccumulator Pteris vittata. New Phytol. 180, 434–441.
Sundaram, S., Rathinasabapathi, B., Ma, L.Q., Rosen, B.P., 2008. An arsenate-activatedGlutaredoxin from the arsenic Hyperaccumulator fern Pteris vittata L. regulates intra-cellular arsenite. J. Biol. Chem. 283, 6095–6101.
Susaya, J., Kim, K.H., Jung, M.C., 2010. The impact of mining activities in alteration of Aslevels in the surrounding ecosystems: an encompassing risk assessment and evalua-tion of remediation strategies. J. Hazard. Mater. 182, 427–438.
Sutton, N.B., Van Der Kraan, G.M., Van Loosdrecht, M.C., Muyzer, G., Bruining, J., Schotting, R.J.,2009. Characterization of geochemical constituents and bacterial populations associatedwith arsenic mobilization in deep and shallow tube wells in Bangladesh. Water Res. 43,1720–1730.
Sylvia, D.M., Fuhrmann, J.J., Hartel, P.G., Zuberer, D.A., 2005. Principles and Applications ofSoil Microbiology, 2nd. ed. Prentice Hall Upper Saddle River, New Jersey.
Taghavi, S., Barac, T., Greenberg, B., Borremans, B., Vangronsveld, J., Van Der Lelie, D., 2005.Horizontal gene transfer to endogenous endophytic bacteria from poplar improvedphyto-remediation of toluene. Appl. Environ. Microbiol. 71, 8500–8505.
Takeuchi, M., Kawahata, H., Gupta, L.P., Kita, N., Morishita, Y., Ono, Y., Komai, T., 2007. Ar-senic resistance and removal by marine and nonmarine bacteria. J. Biotechnol. 127,434–442.
Tauriainen, S., Karp, M., Chang, W., Virta, M., 1997. Recombinant luminescent bacteria formeasuring bioavailable arsenite and antimonite. Appl. Environ. Microbiol. 63,4456–4461.
Thomas, D., Surdinkerjan, Y., 1997. Metabolism of sulfur amino acids in Saccharomycescerevisiae. Microbiol. Mol. Biol. Rev. 61, 503–512.
Tripathi, P., Dwivedi, S., Mishra, A., Kumar, A., Dave, R., Srivastava, S., Shukla, M.K.,Srivastava, P.K., Chakrabarty, D., Trivedi, P.K., Tripathi, R.D., Takeuchi, M., Kawahata, H.,Gupta, L.P., Kita, N.,Morishita, Y., Onoc, Y., Komai, T., 2007. Arsenic resistance and remov-al by marine and non-marine bacteria. J. Biotechnol. 127, 434–442.
Tripathi, R.D., Tripathi, P., Dwivedi, S., Dubey, S., Chatterjee, S., Chakrabarty, D.,Trivedi, P.K., 2012. Arsenomics, omics of arsenic metabolism in plants. Front.Physiol. 3, 275.
Trotta, A., Falaschi, P., Cornara, L., Minganti, V., Fusconi, A., Drava, G., Berta, G., 2006.Arbuscular mycorrhizae increase the arsenic translocation factor in the arsenichyperaccumulating fern Pteris vittata L. Chemosphere 65, 74–81.
Tsai, S.L., Singh, S., Chen, W., 2009. Arsenic metabolism bymicrobes in nature and the im-pact on arsenic remediation. Curr. Opin. Biotechnol. 20, 1–9.
Tsai, S.L., Singh, S., Dasilva, N.A., Chen, W., 2012. Co-expression of Arabidopsis thalianaphytochelatin synthase and Treponema denticola cysteine desulfhydrase for enhancedarsenic accumulation. Biotechnol. Bioeng. 109, 605–608.
Tu, C., Ma, L.Q., 2002. Effects of arsenic concentrations and forms on arsenic uptake by thehyperaccumulator ladder brake. J. Environ. Qual. 31, 641–647.
Tu, C., Ma, L.Q., 2003. Effects of arsenate and phosphate on their accumulation by anarsenic-hyperaccumulator Pteris vittata L. Plant Soil 249, 373–382.
Tu, C., Ma, L.Q., 2005. Effects of arsenic on concentration and distribution of nutrients inthe fronds of the arsenic hyperaccumulator Pteris vittata L. Environ. Pollut. 135,333–340.
Turpeinen, R., Pantsar-Kallio, M., Häggblom, M., Kairesalo, T., 1999. Influence of microbeson the mobilization, toxicity and biomethylation of arsenic in soil. Sci. Total Environ.236, 173–180.
Turpeinen, R., Pantsar-Kallio, M., Häggblom, M., Kairesalo, T., 2002. Role of microbes incontrolling the speciation of arsenic and production of arsines in contaminatedsoils. Sci. Total Environ. 285, 133–145.
U.S. Agency for Toxic Substances and Diseases Registry (ATSDR), 2005. Toxicological Pro-file for Arsenic. http://www.atsdr.cdc.gov/toxprofiles/tp2.pdf.
Virkutyte, J., Sillanpaa, M., Latostenmaa, P., 2002. Electrokinetic soil remediation— criticaloverview. Sci. Total Environ. 289, 97–121.
Wang, C.L., Lum, A.M., Ozuna, S.C., Clark, D.S., Keasling, J.D., 2001. Aerobic sulfide produc-tion and cadmium precipitation by Escherichia coli expressing the Treponemadenticola cysteine desulfhydrase gene. Appl. Microbiol. Biotechnol. 56, 425–430.
Wang, J.R., Zhao, F.J., Meharg, A.A., Raab, A., Feldmann, J., Mcgrath, S.P., 2002. Mechanismsof arsenic hyperaccumulation in Pteris vittata. Uptake kinetics, interactions withphosphate, and arsenic speciation. Plant Physiol. 130, 1552–1561.
Wang, J.Y., Zhang, D.S., Stabnikova, O., Tay, J.H., 2005. Evaluation of electrokinetic removalof heavy metals from sewage sludge. J. Hazard. Mater. B124, 139–146.
Wang, S., Mulligan, C.N., 2006. Natural attenuation processes for remediation of arseniccontaminated soils and groundwater. J. Hazard. Mater. B138, 459–470.
Wang, X., Ma, L.Q., Rathinasabapathi, B., Cai, Y., Liu, Y.G., Zeng, G.M., 2011. Mechanisms ofefficient arsenite uptake by arsenic hyperaccumulator Pteris vittata. Environ. Sci.Technol. 45, 9719–9725.
Wang, X., Rathinasabapathi, B., Oliveira, L.M.D., Guilherme, L.R.G., Ma, L.Q., 2012. Bacteria-mediated arsenic oxidation and reduction in the growth media of arsenichyperaccumulator Pteris vittata. Environ. Sci. Technol. 46, 11259–11266.
Wenzel, W.W., 2009. Rhizosphere processes and management in plant-assisted bioreme-diation (phytoremediation) of soils. Plant Soil 321, 385–408.
Wu, F.Y., Ye, Z.H., Wong,M.H., 2009. Intraspecific differences of arbuscularmycorrhizal fungi intheir impacts on arsenic accumulation by Pteris vittata L. Chemosphere 1258–1264.
Xie, Q.E., Yan, X.L., Liao, X.Y., Li, X., 2009. The arsenic hyperaccumulator fern Pteris vittata L.Environ. Sci. Technol. 43, 8488–8495.
Xie, Z., Luo, Y., Wang, Y., Xie, X., Su, C., 2013a. Arsenic resistance and bioaccumulation ofan indigenous bacterium isolated from aquifer sediments of Datong Basin, NorthernChina. Geomicrobiol J. 30, 549–556.

198 M. Roy et al. / Environment International 75 (2015) 180–198
Xie, W.Y., Huang, Q., Li, G., Rensing, C., Zhu, Y.G., 2013b. Cadmium accumulation in therootless macrophyte Wolffia globosa and its potential for phytoremediation. Int. J.Phytoremediation 15, 385–397.
Xiong, J., Wu, L., Tu, S., Nostrand, J.D.V., He, Z., Zhou, J., Wang, G., 2010. Microbial commu-nities and functional genes associated with soil arsenic contamination and the rhizo-sphere of the arsenic-hyperaccumulating plant Pteris vittata L. Appl. Environ.Microbiol. 76, 7277–7284.
Xiong, J., He, Z., Van Nostrand, J.D., Luo, G., Tu, S., 2012. Assessing the microbial commu-nity and functional genes in a vertical soil profile with longterm arsenic contamina-tion. PLoS ONE 7, e50507.
Xu, X.Y., McGrath, S.P., Zhao, F.J., 2007. Rapid reduction of arsenate in the medium medi-ated by plant roots. New Phytol. 176, 590–599.
Xue, P.Y., Yan, C.Z., 2011. Arsenic accumulation and translocation in the submerged mac-rophyte Hydrilla verticillata (L.f.) Royle. Chemosphere 85, 1176–1181.
Yamamura, S., Watanabe, M., Kanzaki, M., Soda, S., Ike, M., 2008. Removal of arsenic fromcontaminated soils by microbial reduction of arsenate and quinone. Environ. Sci.Technol. 42, 6154–6159.
Yan, X.L., Chen, T.B., Liao, X.Y., Huang, Z.C., Pan, J.R., Hu, T.D., Nie, C.J., Xie, H., 2008. Arsenictransformation and volatilization during incineration of the hyperaccumulator Pterisvittata L. Environ. Sci. Technol. 42, 1479–1484.
Yang, Q., Tu, S., Wang, G., Liao, X., Yan, X., 2012. Effectiveness of applying arsenate reduc-ing bacteria to enhance arsenic removal from polluted soils by Pteris vittata L. Int. J.Phytoremediation 14, 89–99.
Ye, W.L., Khan, A.M., Mcgrath, S.P., Zhao, F.J., 2011. Phytoremediation of arsenic contami-nated paddy soils with Pteris vittatamarkedly reduces arsenic uptake by rice. Environ.Pollut. 159, 3739–3743.
Yuan, C.G., Lu, X.F., Qin, J., Rosen, B.P., Le, X.C., 2008. Volatile arsenic species released fromEscherichia coli expressing the AsIII S-adenosylmethionine methyltransferase gene.Environ. Sci. Technol. 42, 3201–3206.
Zhang, X., Zhao, F.J., Huang, Q., Williams, P.N., Sun, G.X., Zhu, Y.G., 2009. Arsenic uptakeand speciation in the rootless duckweed Wolffia globosa. New Phytol. 182, 421–428.
Zhang, X., Uroic, M.K., Xie,W.Y., Zhu, Y.G., Chen, B.D., McGrath, S.P., Feldmann, J., Zhao, F.J.,2012. Phytochelatins play a key role in arsenic accumulation and tolerance in theaquatic macrophyte Wolffia globosa. Environ. Pollut. 165, 18–24.
Zhao, F.J., Mcgrath, S.P., Meharg, A.A., 2010. Arsenic as a food chain contaminant, mecha-nisms of plant uptake and metabolism and mitigation strategies. Annu. Rev. PlantBiol. 61, 535–559.
Zhu, Y.L., Zayed, A.M., Quian, J.H., De Souza, M., Terry, N., 1999. Phytoaccumulation oftrace elements by wetland plants. J. Environ. Qual. 28, 339–344.
Zhu, Y.G., Rosen, B.P., 2009. Perspectives for genetic engineering for the phytoremediationof arsenic-contaminated environments, from imagination to reality? Curr. Opin.Biotechnol. 20, 220–224.





























