Embed Size (px)
Citation preview

JOURNAL OF VIROLOGY, Oct. 1996, p. 6994–7003 Vol. 70, No. 100022-538X/96/$04.0010Copyright q 1996, American Society for Microbiology
Adenovirus-Mediated Gene Transfer to Ciliated AirwayEpithelia Requires Prolonged Incubation Time
JOSEPH ZABNER,1 BERNHARDT G. ZEIHER,1 ELLIOT FRIEDMAN,2 AND MICHAEL J. WELSH1*
Howard Hughes Medical Institute, Departments of Internal Medicine and Physiology and Biophysics,University of Iowa College of Medicine, Iowa City, Iowa 52242,1 and Universidad
Central de Venezuela, Caracas 1051, Venezuela2
Received 28 March 1996/Accepted 9 June 1996
The efficiency of adenovirus-mediated gene transfer to airway epithelia will be an important factor indetermining whether recombinant adenoviruses can be developed as vectors for transferring cystic fibrosistransmembrane conductance regulator (CFTR) cDNA to patients with cystic fibrosis. Current understandingof the biology of CF lung disease suggests that vectors should express transgene in mature, ciliated airwayepithelia. We evaluated the efficiency of adenovirus-mediated gene transfer to primary cultures of normal andCF human airway epithelia. Our studies showed that the airway cells developed from an undifferentiatedepithelium with markers characteristic of basal cells and a surface covered by short microvilli 3 days afterseeding to a mature epithelium whose apical surface was covered with cilia by 10 to 14 days. The ability ofadenovirus vectors to express a reporter gene and to correct defective cyclic AMP-stimulated Cl2 transport inCF epithelia was correlated inversely with the state of differentiation. However, the inefficiency of adenovirus-mediated gene transfer could be partially corrected when the contact time between vector and epithelium wasprolonged. After prolonged contact, we observed complete correction of the CF Cl2 transport defect indifferentiated CF airway epithelia in culture and of the Cl2 transport defect in the nasal epithelia of micehomozygous for the DF508 mutation. The fact that gene transfer to airway epithelia required prolongedincubation with vector contrasts with the rapid infection observed in cell models such as 293 and HeLa cells,which are commonly used to study adenovirus infection. Gene transfer observed after prolonged incubationmay result from mechanisms different from those that mediate infection of 293 cells. These observationssuggest that interventions that either increase the contact time or alter the epithelium or the vector may berequired to facilitate gene transfer to ciliated respiratory epithelia.
Recombinant adenoviruses are used frequently as vectorsfor gene transfer to a wide variety of cells and tissues, and it ishoped that they may prove useful for gene therapy. In thegenetic disease cystic fibrosis (CF), recombinant adenovirusesare being investigated as vectors to transfer cystic fibrosistransmembrane conductance regulator (CFTR) cDNA to air-way epithelia (4, 9, 22, 39, 41, 42). However, two generalproblems hinder adenovirus-mediated gene transfer for thisapplication. First, the efficiency of gene transfer to airway ep-ithelium is limited (14, 22, 42). Second, immunologic responsescan prevent infection, limit the duration of expression, andcause inflammation (20, 30, 37, 38). In the case of the immu-nologic and inflammatory responses, a number of studies arefocusing on identification of the humoral and cytotoxic mech-anisms involved. The results of such work suggest that alter-ations might be made in the vector and that immunosuppres-sive strategies might be useful. However, the low efficiency ofgene transfer to airway epithelia may prove to be an evengreater obstacle because if infection of airway epithelia byadenovirus is very inefficient, then the therapeutic index isunlikely to be favorable. On the other hand, if gene transferwas very efficient, then the dose of vector could be low andconsequently adverse events would be minimized.On first inspection, several considerations suggested that
recombinant adenoviruses would be attractive vectors for genetransfer to airway epithelia. Infection with type C adenovirus is
common, representing about 3 to 5% of all respiratory infec-tions in children, and almost all adults have antibodies to typeC adenovirus (11). Moreover, transmission of wild-type ade-novirus occurs by aerosolization, and so delivery to the airwaysshould be feasible. The pathology of adenovirus infection hasbeen studied in cotton rats (27) and in autopsy specimens (1),and viral replication in bronchial epithelia has been shown(27). Nevertheless, many questions about the biology of infec-tion with wild-type adenovirus remain. For example, the celltype involved in the initial infection is not known, the efficiencywith which wild-type virus infects airway epithelium has notbeen determined, it is not certain how many cells must beinfected to produce a productive infection, and the mecha-nisms of viral dissemination to other cells are not understood.Previous studies have begun to examine the interaction be-
tween recombinant adenoviruses and human airway epitheliain several model systems. While it seems clear that adenoviruscan infect airway cells and express transgenes, there have beenvariable results in vivo in humans (4, 16, 22, 39) and in vitro (9,14, 40). Some variability may be due to the viral constructs, thereporter, the administration procedure, and the specificity andsensitivity of methods used to assay gene transfer. Perhapseven more importantly, the cellular model that is used mayinfluence the results. The target for CF gene transfer in thesurface airway epithelium is thought to be the ciliated epithe-lial cell, which is the predominant cell type that contacts thelumen in the airways. Therefore, in this study, we used CFairway cells grown on permeable filter supports at the air-liquid interface. Previous studies have shown that under theseconditions, the cells can develop into an epithelium that retainsmany properties of native epithelium (23, 35). We first exam-
* Corresponding author. Mailing address: Howard Hughes MedicalInstitute, University of Iowa College of Medicine, 500 EMRB, IowaCity, IA 52242. Phone: (319) 335-7619. Fax: (319) 335-7623. Electronicmail address: [email protected].
6994
on June 26, 2015 by guesthttp://jvi.asm
.org/D
ownloaded from

ined the state of differentiation of the cells, especially theirability to develop a ciliated surface. Then we tested the abilityof a recombinant adenovirus to express a transgene in the cells.We studied epithelia when their differentiation state variedand when vector was applied for different durations. Becausethe electrophysiologic measurement of Cl2 transport providesa sensitive assay that has direct relevance to the biology of CF,we also used a vector encoding the CFTR Cl2 channel andassayed cyclic AMP (cAMP)-stimulated transepithelial Cl2
transport. Finally, we tested the ability of an adenovirus vectorexpressing CFTR to correct the Cl2 transport defect in CFmice.
MATERIALS AND METHODS
Airway epithelial cells. Airway epithelial cells were obtained from surgicalpolypectomies of CF and non-CF patients. Cells were isolated by enzyme diges-tion as previously described (28). Freshly isolated cells were seeded at a densityof 5 3 105 cells per cm2 onto collagen-coated permeable membranes (0.6 cm2;Millipore-Inserts). The cells were maintained at 378C in a humidified atmo-sphere of 5% CO2 and air. Twenty-four hours after plating, the mucosal mediumwas removed and the cells were allowed to grow at the air-liquid interface (23,36). The culture medium was a mixture of 49% Dulbecco modified Eagle me-dium, 49% Ham’s F12 medium, and 2% Ultraser G (Sepracor Inc., Marlbor-ough, Mass.). Penicillin (100 U/ml) and streptomycin (100 mg/ml) were added tothe medium.Virus preparation. Construction and preparation of Ad2/CFTR-3, Ad2/
CFTR-8, and Ad2/bGal-2 were similar to what we have described previously(33). The viral E1a promoter was used to express CFTR cDNA in Ad2/CFTR-3,and the cytomegalovirus promoter was used to express b-galactosidase in Ad2/bGal-2. In both vectors, genes from the E4 region were replaced with the E4ORF6 (33). Wild-type adenovirus type 2 (Ad2) was obtained from Sam Wad-sworth, Genzyme Corporation.Titers of the recombinant adenoviruses were determined by an infection assay.
Briefly, serial dilutions of the recombinant adenoviruses were plated on 293 cells.The cells were incubated for 7 days and then stained with a polyclonal fluoresceinisothiocyanate (FITC)-labeled antihexon antibody (Chemicon, Temecula, Cal-if.). This method of titration differs from that previously used (40) in that thelength of incubation is longer and titers are three- to fivefold higher than thoseobtained by the previous method.Viral infection. Epithelia were allowed to reach confluence and develop a
transepithelial electrical resistance (Rt), indicating the development of tightjunctions and an intact barrier. All epithelia had Rt values of.800V. Three to 14days after seeding, dilutions of the recombinant viruses (in phosphate-buffer-ed saline [PBS]) ranging from multiplicities of infection (MOI) of 0 to 50were added to the apical surface. Following the indicated incubation time, theviral suspension was removed and the monolayers were rinsed twice withPBS. After infection, the monolayers were incubated at 378C for an additional3 days. Rt was measured with an ohmmeter (EVOM; World Precision Instru-ment Inc., Sarasota, Fla.) before infection and was not altered by application ofvirus.To assess infectivity on 293 cells, 140 ml of 293 medium containing 104 IU of
either wild-type Ad2 or Ad2/CFTR-3 was added in quadruplicate to cells thatwere approximately 70% confluent (48 h postseeding). The virus was removed atintervals from 15 min to 24 h later and subjected to two thorough rinses with 293medium. Thirty hours after infection, the cells were fixed with 50:50 methanol-acetone at 48C for 30 min and stained with a polyclonal FITC-labeled antihexonantibody (Chemicon). The number of fluorescent cells was determined at eachtime point.Determination of b-galactosidase activity. Three days after infection, non-CF
epithelia were rinsed with serum-free medium and incubated for 5 min with0.05% trypsin–0.53 mM EDTA to release the cells from the filter. To detectb-galactosidase activity, we used the chromogenic reagent X-Gal (5-bromo-4-chloro-3-indolyl-b-D-galactopyranoside; Boehringer Mannheim) (29). Cells wereplated onto slides by using a cytocentrifuge (approximately 20,000 cells per slide),fixed with 1.8% formaldehyde–2% glutaraldehyde, and then incubated for 16 hat 378C with 0.313 ml of X-Gal (40 mg/ml) in dimethyl sulfoxide dissolved in 12.5ml of PBS (pH 7.8). Blue staining of the nuclei was assessed by light microscopy,and results are expressed as percentages of cells that were positive.We also measured total b-galactosidase activity by a commercially available
method (Galacto-Light; Tropix, Inc., Bedford, Mass.). Briefly, after being rinsedwith PBS, cells were removed from filters by incubation with 120 ml of lysis buffer(25 mM Tris-phosphate [pH 7.8], 2 mM dithiothreitol, 2 mM 1,2-diaminocyclo-hexane-N,N,N9,N9-tetraacetic acid, 10% glycerol, 1% Triton X-100) for 15 min.Two hundred microliters of 1:100 Galacton diluted in Galacto-Light reactionbuffer was incubated for 1 h with 4 ml of each sample in triplicate. Two secondsprior to the measurement, 300 ml of Galacto-Light Accelerator was added, andlight emission was quantified in a luminometer (Analytical Luminescence Lab-oratory, San Diego, Calif.).
Electron microscopy. Airway epithelia grown on permeable supports werefixed at various times after seeding in 2.5% gluteraldehyde and processed forscanning electron microscopy by using standard procedures. Briefly, the cultureswere postfixed in 1% osmium tetroxide and then dehydrated in a graded seriesof ethanol. The samples were then rinsed twice in hexamethyldisilazane, airdried, and mounted on aluminum stubs with colloidal silver paint. The filterswere sputter-coated with 60:40 gold-palladium and imaged in an Hitachi S-4000field emission scanning electron microscope. Transmission electron microscopyto detect b-galactosidase was performed by using standard techniques as previ-ously described (40).Analysis of cell cytokeratins. Three and ten days after seeding, monolayers
were fixed for 10 min with 4% paraformaldehyde, permeabilized for 10 min with0.2% Triton X-100, and then incubated for 1 h with Superblock (Pierce) to blocknonspecific antibody binding. The monolayers were then incubated with a 1:200dilution of monoclonal anti-cytokeratin 18 (CK-18) linked to FITC (Sigma cloneCY-90) or with a 1:200 dilution of anti-CK-14 (Sigma clone CKB1) for 1 h at378C. The monolayers were then rinsed with PBS. The monolayers stained withanti-CK-14 were incubated with a 1:800 dilution of donkey anti-mouse–Cy3(Jackson ImmunoResearch) for 30 min at 378C. After three rinses with PBS, themonolayers were mounted on slides in Gelmount (Biomeda) and examined byusing epifluorescence with a Leica photomicroscope.On days 3, 6, 10, and 14 after they were seeded onto filters, epithelial mono-
layers were rinsed with serum-free medium and incubated for 5 min with 0.05%trypsin–0.53 mM EDTA to release the cells from the filter. The cells were gentlycollected in suspension and fixed with 1.8% formaldehyde–2% glutaraldehydefor 5 min. The cells were spun at 1,000 rpm and resuspended in PBS at a densityof 107/ml. To assess the percentage of mature ciliated cells, aliquots of the cellsuspension were incubated with 1:100 FITC-labeled monoclonal anti-CK-18antibody or FITC-labeled antihexon antibody for 2 h at room temperature. Toassess the percentage of immature, basal nonciliated epithelial cells, aliquots ofthe cell suspension were incubated with 1:100 monoclonal anti-CK-14 antibodyor no antibody for 2 h. The cells were centrifuged and resuspended with a 1:50Texas red–anti-mouse immunoglobulin G for 1 h. Fluorescence from 10,000individual cells was determined by fluorescence-activated cell sorting (FACS)analysis (FACScan, Lysys II software; Becton Dickinson, San Jose, Calif.). Thepercentage of cells positive for CK-14 or CK-18 was assessed by determining thepercentage of highly fluorescent cells in each group and subtracting the fluores-cence of the control cells. Since both antibodies were mouse monoclonal anti-bodies, studies to evaluate colocalization of CK-14 and CK-18 were not possible.CF mice. We used mice that were homozygous for the DF508 mutation; they
were bred, identified, and housed as previously described (43). For vector ad-ministration, the mice were intubated by inserting a plastic catheter (22G Jelco;Jelco, Tampa, Fla.) through the larynx. They were ventilated with 100% O2 at apressure of 15 cm of H2O. Either Ad2/CFTR-3 or Ad2/bGal-2 (5 3 107 IU in 5ml of saline) was instilled into each nostril while the mice were ventilated. Micewere ventilated for 1 h after application of the vector. Three days after vectoraddition, they were used for X-Gal staining or for measurement of electrophysi-ologic properties.Measurement of transepithelial electrical properties. The epithelial monolay-
ers were mounted in modified Ussing chambers (Jim’s Instruments, Iowa City,Iowa) as previously described (28). Epithelia were bathed on the submucosalsurface with a Ringer’s solution containing 135 mM NaCl, 2.4 mM K2HPO4, 0.6mM KH2PO2, 1.2 mM CaCl2, 1.2 mM MgCl2, 10 mM N-2-hydroxyethylpipera-zine-N9-2-ethanesulfonic acid (HEPES; titrated to pH 7.4 with NaOH), and 10mM dextrose. The mucosal solution was identical with the exception that NaClwas replaced with 135 mM sodium gluconate. Thus, there was a transepithelialCl2 concentration gradient which was used to magnify changes in Cl2 secretion.Amiloride (10 mM) was added to the mucosal solution to inhibit Na1 channelsand thereby transepithelial Na1 transport. The cAMP agonists forskolin (10mM) and 3-isobutyl-1-methylxanthine (100 mM) were added to the mucosal andserosal solution to stimulate transepithelial Cl2 current through CFTR Cl2
channels.The transepithelial electric potential (Vt) across the nasal epithelium was
measured by using techniques similar to those previously described (43). Micewere anesthetized with an intraperitoneal injection of 2,2,2-tribromoethanol (16ml/g). A 23-gauge subcutaneous needle connected with normal saline solution toa Ag-AgCl pellet (E. W. Wright, Guilford, Conn.) was used as a referenceelectrode. The exploring electrode was a 200-mm flexible catheter filled with aRinger’s solution containing 135 mM NaCl, 2.4 mM KH2PO2, 0.6 mM K2HPO4,1.2 mM CaCl2, 1.2 mM MgCl2, and 10 mM HEPES (titrated to pH 7.4 withNaOH) and connected to an Ag-AgCl pellet. Voltage was measured with avoltmeter (Keithley Instruments Inc., Cleveland, Ohio) connected to a strip chartrecorder (Servocorder, Watanabe Instruments, Japan). The exploring catheterwas inserted 4 to 5 mm into the nostril, and the basal Vt was recorded. Once astable baseline was achieved, Ringer’s solution containing 100 mM amiloride(Merck & Co. Inc., West Point, Pa.) was perfused through the second catheterat a rate of 50 ml/min, using a syringe pump (Sage Instruments). After 5 min, theperfusing solution was changed to a solution containing 135 mM sodium glu-conate (substituted for NaCl), 10 mM terbutaline, and 100 mM amiloride. Mea-surements were made after perfusion for 5 min.
VOL. 70, 1996 ADENOVIRUS-MEDIATED GENE TRANSFER TO AIRWAY EPITHELIA 6995
on June 26, 2015 by guesthttp://jvi.asm
.org/D
ownloaded from

RESULTS
Effect of duration in culture on composition of airway epi-thelial monolayers. In vivo, the apical surface of human airwayepithelia is covered by a lawn of cilia. To determine how thesurface of primary cultures of airway epithelia relate to epi-thelia in vivo, we examined the histology of cultures grown atthe air-liquid interface. Because we were primarily interestedin the surface facing the lumen, where the virus first interactswith the epithelia, we used scanning electron microscopy toevaluate the epithelial surface.Three days after seeding, cell monolayers appeared conflu-
ent (Fig. 1A and B). The cells varied in size, and most ap-peared to have short microvilli. However, there were very fewciliated cells. Six days after seeding, most of the cells appearedto have immature cilia (Fig. 1C and D). On days 10 (Fig. 1Eand F) and 14 (Fig. 1G and H) after seeding, the surface of themonolayers was covered completely by cilia, making it difficultto identify individual cells. In some areas, there was also amor-phous material mixed in with the cilia. These data indicate thatthe airway epithelia differentiated and formed a surface cov-ered by cilia by 10 days after seeding. The appearance of thesurface was like that of the in vivo airway (2). Similar resultswere obtained with CF airway epithelia.To evaluate further the cellular composition of the mono-
layers, we stained epithelia with antibodies directed againstCK-14 or CK-18. Both basal cells and immature epithelia ex-press high levels of CK-14, whereas only mature columnarairway epithelia express CK-18 (3, 14). Three days after seed-ing, most of the cells showed high levels of immunostaining forCK-14, but very few cells expressed CK-18 (Fig. 2). By 6 daysafter seeding, most of the cells appeared to express CK-18 andstaining with anti-CK-14 antibody was much less intense.To study more quantitatively the extent of monolayer differ-
entiation, we used FACS to analyze the number of CK-14- andCK-18-producing cells in epithelia on days 3, 6, 10, and 14 afterseeding. Figure 3 show examples of histograms for CK-14- andCK-18-positive cells on days 3 and 6. Three days after seeding,most of the cells were positive for CK-14. With time, theintensity of fluorescence decreased, but most cells remainedweakly positive for CK-14. Thus, determination of the numberof CK-14-positive cells is difficult. In contrast, there was a morestriking change in CK-18 fluorescence (Fig. 3 and 4). Threedays after seeding, a small population of approximately 9% ofthe cells were highly fluorescent for CK-18 antibody. Thesecells may represent the few positive cells observed with immu-nocytochemistry and the few ciliated cells observed with scan-ning electron microscopy. By 6 days after seeding, approxi-mately 85% of the cells were positive, and more than 90% ofthe cells remained positive 10 and 14 days after seeding (Fig.4).These data indicate that shortly after seeding, primary cul-
tures of human airway epithelial cells have properties resem-bling those of basal cells. They then differentiate predomi-nately into ciliated cells, showing first immature and thenmature cilia. By 10 to 14 days after seeding, the epithelium iscovered with cilia.Effect of epithelial differentiation on gene transfer. To assess
the effect of epithelial differentiation on the ability of a recom-binant adenovirus to infect epithelia, we applied Ad2/bGal-2 atan MOI of 50 to the mucosal surface of epithelial monolayers3, 6, 10, or 14 days after seeding. After 6 h of incubation, thevector was removed and the surface was washed twice to re-move unattached vector. Figure 5 shows the b-galactosidaseactivity and the percent of cells that were positive by X-Galstaining in epithelia infected on the different days. Epithelia
infected 6 days after seeding showed significantly less b-galac-tosidase activity and fewer X-Gal-positive cells than epitheliainfected 3 days after seeding. Values of both parameters wererelatively similar for epithelia infected at time points later than6 days.Because the number of cells expressing recombinant protein
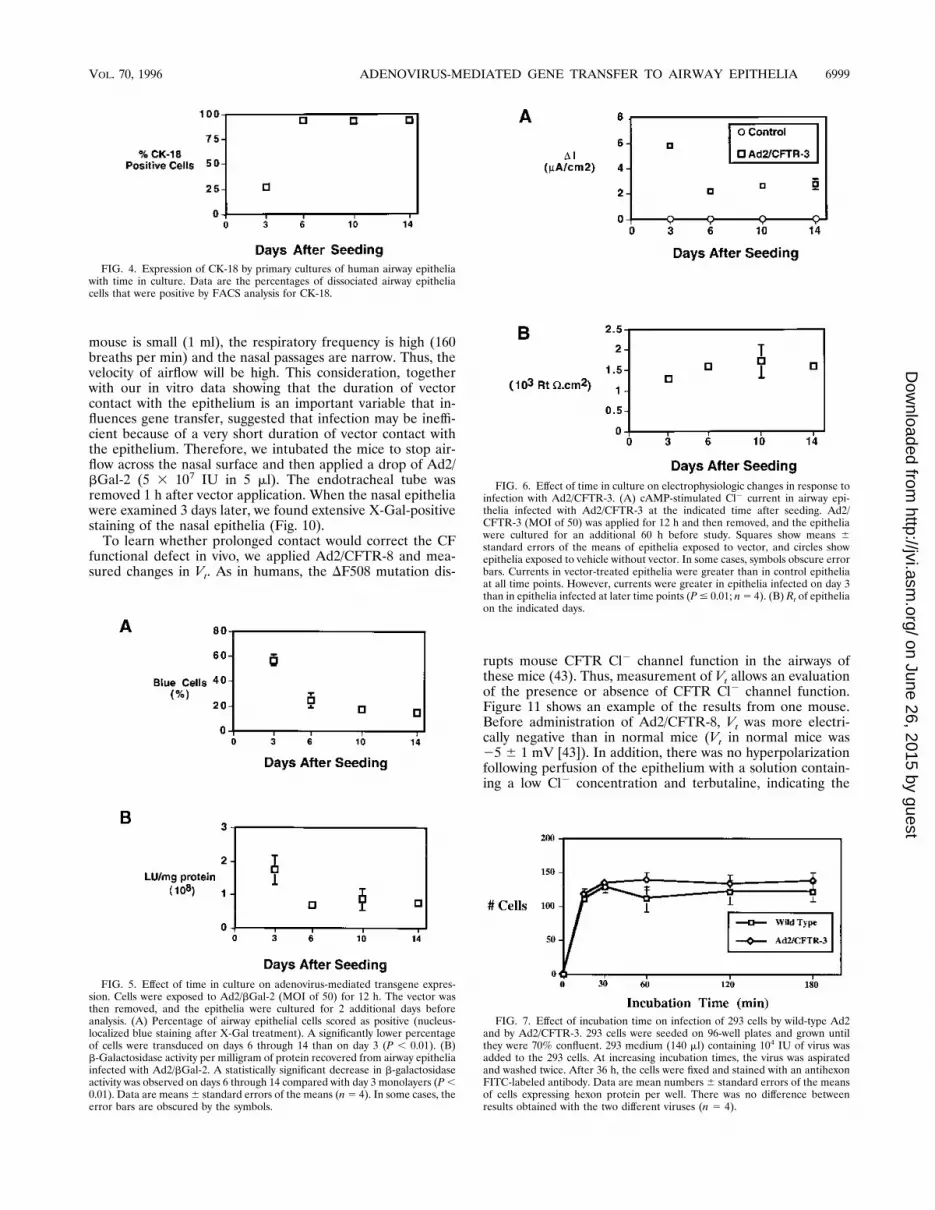
and the level of protein expression in those cells is not linearlyrelated to transepithelial Cl2 transport (19), we studied theeffect of an adenovirus expressing CFTR (Ad2/CFTR-3; MOIof 50) applied to CF airway epithelia at different stages ofdifferentiation. When we measured cAMP-stimulated Cl2 cur-rent, we found a pattern similar to that observed when weassayed b-galactosidase. Figure 6A shows that epithelia treat-ed at all time points had a significant correction of current inresponse to the cAMP agonists forskolin and IBMX. However,there was a greater Cl2 secretory current when vector wasapplied 3 days after the cells were seeded than when it wasapplied to more mature epithelia.Figure 6B shows that Rt for the epithelium was high even at
3 days after seeding and changed little over the subsequent 11days. Thus, the epithelium developed tight junctions and abarrier function before it developed a differentiated ciliatedphenotype and resistance to adenovirus infection. Thus, whiledevelopment of an Rt indicates formation of an epithelium,these results suggest that the presence of a high Rt does notnecessarily reflect the differentiated state of the epithelium,and Rt does not correlate directly with ability to infect theepithelium.These data suggest that as the epithelium differentiates and
matures, it becomes more resistant to gene transfer with re-combinant adenovirus. Both the percentage of cells expressingtransgene and the total activity resulting from transgene ex-pression (b-galactosidase activity or Cl2 secretory current)decreased as the epithelium matured. However, even 14 daysafter seeding, Ad2/CFTR-3 was able to generate a significantCl2 secretory current in CF epithelia.Effect of incubation time. The decreased ability of adenovi-
rus to infect mature compared with immature airway epitheliamay be due to the loss of a receptor required for infection. Oneway to begin to assess this possibility was to examine how theduration of contact between the vector and the epitheliumaffected transgene expression. As a preliminary study, we com-pared the abilities of a recombinant (Ad2/CFTR-3) and awild-type adenovirus to infect 293 cells. We chose 293 cellsbecause they complement the E1 region, which is missing inAd2/CFTR-3. This allowed us to compare the efficiencies ofinfection by using the same detection method (detection ofhexon production) as opposed to having to compare reporterproduction by the recombinant virus and viral protein produc-tion by the wild-type virus. We found that infection of 293 cellsplateaued by 30 min for both the E1-deleted and wild-typeadenoviruses (Fig. 7).We then examined the effects of duration of viral incubation
in immature (3 days after seeding) and mature (10 days afterseeding) epithelia. The incubation time required to infect im-mature airway epithelia appeared to have nearly plateaued by3 h (Fig. 8). In contrast, infection of mature epithelia reacheda maximum after 12 h of incubation. In addition, the absolutelevel of transgene expression was less for the mature epithelia.To learn whether the ciliated cells were expressing transgene,we used transmission electron microscopy to identify cells ex-pressing b-galactosidase. Figure 9 shows an example of elec-tron-dense particles, which are characteristic of b-galactosi-dase (25), at the nuclear membrane in a ciliated epithelial cell.These data indicate that mature ciliated airway epithelia can
be infected by an adenovirus vector. However, the prolonged
6996 ZABNER ET AL. J. VIROL.
on June 26, 2015 by guesthttp://jvi.asm
.org/D
ownloaded from

FIG. 1. Scanning electron photomicrographs of primary cultures of human airway epithelia grown at the air-liquid interface. Photomicrographs of monolayers areshown at the following times after seeding: 3 days (A and B), 6 days (C and D), 10 days (E and F), and 14 days (G and H). In panels A, C, E, and G, the scale barindicates 37.5 mm; in panels B, D, F, and H, the scale bar indicates 5 mm. On day 3, sporadic ciliated cells surrounded by undifferentiated cells with short microvillican be observed. By day 6, most of the cells have cilia (thin arrows in panel D) or immature cilia (thick arrows). By days 10 and 14, nearly all of the cells are ciliated.
6997
on June 26, 2015 by guesthttp://jvi.asm
.org/D
ownloaded from

incubation time that is required suggests that infection occursvia a different mechanism than does infection of immatureepithelia or 293 cells.Infection of nasal epithelia of CF mice. To evaluate the
relevance of these findings in an in vivo model, we used micein which the common DF508 mutation had been introducedinto the mouse CFTR gene (7, 43). We first examined the
effect of applying Ad2/bGal-2 (5 3 107 IU) to the nasal epi-thelium. We placed a drop (5 ml) of vector on the nasal epi-thelium and 3 days later removed the cells and stained themfor X-Gal. As previously reported (14), only very rare X-Gal-positive cells were observed. However, when vector is appliedto the nasal surface, it may be quickly cleared because of rapidairflow across the epithelium. Although the tidal volume of the
FIG. 2. Immunocytochemical staining of human airway epithelia for CK-14 (A and B) and CK-18 (C and D). Monolayers were studied either 3 days (A and C) or10 days (B and D) after seeding. A significant decrease in staining intensity for CK-14 was observed with time (compare panels A and B). On day 3, there were veryfew highly fluorescent cells that stained intensely for CK-18, but by day 10, all of the cells had increased levels of CK-18 fluorescence (compare panels C and D).
FIG. 3. Analysis of CK-14 (A and B)- and CK-18 (C and D)-positive cells from primary cultures of airway epithelia 3 days (A and C) or 6 days (B and D) afterseeding. (A) Histograms of dispersed cells stained with anti-CK-14 antibody followed by Texas red–anti-mouse immunoglobulin G (arrow) compared with control cellsthat were exposed only to the secondary antibody; (B) similar study for cells examined 6 days after seeding; (C) histogram of dispersed cells stained with FITC-labeledanti-CK-18 antibody compared with control cells exposed to FITC-labeled control antibody; (D) similar study of cells examined 6 days after seeding. At 3 days afterseeding, most of the cells are CK-18 negative and CK-14 positive. By 6 days after seeding, the relative intensity of CK-14 staining has decreased and most of the cellsare CK-18 positive.
6998 ZABNER ET AL. J. VIROL.
on June 26, 2015 by guesthttp://jvi.asm
.org/D
ownloaded from
mouse is small (1 ml), the respiratory frequency is high (160breaths per min) and the nasal passages are narrow. Thus, thevelocity of airflow will be high. This consideration, togetherwith our in vitro data showing that the duration of vectorcontact with the epithelium is an important variable that in-fluences gene transfer, suggested that infection may be ineffi-cient because of a very short duration of vector contact withthe epithelium. Therefore, we intubated the mice to stop air-flow across the nasal surface and then applied a drop of Ad2/bGal-2 (5 3 107 IU in 5 ml). The endotracheal tube wasremoved 1 h after vector application. When the nasal epitheliawere examined 3 days later, we found extensive X-Gal-positivestaining of the nasal epithelia (Fig. 10).To learn whether prolonged contact would correct the CF
functional defect in vivo, we applied Ad2/CFTR-8 and mea-sured changes in Vt. As in humans, the DF508 mutation dis-
rupts mouse CFTR Cl2 channel function in the airways ofthese mice (43). Thus, measurement of Vt allows an evaluationof the presence or absence of CFTR Cl2 channel function.Figure 11 shows an example of the results from one mouse.Before administration of Ad2/CFTR-8, Vt was more electri-cally negative than in normal mice (Vt in normal mice was25 6 1 mV [43]). In addition, there was no hyperpolarizationfollowing perfusion of the epithelium with a solution contain-ing a low Cl2 concentration and terbutaline, indicating the
FIG. 4. Expression of CK-18 by primary cultures of human airway epitheliawith time in culture. Data are the percentages of dissociated airway epitheliacells that were positive by FACS analysis for CK-18.
FIG. 5. Effect of time in culture on adenovirus-mediated transgene expres-sion. Cells were exposed to Ad2/bGal-2 (MOI of 50) for 12 h. The vector wasthen removed, and the epithelia were cultured for 2 additional days beforeanalysis. (A) Percentage of airway epithelial cells scored as positive (nucleus-localized blue staining after X-Gal treatment). A significantly lower percentageof cells were transduced on days 6 through 14 than on day 3 (P , 0.01). (B)b-Galactosidase activity per milligram of protein recovered from airway epitheliainfected with Ad2/bGal-2. A statistically significant decrease in b-galactosidaseactivity was observed on days 6 through 14 compared with day 3 monolayers (P,0.01). Data are means6 standard errors of the means (n5 4). In some cases, theerror bars are obscured by the symbols.
FIG. 6. Effect of time in culture on electrophysiologic changes in response toinfection with Ad2/CFTR-3. (A) cAMP-stimulated Cl2 current in airway epi-thelia infected with Ad2/CFTR-3 at the indicated time after seeding. Ad2/CFTR-3 (MOI of 50) was applied for 12 h and then removed, and the epitheliawere cultured for an additional 60 h before study. Squares show means 6standard errors of the means of epithelia exposed to vector, and circles showepithelia exposed to vehicle without vector. In some cases, symbols obscure errorbars. Currents in vector-treated epithelia were greater than in control epitheliaat all time points. However, currents were greater in epithelia infected on day 3than in epithelia infected at later time points (P# 0.01; n5 4). (B) Rt of epitheliaon the indicated days.
FIG. 7. Effect of incubation time on infection of 293 cells by wild-type Ad2and by Ad2/CFTR-3. 293 cells were seeded on 96-well plates and grown untilthey were 70% confluent. 293 medium (140 ml) containing 104 IU of virus wasadded to the 293 cells. At increasing incubation times, the virus was aspiratedand washed twice. After 36 h, the cells were fixed and stained with an antihexonFITC-labeled antibody. Data are mean numbers 6 standard errors of the meansof cells expressing hexon protein per well. There was no difference betweenresults obtained with the two different viruses (n 5 4).
VOL. 70, 1996 ADENOVIRUS-MEDIATED GENE TRANSFER TO AIRWAY EPITHELIA 6999
on June 26, 2015 by guesthttp://jvi.asm
.org/D
ownloaded from

absence of CFTR Cl2 channels (43). (Hyperpolarization ofVt with this maneuver is due to a Cl
2 diffusion voltage gener-ated through apical CFTR Cl2 channels). Three days afteradministration of Ad2/CFTR-8 to the intubated mouse, basalVt had decreased and Vt had hyperpolarized in response toperfusion with a solution containing a low Cl2 concentrationand terbutaline. Figure 12 shows average values of the re-sponse to Ad2/CFTR-8. When vector was administered to in-tubated mice, basal Vt (24.3 6 0.7 mV) and the response to
the low Cl2 solution (213.1 6 1 mV) were in the rangeobserved in normal mice (43). In contrast, application of Ad2/CFTR-8 without intubation had no effect on basal Vt (222.3 61.2 mV) or on the response to perfusion with the low Cl2
solution (20.5 6 1.4 mV).
DISCUSSION
The efficiency of gene transfer clearly depends on the modelsystem and the experimental methods used. Previous work hadshown that when human airway epithelia were cultured onpermeable filter supports at the air-liquid interface, they gen-erated an epithelium with some properties similar to those ofnative airway epithelia (15). Here we have shown that underthe culture conditions that we used, the apical surface is cov-ered with a mat of cilia; this is the site at which the vector firstencounters and must interact with the epithelium. The processof differentiation and the development of a ciliated surfacetakes several days in culture and is paralleled by a change inthe pattern of cytokeratin production. Thus, epithelia culturedin this way should provide a good model for assessing genetransfer to human airways. Our data also indicate that thenasal epithelia of CF mice may be useful for evaluating genetransfer. We obtained similar results with both models; ade-novirus-mediated gene transfer was inefficient, but the ineffi-
FIG. 8. Effect on b-galactosidase activity of increasing the duration for whichvector (Ad2/bGal-2) was in contact with the epithelium. Epithelia were exposedto vector either 3 days or 10 days after seeding.
FIG. 9. Transmission electron photomicrograph of ciliated airway epithelium. Epithelia were exposed to Ad2/bGal-2 (MOI of 50 for 12 h) 10 days after cells wereseeded onto permeable supports. Epithelia were fixed and prepared for microscopy 3 days after exposure to the vector. The cell is identified as ciliated by the presenceof ciliary basal body (small arrow) and cilia above the apical surface. Electron-dense particles at the nuclear membrane are b-galactosidase product (large arrows). Fourof the nine ciliated cells examined by transmission electron microscopy (cells in which the nuclei and cilia could be clearly matched) showed electron-dense particlesat the nuclear membrane. The bar indicates 1 mm.
7000 ZABNER ET AL. J. VIROL.
on June 26, 2015 by guesthttp://jvi.asm
.org/D
ownloaded from

ciency could be at least partly overcome by increasing theincubation time. Other useful models for the study of genetransfer include human airway epithelia grown in xenografts innude mice (8), explants of airway epithelia from humans oranimals (14), and the lungs of animals studied in vivo. Theapplicability to human airway in vivo is less clear for modelssuch as airway epithelia grown on impermeable supports andimmortalized airway cells.These results show that as airway epithelia differentiate and
develop a ciliated surface, they are less readily infected byadenovirus vectors. The difficulty in effecting gene transfer toour in vitro model corresponds to results obtained in vivo:Grubb et al. (14) reported that ciliated airway epithelia in miceare resistant to adenovirus-mediated gene transfer; we showedthat application of an adenovirus vector to the nasal epitheliumof mice was not able to correct the CF electrophysiologicabnormalities, and as discussed below, adenovirus-mediatedgene transfer to human airway epithelia in vivo is inefficient.Thus, the in vitro model of mature airway epithelia seems to bea good model with which to investigate adenovirus-mediatedgene transfer. However, it is possible that additional barriersmay limit gene transfer in vivo. For example, mucus may pro-vide a physical barrier, and mucociliary transport may movethe vector away from the epithelium. Hence, it is difficult tocompare directly the absolute efficiencies in vitro and in vivo.Nevertheless, we were able to overcome the resistance to genetransfer both in vitro and in vivo by increasing the duration ofexposure. These results and the fact that we obtained similarresults with vectors containing two different promoters (Ad2/bGal-2 has the cytomegalovirus promoter and Ad2/CFTR-3has the E1a promoter) suggest that the limited effectiveness isdue to limited virus entry into the ciliated airway cells. Inter-estingly, adenovirus is also inefficient in cultured smooth mus-cle cells, and the deficiency can be partly overcome with pro-longed incubation times (24).It seems surprising that the adenovirus vector did not have
greater efficacy at infecting the epithelia, because airway cellsare thought to be a target of wild-type adenovirus (17). Be-cause the E1-deleted virus appeared to be as efficacious as thewild type at infecting 293 cells, the inefficiency is not likely due
to the fact that a recombinant virus is used. Moreover, moststudies of adenovirus infection have used model cell lines suchas HeLa and 293 cells, often studied in suspension. Theseconsiderations raise important questions about the nature ofnaturally occurring infections of respiratory epithelia by ade-novirus. How do type C adenovirus infect airway epithelia tocomplete their life cycle? Do they require preexisting damageto the epithelia? What cell type is the initial target in a wild-type infection? Is infection of a susceptible cell type followedby viremia and/or local spread of the infection? Future studieswill be needed to address these interesting questions and toprovide further understanding of in vivo infection.The data suggest that the apical surface of mature airway
epithelia may not have the receptors required for efficientinfection. Infection by adenovirus is most effective in cells withreceptors for both the adenovirus fiber and the penton base. InHeLa cells, a high-affinity fiber receptor is believed to bepresent at a density of around 5,000 to 10,000 per cell (26). Thereceptor for fiber has not been identified but is believed to be
FIG. 10. Ciliated nasal airway epithelia form a CF mouse. The photomicro-graph shows a frozen section of nasal mucosa fixed and stained in situ withX-Gal. Blue-stained nuclei identify cells expressing transgene. Ad2/bGal-2 (5 3107 IU) was applied to the nostril of an intubated mouse 3 days before removalof the tissue.
FIG. 11. Vt of a DF508 homozygous mouse before (A) and 3 days after (B)application of Ad2/CFTR-8 (5 3 107 IU). Vector was applied while the mousewas intubated. Bars indicate application of 100 mM amiloride or a solutioncontaining 10 mM terbutaline and a low Cl2 concentration.
FIG. 12. Vt of DF508 homozygous mice before and after administration ofAd2/CFTR-8 or Ad2/bGal-2. Vector (5 3 107 IU) was applied while mice wereintubated or without intubation (Sniffed). (A) Basal voltage before vector appli-cation (day 0) or 3 days after application of Ad2/CFTR-8 or Ad2/bGal-2, asindicated; (B) change in Vt produced by perfusion with a solution containing 10mM terbutaline and a low Cl2 concentration in the presence of 100 mM amilo-ride. Data are means 6 standard errors of the means (n 5 9 for day 0; n 5 12for animals treated with Ad2/CFTR-8; n 5 6 for animals that “sniffed” Ad2/CFTR-8; n 5 6 for animals treated with Ad2/bGal-2). Asterisks indicate P ,0.001 compared with day 0 value.
VOL. 70, 1996 ADENOVIRUS-MEDIATED GENE TRANSFER TO AIRWAY EPITHELIA 7001
on June 26, 2015 by guesthttp://jvi.asm
.org/D
ownloaded from

different for different serotypes of adenoviruses (6, 10, 32). Thereceptors for penton base are cell surface integrins (34). InHeLa and 293 cells, the penton base binds aV integrins, whichmediate internalization (13, 34). Attempts to correlate thepresence of aV integrins with the efficiency of adenovirus-mediated gene transfer have yielded different results (12, 31).Human airway epithelia express aVb5 integrin on basal cellsand on the basolateral surface of columnar cells (5), but thesignificance for adenovirus infection is uncertain. We do notthink that the cilia per se are a physical barrier to infection,because the epithelia developed a resistance to infection by day6, when a significant number of cells either lacked cilia or hadonly cilia progenitors, and infection was similar on days 6 andon days 10 to 14, when nearly all cells were ciliated.How do these results relate to previous studies with hu-
mans? In our first study of gene transfer to the nasal epithe-lium of patients with CF, we found that low input MOIs of anadenovirus vector expressing CFTR partially corrected theelectrophysiologic abnormalities in the nasal epithelium (39).In a second study with humans, we administered vector in aless invasive manner to ciliated airway epithelia on the inferiorsurface of the inferior turbinate in the nose. Despite usinghigher input doses, we found less correction of the CF elec-trophysiologic abnormalities (42). Two other studies in whichan adenovirus vector was applied to nasal epithelium of pa-tients with CF yielded variable results. Hay et al. (16) foundsignificant correction of the CF electrophysiologic abnormali-ties, whereas Knowles et al. (22) found no evidence of elec-trophysiologic correction. Our present data suggest that thevariable results between our first and second studies and vari-ation between results in other studies could have been influ-enced by several factors. First, we reported that the applicatorand vasoconstrictor (cocaine) that we used in our first studydamaged the epithelium (39). Such damage may have exposedmore basal cells and increased the efficiency of gene transfer.Second, the anesthetics that we used in our first study (cocaineand lidocaine) are known to produce ciliostasis (18) and mayhave increased the contact time of the virus with the epithe-lium. Third, in our first study, we applied vector to the medialaspect of the inferior turbinate, which consists primarily ofcuboidal and squamous nonciliated epithelium, which may bemore susceptible to adenovirus infection than is a ciliatedepithelium. These variables may have accounted, in part, formore gene transfer in our first study than in the second study.Similar variation in the methods of vector delivery may haveaccounted for variable results in other studies of humans (16,22).These results suggest that several considerations should be
explored in adenovirus-mediated gene transfer to airway epi-thelia. Because infection increases as the duration of contactwith the epithelium increases, attempts to transiently inhibitciliary clearance could be considered, as well as slow instilla-tion over a prolonged time period. The presence or absence ofspecific receptors for adenovirus on the apical membraneneeds to be determined. It may also be possible to target adifferent serotype of adenovirus to the apical surface of airwayepithelia. Finally, it might be possible to target adenovirus to anew receptor, in a manner analogous to what has been donewith retroviruses (21).
ACKNOWLEDGMENTS
We thank Pary Weber, Tom Moninger, Aurita Puga, Phil Karp, andTheresa Mayhew for excellent assistance. We especially appreciate thehelp of Mary Schroth. We thank Alan E. Smith and Sam Wadsworth,Genzyme (Framingham, Mass.), for the gift of Ad2/CFTR-3, Ad2/CFTR-8, and Ad2/bGal-2.
We appreciate the support of the University of Iowa Gene TransferVector Core. This work was supported by the National Heart Lung andBlood Institute, the Roy J. Carver Charitable Trust, and the CysticFibrosis Foundation. J.Z. is a Parker B. Francis Fellow. M.J.W. is anInvestigator of the Howard Hughes Medical Institute.
REFERENCES
1. Becroft, D. M. O. 1967. Histopathology of fatal adenovirus infection of therespiratory tract in young children. J. Clin. Pathol. 20:561–569.
2. Breeze, R. G., and R. B. Wheeldon. 1977. The cells of the pulmonary airways.Am. Rev. Respir. Dis. 116:705–777.
3. Broers, J. L. V., L. de Leij, M. K. Rot, A. ter Haar, E. B. Lane, I. M. Leigh,S. S. Wagenaar, G. P. Vooijs, and F. C. S. Ramaekers. 1989. Expression ofintermediate filament proteins in fetal and adult human lung tissues. Dif-ferentiation 40:119–128.
4. Crystal, R. G., N. G. McElvaney, M. A. Rosenfeld, C. S. Chu, A. Mastrangeli,J. G. Hay, S. L. Brody, H. A. Jaffe, N. T. Eissa, and C. Danel. 1994. Admin-istration of an adenovirus containing the human CFTR cDNA to the respi-ratory tract of individuals with cystic fibrosis. Nat. Genet. 8:42–51.
5. Damjanovich, L., S. M. Albelda, S. A. Mette, and C. A. Buck. 1992. Distri-bution of integrin cell adhesion receptors in normal and malignant lungtissue. Am. J. Respir. Cell Mol. Biol. 6:197–206.
6. Defer, C., M. T. Belin, M. L. Caillet-Boudin, and P. Boulanger. 1990. Humanadenovirus-host cell interactions: comparative study with members of sub-groups b and c. J. Virol. 64:3661–3673.
7. Deng, C., K. R. Thomas, and M. R. Capecchi. 1993. Location of crossoversduring gene targeting with insertion and replacement vectors. Mol. Cell.Biol. 13:2134–2140.
8. Engelhardt, J. F., E. D. Allen, and J. M. Wilson. 1991. Reconstitution oftracheal grafts with a genetically modified epithelium. Proc. Natl. Acad. Sci.USA 88:11192–11196.
9. Engelhardt, J. F., Y. Yang, L. D. Stratford-Perricaudet, E. D. Allen, K.Kozarsky, M. Perricaudet, J. R. Yankaskas, and J. M. Wilson. 1993. Directgene transfer of human CFTR into human bronchial epithelia of xenograftswith E1-deleted adenoviruses. Nat. Genet. 4:27–34.
10. Gall, J., A. Kass-Eisler, L. Leinwand, and E. Falck-Pedersen. 1996. Adeno-virus type 5 and 7 capsid chimera: fiber replacement alters receptor tropismwithout affecting primary immune neutralization epitopes. J. Virol. 70:2116–2123.
11. Ginsberg, H. S. 1984. The adenoviruses. Plenum Press, New York.12. Goldman, M. J., and J. M. Wilson. 1995. Expression of aVb5 integrin is
necessary for efficient adenovirus-mediated gene transfer in the human air-way. J. Virol. 69:5951–5958.
13. Greber, U. F., M. Willetts, P. Webster, and A. Helenius. 1993. Stepwisedismantling of adenovirus 2 during entry into cells. Cell 75:477–486.
14. Grubb, B. R., R. J. Pickles, H. Ye, J. R. Yankaskas, R. N. Vick, J. F.Engelhardt, J. M. Wilson, L. G. Johnson, and R. C. Boucher. 1994. Ineffi-cient gene transfer by adenovirus vector to cystic fibrosis airway epithelia ofmice and humans. Nature (London) 371:802–806.
15. Gruenert, D. C., W. E. Finkbeiner, and J. H. Widdicombe. 1995. Culture andtransformation of human airway epithelial cells. Am. J. Physiol. 268:L347–L360.
16. Hay, J. G., N. G. McElvaney, J. Herena, and R. G. Crystal. 1995. Modifica-tion of nasal epithelial potential differences of individuals with cystic fibrosisconsequent to local administration of a normal CFTR cDNA adenovirusgene transfer vector. Hum. Gene Ther. 6:1487–1496.
17. Horwitz, M. S. 1990. Adenoviridae and their replication, p. 1679–1721. InB. N. Fields, D. M. Knipe, et al. (ed.), Virology. Raven Press, New York.
18. Ingels, K. J., M. R. Nijziel, K. Graamans, and E. H. Huizing. 1994. Influenceof cocaine and lidocaine on human nasal cilia. Beat frequency and harmonyin vitro. Arch. Otolaryngol. Head Neck Surg. 120:197–201.
19. Johnson, L. G., J. C. Olsen, B. Sarkadi, K. L. Moore, R. Swanstrom, andR. C. Boucher. 1992. Efficiency of gene transfer for restoration of normalairway epithelial function in cystic fibrosis. Nat. Genet. 2:21–25.
20. Kaplan, J. M., J. A. St.George, S. E. Pennington, L. D. Keyes, P. Johnson,S. C. Wadsworth, and A. E. Smith. 1995. Humoral and cellular immuneresponses of non-human primates to long term repeated lung exposure toAd2/CFTR-2. Gene Ther. 3:117–127.
21. Kasahara, N., A. M. Dozy, and Y. W. Kan. 1994. Tissue-specific targeting ofretroviral vectors through ligand-receptor interactions. Science 266:1373–1376.
22. Knowles, M. R., K. W. Hohneker, Z. Zhou, J. C. Olsen, T. L. Noah, P. C. Hu,M. W. Leigh, J. F. Engelhardt, L. J. Edwards, K. R. Jones, et al. 1995. Acontrolled study of adenoviral-vector-mediated gene transfer in the nasalepithelium of patients with cystic fibrosis. New Engl. J. Med. 333:823–831.
23. Kondo, M., W. E. Finkbeiner, and J. H. Widdicombe. 1991. Simple techniquefor culture of highly differentiated cells from dog tracheal epithelium. Am. J.Physiol. 261:L106–L117.
24. March, J. K. L., J. E. Madison, and B. C. Trapnell. 1995. Pharmacokineticsof adenoviral vector-mediated gene delivery to vascular smooth muscle cells:modulation by poloxamer 407 and implications for cardiovascular gene ther-
7002 ZABNER ET AL. J. VIROL.
on June 26, 2015 by guesthttp://jvi.asm
.org/D
ownloaded from

apy. Hum. Gene Ther. 6:41–53.25. Messina, L. M., R. M. Podrazik, T. A. Whitehill, D. Ekhterae, T. E. Brothers,
J. M. Wilson, W. E. Burkel, and J. C. Stanley. 1992. Adhesion and incor-poration of lacZ-transduced endothelial cells into the intact capillary wall inthe rat. Proc. Natl. Acad. Sci. USA 89:12018–12022.
26. Persson, R., C. Wohlfart, U. Svensson, and E. Everitt. 1985. Virus-receptorinteraction in the adenovirus system: characterization of the positive coop-erative binding of virions on hela cells. J. Virol. 54:92–97.
27. Prince, G. A., D. D. Porter, A. B. Jenson, R. L. Horswood, R. M. Chanock,and H. S. Ginsberg. 1993. Pathogenesis of adenovirus type 5 pneumonia incotton rats (Sigmodon hispidus). J. Virol. 67:101–111.
28. Rich, D. P., L. A. Couture, L. M. Cardoza, V. M. Guiggio, D. Armentano,P. C. Espino, K. Hehir, M. J. Welsh, A. E. Smith, and R. J. Gregory. 1993.Development and analysis of recombinant adenoviruses for gene therapy ofcystic fibrosis. Hum. Gene Ther. 4:461–476.
29. Sanes, J. R., J. L. Rubenstein, and J. F. Nicolas. 1986. Use of a recombinantretrovirus to study post-implantation cell lineage in mouse embryos. EMBOJ. 5:3133–3142.
30. Simon, R. H., J. F. Engelhardt, Y. Yang, M. Zepeda, S. Weber-Pendleton, M.Grossman, and J. M. Wilson. 1993. Adenovirus-mediated transfer of theCFTR gene to lung of nonhuman primates: toxicity study. Hum. Gene Ther.4:771–780.
31. Smythe, W. R., H. C. Hwang, A. A. Elshami, K. M. Amin, S. M. Albelda, andL. R. Kaiser. 1995. Differential sensitivity of thoracic malignant tumors toadenovirus-mediated drug sensitization gene therapy. J. Thorac. Cardiovasc.Surg. 109:626–630.
32. Stevenson, S. C., M. Rollence, B. White, L. Weaver, and A. McCleeland.1995. Human adenovirus serotypes 3 and 5 bind to two different cellularreceptors via the fiber head domain. J. Virol. 69:2850–2857.
33. Welsh, M. J., J. Zabner, S. M. Graham, A. E. Smith, R. Moscicki, and S. C.Wadsworth. 1995. Adenovirus-mediated gene transfer for cystic fibrosis. PartA. Safety of dose and repeat administration in the nasal epithelium. Part B.Clinical efficacy in the maxillary sinus. Hum. Gene Ther. 6:205–218.
34. Wickham, T. J., E. J. Filardo, D. A. Cheresh, and G. R. Nemerow. 1994.
Integrin alpha v beta 5 selectively promotes adenovirus mediated cell mem-brane permeabilization. J. Cell Biol. 127:257–264.
35. Widdicombe, J. H. 1990. Use of cultured airway epithelial cells in studies ofion transport. Am. J. Physiol. 258:L13–L18.
36. Yamaya, M., W. E. Finkbeiner, S. Y. Chun, and J. H. Widdicombe. 1992.Differentiated structure and function of cultures from human tracheal epi-thelium. Am. J. Physiol. 262:L713–L724.
37. Yang, Y., F. A. Nunes, K. Berencsi, E. E. Furth, E. Gonczol, and J. M.Wilson. 1994. Cellular immunity to viral antigens limits E1-deleted adeno-viruses for gene therapy. Proc. Natl. Acad. Sci. USA 91:4407–4411.
38. Yei, S., N. Mittereder, K. Tang, C. O’Sullivan, and B. C. Trapnell. 1994.Adenovirus-mediated gene transfer for cystic fibrosis: quantitative evalua-tion of repeated in vivo vector administration to the lung. Gene Ther.1:192–200.
39. Zabner, J., L. A. Couture, R. J. Gregory, S. M. Graham, A. E. Smith, andM. J. Welsh. 1993. Adenovirus-mediated gene transfer transiently correctsthe chloride transport defect in nasal epithelia of patients with cystic fibrosis.Cell 75:207–216.
40. Zabner, J., L. A. Couture, A. E. Smith, and M. J. Welsh. 1994. Correction ofcAMP-stimulated fluid secretion in cystic fibrosis airway epithelia: efficiencyof adenovirus-mediated gene transfer in vitro. Hum. Gene Ther. 5:585–593.
41. Zabner, J., D. M. Petersen, A. P. Puga, S. M. Graham, L. A. Couture, L. D.Keyes, M. J. Lukason, J. A. St George, R. J. Gregory, A. E. Smith, and M. J.Welsh. 1994. Safety and efficacy of repetitive adenovirus-mediated transferof CFTR cDNA to airway epithelia of primates and cotton rats. Nat. Genet.6:75–83.
42. Zabner, J., B. W. Ramsey, D. P. Meeker, M. I. Aitken, R. P. Balfour, R. L.Gibson, J. Launspach, R. A. Moscicki, S. M. Richards, T. A. Standaert, J.Williams-Warren, S. C. Wadsworth, A. E. Smith, and M. J. Welsh. 1996.Repeat administration of an adenovirus vector encoding CFTR to the nasalepithelium of patients with cystic fibrosis. J. Clin. Invest. 97:1504–1511.
43. Zeiher, B. G., E. Eichwald, J. Zabner, J. J. Smith, A. P. Puga, P. B. McCray,M. R. Capecchi, M. J. Welsh, and K. R. Thomas. 1995. A mouse model forthe DF508 allele of cystic fibrosis. J. Clin. Invest. 96:2051–2064.
VOL. 70, 1996 ADENOVIRUS-MEDIATED GENE TRANSFER TO AIRWAY EPITHELIA 7003
on June 26, 2015 by guesthttp://jvi.asm
.org/D
ownloaded from





























