Embed Size (px)
Citation preview

Functional Ecology
2003
17
, 246–259
© 2003 British Ecological Society
246
Blackwell Publishing Ltd.
Variation in coral photosynthesis, respiration and growth characteristics in contrasting light microhabitats: an analogue to plants in forest gaps and understoreys?
K. R. N. ANTHONY*† and O. HOEGH-GULDBERG‡
*
Centre for Coral Reef Biodiversity, School of Marine Biology and Aquaculture, James Cook University, Townsville, Queensland 4811, and
‡
Centre for Marine Studies, University of Queensland, Brisbane, Queensland 4072, Australia
Summary
1.
The often complex architecture of coral reefs forms a diversity of light microhabitats.Analogous to patterns in forest plants, light variation may drive strategies for efficientlight utilization and metabolism in corals.
2.
We investigated the spatial distribution of light regimes in a spur-and-groove reefenvironment and examine the photophysiology of the coral
Montipora monasteriata
(Forskål 1775), a species with a wide habitat distribution. Specifically, we examined thevariation in tissue and skeletal thickness, and photosynthetic and metabolic responsesamong contrasting light microhabitats.
3.
Daily irradiances reaching corals in caves and under overhangs were 1–5 and 30–40% of those in open habitats at similar depth (3–5 m), respectively. Daily rates of netphotosynthesis of corals in cave habitats approximated zero, suggesting more than twoorders of magnitude variation in scope for growth across habitats.
4.
Three mechanisms of photoadaptation or acclimation were observed in cave andoverhang habitats: (1) a 20–50% thinner tissue layer and 40–60% thinner skeletalplates, maximizing light interception per unit mass; (2) a two- to threefold higher photo-synthetic efficiency per unit biomass; and (3) low rates of dark respiration.
5.
Specimens from open and cave habitats displayed a high capacity to acclimate todownshifts or upshifts in irradiance, respectively. However, specimens in caves displayedlimited acclimation to further irradiance reduction, indicating that these live near theirirradiance limit.
6.
Analogous to patterns for some plant species in forest gaps, the morphological plas-ticity and physiological flexibility of
M. monasteriata
enable it to occupy light habitatsthat vary by more than two orders of magnitude.
Key-words
: Energy balance, photoacclimation, reef coral, spur-and-groove
Functional Ecology
(2003)
17
, 246–259
Introduction
Light is the key source of energy for reef-building,zooxanthellate corals (Muscatine 1990). The highgross productivity of coral reefs (see review by Hatcher1990) is, in part, attributable to the high irradiancesin shallow oceanic waters. Numerous studies havefocused on the role of light in the photophysiology andenergetics of reef corals (Falkowski
et al
. 1984; Spencer-Davies 1991; Hoegh-Guldberg & Jones 1999; Anthony& Fabricius 2000; earlier studies reviewed by Falkowski,
Jokiel & Kinzie 1990; Muscatine 1990). Common tomost studies, however, is the notion that light and asso-ciated responses of coral photosynthesis and growth varymainly with depth (Chalker, Dunlap & Oliver 1983;Oliver, Chalker & Dunlap 1983; Barnes & Chalker1990) or turbidity (Anthony & Fabricius 2000; Anthony,Connolly & Willis 2002).
The often complex architecture of coral reefs pro-duces a diversity of light microhabitats within dis-tances of only one to a few metres, for examplebeneath tabular corals (Sheppard 1981), in cavities(Dinesen 1983), and at sites near steep slopes andunder overhangs (Brakel 1979). Based on similaritiesin spatial complexity, the light variation in reef environ-ments may be analogous to that of forest habitats. Plants
†Author to whom correspondence should be addressed. E-mail: [email protected]

247
Habitat light variation and coral photophysiology
© 2003 British Ecological Society,
Functional Ecology
,
17
, 246–259
in canopy gaps and in the understorey can often becharacterized as either sun or shade plants, displayingphysiological as well as morphological adaptations(or acclimations) to their light environment (reviewedby Chazdon
et al
. 1996; Lambers, Chapin & Pons1997). Whereas photophysiological and morphologicalresponses to microsite light variation are well estab-lished for plants, little is known about the temporaland spatial profile of irradiances in structurallycomplex reef habitats, and what the implications arefor the physiology and energetics of coral symbioses.
Species of plants, corals and macro-algae that occurin a wide range of light habitats are faced with con-trasting photophysiological challenges. For example,the growth of plants in forest understoreys (reviewby Chazdon
et al
. 1996) and of phototrophic coralsin shaded (review by Muscatine 1990) or turbid(Anthony & Fabricius 2000) reef habitats may be limitedby light availability. Conversely, in open forest gaps(Kitao
et al
. 2000) or on reef crests (Hoegh-Guldberg& Jones 1999; Jones & Hoegh-Guldberg 2001), highirradiances may cause photoinhibition or photodamage.Given that light limitation and light stress are bothlikely to influence key fitness components such as growthand reproduction, plasticity in light responses islikely to have a selective advantage for coral specieswith wide light-habitat distributions. For instance,species that are able to occupy extreme low or highlight regimes may have reduced competition in thosehabitats. Also, a greater physiological tolerance oflight-habitat dynamics (e.g. after physical disturbances)will increase the physiological potential for coloniza-tion of newly formed light gaps.
We investigated the microhabitat light variation in areef groove environment and the associated variationin growth characteristics (tissue and skeletal thick-ness), photophysiology and respiration of hermatypiccorals, using the common species
Montipora monaste-riata
as a case study. The foliaceous (plate-like) growthform of this species allows comparison of the roles oftissue thickness and photopigment concentration inlight capture, and of the photosynthetic capacity ofcorals and plants. First, we characterized the lightregimes within a groove habitat to allow comparisonwith light regimes of contrasting light environments inother habitats, for example forest gaps and under-storeys. Second, we investigated the implicationsof such contrasting habitats for the energetics of
M. monasteriata
, and to what degree corals can photo-acclimate to changes in growth irradiance. Specifically,we tested the hypothesis that corals in groove habitatsdisplay morphological and physiological adjustmentsto spatial variation in light regimes by varying thethickness and composition of the tissue layer andparameters of the net photosynthesis–irradiance(
P–I
) curve. Such a pattern would be consistent withthat found in forest gaps and understoreys, whereplants generally exhibit light acclimation via twomechanisms: changes in leaf anatomy (e.g. thickness);
and physiological and biochemical changes in cells andchloroplasts (e.g. Chazdon & Kaufmann 1993).
Materials and methods
The field work was conducted in reef grooves along thenorthern front of Wistari Reef (Fig. 1a) near HeronIsland in the southern section of the Great BarrierReef (23
°
27
′
S, 151
°
54
′
E). Reef habitats along north-facing reef slopes were chosen because they are likelyto display the highest variation among light micro-habitats during the day. Specifically, the orientationof groove walls and overhangs is perpendicular tothe path of the sun, shading west-facing groove wallsin the morning and east-facing walls in the afternoon.The depth within the grooves was 2–4 m below thelowest astronomical tide (3 m tidal range) and thewidth of the groves ranged from 3 to 6 m. Based on aninitial survey of light regimes and coral distributionswithin five groove systems, the one that representedmost microhabitat characteristics of all grooves wasselected for further study (Fig. 1).
To characterize the variation in light regimes withinthe groove during the day, a set of five light loggers(392, Dataflow Systems, Cooroy, Australia) withcosine-corrected PAR sensors (400–700 nm wave-length) were deployed along a transect running east–west across the groove floor (Fig. 1b). All sensors hadbeen calibrated under water against a manufacturer-calibrated quantum sensor (Li 192s, LiCor, Lincoln,NE, USA). All sensors were placed next to specimensof
M. monasteriata
(except in the western cave whichwas without scleractinian corals) with the sensororientation similar to that of the colony surface. Theorientation of colonies was generally horizontal in openhabitats and vertical in cave habitats (Fig. 1c), indic-ating the predominant direction of incident light inthose microhabitats. Thus two loggers were placedhorizontally (sensor pointing outwards) in a west- oreast-facing cave (loggers A and E, Fig. 1b). Twologgers were placed under the edge of overhangs inthe eastern and western side of the groove ( loggers Band D), and one logger was placed in the centre of thegroove (logger C). For comparison, a logger was alsodeployed in open habitat 10 m away from the slope at5 m depth (logger F, not shown). The loggers weredeployed for 9 days (11–19 January 2001) during whichthey stored integrated records of irradiance every 5 min.
One of the most prevalent corals in the spur-and-groovesystems of Wistari Reef is
Montipora monasteriata
(Forskål 1775), a species that forms plating colonies
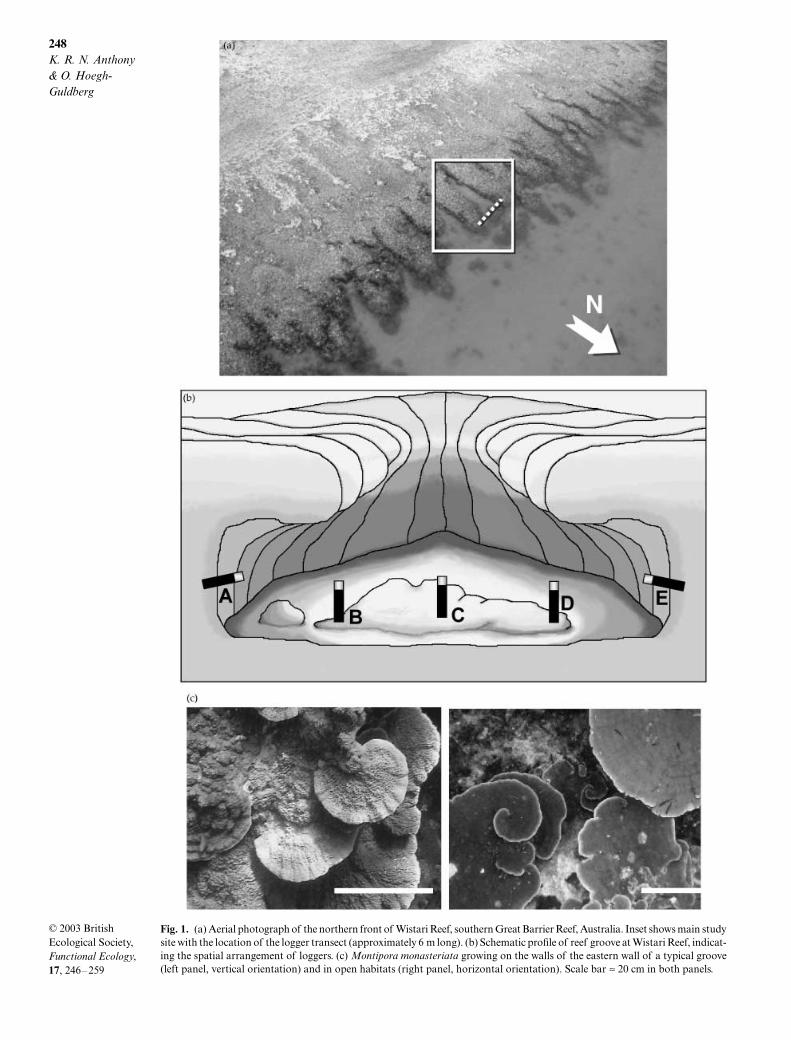
248
K. R. N. Anthony & O. Hoegh-Guldberg
© 2003 British Ecological Society,
Functional Ecology
,
17
, 246–259
Fig. 1. (a) Aerial photograph of the northern front of Wistari Reef, southern Great Barrier Reef, Australia. Inset shows main studysite with the location of the logger transect (approximately 6 m long). (b) Schematic profile of reef groove at Wistari Reef, indicat-ing the spatial arrangement of loggers. (c) Montipora monasteriata growing on the walls of the eastern wall of a typical groove(left panel, vertical orientation) and in open habitats (right panel, horizontal orientation). Scale bar ≈ 20 cm in both panels.

249
Habitat light variation and coral photophysiology
© 2003 British Ecological Society,
Functional Ecology
,
17
, 246–259
up to 50 cm in diameter.
Montipora monasteriata
iscommon throughout the Indo-Pacific (Veron 1986)and is found in most reef microhabitats. To determinethe photophysiological responses, tissue componentsand growth characteristics of
M. monasteriata
fromdifferent light microhabitats, we collected colonyfragments (
≈
8
×
8 cm, using bone-cutters) fromhabitats within the groove. Three habitats could bedefined: (a) open habitats, which were free from anyshading by the walls of the groove; (b) overhangs,which were sites directly underneath the edge of theshelf-like overhang of the groove walls; and (c) caves,which were sites under the overhangs and were per-manently shaded.
To determine
P–I
response curves of
M. monasteriata
from the groove habitats, oxygen-respirometry runswere conducted in the laboratory at Heron IslandResearch Station under two metal halide lamps (each400 watts, EYE, Japan) using a system with six closedchambers. Individual chambers (35 cm long, 11 cmwide, 8 cm high) were designed as flumes and produceda unidirectional flow of 5–6 cm s
−
1
. The system wassubmerged in a jacket of running seawater to buffertemperature fluctuations and to allow periodic flushingof chambers (4 min flushing between every 20 minrecording interval). The chambers were fitted withClark-type oxygen electrodes connected to a signalconditioner (Cheshire Systems, Adelaide, Australia)and a logger (DataTaker 50, Chesterland, Ohio, USA).To control for photosynthesis and respiration of biofilmand micro-organisms, two chambers in each run wereleft empty.
Irradiance was adjusted by elevating or lowering thelamps, exposing the corals to 10 irradiances for 24 mineach (ranging from 0 to 1300
µ
mol photons m
−
2
s
−
1
).Each respirometry run started with a dark incubation at7:00–8:00 am using fully dark-adapted corals, throughstepwise increasing irradiance, and concluded with adark incubation at 02:00–03:00 pm following exposureto maximum irradiance. The second dark incubationwas used to examine the effect of high irradiance onanimal host respiration (
R
′
dark
). We assumed that theincrease in coral metabolism due to its immediatelight history would persist for at least 20 min (theduration of each recording window) after the lightwas turned off.
To normalize
P–I
curve parameters to colony sur-face area, all colonies (and a ruler for scale) werephotographed using a digital camera (Sony DSC-P1).Surface areas were determined from the digital imagesusing the software package Mocha (Jandel Scientific,San Rafael, CA, USA). Each coral was then wrappedin foil and frozen at
−
70
°
C for later morphometricaland biochemical analysis.
P–I
curve parameters andassociated standard errors were estimated by fittingthe model (Jassby & Platt 1976)
(eqn 1)
to the net photosynthesis-irradiance dataset using iter-ative non-linear estimation (Statistica 1999).
P
max
is themaximum rate of photosynthesis (gross);
α
is the ini-tial slope of the
P–I
curve (a proxy for quantum yield);and
R
dark
is the rate of respiration at night. In addition,subsaturation irradiance (
I
k
) was calculated as
I
k
=
P
max
/
α
. Standard errors for parameters in each indi-vidual curve fit were <10% of the parameter estimates,and were not included in the calculation of standarderrors for each coral group.
Daily rates of net photosynthesis (
P
nD
, a proxy forcarbon balance) were estimated by integrating hourlyrates of net photosynthesis (equation 2) over the day,using the 9-day light data for cave west and centralgroove. Because
M. monasteriata
rarely extends itspolyps, we assume that it relies on a minimal amountof heterotrophy and thus that its carbon acquisition ismainly governed by phototrophy. However, excretioncan represent a substantial carbon loss (Crossland1987), and
P
nD
may thus underestimate the carbon bal-ance.
P
nD
was calculated as
, (eqn 2)
where
t
h
and
t
d
are time in hours and days, respectively.Note that irradiance is here parameterized by time ofday. To compare the relative importance of each
P–I
curve parameter with respect to its regulatory effect onthe daily net photosynthesis (
P
nD
) in each habitat, wesubjected equation 2 to a series of sensitivity analyses.Briefly, sensitivity was determined as the relativechange in
P
nD
caused by a 10% change in a given para-meter, while other parameters were kept constant.
For protein analysis, a frozen subsample (2–3 cm
2
)of each colony was solubilized in 10 ml 1
NaOH at90
°
C for 1 h. Subsamples (0·1 ml) of the slurries wereassayed for protein (DC protein assay kit, Bio-RadLaboratories, Regents Park, NSW, Australia) usingbovine serum albumin (Bio-Rad) as a standard.Absorbances were measured at 750 nm wavelength ina spectrophotometer (Agilent 8453, Agilent Technolo-gies, Palo Alto, CA, USA). The surface areas (polypside only) of the remaining skeletal fragments from theprotein assay (and the assays below) were determinedby foil wrapping (Marsh 1970).
Chlorophyll was extracted from coral samples(
≈
4 cm
2
) in darkness using cold (4
°
C) acetone (100%)during three consecutive extractions, each of 4–6 hduration. Absorbances were determined at 630 and
P P h
IP
Rnmax
dark tan max= +α
P P h
I
PRt
t
t
t
t
nD darkh
h
d
d
h
h
d
d
max( )
max
tan = +
=
=
=
=
∑∑19 1
24
1
9 α

250
K. R. N. Anthony & O. Hoegh-Guldberg
© 2003 British Ecological Society,
Functional Ecology
,
17
, 246–259
663 nm, and chlorophyll
a
concentrations were calcu-lated using the formula of Jeffrey & Humphrey (1975).
To determine concentrations of zooxanthellae perunit surface area, the tissue of coral samples was sep-arated from the skeleton using an airbrush connectedto a reservoir of filtered (0·45
µ
m) seawater. A 5% buff-ered formalin solution (4–6 ml) was added to theresulting homogenate (20–30 ml). The concentrationof zooxanthellae was measured using a haemocyto-meter and 10 replicate cell counts per sample. The countswere adjusted to sample volume and normalized tocm
2
colony surface area.To examine morphological mechanisms of photo-
acclimation, the thickness of the skeletal plates of coralsamples was determined under a dissecting micro-scope fitted with an eyepiece graticule. Three replicatepoints were measured along the broken edge approxim-ately 2 cm from the leading edge of the coral colony,the latter to standardize thickness comparisons. Skeletaldensity (
ρ
S
) was estimated as the ratio of skeletal dryweight to skeletal volume (Anthony
et al
. 2002) usingfragments from each coral from the assays above.
To investigate photoacclimative responses in
M.monasteriata
from contrasting light habitats, a trans-plantation experiment between shaded (cave and over-hang) habitats and open groove habitats was conducted.Four large fragments (approximately 10
×
10 cm)were cut from each of six clones in both cave andopen habitats (total of 48 fragments). One fragmentfrom each clone was used for initial photophysiologyassays in the laboratory (see below). The remainingfragments were then transplanted to three habitats:a less-shaded habitat; a more shaded habitat; andtheir native habitat as control (Fig. 2). A full reciprocaltransplantation between light extremes was avoidedbecause such transitions are unlikely to occur naturally.For attachment, a 3–4 mm hole was drilled (underwater)in one corner of each coral colony, which was thensecured with cable ties to a PVC rack fixed horizontally
to a concrete block. The orientation of the coral colon-ies was kept similar to that of conspecifics growingnaturally in each habitat (horizontal in open habitats,vertical in caves). To control for effects of manipulation,additional coral fragments were transplanted back tothe reef substrate.
As a measure of state of photoacclimation, coralswere assayed for apparent electron transport rates(apparent ETR) as a function of irradiance. ApparentETR
vs I
curves provide a proxy for
P–I
curves, but donot allow direct comparison with these (Maxwell& Johnson 2000). Corals were assayed for apparentETR using a pulse amplitude modulation (PAM)chlorophyll fluorometer (Diving PAM, Walz, Effeltrich,Germany) in the laboratory. In addition, maximumquantum yield (
F
v
/
F
m
, dark-adapted) was determined toassess whether photosystem II was adversely affectedby the treatments (Hoegh-Guldberg & Jones 1999).Assays were conducted under the same light set-upand protocol as described above for the respirometryruns. To provide a pretransplantation baseline, one frag-ment per clone (total of 12) was assayed in the laboratoryon day 1 (Fig. 2); and to test for treatment effects alltransplanted corals and controls (total of 48) were assayedin the laboratory on day 21.
Confidence intervals for daily, net rates of photosyn-thesis (
P
nD
, equation 2) were estimated by MonteCarlo simulations of
P
nD
using the 9-day irradiance data-set and the means and standard deviations of indi-vidual
P–I
curve parameters. Briefly, at hourly timestamps, a random-normal generator (programmedin M
L
5·11) sampled randomly from within thenormal distribution defined by the mean and stand-ard deviation for each of the
P–I
curve parametersfor corals from each of the groove habitats. The hourlyoutputs were then entered sequentially into equation 2to produce one replicate estimate of
P
nD
, and the pro-cedure was repeated 500 times to estimate standarddeviation.
Fig. 2. Experimental design of the 3-week transplantation experiment for Montipora monasteriata between light environmentsin a groove at Wistari Reef. The light environments of the five different habitats are depicted in Fig. 3. In each habitat, sixfragments from the different clones were attached to racks. In addition, one fragment from each clone was transplanted to thereef substrate in either the shaded or open habitat to control for effects of rack.
251
Habitat light variation and coral photophysiology
© 2003 British Ecological Society,
Functional Ecology
,
17
, 246–259
Results
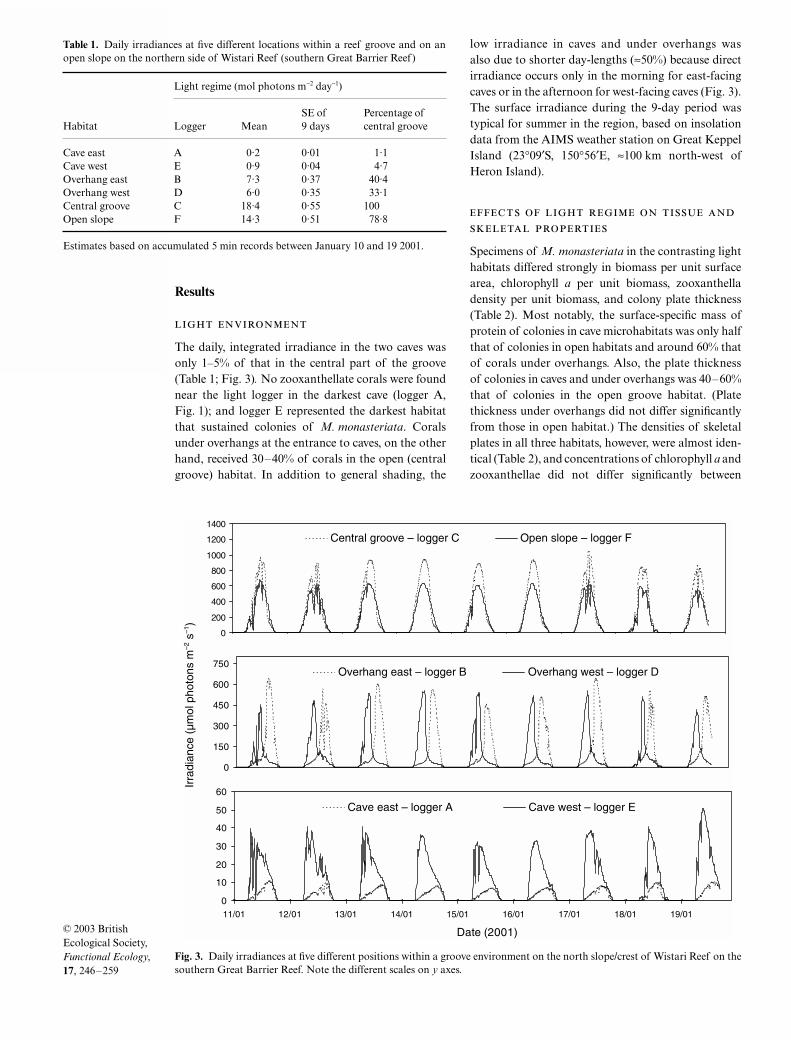
The daily, integrated irradiance in the two caves wasonly 1–5% of that in the central part of the groove(Table 1; Fig. 3)
.
No zooxanthellate corals were foundnear the light logger in the darkest cave (logger A,Fig. 1); and logger E represented the darkest habitatthat sustained colonies of
M. monasteriata
. Coralsunder overhangs at the entrance to caves, on the otherhand, received 30–40% of corals in the open (centralgroove) habitat. In addition to general shading, the
low irradiance in caves and under overhangs wasalso due to shorter day-lengths (
≈
50%) because directirradiance occurs only in the morning for east-facingcaves or in the afternoon for west-facing caves (Fig. 3).The surface irradiance during the 9-day period wastypical for summer in the region, based on insolationdata from the AIMS weather station on Great KeppelIsland (23
°
09
′
S, 150°56′E, ≈100 km north-west ofHeron Island).
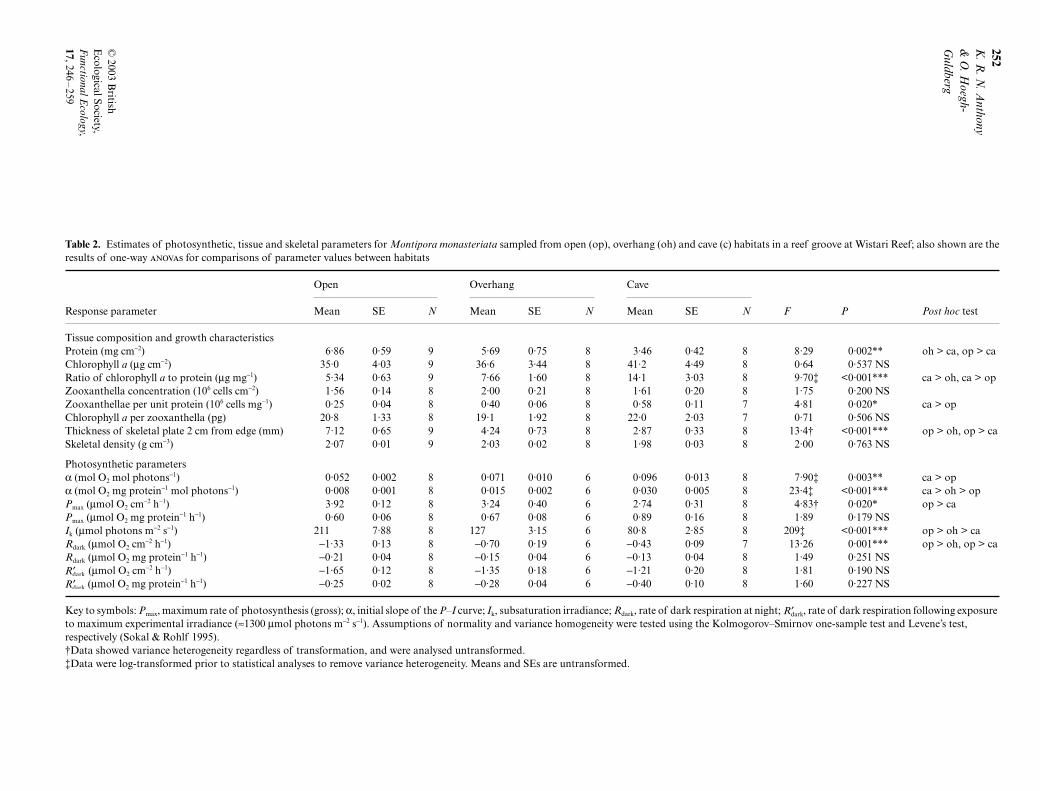
Specimens of M. monasteriata in the contrasting lighthabitats differed strongly in biomass per unit surfacearea, chlorophyll a per unit biomass, zooxanthelladensity per unit biomass, and colony plate thickness(Table 2). Most notably, the surface-specific mass ofprotein of colonies in cave microhabitats was only halfthat of colonies in open habitats and around 60% thatof corals under overhangs. Also, the plate thicknessof colonies in caves and under overhangs was 40–60%that of colonies in the open groove habitat. (Platethickness under overhangs did not differ significantlyfrom those in open habitat.) The densities of skeletalplates in all three habitats, however, were almost iden-tical (Table 2), and concentrations of chlorophyll a andzooxanthellae did not differ significantly between
Fig. 3. Daily irradiances at five different positions within a groove environment on the north slope/crest of Wistari Reef on thesouthern Great Barrier Reef. Note the different scales on y axes.
Table 1. Daily irradiances at five different locations within a reef groove and on anopen slope on the northern side of Wistari Reef (southern Great Barrier Reef)
Habitat
Light regime (mol photons m−2 day−1)
Logger MeanSE of 9 days
Percentage of central groove
Cave east A 0·2 0·01 1·1Cave west E 0·9 0·04 4·7Overhang east B 7·3 0·37 40·4Overhang west D 6·0 0·35 33·1Central groove C 18·4 0·55 100Open slope F 14·3 0·51 78·8
Estimates based on accumulated 5 min records between January 10 and 19 2001.
252K
. R. N
. Anthony
& O
. Hoegh-
Guldberg
© 2003 B
ritish E
cological Society, F
unctional Ecology,
17, 246–
259
Table 2. Estimates of photosynthetic, tissue and skeletal parameters for Montipora monasteriata sampled from open (op), overhang (oh) and cave (c) habitats in a reef groove at Wistari Reef; also shown are theresults of one-way s for comparisons of parameter values between habitats
Open Overhang Cave
Response parameter Mean SE N Mean SE N Mean SE N F P Post hoc test
Tissue composition and growth characteristicsProtein (mg cm−2) 6·86 0·59 9 5·69 0·75 8 3·46 0·42 8 8·29 0·002** oh > ca, op > caChlorophyll a (µg cm−2) 35·0 4·03 9 36·6 3·44 8 41·2 4·49 8 0·64 0·537 NSRatio of chlorophyll a to protein (µg mg−1) 5·34 0·63 9 7·66 1·60 8 14·1 3·03 8 9·70‡ <0·001*** ca > oh, ca > opZooxanthella concentration (106 cells cm−2) 1·56 0·14 8 2·00 0·21 8 1·61 0·20 8 1·75 0·200 NSZooxanthellae per unit protein (106 cells mg−1) 0·25 0·04 8 0·40 0·06 8 0·58 0·11 7 4·81 0·020* ca > opChlorophyll a per zooxanthella (pg) 20·8 1·33 8 19·1 1·92 8 22·0 2·03 7 0·71 0·506 NSThickness of skeletal plate 2 cm from edge (mm) 7·12 0·65 9 4·24 0·73 8 2·87 0·33 8 13·4† <0·001*** op > oh, op > caSkeletal density (g cm−3) 2·07 0·01 9 2·03 0·02 8 1·98 0·03 8 2·00 0·763 NS
Photosynthetic parametersα (mol O2 mol photons−1) 0·052 0·002 8 0·071 0·010 6 0·096 0·013 8 7·90‡ 0·003** ca > opα (mol O2 mg protein−1 mol photons−1) 0·008 0·001 8 0·015 0·002 6 0·030 0·005 8 23·4‡ <0·001*** ca > oh > opPmax (µmol O2 cm−2 h−1) 3·92 0·12 8 3·24 0·40 6 2·74 0·31 8 4·83† 0·020* op > caPmax (µmol O2 mg protein−1 h−1) 0·60 0·06 8 0·67 0·08 6 0·89 0·16 8 1·89 0·179 NSIk (µmol photons m−2 s−1) 211 7·88 8 127 3·15 6 80·8 2·85 8 209‡ <0·001*** op > oh > caRdark (µmol O2 cm−2 h−1) −1·33 0·13 8 −0·70 0·19 6 −0·43 0·09 7 13·26 0·001*** op > oh, op > caRdark (µmol O2 mg protein−1 h−1) −0·21 0·04 8 −0·15 0·04 6 −0·13 0·04 8 1·49 0·251 NS
(µmol O2 cm−2 h−1) −1·65 0·12 8 −1·35 0·18 6 −1·21 0·20 8 1·81 0·190 NS (µmol O2 mg protein−1 h−1) −0·25 0·02 8 −0·28 0·04 6 −0·40 0·10 8 1·60 0·227 NS
Key to symbols: Pmax, maximum rate of photosynthesis (gross); α, initial slope of the P–I curve; Ik, subsaturation irradiance; Rdark, rate of dark respiration at night; , rate of dark respiration following exposure to maximum experimental irradiance (≈1300 µmol photons m−2 s−1). Assumptions of normality and variance homogeneity were tested using the Kolmogorov–Smirnov one-sample test and Levene’s test, respectively (Sokal & Rohlf 1995).†Data showed variance heterogeneity regardless of transformation, and were analysed untransformed.‡Data were log-transformed prior to statistical analyses to remove variance heterogeneity. Means and SEs are untransformed.
′Rdark
′Rdark
′Rdark
253Habitat light variation and coral photophysiology
© 2003 British Ecological Society, Functional Ecology, 17, 246–259
groups when normalized to surface area. Overall,these results indicate that growth in the shaded micro-habitats maximizes the ratio of surface area to coralbiomass and skeleton, as well as the content of algaeand photopigment per unit biomass, in combinationmaximizing light interception per unit biomass andper unit mass of photosynthetic pigments.
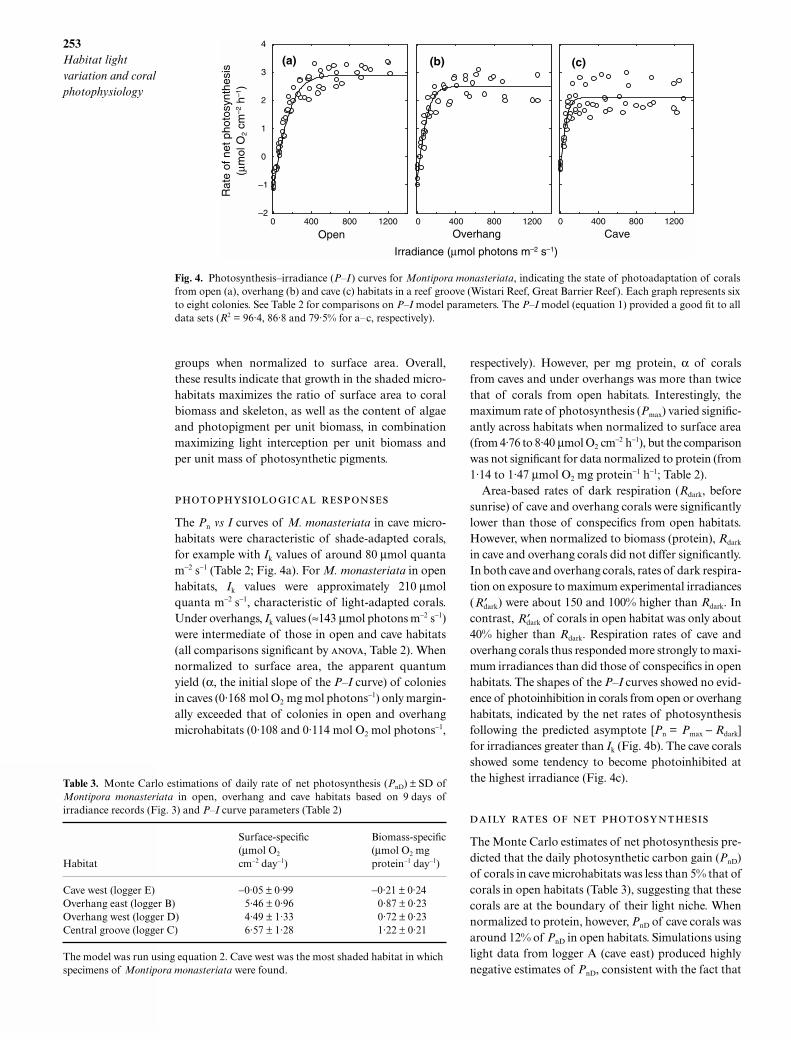
The Pn vs I curves of M. monasteriata in cave micro-habitats were characteristic of shade-adapted corals,for example with Ik values of around 80 µmol quantam−2 s−1 (Table 2; Fig. 4a). For M. monasteriata in openhabitats, Ik values were approximately 210 µmolquanta m−2 s−1, characteristic of light-adapted corals.Under overhangs, Ik values (≈143 µmol photons m−2 s−1)were intermediate of those in open and cave habitats(all comparisons significant by , Table 2). Whennormalized to surface area, the apparent quantumyield (α, the initial slope of the P–I curve) of coloniesin caves (0·168 mol O2 mg mol photons−1) only margin-ally exceeded that of colonies in open and overhangmicrohabitats (0·108 and 0·114 mol O2 mol photons−1,
respectively). However, per mg protein, α of coralsfrom caves and under overhangs was more than twicethat of corals from open habitats. Interestingly, themaximum rate of photosynthesis (Pmax) varied signific-antly across habitats when normalized to surface area(from 4·76 to 8·40 µmol O2 cm−2 h−1), but the comparisonwas not significant for data normalized to protein (from1·14 to 1·47 µmol O2 mg protein−1 h−1; Table 2).
Area-based rates of dark respiration (Rdark, beforesunrise) of cave and overhang corals were significantlylower than those of conspecifics from open habitats.However, when normalized to biomass (protein), Rdark
in cave and overhang corals did not differ significantly.In both cave and overhang corals, rates of dark respira-tion on exposure to maximum experimental irradiances( ) were about 150 and 100% higher than Rdark. Incontrast, of corals in open habitat was only about40% higher than Rdark. Respiration rates of cave andoverhang corals thus responded more strongly to maxi-mum irradiances than did those of conspecifics in openhabitats. The shapes of the P–I curves showed no evid-ence of photoinhibition in corals from open or overhanghabitats, indicated by the net rates of photosynthesisfollowing the predicted asymptote [Pn = Pmax − Rdark]for irradiances greater than Ik (Fig. 4b). The cave coralsshowed some tendency to become photoinhibited atthe highest irradiance (Fig. 4c).
The Monte Carlo estimates of net photosynthesis pre-dicted that the daily photosynthetic carbon gain (PnD)of corals in cave microhabitats was less than 5% that ofcorals in open habitats (Table 3), suggesting that thesecorals are at the boundary of their light niche. Whennormalized to protein, however, PnD of cave corals wasaround 12% of PnD in open habitats. Simulations usinglight data from logger A (cave east) produced highlynegative estimates of PnD, consistent with the fact that
Fig. 4. Photosynthesis–irradiance (P–I ) curves for Montipora monasteriata, indicating the state of photoadaptation of coralsfrom open (a), overhang (b) and cave (c) habitats in a reef groove (Wistari Reef, Great Barrier Reef). Each graph represents sixto eight colonies. See Table 2 for comparisons on P–I model parameters. The P–I model (equation 1) provided a good fit to alldata sets (R2 = 96·4, 86·8 and 79·5% for a–c, respectively).
′Rdark
′Rdark
Table 3. Monte Carlo estimations of daily rate of net photosynthesis (PnD) ± SD ofMontipora monasteriata in open, overhang and cave habitats based on 9 days ofirradiance records (Fig. 3) and P–I curve parameters (Table 2)
Habitat
Surface-specific (µmol O2 cm−2 day−1)
Biomass-specific(µmol O2 mg protein−1 day−1)
Cave west (logger E) −0·05 ± 0·99 −0·21 ± 0·24Overhang east (logger B) 5·46 ± 0·96 0·87 ± 0·23Overhang west (logger D) 4·49 ± 1·33 0·72 ± 0·23Central groove (logger C) 6·57 ± 1·28 1·22 ± 0·21
The model was run using equation 2. Cave west was the most shaded habitat in which specimens of Montipora monasteriata were found.
254K. R. N. Anthony & O. Hoegh-Guldberg
© 2003 British Ecological Society, Functional Ecology, 17, 246–259
no hermatypic (photosymbiotic) corals could be foundin such dark microhabitats. Surprisingly, PnD underoverhangs ranged from 60 to 83% of that in openhabitats. The low standard deviation for all estimates
of PnD was due, in part, to the low variability in dailyirradiance during the 9-day period.
The results of sensitivity analyses indicated that PnD
of all coral groups was more sensitive to changes inRdark than to any other P–I curve parameter, but moreso for corals in caves than for those under overhangsand in open habitats (Table 4). Specifically, a 10%change in Rdark of cave specimens produced an almost10-fold greater effect on PnD than did changes in Pmax.In contrast, changes in Rdark and Pmax of corals in openhabitats produced largely similar effects (although inopposite directions). Also, PnD of the cave corals washighly sensitive to changes in apparent quantum yield(α), whereas the opposite was the case for corals inopen habitats.
Groups of M. monasteriata transplanted from cave andopen habitats showed acclimation responses mainlythrough changes in maximum apparent electrontransport rate (ETRmax) and subsaturation of ETR(Ik-ETR, Fig. 5). However, during the 3-week experi-ment there were no significant changes in chlorophylla content (Fig. 6). Three main points are worth noting.First, the apparent ETRmax of corals transplantedfrom open habitats to under the overhang reachedvalues similar to those of conspecifics transplantedfrom shaded habitats to under the overhang (compari-son not significant by , Table 5). Second, thepoint of subsaturation (Ik-ETR) for open-habitat corals
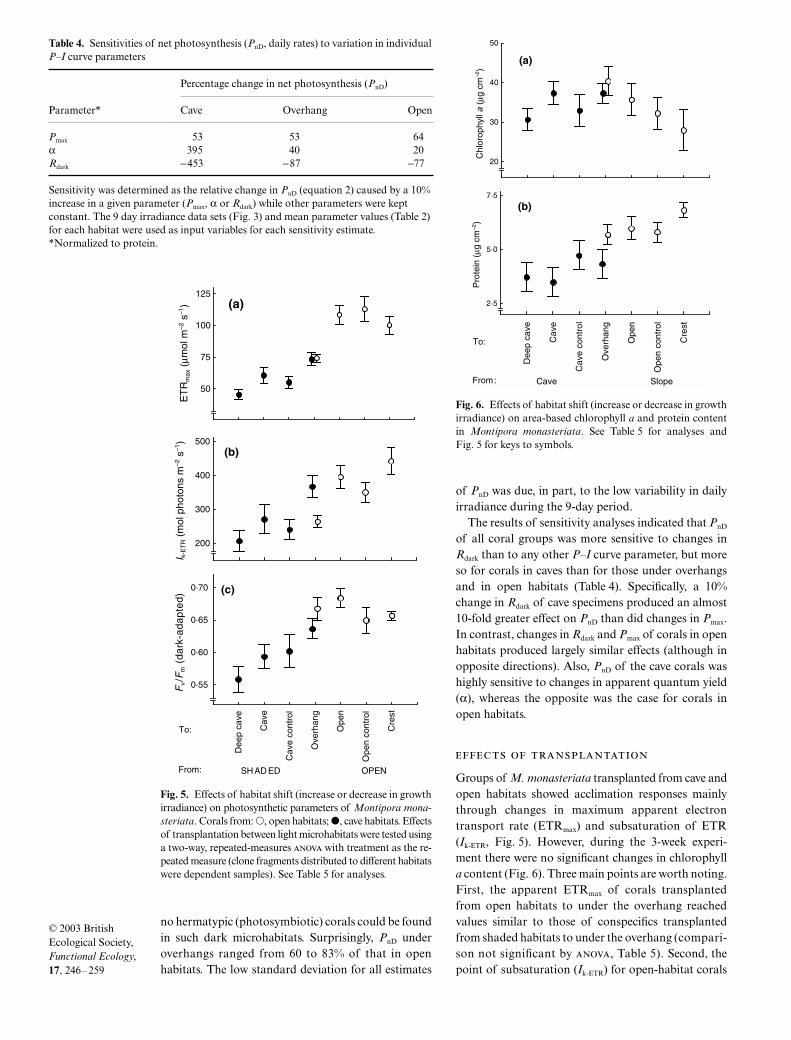
Fig. 6. Effects of habitat shift (increase or decrease in growthirradiance) on area-based chlorophyll a and protein contentin Montipora monasteriata. See Table 5 for analyses andFig. 5 for keys to symbols.
Table 4. Sensitivities of net photosynthesis (PnD, daily rates) to variation in individualP–I curve parameters
Percentage change in net photosynthesis (PnD)
Parameter* Cave Overhang Open
Pmax 53 53 64α 395 40 20Rdark −453 −87 −77
Sensitivity was determined as the relative change in PnD (equation 2) caused by a 10% increase in a given parameter (Pmax, α or Rdark) while other parameters were kept constant. The 9 day irradiance data sets (Fig. 3) and mean parameter values (Table 2) for each habitat were used as input variables for each sensitivity estimate.*Normalized to protein.
Fig. 5. Effects of habitat shift (increase or decrease in growthirradiance) on photosynthetic parameters of Montipora mona-steriata. Corals from: �, open habitats; �, cave habitats. Effectsof transplantation between light microhabitats were tested usinga two-way, repeated-measures with treatment as the re-peated measure (clone fragments distributed to different habitatswere dependent samples). See Table 5 for analyses.

255Habitat light variation and coral photophysiology
© 2003 British Ecological Society, Functional Ecology, 17, 246–259
moved to under the overhang was significantlylower than that of cave corals moved to under theoverhang (Table 5), signifying overcompensation bythe open-habitat corals in terms of photoacclimation.Third, maximum quantum yield (Fv/Fm, dark-adapted)varied significantly between habitat groups and amongtransplant groups, but there was also significantinteraction between these factors (Table 5), the latterindicating that the two groups responded differentlyto light variation. Specifically, Fv/Fm of open-habitatcorals was approximately 20% higher than that of caveconspecifics. However, Fv/Fm of transplanted cave cor-als was maximized under the overhang (near logger D)and minimized in the darkest cave near logger A(approximately 15% relative decrease), indicating thatphotosystem II of zooxanthellae in the dark-cavetransplants was stressed relative to that of symbiontsunder the overhang. Conversely, Fv/Fm did not varysignificantly among transplant groups in the openhabitat.
Within the shaded and open habitat groups, thethree photophysiological parameters did not vary sig-nificantly among dark cave, cave and control trans-plants, or among open control and crest. The mostshaded group had marginally lower apparent ETRmax,Ik-ETR and Fv/Fm than the remaining groups; however,comparisons were not significant by . These resultsindicate, first, that specimens in the most shaded,native cave habitats (near logger E) have limited abilityto photoacclimate to further reductions in lightregime, but acclimate well to a light increase (cf. gapopening). Second, variations in light habitats in theopen regime (except under overhangs) produced onlyminor variation in photophysiological parameters.
There was no main effect of habitat and transplan-tation on the concentration of chlorophyll a (Fig. 6;Table 5). However, the interaction term for these fac-tors was significant, indicating that the transplantationhad different effects on the two groups. Specifically,chlorophyll a tended to decrease with an increase in light
habitat (overhang to crest habitats) in the open habitat,but was relatively invariant in the shaded regime (over-hang to cave habitats). Consistent with the genericopen-vs-shade comparison above, the protein concen-trations of the open-habitat corals were significantlyhigher than those of the shade-habitat conspecifics,but transplant groups did not vary significantly.
Discussion
The two orders of magnitude lower growth irradiancein cave compared to open microhabitats of the grooveenvironment corresponds to the light attenuationoccurring over more than 50 m depth of open water inoffshore locations (Kirk 1994). Also, such irradiancevariation between the groove microhabitats approx-imates that found among canopy, gap and understoreyin rainforests (Chazdon & Fetcher 1984; Pearcy 1987).Our results suggest that the broad light niche of M.monasteriata is a consequence of physiological flexibil-ity and morphological plasticity, consistent with thepatterns found for leaves of forest plants in gaps andunderstoreys (reviewed by Chazdon et al. 1996).Combined physiological, cellular and morphologicalstrategies for optimizing light capture in spatially andtemporally varying light regimes have been studiedextensively in rainforest plants (Oberbauer & Strain1986; Pearcy 1987; Chazdon & Kaufmann 1993). Incorals, previous studies have focused mainly on cellu-lar responses of the symbionts (Chang, Prezelin &Trench 1983; Iglesias-Prieto & Trench 1994).
Variations in morphological and biochemical character-istics of M. monasteriata across groove microhabitatswere largely analogous to those reported for sun andshade plants. Specifically, cave specimens had highchlorophyll a content per unit biomass and low biomass
Table 5. Results of for effects of transplantation on apparent ETR vs I curve parameters and tissue properties forMontipora monasteriata among light microhabitats within a reef groove (see Fig. 2 for details of design)
Habitat Transplantation Habitat × Transplantation
F P F P F P
Response parameters (1,10) (3,30) (3,30)
ETRmax (µmol electrons m−2 s−1) 32·47 <0·001*** 38·96 <0·001*** 2·96 0·075 NSIk-ETR (µmol quanta m−2 s−1) 18·91 0·001** 19·33 <0·001*** 3·51 0·027*Fv/Fm (dimensionless) 9·51 0·012* 26·00 <0·001*** 4·09 0·032*Chlorophyll a (µg cm−2) 0·388 0·547 NS 0·93 0·441 NS 3·14 0·040*Protein (mg cm−2) 21·45 <0·001*** 2·76 0·056 NS 0·86 0·475 NS
The factor Transplantation was analysed as repeated measures because specimens transplanted across microhabitats originated from one colony (N = 6 colonies per transplant group). Because one to three control samples were lost from each habitat group, their cells were replaced by the means of the remaining control samples to maintain a balanced design. Degrees of freedom are given in parentheses (effect, error). See Figs 5 and 6 for comparison of transplant groups.†Data showed variance heterogeneity (Levene’s test) regardless of transformation, and were analysed untransformed.

256K. R. N. Anthony & O. Hoegh-Guldberg
© 2003 British Ecological Society, Functional Ecology, 17, 246–259
per unit surface area, and displayed a growth patterndirected towards expanding colony surface arearather than volume (indicated by thin skeletal plates).Also, colonies in caves and under overhangs formedvertical or oblique plates, further maximizing the cap-ture of incident light. Similarly, leaves of shade plantsare generally thin, and have a relatively large amountof photosynthetic material and a low proportionof nonphotosynthetic tissue (Chazdon & Kaufman1993; reviewed by Lambers et al. 1997). Further, under-storey plants may display leaf orientations that maximizelight capture (Takenaka, Takahashi & Kohyama2001). In contrast to patterns found for chloroplasts inleaves of shade plants (Lambers et al. 1997) and zoo-xanthellae of other species (Chang et al. 1983; Porteret al. 1984), shaded colonies of M. monasteriata in thisstudy did not have a significantly higher chlorophyll aconcentration per algal cell. Recent studies of cul-tured zooxanthellae from Montipora, however, showonly 20% variation in chlorophyll a content across abroad range of light regimes (Iglesias-Prieto & Trench1994). It is possible that different within-colony distri-butions of zooxanthellae in the two habitats may havemasked variations in cellular pigments. Specifically,the undersides of colonies in open habitats had tissueand contained zooxanthellae, but most corals in cavesdid not. Morphological plasticity to maximize lightinterception and use has been suggested previouslyfor corals across depth ranges (Barnes 1973), inparticular in inshore turbid habitats (Willis 1985).These studies, however, drew inferences from compari-sons of colony shape and did not measure photophysio-logical parameters.
Phototrophic organisms in shaded and semishadedmicrohabitats, such as reef grooves and forest gaps andunderstories, are facing two major challenges: maximi-zation of irradiance capture, and protection from shortperiods of peak irradiances. First, in cave habitats (thisstudy) irradiance is severely reduced throughout theday. This is also true for forest understories, except forthe occurrence of sunflecks (Pearcy 1994; Watlinget al. 1997). To sustain a positive (or zero) energy bal-ance in habitats that are largely permanently shaded,phototrophs must maximize their capture and utiliza-tion of weak irradiance and minimize their energycosts (Chazdon & Kaufmann 1993; Mulkey, Wright& Smith 1993). In this study, cave specimens ofM. monasteriata demonstrated all such responses:thinning and widening of tissue and skeleton normalto the direction of irradiance; a high apparent quan-tum yield (α); low subsaturation irradiance (Ik); andlow rates of dark respiration (Rdark). Moreover, on a per-biomass basis, the photosynthetic capacity (Pmax) of cavecorals was slightly higher than that of conspecifics inopen habitats, an opposite response to that reported
for forest plants (Mulkey et al. 1993). Second, for cor-als under reef overhangs or for plants in forest gaps,the effective day-length may be shortened significantlydepending on the orientation of the reef walls relativeto the path of the sun (Brakel 1979) or the size of thegap (Chazdon & Fetcher 1984). Under overhangsin the reef groove of this study, the daily light vs timeprofile was compressed to only 2–3 h above the sub-saturating irradiances. Photosynthetic responses insuch habitats are therefore likely to be a compromisebetween maximum light capture during the shadedwindow of the day, and protection from excess lightduring the unshaded window. The lack of photoinhibi-tion at irradiances up to 1300 µmol m−2 s−1 in specimensfrom under overhangs (Fig. 4b) suggests that thesecorals are capable of avoiding photosynthetic stressduring short periods of irradiance, allowing a relativelyhigh photosynthetic efficiency (high α, low Ik). As aresult, despite 70% lower growth irradiances underoverhangs relative to open habitats; the daily rates ofnet photosynthesis (PnD) in corals under overhangswere 60–83% of those in open habitats.
Our study is unique in demonstrating that specimensof a photosymbiotic coral species, separated by only afew metres, inhabit light regimes that differ by twoorders of magnitude, resulting in contrasting scopesfor growth and reproduction. Despite the photophysi-ological, metabolic and morphological plasticity ofM. monasteriata, daily rates of net photosynthesis inthe most shaded habitats are likely to cover only main-tenance costs and a minimal growth rate. Such con-trasts in resources and scope for energy investmentacross a wide niche suggest an opportunistic strategyof habitat use whereby only part of the local popu-lation maintains high productivity (open-habitatcorals), while the remainder of the population (cavecorals) escapes competition at the expense of produc-tivity. Several studies have suggested similar diversi-fication of scope for growth and reproduction withinpopulations of rainforest plants along successionalgradients (Strauss-Debenedetti & Bazzaz 1996), butnot previously in coral populations.
The results of the sensitivity analyses indicate thata reduced rate of dark respiration (Rdark) is the keymechanism for maintaining a positive rate of daily netphotosynthesis in shaded and semishaded habitats.Because dark respiration represents a relatively hightax on the daily energy budget of corals with low ratesof energy acquisition and/or short periods of irradi-ance, even a small reduction in the hourly rate of darkrespiration has major implications for the energy bal-ance in marginal light habitats. The daily rates of netphotosynthesis of corals in caves were also highly sen-sitive to changes in apparent quantum yield (α), butthis pattern was reversed for corals in open habitatsand under overhangs. In marginal light habitats such

257Habitat light variation and coral photophysiology
© 2003 British Ecological Society, Functional Ecology, 17, 246–259
as caves, mechanisms for downregulating dark respira-tion and maximizing photosynthetic efficiency arethus likely to be more critical for coral energetics thanmechanisms for maximizing Pmax. The lower area-based Rdark of corals in caves and under overhangs waslargely explained by their lower tissue mass (60% pro-tein relative to the open-habitat conspecifics), as alsofound for shade plants (Chazdon & Kaufmann 1993).Other factors explaining the lower dark respiration ofshaded corals are their thinner tissue layer and lowerlevels of respiratory substrate (e.g. glycerol transloca-tion from zooxanthellae to host), and lower growthrates, analogous to the pattern found in vascularplants (Lambers et al. 1997). The greater increase inrate of respiration by corals from caves and overhangsfollowing exposure to high irradiance may indicate ahigh capacity for metabolic activity or growth onceirradiance increases. Such light-induced metabolicactivity has been observed for plants in forest under-storeys following the opening of tree-fall gaps (Strauss-Debenedetti & Bazzaz 1996), and may also, tentatively,facilitate rapid growth following disturbances thatenhance the local light regime (e.g. gap openings) incoral reef habitats (Stimson 1985). It is likely, however,that the strong light response of respiration ratesin shade corals represents a stress response by bothzooxanthellae and coral host, in that maximumexperimental irradiance was several-fold that experi-enced naturally by the cave corals.
The results of the transplantation from shaded tosemi-open habitat illustrated a high capacity inM. monasteriata to acclimate to upshifts and down-shifts in growth irradiance such as those likely to occurfollowing physical disturbances on reefs (Stimson1985) or resulting from competition for space (Lang& Chornesky 1990). Specimens transplanted fromopen habitats to underneath the overhang (about 70% re-duction in growth irradiance) acclimated fully within3 weeks of transplantation. [We argue that, based onthe results of previous transplantation experimentsacross light regimes (Anthony & Hoegh-Guldberg2003), 3 weeks is ample time for full biochemical photo-acclimation to occur.] Most notably, colonies fromthe open habitat reached significantly lower points ofsubsaturation (Ik-ETR) than did colonies transplantedfrom caves to under the overhang (Fig. 5; Table 5),indicating different photoacclimative responses follow-ing upshifts or downshifts in growth irradiance. Therelative overcompensation of Ik-ETR by cave specimensmoved out under overhangs is likely to signify a pro-tective mechanism against photodamage during peakirradiances. Conversely, the relatively large reductionin Ik-ETR of specimens transplanted from the openhabitat to underneath the overhang may indicate anadaptive mechanism to offset energy deficits. Based
on the respirometry results, predicted Ik-ETR for over-hang corals would be intermediary between caveand open-habitat corals. The difference in acclimatedIk-ETR of the two groups under the overhang cannotbe explained by differences in tissue thickness, as therelatively thick tissue of open-habitat corals wouldcounteract a reduction in their Ik, and area-based pro-tein did not change significantly during the experi-ment. The latter is expected, given the relatively shortduration (3 weeks) of the experiment. Further, thehypothesis that differences in Ik-ETR were attributable todifferences in symbiont taxonomy (Iglesias-Prieto &Trench 1994) could not be supported in this study, assymbionts of all coral samples belonged to the sameclade (C17 of Symbiodinium sp., T. LaJeunesse, unpub-lished results). Colonies of M. monasteriata found intheir most shaded microhabitat (western cave) showedonly limited acclimation to further reduction ingrowth irradiance, as indicated by a lack of change inIk-ETR. Also, the slight reductions (≈15%) in maximumelectron transport rate, maximum quantum yield andconcentrations of chlorophyll a collectively indicate adegree of photosynthetic stress induced by light limi-tation. Conversely, colonies moved from open-slopehabitat to reef crest showed a slight increase in Ik-ETR
(relative to open-habitat controls), signifying photo-acclimation to increased growth irradiance.
In conclusion, these results demonstrate that thehabitat distribution of M. monasteriata in spur-and-groove reef environments spans a continuum from extremelow- to high-light environments, analogous to the habitatdistribution of some plant species in forest gaps andunderstories. The wide light niche of M. monasteriatais facilitated by physiological as well as morphologicalacclimations, but results in contrasting scope forgrowth across light environments. The capacity of cavespecimens to acclimate well to increasing irradiances(and to respond to high irradiance by increased meta-bolism) suggests that M. monasteriata may share thestrategy of understorey plants that rely on future oppor-tunities to invade more favourable habitats (Strauss-Debenedetti & Bazzaz 1996). The adaptive significanceof extreme niche widths for the population biology ofcorals is unclear and deserves further attention.
Acknowledgements
This study was funded by the Australian ResearchCouncil (grant no. A00105071 to K.R.N.A. andA10009109 to O.H.G.). We thank Roberto Iglesias-Prieto, Ross Jones, Pia Anthony and Melissa Varoy fortheir critical reading and valuable comments. This iscontribution number 71 of the Centre for Coral ReefBiodiversity at James Cook University.
References
Anthony, K.R.N. & Fabricius, K.E. (2000) Shifting roles ofheterotrophy and autotrophy in coral energetics under

258K. R. N. Anthony & O. Hoegh-Guldberg
© 2003 British Ecological Society, Functional Ecology, 17, 246–259
varying turbidity. Journal of Experimental Marine Biologyand Ecology 252, 221–253.
Anthony, K.R.N. & Hoegh-Guldberg, O. (2003) Kinetics ofcoral photoacclimation. Oecologia 134, 23–31.
Anthony, K.R.N., Connolly, S.R. & Willis, B.L. (2002) Com-parative analysis of tissue and skeletal growth in corals.Limnology and Oceanography 47, 1417–1429.
Barnes, D.J. (1973) Growth in colonial scleractinians. Bulletinof Marine Science 23, 280–298.
Barnes, D.J. & Chalker, B.E. (1990) Calcification and photo-synthesis in reef-building corals and algae. Ecosystems ofthe World: Coral Reefs, Vol. 25 (ed. Z. Dubinsky), pp. 109–131. Elsevier, Amsterdam.
Brakel, W.H. (1979) Small-scale spatial variation in lightavailable to coral reef benthos: quantum irradiance meas-urements from a Jamaican reef. Bulletin of Marine Science29, 406–413.
Chalker, B.E., Dunlap, W.E. & Oliver, J.K. (1983) Bathy-metric adaptations of reef-building corals at Davies Reef,Great Barrier Reef, Australia. II. Light saturation curvesfor photosynthesis and respiration. Journal of Experi-mental Marine Biology and Ecology 73, 37–56.
Chang, S.S., Prezelin, B.B. & Trench, R.K. (1983) Mechan-isms of photoadaptation in three strains of the symbioticdinoflagellate Symbiodinium microadriaticum. MarineBiology 76, 219–229.
Chazdon, R.L. & Fetcher, N. (1984) Photosynthetic lightenvironments in a lowland tropical rainforest in CostaRica. Journal of Ecology 72, 553–564.
Chazdon, R.L. & Kaufmann, S. (1993) Plasticity of leaf ana-tomy of two rain forest shrubs in relation to photosyntheticlight acclimation. Functional Ecology 7, 385–394.
Chazdon, R.L., Pearcy, R.W., Lee, D.W. & Fetcher, N. (1996)Photosynthetic responses of tropical forest plants to contrast-ing light environments. Tropical Forest Plant Ecophysiology(eds S.S. Mulkey, R.L. Chazdon & A.P. Smith), pp. 5–55.Chapman & Hall, London.
Crossland, C.J. (1987) In situ release of mucus and DOC-lipidfrom the coral Acropora variabilis and Stylophora pistillata.Coral Reefs 6, 35–42.
Dinesen, Z.D. (1983) Shade dwelling corals of the GreatBarrier Reef, Australia. Marine Ecology Progress Series 10,173–186.
Falkowski, P.G., Dubinsky, Z., Muscatine, L. & Porter, J.(1984) Light and the bioenergetics of a symbiotic coral.Bioscience 34, 705–709.
Falkowski, P.G., Jokiel, P.L. & Kinzie Iii, R.A. (1990) Irradi-ance and corals. Ecosystems of the World: Coral Reefs, Vol.25 (ed. Z. Dubinsky), pp. 89–107. Elsevier, Amsterdam.
Hatcher, B.G. (1990) Coral reef primary productivity: a hier-archy of pattern and process. Trends in Ecology and Evolution5, 149–155.
Hoegh-Guldberg, O. & Jones, R.J. (1999) Photoinhibitionand photoprotection in symbiotic dinoflagellates fromreef-building corals. Marine Ecology Progress Series 183,73–86.
Iglesias-Prieto, R. & Trench, R.K. (1994) Acclimation andadaptation to irradiance in symbiotic dinoflagellates. I.Responses of the photosynthetic unit to changes in photonflux density. Marine Ecology Progress Series 113, 163–175.
Jassby, A.D. & Platt, T. (1976) Mathematical formulation ofthe relationship between photosynthesis and light for phyto-plankton. Limnology and Oceanography 21, 540–547.
Jeffrey, S.W. & Humphrey, G.F. (1975) New spectrophoto-metric equations for determining chlorophyls a, b, c1 andc2 in higher plants, algae and natural phytoplankton. Bio-chemie und Physiologie der Pflanzen 167, 191–194.
Jones, R.J. & Hoegh-Guldberg, O. (2001) Diurnal changes inthe photochemical efficiency of the symbiotic dinoflagellates(Dinophyceae) of corals: photoprotection, photoactivation
and the relationship to coral bleaching. Plant, Cell andEnvironment 24, 89–99.
Kirk, J.T.O. (1994) Light and Photosynthesis in AquaticEcosystems, 2nd edn. Cambridge University Press,Cambridge, UK.
Kitajima, K. (1994) Relative importance of photosynthetictraits and allocation patterns as correlates of seedlingshade tolerance of 13 tropical trees. Oecologia 98, 419–428.
Lambers, H., Chapin Iii, F.S. & Pons, T.L. (1997) Plant Phys-iological Ecology. Springer, Berlin.
Lang, J.C. & Chornesky, E.A. (1990) Competition betweenscleractinian reef corals: a review of mechanisms andeffects. Coral Reefs, Vol. 25 (ed. Z. Dubinsky), pp. 209–252. Elsevier, Amsterdam.
Marsh, J.A. (1970) Primary productivity of reef-buildingcalcareous red algae. Ecology 51, 255–263.
Maxwell, K. & Johnson, G.N. (2000) Chlorophyll fluores-cence – a practical guide. Journal of Experimental Botany51, 659–668.
Mulkey, S.S., Wright, S.J. & Smith, A.P. (1993) Comparativephysiology and demography of three neotropical forestshrubs – alternative shade-adaptive character syndromes.Oecologia 96, 526–536.
Muscatine, L. (1990) The role of symbiotic algae in carbonand energy flux in reef corals. Ecosystems of the World:Coral Reefs, Vol. 25 (ed. Z. Dubinsky), pp. 75–87. Elsevier,Amsterdam.
Oberbauer, S.F. & Strain, B.R. (1986) Effects of canopy posi-tion and irradiance on the leaf physiology and morphologyof Pentaclethra macroloba (Mimisaceae). American Journalof Botany 73, 409–416.
Oliver, J.K., Chalker, B.E. & Dunlap, W.E. (1983) Bathymetricadaptations of reef-building corals at Davies Reef, GreatBarrier Reef, Australia. I. Long-term growth responses ofAcropora formosa. Journal of Experimental Marine Biologyand Ecology 73, 11–35.
Pearcy, R.W. (1987) Photosynthetic gas exchange responsesof Australian tropical forest trees in canopy, gap andunderstory micro-environments. Functional Ecology 1,169–178.
Pearcy, R.W. (1994) Photosynthetic response to sunflecksand light gaps: mechanisms and constraints. Photoinhibi-tion of Photosynthesis: from Molecular Mechanisms to theField (eds N.R. Baker & J.R. Bowyer), pp. 255–271. BIOSScientific, Oxford.
Porter, J.W., Muscatine, L., Dubinsky, Z. & Falkowski, P.G.(1984) Primary production and photoadaptation in light-adapted and shade-adapted colonies of the symbioticcoral, Stylophora pistillata. Proceedings of the Royal Societyof London Series B 222, 161–180.
Sheppard, C.R.C. (1981) Illumination and the coral com-munity beneath tabular Acropora species. Marine Biology64, 53–58.
Sokal, R.R. & Rohlf, F.J. (1995) Biometry, 3rd edn. W.H.Freeman, New York.
Spencer-Davies, P. (1991) Effect of daylight variations on theenergy budget of shallow-water corals. Marine Biology108, 137–144.
Statistica (1999) Release 5.1. StatSoft Inc., Tulsa, OK, USA.Stimson, J. (1985) The effect of shading by the table coral
Acropora hyacinthus on understory corals. Ecology 66, 40–53.
Strauss-Debenedetti, S. & Bazzaz, F.A. (1996) Photosynthesiccharacteristics of tropical trees along successional gradients.Tropical Forest Plant Ecophysiology (eds S.S. Mulkey,R.L. Chazdon & A.P. Smith), pp. 162–186. Chapman &Hall, London.
Takenaka, A., Takahashi, K. & Kohyama, T. (2001) Optimalleaf display and biomass partitioning for efficient light

259Habitat light variation and coral photophysiology
© 2003 British Ecological Society, Functional Ecology, 17, 246–259
capture in an understorey palm, Licuala arbuscula. Func-tional Ecology 15, 660–668.
Veron, J.E.N. (1986) Corals of Australia and the Indo-Pacific,2nd edn. University of Hawaii Press, Honolulu, Hawaii.
Watling, J.R., Robinson, S.R., Woodrow, I.E. & Osmond,C.B. (1997) Responses of rainforest understorey plants toexcess light during sunflecks. Australian Journal of PlantPhysiology 24, 17–25.
Willis, B.L. (1985) Phenotypic plasticity versus phenotypicstability in the reef corals Turbinaria mesenterina andPavona cactus. Proceedings of the Fifth International CoralReef Congress, Tahiti, Vol. 4, pp. 107–112. AntenneMuseum National d’Histoire Naturel, Moorea.
Received 22 July 2002; revised 28 November 2002; accepted 1 December 2002





























