Embed Size (px)
Citation preview

Physiologia Plantarum 2009 Copyright © Physiologia Plantarum 2009, ISSN 0031-9317
Alternative oxidase involvement in Daucus carota somaticembryogenesisAntonio Miguel Fredericoa, Maria Doroteia Camposa, Helia Guerra Cardosoa, Jafargholi Imanib andBirgit Arnholdt-Schmitta,∗
aEU Marie Curie Chair, ICAAM, University of Evora, PortugalbResearch Center for BioSystems, Land Use and Nutrition, IPAZ, Justus-Liebig University of Giessen, Germany
Correspondence*Corresponding author,e-mail: eu [email protected]
Received 9 June 2009; revised 24 July2009
doi:10.1111/j.1399-3054.2009.01278.x
Plant alternative oxidase (AOX) is a mitochondrial inner membrane enzymeinvolved in alternative respiration. The critical importance of the enzymeduring acclimation upon stress of plant cells is not fully understood and is stillan issue of intensive research and discussion. Recently, a role of AOX wassuggested for the ability of plant cells to change easily its fate upon stress.In order to get new insights about AOX involvement in cell reprogramming,quantitative real-time polymerase chain reaction (PCR) and inhibitor studieswere performed during cell redifferentiation and developmental stages ofDaucus carota L. somatic embryogenesis. Transcript level analysis shows thatD. carota AOX genes (DcAOX1a and DcAOX2a) are differentially expressedduring somatic embryogenesis. DcAOX1a shows lower expression levels,being mainly down-regulated, whereas DcAOX2a presented a large up-regulation during initiation of the realization phase of somatic embryogenesis.However, when globular embryos start to develop, both genes are down-regulated, being this state transient for DcAOX2a. In addition, parallel studieswere performed using salicylhydroxamic acid (SHAM) in order to inhibit AOXactivity during the realization phase of somatic embryogenesis. Embryogeniccells growing in the presence of the inhibitor were unable to developembryogenic structures and its growth rate was diminished. This effect wasreversible and concentration dependent. The results obtained contribute tothe hypothesis that AOX activity supports metabolic reorganization as anessential part of cell reprogramming and, thus, enables restructuring and denovo cell differentiation.
Introduction
Somatic embryogenesis (SE) is the most prominentexample of cell reprogramming. Daucus carota L. hasbeen the first species where totipotency through SEhad been proven (see discussion in Costa et al. 2009,Raghavan 2006, Steward et al. 1958). In D. carota, SE
Abbreviations – 2,4-D, 2,4-dichlorophenoxyacetic acid; AOX, alternative oxidase; AtAOX, Arabidopsis thaliana alternativeoxidase; DcAOX, Daucus carota alternative oxidase; DEPC, diethyl pyrocarbonate; DMSO, dimethyl sulfoxide; IBA, indole-3-butyric acid; MMLV-RT, moloney murine leukemia virus-reverse transcriptase; QTL, quantitative trait locus; qRT-PCR,quantitative real-time polymerase chain reaction; SE, somatic embryogenesis; SHAM, salicylhydroxamic acid; SNP, singlenucleotide polymorphism.
is auxin induced and the realization of the inducedprogram depends on the depletion of auxin in themedium (Grieb et al. 1997). Energetic or metabolicrestrictions for the realization process can be expectedto be as important as the induction itself for successfulembryo formation. However, genotypic differences existeven in D. carota as a very ‘easy to induce’ plant
Physiol. Plant. 2009

species. Thus, a strong genetic control for the capacityof cells to form somatic embryos is given. All D.carota genotypes seem to be inducible, but differ inthe timing of embryo formation (Arnholdt-Schmitt B,personal communication). Identification of markers foreasy-embryo formation from D. carota is expectedto help developing functional markers for efficientbiotechnological propagation of recalcitrant genotypes.
Plants as sessile organisms can respond with highflexibility to environmental constraints and opportu-nities. This plasticity includes adaptive biochemical,physiological and morphological reactions and is relatedto high flexibility in metabolism (Foyer and Noctor 2009,McDonald and Vanlerberghe 2006, Plaxton and Podesta2006) and adaptive reprogramming of gene activities(e.g. Clifton et al. 2006, Scheible et al. 2004). SE can beinterpreted as a stress-induced morphogenic response(Feher 2005, Kikuchi et al. 2006, Pasternak et al. 2002,Potters et al. 2007). Such reactions depend on biochem-ical sensors and networks for the integration of bioen-ergetics, signal transduction cascades and metabolism(Sweetlove et al. 2007, Zeng et al. 2007). Mitochondriaare increasingly accepted to play a central role as a phys-ical platform for networks, signal perception and signalcanalization (Amirsadeghi et al. 2007, Fernie et al. 2004,Kohler and Villar 2008, Noctor et al. 2007, Raghaven-dra and Padmasree 2003, Rhoads and Subbaiah 2007,Sweetlove et al. 2007). The significance of mitochon-dria for cell fate decisions through dedifferentiation andde novo differentiation is recognized (Amirsadeghi et al.2007, Sheahan et al. 2005). The alternative respirationpathway is localized in mitochondria and is increasinglygetting into the focus of research on stress acclimationand adaptation. Clifton et al. (2005, 2006) pointed to theimportance of this pathway as an early sensoring systemfor cell programming. AOX is the critical componentin the alternative pathway that transfers electrons fromreduced ubiquinone directly to oxygen. The enzymeis related to all types of abiotic and biotic stress andknown to be involved in growth responses and devel-opment (Giraud et al. 2008, Ho et al. 2007, Sieger et al.2005, Sugie et al. 2007, Umbach et al. 2005). AOX isproposed as a functional marker candidate for efficientcell reprogramming upon stress (Arnholdt-Schmitt et al.2006, see also: www.aox2008.uevora.pt). Induced sin-gle nucleotide polymorphism (SNP) was shown to affecta quantitative trait locus (QTL) for thermo tolerance inOryza sativa L. (Abe et al. 2002). AOX genes demon-strate a rich natural source for polymorphic within-genesequences in various species that might be exploredin the future for functional marker candidates (Cardosoet al. 2009, Ferreira et al. 2009, Holtzapffel et al. 2003,Santos Macedo et al. 2009).
Recently, AOX genes have been isolated fromD. carota (Campos et al. 2009, Cardoso et al. 2009,Costa et al. 2009). The D. carota AOX (DcAOX)multigene family consists of two genes from thesubfamily AOX1 and two genes from the subfamilyAOX2. All four genes were expressed in varioustissues and during development and growth. However,both genes of each subfamily demonstrated differentialactivities among each other. Typically, higher transcriptaccumulation and responsiveness were found for thetwo genes DcAOX1a and DcAOX2a and co-regulation ofboth was demonstrated during de novo growth induction(Campos et al. 2009).
The paper presents indications for the importanceof AOX activity in SE and the specific involvement ofDcAOX genes during realization. DcAOX2a is outlinedas a promising gene candidate for functional markerdevelopment related to the easy-embryo formation ofD. carota.
Material and methods
Plant material
Embryogenic cell line selection
Seeds from D. carota cv. Rotin were surface desinfectedfor 5 min with ethanol at 75% (v/v), followed byan immersion in a solution at 20% (v/v) commercialblench for 20 min and subsequently washed twicewith sterilized bidistilled water. Seeds were placedin Petri dishes (Ø = 9 cm) onto solidified B+
5 culturemedium (modified from Gamborg et al. 1968, see inGrieb et al. 1997) supplemented with 2.3 μM 2,4-dichlorophenoxyacetic acid (2,4-D) in order to induceembryogenic calli development (induction phase).
Selected embryogenic calli were isolated from theoriginal seed material and propagated individually ascell lines. These stock cell lines were maintained inthe dedifferentiated state in the same B+
5 medium asit was used for embryogenic calli induction and sub-cultured every 2 weeks. The cultures were maintainedat 25◦C and under a 16 h photoperiod (approximately95–100 μmol m−2 s−1 of light intensity provided by flu-orescent lamps; Philips, Amsterdam, The Netherlands).To select the cell line with the highest efficiency forthe development of somatic embryos, the obtained lineswere subcultured on solid B−
5 medium under the sameconditions (realization phase). B−
5 contains the samebasal components as B+
5 , but lacks 2,4-D. Selectionparameters were: required time to develop somaticembryos and somatic embryos quality (normal andabnormal development). The cell line L5.S.R. showedthe highest rank related to rapid induction and high
Physiol. Plant. 2009

degrees of normal development and was selected tostudy AOX activities involvement in D. carota SE duringthe realization phase in cell suspension cultures.
L5.S.R. suspension culture and realization experiments
The cell line L5.S.R. was used to initiate cell suspensioncultures in Erlenmeyer flasks containing 33% (v/v) B+
5liquid culture medium supplemented with 0.9 μM 2,4-D. Subcultures were made every 10 days. Cell diameterhomogeneity was maintained through periodic sieving atsubculture, using 95 μm mesh pore polyester precisionwoven screens (Sefar Petex, Thal, Switzerland). Theembryogenic competence was evaluated periodicallyas described above. The cultures were kept at 28◦C,continuous light (approximately 95–100 μmol m−2 s−1
provided by fluorescent lamps; Philips, Amsterdam, TheNetherlands) and 90–95 rpm on an orbital shaker (SM-30, Edmund Buhler, Hechingen, Germany).
For realization experiments, the L5.S.R. cell line wastaken during the exponential growth phase, between4 and 6 days after subculture and sieved to achievehomogeneity (see Fig. 1). Flow-through cells werewashed twice by centrifugation at 300 G with fresh B−
5medium. Experiments were performed by using equalpacked cell volumes of 0.5% (v/v) in Erlenmeyer flasksfilled with 33% (v/v) B−
5 culture medium at the physicalconditions referred above.
To investigate the influence of the AOX inhibitorsalicylhydroxamic acid (SHAM) on cell growth rates andthe ability of embryogenic cells to develop embryogenicstructures, a realization experiment was performedduring 12 days. Seven repetitions were performed: threewere used to evaluate the development of embryogenicstructures (4th, 8th and 12th day after inoculation) andfour were taken to determine dry weights at the 12th dayafter inoculation.
To quantify AOX transcript levels during the real-ization process, 12 samples were collected during a10 days’ time course experiment (see Fig. 1). Cells and/orembryos were collected through sieving with 54 μmmesh pore polyester precision woven screens (SefarPetex, Thal, Switzerland), immediately disrupted in liq-uid nitrogen and stored at –80◦C until RNA extraction.
The use of SHAM in the AOX inhibition studies
A 500 mM SHAM (Aldrich, St. Louis, MO, USA)stock solution was prepared freshly. Dimethyl sulfox-ide (DMSO) (Fluka BioChemika, St. Louis, MO, USA)(50% v/v) was the primary solvent and bidistilled waterwas added (50% v/v). The pH of the SHAM stock solutionwas adjusted to culture medium pH (5.72) and sterilized
by filtration using a 0.22 μm mesh pore filter (Sarstedt,Numbrecht, Germany).
SHAM concentrations were optimized by consider-ing its effect on cell growth and embryo development.The final SHAM concentrations tested were: 0.2 mM,0.4 mM, 0.6 mM, 0.8 mM, 1 mM and 1.5 mM. Thecontrol treatment was performed without SHAM sup-plementation.
To exclude any negative effect of the solvent DMSO asit was observed in other cell culture systems (Randhawa2008, Rodrıguez-Burford et al. 2003, Wang et al. 2002),parallel realization experiments were performed bysupplementing suspension cultures only with DMSO.
Hydroxamic acids such as SHAM are able to chelateferric iron (Rich et al. 1978) and browning was observedin the D. carota suspension cultures after SHAMapplication. Therefore, control experiments withoutiron in the medium were conducted to exclude thepossibility of negative effects by iron depletion duringthe realization experiments.
AOX qRT-PCR studies
RNA extraction and cDNA synthesis
Total RNA was extracted by using the RNeasy® plantmini kit (Qiagen, Hilden, Germany) according to themanufacturer’s instruction. After DNase I (Fermentas,Ontario, Canada) treatment, RNA integrity was analyzedon 1.4% DEPC (diethyl pyrocarbonate) (Sigma, St. Louis,MO, USA) agarose gel and total RNA was quantifiedin a spectrophotometer (DU 530, Beckman CoulterInc., Fullerton, CA, USA) using a 50 μl microcell. Forfirst-strand cDNA synthesis, 2 μg of DNA-free RNAwas used with the moloney murine leukemia virus-reverse transcriptase (MMLV-RT) and oligo (dT)18 primeraccording the RETROscript® kit (Ambion, Austin, TX,USA) manufacturer’s instruction.
qRT-PCR primers design
For AOX transcript quantification by qRT-PCR, spe-cific primers were designed. The forward (5′-CTTCAACGCCTACTTCCTTG) and the reverse primer(5′-ATCTCGCAATGTAGAGTCAGC) were designed forthe amplification of a 196 bp size fragment of DcAOX1a(EU286573). For the amplification of the DcAOX2a(EU286575) 200 bp size fragment, the forward (5′-TCTTCAATGCTTTCTTTGTTCTT) and the reverse primer(5′-GACATCTTTTAGTTTGGCATCTTT) were applied.As housekeeping, the D. carota ubiquitin-carboxylextension gene (U68751) was used. The specific primersto amplify a 152 bp size fragment for this gene were:
Physiol. Plant. 2009
30 μm
50 μm
200 μm
200 μm
To
rped
oC
oty
led
on
ary
50 μmGlo
bu
lar
100 μm
Hea
rt
50 μm
T-2 – Flow-through cells sieved with a 200 μm mesh pore screen in B5+
T-1 – Flow-through cells sieved with a 95 μm mesh pore screen in B5+
T0 – Washed cells with B5- by centrifugation (SE start)
T1 – 1 hour after inoculation in B5-
T2 – 4 hours after inoculation in B5-
T3 – 12 hours after inoculation in B5-
T4 – 24 hours after inoculation in B5-
T5 – 2 days after inoculation in B5-
T6 – 3 days after inoculation in B5-
T7 – 5 days after inoculation in B5-
T8 – 7 days after inoculation in B5-
T9 – 10 days after inoculation in B5-
Pre
-glo
bu
lar
cells
D. c
aro
ta c
v. R
oti
n s
om
atic
em
bry
og
enes
is r
ealiz
atio
n s
tag
es
Physiol. Plant. 2009

forward (5′-AAGCCCAAGAAGATCAAGCA) and reverse(5′-TCAAAATGATTGGCCATGAA).
qRT-PCR conditions
qRT-PCR was performed in a Mx3000P® qPCR sys-tem (Stratagene, La Jolla, CA, USA). About 15 μl PCRwas applied using 10 ng of cDNA and SYBR® GreenJumpStart™ Taq ReadyMix™ (Sigma, St. Louis, MO, USA)in 96-well plates. Specific primers were adjusted to thefinal concentration of 166 nM. An initial step at 95◦Cduring 7 min for initial template denaturation was runfollowed by 40 cycles consisting each one in 95◦Cduring 30 s for denaturation; 60, 52 or 51◦C during30 s for primers annealing (DcAOX1a, DcAOX2a andubiquitin, respectively); and finally the extension step at72◦C during 30 s. To analyze dissociation curve profiles,an additional step at 95◦C during 1 min was added, fol-lowed by a constant increase of the temperature between60 and 95◦C. For each sample, PCRs were ran intriplicate.
Data analyses
The statistical analyses and graphics applied to calculatethe effect of SHAM on growth rates were performedwith SPSS 16.0 software (SPSS Inc., Chicago, IL, USA).Growth of cells and embryo structures was measuredas dry weights. Dry weights are presented in a boxplotanalysis from four repetitions of each applied treatmentincluding the control. Maximum and minimum dryweights, the median and the 50% quartile were thestatistical parameters used to illustrate the results.
Somatic embryo stages were named in accordancewith Yeung (1995). Images representing the cells andembryo developmental stages (see Fig. 1 and 3) weretaken using a digital camera (PowerShot A630, CanonInc., Tokyo, Japan). The images representing cells orembryos were selected as representing the most commondevelopmental stage observed in three samples collectedfrom different suspension cultures of each treatmentor experiment using an inverted microscope (CK40,Olympus, Tokyo, Japan). The effect of SHAM on embryostructure development was evaluated by the presence orabsence of embryo structures.
Statistical calculations associated with the 2−��CT
methodology (Livak and Schmittgen 2001) for qRT-PCR were applied for the triplicated CT values
obtained for each sample using the freeware qCal-culator version 1.0 (freely available at: www.gene-quantification.de/qCalculator.zip). The sample collectedat T-2 was selected as the calibrator. The results are pre-sented as the average of the triplicated CT values fromeach sample. Standard deviation was residual among thesample triplicates.
Results
The effect of SHAM on the realization of SE
The effect of AOX pathway inhibition on SE was studiedby applying SHAM permanently to an embryogenic cellline during the realization phase of the embryogenicprogramme induced by auxin. Fig. 2 demonstrates thedry weights of the embryogenic cells and structuresachieved after 12 days with and without SHAMtreatments. Under the control treatment, the dry weightswere in average 9.6 times higher (approximately 106 mg)than for the initially inoculated cells (approximately11 mg) (data not shown). When the culture mediumwas supplemented with SHAM, the highest value ofthe dry weight at the 12th day was always lowerthan in the control treatment. At the three lowestSHAM concentrations tested (0.2 mM, 0.4 mM and0.6 mM), the variation in dry weight between the fourrepetitions was higher in comparison with the threeupper concentrations (0.8 mM, 1 mM and 1.5 mM).From the six SHAM concentrations tested, the highestdry weight variation was found at 0.6 mM, ranging from32 to 86 mg dry weight (Fig. 2). However, the lowest dryweight variation was detected at 1.5 mM SHAM, rangingfrom 16 mg dry weight to 23 mg.
In Fig. 3, the development of embryo structuresat the 4th, 8th and 12th day of the experiment isshown at different concentrations of SHAM. WhenSHAM was supplied at the lower concentrations of0.2 and 0.4 mM, a small delay in the appearanceof embryogenic structures (globular and heart shapedembryos) was observed at the fourth and eighth day inrelation to the control treatment. However, the abilityto develop embryogenic structures was not suppressed.At the 12th day, it was not possible to detect by theapplied methodology any difference among these twoSHAM-treated suspension cultures and the control inrelation to the development of embryogenic structures.At concentrations higher than 0.6 mM, the ability of the
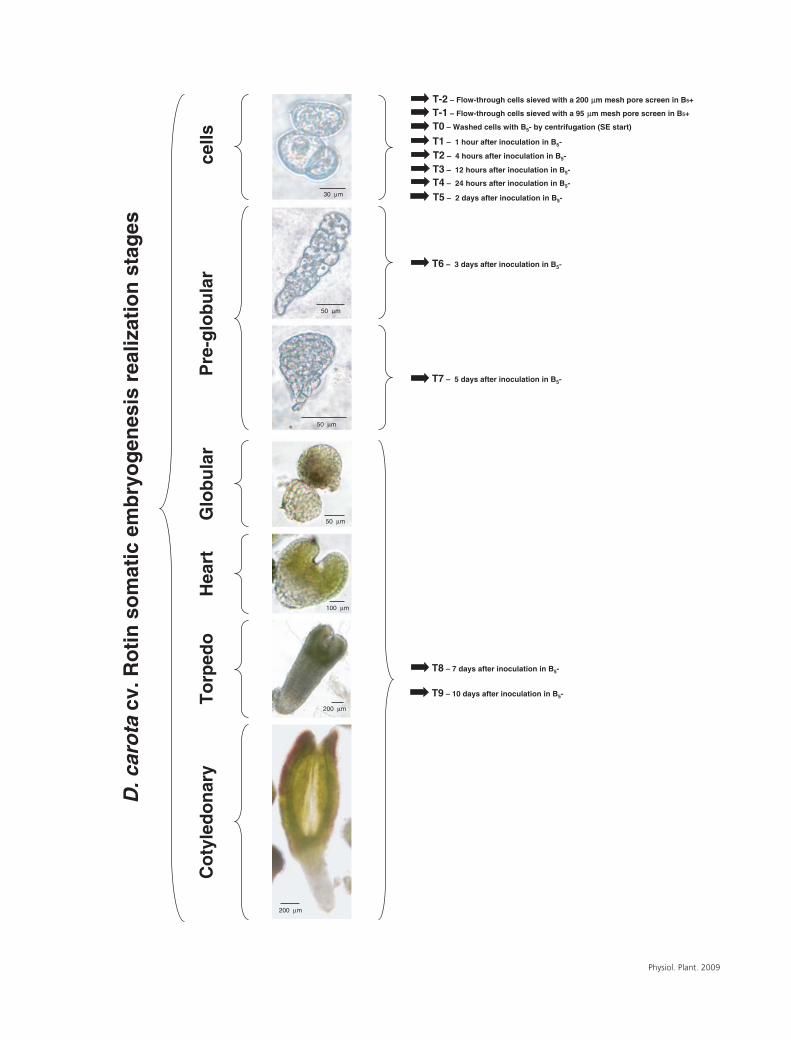
Fig. 1. Experimental design to study AOX transcript accumulation during SE realization (Daucus carota L. cv. Rotin, cell line L5.S.R.). SE developmentalstages are schematically presented in relation to the sampling times for qRT-PCR analyses. The images are showing the most advanced stages ofdevelopment in the suspension culture at the time of sampling. Samples T-2 and T-1 were harvested from B+
5 medium. All other samples were takenfrom B−
5 medium. T0 represents the beginning of the SE realization phase.
Physiol. Plant. 2009

5101520253035404550556065707580859095
100105110115120
0Control 0.2 0.4 0.80.6 1.0 1.5
SHAM (mM )
Dry
wei
ght (
mg)
Fig. 2. The effect of SHAM on dry weights of cells and embryogenicstructures during SE realization (Daucus carota L. cv. Rotin, cell lineL5.S.R.). Cells were inoculated in liquid B−
5 medium supplementedwith different SHAM concentrations ranging from 0.2 to 1.5 mM. Thedry weight of cells and embryos were measured after 12 days in theSE realization medium. A boxplot analysis was performed for fourrepetitions.
embryogenic cells to form embryogenic structures wascompletely suppressed. No globular-shaped structureswere visible in all three repetitions of all treatments.
SHAM-treated cells that are suppressed in theirdevelopment to form embryogenic structures at higherconcentrations do not lose permanently this ability. Ifsubcultured in B−
5 medium without SHAM they canrecover and develop embryogenic structures (data notshown).
When applied alone DMSO, at the applied volumes,did not have any measurable or visible effect on cellgrowth rates or the ability to develop embryogenicstructures. Also, the lack of medium iron did not affectnormal suspension culture growth or embryo structureformation during the realization phase (data not shown).Globular, heart and torpedo-shaped embryos developedas in untreated iron-supplemented suspension cultures(data not shown).
Transcription of DcAOX1a and DcAOX2a duringthe realization phase
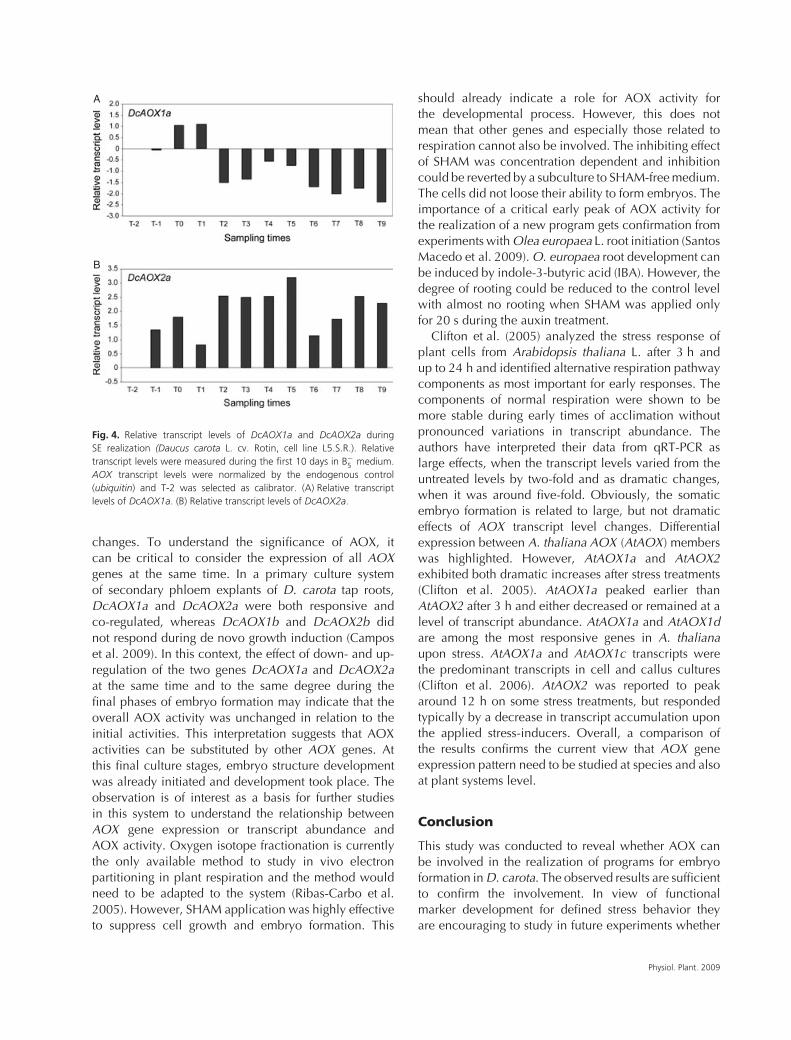
Figure 4 shows the result of qRT-PCR analysis ofDcAOX1a and DcAOX2a expression during the realiza-tion phase of SE. The accumulation of transcripts is givenin reference to the initial level for each gene (see T-2 inFig. 1). The two genes were differentially expressed. Thelevel of transcript accumulation of DcAOX1a was lowerduring the realization of the embryogenic program than
at the beginning of the experiment (Fig. 4A), whereasDcAOX2a was up-regulated and remained high duringthe experiment in comparison with the initial level at thetransfer to B−
5 medium (Fig. 4B).Sieving had no direct effect on DcAOX1a transcript
levels. However, a one-fold increase was observed when2,4-D was removed from the culture medium and thecells were centrifuged (T0, Fig. 4A). This minimal changeremained visible also 1 h later (T1, Fig. 4A). DcAOX2atranscripts accumulated the same amount of transcriptsas DcAOX1a already at sieving (T-1, one-fold change,Fig. 4) and remained at almost the same level duringcentrifugation (T0, 1.5-fold, Fig. 4B).
Four hours after inoculation in B−5 medium, DcAOX1a
transcript accumulation level decreased largely (> 2.5-fold) (T2, Fig. 4A). DcAOX2a decreased also, but onlyslightly and earlier at 1 h after inoculation (T1, Fig. 4B).The clear difference in regulation between the twogenes starts to become obvious 4 h after inoculation (T2,Fig. 4A, B). DcAOX2a showed a 2.5-fold up-regulationin comparison with the experimental start. Contrarily,DcAOX1a is clearly down-regulated (approximately1.5-fold) at that stage. Both genes show a slightlyincreased transcript accumulation between 24 and 48 hand a down-regulation at 72 h (T6, Fig. 4A and B)after inoculation, which is especially pronounced forDcAOX2a (two-fold). This stage is characterized bythe appearance of globular structures (see T6, Fig. 1).Down-regulation of DcAOX1a resulted in a level oftranscripts that remained more or less stable during thefollowing periods until the end of the experiment. Thelarge decrease in transcripts at T6 observed for DcAOX2awas transient with a subsequent up-regulation during thefollowing 5–10 days (up to 2.5-fold at day 7).
Discussion
Campos et al. (2009) reported that both DcAOX1aand DcAOX2a showed the highest level of transcriptaccumulation in tissues and organs and had been co-regulated during the first days of de novo growth induc-tion of secondary phloem tissues from D. carota tap roots.However, co-regulation seems to be system- and devel-opment dependent. Here, we present the expression ofthese two genes in another biological system duringthe realization of SE from auxin-induced embryogeniccells. The initiation of somatic embryo formation fromauxin-induced embryogenic cells is accompanied byimportant differential AOX transcription of both genes.However, the two genes revealed also phases of co-regulation. Similar regulation was observed at criticalpoints for transitions such as directly when auxin isremoved from the medium as well as when globular
Physiol. Plant. 2009

0.2 mM
0.4 mM
0.6 mM
0.8 mM
1 mM
1.5 mM
control
100 μm
100 μm 100 μm
100 μm
100 μm
100 μm
100 μm
100 μm
4th day 8th day 12th day
100 μm
100 μm
100 μm
100 μm
100 μm 100 μm
100 μm 100 μm
100 μm 100 μm
100 μm
Treatments
4th day 8th day 12th dayTreatments
100 μm
100 μm
100 μm 100 μm
100 μm
Fig. 3. The effect of SHAM on embryo structure formation during SE realization (Daucus carota L. cv. Rotin, cell line L5.S.R.). The figure showsrepresentative photos taken at various times during the SE realization phase of cell line L5.S.R. in B−
5 medium: initial cell suspension before inoculationinto B−
5 medium (left side); cells and embryogenic structures at the 4th, 8th and 12th day, during SE realization in the control treatment withoutSHAM and at increasing supplementations of SHAM from 0.2 to 1.5 mM.
structures start to be formed. Differential regulation wasespecially obvious during the first hours of acclima-tion to the auxin-depleted medium. The transcript levelof DcAOX2a was 2.5-fold up-regulated and DcAOX1awas around 1.5-fold down-regulated in comparison withthe initial transcript levels of each gene. Interestingly, thedown-regulation of DcAOX1a and the up-regulation ofDcAOX2a between the 5th and the 10th days of embryoformation are ranging both around a two-fold change intranscript accumulation, but in the opposite direction.
Up-regulation of both genes upon sieving and thedepletion of auxin can be interpreted as a general stressresponse. It shows the immediate reaction upon changesof the environmental conditions. However, acclimation
to auxin-depletion during the very first hours seems to berelated to a more specific response of both AOX genes.Campos et al. (2009) studied AOX transcript levelsstarting from 1.5 days after de novo growth inductionand found co-regulation of both genes. However, themain point where differential expression between bothgenes seems to play a role during embryo formation isalready earlier during the first 12 h. During that time,the 2.5-fold up-regulated and 1.5-fold down-regulatedtranscript levels from both genes together count in thebalance for an overall up-regulation of AOX transcripts(one-fold). This observation can be important in viewof the current discussion about the effect of transcriptabundance variability as a response upon environmental
Physiol. Plant. 2009
A
B
Fig. 4. Relative transcript levels of DcAOX1a and DcAOX2a duringSE realization (Daucus carota L. cv. Rotin, cell line L5.S.R.). Relativetranscript levels were measured during the first 10 days in B−
5 medium.AOX transcript levels were normalized by the endogenous control(ubiquitin) and T-2 was selected as calibrator. (A) Relative transcriptlevels of DcAOX1a. (B) Relative transcript levels of DcAOX2a.
changes. To understand the significance of AOX, itcan be critical to consider the expression of all AOXgenes at the same time. In a primary culture systemof secondary phloem explants of D. carota tap roots,DcAOX1a and DcAOX2a were both responsive andco-regulated, whereas DcAOX1b and DcAOX2b didnot respond during de novo growth induction (Camposet al. 2009). In this context, the effect of down- and up-regulation of the two genes DcAOX1a and DcAOX2aat the same time and to the same degree during thefinal phases of embryo formation may indicate that theoverall AOX activity was unchanged in relation to theinitial activities. This interpretation suggests that AOXactivities can be substituted by other AOX genes. Atthis final culture stages, embryo structure developmentwas already initiated and development took place. Theobservation is of interest as a basis for further studiesin this system to understand the relationship betweenAOX gene expression or transcript abundance andAOX activity. Oxygen isotope fractionation is currentlythe only available method to study in vivo electronpartitioning in plant respiration and the method wouldneed to be adapted to the system (Ribas-Carbo et al.2005). However, SHAM application was highly effectiveto suppress cell growth and embryo formation. This
should already indicate a role for AOX activity forthe developmental process. However, this does notmean that other genes and especially those related torespiration cannot also be involved. The inhibiting effectof SHAM was concentration dependent and inhibitioncould be reverted by a subculture to SHAM-free medium.The cells did not loose their ability to form embryos. Theimportance of a critical early peak of AOX activity forthe realization of a new program gets confirmation fromexperiments with Olea europaea L. root initiation (SantosMacedo et al. 2009). O. europaea root development canbe induced by indole-3-butyric acid (IBA). However, thedegree of rooting could be reduced to the control levelwith almost no rooting when SHAM was applied onlyfor 20 s during the auxin treatment.
Clifton et al. (2005) analyzed the stress response ofplant cells from Arabidopsis thaliana L. after 3 h andup to 24 h and identified alternative respiration pathwaycomponents as most important for early responses. Thecomponents of normal respiration were shown to bemore stable during early times of acclimation withoutpronounced variations in transcript abundance. Theauthors have interpreted their data from qRT-PCR aslarge effects, when the transcript levels varied from theuntreated levels by two-fold and as dramatic changes,when it was around five-fold. Obviously, the somaticembryo formation is related to large, but not dramaticeffects of AOX transcript level changes. Differentialexpression between A. thaliana AOX (AtAOX) memberswas highlighted. However, AtAOX1a and AtAOX2exhibited both dramatic increases after stress treatments(Clifton et al. 2005). AtAOX1a peaked earlier thanAtAOX2 after 3 h and either decreased or remained at alevel of transcript abundance. AtAOX1a and AtAOX1dare among the most responsive genes in A. thalianaupon stress. AtAOX1a and AtAOX1c transcripts werethe predominant transcripts in cell and callus cultures(Clifton et al. 2006). AtAOX2 was reported to peakaround 12 h on some stress treatments, but respondedtypically by a decrease in transcript accumulation uponthe applied stress-inducers. Overall, a comparison ofthe results confirms the current view that AOX geneexpression pattern need to be studied at species and alsoat plant systems level.
Conclusion
This study was conducted to reveal whether AOX canbe involved in the realization of programs for embryoformation in D. carota. The observed results are sufficientto confirm the involvement. In view of functionalmarker development for defined stress behavior theyare encouraging to study in future experiments whether
Physiol. Plant. 2009

within-gene polymorphisms can be related to changes indifferential gene regulation and expression patterns thatcan influence the efficiency of the realization of embryoformation. DcAOX2a can be highlighted as a promisinggene marker candidate. The genomic sequence ofDcAOX2a is already known (Campos et al. 2009,Cardoso et al. 2009) and important polymorphisms inintron 3 have been indicated. Interestingly, Cardoso et al.(2009) predicted a pre-miRNA site in the polymorphicsequence in intron 3 that seems to be related toembryogenesis. Studies are in progress to validate thissite as a functional marker candidate in the presentedsystem. In addition, DcAOX2a was mapped to linkagegroup 7 of the segregating population DM19 and will beexplored for a correlation to other markers present in thesame linkage group (Cardoso et al. 2009).
Acknowledgements – The authors express a special thankto Virgınia Sobral for her excellent technical assistance inthe biotechnological tasks and her engaged collaboration.The authors also thank our colleagues at ICAAM, Universityof Evora, Augusto Peixe and Maria A. Zavattieri, forbeing always available for fruitful discussions, collaborationand support. The authors feel grateful to the EuropeanCommission for providing the Marie Curie Chair toB. A.-S. The authors appreciate the financial supportprovided by the Fundacao para a Ciencia e a Tecnologiathrough fellowships to A.M.F. (FCT:SFRH/BI/33048/2007),M.D.C. (GRICES/FCT:SFRH/BI/15991/2006) and to H.G.C.(FCT:SFRH/BPD/27016/2006). Collaboration between thePortuguese and the German group was supported by abilateral cooperation contract through GRICES/FCT andDAAD. The authors thank also Karl-Heinz Kogel and hisgroup from IPAZ, Germany, especially Gregor Langen, forgiving support to this collaboration.
References
Abe F, Saito K, Miura K, Toriyama K (2002) A singlenucleotide polymorphism in the alternative oxidasegene among rice varieties differing in low temperaturetolerance. FEBS Lett 527: 181–185
Amirsadeghi S, Robson CA, Vanlerberghe GC (2007) Therole of the mitochondrion in plant responses to bioticstress. Physiol Plant 129: 253–266
Arnholdt-Schmitt B, Costa JH, Fernandes de Melo D(2006) AOX - A functional marker for efficient cellreprogramming under stress? Trends Plant Sci 11:281–287
Campos MD, Cardoso HG, Linke B, Costa JH, Fernandesde Melo D, Justo L, Frederico AM, Arnholdt-Schmitt B(2009) Differential expression and co-regulation ofcarrot AOX genes (Daucus carota). Physiol Plant doi:10.1111/j.1399-3054.2009.01282.x
Cardoso HG, Campos MD, Costa AR, Campos MC,Nothnagel T, Arnholdt-Schmitt B (2009) Carrotalternative oxidase gene AOX2a demonstrates allelicand genotypic polymorphisms in intron 3. Physiol Plantdoi: 10.1111/j.1399-3054.2009.01299.x
Clifton R, Lister R, Parker KL, Sappl PG, Elhafez D,Millar AH, Day DA, Whelan J (2005) Stress-inducedco-expression of alternative respiratory chaincomponents in Arabidopsis thaliana. Plant Mol Biol 58:193–212
Clifton R, Millar AH, Whelan J (2006) Alternative oxidasesin Arabidopsis: A comparative analysis of differentialexpression in the gene family provides new insights intofunction of non-phosphorylating bypasses. BiochimBiophys Acta 1757: 730–741
Costa JH, Cardoso HC, Campos MD, Zavattieri A,Frederico AM, Fernandes de Melo D,Arnholdt-Schmitt B. (2009). Daucus carota L. - an oldmodel for cell reprogramming gains new importancethrough a novel expansion pattern of AOX genes. PlantPhysiol Biochem 47: 753–759
Feher A (2005) Why somatic plant cells start to formembryos? In: Mujib A, Samaj J (eds) SomaticEmbryogenesis. Plant Cell Monographs, Vol. 2, Springer,Heidelberg, pp 85–101
Fernie AR, Carrari F, Sweetlove LJ (2004) Respiratorymetabolism: glycolysis, the TCA cycle andmitochondrial electron transport. Curr Opin Plant Biol7: 254–261
Ferreira A, Cardoso HG, Santos Macedo E, Breviorio D,Arnholdt-Schmitt B (2009) Intron polymorphism patternin AOX1b of wild St. John’s Wort (Hypericumperforatum) allows discrimination between individualplants. Physiol Plant doi:10.1111/j.1399-3054.2009.01291.x
Gamborg O, Miller RA, Ojima K (1968) Nutrientrequirements of suspension cultures of soybean rootcells. Exp Cell Res 50: 151–158
Giraud E, Ho LHM, Clifton R, Carroll A, Estavillo G,Tan Y-F, Howell KA, Ivanova A, Pogson BJ, Millar AH,Whelan J (2008) The absence of alternative oxidase1a inArabidopsis results in acute sensitivity to combined lightand drought stress. Plant Physiol 147: 595–610
Grieb B, Schafer F, Imani J, Mashayekhi KN,Arnholdt-Schmitt B, Neumann KH (1997) Changes insoluble proteins and phytohormone concentrations ofcultured carrot petiole explants during induction ofsomatic embryogenesis (Daucus carota L.). J Appl Bot71: 94–103
Foyer CH, Noctor G (2009) Redox regulation inphotosynthetic organisms: Signaling, acclimation,and practical implications. Antioxid Red Signal 11:181–905
Ho LHM, Giraud E, Lister R, Thirkettle-Watts D, Low J,Clifton R, Howell KA, Carrie C, Donald T, Whelan J
Physiol. Plant. 2009

(2007) Characterization of the regulatory and expressioncontext of an alternative oxidase gene provides insightsinto cyanide-insensitive respiration during growth anddevelopment. Plant Physiol 143: 1519–1533
Holtzapffel RC, Castelli J, Finnegan PM, Millar AH,Whelan J, Day DA (2003) A tomato alternative oxidaseprotein with altered regulatory properties. BiochimBiophys Acta 1606: 153–162
Kikuchi A, Sanuki N, Higashi K, Koshiba T, Kamada H(2006) Abscisic acid and stress treatment are essentialfor the acquisition of embryogenic competence bycarrot somatic cells. Planta 223: 637–645
Kohler C, Villar CB (2008) Programming of geneexpression by Polycomb group proteins. Trends CellBiol 18: 236–243
Livak KJ, Schmittgen TD (2001) Analysis of relative geneexpression data using real-time quantitative PCR and the2[-Delta DeltaC(T)] Method. Methods 25: 402–408
McDonald AE, Vanlerberghe GC (2006) The organizationand control of plant mitochondrial metabolism. In:Plaxton WC, McManus MT (eds) Control of PrimaryMetabolism in Plants. Annual Plant Reviews, Vol 22,Blackwell, Oxford, pp 290–324
Noctor G, De Paepe R, Foyer CH (2007) Mitochondrialredox biology and homeostasis in plants. Trends PlantSci 12: 125–134
Pasternak T, Prinsen E, Ayaydin F, Miskolczi P, Potters G,Asard H, van Onckelen H, Dudits D, Feher A (2002)The role of auxin, pH and stress in the activation ofembryogenic cell division in leaf protoplast-derivedcells of alfalfa (Medicago sativa L.). Plant Physiol 129:1807–1819
Plaxton WC, Podesta FE (2006) The functionalorganisation and control of plant respiration. Crit RevPlant Sci 25: 159–198
Potters G, Pasternak TP, Guisez Y, Palme KJ, Jansen MAK(2007) Stress-induced morphogenic responses: growingout of trouble? Trends Plant Sci 12: 98–105
Raghavan V (2006) Can carrot and Arabidopsis serve asmodel systems to study the molecular biology of somaticembryogenesis? Curr Sci 90: 1336–1343
Raghavendra AS, Padmasree K (2003) Beneficialinteractions of mitochondrial metabolism withphotosynthetic carbon assimilation. Trends Plant Sci 8:546–553
Randhawa MA (2008) Dimethyl sulfoxide (DMSO) inhibitsthe germination of Candida albicans and theArthrospores of Trichophyton mentagrophytes. NipponIshinkin Gakkai Zasshi 49: 125–128
Rhoads DM, Subbaiah CC (2007) Mitochondrial retrograderegulation in plants. Mitochondrion 7: 177–194
Ribas-Carbo M, Taylor NL, Giles L, Busquets S,Finnegan PM, Day DA, Hambers H, Medrano H,Berry JA, Flexas J (2005) Effects of water stress on
respiration in soybean leaves. Plant Physiol 139:466–473
Rich PR, Wiegand NK, Blum H, Moore AL, Bonner WD(1978) Studies on the mechanism of inhibition of redoxenzymes by substituted hydroxamic acids. BiochimBiophys Acta 525: 325–337
Rodrıguez-Burford C, Oelschlager DK, Talley LI, BarnesMN, Partridge EE, Grizzle WE (2003) The use ofdimethylsulfoxide as a vehicle in cell cultureexperiments using ovarian carcinoma cell lines. BiotechHistochem 78: 17–21
Santos Macedo E, Cardoso HG, Hernandez A, Peixe AA,Polidoros A, Ferreira A, Cordeiro A, Arnholdt-Schmitt B(2009) Physiological responses and gene diversityindicate olive alternative oxidase AOX as a potentialsource for markers involved in efficient adventitiousrooting induction. Physiol Plant doi:10.1111/j.1399-3054.2009.01302.x
Scheible WR, Morcuende R, Czechowski T, Fritz C,Osuna D, Palacios-Rojas N, Schindelasch D, Thimm O,Udvardi M, Stitt M (2004). Genome-widereprogramming of primary and secondary metabolism,protein synthesis, cellular growth processes, and theregulatory infrastructure of Arabidopsis in response tonitrogen. Plant Physiol 136: 2483–2499
Sheahan MB, McCurdy DW, Rose RJ (2005) Mitochondriaas a connected population: ensuring continuity of themitochondrial genome during plant celldedifferentiation through massive mitochondrial fusion.Plant J 44: 744–755
Sieger SM, Kristensen BK, Robson CA, Amirsadeghi S,Eng EWY, Abdel-Mesih A, Moller IM, Vanlerberghe GC(2005) The role of alternative oxidase in modulatingcarbon use efficiency and growth during macro-nutrient stress in tobacco cells. J Exp Bot 56: 1466–1515
Steward F, Mapes M, Mears K (1958) Growth andorganized development in cultured cells. II.Organization in cultures grown from freely suspendedcells. Am J Bot 45: 705–707
Sugie A, Murai K, Takumi, S (2007) Alteration ofrespiration capacity and transcript accumulation level ofalternative oxidase genes in necrosis lines of commonwheat. Genes Genet Syst 82: 231–239
Sweetlove LJ, Fait A, Nunes-Nesi A, Williams T, Fernie AR(2007) The mitochondrion: an integration point ofcellular metabolism and signalling. Crit Rev Plant Sci26: 17–43
Umbach AL, Fiorani F, Siedow JN (2005) Characterizationof transformed Arabidopsis with altered alternativeoxidase levels and analysis of effects on reactive oxygenspecies in tissue. Plant Physiol 139: 1806–1820
Wang X, Yuan YJ, Li JC, Wu JC, Yang WL (2002).Biological changes in suspension cultures of Taxuscuspidata induced by dimethyl sulfoxide and ethanol.J Mol Catal B-Enzym 18: 211–217
Physiol. Plant. 2009

Yeung E (1995) Structural and developmental patterns insomatic embryogenesis. In: Thorpe TA (ed) In vitroEmbryogenesis in Plants. Current Plant Scienceand Biotechnology in Agriculture, Vol 20,Kluwer, Dordrecht, pp 205–247
Zeng F, Zhang X, Cheng L, Hu L, Zhu L, Cao J,Guo X (2007) A draft gene regulatory network forcellular totipotency reprogramming duringplant somatic embryogenesis. Genomics 90:620–628
Edited by V. Hurry
Physiol. Plant. 2009





























