Embed Size (px)
Citation preview

This article was downloaded by: [Ivana Svitkova]On: 29 October 2012, At: 06:12Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Plant Biosystems - An International Journal Dealingwith all Aspects of Plant Biology: Official Journal of theSocieta Botanica ItalianaPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/tplb20
An expert-based classification of high-altitudearctic-alpine vegetation of the class Carici rupestris-Kobresietea Ohba 1974: Achievements and obstaclesI. Svitková a & J. Šibík aa Institute of Botany, Slovak Academy of Sciences, Dúbravská cesta 9, SK-845 23, Bratislava,Slovakia
Accepted author version posted online: 09 Oct 2012.Version of record first published: 26Oct 2012.
To cite this article: I. Svitková & J. Šibík (): An expert-based classification of high-altitude arctic-alpinevegetation of the class Carici rupestris-Kobresietea Ohba 1974: Achievements and obstacles, Plant Biosystems - AnInternational Journal Dealing with all Aspects of Plant Biology: Official Journal of the Societa Botanica Italiana,DOI:10.1080/11263504.2012.736422
To link to this article: http://dx.doi.org/10.1080/11263504.2012.736422
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form toanyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug doses shouldbe independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims,proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly or indirectly inconnection with or arising out of the use of this material.

An expert-based classification of high-altitude arctic-alpine vegetationof the class Carici rupestris-Kobresietea Ohba 1974: Achievements andobstacles
I. SVITKOVA & J. SIBIK
Institute of Botany, Slovak Academy of Sciences, Dubravska cesta 9, SK-845 23 Bratislava, Slovakia
AbstractThe specific species-rich high-altitude vegetation of the class Carici rupestris-Kobresietea bellardii Ohba 1974 (CK), with theoccurrence of many arctic-alpine and endemic species, was chosen for a case study. The analyses were based on a dataset of37,204 phytosociological releves from the Slovak Vegetation Database. The traditional classification of the class CK, basedon cluster analyses, was reproduced satisfactorily by means of formalised classification, based on the formal definitionscreated by the Cocktail method together with the frequency-positive fidelity index affiliation. Unequivocal assignmentcriteria for all eight associations of both alliances [Oxytropido-Elynion Br.-Bl. (1948) 1949 and Festucion versicoloris Krajina1933] of the class CK were formulated. The formal delimitations followed the traditional ones very well. It was demonstratedthat the results of applying the formal definitions created on the basis of a large, geographically stratified dataset capturing theoccurrence of all vegetation types in Slovakia were highly similar in comparison with the traditional classification based onthe results of cluster analysis. The reliability and the pros and cons of the expert system are also discussed.
Keywords: Alpine communities, Cocktail method, frequency-positive fidelity index, syntaxonomy, Western Carpathians
Introduction
Phytosociological research has a long history in
Europe (cf. Schaminee et al. 2011), and hence in
Slovakia as well, where it goes back to the beginning
of the 20th century. Although the description of the
vegetation (including its characteristics, structure,
dynamics and variability, and its consequent classi-
fication in a particular region – e.g. country,
mountain range, continent) is quite often not among
the main goals of scientific research, it serves as an
irreplaceable tool for the generation of a conceptual
basis for phytosociology and habitat mapping (Chy-
try 2000). Phytosociological studies produced local
and regional overviews of vegetation types based on
the classification of the sampled releves (Schaminee
et al. 2009). Local studies were progressively
supplanted by vegetation surveys of larger regions.
The initial methods of manual and intuitive evalua-
tion and comparison of vegetation were later
replaced with the development of more powerful
computers and software for vegetation data analysis,
by more unbiased statistical analyses and methods
(Mucina & van der Maarel 1989; Chytry & Tichy
2003; Podani 2005, 2006; Jarolımek & Sibık 2008;
Carni et al. 2011).
During the last decade, several studies summarised
the vegetation overviews in individual countries
(Rodwell 1990, 1991, 1992; Grabherr & Mucina
1993; Valachovi�c et al. 1995; Jarolımek et al. 1997;
Valachovi�c 2002; Kliment & Valachovi�c 2007) and
were followed by supra-regional syntheses (Dubravko-
va et al. 2010; Michl et al. 2010; Sibık et al. 2010).
Simultaneous with these studies, there was an
effort to create methods that do not have the
disadvantages of traditional numerical classifications,
which in some cases do not take into account the
general diagnostic value of the species or the fact that
it is based on the current structure of the analysed
datasets only and hence it is valid only in the used
partial datasets. These new methods are usually
referred to as formalised approaches and should aim
at precise definitions of classification criteria and
algorithms (Chytry 2000). The Cocktail method
Correspondence: I. Svitkova, Institute of Botany, Slovak Academy of Sciences, Dubravska cesta 9, SK-845 23 Bratislava, Slovakia. Email:
Plant Biosystems, 2012; 1–18, iFirst article
ISSN 1126-3504 print/ISSN 1724-5575 online ª 2012 Societa Botanica Italiana
http://dx.doi.org/10.1080/11263504.2012.736422
Dow
nloa
ded
by [
Ivan
a Sv
itkov
a] a
t 06:
12 2
9 O
ctob
er 2
012

(Bruelheide 1995, 2000) has become one of the most
used methods in classification of large datasets to
determine the major vegetation types by providing
unequivocal criteria for assignment of releves to the
plant associations (Bruelheide & Chytry 2000). The
first complex national vegetation overview based on
the formalised criteria was the series Vegetation of the
Czech Republic (Chytry 2007, 2009), which followed
numerous studies dealing with defining formalised
criteria for particular vegetation types in different
regions (Ko�cı et al. 2003; Boublık et al. 2007; Dıte
et al. 2007, Role�cek 2007, Silc & Carni 2007,
Hrivnak et al. 2008, Boublık 2010, Rozbrojova et al.
2010).
The high-altitude vegetation of Slovakia (the
Western Carpathians) was analysed only recently
(Kliment & Valachovi�c 2007), using traditional
methods based on cluster analyses. The specifics of
high-altitude vegetation, its patchiness and abrupt
changes in individual ecological gradients causing
great variability were all taken into account when
performing the cluster analyses, which were often
achieved in several steps by experts specialising in
individual vegetation types (cf. Kliment et al. 2010).
Recently, alongside the vegetation surveys of
Slovakia, the preparation of an expert system for
the identification of individual vegetation units in
Slovakia has started, using external a priori defined
criteria created by the Cocktail method. The first
publication to use this system is a survey of meadow
and pasture vegetation (Janisova et al. 2007). Since
the time when the formal definitions were created for
individual vegetation units of meadows and pastures
(Janisova et al. 2007), a number of releves, primarily
of high-altitude vegetation, have increased signifi-
cantly in the Slovak Vegetation Database (SVD, cf.
Sibıkova et al. 2009). Moreover, the quality of data
already stored in the database increased dramatically
as a result of the more accurate post-hoc identification
of the geographical coordinates of older releves, more
precise control of data input, and other changes.
Hence, it was necessary to take this fact into account in
this study and to critically evaluate the constituents of
some previously defined species groups.
The class Carici rupestris-Kobresietea bellardii Ohba
1974 (CK) represents a specific species-rich high-
altitude vegetation, which includes many arctic-alpine
and endemic species (Sibıkova et al. 2010). It was
chosen as a case study due to several facts. It is a
natural type of vegetation, only minimally influenced
by human activities and individual associations are
well determined. The basic question of our analyses
was if the application of formal definitions created on
the basis of an extensive dataset capturing the
occurrence of all vegetation types in Slovakia will yield
similar results in comparison with the traditional
classification based on the results of cluster analysis.
The main goals of this study can be summarised as
follows: (i) to create unequivocal definitions of the
traditional associations using the combinations of
created species groups and to characterise the
successfully delimited associations; (ii) to test the
applicability of the definitions and compare the results
with the traditional classifications taking into account
the reliability of the expert system.
Material and methods
The analyses were based on a dataset of 37,204
phytosociological releves from the SVD (Jarolımek &
Sibık 2008, www.ibot.sav.sk/cdf; Sibıkova et al.
2009; Sibık 2012) and from the private databases
of the authors. The releves were exported and further
analysed in the JUICE software (Tichy 2002). Due
to uneven distribution of releves across the country,
geographically stratified resampling of the dataset
was performed in JUICE software (Tichy 2002)
using a 1.25 longitudinal6 0.75 latitudinal minute
geographical grid (cf. Knollova et al. 2005). The
resulting dataset of 21,142 releves, including mosses
and lichens, was used for all analyses.
Species found in several layers (especially woody
species) were merged into one layer in each releve.
Taxa determined only to the genus level were
excluded from the analysis. Taxa determined to the
subspecies and variety level were merged to the
species level or within broadly conceived taxa. The
nomenclature of the vascular plants generally follows
the checklist by Marhold and Hindak (1998). The
detailed methods of preparing the dataset follow the
one used by Jarolımek and Sibık (2008).
The species groups (Table I) were created in the
geographically stratified dataset of 21,142 releves by
the Cocktail method (Bruelheide 1995, 2000), using
the phi coefficient (Chytry et al. 2002). The species
groups defined by Janisova et al. (2007) were tested
in our dataset and critically evaluated due to
substantial expansion of releves from high altitudes
in our dataset in comparison with dataset of Janisova
et al. (2007). The phi coefficient was used to
measure species’ tendencies to occur together in
the releves included in the dataset. The aim was to
define such species groups, where individual species
have the highest fidelity values. The defined species
groups had at least three members and were
considered present when at least half of the members
were present. The formal definitions (Table II) of the
associations were created by combining the species
groups using the logical operators AND, OR and
AND NOT (Bruelheide 1997) and the cover value of
diagnostically important taxa. Detailed descriptions
of the individual steps involved in creating the
species groups and definitions can be found in the
study by Ko�cı et al. (2003).
2 I. Svitkova and J. Sibık
Dow
nloa
ded
by [
Ivan
a Sv
itkov
a] a
t 06:
12 2
9 O
ctob
er 2
012

The resulting formal definitions were then tested
and applied to the large dataset of 37,204 phytoso-
ciological releves. To assign the releves that were
assigned to more than one association after applying
the formal definition, as well as the releves that did
not suit any of the definitions and were previously
assigned to the class CK (by the original author or by
Petrık et al. 2006), the frequency-positive fidelity
index (FPFI), which compares the similarity of the
species composition of a selected releve to a group of
releves (Tichy 2005), was used. Only those releves
with a FPFI value of more than 5 were finally
assigned to the association.
A synoptic table (see Supplementary material) was
created and the diagnostic species of the formally
defined associations were determined using the
Table I. Species groups used for the Cocktail classification of the high-altitude arctic-alpine vegetation of the class Carici rupestris-Kobresietea
Ohba 1974 from the Western Carpathians.
Species group No. of releves Species (F6 100)
Androsace chamaejasme 41 Androsace chamaejasme (112.95), Hedysarum hedysaroides (104.27), Oxytropis halleri (90.78)
Carex firma 93 Carex firma (107.13), Dryas octopetala (103.45), Ranunculus alpestris (100.04)
Cerastium eriophorum 69 Cerastium eriophorum (114.52), Carex fuliginosa (109.84), Luzula spicata (102.05)
Doronicum stiriacum 138 Salix retusa agg. (101.90), Doronicum stiriacum (95.74), Pulsatilla scherfelii (90.06)
Draba siliquosa 12 Draba siliquosa (139.30), Plagiobryum demissum (98.89), Agrostis alpina (73.94)
Elyna myosuroides 10 Elyna myosuroides (117.79), Physconia muscigena (112.77), Dactylina madreporiformis (99.50)
Oreochloa disticha 451 Oreochloa disticha (126.08), Campanula alpina (120.97), Juncus trifidus (118.90), Festuca supina
(113.89)
Oxyria digyna 51 Oxyria digyna (113.70), Cardaminopsis neglecta (101.36), Saxifraga androsacea (98.13)
Oxytropis carpatica 24 Oxytropis carpatica (100.09), Helianthemum alpestre (85.29), Arenaria tenella (72.82)
Primula auricula 152 Primula auricula (111.24), Gentiana clusii (102.93), Trisetum alpestre (101.66)
Saxifraga wahlenbergii 26 Saxifraga wahlenbergii (116.29), Pritzelago alpina (76.86), Salix reticulata (75.23)
Silene acaulis 118 Lloydia serotina (123.22), Ligusticum mutellinoides (117.27), Minuartia sedoides (117.22), Silene
acaulis (111.75), Pedicularis oederi (101.21)
Vaccinium myrtillus 1611 Vaccinium myrtillus (119.89), Avenella flexuosa (99.35), Homogyne alpina (98.81), Vaccinium
vitis-idaea (90.29),
Species of each group are ranked by decreasing fidelity to the group, calculated as the phi coefficient in a stratified data set of 21,142 releves
of all vegetation types of Western Carpathians. The F values (multiplied by 100) of the species are given. The number of releves, in which the
group occurs, is the number of releves containing at least half of the group’s species.
Table II. Cocktail definitions of the associations from the class Carici rupestris-Kobresietea Ohba 1974.
Association Cocktail definition
Oxytropido-Elynion
1. Oxytropido carpaticae-
Elynetum
myosuroides
{5### Elyna myosuroides4AND(5### Silene acaulis4OR5### Androsace
chamaejasme4)}AND5Elyna myosuroidesUP054
2. Drabo siliquosae-Festucetum
versicoloris
(5Festuca versicolorUP254AND5### Draba siliquosa4)NOT5Silene acaulisUP124
3. Pyrolo carpaticae-Salicetum
reticulatae
{(5Salix reticulataUP054AND5### Saxifraga wahlenbergii4)AND5### Cerastium
eriophorum4}NOT(5### Oxyria digyna4OR5Carex rupestrisUP054)
4. Festucetum versicoloris {(5### Silene acaulis4AND5Festuca versicolorUP054)AND5### Androsace
chamaejasme4}NOT[{(5Dryas octopetalaUP054OR5Elyna myosuroidesUP054)OR(5Salix
reticulataUP124OR5Oreochloa distichaUP054)}OR(5Carex firmaUP054OR5### Draba
siliquosa4)]
5. Festuco versicoloris-
Oreochloetum distichae
[{(5Festuca supinaUP124OR5Oreochloa distichaUP204)AND5### Oreochloa
disticha4}AND5### Androsace chamaejasme4]NOT5Salix reticulataUP25>
Festucion versicoloris
6. Silenetum acaulis (5### Silene acaulis4AND5Silene acaulisUP054)NOT{((5Festuca
versicolorUP054OR5Oxyria digynaUP124)OR(5### Vaccinium myrtillus4OR5###
Androsace chamaejasme4))OR(5### Oxytropis carpatica4OR5### Saxifraga wahlenbergii4)}
7. Agrosti alpinae-Festucetum
versicoloris
(5### Oreochloa disticha4AND5Festuca versicolorUP054)NOT[((5### Androsace
chamaejasme4OR5Vaccinium vitis-idaeaUP054)OR(5### Saxifraga wahlenbergii4OR5###
Primula auricula4))OR(5Salix alpinaUP254OR5### Draba siliquosa4)]
8. Salicetum kitaibelianae [5Salix retusa agg.UP254OR{(5### Doronicum styriacum4AND5### Oreochloa
disticha4)AND5Salix retusa agg.UP054}]NOT{(5### Carex firma4OR5Festuca
versicolorUP12>)OR(5Juncus trifidusUP05>OR5### Androsace chamaejasme4)}
Classification of high-altitude arctic-alpine vegetation 3
Dow
nloa
ded
by [
Ivan
a Sv
itkov
a] a
t 06:
12 2
9 O
ctob
er 2
012

fidelity calculations in the JUICE software (Tichy
2002), using the final dataset of 422 releves assigned
to the association level by the created formal
definitions and FPFI calculations. Species with phi
values higher than 0.3 and Fisher’s exact test
significance higher than 0.001 (p5 0.001) were
designated as diagnostic. The similarity of synoptic
tables was also calculated by the JUICE software
(Tichy 2002). In the synoptic table, the diagnostic
and most frequent species of all the eight associations
of the class CK are shown.
The main gradients in floristic composition were
analysed by non-metric dimensional scaling (NMDS)
in the R program (R Development Core Team 2007)
via JUICE 7.0. Non-metric dimensional scaling was
used to analyse the matrix of dissimilarities between
samples and was performed on a matrix of Bray–
Curtis dissimilarities between releves. The NMDS was
visualised using the function ‘‘Ordispider’’ in the R
program ‘‘Vegan’’ package (Oksanen et al. 2009),
which draws spider diagrams with each point (releve)
connected to the cluster centroid by a line. A three-
dimensional ordination diagram with sites was drawn.
Statistica 8.0 (StatSoft Inc. 2006, http://www.stat
soft.com/) was used for the construction of box and
whisker plots comparing the individual groups
(vegetation units) using Ellenberg’s indicator values
(Ellenberg et al. 1992). Because the original Ellen-
berg’s indicator values do not list many species
occurring in Slovakia (especially Carpathian and
West Carpathian endemics and subendemics), the
indicator values for such species were added into the
dataset was following the study by Sibıkova et al.
(2010). The Tukey post-hoc test was used for
multiple comparisons following one-way analysis of
variance (ANOVA). Box and whisker plots were also
constructed for the frequency of arctic-alpine ele-
ments and endemics in relation to a particular
vegetation type and for the Shannon–Wiener index
of diversity and the number of vascular plants in
releve. Geographical elements were defined accord-
ing to Sibıkova et al. (2010). Spearman’s correlation
coefficient (non-parametric measure of correlation),
which assesses how well an arbitrary monotonic
function describes the relationship between two
variables, was used (Table III).
The nomenclature of higher syntaxa follows
Jarolımek and Sibık (2008) and Kliment et al.
(2010). The subspecies are marked by an asterisk.
Results
Thirteen species groups were defined in the stratified
dataset by means of the Cocktail method and,
together with the dominance criteria, used in formal
definitions of the associations of the class CK
(Table I). All eight associations from both alliances
of the class CK were satisfactorily reproduced by the
Cocktail definitions.
Some of the species groups are typical for individual
associations (e.g. Elyna myosuroides group for the
association Oxytropido carpaticae-Elynetum myosur-
oides or Draba siliquosa group for the association
Drabo siliquosae-Festucetum versicoloris), for individual
alliances (e.g. Androsace chamaejasme group for the
alliance Oxytropido-Elynion) or for the class Carici
rupestris-Kobresietea bellardii (e.g. Silene acaulis
group). Some species groups were typical for other
classes (e.g. Oreochloa disticha or Vaccinium myrtil-
lus groups) and were used in the definitions to
differentiate marginally overlapping habitats.
The created definitions were applied to a dataset
with 490 releves assigned by Petrık et al. (2006) or
the original authors to the class CK (at the
association, alliance or class level). Of them, based
on the criteria, 380 (77.6%) releves were assigned to
individual associations. Seven releves were assigned
to more than one vegetation unit. After the calcula-
tion of FPFI, six of them were assigned to only one
association. The definitions were also applied to the
unstratified dataset to see whether there are any
releves which would meet the conditions of the
definitions and which were previously classified
within some other higher vegetation units. In this
step, 18 releves from other higher vegetation units
fitted the created definitions and were assigned to
one of the eight associations of CK.
The results, however, matched the traditional
classification based on the cluster analyses. For the
remaining 104 unassigned releves, the FPFI index
was calculated following Ko�cı et al. (2003); accord-
ing to the results, 18 releves were assigned to
individual associations and 86 releves remained
unassigned according to our criteria.
The comparison of synoptic tables generated by
the application of the traditional, expert-based
classification and the classification based on the
Cocktail definitions indicated that their results
agreed at the level of 83.85%.
Table III. Correlations of individual variables in selected vegetation types. The non-parametric Spearman’s correlation test was used.
Significance level: p5 0.001.
Variable Arctic-Alpine Endemics Light Temperature Continentality Moisture Soil reaction Nutrients
Arctic-Alpine – 0.631885 0.633533 70.200291 0.704428 70.277126 0.752559 n.s.
Endemics 0.631885 – 0.386910 n.s. 0.532203 n.s 0.605137 0.264859
4 I. Svitkova and J. Sibık
Dow
nloa
ded
by [
Ivan
a Sv
itkov
a] a
t 06:
12 2
9 O
ctob
er 2
012

Dissimilarities between vegetation types are de-
picted in Figure 1, which combines the results of our
analysis and NMDS ordination. The main differ-
ences are shown along the first axis, where the releves
from the association Salicetum kitaibelianae are the
most remote in comparison with other communities
from the class CK. The Salicetum kitaibelianae seems
to be the most aberrant syntaxon of the studied
group. This community shows transitional charac-
teristics (floristic and ecological) of several high-rank
syntaxa, including the alliances Festucion versicoloris
and Loiseleurio-Vaccinion. It occupies scree habitats
below the steep rocky cliffs on mylonite or acidic
granite bedrock. The dominant species Salix kitai-
beliana represents microspecies from the broadly
defined group Salix retusa agg. (Dubravcova & Sibık
2006; Petrık et al. 2006; Sibık et al. 2006). According
to the positions of individual centroids (representing
the plant communities) within the first axes, we can
assume a correlation with the gradient of soil reaction
related to the characteristics of the substratum in the
habitats of individual associations, such as the
calcium and magnesium availability in the soil (cf.
Petrık et al. 2006).
Box plot diagrams (Figure 2) show the distribu-
tions of the occurrence of arctic-alpine and endemic
species as well as the richness of vascular plants and
the Shannon–Wiener index of biodiversity in relation
to individual vegetation types from the class CK. A
comparison of groups according to Ellenberg’s
indicator values is shown in box plot diagrams, too
(Figure 3). The significant differences between the
associations are depicted by the letters a–d.
To interpret the correlations between arctic-
alpine and endemic species with Ellenberg’s in-
dicator values, as an indirect assessment of environ-
mental factors in selected vegetation types, the non-
parametric Spearman’s correlation test was used
(Table III). The results show that the occurrence of
both the arctic-alpine and the endemic species
increased significantly with increasing light require-
ments, continentality and soil reaction; the occur-
rence of endemic species also correlates positively
with the amount of nutrients. There were significant
negative correlations between the occurrence of the
arctic-alpine species and both moisture and tem-
perature as a result of their occurrence in colder
habitats.
Oxytropido-Elynion Br.-Bl. (1948) 1949
Alpine xero-cryophilous wind-exposed swards and
carpets on calcium-rich soils. Contact vegetation:
Figure 1. Non-metric multi-dimensional scaling (NMDS) ordination diagram of vegetation plots from the class Carici rupestris-Kobresietea
(CK, see legend below). Each spider connects individual plots (releves) to the cluster centroid by a line. Note: Oxytropido-Elynion: 1
Oxytropido carpaticae-Elynetum myosuroides, 2 Drabo siliquosae-Festucetum versicoloris, 3 Pyrolo carpaticae-Salicetum reticulatae, 4 Festucetum
versicoloris, 5 Festuco versicoloris-Oreochloetum distichae; Festucion versicoloris: 6 Silenetum acaulis, 7 Agrosti alpinae-Festucetum versicoloris, 8
Salicetum kitaibelianae.
Classification of high-altitude arctic-alpine vegetation 5
Dow
nloa
ded
by [
Ivan
a Sv
itkov
a] a
t 06:
12 2
9 O
ctob
er 2
012

Potentillion caulescentis, Caricion firmae, Seslerion
tatrae. Distributional area: Belianske Tatry Mts, rare
Zapadne Tatry Mts.
1. Oxytropido carpaticae-Elynetum myosur-
oides (Puscaru et al. 1956) Coldea 1991
Formal definition: Table II.
Diagnostic species: see Supplementary material, col-
umn 1.
Brief characteristics: Open, species-rich plant com-
munity dominated by Elyna myosuroides and with
high frequencies of Festuca *versicolor and Silene
acaulis with relation to the phytocoenoses of the
rocky fissures of the alliance Potentillion caulescentis. It
occupies extremely windswept rocky ridges and
edges on quartzite, horn-stones and marls in the
alpine belt of the Belianske Tatry Mts.
Comparison: The releves traditionally assigned to this
association met the formal definition.
2. Drabo siliquosae-Festucetum versicoloris
Petrık in Petrık et al. 2006
Formal definition: Table II.
Diagnostic species: see Supplementary material, col-
umn 2.
Brief characteristics: Dense, species-rich, mat-forming
plant community occurring on the terraces on
convex parts of the steep slopes on calcium-rich
shale and horn-stones in the alpine belt of the
Belianske Tatry Mts with a close relation to some
phytocoenoses of the alliance Seslerion tatrae.
Comparison: The releves traditionally assigned to this
association met the formal definition.
3. Pyrolo carpaticae-Salicetum reticulatae Pet-
rık in Petrık et al. 2006
Formal definition: Table II.
Diagnostic species: see Supplementary material, col-
umn 3.
Figure 2. Box plot diagrams of the distribution of arctic-alpine and endemic species, number of vascular plants and Shannon-Wiener
diversity index in relation to individual associations. Significant differences between the associations are depicted by the letters a–d, and these
differences were tested by Tukey’s post-hoc test (p50.05). The box length is the interquartile range. The square inside the boxes indicates
the median. For legend for associations see Figure 1.
6 I. Svitkova and J. Sibık
Dow
nloa
ded
by [
Ivan
a Sv
itkov
a] a
t 06:
12 2
9 O
ctob
er 2
012

Brief characteristics: Dense, species-rich plant com-
munity with the typical character of arctic-alpine
tundra seen in the low carpets of Salix reticulata and
Dryas octopetala. The stands occur on windward
marly (or less often calcareous) slopes with relatively
long snow cover in the ridge parts of the Belianske
Tatry Mts. The stands have close relations to the
association Arenario tenellae-Caricetum firmae (alli-
ance Caricion firmae).
Comparison: The formal definition did not include a
small group of releves with high frequency or
dominancy of Carex firma. These releves might have
closer relations to the stands from the alliance
Caricion firmae.
Figure 3. Box and whisker plots of the Ellenberg’s indicator values in relation to individual associations. Letters a–d indicate homogenous
groups according to post-hoc comparisons using ANOVA (Tukey test, p50.05). The box length is the interquartile range. The square
inside the boxes indicates the median. For legend for associations see Figure 1.
Classification of high-altitude arctic-alpine vegetation 7
Dow
nloa
ded
by [
Ivan
a Sv
itkov
a] a
t 06:
12 2
9 O
ctob
er 2
012

4. Festucetum versicoloris Domin 1929
Formal definition: Table II.
Diagnostic species: see Supplementary material, col-
umn 4.
Brief characteristics: Species-rich, cushion- and mat-
forming plant community dominated by Festuca
*versicolor and Silene acaulis. At lower altitudes and
on more leeward slopes, it is closely related to the
phytocoenoses of the alliance Seslerion tatrae (Seslerio
tatrae-Festucetum versicoloris). It occupies the rocky
slopes on calciphilous slates, horn-stones and marls
in the alpine belt of the Belianske Tatry Mts.
Comparison: The formal definition did not include a
small group of releves of dense phytocoenoses with
the occurrence of acidophilous species, such as
Luzula alpinopilosa or Doronicum stiriacum, or the
high frequency of Cetraria islandica previously as-
signed to subass. caricetosum atratae Petrık et al. 2006.
These releves have relatively close relations to the
association Festuco versicoloris-Oreochloetum distichae
(cf. Petrık et al. 2006; Sibık et al. 2007), but they do
not meet the definition of this association either.
5. Festuco versicoloris-Oreochloetum distichae
Pawłowski et Stecki 1927 corr. Petrık et al.
2006 nom. invers. propos.
Formal definition: Table II.
Diagnostic species: see Supplementary material, col-
umn 5.
Brief characteristics: Species-rich, cushion- and mat-
forming plant community dominated by Festuca
supina and Silene acaulis, with a high frequency of
Oreochloa disticha and the occurrence of acidophilous
species. It has close relations to the plant commu-
nities of the alliance Juncion trifidi. The stands occur
on windward flat ridges on marls in the alpine belt of
the Belianske Tatry Mts and Zapadne Tatry –
Cervene vrchy Mts and represent typical arctic-
alpine tundra (Petrık & Sibık 2010).
Comparison: The releves traditionally assigned to this
association met the formal definition except for very
few releves lacking plant species from the Androsace
chamaejasme group.
Festucion versicoloris Krajina 1933
Alpine xero-cryophilous tussocky, cushion-like and
dwarf-shrub vegetation on siliceous-neutral soils on
mylonites or other type of metamorphic bedrock.
Contact vegetation: Androsacion alpinae, Juncion trifi-
di. Distributional area: Mylonite zones of the Tatra
Mts.
6. Silenetum acaulis Krajina 1933
Formal definition: Table II.
Diagnostic species: see Supplementary material, col-
umn 6.
Brief characteristics: Cushion-forming plant commu-
nity dominated by Silene acaulis and Saxifraga species
with a relation to scree phytocoenoses from the
alliance Androsacion alpinae. The stands of this
community occur on extreme south-facing mylonite
screes in the subnivale belt of the Vysoke and
Zapadne Tatry Mts or on north-facing mylonite
screes in the alpine belt.
Comparison: The formal definition did not include a
group of releves from stable soil substrates with
occurrence of species such as Agrostis alpina, Agrostis
rupestris or Carex *silicicola, which were previously
assigned to the subass. agrostietum alpinae (Krajina
1933) Dubravcova et Sibık 2006. These releves have
relatively close relations to the association Agrosti
alpinae-Festucetum versicoloris (cf. Dubravcova and
Sibık 2006, Sibık et al. 2007), but they do not meet
the definition of this association either.
7. Agrosti alpinae-Festucetum versicoloris
Pawłowski in Pawłowski et al. 1928 nom.
invers. propos.
Formal definition: Table II.
Diagnostic species: see Supplementary material, col-
umn 7.
Brief characteristics: Tussocks forming open plant
community dominated by Festuca *versicolor of very
steep rocky slopes and ridges on mylonite screes in
the alpine belt of the Vysoke and less often Zapadne
Tatry Mts.
Comparison: The formal definition corresponds well
with the traditional classification.
8. Salicetum kitaibelianae Krajina 1933
Formal definition: Table II.
Diagnostic species: see Supplementary material, col-
umn 8.
Brief characteristics: Relatively heterogeneous scree
plant community dominated by Salix kitaibeliana (S.
retusa agg.) of the alpine belt of the Vysoke and
Zapadne Tatry Mts. The stands occur on mylonite
and granite screes below the steep rocky slopes.
Comparison: The formal definition corresponds well
with the traditional classification.
Discussion
Comparison of two classifications
Ko�cı et al. (2003) and Role�cek (2007) tested the
created definitions in smaller datasets, which in-
cluded only the releves assigned by the original
authors to the studied vegetation unit (class).
8 I. Svitkova and J. Sibık
Dow
nloa
ded
by [
Ivan
a Sv
itkov
a] a
t 06:
12 2
9 O
ctob
er 2
012

However, there are many overlapping biotopes where
phytocoenoses from different classes are in contact.
It may be that the stands on one side of a gradient
being classified, e.g. to the alliance Oxytropido-
Elynion (CK), are on the other side of the gradient
similar to the plant communities of the alliance
Caricion firmae (Elyno-Seslerietea – ES). Sometimes
the releve assigned to an alliance as a marginal releve
or as a successional stage of the community of the
first alliance meets the definitional criteria of the
plant community from the second alliance (and
class) and vice versa. Hence, we did not test the
created definitions in a smaller dataset of the class
CK but in the large, unstratified dataset comprised of
releves from all vegetation units. The calculation of
FPFI coefficients for the releves assigned to more
than one vegetation unit, and for unassigned releves
previously (i.e. by prior authors) classified within the
class CK, was on the other hand performed in the
smaller dataset.
Several authors have tried to test whether they can
successfully use the Cocktail method to either create
formal definitions to classify or revise studied
vegetation units (Lososova 2004; Hajkova et al.
2006; Havlova 2006) or apply the created formal
definitions in an attempt to reproduce the traditional
classifications (Ko�cı et al. 2003; Dıte et al. 2007;
Role�cek 2007). The null hypothesis of our analyses
was that the results of applying the formal definitions
created on the basis of a large, geographically
stratified dataset including the occurrence of all
vegetation types of Slovakia would not differ from the
results of the traditional classification. As a case
study, the specific species-rich high-altitude vegeta-
tion with the occurrence of many arctic-alpine and
endemic species (Figure 2), was chosen. The
distinctiveness (and the presence) of the class CK
in the Western Carpathians was rejected by some
authors from other regions, especially from Oro-
Mediterranean (cf. Oriolo 2001; Peyre & Font
2011). The fact is that for the class CK, it is very
difficult to distinguish the characteristic species s. str.
They are usually circumpolar distributed arctic-
alpine species which naturally occur also in contact
phytocoenoses. The difference is that their distribu-
tional optimum covers extreme habitats, which
served as refugia for these species during the
postglacial periods. Therefore, we evaluate the
characteristic taxa in broader context, not only from
regional point of view (which often means marginal),
but taking into account the overall distributional
range of the plant communities of the class CK,
where these species have their natural optimum (cf.
Ohba 1974). On the other hand, like some other
studies (Coldea 1997; Sibık et al. 2007; Sibıkova
et al. 2010; Lancioni et al. 2011), this study supports
its delimitation.
It was possible to satisfactorily reproduce the
traditional classification of the class CK by means
of formal classification based on the formal defini-
tions created by the Cocktail method together with
the FPFI affiliation of the unclassified releves. We
were able to formulate the unequivocal assignment
criteria for all eight associations of both alliances of
the class CK. The formal delimitations agree very
well with the traditional ones. The positive criteria
species groups used in the formal definition contain
many species previously used as diagnostic (char-
acteristic, differential or constant) for individual
associations (Table I; see also Supplementary mate-
rial). Of the species used in the positive criteria
species groups, 32% were arctic-alpine taxa and 8%
were endemic, demonstrating the relic character of
the stands.
The statistical comparison of both synoptic tables
of associations – the first created from the dataset of
464 releves with the original assignment based on the
traditional expert classification using the cluster
analyses (Petrık et al. 2006, Kliment et al. 2010)
and the second (see Supplementary material) based
on formal classification – shows that we can accept
the null hypotheses.
Formalised classification – how reliable is the expert
system?
Several projects have recently been undertaken in
Slovakia to study meadows and pastures. One of
these studies yielded an expert system for grassland
vegetation (Janisova et al. 2007). Hence, we aimed to
continue creating new species groups and definitions
based on the existing expert system and to develop
the existing database. But the dataset has been
improved, up to the present, mainly with the
addition of new high-altitude releves. Therefore,
even after the geographical stratification, the new
dataset was considerably larger (21,142 releves vs.
the former 16,640). As we realised, the creation of
the species groups is strongly database dependent,
and after improving the dataset and adding a large
number of new releves, it is not possible to apply the
results from previous studies or from an expert
system based on a different dataset. Whereas the
results of the cluster or divisive classifications are
more dependent on the analysed dataset and there
are possible changes in the results after adding even a
small number of releves, the differences in the
process of creating the species groups used to define
the associations by the Cocktail method are evident
only after substantial modification of the dataset. In
our case, an increase in releves of 20% in the
stratified dataset, mainly as a result of adding the
high-altitude vegetation, affected the structures of
previously defined species groups. It is clear that due
Classification of high-altitude arctic-alpine vegetation 9
Dow
nloa
ded
by [
Ivan
a Sv
itkov
a] a
t 06:
12 2
9 O
ctob
er 2
012

to the increase in the number of releves of various
vegetation types, which were insufficiently included
in the former dataset; the representativeness of the
new dataset and hence the results obtained might be
considered as more objective. Based on these results,
we propose the occasional testing of the defined
species groups and formal definitions in improved,
more representative datasets, assuming that, e.g.
more data on the scarce vegetation types will be
included or the amount of data from sparsely
covered areas will increase. No classification of the
vegetation should be considered as definitive if all of
the vegetation types and the whole studied region are
not covered sufficiently and representatively.
Role�cek (2007) also pointed out some limitations
caused by the rules of using the species groups – in
particular, it might be difficult to define a vegetation
unit because suitable diagnostic species are already
included in other species groups used for different
vegetation types. This issue makes it more difficult to
continue creating the expert system (even using the
same dataset) after one part of the system for one
vegetation group has been finished. We suggest that
the strict ‘‘rule of priority’’ of previously used species
in particular species groups should be replaced by a
‘‘rule of consensus’’ and that the inclusion of
particular species into a species group should be
considered by experts on related (from the syneco-
logical or syndynamical point of view) vegetation
types.
When creating the species groups, several other
problems were encountered. They can be divided
into four main cases. In the first case, species that
belonged to previously defined species groups for
grassland vegetation (Janisova et al. 2007) were not
coherent in our dataset, i.e. particular species groups
were not observed and new groups were needed. In
the second case, after adding new species, the
broader species group was divided into two new
species groups, which had higher positive fidelity
values in the dataset than the former group did. In
the third case, almost identical species groups arose
with a difference in one species; the new group again
had higher positive fidelity values than the previous
one. The last case covers such species, which we did
not involve in any of the species groups due to its
weak diagnostic affinity, but these species were
previously used to form a species group – e.g. Sesleria
albicans. Janisova et al. (2007) used this species to
create a xero-thermophilous species group (Sesleria
albicans group: S. albicans, Genista pilosa, Leontodon
incanus) to define the xero-thermophilous grasslands.
S. albicans, however, has a wide vertical distributional
area and is typical also of high-altitude calcareous
grasslands. In subalpine belt, it often occurs with L.
incanus as a diagnostic species of several high-altitude
vegetation units (Kliment et al. 2007).
The question is to what extent we can rely on the
expert system (or use this method as a classification
tool) in cases when we expect that a large number of
releves will be added to the database in the near
future (i.e. with a horizon of 5 years), making the
dataset significantly larger and different. After every
significant extension of the database (especially from
few vegetation types), there is a significant chance
that it will not be possible to use the previously
generated species groups created from a smaller
dataset, and hence, the Cocktail definitions will no
longer be applicable. The authors are aware that this
new method and the formal definitions of vegetation
types do not have to be considered as final because
the ecological and geographical representativeness
will probably improve over time (Role�cek 2007), but
this method is, together with other classification
methods, a highly useful and complementary tool to
obtain objective results.
According to Mucina, even objective methods
need not yield objective results (Mucina, in verb., 3
May 2008, 17th International Workshop ‘‘European
Vegetation Survey’’, Brno). It would be interesting to
compare the results of several scientists, working
independently, with comparable knowledge back-
grounds in terms of how different the results would
be after processing the same dataset. The team of
experts that is behind the formation of species groups
and the fact that they are the experts on every
vegetation type are important to prevent the possi-
bility of ‘‘stealing’’ a species for a particular species
group or of incorrect inclusion of any species in a
suboptimal species group. However, there might not
have been full agreement even when using the
methods of numerical classification. Experts on
individual vegetation types could obtain similar (or
identical) results by means of numerical classification
(e.g. cluster analysis) when performing subsequent
partial and subjectively chosen analyses; hence, they
could create a specific ‘‘expert system’’, but this
system would be more difficult to replicate or define
by another person without specific (expert) knowl-
edge.
The expert system is also dependent on the quality
of the entries in the dataset. Sibıkova et al. (2009)
indicated that the SVD should be improved in terms
of the completeness and quality of its data. The
missing entries in particular fields, such as the
coordinates, are most significant. Both missing
coordinates and inaccurately or incompletely deter-
mined post-hoc coordinates have strong impacts on
the state (quality) of a geographically stratified
dataset and on its subsequent processing. The
releves from the regions where many vegetation
types co-occur on relatively small areas with rugged
relief that causes substantial changes in the ecological
conditions of the habitats are especially sensitive to
10 I. Svitkova and J. Sibık
Dow
nloa
ded
by [
Ivan
a Sv
itkov
a] a
t 06:
12 2
9 O
ctob
er 2
012

the accuracy of post-hoc determination of the
coordinates. The high-altitude mountain regions
are a good example of such habitats. If the
coordinates were determined inaccurately (e.g. as a
hypothetical centre of a valley in a mountain range),
they are useful only for a rough draft of the
distribution of individual vegetation units on, e.g.
the national level, but they are not suitable for
objective geographical stratification of the dataset.
The stratification of the releves from one vegetation
unit and one area (e.g. mountain valley) with such
inaccurately determined post-hoc coordinates leads
to the loss of information about variability of that
vegetation unit; hence, the created species groups
were not generally valid and were strongly dependent
on the quality of the database. Currently, Senko and
Sibık (in prep.) are addressing the precise post-hoc
determination of old phytosociological releves.
Another issue concerns the objective way to create
the definitions. By means of logical operators
together with the dominance threshold criteria, it is
possible to create a more or less complicated formal
definition for almost every vegetation unit or
assembly of releves. The questions are to what extent
the definitions would be applicable in praxis and to
what extent they would be database specific. Role�cek
(2007) believes that there is more to lose than to gain
from complicated definitions because they are
difficult to comprehend and usually strongly data-
base specific. But when the aim is to cover not only
the narrowly comprehended (basic) core of the
vegetation unit but also its whole content, i.e. to
cover the whole ecological scale of the unit including
the subunits and not only the releves of nominative
subassociation, it is often difficult to create a simple
definition, such as A AND B or A NOT B. This is
the case of vegetation units that cover more diverse
habitats and ecological conditions, including the
mountain ecosystems. As Dıte et al. (2007) indi-
cated, it is possible to simplify the formal definitions
by omitting the negative members of the definition.
This is also applicable to cases when the definitions
are created or tested in the unstratified dataset
containing only the releves from the specific well
determined vegetation units or in cases when only
the releves from one class (that does not include
transitional vegetation units) are analysed.
Role�cek (2007) raised two important issues – if the
large dataset of a large spatial and temporal scale is
really representative and if it is suitable for specific
purposes (classification) in a particular time and
area. In our opinion, the application of any classifica-
tion method, even the most objective one, no matter
how large the dataset may be, reflects to a large
extent the current state (i.e. the quality) of the
dataset (database) and the professionalism (i.e. the
level of expertise) of the botanist processing the data.
As Chytry (2000) pointed out in his crucial article
about formalised approaches to classification, the
imperfectly formalised approaches (e.g. cluster ana-
lysis) are essential in fine-scale classifications at the
regional and landscape levels, whereas the formalised
approach is superior for large-scale vegetation
surveys. The two methods are hence complementary
and, in general, it is not viable to put one above the
other. On the contrary, it is important to consider the
presumptive benefits of each of the approaches. The
feasibility of the formal classification is strongly
dependent on the quality and representativeness of
the database created for the area where the formal
definitions are to be valid. Hence, these methods are
widely used only in a few countries, but their rapid
spread is expected in the near future, along with the
development of high-quality national and supra-
national databases.
Acknowledgements
We are indebted to American Journal Experts
(www.journalexperts.com) for English proofreading.
This work was supported by the grant agency VEGA,
grant no. 2/0121/09 and 2/0090/12.
References
Boublık K. 2010. Formalized classification of the vegetation of
Abies alba-dominated forests in the Czech Republic. Biologia
65/5: 822–831.
Boublık K, Petrık P, Sadlo J, Hedl R, Willner W, Cerny T, et al.
2007. Calcicolous beech forests and related vegetation in the
Czech Republic: A comparison of formalized classifications.
Preslia 79: 141–161.
Bruelheide H. 1995. Die Grunlandgesellschaften des Harzes und
ihre Standortsbedingungen. Mit einem Beitrag zum Glieder-
ungsprinzip auf der Basis von statistisch ermittelten Arten-
gruppen. Diss Bot 244: 1–338.
Bruelheide H. 1997. Using formal logic to classify vegetation.
Folia Geobot Phytotax 32: 41–46.
Bruelheide H. 2000. A new measure of fidelity and its application
to defining species groups. J Veg Sci 11: 167–178.
Bruelheide H, Chytry M. 2000. Towards unification of national
vegetation classifications: A comparison of two methods for
analysis of large data sets. J Veg Sci 11: 295–306.
Carni A, Juvan N, Kosir P, Marinsek A, Pausi�c A, Silc U. 2011.
Plant communities in gradients. Plant Biosyst 145: 54–64.
Chytry M. 2000. Formalizovane prıstupy k fytocenologicke
klasifikaci vegetace. Preslia 72: 1–29.
Chytry M, editor. 2007. Vegetace Ceske republiky 1. Travinna a
kerı�ckova vegetace (Vegetation of the Czech Republic 1.
Grassland and heathland vegetation). Praha: Academia. 528 p.
Chytry M, editor. 2009. Vegetace Ceske republiky 2. Ruderalnı,
plevelova, skalnı a sut’ova vegetace (Vegetation of the Czech
Republic 1. Ruderal, weed, rock and scree vegetation). Praha:
Academia. 524 p.
Chytry M, Tichy L. 2003. Diagnostic, constant and dominant
species of vegetation classes and alliances of the Czech
Republic: A statistical revision. Folia Facultatis Scientiarum
Naturalium Universitatis Masarykianae Brunensis. Biologia
108: 1–231.
Classification of high-altitude arctic-alpine vegetation 11
Dow
nloa
ded
by [
Ivan
a Sv
itkov
a] a
t 06:
12 2
9 O
ctob
er 2
012

Coldea G. 1997. Carici rupestris-Kobresietea bellardii Ohba 1974. In:
Coldea G, editor. Les associations vegetales de Roumanie. 1.
Les associations herbacees naturelles. Presses Universitaires de
Cluj, pp. 202–207.
Dıte D. Hajek M, Hajkova P. 2007. Formal definitions of
Slovakian mire plant associations and their application in
regional research. Biologia 62: 400–408.
Dubravkova D, Chytry M, Willner W, Illyes E, Janisova M,
Kallayne SJ. 2010. Dry grasslands in the Western Carpathians
and the northern Pannonian Basin: A numerical classification.
Preslia 82: 165–221.
Dubravcova Z, Sibık J. 2006. K variabilite spolo�censtiev zvazu
Festucion versicoloris (trieda Carici rupestris-Kobresietea). Bull
Slov Bot Spolo�cn 28: 223–238.
Ellenberg H, Weber HE, Dull R, Wirth W, Werner W, Paulißen
D. 1992. Zeigerwerte von Pflanzen in Mitteleuropa. Ed. 2.
Scripta Geobot 18: 1–258.
Grabherr G, Mucina L, editors. 1993. Die Pflanzengesellschaften
Osterreichs 2. Jena: Gustav Fischer Verlag. 523 p.
Hajkova P, Hajek M, Apostolova I. 2006. Diversity of wetland
vegetation in the Bulgarian high mountains, main gradients
and context-dependence of the pH role. Plant Ecol 184: 111–
130.
Havlova M. 2006. Syntaxonomical revision of the Molinion
meadows in the Czech Republic. Preslia 78: 87–101.
Hrivnak R, Hajek M, Blanar D, Kochjarova J, Hajkova P. 2008.
Mire vegetation of the Muranska Planina Mts – formalised
classification, ecology, main environmental gradient
and influence of geographical position. Biologia 63/3: 368–
377.
Janisova M, Hajkova P, Hegedusova K, Hrivnak R, Kliment J,
Michalkova D, et al. 2007. Travinnobylinna vegetacia Slo-
venska – elektronicky expertny system na identifikaciu syntax-
onov. Bratislava: Botanicky ustav SAV. 263 p.
Jarolımek I, Sibık J, editors. 2008. Diagnostic, constant and
dominant species of the higher vegetation units of Slovakia.
Bratislava: Veda. 332 p.
Jarolımek I, Zaliberova M, Mucina L, Mochnacky S. 1997.
Rastlinne spolo�censtva Slovenska. 2. Synantropna vegetacia.
Bratislava: Veda. 420 p.
Kliment J, Bernatova D, Jarolımek I, Petrık A, Sibık J, Uhlırova J.
2007. Elyno-Seslerietea Br.-Bl. 1948. In: Kliment J, Valachovi�c
M, editors. Rastlinne spolo�censtva Slovenska. 4. Vysokohorska
vegetacia. Bratislava: Veda. p. 147–208.
Kliment J, Sibık J, Sibıkova I, Jarolımek I, Dubravcova Z,
Uhlırova J. 2010. High-altitude vegetation of the Western
Carpathians – A syntaxonomical review. Biologia 65/6: 965–
989.
Kliment J, Valachovi�c M, editors. 2007. Rastlinne spolo�censtva
Slovenska. 4. Vysokohorska vegetacia. Bratislava: Veda.
388 p.
Knollova I, Chytry M, Tichy L, Hajek O. 2005. Stratified
resampling of phytosociological databases: some strategies for
obtaining more representative data sets for classification
studies. J Veg Sci 16: 479–586.
Ko�cı M, Chytry M, Tichy L. 2003. Formalized reproduction of an
expert-based phytosociological classification: A case study of
subalpine tall-forb vegetation. J Veg Sci 14: 601–610.
Lancioni A, Facchi J, Taffetani F. 2011. Syntaxonomical analysis
of the Kobresio myosuroidis-Seslerietea caeruleae and Carici
rupestris-Kobresietea bellardii classes in the central southern
Apennines. Fitosociologia 48/1: 3–21.
Lososova Z. 2004. Weed vegetation in southern Moravia (Czech
Republic): A formalized phytosociological classification. Pre-
slia 76: 65–85.
Marhold K, Hindak F, editors. 1998. Zoznam nizsıch a vyssıch
rastlın Slovenska. (Checklist of non-vascular and vascular
plants of Slovakia). Bratislava: Veda. 687 p.
Michl T, Dengler J, Huck S. 2010. Montane-subalpine tall-herb
vegetation (Mulgedio-Aconitetea) in central Europe: Large-scale
synthesis and comparison with northern Europe. Phytocoeno-
logia 40: 117–154.
Mucina L, van der Maarel E. 1989. Twenty years of numerical
syntaxonomy. Vegetatio 81: 1–15.
Ohba T. 1974. Vergleichende Studien uber die alpine Vegetation
Japans. 1. Carici rupestris-Kobresietea bellardii. Phytocoenologia
1/3: 331–401.
Oksanen J, Kindt R, Legendre P, O’Hara B, Simpson GL,
Solymos P, Stevens MHH, Wagner H. 2009. Vegan: Com-
munity Ecology Package. R package version 1.15–4. Available:
http://cran.r-project.org/web/packages/vegan/index.html. Ac-
cessed Nov 2010, 22.
Oriolo G. 2001. Naked rush swards (Oxytropido-Elynion Br.-Bl.
1949) on the Alps and the Apennines and their syntaxonomical
position. Fitosociologia 38: 91–101.
Petrık A, Dubravcova Z, Jarolımek I, Kliment J, Sibık J, Valachovi�cM. 2006. Syntaxonomy and ecology of plant communities of
the Carici rupestris-Kobresietea bellardii in the Western Car-
pathians. Biologia 61/4: 393–412.
Petrık A, Sibık J. 2010. Asociacia Festuco versicoloris-Oreochloetum
distichae – vysokohorska tundra v Belianskych Tatrach. Naturae
Tutela 14/2: 147–154.
Peyre G, Font X. 2011. Syntaxonomic revision and floristic
characterization of the phytosociological alliances correspond-
ing to subalpine and alpine grasslands of the Pyrenees and
Cantabrian Mountains (classes Caricetea curvulae, Carici-
Kobresietea and Festuco Seslerietea). Plant Biosyst 145: 220–232.
Podani J. 2005. Multivariate exploratory analysis of ordinal data in
ecology: Pitfalls, problems and solutions. J Veg Sci. 16: 497–510.
Podani J. 2006. Braun-Blanquet’s legacy and data analysis in
vegetation science. J Veg Sci 17/1: 113–117.
R Development Core Team. 2007. R: A language and environ-
ment for statistical computing. Vienna: R Foundation for
Statistical Computing. Available: http://www.R-project.org.
Accessed Nov 2010, 22.
Rodwell JS, editor. 1990. British plant communities. Volume 1.
Woodlands and scrub. Cambridge: Cambridge University
Press. 400 p.
Rodwell JS, editor. 1991. British plant communities. Volume 2.
Mires and heaths. Cambridge: Cambridge University Press.
640 p.
Rodwell JS, editor. 1992. British plant communities. Volume 3.
Grasslands and montane communities. Cambridge: Cam-
bridge University Press. 552 p.
Role�cek J. 2007. Formalized classification of thermophilous oak
forests in the Czech Republic: What brings the cocktail
method? Preslia 79: 1–21.
Rozbrojova Z, Hajek M, Hajek O. 2010. Vegetation diversity of
mesic meadows and pastures in the West Carpathians. Preslia
82: 307–332.
Schaminee JHJ, Hennekens SM, Chytry M, Rodwell JS. 2009.
Vegetation-plot data and databases in Europe: an overview.
Preslia 81: 173–185.
Schaminee JHJ, Janssen JAM, Hennekens SM, Ozinga WA. 2011.
Large vegetation databases and information systems: New
instruments for ecological research, nature conservation and
policy making. Plant Biosyst 145: 85–90.
Senko D, Sibık J. in prep. Expert identification of geographical
coordinates of older phytosociological releves using by GIS.
StatSoft Inc. 2006. Electronic statistics textbook. Tulsa: Statsoft.
Available: http://www.statsoft.com/textbook/stahme.html. Ac-
cessed Nov 2010, 22.
Sibık J. 2012. Slovak vegetation database. Biodiversity & Ecology
4: 429–429. Special volume: In: Dengler J, Oldeland J, Jansen
F, Chytry M, Ewald J, Finckh M, et al., editors. Vegetation
databases for the 21st century.
12 I. Svitkova and J. Sibık
Dow
nloa
ded
by [
Ivan
a Sv
itkov
a] a
t 06:
12 2
9 O
ctob
er 2
012

Sibık J, Kliment J, Dubravcova Z, Belohlavkova R, Paclova L.
2006. Syntaxonomy and nomenclature of the alpine heaths
(the class Loiseleurio-Vaccinietea) in the Western Carpathians.
Hacquetia 5/1: 37–71.
Sibık J, Petrık A, Valachovi�c M, Dubravcova Z. 2007. Carici
rupestris-Kobresietea bellardii. In: Kliment J, Valachovi�c M,
editors. Rastlinne spolo�censtva Slovenska. 4. Vysokohorska
vegetacia. Bratislava: Veda. p. 209–249.
Sibık J, Sibıkova I, Kliment J. 2010. The subalpine Pinus mugo-
communities of the Carpathians with a European perspective.
Phytocoenologia 40/2–3: 155–188.
Sibıkova I, Sibık J, Jarolımek I, Kliment J. 2009. Current
knowledge and phytosociological data on the high-altitude
vegetation in the Western Carpathians – A review. Biologia 64/
2: 215–224.
Sibıkova I, Sibık J, Hajek M, Kliment J. 2010. The distribution of
arctic-alpine elements within high-altitude vegetation of the
Western Carpathians in relation to environmental factors, life
forms and phytogeography. Phytocoenologia 40/2–3: 189–203.
Silc U, Carni A. 2007. Formalized classification of the weed
vegetation of arable land in Slovenia. Preslia 79: 283–302.
Tichy L. 2002. JUICE, software for vegetation classification. J Veg
Sci 13: 451–453.
Tichy L. 2005. New similarity indices for the assignment of releves
to the vegetation units of an existing phytosociological
classification. Plant Ecol 179: 67–72.
Valachovi�c M, editor. 2001. Rastlinne spolo�censtva Slovenska. 3.
Vegetacia mokradı. Bratislava: Veda. 435 p.
Valachovi�c M, Otahelova H, Stanova V, Maglocky S. 1995.
Rastlinne spolo�censtva Slovenska 1. Pionierska vegetacia.
Bratislava: Veda. 185 p.
Supplementary material. Shortened synoptic table of the high-altitude arctic-alpine vegetation of the
class Carici rupestris-Kobresietea Ohba 1974 in the Western Carpathians.
Diagnostic taxa (F4 0.30) with the percentage values of their frequency are sorted according to their fidelity to
the vegetation type (phi koeficient6 100, upper index). The species with probability of random distribution in
the vegetation type higher than 0.001 yelded by Fisher’s exact test were excluded from the list of diagnostic
species. Explanations: dg¼Taxa characterized by Sibık et al. (2007) as diagnostic (differential, characteristic or
transgressive) for any syntaxa of the class Carici rupestris-Kobresietea; aa¼ arctic-alpine taxon according to
Sibıkova et al. (2010). Species printed in bold are characteristic species s.str. of the class Carici rupestris-
Kobresietea according to Kliment & Valachovi�c (2007).
Group No. 1 2 3 4 5 6 7 8
No. of releves 10 11 31 24 19 86 100 141
Oxytropido carpaticae-Elynetum myosuroides
aa Elyna myosuroides dg 10088.7 .— .— 8— 11— .— 4— .—
Physconia muscigena (E0) dg 8073.9 .— .— 2512.9 .— 1— 1— .—
Primula auricula dg 5065.0 .— .— 4— .— .— .— .—
Fulgensia bracteata (E0) dg 4060.7 .— .— .— .— .— .— .—
Leontopodium alpinum 4060.7 .— .— .— .— .— .— .—
Trisetum alpestre 7056.5 18— .— 3318.4 .— 2— .— 1—
Ochrolechia upsaliensis (E0) 4056.2 .— .— .— 5— .— .— .—
Psora decipiens (E0) 5054.7 .— .— 2115.9 .— .— .— .—
Squamarina gypsacea (E0) 4052.2 .— .— 8— .— 2— .— .—
Hypnum vaucheri (E0) 4049.7 .— .— 8— .— 2— 4— .—
Caloplaca ammiospila (E0) 4049.0 .— .— 12— .— 2— 1— .—
Mycobilimbia lobulata (E0) dg 5048.2 9— .— 2517.6 .— 1— .— .—
Cladonia pyxidata agg. (E0) dg 9047.6 27— 16— 12— 26— 38— 22— 19—
Distichium inclinatum (E0) 3046.1 .— 6— .— .— .— .— .—
Cetraria muricata (E0) 3044.6 .— 3— 4— .— .— 1— .—
Anaptychia bryorum (E0) 2042.4 .— .— .— .— .— .— .—
Campanula cochleariifolia 3040.1 .— .— 12— .— 2— .— .—
Lecanora epibryon (E0) 3039.4 .— .— 8— 5— 2— .— .—
Phaeorrhiza nimbosa (E0) 3036.6 .— .— 2122.4 .— .— .— .—
Oxytropis carpatica dg 5035.7 9— 19— 4227.0 5— .— .— .—
Myurella julacea (E0) 6034.2 5529.2 10— 5025.1 .— 3— 1— .—
Drabo siliquosae-Festucetum versicoloris
Draba siliquosa dg 20— 10088.2 .— 4— .— .— .— .—
Plagiothecium laetum (E0) dg .— 7383.2 .— .— .— .— .— 1—
Helianthemum grandiflorum dg .— 9178.3 .— 21— 11— .— 3— .—
Linum extraaxillare dg .— 7377.3 .— .— 5— .— 5— .—
Plagiobryum demissum (E0) dg 10— 8276.3 10— 4— .— .— .— 1—
Parnassia palustris dg .— 10063.5 16— 4213.3 37— 3— 7— 4—
Astragalus australis dg 30— 8263.3 10— 21— .— .— .— .—
(continued)
Classification of high-altitude arctic-alpine vegetation 13
Dow
nloa
ded
by [
Ivan
a Sv
itkov
a] a
t 06:
12 2
9 O
ctob
er 2
012
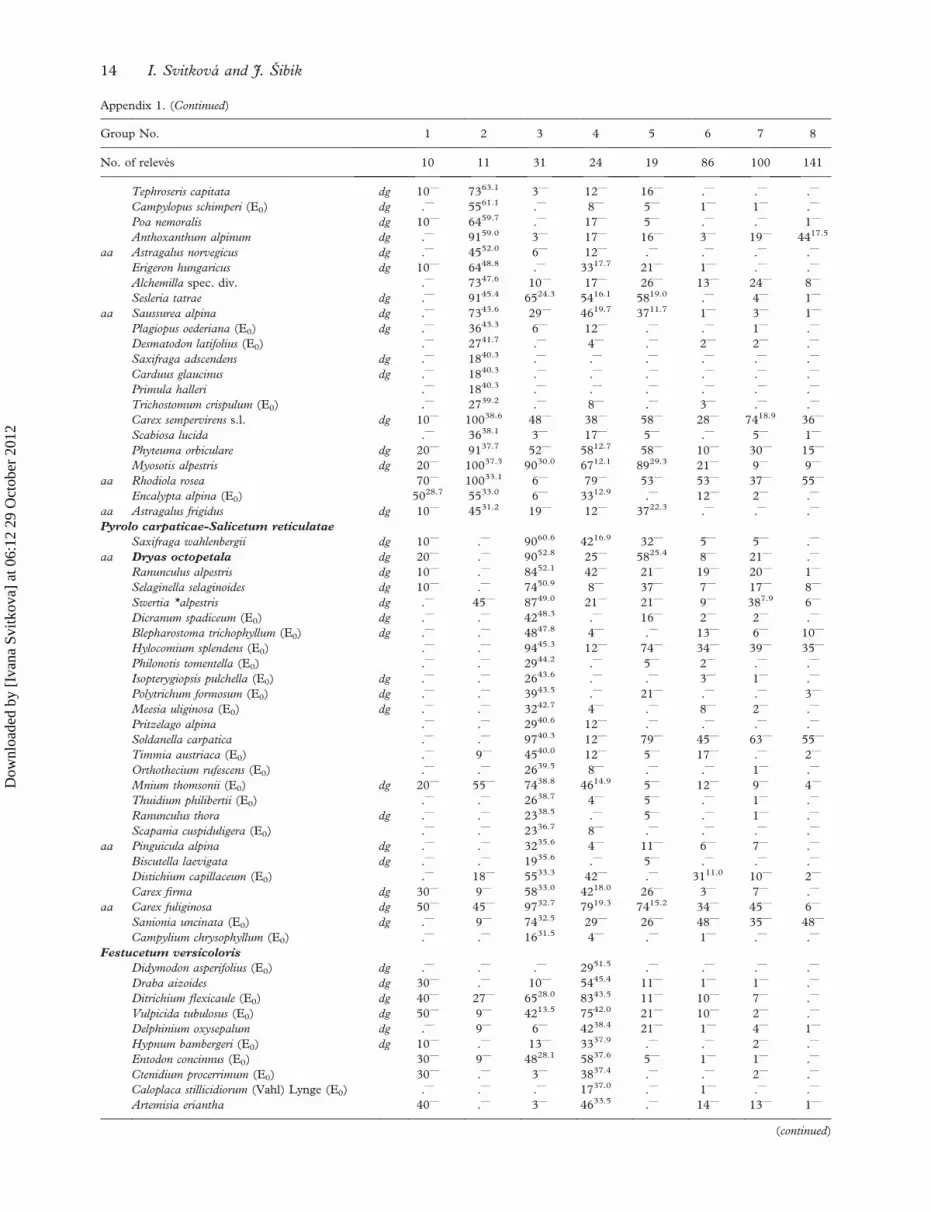
Appendix 1. (Continued)
Group No. 1 2 3 4 5 6 7 8
No. of releves 10 11 31 24 19 86 100 141
Tephroseris capitata dg 10— 7363.1 3— 12— 16— .— .— .—
Campylopus schimperi (E0) dg .— 5561.1 .— 8— 5— 1— 1— .—
Poa nemoralis dg 10— 6459.7 .— 17— 5— .— .— 1—
Anthoxanthum alpinum dg .— 9159.0 3— 17— 16— 3— 19— 4417.5
aa Astragalus norvegicus dg .— 4552.0 6— 12— .— .— .— .—
Erigeron hungaricus dg 10— 6448.8 .— 3317.7 21— 1— .— .—
Alchemilla spec. div. .— 7347.6 10— 17— 26— 13— 24— 8—
Sesleria tatrae dg .— 9145.4 6524.3 5416.1 5819.0 .— 4— 1—
aa Saussurea alpina dg .— 7343.6 29— 4619.7 3711.7 1— 3— 1—
Plagiopus oederiana (E0) dg .— 3643.3 6— 12— .— .— 1— .—
Desmatodon latifolius (E0) .— 2741.7 .— 4— .— 2— 2— .—
Saxifraga adscendens dg .— 1840.3 .— .— .— .— .— .—
Carduus glaucinus dg .— 1840.3 .— .— .— .— .— .—
Primula halleri .— 1840.3 .— .— .— .— .— .—
Trichostomum crispulum (E0) .— 2739.2 .— 8— .— 3— .— .—
Carex sempervirens s.l. dg 10— 10038.6 48— 38— 58— 28— 7418.9 36—
Scabiosa lucida .— 3638.1 3— 17— 5— .— 5— 1—
Phyteuma orbiculare dg 20— 9137.7 52— 5812.7 58— 10— 30— 15—
Myosotis alpestris dg 20— 10037.3 9030.0 6712.1 8929.3 21— 9— 9—
aa Rhodiola rosea 70— 10033.1 6— 79— 53— 53— 37— 55—
Encalypta alpina (E0) 5028.7 5533.0 6— 3312.9 .— 12— 2— .—
aa Astragalus frigidus dg 10— 4531.2 19— 12— 3722.3 .— .— .—
Pyrolo carpaticae-Salicetum reticulatae
Saxifraga wahlenbergii dg 10— .— 9060.6 4216.9 32— 5— 5— .—
aa Dryas octopetala dg 20— .— 9052.8 25— 5825.4 8— 21— .—
Ranunculus alpestris dg 10— .— 8452.1 42— 21— 19— 20— 1—
Selaginella selaginoides dg 10— .— 7450.9 8— 37— 7— 17— 8—
Swertia *alpestris dg .— 45— 8749.0 21— 21— 9— 387.9 6—
Dicranum spadiceum (E0) dg .— .— 4248.3 .— 16— 2— 2— .—
Blepharostoma trichophyllum (E0) dg .— .— 4847.8 4— .— 13— 6— 10—
Hylocomium splendens (E0) .— .— 9445.3 12— 74— 34— 39— 35—
Philonotis tomentella (E0) .— .— 2944.2 .— 5— 2— .— .—
Isopterygiopsis pulchella (E0) dg .— .— 2643.6 .— .— 3— 1— .—
Polytrichum formosum (E0) dg .— .— 3943.5 .— 21— .— .— 3—
Meesia uliginosa (E0) dg .— .— 3242.7 4— .— 8— 2— .—
Pritzelago alpina .— .— 2940.6 12— .— .— .— .—
Soldanella carpatica .— .— 9740.3 12— 79— 45— 63— 55—
Timmia austriaca (E0) .— 9— 4540.0 12— 5— 17— .— 2—
Orthothecium rufescens (E0) .— .— 2639.5 8— .— .— 1— .—
Mnium thomsonii (E0) dg 20— 55— 7438.8 4614.9 5— 12— 9— 4—
Thuidium philibertii (E0) .— .— 2638.7 4— 5— .— 1— .—
Ranunculus thora dg .— .— 2338.5 .— 5— .— 1— .—
Scapania cuspiduligera (E0) .— .— 2336.7 8— .— .— .— .—
aa Pinguicula alpina dg .— .— 3235.6 4— 11— 6— 7— .—
Biscutella laevigata dg .— .— 1935.6 .— 5— .— .— .—
Distichium capillaceum (E0) .— 18— 5533.3 42— .— 3111.0 10— 2—
Carex firma dg 30— 9— 5833.0 4218.0 26— 3— 7— .—
aa Carex fuliginosa dg 50— 45— 9732.7 7919.3 7415.2 34— 45— 6—
Sanionia uncinata (E0) dg .— 9— 7432.5 29— 26— 48— 35— 48—
Campylium chrysophyllum (E0) .— .— 1631.5 4— .— 1— .— .—
Festucetum versicoloris
Didymodon asperifolius (E0) dg .— .— .— 2951.5 .— .— .— .—
Draba aizoides dg 30— .— 10— 5445.4 11— 1— 1— .—
Ditrichium flexicaule (E0) dg 40— 27— 6528.0 8343.5 11— 10— 7— .—
Vulpicida tubulosus (E0) dg 50— 9— 4213.5 7542.0 21— 10— 2— .—
Delphinium oxysepalum dg .— 9— 6— 4238.4 21— 1— 4— 1—
Hypnum bambergeri (E0) dg 10— .— 13— 3337.9 .— .— 2— .—
Entodon concinnus (E0) 30— 9— 4828.1 5837.6 5— 1— 1— .—
Ctenidium procerrimum (E0) 30— .— 3— 3837.4 .— .— 2— .—
Caloplaca stillicidiorum (Vahl) Lynge (E0) .— .— .— 1737.0 .— 1— .— .—
Artemisia eriantha 40— .— 3— 4633.5 .— 14— 13— 1—
(continued)
14 I. Svitkova and J. Sibık
Dow
nloa
ded
by [
Ivan
a Sv
itkov
a] a
t 06:
12 2
9 O
ctob
er 2
012

Appendix 1. (Continued)
Group No. 1 2 3 4 5 6 7 8
No. of releves 10 11 31 24 19 86 100 141
aa Saxifraga oppositifolia dg 30— .— 13— 5832.3 .— 5226.8 21— 6—
Crepis jacquinii dg .— 18— 10— 2931.4 .— 2— .— .—
Poa alpina dg 50— 45— 7723.4 8831.0 6816.6 27— 14— 3—
Festuco versicoloris-Oreochloetum distichae
Senecio *carpathicus dg .— .— .— .— 4254.6 1— 3— 6—
Trollius altissimus .— .— 3— .— 3250.4 .— .— .—
Dianthus glacialis dg .— .— 26— 21— 6849.7 17— 9— 2—
aa Saxifraga hieraciifolia .— .— 4521.4 8— 7447.5 339.8 9— 6—
Ptilidium ciliare (E0) .— .— 4225.8 8— 6347.4 1— 8— 10—
Bistorta major dg .— 55— 19— 4— 7945.4 5— 25— 23—
Dicranum fuscescens (E0) .— .— 19— .— 4244.2 5— 4— 1—
aa Carex atrata agg. dg 10— 45— 5814.8 29— 9543.2 27— 34— 13—
aa Potentilla crantzii dg .— 45— 3— 12— 6341.6 14— 15— 3—
Luzula sudetica dg .— 45— 5225.4 17— 6840.4 .— 3— 1—
Hedysarum hedysaroides dg 30— 6418.0 7426.1 5813.9 8937.9 2— 1— 4—
Lophozia obtusa (E0) .— .— .— .— 1637.5 .— .— .—
Rhodax alpestris dg 50— 18— 6120.9 7128.5 7934.9 .— .— .—
Salix hastata dg .— .— .— .— 1634.8 .— 2— .—
Dicranum bonjeanii (E0) .— .— 3— .— 1633.3 .— .— .—
Botrychium lunaria .— .— .— 4— 1632.2 .— .— .—
Silenetum acaulis
Gentiana frigida dg .— .— 16— 12— .— 7151.3 36— 11—
Novosieversia reptans dg .— .— .— .— .— 3650.3 7— 1—
Saxifraga bryoides dg 10— .— 6— 8— 5— 5348.9 8— 3—
Cardaminopsis neglecta dg .— .— .— .— .— 4748.5 25— 3—
Leucanthemopsis alpina dg .— .— .— .— .— 3742.6 10— 13—
aa Oxyria digyna dg .— .— 3— 4— .— 3439.1 13— 3—
Pogonatum urnigerum (E0) dg .— .— 6— 12— 11— 4936.2 21— 19—
Saxifraga androsacea dg .— .— 32— .— 5— 4034.7 8— 2—
Solorina crocea (E0) .— .— .— .— .— 2334.4 6— 6—
Cetraria nivalis (E0) dg 10— .— 3— 21— 32— 5533.7 23— 12—
aa Erigeron uniflorus dg .— .— .— .— .— 3333.3 21— 13—
Poa laxa dg 10— .— .— .— .— 3031.1 20— 4—
aa Salix herbacea dg .— .— .— .— .— 2630.5 18— 6—
Agrosti alpinae-Festucetum versicoloris
Callianthemum coriandrifolium dg .— .— .— .— .— 7— 2436.1 4—
Anemone narcissiflora dg .— 36— 3— 4— 5— 6— 4330.5 18—
Salicetum kitaibelianae
Salix retusa agg. (incl. S. kitaibelliana) dg 10— .— 35— 8— 21— 31— 68— 10052.3
Vaccinium myrtillus dg .— .— 3— .— 5— .— 14— 3944.4
Pulsatilla scherfelii dg .— 9— .— .— 32— 9— 50— 6843.7
Gentiana punctata dg .— .— .— .— .— .— 4— 2340.7
Avenella flexuosa dg .— 9— .— .— .— .— 6— 2838.2
Doronicum stiriacum .— 9— 13— 12— 32— 59— 35— 7437.4
Valeriana tripteris dg .— .— .— .— .— .— .— 1637.3
Solidago *minuta dg .— 18— 3— .— .— .— 5— 3337.2
Oreogeum montanum dg .— .— .— .— .— 5— 2— 2135.8
Cladonia arbuscula s.l. (E0) .— .— .— .— 11— 16— 17— 3935.6
Ligusticum mutellina .— .— .— .— 5— 6— 28— 3534.0
Senecio *carniolicus .— .— .— .— .— 3— 1— 1733.5
Homogyne alpina .— .— 6— 4— .— 6— 12— 3033.5
Ranunculus pseudomontanus .— .— .— .— 5— 9— 23— 3332.5
Diagnostic taxa for more syntaxa
Dactylina madreporiformis (E0) dg 8055.1 9— 10— 6238.8 5— .— .— .—
Arenaria tenella dg 7046.8 18— 10— 5431.9 5— 5— .— .—
Oxytropis halleri dg 7033.9 27— 26— 7941.5 32— .— .— .—
aa Aster alpinus dg 5033.4 7356.3 .— 12— .— .— .— .—
Agrostis alpina dg 6032.0 8251.3 3— 21— .— 5— 20— 1—
Ranunculus breyninus dg 10031.0 10031.0 52— 9627.7 8922.8 12— 26— 4—
Bupleurum ranunculoides dg 5030.6 8261.5 .— 17— .— .— .— .—
Thymus pulcherrimus dg 10— 5538.6 3— 5438.2 11— .— .— .—
(continued)
Classification of high-altitude arctic-alpine vegetation 15
Dow
nloa
ded
by [
Ivan
a Sv
itkov
a] a
t 06:
12 2
9 O
ctob
er 2
012

Appendix 1. (Continued)
Group No. 1 2 3 4 5 6 7 8
No. of releves 10 11 31 24 19 86 100 141
Rhytidiadelphus triquetrus (E0) dg .— .— 9460.7 .— 6334.0 2— 15— 22—
Pyrola carpatica dg .— .— 7156.8 .— 5337.8 .— 2— 2—
aa Salix reticulata dg .— .— 10053.6 42— 9549.3 9— 16— 4—
Saxifraga aizoides dg 10— 18— 7445.8 6235.3 5— 3— 10— 1—
Minuartia pauciflora (Kit. ex Kanitz) Dvor. dg 30— 45— 10038.7 9232.4 63— 34— 20— 6—
Cardaminopsis halleri dg .— 27— 4831.2 8— 5335.5 .— 1— .—
aa Androsace chamaejasme dg 10031.4 9124.4 7714.0 10031.4 10031.4 2— 3— .—
Other taxa with higher fidelity or frequency
aa Cerastium eriophorum dg 9017.6 10025.7 9420.5 9622.3 9521.4 34— 35— 4—
Minuartia sedoides dg 80— 45— 9722.0 9621.2 10024.6 9117.0 37— 16—
Festuca versicolor dg 100— 100— 9716.7 10019.7 9514.8 29— 9918.7 10—
Galium anisophyllon dg 80— 10028.5 8718.4 9222.0 10028.5 9— 29— 13—
aa Pedicularis oederi dg 40— 100— 10023.7 9216.7 84— 60— 752.7 23—
Campanula tatrae 80— 100— 65— 9620.2 74— 41— 731.0 47—
aa Silene acaulis dg 90— 55— 10017.2 96— 95— 10017.2 896.1 39—
Oreochloa disticha dg 20— 45— 23— 25— 79— 77— 78— 72—
Primula minima 70— 18— 58— 58— 68— 8619.9 7914.5 44—
Viola biflora 20— 36— 10— 4— 26— 12— 3513.8 19—
aa Bistorta vivipara dg 100— 100— 100— 100— 95— 65— 78— 82—
Festuca supina dg 50— 45— 52— 58— 79— 9220.3 76— 80—
Saxifraga paniculata 80— 91— 8411.9 10025.1 63— 50— 51— 36—
Tortella tortuosa 80— 9125.0 719.7 9225.6 21— 53— 50— 9—
aa Bartsia alpina 20— 64— 74— 21— 21— 34— 56— 6717.3
aa Juncus trifidus .— 82— 23— 38— 84— 35— 7216.2 7215.9
aa Astragalus alpinus 10— 27— 3— 8— 11— .— .— .—
Bellidiastrum michelii .— 9— 16— 4— 11— 2— 6— 1—
Cerastium *glandulosum .— 18— 19— 8— 37— 13— 6— 6—
Saxifraga moschata dg 20— 100— 71— 79— 9523.2 8616.3 49— 25—
aa Luzula spicata dg 30— 73— 42— 58— 53— 7117.6 40— 15—
Saxifraga carpathica .— .— .— 4— .— 1019.6 5— .—
aa Gentiana nivalis dg 30— 3626.3 6— 2513.5 5— .— .— 1—
Coeloglossum viride dg .— .— 3229.6 .— 16— .— 11— 16—
Draba dubia 20— .— .— 8— .— 1— 3— .—
Gentianella lutescens 40— 18— .— 12— .— 6— 8— 5—
Geranium sylvaticum .— 18— .— .— .— .— 1— 5—
Aconitum firmum .— 18— 6— 4— 16— 1— 2— 6—
aa Pedicularis verticillata .— 73— 45— 29— 21— 57— 46— 51—
Hypnum cupressiforme 30— .— 10— 17— 11— 8— 12— 3—
Androsace lactea 20— .— .— 12— .— 1— .— .—
Euphrasia salisburgensis dg 5025.5 5529.7 6— 5429.3 5— 2— 2— 1—
Achillea *sudetica .— 18— 3— .— 5— .— 4— 1—
aa Carex atrofusca dg .— .— 1029.3 .— .— .— .— .—
Huperzia selago dg .— .— 23— 12— .— 53— 48— 46—
Thymus alpestris 50— 64— 13— 42— 21— 8— 29— 16—
Saussurea pygmaea dg 10— 9— .— 12— .— 17— 2821.4 8—
Campanula alpina 10— .— 48— 38— 84— 53— 8226.7 58—
Saxifraga retusa dg .— .— 3— .— .— 1923.3 1822.3 1—
Leontodon pseudotaraxaci dg 10— 27— 26— 29— 16— 6— 268.0 4—
Luzula luzuloides .— 27— .— .— 5— .— 4— 9—
Plantago atrata .— .— .— .— 5— .— .— .—
Sedum alpestre .— .— .— .— .— 13— 4— 12—
aa Chamorchis alpina 10— 9— 2320.0 8— 11— 2— 2— .—
Euphrasia tatrae .— 27— 3— .— 5— 2— 3— 3129.2
Hieracium alpinum .— .— 3— .— 32— 8— 27— 4029.2
Polytrichum alpinum .— .— .— .— .— 37— 39— 4128.3
Pyrola minor .— .— .— .— .— .— .— 927.4
Vaccinium gaultherioides .— .— 6— 4— .— 3— 12— 2326.2
Agrostis rupestris .— 9— .— .— 16— 5— 27— 3124.5
Salix silesiaca .— .— .— .— .— .— .— 623.7
Festuca picturata .— 45— .— 4— 5— .— 4— 2921.8
Luzula alpinopilosa .— 9— 23— 8— 68— 55— 36— 5821.0
(continued)
16 I. Svitkova and J. Sibık
Dow
nloa
ded
by [
Ivan
a Sv
itkov
a] a
t 06:
12 2
9 O
ctob
er 2
012

Appendix 1. (Continued)
Group No. 1 2 3 4 5 6 7 8
No. of releves 10 11 31 24 19 86 100 141
Veratrum *lobelianum .— .— .— .— .— .— 1— 620.1
Avenula versicolor dg 10— 9— 10— 17— 32— 6— 40— 4219.9
Vaccinium vitis-idaea dg .— .— 42— 4— 32— 12— 4822.8 4519.8
Potentilla aurea .— 9— 3— .— 32— .— 16— 2416.8
aa Lloydia serotina dg 90— 91— 878.9 9616.8 89— 78— 71— 16—
aa Ranunculus glacialis .— .— .— .— .— 1529.7 5— 1—
aa Ligusticum mutellinoides dg 70— 27— 8418.0 71— 9526.4 7410.7 48— 16—
aa Draba fladnizensis 10— 9— .— 2127.5 .— .— .— .—
Cardaminopsis arenosa agg. .— 36— 2311.6 3323.9 5— 1— 1— .—
Schistidium apocarpum agg. 40— .— 10— 3829.8 .— 3— 2— 3—
Cortusa matthioli .— 18— 3— .— 5— .— .— 1—
aa Antennaria carpatica dg 50— 9— 32— 46— 21— 15— 23— 21—
aa Carex capillaris dg 5024.0 5528.1 13— 4216.6 26— .— .— .—
aa Comastoma tenellum dg 6029.8 5525.1 19— 5424.7 16— .— 1— .—
aa Tofieldia pusilla dg .— .— 1029.3 .— .— .— .— .—
aa Carex rupestris dg .— .— 3— .— .— 2— 1529.7 .—
Other arctic-alpine taxa or rare taxa of West Carpathians’ flora
aa Kobresia simpliciuscula 20— .— .— 4— 5— 1— .— .—
Bellardiochloa variegata .— 9— .— .— .— .— .— .—
aa Arctous alpina dg .— .— 6— .— .— .— .— .—
Draba tomentosa .— .— .— 4— .— .— 1— .—
Pilosella alpicola .— .— .— .— .— .— 3— 4—
aa Arabis alpina .— .— .— .— .— .— 3— 1—
aa Dichodon cerastoides .— .— .— .— .— .— 1— 1—
Pulsatilla vernalis dg .— .— .— .— .— .— 2— .—
aa Veronica alpina .— .— .— .— .— 8— 1— 2—
Armeria alpina .— .— .— .— .— 1— 1— .—
Cerastium uniflorum .— .— .— .— .— 1— 1— 1—
aa Omalotheca supina .— .— .— .— .— 2— .— .—
Woodsia alpina .— .— .— .— .— 1— .— .—
Bryophytes and Lichens (E0)
Cetraria islandica 10— 27— 9025.4 50— 95— 65— 54— 65—
Thamnolia vermicularis 90— 9— 52— 8827.8 58— 6611.7 31— 13—
Rhytidium rugosum 30— 45— 52— 46— 58— 22— 36— 10—
Campylium stellatum 20— .— 5529.6 33— 5— 4419.9 15— 6—
Alectoria ochroleuca 30— .— 10— 8— 11— 34— 23— 26—
Ctenidium molluscum 20— 9— 19— 17— .— .— 7— .—
Plagiochila porelloides 20— 4525.7 4525.4 29— 5— .— 6— .—
Polytrichum alpinum 20— 18— 3514.3 3816.2 4220.5 1— 8— .—
Cladonia rangiferina .— .— 3— .— 53— 14— 21— 3822.8
Lecidoma demissum 10— 9— .— 2121.7 11— 1— 1— .—
Cratoneuron commutatum .— .— .— .— .— 622.6 .— .—
Pohlia cruda .— 36— 5229.4 .— .— 4220.2 20— 13—
Blindia acuta .— .— .— .— .— 1226.4 4— .—
Dicranum majus .— .— .— .— .— 724.8 .— .—
Alectoria nigricans .— .— .— .— .— 824.9 1— .—
Polytrichum piliferum 10— .— .— .— .— 3129.2 14— 18—
Hypnum hamulosum .— 36— 19— 2920.6 .— 3— 5— .—
Stegonia latifolia 20— .— .— 1721.8 .— .— .— .—
Anthelia juratzkana .— .— .— 4— .— 1323.4 2— 3—
Cetraria cucullata 20— .— 3— 21— 53— 4823.4 15— 17—
Bryum elegans .— .— .— .— .— 622.6 .— .—
Amphidium lapponicum .— .— .— .— .— 724.8 .— .—
Hypogymnia physodes 20— .— .— 4— .— .— 2— .—
Hypnum revolutum 20— .— 6— 8— .— .— .— .—
Gymnomitrion concinnatum .— .— .— .— .— 1226.4 4— .—
Pohlia drummondii .— .— .— .— .— 826.8 .— .—
Solorina bispora 10— 9— 3— 2529.1 .— 1— 2— .—
Geheebia gigantea 10— .— 16— 2122.7 .— 2— 1— .—
Encalypta ciliata 20— 2729.8 .— 8— .— .— 1— .—
Tritomaria quinquedentata .— .— 3528.9 17— 11— 14— 5— 9—
(continued)
Classification of high-altitude arctic-alpine vegetation 17
Dow
nloa
ded
by [
Ivan
a Sv
itkov
a] a
t 06:
12 2
9 O
ctob
er 2
012

Appendix 1. (Continued)
Group No. 1 2 3 4 5 6 7 8
No. of releves 10 11 31 24 19 86 100 141
Cirriphyllum cirrosum .— .— 23— 17— 5— 1913.2 4— 3—
Ptychodium plicatum .— .— 1929.0 8— 5— .— .— .—
Racomitrium lanuginosum .— .— 19— 4— 5— 28— 3022.2 4—
Paraleucobryum enerve .— .— .— .— .— 14— 5— 6—
Tayloria lingulata .— .— 1029.3 .— .— .— .— .—
Bartramia ithyphylla .— .— 3— .— .— 16— 6— 7—
Cynodontium polycarpon .— 9— .— 21— .— 5— 5— 4—
Cladonia uncialis .— .— .— .— 37— 20— 22— 10—
Cetraria ericetorum .— .— 3— .— 16— 1— 2— 1—
Cladonia coccifera 20— 9— 3— 12— 11— 26— 20— 11—
Encalypta streptocarpa 10— 18— .— .— .— .— .— .—
Encalypta rhaptocarpa 20— 9— .— 12— .— 3— 1— .—
Dicranum scoparium .— .— 29— .— 26— 10— 20— 18—
Hylocomium pyrenaicum .— .— 16— .— 11— .— 4— 3—
Pseudoleskea catenulata 20— 9— 10— 8— .— .— .— .—
Myurella tenerrima 10— 18— 10— .— .— 6— .— .—
Polytrichum strictum .— .— 6— 12— 26— 5— 4— 4—
Pleurozium schreberi .— .— 26— .— 42— 14— 19— 26—
Megaspora verrucosa 20— .— 3— 8— .— 2— .— .—
Pseudevernia furfuracea 20— .— .— 12— .— 2— 4— .—
Racomitrium canescens 10— .— 19— 8— 5— 1— 1— 5—
Cladonia gracilis s.l. .— 18— 6— 17— 42— 27— 26— 31—
Aulacomnium palustre .— .— 3— 4— 16— 1— 2— .—
Rhytidiadelphus squarrosus .— .— 13— .— 11— .— 1— 2—
18 I. Svitkova and J. Sibık
Dow
nloa
ded
by [
Ivan
a Sv
itkov
a] a
t 06:
12 2
9 O
ctob
er 2
012





























