Embed Size (px)
Citation preview

Send Orders for Reprints to [email protected]
Current Medicinal Chemistry, 2013, 20, 4575-4��� 4575
Antioxidative Peptides: Trends and Perspectives for Future Research
A.C. Freitas*,1,2, J.C. Andrade3, F.M. Silva3, T.A.P. Rocha-Santos1,2, A.C. Duarte2 andA.M. Gomes4
1ISEIT/ Viseu, Instituto Piaget, Estrada do Alto do Gaio, Galifonge, 3515-776 Lordosa, Viseu, Portugal; 2CESAM & Departamento de Química, Universidade de Aveiro, P-3810-193 Aveiro, Portugal; 3Centro de Investigação em Ciências da Saúde (CICS), Instituto Superior de Ciência da Saúde –Norte, CESPU. Rua Central de Gandra, 1317. 4585-116 Gandra PRD, Portugal; 4CBQF - Centro de Biotecnologia e Química Fina – Laboratório Associado, Escola Superior de Biotecnologia, Universidade Católica Portuguesa/Porto, Rua Dr. António Bernardino Almeida, 4200-072 Porto, Portugal
Abstract: In recent years, much attention has been given to dietary antioxidants, especially polyphenols. Several peptides derived from protein molecules have also been found to show antioxidant capacity along with other biological properties and thus there is an increasing interest in these compounds as health promoters. This review summarizes and discusses the main sources of antioxidative peptides with focus on food-derived peptides (animal, plant and marine sources), methods of preparation, antioxidant capacity evaluation as well as their proposed mechanisms of action. A discussion of the poten-tial health effects and comments on the different applications for these antioxidants and their potential research interest arealso subject of this review.
Keywords: Antioxidative peptides, food-derived peptides, preparation methods, antioxidant capacity evaluation, mechanism of action, applications, health.
1. INTRODUCTION
Over the last decades the perception, interest and re-search on bioactive compounds has been huge. In parallel to the increased potential of modern analytical strategies to deepen knowledge on bioactive compounds and their action in health, the awareness of people toward healthy and dis-ease risk prevention foods has concomitantly increased. It is well established that the lifestyle of the so-called modern societies, characterized by stress and inadequate diets among other factors, has been leading to severe disturbances in health of mankind. All over the world a large amount of studies and reports on bioactive compounds have been pro-duced, namely on bioactive peptides derived from food and other sources. Bioactive peptides are recognized as specific fragments of proteins that are inactive within the parent pro-tein, but once released impart positive functions or benefits to human health [1, 2]. According to Samaranayaja & Li-Chan [3], the inclusion of dietary antioxidants has been rec-ognized as a potential way to promote health since they are able to increase the body’s antioxidant load. Additionally, peptides with antioxidative properties have also been recog-nized with great potential to be used as nutraceuticals, as pharmaceuticals as well as substitutes for synthetic antioxi-dants [4].
Bioactive peptides, short-chain molecules based on less than 20 amino acid residues [4, 5], are considered physio-
*Address correspondence to this author at the ISEIT/ Viseu, Instituto Piaget, Estrada do Alto do Gaio, Galifonge, 3515-776 Lordosa, Viseu, Portugal; Tel: +351 232 910 017; Fax: +352 232 910 13; E-mail: [email protected]; [email protected]
logically active due to their biological properties such as blood pressure reduction (antihypertensive activity), scav-enging and/or neutralization of oxidative compounds (anti-oxidant activity), prevention of tumor formation (anticancer activity), inhibition of microbial growth (anti-bacterial activ-ity), blood lipid (cholesterol-lowering activity) and enhanced calcium and mineral absorption [6]. According to Mine et al.[7], biologically active peptides are some of the recom-mended functional ingredients, however it is still considered a challenge to know about the role and destiny of dietary antioxidative peptides once they enter the circulation system and cross the cell membrane. The majority of bioactive pep-tides are less than 1 kDA in weight and vary essentially from dipeptides to heptapeptides; the latter may reach 3 kDA [6]. A low molecular weight is a requirement because they need to be resistant to the digestion process throughout the gastro-intestinal tract (GIT) and be able to be absorbed intact into the blood system to reach the target organs.
A regular cell metabolism is dictated by an antioxidant-oxidation balance which is of utmost importance to the main-tenance of homeostasis of healthy cells, tissues and organs. Oxidation is a chemical phenomenon always present in the biological processes resulting from respiration, nutrient me-tabolism, detoxification phenomena among others. Therefore endogenous compounds with antioxidant properties (Fig. 1)are produced by the cells to protect them from oxidation damages. Oxidation stress is a major mediator of tissue and cell injuries such as those implicated in pathophysiological mechanisms of various diseases such as atherosclerosis, re-nal diseases, chronic inflammation, carcinogenesis, diabetes mellitus and other neurodegenerative disorders [9, 10].
1875-533X/13 $58.00+.00 © 2013 Bentham Science Publishers

4576 Current Medicinal Chemistry, 2013, Vol. 20, No. 36 Freitas et al.
When the antioxidant-oxidation balance is disturbed by several factors such as age, genetic susceptibility, stress, inadequate diets and other environmental factors, the pres-ence of exogenous antioxidative peptides in the cells can enhance their antioxidant capacity (Fig. 1). According to Karadag et al. [11], antioxidant capacity is related with com-pounds that are able to protect a biological system or food from the adverse effect of processes or reactions involving reactive oxygen species (ROS) or reactive nitrogen species (RNS) species. Studies on antioxidative peptides has indi-cated that their antioxidant action is related to their ability to quench and scavenge ROS, RNS and other free radicals as well as their ability to inhibit ROS-induced oxidation of bio-logical macromolecules such as proteins, lipids and DNA. The treatment with antioxidative peptides may provide pro-tection since DNA damage is a known causative factor of various chronic diseases including cancer, Alzheimer’s and Parkinson’s diseases [6, 12, 13]. The nature of the peptide is also of biological importance; for example, the ability of hydrophobic peptides to interact with cell membranes could provide in situ protection against lipid peroxidation and cel-lular damage [6]. According to Xiong [8], peptidyl antioxi-dants tend to have multifunctional properties, in opposition to non-protein antioxidants; besides the ability to stabilize free radicals and oxidation initiators some of the peptidyl antioxidants demonstrate antihypertensive, anticancer, an-timicrobial, immunomodulatory or opioid activities.
Many recent reviews have covered specific issues related with antioxidative peptides from different sources and pro-moting different mechanisms; based on such availability an overall perspective focusing, in particular, recent studies published over the last two years, supported by a discussion on which are the next steps in research is in order. Hence, the present review with special focus on food-derived antioxida-
tive peptides presents an overall view on their main sources and extraction/production procedures, discusses the advan-tages and disadvantages of the many methodologies avail-able for assessment of antioxidant capacity and discusses latest developments on proposed mechanisms of action that support potential health effects. Applications of these anti-oxidative peptides and future research trends will also be subject of this review. �
2. SOURCES OF ANTIOXIDATIVE PEPTIDES
The role of protein in the diet has been acknowledged worldwide and currently dietary proteins are one of the ma-jor sources of bioactive compounds with beneficial physio-logical impact on human’s and animal’s health. According to Kim et al. [4], dairy and marine-derived products seem to be the major sources of food-derived bioactive proteins and peptides but other sources such as eggs and plant-derived products including legumes (for example soy), nuts and ce-reals (for example wheat, rice, maize) are equally of impor-tance for providing bioactive peptides. Synthetic peptides in turn are also being produced especially for pharmaceutical purposes since they can exhibit therapeutic roles in the treatment of several diseases.
Some protein hydrolysates and peptides such as Tyr-Gly-Ser from peanut [14], Val-His-Leu-Lys-Pro from bovine whey [15], Val-Thr-Pro-Tyr from sea cucumber [16] and Glu-Cys-His, a synthetic peptide [17] have been shown to have potential antioxidant capacity, and will be discussed in the following sections.
Once confirmed the potential of bioactive peptides as an-tioxidative agents, it is important to consider other funda-mental aspects including i) bioavailability and ii) safety. Concerning bioavailability of bioactive peptides it is impor-
Fig. (1). Schematic representation of antioxidation-oxidation balance between oxygen and nitrogen reactive species and antioxidant cell ca-pacity that can be increased by exogenous antioxidative peptides. (Based on Kim et al. [4], Aluko [6], and Xiong [8]).
Regular cell metabolismRegular cell metabolism- Endogenous antioxidative compounds: selenium, �-tocophenol, vitamin C;
Enzymes: id di t t l l t thi
- Reactive oxygen species(ROS: .OH; .OOR, O2
-.)R i i i
Able to destroy
- Enzymes: superoxide dismutase, catalase, glutathione peroxidase;- Endogenous antioxidative peptides: glutathione, carnosine, anserine, belenine, etc.
E d ti id ti i id
- Reactive nitrogen species (RNS: ONOO-, NO) Balance
cellular components - Endogenous antioxidative aminoacids: histidine, cysteine, methionine, tyrosine.
Internal mechanisms tomitigate oxidation
- Age;
Factors leading to increases of ROS and RNS
Enhancement of balance between ROS/RNS levels and the antioxidative cell
Exogenous antioxidative
i) Scavenging or quenching of ROS/RNS and free radicals; ii) I hibiti f ROS i d d
- Genetic susceptibility;- Stress;- Inadequate diets;- Environmental factors
and the antioxidative cell capacity peptides
ii) Inhibition of ROS-induced oxidation of biological molecules (lipids, proteins, DNA)

Antioxidative Peptides: Trends and Perspectives Current Medicinal Chemistry, 2013, Vol. 20, No. 36 4577
tant to understand the relation between in vitro and in vivoantioxidant properties since it is well known that peptides upon oral ingestion are subjected to degradation and modifi-cation reactions through GIT, vascular system and liver [2]. Several reports indicate evidences that small peptides with 2-7 aminoacids or with less than 1 KDa are absorbed intact exhibiting their biological functions in their target organs or tissues [2, 6]. Peptide composition influences its resistance to digestive enzymes; apparently tripeptides with Pro-Pro at the C-terminal are resistant to proline-specific peptidases [18]. One of the major concerns associated with the use of pep-tides as antioxidants is the potential issue of allergenicity, especially with those that originate from proteins derived from dairy, soy, nuts, and eggs [4]. Since the majority of bioactive peptides are products of protein hydrolysis, this process should guarantee that the peptides associated with biological activity do not include the sequences of their par-ent protein responsible for allergic responses, in order to guarantee the safety of the consumers. Therefore, suitable research strategies based on toxicological studies using invitro and animal models, are fundamental and need to be carried out on new bioactive peptides to discard any possible cytotoxic effects.
2.1. Endogenous Antioxidative Peptides Among endogenous antioxidant compounds that comprise
the human antioxidant defence system several peptides are included. Probably the best known peptide is the glutathione (GSH) tripeptide (Table 1). This peptide is present in all ani-mal cells, quite often at high concentrations acting as an elec-tron donor protecting the cell from free radicals [19]. Besides antioxidant activity, GSH has an important role in a multitude of other cellular processes, including cell differentiation, pro-liferation, and apoptosis [20]. It becomes more and more evi-dent that disturbances in GSH homeostasis, such as its deple-tion, is associated with several disease states, including aging, cancer, cystic fibrosis, inflammatory, immune, cardiovascular, neurodegenerative and metabolic diseases [20].
Another well-known endogenous antioxidant is carnosine and related peptides, homocarnosine, anserine and balenine (Table 1), which are common dipeptides found in mammals, birds, and fish [21]. Carnosine is a multifunctional antioxi-dant that chelates pro-oxidative metals such as copper, can inactivate free radicals as well as form conjugates with alde-hyde lipid oxidation products that are potentially toxic [22]. Carnosine has been suggested to be useful for treating neu-rodegenerative diseases or preventing accumulation of aging features [23].
Cyclo-(His-Pro) (CHP) is a naturally occurring cyclic dipeptide consisting of histidyl and proline residues and is a metabolite of thyrotrophin releasing hormone, but it is also produced directly from amino acid or peptide sources [24]. It crosses the blood brain barrier and improves recovery in models of traumatic injury to the brain or spinal cord [25]. Invitro studies on rat pheochromocytoma PC12 cells have shown that it is a protective factor against stress stimuli and that it enhances the expression of small heat shock proteins and antioxidant protection at the cellular level [25]. Low concentrations (pmol/g) of this peptide can also be found in some foods such as tuna, dried shrimp, ham or bread [26].
The human tripeptide GHK isolated from human plasma has high affinity for copper ions and easily forms a copper complex or GHK-Cu. GHK possesses several regenerative and protective actions including antioxidant, anti-inflammatory, and wound healing properties. It is currently used in wound healing and anti-aging skin care, and recently, was proposed as a possible therapeutic agent against age-associated neurodegeneration and cognitive decline [27]. GHK has been proven to quench �,�-4-hydroxy-trans-2-nonenal, a toxic product of fatty acids’ lipid peroxidation that plays an important role in the pathogenesis of several age-related conditions including Alzheimer’s disease, neu-ropathy, and retinopathy [28]. GHK was also able to quench acrolein - another toxic product of lipid peroxidation in-
Table 1. Amino Acids Sequence of Endogenous Antioxidant Peptides
Peptide Sequence
Gluthatione �Glu-Cys-Gly
Carnosine �-Ala-His
Anserine �-Ala-1-CH3-His
Balenine �-Ala-3-CH3-His
Cyclo-(His-Pro) His-Pro
Human tripeptide GHK Gly-His-Lys
Luteinizing hormone-releasing hormone pyroGlu-His-Trp-Ser-Tyr-Gly-Leu-Arg-Pro-Gly-NH2
Leu-enkephalin Tyr-Gly-Gly-Phe-Leu
Angiotensin II Asp-Arg-Val-Tyr-Ile-His-Pro-Phe
Oxytocin Cys-Tyr-Ile-Gln-Asn-Cys-Pro-Leu-Gly-NH2
Vasoactive intestinal peptide His-Ser-Asp-Ala-Val-Phe-Thr-Asp-Asn-Tyr-Thr-Arg-Leu-Arg-Lys-Gln-Met-Ala-Val-Lys-Lys-Tyr-Leu-Asn-Ser-Ile-Leu-Asn

4578 Current Medicinal Chemistry, 2013, Vol. 20, No. 36 Freitas et al.
volved in the development of many age-related degenerative disorders [29].
There are several other endogenous peptidic molecules which primary discovered function was not related to anti-oxidant activity but that were shown later to also have this capacity. Such is the case of the secretory peptide hormones: luteinizing hormone-releasing hormone, enkephalin, angio-tensin II, and oxytocin (Table 1). These hormones were shown to scavenge free peroxyl radicals, prevent the oxida-tion of low-density lipoproteins, and inhibit lipid peroxida-tion in brain membranes [30].
The vasoactive intestinal peptide (VIP) is a 28-amino acid neuropeptide widely expressed throughout the body exerting a wide array of biological effects including the regu-lation of endocrine and exocrine secretions, cell proliferation and differentiation, and immune responses. Recently VIP was shown to also have a role in the oxidation stress control; VIP decreases the intracellular ROS levels reached by H2O2-induced oxidative stress in HK2 human renal cells [9]. In another study, VIP significantly protected neuronal tissue from oxidative stress and apoptosis by reducing lipid peroxi-dation and DNA fragmentation on 6-hydroxydopamine (6-OHDA) challenged rats [31].
2.2. Animal-Derived Peptides
The nutritional and functional properties of food proteins, namely those of animal origin, have long been recognized. In the last years there has been an increasing interest in bioac-tivities of peptides released from protein hydrolysis. These bioactive peptides have been shown to display a wide range of physiological functions including antihypertensive, anti-oxidative, opioid agonistic, immunomodulatory, antimicro-bial, osteoprotective, antithrombotic and antilipidemic ef-fects [32, 33].
Milk proteins are considered the most important source of bioactive peptides [34-36] and also one of the most stud-ied sources. The potential of the milk-derived bioactive pep-tides is already being explored as there are several products currently used as supplements in food, nutraceutical and pharmaceutical preparations [37].
In different research studies published in the last two years related with antioxidative peptides and summarized in Table 2, it can be seen that milk proteins are still the major protein source studied. Studies involved both isolated pro-teins (casein or whey proteins) [15, 39, 41] or whole milk [40, 42]. Some of the research works only covered the char-acterization of the antioxidative capacity of the hydrolysates while others probed further and identified the implicated peptides. It is interesting to note that one of the identified peptides, namely Val-His-Leu-Lys-Pro [15], protected fibro-blast MRC-5 cells against hydrogen peroxide-induced oxida-tive damage, showing in this way that the antioxidative properties placed in evidence by chemical assays were also verified at cellular level.
Meat or muscle proteins are also being increasingly stud-ied as sources of bioactive peptides (for reviews see Ber-nardini et al. [49] and Udenigwe and Howard [50]). Besides carnosine and anserine, other antioxidant peptides have been
found [33, 47, 51, 52]. In some cases, besides antioxidative capacity, anti-hypertensive capacity was also found [40, 47].
The in vivo physiological activities of antioxidative pep-tides from animal sources have been put into evidence re-cently. An antioxidative peptide isolated from duck skin by-products, was shown to protect the liver of rats against oxi-dative damage [45]. In another report, porcine plasma pro-tein hydrolysates were demonstrated to have hepatoprotec-tive and antioxidant effects on carbon tetrachloride-induced liver damage in rats [53].
Despite these new developments in animal-derived pep-tides physiological roles further studies still are required to better understand such role. Undoubtedly, food consumption trends and subsequent animal-derived by-product generation may contribute with interesting raw materials for research on new robust bioactive peptides with associated antioxidative properties. Not only identification and safety confirmation strategies are required thereof but, above all, study of the implicated physiological mechanisms of action is in order.
2.3. Plant-Derived Antioxidant Peptides
As previously mentioned all proteins may be considered as precursors of bioactive peptides and plant-derived proteins are no exception. Most of the main vegetable sources of die-tary proteins such as cereals, pulses or soybeans have been evaluated in the search for bioactive peptides.
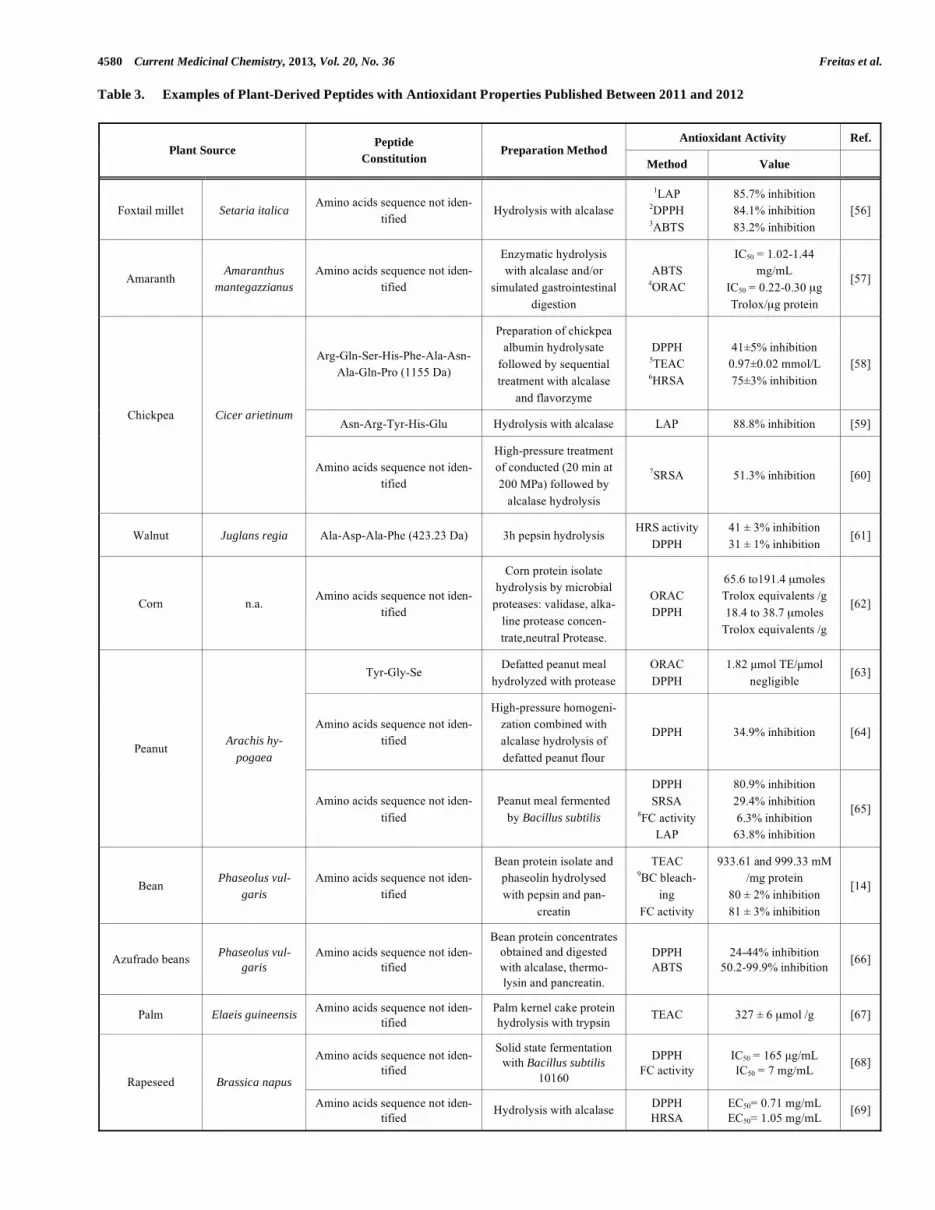
General reviews on food-derived antioxidative peptides have been presented recently [54, 55]. In (Table 3) are sum-marized research works on antioxidative peptides derived from plant proteins published from 2011 onwards. It can be observed that the main approach used to produce these pep-tides was via enzymatic hydrolysis. In some other cases, for example with chickpea or peanuts, high pressure treatments have been combined with the enzymatic hydrolysis and have resulted in an increase in the antioxidant ability of the hydro-lysed fractions obtained when compared to those obtained by enzymatic hydrolysis alone; no peptides were purified and indentified in both these studies [60, 64]. Identification of sev-eral antioxidative peptides were, however, reported by several authors that studied enzymatic hydrolysis of chickpea, walnut, peanut, Jujube fruit, wheat and barley reporting diverse anti-oxidant capacities via assays such as 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging capacity, Trolox equivalent antioxidant capacity (TEAC), hydroxyl radical (HR) scavenging activity, among others [58-61, 63, 70-72].
Three of the recently published reports used microbial fermentation for production of antioxidative protein fractions from rapeseed, diverse cereal flours and peanuts [65, 68, 73]. In one of these cases, fermentation of several cereal flours (wheat, rye, oat, spelt, barley, maize, rice, kamut) with se-lected Lactobacillus strains yielded 25 peptides of 8 to 57 amino acid residues in size [73]. It is interesting to note that in another work of the same group [74], Lactobacillus fer-mentation of cereal and non-conventional flours resulted in release of lunasin, a 43-amino acid peptide, with antioxidant, anti-inflammatory and anticancer properties [75, 76]. Com-pared to control doughs, the concentration of lunasin in-creased up to 2–4 times during fermentation. Lunasin can also be found naturally in several seeds including cereal grains and soybeans [76].

Antioxidative Peptides: Trends and Perspectives Current Medicinal Chemistry, 2013, Vol. 20, No. 36 4579
Table 2. Examples of Animal-Derived Peptides with Antioxidant Properties Published Between 2011 and 2012
Antioxidant Activity Ref. Animal Source
Peptide Constitution
Preparation Method Method Value
Human Milk Tyr-Gly-Tyr-Thr-Gly-Ala Ile-Ser-Glu-Leu-Gly-Trp
Hydrolysis with pepsin followed by pancreatin
1ORAC 5169 �M Trolox equiv/mmol and 4479 �lM Trolox equiv
/mmol [38]
Leu-Gln-Lys-Trp Leu-Asp-Thr-Asp-Tyr-Lys-Lys
Whey protein concentrate hydrolyzed using Corolase
PP® and thermolysin ORAC
2.57 mmol Trolox equiv/mg protein
[39]
Bovine whey
Val-His-Leu-Lys-Pro Whey protein hydrolysed
by alcalase 2FRAP
185.0 �M Trolox equiv Protec-tion of lung fibroblast MRC-5
cells against hydrogen peroxide-induced oxidative damage
[15]
Yoghurt Amino acids sequence not identi-
fied Fermentation by starter
cultures 3TEAC
219.3 – 2813 mM Trolox equiv/g
[40]
Ovine casein - Ovine caseinate hydrolysis with Bacillus spp. P7 pro-
tease
4ABTS 5DPPH
6FC activity
83.4% inhibition 30.9% inhibition 83.3% inhibition
[41]
Ewe cheese 17 peptides identified in water soluble extracts of Roquefort-
type cheese
Water soluble extracts Different cheese types
TEAC FC activity
7TBARS
0.71-2.02 mM TE 7.1-55.1% inhibition
26.0-51.8% inhibition [42]
Chicken breast Amino acids sequence not identi-
fied Hydrolysis by papain DPPH EC50 =1.28 mg/mL [43]
Hen egg white lysozyme Arg-Gly-Tyr Trp-Ile-Arg Val-Ala-Trp
simulated gastrointestinal digestion
DPPH 8LPIA
1.08,1.32 and 0.76% inhibition 75.3, 79.8 and 83.6% inhibition
[44]
Duck skin by-products His-Thr-Val-Gln-Cys-Met-Phe-
Gln (941.4 Da). Hydrolysis with pepsin
9HRSA DPPH
10SRSA
IC50= 32.6 �g/mL IC50= 22.7 �g/mL IC50= 49.8 �g/mL
[45]
Ostrich egg white (Struthio camelus)
Leu-Thr-Glu-Gln-Glu-Ser-Gly-Val-Pro-Val-Met-Lys (1317.65
Da)
Hydrolysis using various proteases (�-chymotrypsin, pepsin, trypsin and papain)
LPIA DPPH ABTS
FC activity HRSA
86.4% inhibition 81.0% inhibition 37.6% inhibition 20.0% inhibition 28.6% inhibition
[46]
Brisket muscle (Bos Taurus)
Fifteen peptides identified in the 10 kDa filtrate
Hydrolysis with papain DPPH FRAP
FC activity
15±5 % inhibition 7±3 �g Trolox equiv/5mg sam-
ple 82±3% inhibition
[47]
Bovine tendon collagen
Gly-Glu-Thr-Gly-Pro-Ala-Gly-Pro-Ala-Gly-Pro-Ile-Gly-Pro-
Val-Gly-Ala-Arg-Gly-Pro-Ala-Gly-Pro-Gln-Gly-Pro-Arg-Gly-Asp-Lys-Gly-Glu-Thr-Gly-Glu-
Gln
Hydrolysis of collagen with bacterial crude protease
DPPH FC activity
EC50= 388 �mg/mL EC50= 163 �mg/mL
[48]
1Oxygen radical absorbance capacity (ORAC) assay; 2Ferric reducing/antioxidant power (FRAP) assay; 3Trolox equivalent antioxidant capacity (TEAC);42,2’-azinobis(3-ethylbenzothiazoline-6-sulfonic acid (ABTS); 52,2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging activity assay; 6Fe2+ chelating (FC) activity; 7Thiobarbituric acid reactive substances (TBARS) assay; 8Lipid peroxidation inhibition activity (LPIA) assay; 9Hydroxyl radical scavenging activity (HRSA) assay; 10Superoxide radical scavenging activity (SRSA) assay.
2.4. Marine-Derived Peptides
Bioactive peptides isolated from fish, molluscs, crusta-ceans, macroalgae, microalgae proteins have been shown to
possess anticoagulant, anticancer, hypocholesterolemic, an-timicrobial, antioxidant activities among others [5, 77, 78]. In fact, peptides derived from marine sources have been ex-tensively researched due to the great potential of marine
4580 Current Medicinal Chemistry, 2013, Vol. 20, No. 36 Freitas et al.
Table 3. Examples of Plant-Derived Peptides with Antioxidant Properties Published Between 2011 and 2012
Antioxidant Activity Ref. Plant Source
Peptide Constitution
Preparation Method Method Value
Foxtail millet Setaria italica Amino acids sequence not iden-
tified Hydrolysis with alcalase
1LAP 2DPPH 3ABTS
85.7% inhibition 84.1% inhibition 83.2% inhibition
[56]
Amaranth Amaranthus
mantegazzianus Amino acids sequence not iden-
tified
Enzymatic hydrolysis with alcalase and/or
simulated gastrointestinal digestion
ABTS 4ORAC
IC50 = 1.02-1.44 mg/mL
IC50 = 0.22-0.30 �g Trolox/�g protein
[57]
Arg-Gln-Ser-His-Phe-Ala-Asn-Ala-Gln-Pro (1155 Da)
Preparation of chickpea albumin hydrolysate
followed by sequential treatment with alcalase
and flavorzyme
DPPH 5TEAC 6HRSA
41±5% inhibition 0.97±0.02 mmol/L 75±3% inhibition
[58]
Asn-Arg-Tyr-His-Glu Hydrolysis with alcalase LAP 88.8% inhibition [59] Chickpea Cicer arietinum
Amino acids sequence not iden-tified
High-pressure treatment of conducted (20 min at 200 MPa) followed by
alcalase hydrolysis
7SRSA 51.3% inhibition [60]
Walnut Juglans regia Ala-Asp-Ala-Phe (423.23 Da) 3h pepsin hydrolysis HRS activity
DPPH 41 ± 3% inhibition 31 ± 1% inhibition
[61]
Corn n.a. Amino acids sequence not iden-
tified
Corn protein isolate hydrolysis by microbial
proteases: validase, alka-line protease concen-trate,neutral Protease.
ORAC DPPH
65.6 to191.4 �moles Trolox equivalents /g 18.4 to 38.7 �moles
Trolox equivalents /g
[62]
Tyr-Gly-Se Defatted peanut meal
hydrolyzed with protease ORAC DPPH
1.82 �mol TE/�mol negligible
[63]
Amino acids sequence not iden-tified
High-pressure homogeni-zation combined with alcalase hydrolysis of defatted peanut flour
DPPH 34.9% inhibition [64] Peanut
Arachis hy-pogaea
Amino acids sequence not iden-tified
Peanut meal fermented by Bacillus subtilis
DPPH SRSA
8FC activity LAP
80.9% inhibition 29.4% inhibition 6.3% inhibition
63.8% inhibition
[65]
Bean Phaseolus vul-
garis Amino acids sequence not iden-
tified
Bean protein isolate and phaseolin hydrolysed with pepsin and pan-
creatin
TEAC 9BC bleach-
ing FC activity
933.61 and 999.33 mM /mg protein
80 ± 2% inhibition 81 ± 3% inhibition
[14]
Azufrado beans Phaseolus vul-garis
Amino acids sequence not iden-tified
Bean protein concentrates obtained and digested with alcalase, thermo-lysin and pancreatin.
DPPH ABTS
24-44% inhibition 50.2-99.9% inhibition
[66]
Palm Elaeis guineensis Amino acids sequence not iden-tified
Palm kernel cake protein hydrolysis with trypsin
TEAC 327 ± 6 �mol /g [67]
Amino acids sequence not iden-tified
Solid state fermentation with Bacillus subtilis
10160
DPPH FC activity
IC50 = 165 �g/mL IC50 = 7 mg/mL
[68] Rapeseed Brassica napus
Amino acids sequence not iden-tified
Hydrolysis with alcalase DPPH HRSA
EC50= 0.71 mg/mL EC50= 1.05 mg/mL
[69]

Antioxidative Peptides: Trends and Perspectives Current Medicinal Chemistry, 2013, Vol. 20, No. 36 4581
(Table 3) contd….
Antioxidant Activity Ref. Plant Source
Peptide Constitution
Preparation Method Method Value
Jujube fruit Zizyphus jujuba Val-Gly-Gln-His-Thr-Arg
(678.36 Da) GlyTrpLeuLys (482.27 Da)
Enzymatic hydrolysis using papain, trypsin and
a mixture of both
DPPH FC activity
EC50= 0.6 and 0.5 mg/ml
EC50= 1.18 and 0.86 mg/ml
[70]
Wheat Triticum spp. Val-His-His-His Wheat germ autolysis by
endogenous proteases DPPH
10CC activity76% inhibition 65% inhibition
[71]
Barley Hordeum vulgare
Gln-Lys-Pro-Phe-Pro-Gln-Gln-Pro Pro-Phe,
Pro-Gln-Ile-Pro-Glu-Gln-Phe, Leu-Arg-Thr-Leu-Pro-Met Ser-Val-Asn-Val-Pro-Leu
Extraction of barley glutelin and hydrolysis
with alcalase
DPPH SRSA
FC activity
61.9% inhibition 48.0 % inhibition 93.5 % inhibition
[72]
Cereal flours Twenty five peptides with 8 to 57 amino acid residues were
identified
Sourdough fermentation by selected lactic acid
bacteria LAP
Protection of oxida-tion-induced cells.
[73]
1 Linolenic acid peroxidation (LAP) assay; 22,2-diphenyl-2-picrylhydrazyl (DPPH) radical scavenging activity assay; 32,2’-azinobis(3-ethylbenzothiazoline-6-sulfonic acid (ABTS) radical scavenging capacity; 4Oxygen radical absorbance capacity (ORAC) assay; 5 Trolox equivalent antioxidant capacity (TEAC); 6Hydroxyl radical scavenging activity (HRSA) assay; 7Superoxide radical scavenging activity (SRSA) assay; 8Fe2+ chelating (FC) activity; 9�-carotene bleaching method; 10Cupper chelating (CC) activity.
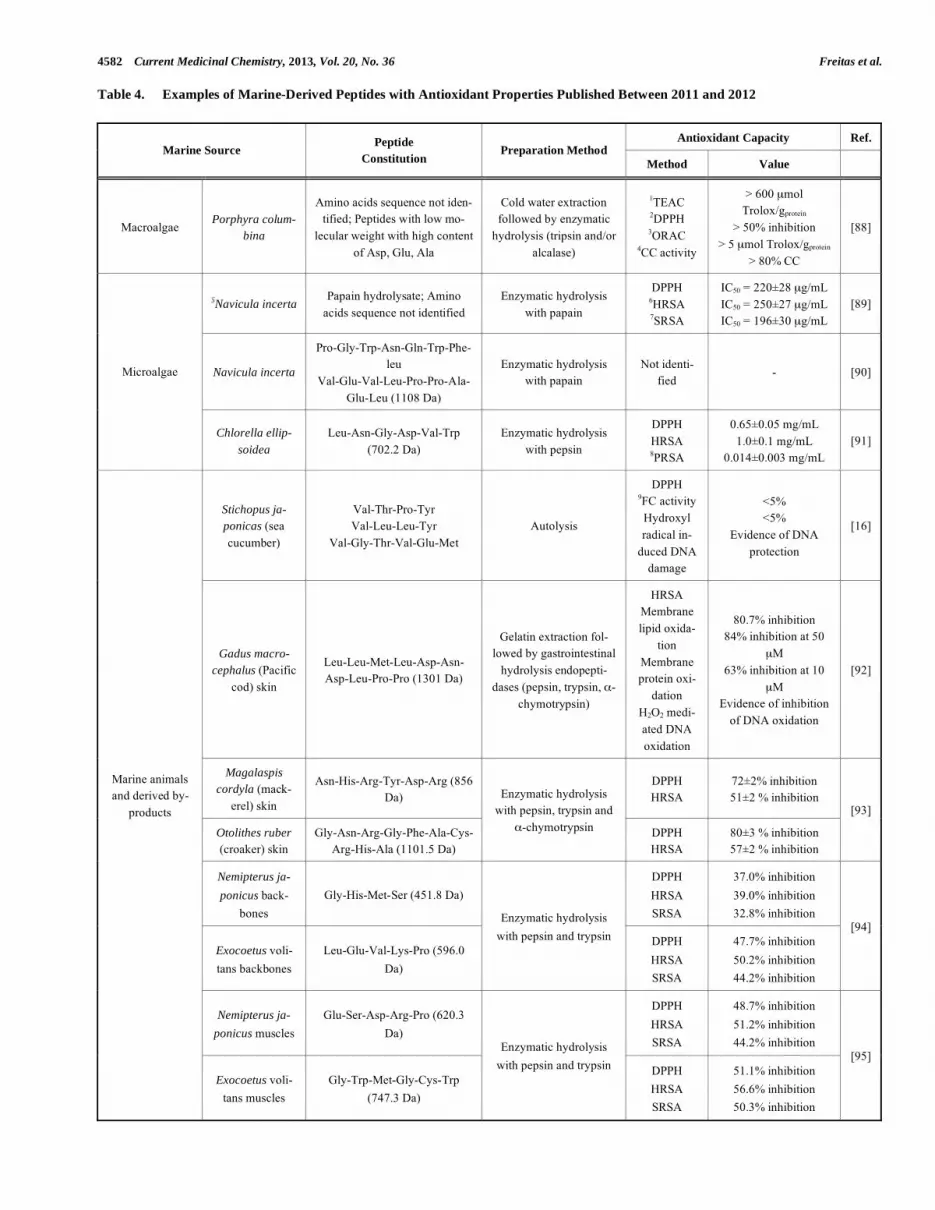
environment which is characterized by high diversity of organisms of interest. Proving this huge interest in marine-derived peptides are the several reviews published in be-tween 2010 and 2012 on proteins and bioactive peptides from marine algae [79, 80], bioactive peptides from marine organisms, in general [5, 81], fish-derived antioxidant pep-tides [82] as well as from marine processing waste and shell-fish [83].
According to Ngo et al. [84], among the marine fauna and flora, marine algae are considered a rich source of natu-ral antioxidants. Several authors believe that their potential could be derived from their protecting mechanisms to sur-vive in the marine habitat (light intensities fluctuations, dis-solved oxygen level fluctuations, UV radiations, tempera-ture, salinity, nutrient levels) which can lead to free radicals and other ROS formation in algae cells [79, 85]. Stimulation of enzymatic (ascorbate-glutathione cycle) and non-enzymatic (glutathione) antioxidant systems to scavenge and detoxify oxidative free-radicals due to UVB radiation in Ulva fasciata has been reported [86]. Several other reports confirm the role of many secondary metabolites with anti-oxidant properties produced from macro- and microalgae such as polyphenols, polysaccharides, pigmented compounds and peptides [87]. More focus will be directed to recently found peptides since it is the main interest of the present manuscript. In terms of macroalgae, numerous studies have been carried out to date reporting the antioxidant potential of macroalgae protein hydrolysates. An intensive description of antioxidant potential of diverse macroalgae protein hydro-lysates is given by Harnedy and Fitzgerald [80], however, almost no reports exist on purified peptides exhibiting anti-oxidant activity. In Table 4 a recent study is displayed on the antioxidatnt potential of enzymatic extracts rich in low mo-lecular weight peptides from Porphyra columbina but no purification and identification of amino acids sequence is described. Nevertheless, macroalgae peptides exhibiting
other biological properties such as ACE-inhibitory and anti-hypertensive activities have been identified namely from Palmaria palmate [98] as well as from Undaria pinnatifida,Hizikia fusiformis and Porphyra yezoensis described in Harnedy and Fitzgerald’s [80] review. From microalgae, antioxidative peptides have been identified and in Table 4two selected examples with demonstrated in vitro antioxidant activity are presented. The high protein content found in some species of microalgae makes them of potential interest for the exploration as a source of bioactive peptides.
The fish-derived protein hydrolysates have been deeply explored and studied [92-95]. According to these reports many by-products resulting from fish processing are able to furnish antioxidant and other bioactive peptides. Further-more, it is demonstrated that more investment should be ap-plied in the use of these by-products to produce healthy components. In an era where the natural sources have to be managed properly for sustainability purposes it is mandatory to explore their use for food and pharmaceutical purposes.
Several marine-derived antioxidant peptides have been described in literature. Table 4 lists some of the more recent examples published in the last two years from marine ani-mals and their by-products. Besides antioxidant capacity evidences from in vitro assays such as 2,2-diphenyl-1-picrylhydrazyl DPPH radical scavenging capacity and HR scavenging activity, two of the most used assays, evidence of DNA protection from oxidation damage [16, 92], activity against polyunsaturated fatty acids (PUFA) peroxidation using the �-linoleic acid model [93, 96] have also been re-ported. According to Himaya et al. [92], the peptide treat-ment in RAW264.7 cells up-regulated the m-RNA expres-sion of cellular antioxidative enzymes such as superoxide dismutase, glutathione peroxidase and catalase which is indi-cation of enhancement of the intracellular antioxidant mechanisms. In the majority of the peptides sequences de-scribed in Table 4, the presence of hydrophobic amino acids
4582 Current Medicinal Chemistry, 2013, Vol. 20, No. 36 Freitas et al.
Table 4. Examples of Marine-Derived Peptides with Antioxidant Properties Published Between 2011 and 2012
Antioxidant Capacity Ref. Marine Source
Peptide Constitution
Preparation Method Method Value
Macroalgae Porphyra colum-
bina
Amino acids sequence not iden-tified; Peptides with low mo-
lecular weight with high contentof Asp, Glu, Ala
Cold water extraction followed by enzymatic
hydrolysis (tripsin and/oralcalase)
1TEAC 2DPPH 3ORAC
4CC activity
> 600 �mol Trolox/gprotein
> 50% inhibition > 5 �mol Trolox/gprotein
> 80% CC
[88]
5Navicula incerta Papain hydrolysate; Amino
acids sequence not identified Enzymatic hydrolysis
with papain
DPPH 6HRSA 7SRSA
IC50 = 220±28 �g/mL IC50 = 250±27 �g/mL IC50 = 196±30 �g/mL
[89]
Navicula incerta
Pro-Gly-Trp-Asn-Gln-Trp-Phe-leu
Val-Glu-Val-Leu-Pro-Pro-Ala-Glu-Leu (1108 Da)
Enzymatic hydrolysis with papain
Not identi-fied
- [90]Microalgae
Chlorella ellip-soidea
Leu-Asn-Gly-Asp-Val-Trp (702.2 Da)
Enzymatic hydrolysis with pepsin
DPPH HRSA 8PRSA
0.65±0.05 mg/mL 1.0±0.1 mg/mL
0.014±0.003 mg/mL [91]
Stichopus ja-ponicas (sea cucumber)
Val-Thr-Pro-Tyr Val-Leu-Leu-Tyr
Val-Gly-Thr-Val-Glu-Met Autolysis
DPPH 9FC activity Hydroxyl radical in-
duced DNA damage
<5% <5%
Evidence of DNA protection
[16]
Gadus macro-cephalus (Pacific
cod) skin
Leu-Leu-Met-Leu-Asp-Asn-Asp-Leu-Pro-Pro (1301 Da)
Gelatin extraction fol-lowed by gastrointestinal
hydrolysis endopepti-dases (pepsin, trypsin, �-
chymotrypsin)
HRSA Membrane lipid oxida-
tion Membrane protein oxi-
dation H2O2 medi-ated DNA oxidation
80.7% inhibition 84% inhibition at 50
�M63% inhibition at 10
�MEvidence of inhibition
of DNA oxidation
[92]
Magalaspis cordyla (mack-
erel) skin
Asn-His-Arg-Tyr-Asp-Arg (856Da)
DPPH HRSA
72±2% inhibition 51±2 % inhibition
Otolithes ruber(croaker) skin
Gly-Asn-Arg-Gly-Phe-Ala-Cys-Arg-His-Ala (1101.5 Da)
Enzymatic hydrolysis with pepsin, trypsin and
�-chymotrypsin DPPH HRSA
80±3 % inhibition 57±2 % inhibition
[93]
Nemipterus ja-ponicus back-
bones Gly-His-Met-Ser (451.8 Da)
DPPH HRSA SRSA
37.0% inhibition 39.0% inhibition 32.8% inhibition
Exocoetus voli-tans backbones
Leu-Glu-Val-Lys-Pro (596.0 Da)
Enzymatic hydrolysis with pepsin and trypsin DPPH
HRSA SRSA
47.7% inhibition 50.2% inhibition 44.2% inhibition
[94]
Nemipterus ja-ponicus muscles
Glu-Ser-Asp-Arg-Pro (620.3 Da)
DPPH HRSA SRSA
48.7% inhibition 51.2% inhibition 44.2% inhibition
Marine animals and derived by-
products
Exocoetus voli-tans muscles
Gly-Trp-Met-Gly-Cys-Trp (747.3 Da)
Enzymatic hydrolysis with pepsin and trypsin DPPH
HRSA SRSA
51.1% inhibition 56.6% inhibition 50.3% inhibition
[95]

Antioxidative Peptides: Trends and Perspectives Current Medicinal Chemistry, 2013, Vol. 20, No. 36 4583
(Table 4) contd….
Antioxidant Capacity Ref. Marine Source
Peptide Constitution
Preparation Method Method Value
Magalaspis cordyla (mack-
erel) viscera Ala-Cys-Phe-Leu (518.5 Da)
Enzymatic hydrolysis with pepsin, trypsin and
�-chymotrypsin
DPPH HRSA
89.2% inhibition 59.1% inhibition
[96]
Tilapia (Oreo-chromis niloti-cus) frame pro-
tein
Asp-Cys-Gly-Tyr (456.1 Da) Asn-Tyr-Asp-Glu-Tyr (702.6
Da)
Enzymatic hydrolysis trypsin provided highest degree of hydrolysis and
antioxidant activity
HRSA IC50 =27.6 �g/mL IC50 = 38.4 �g/mL
[97]
1Trolox equivalent antioxidant capacity (TEAC); 22,2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging activity assay; 3Oxygen radical absorbance capacity (ORAC) assay; 4Copper-chelating (CC) activity; 5Benthic diatom; 6Hydroxyl radical scavenging activity (HRSA) assay; 7Superoxide radical scavenging activity (SRSA) assay; 8Peroxyl radical scavenging activity (PRSA) assay; 9Fe2+ chelating (FC) activity.
such as valine, leucine and proline is common which con-tributes greatly to their potential antioxidant activity [99] and have a major role in the inhibition of lipid peroxidation [93].
According to Kim and Wijesekara [5] and Harnedy and Fitzgerald [83] most of the biological activities of marine-derived bioactive peptides, including those with antioxidant properties, have been observed in in vitro or in animal model systems. Further research studies are needed to investigate their activities in vivo and in human subjects which would increase and confirm the real added-value of marine-derived bioactive peptides.
2.5. Synthetic Peptides
The therapeutic potential of several natural antioxidative peptides have led to attempts to synthesize peptides with improved characteristics related to activity, barrier perme-ability or digestion resistance.
Gu et al. [17] designed new glutathione (Glu-Cys-Gly) based peptides in order to obtain stronger antioxidant activity. The antioxidant amino acids, His and Tyr, were included in the new designed peptides ECH (Glu-Cys-His) and YECG (Tyr-Glu-Cys-Gly) and the antioxidant activity was evaluated using TEAC and ORAC assays. The protecting effect on hy-drogen peroxide (H2O2)-induced cytotoxicity in PC12 cells was also measured. The ECH peptide, with His located at the C-terminal position of the peptide, demonstrated good electron donating capacity, while with Tyr at the N-terminal position, YECG, exhibited strong oxygen radical absorbance capacity. This latter peptide also conferred best resistance to H2O2-induced cells necrosis.
Recently, a novel class of synthetic antioxidant peptides was reported designated as Szeto-Schiller (SS) peptides [100]. The structural motif of these SS tetrapeptide centres on alternating aromatic residues and basic amino acids, with 2,6-dimethyltyrosine residues, originally prepared to act as opioid analgesics. Physiologically, they are taken up by iso-lated cells and mitochondria due to their positive charge at physiological pH values and show antioxidant properties [101]. Three of the most extensively studied peptides are shown in (Fig. 2). Those that possess a tyrosine residue, such as SS-02 and SS-31, can dose-dependently scavenge hydro-gen peroxide (H2O2), hydroxyl radical, and peroxynitrite [100, 102]. By scavenging hydroxyl radical, SS-31 and SS-02 also inhibit lipid peroxidation [102]. SS20, which does not contain a tyrosine residue, lacks scavenging activity but can still reduce mitochondrial H2O2 production [100]. Their antioxidant action can be attributed to the tyrosine, or di-methyltyrosine (Dmt) residues. Tyrosine can scavenge oxyradicals forming relatively unreactive tyrosyl radicals, which can be followed by radical-radical coupling to give dityrosine, or react with superoxide to form tyrosine hydrop-eroxide. Dimethyltyrosine is more effective than tyrosine in scavenging of ROS. The specific location of the tyrosine or Dmt residue is not important as SS-31 was found to be as effective as SS-02 in scavenging H2O2 and inhibiting LDL oxidation. However, replacement of Dmt with phenylalanine in SS-02 eliminated the scavenging ability [103].
The synthetic tripeptide pinealon (Glu-Asp-Arg) which has been synthesized after analysis of amino acid composi-tion of bovine brain extracts was tested with different cells,
Fig. (2). Structures of antioxidant SS tetrapeptides. SS-02: Dmt-D-Arg-Phe-Lys -NH2; SS-20: Phe-D-Arg-Phe-Lys-NH2; SS-31: D-Arg-Dmt-Lys-Phe-NH2; Dmt. 2’,6’-dimethyltyrosine.
HN
NH2HNNH2
HN
NH2HNNH2
NHHOOC
O
NH
O
O
NHNH2
NHHOOC
O
NH
O
O
NHNH2
HO
O O
HO
O O
SS-02 SS-20 SS-31

4584 Current Medicinal Chemistry, 2013, Vol. 20, No. 36 Freitas et al.
cerebellar granule cells, neutrophils, and pheochromocy-tomas. This peptide demonstrated dose-dependent restriction of ROS accumulation, induced by oxidative stress stimulated by receptor-dependent or -independent processes [104].
Within the nutraceutical and pharmaceutical applications a more controlled production system is always required and based on the abovementioned very recent studies synthesis of peptides will continue to emerge as one of the most prom-ising areas of research as far as antioxidative capacity is con-cerned.
3. PREPARATION OF ANTIOXIDATIVE PEPTIDES
The production of antioxidative peptides has been searched intensively in the last years. The two main ap-proaches are based on enzymatic or chemical actions. The enzymatic action can be divided in enzymatic hydrolysis using exogenous or endogenous enzymes or enzymatic pathways occurring in processes such as microbial fermenta-tions. The enzymatic approach is considered by some authors as preferable for producing bioactive peptides with antioxi-dant properties because the enzymatic processes produce a well-defined peptides profile with high degree of productiv-ity with no undesirable by-products [4, 6]. The chemical action in turn, could be divided into chemical hydrolysis or chemical synthesis. Chemical acid or alkali digestion for example can produce some unwanted by-products such as lysinoalanine and amino acids with D-form with negative impact on human health or can destroy L-forms of amino acids required for nutritional and physiological properties of the peptides [6].
3.1. Enzymatic Preparation
According to the literature, and as previously mentioned, several food protein sources including milk, fish, seafood, soybean, egg, zein and wheat have been used to produce antioxidative peptides [2, 3] by enzymatic hydrolysis being inclusively the most commonly used method for the produc-tion of bioactive peptides. Proteolytic enzymes from mi-crobes, plants and animals can be used singly or as a combi-nation of two or more enzymes to cleave proteins or large polypeptides into small and more soluble bioactive peptides [4, 6]. In Table 5 are listed the different enzymes used for producing antioxidative peptides.
The use of exogenous enzymes is a preferred methodol-ogy over the use of the endogenous enzymes present in the food source itself (autolytic process), since a better control of the hydrolysis process is possible and enables to obtain a more consistent molecular weight profile and peptide com-position in a shorter time of hydrolysis. There are several commercially available enzymes derived from microorgan-isms (e.g. alcalase) and other enzymes from animal (e.g. pepsin) and plant sources (e.g. papain) that are food grade and have been used in the production of antioxidative pep-tides [3]. Enzymatic hydrolysis depends on several physico-chemical factors such as pH, temperature and substrate con-centration, which should be adjusted to optimal activity of the selected enzyme [112]. Furthermore, the choice of a suit-able enzyme and hydrolysis time is also crucial for obtaining peptides with antioxidant capacity. After selection of proper enzymes, several conditions can be tested in order to obtain
maximum recovery of antioxidative components. Therefore, the influencing factors in enzymatic preparation should be optimized preferably using statistical methods of optimiza-tion to find the optimal reaction conditions [112]. Addition-ally, the hydrolysates can be further processed using tech-niques such as ion-exchange membranes, ultra-filtration and column chromatographic methods in order to obtain a more uniform product with the desired range of molecular mass [3].
With a focus on the most recently published studies con-cerning antioxidative peptides production, Zhang and co-workers [59] used in their research, alcalase to produce the antioxidative peptide Asn-Arg-Tyr-His-Glu from chickpea (Cicer arietium L.) protein hydrolysates. The chickpea pro-tein hydolysates were separated using a Sephadex G-25 gel column and the fractions measured by spectrophotometric measurements at 280 nm. Then, the fraction with highest antioxidant capacity was pooled, and several times lyophi-lised and further loaded onto a semi-preparative C18 re-versed phase (RP) - high performance liquid chromatography (HPLC) column of an HPLC coupled to diode-array detector (DAD). After purification the structural properties including molecular mass, amino acid composition and sequence were determined and antioxidative capacity evaluated. Ren et al.[105] and Tang et al. [106] also used alcalase in the prepara-tion of Pro-Ser-Lys-Tyr-Glu-Pro-Phe-Val from grass carp muscle and Tyr-Ala and Leu-Met-Cys-His from zein hydro-lysates, respectively. Tang et al. [106] also verified that the antioxidant action of zein hydrolysate was not only related to the molecular mass, but also to the hydrophobicity character-istic of some amino acids present in the peptide sequence and showed that it is possible to produce natural antioxida-tive peptides from zein by enzymatic hydrolysis and differ-ent separation and purification techniques. Peptides with antioxidant capacity were prepared from human milk by Tsopmo et al. [38] by conducting a two-step gastric diges-tion using pepsin (firstly 37 ºC, pH 4, 30 min, secondly pH 6, 30 min) followed by pancreatin (pH 7). Incubation was carried out for 2h at 37 ºC, and enzymes were inactivated by heating for 15 min at 90 ºC. Purification and separation steps were also conducted using ultra-filtration and RP-HPLC. Chemical screening of the resulting fractions using the ORAC antioxidant assay yielded a peptide fraction with an-tioxidant capacity. Tandem mass spectrometry (MS) allowed the identification of twenty peptides, where the Tyr-Gly-Tyr-Thr-Gly-Ala and Ile-Ser-Glu-Leu-Gly-Trp peptides were the most active. Contreras et al. [39] used Corolase PP® and thermolysin to produce hydolysates from whey protein con-centrate with antioxidant capacity. The authors performed a central composite circumscribed design to optimise the main experimental variables involved in the process such as type of enzyme, and hydrolysis conditions. Therefore the enzyme to substrate ratio, time and temperature were evaluated using response surface methodology. The antioxidative peptides Leu-Gln-Lys-Tr and Leu-Asp-Thr-Asp-Tyr-Lys-Lys were identified using a RP-HPLC-MS/MS.
Depending on the raw material, the use of endogenous enzymes (autolysis) such as trypsin, chymotrypsin, pepsin, and other enzymes of digestive tract, lysosomal proteases or catheptic enzymes may also contribute to the discovery of antioxidative peptides. Hu et al. [71] incubated wheat germ

Antioxidative Peptides: Trends and Perspectives Current Medicinal Chemistry, 2013, Vol. 20, No. 36 4585
Table 5. Examples of Antioxidative Peptides Resulted from Enzymatic Hydrolysis
Antioxidative Peptide Source Antioxidant Capacity Method Enzymes Ref.
Pro-Ser-Lys-Tyr-Glu-Pro-Phe-Val
Grass carp muscle 1HRSA 2LPIA
Alcalase [105]
Tyr-Ala Leu-Met-Cys-His
Zein
3SRSA 4DPPH 5ABTS
Alcalase [106]
Pro-Ser-Tyr-Val Loach (Misgurnus anguillicauda-tus)
6CCLPIA HRSA DPPH
Papain [107]
Leu–Pro–Thr–Ser–Glu–Ala–Ala–Lys–Tyr Pro–Met–Asp–Tyr–Met–Val–Thr
Tuna (Thunnus obesus)7AADPPH
Orientase Protease XXIII
[108]
Phe-Arg-Asp-Glu-His-Lys-Lys Lys-His-Asp-Arg-Gly-Asp-Glu-Phe
Rice endosperm
SRSA HRSA DPPH Linoleic acid autoxidation
Alcalase, Chymotrypsin, Neu-trase, Papain and Flavorase
[109])
Val-His-His-His Wheat germ CC activity HRSA DPPH
Autolysis [71]
Leu-Gln-Lys-Trp Leu-Asp-Thr-Asp-Tyr-Lys-Lys
Whey protein concentrate 8ORAC Corolase PP® [39]
Gly-Pro-Glu-Pro-Thr-Gly-Pro-Thr-Gly-Ala-Pro-Gln-Trp-Leu-Arg
Sea Cucumber (Stichopus japoni-cus)
SRSA DPPH
Protease Papain Pepsin Trypsin
[110]
Tyr-Gly-Tyr-Thr-Gly-Ala Ile-Ser-Glu-Leu-Gly-Trp
Human milk ORACPepsin Pancreatin
[38]
Asn-His-Arg-Tyr-Asp-Arg Fish horse mackerel (Magalaspis cordyla) skin protein hydrolysate
Gly-Asn-Arg-Gly-Phe-Ala-Cys-Arg-His-Ala
Fish croaker (Otolithes ruber)skin protein hydrolysate
HRSA DPPH
Pepsin Trypsin �-chymotrypsin
[93]
Pro-Ala-Gly-Pro-Phe Rapeseed protein hydrolysate DPPH
Pectinase Cellulase �-glucanase Alcalase
[111]
Asn-Arg-Tyr-His-Glu Chickpea (Cicer arietium L.) SRSA HRSA DPPH
Alcalase [59]
1Hydroxyl radical scavenging activity (HRSA) assay; 2Lipid peroxidation inhibition activity (LPIA) assay; 3Superoxide radical scavenging activity (SRSA) assay; 42,2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging activity assay; 52,2’-azinobis(3-ethylbenzothiazoline-6-sulfonic acid (ABTS) radical scavenging capacity assay; 6Copper-chelating (CC) activity assay; 7Antioxidant activity (AA) by ferric thiocyanate method; 7Oxygen radical absorbance capacity (ORAC) assay.
and its proteins were hydrolysed by endogenous proteases to produce peptides. A resulting peptide Val-His-His-His with antioxidant action was purified using ultrafiltration, Se-phadex G-25 gel filtration column and consecutive chroma-tographic methods and identified using RP-HPLC connected on-line to an electrospray ionization MS. Further, the authors concluded that this process is an economical and convenient method to isolate antioxidative peptides from wheat germ.
3.2. Microbial Fermentation
According to Xiong et al. [8], the use of direct microbial fermentation in alternative to purified enzymes is an ap-proach used in many countries to produce health products. It is sought that fermentation may increase the nutraceutical value of foods and the storability time due to microbial pro-teases which are responsible for fragmentation of proteins to bioactive peptides [2]. Therefore microbial fermentation by

4586 Current Medicinal Chemistry, 2013, Vol. 20, No. 36 Freitas et al.
lactic acid bacteria or fungi, especially of milk or soybean, can be used as a source of proteolytic enzymes that can ef-fectively hydrolyse proteins.
Farvin et al. [113] studied the factors contributing to the oxidative stability of fish-oil-enriched yoghurt emphasizing the antioxidative effects of peptides released during yoghurt fermentation. According to evaluation of antioxidant capac-ity of lower molecular mass fractions 3-10 kDa and <3 kDa testing the inhibition of oxidation in liposome model system, DPPH radical-scavenging activity, iron-chelating (IR) activ-ity, and reducing power (RP), the higher oxidative stability of the yoghurt may result from the released antioxidative peptides during milk lactic acid fermentation and/or by the lower oxygen content of yoghurt which reduced the potential of fish oil oxidative deterioration. In order to pinpoint which fraction better suited this antioxidant potential, Farvin et al.[114] performed characterisation of peptide fractions and concluded that the 3-10 kDa fraction possessed many peptide sequences that were correlated with antioxidative peptides. Additionally, enriched yoghurt contained a considerable amount of free amino acids, such as histidine, tyrosine, threonine and lysine, which have also been reported to have antioxidant properties. Therefore the authors suggested that the naturally-occurring antioxidative peptides released dur-ing fermentation can be further used as an ingredient in en-riched foods with fish oils increasing their oxidative stability.
In alternative to the more common substrates for fermen-tation, He et al. [68] produced antioxidative peptides from rapeseed by solid state fermentation with Bacillus subtilis.After 3 days of fermentation the rapeseed peptides were pre-sent in five fractions within the 180-5500 Da size range; the most abundant amino acids in those peptides were glutamic acid, lysine and proline while serine, tryptophan and cysteine were present in least amounts. The authors also found a con-centration-dependent effect on antioxidant activities of rape-seed peptides and concluded that fermentation was a suitable process for converting the rapeseed protein into antioxidative peptides able to be used for formulation of functional foods and nutraceuticals; indeed these results open new avenues to be crossed as far as alternative substrates for fermentation are concerned.
3.3. Chemical Synthesis
In order to study and validate the functional properties of identified and purified bioactive peptides, several authors promote their chemical synthesis with identical or similar structures. Tsopmo et al. [38] synthesized eight small mo-lecular mass peptides from 4 to 6 amino acids mimicking some of those peptides found in human milk hydrolysates, in order to screen for their antioxidant properties using ORAC and linoleic acid emulsion assays. Other studies have been performed based on synthesized peptides to deepen knowl-edge about structure-function relationships of peptide se-quences and their biological properties. As already men-tioned Gu et al. [17] designed and synthesize two novel pep-tides ECH (Glu-Cys-His) and YECG (Tyr-Glu-Cys-Gly) based on endogenous glutathione peptide (Table 1) by solid-phase synthesis with the fluorenylmethoxycarbonyl (Fmoc)-strategy in order to study their antioxidant properties. This study was able to demonstrate that the presence of antioxi-
dant amino acids His/Tyr and their position in the peptide sequence was responsible for different antioxidant capacities between both novel and synthesized peptides as previously discussed in this review.
Advances in solid-phase peptide chemical synthesis with higher yields, higher acylation rates, lower levels of racemi-zation and higher solubility in solvents, in comparison to traditional solution-phase chemical synthesis has encouraged the chemical synthesis of large complex peptides as well as small peptides [115]. According to Zompra et al. [116], chemical synthesis of active peptides allows all possible se-quences, enables the introduction of biochemical and bio-physical probes, is faster and can be scaled-up more easily in comparison to biological methods.
Since peptide-based drugs could be an important serious approach for addressing new therapeutic challenges and con-sidering that the number of approved drugs has decreased in the last years, these facts have triggered a great interest in peptide-based drugs, some of them already available on the market [117]. Zompra et al. [116] in their review discuss the preparation of peptides at an industrial large-scale, the pep-tides approved by US FDA or European Medicines Agency (EMEA) available on the market as well as their characteris-tics and synthetic strategies for their industrial preparation. According to the authors more than 40 peptides are available on the pharmaceutical market for diverse treatment purposes. Vlieghe et al. [117] also discuss the chemical strategies to improve peptide biological, activity, specificity and stability and give important guidelines to be pursued in optimization strategies. According to these authors chemical optimization strategy of a therapeutic peptide should be based on struc-ture–activity relationship and/or quantitative structure–activity relationship of newly synthesized peptide deriva-tives. Such strategy may help improve their bioavailability, minimize elimination and biodegradation as well as increase selectivity or affinity to its receptor or target.
The identification and production of more active peptides for therapeutic applications may also be sought via the phage display technique. This technology has been frequently used over the past years as an important tool to display peptides or proteins on the surface of bacteriophages enabling the isola-tion of genes that code for specific peptides of interest by screening appropriate ligands. In this manner, the genes that code for the displayed peptide can be easily cloned from phages that express the peptide or protein, even in very small quantities. These identified peptides may target biomolecules either by antagonizing numerous receptors, or mimicking the binding sites of many physiological receptors [118]. Phage display libraries may be used to isolate specific antibodies against targeted antigens (used to improve vaccine efficacy) but may also be used to identify candidate peptides targeting antioxidative mechanisms or tumor suppressing potential. Paduano et al. [119] used this approach to screen a combina-torial phage display library to identify synthetic cyclic nonamer agonist peptides active as inhibitors of the Tyrosine Phosphatase Receptor and endowed with tumor suppressor activity. Xue et al. [118] used phage display to identify and isolate a multifunctional heptapeptide capable of inhibiting �-amyloid aggregates (A�, responsible for Alzheimer’s dis-ease pathogenesis) cytotoxicity in vitro, increasing the mi-

Antioxidative Peptides: Trends and Perspectives Current Medicinal Chemistry, 2013, Vol. 20, No. 36 4587
croglial phagocytosis of A�, decreasing the A�-induced gen-eration of ROS and NO and attenuating memory deficits in transgenic mice. Taddei et al. [120] had also previously ap-plied phage display technology to identify 25 peptides that bound human �� (40 or 42) using two random peptide li-braries containing 6-mer and 15-mer peptide inserts; among the 25 peptides identified two were shown to significantly reduce the A�42’s superoxide-dismutase-like activity. The invivo therapeutic potential remains to be determined. How-ever, these reports together with other studies not covered in this paper suggest that phage display technique may have great potential in identifying candidate peptides important for rational therapeutic drug development and encourage its use for the production of more selective and effective pep-tides that may be tested in preclinical disease models.
4. MECHANISM OF ACTION
Antioxidative peptides have demonstrated higher anti-oxidant properties than intact proteins and they are able to favor antioxidant cell capacity because, in general, they can act as: i) a radical scavenger; ii) a proton donor; iii) a metal-ion chelator and ferric-reducing capacity or as iv) a physical barrier to prevent ROS generation or their access to biologi-cal targets. Food-derived peptides can also enhance antioxi-dant cell capacity by induction of gene expression of proteins that are able to protect cellular components from oxidative stress-induced deterioration [33]. Antioxidant capacity of proteins and peptides has been reviewed by Elias et al. [78].
According to Aluko [6], there are two basic structural features that turn peptides into physiological antioxidative agents: i) presence of excess or donor electrons and/or aro-matic rings – the excess electrons are donated and used to neutralize free radicals while the aromatic ring ensures that loss of electrons does not turn peptide into a free radical; ii) possess a hydrophobic character in order to enable transloca-tion through cell membrane to cytoplasm and mitochondria where free radicals are generated as well as to increase their accessibility to hydrophobic targets such as polyunsaturated chain of fatty acids.
Many studies have been performed in the last years to understand how the predominance of particular amino acids in the peptides, their order along peptide chain as well as the chain length influences the antioxidative potential of the pep-tides [8]. In Table 6 it is summarized the role of amino acid composition of food-derived peptides in their antioxidant properties which also depends on the nature of ROS/free radical and on the reaction medium [6, 33]. The chemical and physical properties arising from amino acids sequence in the peptide are, at least in part, responsible for the higher ability of peptides to act as antioxidant agents in comparison to free amino acids; for example the stability of resultant peptide radical does not allow the initiation or propagation of subsequent oxidative reactions after its formation [78]. Al-though many studies demonstrate the importance of amino acid composition, sequence and size toward the antioxidative potential of peptides including those by Chen et al. [122] and Saito et al. [127], who studied 28 and 114 peptides respec-tively, and more recently by Tang et al. [106], more investi-gation is still needed to fully understand the influence of amino acid sequence arrangement on the antioxidative pep-
tide properties [6]. According to Samaranayaka and Li-Chan [3] the studies should first identify the target use of a specific antioxidative peptide of interest followed by design of the assay and selection of proper model systems considering the mechanisms of action in the predicted application, in order to be able to determine the real antioxidative potential.
5. EVALUATION OF ANTIOXIDANT CAPACITY
The diversity of developed assays for antioxidant evalua-tion makes the interpretation of many reported results a complex task. The observation of diverse expression of re-sults from the different antioxidant capacity evaluation as-says reported in (Tables 2, 3 and 4) shows that comparisons between studies are remarkably difficult because no direct relationship can be postulated between values expressed as either percentage of inhibition or values of IC50 or even as �moles Trolox equivalents for the same assay such as the DPPH radical scavenging capacity assay (Table 2). How-ever, no single assay seems to be able to accurately reflect the mechanism of action of all radicals and antioxidants es-pecially in complex systems like foods or cells because the response of antioxidants to different radicals or oxidants may be totally different; some antioxidants may not be particu-larly good quenchers of peroxyl radicals but may be good singlet-oxygen scavengers [11, 128].
Karadag et al. [11] presented a complete review on the most widely used methods based on chemical assays to de-termine antioxidant capacity, describing the general princi-ples, applications as well as their advantages and limitations. According to these authors the major antioxidant capacity methods are based on hydrogen atom transfer or on electron transfer reaction-based methods where the bond dissociation energy and ionization potential are the two major facts be-hind the mechanism and efficiency of the evaluated antioxi-dants. There are plenty of methods that differ from each other in terms of reaction mechanism, oxidant and target, reaction conditions and correspondent expression of results; ORAC, Total radical trapping antioxidant parameter (TRAP), �-Carotene or Crocin-Blaeching assay, Total phe-nol assay, TEAC, DPPH radical scavenging capacity assay, Ferric reducing antioxidant power (FRAP), Total antioxidant potential assay using Cu2+ also known as CUPRAC, Total oxidant scavenging capacity (TOSC) assay and other meth-ods such as chemiluminescence, photochemiluminescence as well as assays to measure other ROS/RNOS species are fully described by Karadag and co-workers [11].
Although antioxidative protein hydrolysates or peptides are indeed potential sources to control various oxidative processes, it is difficult to compare results from various stud-ies because of the diversity of in vitro assays and the incon-sistency of conditions used to evaluate their antioxidative capacity [3, 79]. According to Samaranayaka and Li-Chan [3], specific methods have not yet been developed to deter-mine the antioxidative capacity of peptides. Therefore, in most of the studies the evaluation of antioxidant capacity is done by various methods of measurement classically applied to non-peptidic antioxidants; this fact becomes evident in the examples displayed in Tables 2-5. ORAC, DPPH radical scavenging capacity assay, TEAC as well as radicals scav-enging activities are the most used assays to evaluate

4588 Current Medicinal Chemistry, 2013, Vol. 20, No. 36 Freitas et al.
Table 6. The Influence of Amino Acid Composition Food-Derived Peptides in Their Antioxidant Properties
Peptide Constitution
Amino Acid Type Amino Acids Achievements References
Electrically charged - His
Imidazole group of histidine residue contribute to: - Increasing of metal ion chelation; - Quenching of active oxygen; - Scavenging of hydroxyl radical. Imidazole group are able to participate in hydrogen atom and single electron trans-fer reactions in order to neutralize free radicals or bind metal ions.
[6, 33]
-His Histidine-containing compounds were related to antioxidation abilities. [121]
- Pro, Leu Addition of hydrophobic amino acids to the dipeptide His-His N-terminus in-creases its antioxidant properties and promotes synergy with non-peptide antioxi-dants.
[6, 122, 123]
- Trp Hydroxyl scavenging ability. [6]
-Val, Leu; Gly
Representing about 55% of the sequence may contribute to lipid peroxidation inhibition by increasing solubility of peptides in lipid phase; the presence of pep-tides in the water-lipid interface facilitates the scavenging of generated free radi-cals.
[124]
Presence of hydrophobic amino acids
-Gly, Pro High antioxidant levels in marine gelatin peptides rich in Gly and Pro. [99]
Sulfur containing amino acids
- Met, Cys Proton-donating ability of sulfhydril group enables scavenging of radicals; Residues of Cys in peptides can be used for glutathione syntheses (�-L-glutamyl-L-cysteinylglycine).
[6, 125]
Acidic amino acids - Glu, Asp
Electron-donating ability enables the scavenging radicals and ferric reducing ca-pacity. Metal-ion binders that limit the propagation of lipid peroxidation caused by transi-tional metal ions and change the redox cycling capacity.
[6, 8]
Amino acids with electron-dense aro-matic rings
- Phe, Tyr The rings enables the chelation of pro-oxidant metal ions; Phenylalanine is able to scavenge hydroxyl radicals to form more stable para-, meta- or ortho- substituted hydroxylated derivatives.
[6, 126]
Diverse - His, Tyr, Trp, Met, Cys, ProCorrelated with radical quenching of several antioxidative peptides derived from different food sources namely whey and soy.
[8]
antioxidant capacities of peptides independently of its source and applicability. The use of model systems providing the chemical, physical and environmental conditions mimicking the matrix involving antioxidative peptides is of utmost im-portance to validate their antioxidant capacity [129]. It must also be emphasized that in vitro measurements of antioxidant capacity cannot be directly related to their potential in vivo.Extrapolation of the results from in vitro chemical assays to the food or biological system should be done with caution.
Several factors such as bioavailability, reactivity, stabil-ity in carrier food, fate of peptides during GI digestion as well as permeability through cellular membranes have im-pact on the biological efficacy of antioxidative peptides [2, 3]. For example, it is advisable to use biomarkers of lipid and protein peroxidation as well as of DNA oxidative damage to assess oxidative stress in vivo [130].
6. POTENTIAL HEALTH EFFECTS OF ANTIOXIDA-TIVE PEPTIDES
As previously discussed, oxidative stress can impart negative effects constituting a possible factor for the initia-
tion or development of chronic or degenerative diseases as has been indicated and discussed throughout this review. Since the formation of cancer cells can be directly induced by free radicals, natural compounds with antioxidative prop-erties can be seen as chemopreventive agents in the treatment of cancer. Bioactive peptides and amino acids may act as alternative molecules to small molecular drugs possessing the advantage of high bioavailability and bio-specificity for the targets in comparison to conventional drugs; low toxicity, structural diversity and no accumulation of peptides or amino acids make them eligible for therapeutic purposes [79].
The in vitro biological assays for the antioxidative capac-ity evaluation (Tables 2-4) as well as in vivo studies involv-ing antioxidative peptides has shown their great potential to prevent or minimize the oxidative damages caused by ROS or RNOS. According to Mendis et al. (2005) two purified antioxidant peptides (Phe-Asp-Ser-Gly-Pro-Ala-Gly-Val-Leu and Asn-Gly-Pro-Leu-Gln-Ala-Gly-Gln-Pro-Gly-Glu-Arg) from jumbo skid skin gelatin enhanced the viability of radical-mediated oxidation-induced human lung fibroblasts

Antioxidative Peptides: Trends and Perspectives Current Medicinal Chemistry, 2013, Vol. 20, No. 36 4589
radical by 77 and 72%, respectively. According to Ngo et al.[84], radical scavenging compounds such as bioactive pep-tides can be used as functional ingredients to reduce cancer formation. Suarez-Jimenez et al. [131] recently reviewed the potential role of bioactive peptides and depsipeptides in the prevention and treatment of cancer. A peptide (Val-Glu-Cys-Tyr-Gly-Pro-Asn-Arg-Pro-Gln-Phe) isolated from algae pro-tein waste has demonstrated strong dose-dependent antipro-liferation and promotion of cell cycle arrest in human gastric cancer cells besides higher antioxidant capacity than natural antioxidants [132]. On a different tone, marine oligopeptide preparations from Chum salmon have been shown to in-crease lymphocyte proliferation, natural killer cell activity as well as the secretion of cell cytokines in mice when intro-duced in murine diets and may therefore enhance the im-mune function in its hosts, including cellular immunity [133].
7. POTENTIAL APPLICATIONS
Antioxidative peptides derived from diverse sources can be used as functional ingredients to control oxidation dete-rioration of foods or as versatile raw materials for producing functional foods, nutraceuticals as well as pharmaceuticals and according to some authors even for cosmeceutical prod-ucts [79, 84].
It is well known that lipid oxidation by ROS causes dete-rioration of some foods resulting in a decrease of their nutri-tional value and impacts negatively in terms of safety and quality, in particular affects appearance of the food [84]. The use of synthetic antioxidants such as propyl gallate, or buty-lated hydroxytoluene have has been used to retard the per-oxidation process in food systems as well as in pharmaceuti-cals [4, 84]. Since the use of synthetic antioxidants can cause adverse effects on health and their use must be minimal and controlled, the need and search for natural antioxidants as safer alternatives to synthetic compounds is of utmost impor-tance in the food industry. In fact, the use of natural antioxi-dative peptides to control oxidation processes in food prod-ucts is an application of utmost interest especially becausethe use of natural antioxidants available in food has attracted significant interest due to their presumed safety, nutritional, and therapeutic values. Bioactive peptides generated using food-grade enzymes (proteinases and peptidases) have prom-ising application as natural functional food ingredients [80]. In cooked ground beef caseinophosphopeptides and casein hydrolysates inhibited 75 and 39% of thiobarbituric acid reactive substances formation after 4 days of storage [134]. Reduction of lipid oxidation during 6 weeks cold storage (4ºC) and 8 months frozen storage (-18 ºC) of smoked salmon with a brine solution containing salmon fish protein hydrolysate was also observed [135].
Although the antioxidant properties of some peptides confer them a great potential as food additives or contribute to create new functional foods or nutraceuticals, we have to be aware that in some cases they may not be totally suitable for these applications. In its use some crucial requisites should be evaluated and guaranteed such as: i) absence of allergenic potential, ii) of safe use, iii) effective at low con-centration, and iv) be compatible with food and nutraceutical matrices presenting organoleptic concerns related with color,
odor, or taste. The bitterness of some protein hydrolysates may limit the use of some bioactive peptides as food addi-tives. Additionally, it is known that the bioactivity of the peptides could be affected by food processing conditions or by components present in the matrices. However the use of natural antioxidative peptides should be encouraged because they are considered mainly toxicologically safe, with fewer side effects than the synthetic compounds and are better ab-sorbed in the intestinal tract [3, 136].
Nowadays the non-availability of large scale technologies and high cost of purification techniques are limiting factors to the commercialization of food derived bioactives; whole hydrolysates with unpurified bioactive peptides are the main components of functional foods [136]. According to Schaaf-sma [137], various vegetable protein hydrolysates are being incorporated into specific foods in the United States of America as food additives as well as food ingredients in sev-eral countries. In turn, pure peptides have found more appli-cations for pharmaceutical products.
Several functional foods and natural health products are available on the market but the majority of these products bet on antihypertensive and antihypercholesterolemic effects with low molecular mass food-derived peptides such as Cal-pis AMEEL S (Japan) or Evolus (Finland) fermented milks [115]. Among the 14 functional foods or food ingredients reported by Hartmann and Meisel [138] none have antioxi-dant health claims. Harnedy and Fitzgerald [83] in their re-view on bioactive peptides from marine sources presented 12 commercially available marine protein hydrolysate and pep-tide products where only one product (Fortidium Li-quamen®), a white fish autolysate, is commercialized as hav-ing multifunctional effects including an antioxidant potential. Therefore it seems that not much has been invested in prod-ucts with antioxidative peptides; this can be further sup-ported by Samaranayaka and Li-Chan [3] who state that only a limited number of studies have been performed to assess the biological antioxidative potential of peptide hydrolysates or isolated peptides. This context may also explain the rea-son for the European Food Safety Authority’s position con-cerning antioxidants in general in terms of missing informa-tion to uphold health claims concerning antioxidative poten-tial.
Cosmeceuticals containing antioxidative compounds are another application that has been explored. Cosmeceuticals are topical products with benefits for human skin such as improvement of appearance and anti-ageing properties [139]; antioxidative compounds may neutralize free radicals lessen-ing or preventing the signs of aging skin [3]. The role of topical peptides in preventing or treating aged skin is re-viewed by Gorouch and Maibach [140]. The use of antioxi-dative peptides in cosmeceuticals is however not yet very generalized; as in food products bioactive peptides are used in skin care products with other purposes than to prevent oxidation. According to Zhang and Falla [141], 25 peptides are currently used in cosmetic products to modulate cell pro-liferation, cell migration, inflammation among others. How-ever, the use of antioxidative peptides to protect skin from endogenous or exogenous oxidative aggressions should be better explored; for example silk protein, sericin, extracted

4590 Current Medicinal Chemistry, 2013, Vol. 20, No. 36 Freitas et al.
from Bombyx mori silkworm can reduce UV-B induced acute damage and tumor promotion in mouse skin [142].
According to Hartmann and Meisel [138], food-derived peptides also have great potential to be incorporated in pharmaceutical products. However the chemical synthesis of peptides is still considered as gold standard in industrial pep-tide preparation. Thirty five peptide drugs are manufactured using a synthetic approach [116] and according to these authors the native isolation and enzymatic approach are mostly used for the production of short peptide sequences for food additives. The plausibility of manufacturing pharma-ceutical-grade bioactive peptides from food proteins was reviewed by Agyei and Danquah [136] whereas science and market of synthetic therapeutic peptides was reviewed by Vlieghe et al. [117]. The high biological activity, high speci-ficity and low toxicity are characteristics that encourage the pharmaceutical companies to develop peptide drugs to treat several diseases [116].
8. CONCLUSIONS AND FUTURE TRENDS
Research continues to reveal a plethora of antioxidative peptides derived from different sources with the potential of positively impacting on human health. These peptides are made available through proteolysis by digestive enzymes, via microbial fermentation or via synthetic production strate-gies. Several of these peptides have been shown to exhibit a range of antioxidative mechanisms via both in vitro and invivo studies and much has been hypothesized concerning their physiological importance and influence on health, in particular prevention against oxidation-related diseases. Nonetheless, no standardised procedure is yet available for the evaluation of antioxidant capacity of such peptides and comparison among different peptides from different sources is a difficult task; furthermore, it is well-established that invitro antioxidant activity cannot be directly correlated to health-promoting activity. Studies on direct antioxidant ac-tivity of bioactive peptides in the human body are lacking yet are essential to establish such relationships. Although of spe-cial interest for different types of applications including food quality promotion, cosmetics and pharmaceutical prepara-tions, these have not been as explored as expected and a long course still has to be pursued if antioxidative peptides are to conquer the added-value market either as functional foods, nutraceuticals or pharmaceuticals. Independent of the appli-cation, be it food or drug, hurdles such as stability in product or biostability of peptides, lack of oral bioavailability, resis-tance to proteolytic attack need to be overcome to guarantee the physiological effectiveness of the antioxidative peptides. When pharmaceutical applications are being considered studies need to probe further on the pharmacokinetics (ab-sorption, distribution, metabolism and excretion) and phar-macodynamics of the different compounds by (i) establishing effective dose, route and mode of administration including improved delivery systems; (ii) overcoming technological challenges during formulation; (iii) pinpointing mechanism of action besides safety properties; (iv) regulatory concerns; (v) quality control and (vi) post-market surveillance. None-theless, with the onset of European legislation concerning health claims associated with functional foods since 2006, the development of a functional food also needs to undergo studies that are quite close to those demanded by a pharma-
ceutical product; although not as extensive and overwhelm-ing some of the efficacy, delivery and safety issues are also required supported by an integrated scientific basis.
CONFLICT OF INTEREST
The authors confirm that this article content has no con-flicts of interest.
ACKNOWLEDGEMENTS
This work was supported by Portuguese Science Founda-tion (Fundação para a Ciência e Tecnologia), through indi-vidual research grants references SFRH/BPD/73781/2010 and SFRH/BPD/65410/2009 under QREN - POPH funds, co-financed by the European Social Fund and Portuguese National Funds from MCTES.
DISCLOSURE
This manuscript has been submitted for publication in Current Medicinal Chemistry. It is not to be reproduced or cited without the written permission of the authors.
REFERENCES [1] Kanan, A.; Hettiarachchy, N.; Marshall, M. In: Bioactive Food
Proteins and Peptides; Hettiarachchy N.S., Ed.; CRC Press Taylor & Francis Group: Boca Raton, 2012; pp. 1-28.
[2] Sarmadi, B.H.; Ismail, A. Antioxidative peptides from food pro-teins: A review. Peptides, 2010, 31, 1949–1956.
[3] Samaranayaka, A.G.P.; Li-Chan, E.C.Y. Food-derived peptide antioxidants: a review of their production, assessment and potential applications. J. Functional Foods, 2011, 3, 229-254.
[4] Kim, S.; Wijesekara, I.; Park, E.Y.; Matsumura, Y.; Nakamura, Y.; Sato, K. In: Bioactive Food Proteins and Peptides; Hettiarachchy N.S., Ed.; CRC Press Taylor & Francis Group: Boca Raton, 2012;pp. 97-115.
[5] Kim, S.; Wijesekara, I. Development and biological activities of marine-derived bioactive peptides: A review. J. Functional Foods,2010, 2, 1-9.
[6] Aluko, R.E. Functional Foods and Nutraceuticals; Springer; New York, 2012.
[7] Mine, Y.; Li-Chan, E.C.Y.; Jiang, B. In: Bioactive Proteins and Peptides as Functional foods and Nutraceuticals; Mine, Y.; Li-Chan, E.; Jiang, B., Ed.; Wiley-Blackwell: Iowa, 2010; pp. 5-11.
[8] Xiong, Y.L. In: Bioactive Proteins and Peptides as Functional foods and Nutraceuticals; Mine, Y.; Li-Chan, E.; Jiang, B., Ed.; Wiley-Blackwell: Iowa, 2010; pp. 29-42.
[9] Vacas, E.; Bajo, A.M.; Schally, A.V.; Sánchez-Chapado, M.; Prieto, J.C.; Carmena, M.J. Antioxidant activity of vasoactive intes-tinal peptide in HK2 human renal cells. Peptides, 2012, in press; doi:10.1016/j.peptides.2012.09.011.
[10] Hernández-Ledesma, B.; Hsieh, C.; Lumen, B.O. Antioxidant and anti-inflammatory properties of cancer preventive peptide lunasin in RAW 264.7 macrophages. Biochem. Biophysical Res. Comm., 2009, 390, 803–808.
[11] karadag, A.; Ozcelik, B.; Saner, S.; Review of methods to deter-mine antioxidant capacities. Food Anal. Methods, 2009, 2, 41-60.
[12] Lee, H.P.; Zhu, X.; Casadeus, G.; Castellani, R.J.; Nunomura, A.; Smith, M..; Lee, H.; Perry, G. Antioxidant approaches for the treatment of Alzheimers disease. Expert Review Neurotherapeutics,2010, 10, 1201-1208.
[13] Borek, C. Dietary Antioxidants and human cancer. Integr. Cancer Ther., 2004, 3, 333-341.
[14] Carrasco-Castilla, J.; Hernández-Álvarez, A.J.; Jiménez-Martínez, C.; Jacinto-Hernández, C.; Alaiz, M.; Girón-Calle, J.; Vioque, J.; Dávila-Ortiz, G. Antioxidant and metal chelating activities of pep-tide fractions from phaseolin and bean protein hydrolysates. Food Chem., 2012, 135, 1789-1795.

Antioxidative Peptides: Trends and Perspectives Current Medicinal Chemistry, 2013, Vol. 20, No. 36 4591
[15] Kong, B.; Peng, X.; Xiong Y.L.; Zhao, X. Protection of lung fibro-blast MRC-5 cells against hydrogen peroxide-induced oxidative damage by 0.1–2.8 kDa antioxidative peptides isolated from whey protein hydrolysate. Food Chem., 2012, 135, 540–547.
[16] Zheng, J.; Wu, H.; Zhu, B.; Dong, X.; Zhang, M.; Li, Y. Identifica-tion of antioxidative oligopeptides derived from autolysis hydro-lysates of sea cucumber (Stichopus japonicus) guts. Eur. Food Res. Technol., 2012, 234, 895-904.
[17] Gu, L.; Zhao, M.; Li, W.; You, L.; Wang, J.; Wang, H.; Ren, J. Chemical and cellular antioxidant activity of two novel peptides designed based on glutathione structure. Food Chemical Toxic.,2012, 50, 4085–4091.
[18] Fitzgerald R.J., Meisel, H.; Milk protein-derived peptide inhibitors of angiotensin-I-converting enzyme. Br. J. Nutr., 2000, 84, S33-S37
[19] Sen, C.K. Nutritional biochemistry of cellular glutathione. Nutr. Biochem., 1997, 8, 660-672.
[20] Ballatori, N.; Krance, S.M.; Notenboom, S.; Shi S.; Tieu, K.; Hammond, C.L. Glutathione dysregulation and the etiology and progression of human diseases. Biological Chem., 2009, 390, 191–214.
[21] Hipkiss, A.R. In: Advances in Food and Nutrition Research; Steve L. T., Ed.; Academic Press 2009; Vol. 57, pp 87-154.
[22] Decker, E.A.; Livisay, S.A.; Zhou S. A re-evaluation of the anti-oxidant activity of purified carnosine. Bioch. (Moscow), 2000, 65,766-770.
[23] Boldyrev, A.A.; Stvolinsky, S.L.; Fedorova, T.N.; Suslina Z.A. Carnosine as a natural antioxidant and geroprotector: from molecu-lar mechanisms to clinical trials. Rejuvenation Res., 2010, 13, 156-158.
[24] Yamada, M.; Shibusawa, N.; Hashida, T.; Satoh, T.; Monden, T.; Prasad, C.; Mori, M. Abundance of cyclo(His-Pro)-like immunore-activity in the brain of TRH-deficient mice. Endocrinology, 1999,140, 538-541.
[25] Minelli, A.; Bellezza, I.; Grottelli, S.; Galli, F. Focus on cyclo(His-Pro): history and perspectives as antioxidant peptide. Amino Acids,2008, 35, 283–289.
[26] Hilton, C.W.; Prasad, C.; Vo, P.; Mouton, C. Food contains the bioactive peptide, cyclo(His-Pro). J. Clin. Endocrinol. Metab.,1992, 75, 375-378.
[27] Pickart, L.; Vasquez-Soltero, J.M.; Margolina, A. The human tripeptide GHK-Cu in prevention of oxidative stress and degenera-tive conditions of aging: implications for cognitive health. Oxida-tive Medicine and Cellular Longevity, 2012, Article ID 324832, 8 pages.
[28] Beretta, G.; Artali, R.; Regazzoni, L.; Panigati, M.; Facino, R. M. Glycyl-histidyl-lysine (GHK) is a quencher of �,�-4-hydroxy-trans-2-nonenal: a comparison with carnosine. Insights into the mecha-nism of reaction by electrospray ionization mass spectrometry, 1H NMR, and computational techniques. Chemical Res. Toxicol.,2007, 20, 1309–1314.
[29] Beretta, G.; Arlandini, E.; Artali, R.; Anton, J. M.; Facino, R. M. Acrolein sequestering ability of the endogenous tripeptide glycyl-histidyl-lysine (GHK): characterization of conjugation products by ESI-MSn and theoretical calculations. J. Pharm. Biomedical Anal.,2008, 47, 596–602.
[30] Moosmann, B.; Behl C. Secretory peptide hormones are biochemi-cal antioxidants: structure-activity relationship. Mol. Pharmacol-ogy, 2002, 61, 260-268.
[31] Tunçel, N.; Korkmaz, O. T.; Tekin, N.; ener, E.; Akyüz, F.; Inal, M. Antioxidant and anti-apoptotic activity of vasoactive intestinal peptide (VIP) against 6-hydroxy dopamine toxicity in the rat cor-pus striatum. J Mol Neurosci. 2012, 46, 51-57.
[32] Moller, N.P.; Scholz-Ahrens, K.E:, Roos, N.; Schrezenmeir, J. Bioactive peptides and proteins from foods: indication for health effects. Eur. J. Nutr., 2008, 47, 171–182.
[33] Udenigwe, C.C.; Aluko, R.E. Food protein-derived bioactive pep-tides: Production, processing, and potential health benefits. J. Food Sci., 2012, 71, R11-R24.
[34] Korhonen, H. Milk-derived bioactive peptides: From science to applications. J. Functional Foods, 2009, 1, 177–187.
[35] Madureira, A.R.; Tavares, T.; Gomes, A.M.; Pintado, M.E.; Mal-cata, F.X. Invited review: Physiological properties of bioactive pep-
tides obtained from whey proteins. J. Dairy Sci., 2010, 93, 437-455.
[36] Power, O.; Jakeman, P.; FitzGerald, R.J. Antioxidative peptides: enzymatic production, in vitro and in vivo antioxidant activity and potential applications of milk-derived antioxidative peptides. Amino Acids, 2012, in press Doi:10.1007/s00726-012-1393-9.
[37] Phelan, M.; Aherne, A.; Fitzgerald, R.J.; O’Brien, N.M. Casein-derived bioactive peptides: Biological effects, industrial uses, safety aspects and regulatory status. Int. Dairy. J., 2009, 19, 643-654.
[38] Tsopmo, A.; Romanowski, A.; Banda, L.; Lavoie, J.C.; Jenssen, H.; Friel, J.K.; Novel anti-oxidative peptides from enzymatic digestion of human milk. Food Chem., 2011, 126, 1138-1143.
[39] Contreras, M.M.; Hernández-Ledesma, B.; Amigo, L.; Martín-Álvarez; P.J.; Recio, I. Production of antioxidant hydrolyzates from a whey protein concentrate with thermolysin: Optimization by re-sponse surface methodology. LWT - Food Sci. Technol., 2011, 44,9-15.
[40] Alolu, H.S.; Öner, Z. Determination of antioxidant activity of bioactive peptide fractions obtained from yogurt. J. Dairy Sci.,2011, 94, 5305–5314.
[41] Corrêa, A.P.F.; Daroit, D.J.; Coelho, J.; Meira, S.M.M.; Lopes, F. C.; Segalin, J.; Rissoc, P.H.; Brandelli, A.. Antioxidant, antihyper-tensive, and antimicrobial properties of ovine milk caseinate hydro-lyzed with a microbial protease. J. Sci. Food Agric., 2011, 91,2247–2254.
[42] Meira, S.M.M.; Daroit, D.J.; Helfer, V.E.; Corrêa, A.P.F.; Segalin, J.; Carro, S.; Brandelli, A. Bioactive peptides in water-soluble ex-tracts of ovine cheeses from Southern Brazil and Uruguay. Food Res. Int., 2012, 48, 322–329.
[43] Sun, Y.; Pan, D.; Guo, Y.; Li, J. Purification of chicken breast protein hydrolysate and analysis of its antioxidant activity. Food Chemical Toxicol., 2012, 50, 3397–3404.
[44] Rao, S.; Sun, J.; Liu, Y.; Zeng, H.; Su Y.; Yang, Y. ACE inhibitory peptides and antioxidant peptides derived from in vitro digestion hydrolysate of hen egg white lysozyme. Food Chem., 2012, 135,1245–1252.
[45] Lee, D.; Kim, Y.; Hwang, J.; Kim, E.; Moon, S.; Jeon, B.; Jeon, T.; Kim, J.M; Park, P. Purification and characterization of a novel an-tioxidative peptide from duck skin by-products that protects liver against oxidative damage. Food Res. Int., 2012, 49, 285–295.
[46] Tanzadehpanah, H.; Asoodeh, A.; Chamani, J. An antioxidant peptide derived from ostrich (Struthio camelus) egg white protein hydrolysates. Food Res. Int., 2012, 49, 105–111.
[47] Bernardini, R.; Mullen, A.M.; Bolton, D.; Kerry, J.; O'Neill, E.; Hayes, M. Assessment of the angiotensin-I-converting enzyme (ACE-I) inhibitory and antioxidant activities of hydrolysates of bo-vine brisket sarcoplasmic proteins produced by papain and charac-terisation of associated bioactive peptidic fractions. Meat Sci.,2012, 90, 226–235.
[48] Banerjee, P.; Suseela, G.; Shanthi, C. Isolation and Identification of cryptic bioactive regions in bovine achilles tendon collagen. Pro-tein J., 2012, 31, 374–386.
[49] Bernardini, R.; Harnedy, P.; Bolton, D.; Kerry, J.; O’Neill, E.; Mullen, A.M.; Hayes, M. Antioxidant and antimicrobial peptidic hydrolysates from muscle protein sources and by-products. Food Chem., 2011, 124, 1296–1307.
[50] Udenigwe, C.C; Howard, A. Meat proteome as source of functional biopeptides. Food Res. Int., 2012, in press, Doi:10.1016/j.foodres. 2012.10.002.
[51] Broncano, J.M.; Otte, J.; Petrón, M.J.; Parra, V.; Timón, M.L. Isolation and identification of low molecular weight antioxidant compounds from fermented “chorizo” sausages. Meat Sci., 2012,90, 494–501.
[52] Escudero, E.; Aristoy, M.; Nishimura, H.; Arihara, K.; Toldrá, F. Antihypertensive effect and antioxidant activity of peptide fractions extracted from Spanish dry-cured ham. Meat Sci., 2012, 91, 306–311.
[53] Liu, Q.; Kong, B.; Li, G.; Liu, N.; Xia, X. Hepatoprotective and antioxidant effects of porcine plasma protein hydrolysates on car-bon tetrachloride-induced liver damage in rats. Food Chemical Toxic., 2011, 49, 1316–1321.

4592 Current Medicinal Chemistry, 2013, Vol. 20, No. 36 Freitas et al.
[54] Bahareh, H.; Sarmadi, A.I. Antioxidative peptides from food pro-teins: A review. Peptides, 2010, 31, 1949–1956.
[55] Anusha, G.P.; Samaranayaka, E.; Li-Chan, C.Y. Food-derived peptidic antioxidants: A review of their production, assessment, and potential applications. J. Functional Foods, 2011, 3, 229 –254.
[56] Mohamed, T.K.; Issoufou, A.; Zhou, H. Antioxidant activity of fractionated foxtail millet protein hydrolysate. Int. Food Res. J.,2012, 19, 207-213.
[57] Delgado, O.M.C.; Tironi, V. A.; Añón, M. C. Antioxidant activ-ity of amaranth protein or their hydrolysates under simulated gas-trointestinal digestion. LWT - Food Sci. Technol., 2011, 44,1752-1760.
[58] Kou, X.; Gao, J.; Xue, Z.; Zhang, Z.; Wang, H.; Wang, X. Purifica-tion and identification of antioxidant peptides from chickpea (Cicer arietinum L.) albumin hydrolysates. LWT - Food Sci. Technol., 2012, 50, 591-598.
[59] Zhang, T.; Li, Y.; Miao, M.; Bo Jiang, B. Purification and char-acterisation of a new antioxidant peptide from chickpea (Cicer arietium L.) protein hydrolysates. Food Chem., 2011, 128, 28–33.
[60] Zhang, T.; Jiang, B., Miao, M.; Mu, W., Li, Y. Combined effects of high-pressure and enzymatic treatments on the hydrolysis of chick-pea protein isolates and antioxidant activity of the hydrolysates. Food Chem., 2012, 135, 904–912.
[61] Chen, N.; Yang, H.; Sun, Y.; Niu, J.; Liua, S. Purification and identification of antioxidant peptides from walnut (Juglans regiaL.) protein hydrolysates. Peptides, 2012, 38, 344-349.
[62] Zhou, K.; Sun, S.; Canning, C. Production and functional charac-terisation of antioxidative hydrolysates from corn protein via en-zymatic hydrolysis and ultrafiltration. Food Chem., 2012, 135,1192–1197.
[63] Zheng, L.; Su, G.; Ren, J.; Gu, L.; You, L.; Zhao, M. Isolation and characterization of an oxygen radical absorbance activity peptide from defatted peanut meal hydrolysate and its antioxidant proper-ties. J. Agric. Food Chem., 2012, 60, 5431�5437.
[64] Dong, X.; Zhao, M.; Shi, J.; Yang, B.; Li, J.; Luo, D.; Jiang, G.; Jiang, Y. Effects of combined high-pressure homogenization and enzymatic treatment on extraction yield, hydrolysis and function properties of peanut proteins. Innovative Food Sci. Emerging Technol., 2011, 12, 478–483.
[65] Zhang, Y.; Zhang, H.; Wang, L.; Guo, X.; Qi, X.; Qian, H. Influ-ence of the degree of hydrolysis (DH) on antioxidant properties and radical-scavenging activities of peanut peptides prepared from fermented peanut meal. Eur. Food Res. Technol., 2011, 232,941–950.
[66] Valdez-Ortiz, A.; Fuentes-Gutiérrez, C.I.; Germán-Báez, L.J.; Gutiérrez-Dorado, R.; Medina-Godoy, S. Protein hydrolysates ob-tained from Azufrado (sulphur yellow) beans (Phaseolus vulgaris):Nutritional, ACE-inhibitory and antioxidative characterization. LWT - Food Sci. Technol., 2012, 46, 91-96.
[67] Ng, K.L.; Ayob, M.K.; Said, M.; Osman, M.A.; Ismail, A. Optimi-zation of enzymatic hydrolysis of palm kernel cake protein (PKCP) for producing hydrolysates with antiradical capacity. Ind. Crops and Products, 2013, 43, 725– 731.
[68] He, R.; Ju, X.; Yuan, J.; Wang, L.; Girgih, A.T.; Aluko, R-E. Anti-oxidant activities of rapeseed peptides produced by solid state fer-mentation. Food Res. Int., 2012, 49, 432–438.
[69] Pan, M.; Jiang, S.; Pan, J.L. Antioxidant activities of rapeseed protein hydrolysates. Food Bioprocess Technol., 2011, 4, 1144-1152.
[70] Memarpoor-Yazdi, M.; Mahaki, H.; Zare-Zardini, H. Antioxidant activity of protein hydrolysates and purified peptides from Zizy-phus jujuba fruits. J. Functional Foods, 2013, 5, 62-70.
[71] Hu, L.; Song, R.; Gu, Z. An antioxidant peptide produced by autolysis reactions from wheat germ. African J. Biotechnol., 2012,11, 3640-3648.
[72] Xia, Y.; Bamdad, F.; Gänzle, M.; Chen, L. Fractionation and char-acterization of antioxidant peptides derived from barley glutelin by enzymatic hydrolysis. Food Chem., 2012, 134, 1509–1518.
[73] Coda R.; Rizzello, C.G.; Pinto, D.; Gobbetti, M. Selected lactic acid bacteria synthesize antioxidant peptides during sourdough fermentation of cereal flours. Appl. Env. Microbiol., 2012, 78,1087–1096.
[74] Rizzello, C.G.; Nionelli, L.; Coda, R., Gobbetti, M. Synthesis of the cancer preventive peptide Lunasin by lactic acid bacteria dur-ing sourdough fermentation. Nutrition and Cancer, 2012, 64,111–120.
[75] Hernández-Ledesma, B.; Hsieh, C.C.; de Lumen, B.O. Antioxidant and anti-inflammatory properties of cancer preventive peptide lunasin in RAW 264.7 macrophages. Bioch. Biophysical Res. Comm., 2009, 390, 803–808.
[76] Hernández-Ledesma, B.; Hsieh, C.C.; de Lumen, B.O. Lunasin, a novel seed peptide for cancer prevention. Peptides, 2009, 30, 426–430.
[77] Lordan, S.; Ross, R.P.; Stanton, C. Marine bioactives and func-tional food ingredients: Potential to reduce incidence of chronic diseases. Mar. Drugs, 2011, 9, 1056-1100.
[78] Elias, R.J.; Kellerby, S.S.; Decker, E.A. Antioxidant activity of proteins and peptides. Crit. Rev. Food Sci. Nutr., 2008, 48, 430-441.
[79] Samarakoon, K.; Jeon, Y. Bio-functionalities of proteins derived from marine algae – A review. Food Res. Int., 2012, 48, 948-960.
[80] Harnedy, P.A.; Fitzgerald, R.J. Bioactive proteins, peptides, and amino acids from macroalgae. J. Phycol., 2011, 47, 218-232.
[81] Ngo, D.; Vo, T.; Ngo, D.; Wijesekara, I.; Kim, S. Biological activi-ties and potential health benefits of bioactive peptides derived from marine organisms. Int. J. Biological Macromolecules, 2012, 51,378-383.
[82] Najafian, L.; Babji, A.S. A review of fish-derived antioxidant and antimicrobial peptides: Their production, assessment, and applica-tions. Peptides, 2012, 33, 178-185.
[83] Harnedy, P.A.; Fitzgerald, R.J. Bioactive peptides from marine processing waste and shellfish: a review. J. Functional Foods,2012, 4, 6-24.
[84] Ngo, D.; Wijesekara, I.; Vo, T.; Ta, Q.V.; Kim, S. Marine food-derived functional ingredients as potential antioxidants in the food industry: An overview. Food Res. Int., 2011, 44, 523-529.
[85] Guedes, A.C.; Amaro, H.M.; Malcata, F.X. Microalgae as source of high added-value compounds – A brief review of recent work. Bio-technol. Progress, 2011, 27, 597-613.
[86] Shiu, C.T.; Lee, T.M: Ultraviolet-B-induced oxidative stress and responses of the ascorbate-glutathione cycle in a marine macroal-gae Ulva fasciata. J. Exp. Botany, 2005, 56, 2851-2865.
[87] Tierney, M.S.; Croft, A.K., Hayes, M. A review of antihypertensive and antioxidant activities in macroalgae. Botânica Marina, 2010,53, 387-408.
[88] Cian, R.E.; Martínez-Augustin, O.; Drago, S. Bioactive properties of peptides obtained by enzymatic hydrolysis from protein byprod-ucts of Porphyra Columbina. Food Res. Int., 2012, 49, 364-372.
[89] Kang, K.; Qian, Z.; Ryu B.; Kim S. Characterzation of growth and protein contents from microalgae Navicula incerta with the investi-gation of antioxidant activity of enzymatic hydrolysates. Food Sci. Biotechnol., 2011, 20, 183-191
[90] Kang, K.; Qian, Z.; Ryu B.; Karadeniz, F.; Kim, D.; Kim S. Anti-oxidant peptides from protein hydrolysate of microalgae Navicula incerta and their protective effects in Hepg2/CYP2E1 cells induced by ethanol. Phytotherapy Res., 2012, 26, 1555-1563.
[91] Ko, S.; Kim, D.; Jeon, Y. Protective effect of a novel antioxidative peptide purified from a marine Chlorella ellipsoidea protein against free radical-induced oxidative stress. Food Chemical Toxicol.,2012, 50, 2294-2302.
[92] Himaya, S.W.A.; Ngo, D.; Ryu, B.; Kim, S. An active peptide purified grom gastrointestinal enzyme hydrolysate of Pacific cod skin gelatin attenuates angiotensin-1 converting enzyme (ACE) ac-tivity and cellular oxidative stress. Food Chem., 2012, 132, 1872-1882.
[93] Kumar, N.S.S.; Nazeer, R.A.; Jaiganesh, R. Purification and identi-fication of antioxidant peptides from the skin protein hydrolysate of two marine fishes, horse mackerel (Magalaspis cordyla) and croaker (Otolithes ruber). Amino acids, 2012, 42, 1641-1649.
[94] Shabena, Y.N.; Nazeer, R.A. Identification of active peptides from backbones of Nemipterus japonicas and Exocoetus volitans by electrospray ionization-mass spectrometry. Int. J. Food Sci. Tech-nol., 2011, 46, 1993-1996.
[95] Naquash, S.Y.; Nazeer, R.A. Optimization of enzymatic hydrolysis conditions for the production of antioxidant peptides from muscles

Antioxidative Peptides: Trends and Perspectives Current Medicinal Chemistry, 2013, Vol. 20, No. 36 4593
of Nemipterus japonicas and Exocoetus volitans using response surface methodology. Amino Acids, 2012, 43, 337-345.
[96] Kumar, N.S.S.; Nazeer, R.A.; Jaiganesh, R. Purification and bio-chemical characterization of antioxidant peptide from horse mack-erel (Magalaspis cordyla) viscera protein. Peptides, 2011, 32,1496-1501.
[97] Fan, J.; He, J.; Zhuang, Y.; Sun, L. Purification and identification of antioxidant peptides from enzymatic hydrolysates of Tilapia (Oreochromis niloticus) frame protein. Molecules, 2012, 17,12836-12850.
[98] Fitzgerald, C.; Mora-Soler, L.; Gallagher, E.; O’Connor, P.; Prieto, J.; Soler-Vila, A.; Hayes, M. Isolation and characterization of bio-active pro-peptides with in vito rennin inhibitory activities from macroalga Palmaria palmate. J. Agric. Food Chem., 2012, 60,7421-7427.
[99] Mendis, E.; Rajapakse, N.; Byun, H.; Kim, S. 2005. Investigation of jumbto squid (Dosidicys gigas) skin gelatin peptides for their in vitro antioxidant effects. Life Sciences, 2005, 77, 2166-2178.
[100] Zhao, K.; Zhao, G.M.; Wu, D.; Soong, Y.; Birk, A.V.; Schiller, P.W.; Szeto, H.H. Cell-permeable peptide antioxidants targeted to inner mitochondrial membrane inhibit mitochondrial swelling, oxi-dative cell death, and reperfusion injury. J. Biol. Chem., 2004, 279,34682-34690.
[101] Zhao, K.; Luo, G.; Zhao, G.M.; Schiller, P.W.; Szeto, H.H. Tran-scellular transport of a highly polar 3+ net charge opioid tetrapep-tide. J. Pharmacol. Exp. Ther., 2003, 304, 425–432.
[102] Szeto, H.H. Mitochondria-targeted cytoprotective peptides for ischemia-reperfusion injury. Antioxid. Redox. Signal., 2008, 10,601–619.
[103] Szeto, H.H. Cell-permeable, mitochondrial-targeted, peptide anti-oxidants. The AAPS J., 2006, 8, E277-E283.
[104] Khavinson, V.; Ribakova, Y.; Kulebiakin, K.; Vladychenskaya, E.; Kozina, L.; Arutjunyan, A.; Boldyrev, A. Pinealon increases cell viability by suppression of free radical levels and activating proliferative processes. Rejuvenation Research, 2011, 14, 535-541.
[105] Ren, J.; Zhao, M.; Shi, J.; Wang, J.; Jiang, Y.; Cui, C.; Kakuda, Y.; Xue, S.J. Purification and identification of antioxidant peptides from grass carp hydrolysates by consecutive chromatography and electrospray ionization-mass spectrometry. Food Chem., 2008, 108,727–736.
[106] Tang, X.; He, Z.; Dait, Y.; Xiong, Y. L.; Xie, M.; Chen, J. Peptide fractionation and free radical scavenging activity of zein hydro-lysate. J. Agr. Food Chem., 2010, 58, 587–593.
[107] You, L.; Zhao, M.; Regenstein, J.M.; Ren, J. Purification and iden-tification of antioxidative peptides from loach (Misgurnus anguilli-caudatus) protein hydrolysate by consecutive chromatography and electrospray ionization-mass spectrometry. Food Res. Int., 2010,43, 1167–1173.
[108] Hsu, K.C. Purification of antioxidative peptides prepared from enzymatic hydrolysates of tuna dark muscle by-product. Food Chem., 2010, 122, 42-48.
[109] Zhang, J.; Zhang, H.; Wang, L.; Guo, X.; Wang, X.; Yao, H. Isola-tion and identification of antioxidative peptides from rice en-dosperm protein enzymatic hydrolysate by consecutive chromatog-raphy and MALDI-TOF/TOF MS/MS. Food Chem., 2010, 119,226-234.
[110] Zhou, X.; Wang, C.; Jiang, A. Antioxidant peptides isolated from sea cucumber Stichopus japonicas. Eur. Food Res. Technol., 2012,234, 441-447.
[111] Zhang, S.B.; Wang, Z.; Xu, S.Y.;Gao, X.F. Purification and charac-terization of a radical scavenging peptide from rapseed protein hy-drolysates. J. Am. Oil Chem. Soc., 2009, 86, 959-966.
[112] Wijesinghe, W.A.J.P.; Jeon, Y.-J.; Enzyme-assisted extraction (EAE) of bioactive components: a useful approach for recovery of industrially important metabolites from seaweeds: A review. Fi-toterapia, 2012, 83, 6-12.
[113] Farvin, K.H.S.; Baron, C.P.; Nielsen, N.S.; Jacobsen, C. Antioxi-dant activity of yoghurt peptides: Part 1-in vitro assays and evalua-tion in �-3 enriched milk. Food Chem, 2010, 123,1081-1089.
[114] Farvin, K.H.S.; Baron, C.P.; Nielsen, N.S.; Otte, J., Jacobsen, C. Antioxidant activity of yoghurt peptides: Part 2 – Characterisation of peptide fractions. Food Chem, 2010, 123,1090-1097.
[115] Zambrowicz, A.; Timmer, M.; Polanowski, A.; Lubec, G.; Trziska T. Manufacturing of peptides exhibiting biological activity. Amino acids, 2013, 44, 315-320.
[116] Zompra, A.A.; Galanis, A.S.; Werbitzky, O.; Albericio, F. Manufacturing peptides as active pharmaceutical ingredients. Fu-ture Medical Chem., 2009, 1, 361-377.
[117] Vlieghe, P.; Lisowski, V.; Martinez, J.; Khrestchatisky, M. Syn-thetic therapeutic peptides: science and market. Drug Discovery Today, 2010, 15, 40 - 56.
[118] Xue, D.; Zhao, M.; Wang, Y-J.; Wang , L.; Yang, Y.; Wang, S-W.; Zhang, R.; Zhao Y.; Liu, R-T. A multifunctional peptide rescues memory deficits in Alzheimer's disease transgenic mice by inhibit-ing A�42-induced cytotoxicity and increasing microglial phagocy-tosis. Neurobiology of Disease, 2012, 46, 701–709
[119] Paduano, F.; Ortuso, F.; Campiglia, P.; Raso, C.; Iaccino, E.; Gas-pari, M.; Gaudio, E.; Mangone, G.; Carotenuto, A.; Bilotta, A.; Narciso, D.; Palmieri, C.; Agosti, V.; Artese, A.; Gomez-Monterrey, I.; Sala, M.; Cuda, G.; Iuliano, R.; Perrotti, N.; Scala, G.; Viglietto, G.; Alcaro, S.; Croce, C.M.; Novellino, E.; Fusco, A.; Trapasso, F. Isolation and Functional Characterization of Peptide Agonists of PTPRJ, a Tyrosine Phosphatase Receptor Endowed with Tumor Suppressor Activity. ACS Chemical Biol., 2012, 7,1666�1676.
[120] Taddei, K.; Laws, S.M.; Verdile, G.; Munns, S.; D’Costa, K.; Har-vey, A.R.; Martins, I.J.; Hill, F.; Levy, E.; Shaw, J.E.; Martins R.N. Novel phage peptides attenuate beta amyloid-42 catalysed hydro-gen peroxide production and associated neurotoxicity. Neurobiol-ogy of Aging, 2010, 31, 203-214.
[121] Wu, H.; Shiau, C.; Chen, H.; Chiou, T. Antioxidant activities of carnosine, anserine, some free amino acids and their combination. J. Food Drug Anal., 2003, 11, 148-153.
[122] Chen, H.M.; Muramoto, K.; Yamauchi, F.; Nokihara, K. Antioxi-dant activity of designed peptides based on the antioxidant peptide isolated from digests of a soybean protein. J. Agric. Food Chem.,1996, 44, 2619-2623.
[123] Kitts, D.D.; Weiler, K. Bioactive proteins and peptides from food sources. Applications of bioprocesses used in isolation and recov-ery. Curr. Pharm. Des., 2003, 9, 1309-1323.
[124] Ranathunga, S.; Rajapakse, N.; Kim, S. Purification and characterization of antioxidative peptide derived from muscle of conger eel (Conger myriaster). Eur. Food Res. Technol., 2006, 222,310-315.
[125] Meisel, H. Biochemical properties of peptides encrypted in bovine milk protein. Curr. Med. Chem., 2005, 12, 1905-1919.
[126] Sun, J.Z.; Kaur, H.; Halliwell, B.; Li, X.Y.; Bolli, R. Use of aro-matic hydroxylation of phenylalanine to measure production of hy-droxyl radicals after myocardial ischemia in vivo. Direct evidence for a pathogenic role of the hydroxyl radical in myocardial stun-ning. Circ. Res., 1993, 73, 534-549.
[127] Saito, K.; Jin, D.H.; Ogawa, T.; Muramoto, K.; Hatakeyama, E.; Yasuhara, T.; okihara, K. Antioxidative properties of tripeptide prepared by the combinatorial chemistry. J. Agric. Food Chem.,2003, 51, 3668-3674.
[128] Prior, R.L.; Wu, X.; Achaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agric. Food Chem., 2005, 53, 4290-4302.
[129] Decker, E.A.; Warner, K.; Richards, M.P.; Shahidi, F. Measuring antioxidant effectiveness in Food. J. Agric. Food Chem., 2005, 53,4303-4310.
[130] Wood, L.G. Gibson, P.G.; Garg, M.L. A review of the methodol-ogy for assessing in vivo antioxidant capacity. J. Sci. Food Agric.,2006, 86, 2057-2066.
[131] Suarez-Jimenez, G.; Burgos-Hernandez, A.; Esquerra-Brauer, J. Bioactive peptides and depsipeptides with anticancer potential: Sources from marine animals. Mar. Drugs, 2012, 10, 963-986.
[132] Sheih, I.; Fang, T.J.; Wu, T.; Lin P. Anticancer and antioxidant activities of the peptide fraction from algae protein waste. J. Agric. Food Chem., 2010, 58, 1202-1207
[133] Yang, R.; Zhang, Z.; Pei, X.; Han, X.; Wang, J.; Wang, L.; Long Z.; Shen, X.; Li; Y. Immunomodulatory effects of marine oligopep-tide preparation from Chum Salmon (Oncorhynchus keta) in mice. Food Chem., 2009; 113, 464-470.

4594 Current Medicinal Chemistry, 2013, Vol. 20, No. 36 Freitas et al.
[134] Díaz, M.; Decker, E.A. Antioxidant mechanisms of caseinophos-phopeptides and casein hydrolysates and their application in ground beef. J. Agric. Food Chem., 2004, 2, 8208-8213.
[135] Hagen, H.; Sandnes, F. Process for improvement of meat quality in fish, protein hydrolysate and method of producing a protein hydro-lysate. International Patent WO 2004/071202, 2004.
[136] Agyei, D.; Danquah, M.K. Industrial-scale manufacturing of phar-maceutical-grade bioactive peptides. Biotechnol. Adv., 2011, 29,272-277.
[137] Schaafsma, G. Safety of protein hydrlysates, fractions thereof and bioactive peptides in human nutrition. Eur. J. Clinical Nutr., 2009,63, 1161-1168.
[138] Hartmann, R.; Meisel, H. Food-derived peptides with biological activity: from research to food applications. Curr. Opinion Bio-technol., 2007, 18, 163-169.
[139] Kim, S., Ravichandran, Y.D.; Han S.B.; Kim, Y.T. Prospective of the cosmeceuticals derived from marine organisms. Biotecnhol. Bioproces Eng., 2008, 13, 511-523.
[140] Gorouhi, F.; Maibach, H.I. Role of topical peptides n preventing or treating age skin. Int. J. Cosmetic Sci., 2009, 31, 327-345.
[141] Zhang, L.; Falla, T.J. Cosmeceuticals and peptides. Clinics in Der-matology, 2009, 27,485-494.
[142] Zhaorigetu, S.; Yanaka, S.; Sasaki, N.; Watanabe, H.; Kato, N. Inhibitory effects of silk protein, sericin on UVB-induced acute damage and tumor promotion by reducing oxidative stress in the skin of hairless mouse. J. Photochem. Photobiol., 2003, 71, 11-17.
Received: November 08, 2012 Revised: January 2�, 2013 Accepted: February 03, 2013





























