Embed Size (px)
Citation preview

A Role for the Endocannabinoid System in Exercise-Induced SpatialMemory Enhancement in Mice
Talita H. Ferreira-Vieira,1 Cristiane P. Bastos,1 Grace S. Pereira,1
Fabricio A. Moreira,2 and Andr�e R. Massensini1*
ABSTRACT: It is well known that physical exercise has positive effects oncognitive functions and hippocampal plasticity. However, the underlying mech-anisms have remained to be further investigated. Here we investigated thehypothesis that the memory-enhancement promoted by physical exercise relieson facilitation of the endocannabinoid system. We observed that the spatialmemory tested in the object location paradigm did not persist in sedentarymice, but could be improved by 1 week of treadmill running. In addition, exer-cise up-regulated CB1 receptor and BDNF expression in the hippocampus. Toverify if these changes required CB1 activation, we treated the mice with theselective antagonist, AM251, before each period of physical activity. In linewith our hypothesis, this drug prevented the exercise-induced memoryenhancement and BDNF expression. Furthermore, AM251 reduced CB1 expres-sion. To test if facilitating the endocannabinoid system signaling would mimicthe alterations observed after exercise, we treated sedentary animals during 1week with the anandamide-hydrolysis inhibitor, URB597. Mice treated withthis drug recognized the object in a new location and have increased levels ofCB1 and BDNF expression in the hippocampus, showing that potentiating theendocanabinoid system equally benefits memory. In conclusion, the favorableeffects of exercise upon spatial memory and BDNF expression depend on facili-tation of CB1 receptor signaling, which can be mimic by inhibition of ananda-mide hydrolysis in sedentary animals. Our results suggest that, at least in part,the promnesic effect of the exercise is dependent of CB1 receptor activationand is mediated by BDNF. VC 2013 Wiley Periodicals, Inc.
KEY WORDS: fatty acid amide hydrolase; cannabinoid receptor;AM251; hippocampus; treadmill
INTRODUCTION
The herb Cannabis sativa (hemp) has been consumed for centuriesdue to its well known psychoactive effects, mediated mainly by D9-tetra-
hydrocannabinol (D9-THC). Extensive studies aroundthe mechanism underlying the actions of D9-THCand its synthetic derivate (cannabinoids) led to thediscovery of the endogenous cannabinoid system ofthe brain (Bisogno et al., 2005; Wang and Ueda,2009). This system comprises at least two endogenousligands (endocannabinoids), arachidonoyl ethanola-mide (also termed anandamide; Devane et al., 1992)and 2-arachidonoyl glycerol (2-AG; Mechoulam et al.,1995); the biochemical processes for their synthesisand inactivation, mediated by the enzymes fatty acidamide hydrolase (FAAH) and mono-acyl glycerollipase, respectively (Piomelli, 2003; Di Marzo, 2008);and the cannabinoid (CB) receptors, which includeCB1, CB2, and possibly others (Di Marzo et al.,2004; Piomelli, 2003; Di Marzo, 2008). CB2 recep-tors are often found in immune cells and usuallyrelated to inflammatory responses (Munro et al.,1993; Galiegue et al., 1995). On the other hand, CB1
is the most abundant G-protein coupled receptor ofthe mammalian central nervous system (Matsudaet al., 1990) and seems to mediate the majority ofcentral actions of cannabinoids (Cabral andMarciano-Cabral, 2005; Di Marzo, 2009).
Given the high density of CB1 receptors in the hip-pocampus (Matsuda et al., 1990; Davies et al., 2002)and its role in learning and memory (Broadbentet al., 2004) it is not surprising that this brain regionis strongly modulated by exo- and endo-cannabinoids(Wise et al., 2009b). While marijuana and syntheticCB1 agonists are well known for their amnestic effects(Egashira et al., 2002; Suenaga et al., 2008), somestudies have indicated that increasing the endogenouslevels of anandamide (FAAH inhibition or deletion)might enhance cognitive functions. For instance,FAAH knockout mice exhibit enhanced acquisition ofan aversive, but not of an appetitive, Barnes maze task(Wise et al., 2009a). Moreover, the FAAH inhibitor,URB597, had a significant enhancing effect on mem-ory acquisition using a passive-avoidance task (Maz-zola et al., 2009). Similarly, genetic deletion orpharmacological inhibition of FAAH facilitates theacquisition and extinction of a spatial memory task(Varvel et al., 2007). Nevertheless, the role of theendocannabinoids in cognition, especially in memoryimprovement, has not been well established andremains controversial (Panlilio et al., 2013).
1 Department of Physiology and Biophysics, Institute of Biological Scien-ces, Universidade Federal de Minas Gerais, Belo Horizonte, MG, Brazil;2 Department of Pharmacology, Institute of Biological Sciences, Univer-sidade Federal de Minas Gerais, Belo Horizonte, MG, BrazilGrant sponsors: CNPq, FAPEMIG, CAPES, and Pr�o Reitoria de Pesquisa/UFMG.*Correspondence to: Andr�e Ricardo Massensini, Ph.D., N�ucleo de Neuro-ciencias, Departamento de Fisiologia e Biof�ısica, Instituto de Ciencias Biol-gicas, Universidade Federal de Minas Gerais, Av. Antonio Carlos 6627,31270-901, Belo Horizonte, MG, Brazil. E-mail: [email protected] (or)Fabricio A. Moreira, Ph.D., Laborat�orio de Neuropsicofarmacologia, Depar-tamento de Farmacologia, Instituto de Ciencias Biol�ogicas, UniversidadeFederal de Minas Gerais, Av. Antonio Carlos 6627, 31270-901, Belo Hori-zonte, MG, Brazil. E-mail: [email protected] for publication 9 September 2013DOI 10.1002/hipo.22206Published online 18 October 2013 in Wiley Online Library(wileyonlinelibrary.com).
VC 2013 WILEY PERIODICALS, INC.
HIPPOCAMPUS 24:79–88 (2014)

Accumulating evidence indicates that several factors, such asstress (Fuchs and Flugge, 1998; Mitra et al., 2006), environ-mental enrichment (van Praag et al., 2000), or physical exercise(Bekinschtein et al., 2011), can induce plastic changes in thehippocampus and thus interfere with cognitive functions. Atthe molecular level, physical exercise increases the expression ofneurotrophins, including the brain derived neurotrophic factor(BDNF), in the hippocampus (Neeper et al., 1996; Johnsonet al., 2003; Soya et al., 2007). It also promotes cell prolifera-tion and neurogenesis (van Praag et al., 1999; Olson et al.,2006; Bekinschtein et al., 2011) and facilitates the occurrenceof long-term potentiation (LTP) of synaptic activity (van Praaget al., 1999; O’Callaghan et al., 2007). Physical exercise alsoimproves behavioral performances in several tasks (Vaynmanand Gomez-Pinilla, 2005; Hillman et al., 2008), includingmemory tests (Clarkson-Smith and Hartley, 1989; Suijo et al.,2012). Our group, for instance, demonstrated that rats under-going exercise training for 8 weeks shows improvement inshort-term spatial memory when compared to sedentary ani-mals (Drumond et al., 2012). Similarly, treadmill training for7 days enhances object recognition memory as well as long-term potentiation in dentate gyrus (O’Callaghan et al., 2007).This protocol also increases the expression of BDNF in thehippocampus and perirhinal cortex, suggesting that 7 days ofexercise are sufficient to induce plastic changes in the hippo-campus and promote cognitive improvement (Griffin et al.,2009).
Despite this, the molecular mechanisms by which exercisemodifies the brain and behavior have not been well defined.Evidence has emerged suggesting that the endocannabinoid sys-tem may, at least in part, mediate the neurobiological effectsprovided by the exercise. Sparling et al. (2003) evaluated vol-unteers who exercised on a treadmill or stationary bike for 6months. They demonstrated that individuals who performedphysical activity, regardless of the sport, had higher levels ofanandamide in the bloodstream when compared to sedentaryindividuals (Sparling et al., 2003). In rodents, 8 days of exer-cise is sufficient to increase the levels of anandamide (Hillet al., 2010). Recently, Raichlen et al. (2013) also examinedcirculating levels of endocannabinoids in runners. They showedthat endocannabinoid signaling is intensity dependent so thatthe increase circulating anandamide occur only at moderatesintensities (Raichlen et al., 2013). Another study found similarresults after evaluating 11 cyclists, in addition to demonstrate apositive correlation between anandamide and BDNF concen-tration (Heyman et al., 2012). These findings suggest thatendogenous activation of the endocannabinoid system, duringphysical exercise, might be one of the factors involved inimprovement of cognitive functions after exercise. Thus, itseems plausible to speculate that the endocannabinoid systemcould actually facilitate some cellular events related to memoryenhancement.
Consistent with this hypothesis, hippocampal cell prolifera-tion and neurogenesis induced by voluntary exercise depend onCB1 receptor signaling (Hill et al., 2010; Wolf et al., 2010).The endocannabinoid system seems also to be crucial for the
induction of learning and activity-dependent synaptic plasticity(Madronal et al., 2012). The mechanism though which CB1
receptors influence neuronal plasticity appears to involveBDNF (Aso et al., 2008).
Considering these evidences, this study was designed to testthe hypothesis that endocannabinoid signaling mediates thepromnesic effect of physical exercise. To address this question,spatial memory, hippocampal CB1 receptors and BDNF expres-sion were evaluated in mice after treadmill running. Further-more, we also investigated whether the memory-improvementpromoted by this protocol would be prevented by CB1 recep-tor antagonism. Finally, we tested whether the effects of exer-cise would be mimicked by anandamide-hydrolysis inhibitionin sedentary animals.
MATERIALS AND METHODS
Subjects
Male Swiss mice (7–8 weeks of age) supplied by the Centrode Bioterismo da Universidade Federal de Minas Gerais(CEBIO-UFMG vivarium) were housed under controlled envi-ronmental conditions (temperature 26 6 1�C and humidity 40–50%) with a 12:12-h light–dark cycle and free access to foodand water. All experiments were conducted under the ProtocolLicense no 09/2010 and approved by the University’s EthicsCommittee for Animal Experimentation (CETEA–UFMG). Allefforts were made to minimize the number of animals used andtheir suffering. The CETEA directives are in compliance withNational Institute of Health Guide for Care and Use of Labora-tory Animals (National Research Council, 1985).
Training Protocol
Seventy-two mice were familiarized to motorized treadmillfor three consecutive days for 5 min/day at speed of 6 m/minand 5� of inclination. Subsequently, the animals were randomlydivided into two groups (sedentary or trained, n 5 36 pergroup), and then subdivided into three subgroups accordinglyto treatment (vehicle, AM251, or URB597, n 5 12/subgroup)that the animals received. The exercise protocol consisted of 7consecutive days of forced running at 11 m/min, 1 session/day,for 30 min, as modification in protocol proposed by O’Calla-ghan et al. (2007). Sedentary mice were placed on treadmillfor 5 min/day at a speed of 5 m/min. The protocol of forcedphysical activity was the first choice by reason of mice are lesswilling to run after blockage of endocannabinoid signaling(Dubreucq et al., 2010; Chaouloff et al., 2011). Thus, sincewe aimed to study how CB1 receptor would interfere with theeffects of exercise on memory acquisition, voluntary exercise(e.g., wheel running) would not be a good option. In this pro-tocol, the total running distance of each animal was carefullycontrolled, as explained above. Nevertheless, our protocol doesnot involve punishing stimuli to induce running.
80 FERREIRA-VIEIRA ET AL.
Hippocampus

Drug Administration
Animals received daily injections of the AM251 (Tocris Bio-sciences, Bristol, UK), CB1 receptor antagonist (1 mg/kg),URB597 (Cayman Chemical, Ann Arbor), Fatty Acid AmideHydrolase inhibitor (0.5 mg/kg), or vehicle (1:1:18 solution ofethanol; cremophor EL; sterile saline 0.9%; Patel and Hillard,2006; Moreira et al., 2008), for 7 consecutive days, 30 minbefore start training. All drugs were administered intraperitone-ally (i.p.), using 261/2 gauge (13 mm 3 0.45 mm) needles at avolume of 10 mL/kg. All solutions were prepared immediatelybefore use and were kept on ice and protected from light dur-ing the experimental sessions.
Behavioral Tests
All the behavioral tests were performed 24 h after the lasttraining day during the light phase. Half of each group of ani-mals performed the tests to evaluate spatial and working mem-ory and another half performed the tests to assess behavioranxiety-like and locomotion.
Spatial object recognition test
To access spatial hippocampal-dependent memory (Galaniet al., 1998), mice were habituated to a white rectangular box(41 cm 3 34 cm 3 16 cm), that was equally illuminated, dur-ing 5 min. Twenty-four hours later, mice were re-habituatedfor 1 min, and after, two identical objects were placed in theleft (position A) and right (position B) corner of the box (�5cm from the walls). In the sample phase, mice were allowed tofreely investigate the objects until they accumulated a total of30 s of exploration (when the front paws or nose touched theobjects). Thirty minutes later (choice phase) one of the objectshas changed location (position C) and the time spent exploringeach object was recorded. Animals remained in the box untilthey accumulated a total of 30 s of object exploration. Thetime spent to accumulate 30 s of exploration (i.e., elapsedtime) during both the sample and choice phases was recorded.The novel object location was counterbalanced across mice.The box and objects were cleaned with 70% ethanol betweendifferent mice at each test to prevent a build-up of olfactorycues. The height of the objects was sufficient to prevent micefrom climbing up on them (Gresack and Frick, 2006).
Y-maze
The immediate working memory was assessed by recordingspontaneous alternation behavior during a single session in theY-maze (made in white wood with 30 cm long by 6 cm wideby 20 cm high), positioned at the exact same location for allprocedures (Lazaroni et al., 2012). Each naive mouse wasplaced at the end of one arm and allowed to freely movethrough the maze during a session of 8 min. The series of armentries was visually recorded. An arm choice was added torecords only when both forepaws and hind paws fully enteredinto the arm. The Y-maze was cleaned between each animal
tested with 70% alcohol to remove any residual odors. Alterna-tion was defined as a successive entrance onto the three differ-ent arms. The number of entrance sequences (e.g., ABC, BCA)was defined as the number of alternations. The percentage ofalternation was calculated according to the following formula:[total alternation/(total arms entered 2 2)] 3 100 (Lazaroniet al., 2012). Consequently, the following hypotheticalsequence of arms entered by a mice: A, C, A, B, C, A, C, B,A, C would yield an alternation score of 75% ([6 alternations/(10 – 2) arms entered] 3 100). Random selection of goal armsyields an alternation score of 50%.
Elevated plus maze (EPM)
Elevated plus maze is a method for the assessment of uncon-ditioned anxiety-like behavior in rats and mice (Lister, 1987).The apparatus consists of two opposing open arms and twoclosed arms, crossed in the middle perpendicularly to eachother and a center platform common to all arms. The mazewas raised 30 cm above the floor and the animal was allowedto move freely between the arms during 5 min. Since animalshave innate fear of elevated open places they stay for a shortertime in the open arms as compared to the closed arms whenallowed to freely explore the maze. The number of entries andthe time spent in the open arms were used as indices of openspace-induced anxiety in mice. The number of visits in closedarms as well as the time spent in the open arms was calculated(Moreira et al., 2008). An arm entry was defined as a mousehaving fully entered an arm of the maze with all its paws.
Open field test
Spontaneous locomotor activity was measured in the openfield test (Sousa et al., 2006). The open field is made of anopen 50 3 50 cm box with floor subdivided into 16 squares.Each animal was individually positioned in the center of thearena during a single session. The number of segments crossedand the time spent in central square of arena were counted andanalyzed, respectively during 300 s.
Biochemical Analysis
Western blotting
Immediately after the behavioral tests (spatial recognitiontest and Y-maze), animals were euthanized and their hippo-campi quickly collected, frozen in liquid nitrogen and stored at280�C until use. The samples were then homogenized on icein RIPA buffer (150 mM NaCl, 50 mM Tris pH 5 7.4), 1mM EDTA, 1% Nonidet P40, 1 mM phenylmethylsulfonylfluoride, 0.5% sodium deoxycholate). The homogenates werecentrifuged at 2040.35 G (Eppendorf Centrifuge 5415R, Ger-many) at 4�C for 15 min and the protein concentration of thesupernatant was determined by Bradford method (Bradford,1976) using bovine serum albumin (1 mg/mL) as the standard.Samples containing 50 lg of total proteins were denatured inprotein sample buffer (100 mM Tris-HCl pH 5 6.8, 4%SDS, 0.2% bromophenol blue, 20% glycerol, 20% H2O,
ENDOCANNABINOID SYSTEM MEDIATES PROMNESIC EFFECT OF EXERCISE 81
Hippocampus
0.5% b-mercaptoethanol) at 100�C for 4 min and loaded,separated on 12 or 15% SDS-polyacrylamide gels and trans-ferred to polyvinylidene dixuoride membrane (PVDF,Immobilon-P, Millipore, MA) in a Bio-Rad electrophoresissystem (Bio-Rad, Hercules). After blocking, for 24 h, withTBST (Tris-buffered saline with 0.1% Tween) containing 5%non-fat milk and 0.3% BSA, the membranes were kept at4�C overnight with primary antibodies against specific pro-teins, b-actin (1:2500, Santa Cruz Biotechnology, sc-69879),CB1 (1:1000, Calbiochem, #209550), and BDNF (1:500,Santa Cruz Biotechnology, sc-546), respectively. For thechemiluminescent visualization, membranes were incubatedwith anti-rabbit or anti-mouse antibody conjugated withHRP, at room temperature for 1 h. The target protein bandswere detected using the ECL Western blotting detection sys-tem (Pierce) and the density of the b-actin, CB1 and BDNFblotting bands were quantified by ImageJ software (Version1.44p, National Institute Health).
Statistical Analysis
All data are shown as the mean 6 SEM. Graph Pad Prism 4statistics software was used to perform all analyses. Differencesbetween groups were assessed by two-way analysis of variance(ANOVA). Post-hoc comparisons were performed with Bonfer-roni test. In the spatial object recognition test, the preferencefor one object over another during the sample and choicephases was assessed using one-sample t-tests (Baker and Kim,2002; Gresack and Frick, 2006; Suarez et al., 2012). To deter-mine if alternation scores were significantly above the chance(50%), in the Y-maze test, we also used one-sample t-test (Laz-aroni et al., 2012). The significance level was set for all analy-ses at 0.95% (P < 0.05).
RESULTS
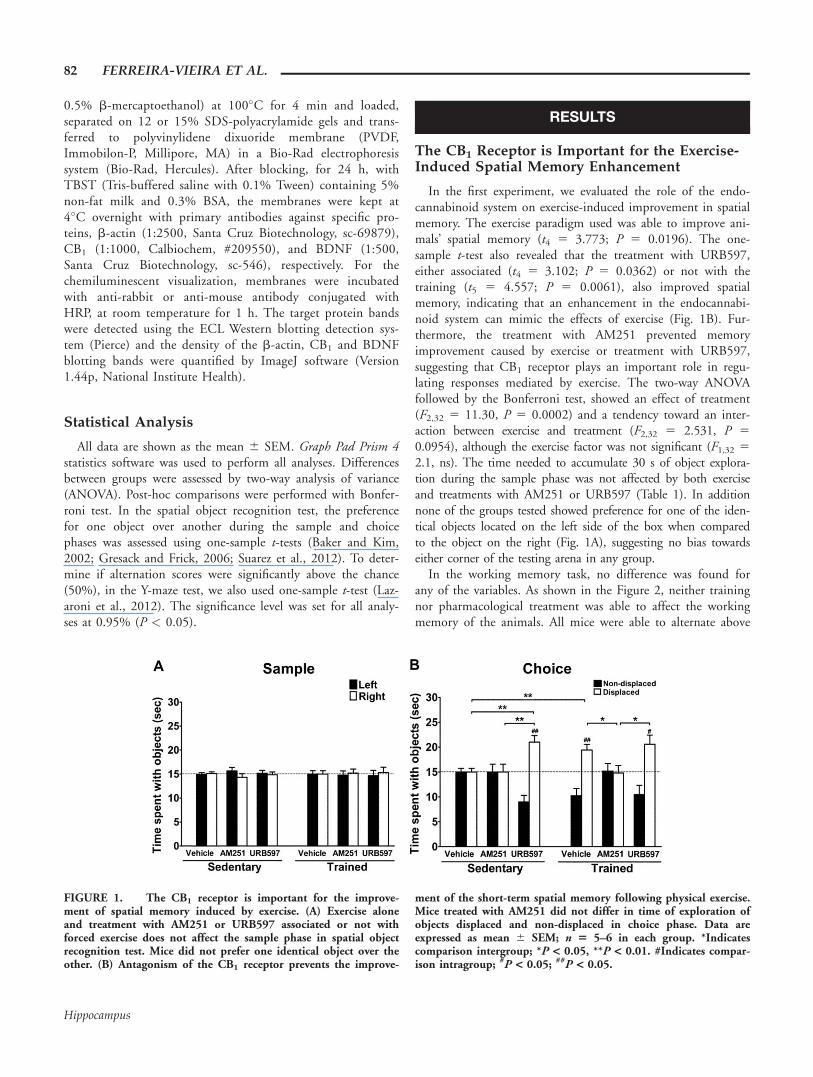
The CB1 Receptor is Important for the Exercise-Induced Spatial Memory Enhancement
In the first experiment, we evaluated the role of the endo-cannabinoid system on exercise-induced improvement in spatialmemory. The exercise paradigm used was able to improve ani-mals’ spatial memory (t4 5 3.773; P 5 0.0196). The one-sample t-test also revealed that the treatment with URB597,either associated (t4 5 3.102; P 5 0.0362) or not with thetraining (t5 5 4.557; P 5 0.0061), also improved spatialmemory, indicating that an enhancement in the endocannabi-noid system can mimic the effects of exercise (Fig. 1B). Fur-thermore, the treatment with AM251 prevented memoryimprovement caused by exercise or treatment with URB597,suggesting that CB1 receptor plays an important role in regu-lating responses mediated by exercise. The two-way ANOVAfollowed by the Bonferroni test, showed an effect of treatment(F2,32 5 11.30, P 5 0.0002) and a tendency toward an inter-action between exercise and treatment (F2,32 5 2.531, P 5
0.0954), although the exercise factor was not significant (F1,32 5
2.1, ns). The time needed to accumulate 30 s of object explora-tion during the sample phase was not affected by both exerciseand treatments with AM251 or URB597 (Table 1). In additionnone of the groups tested showed preference for one of the iden-tical objects located on the left side of the box when comparedto the object on the right (Fig. 1A), suggesting no bias towardseither corner of the testing arena in any group.
In the working memory task, no difference was found forany of the variables. As shown in the Figure 2, neither trainingnor pharmacological treatment was able to affect the workingmemory of the animals. All mice were able to alternate above
FIGURE 1. The CB1 receptor is important for the improve-ment of spatial memory induced by exercise. (A) Exercise aloneand treatment with AM251 or URB597 associated or not withforced exercise does not affect the sample phase in spatial objectrecognition test. Mice did not prefer one identical object over theother. (B) Antagonism of the CB1 receptor prevents the improve-
ment of the short-term spatial memory following physical exercise.Mice treated with AM251 did not differ in time of exploration ofobjects displaced and non-displaced in choice phase. Data areexpressed as mean 6 SEM; n 5 5–6 in each group. *Indicatescomparison intergroup; *P < 0.05, **P < 0.01. #Indicates compar-ison intragroup; #P < 0.05; ##P < 0.05.
82 FERREIRA-VIEIRA ET AL.
Hippocampus
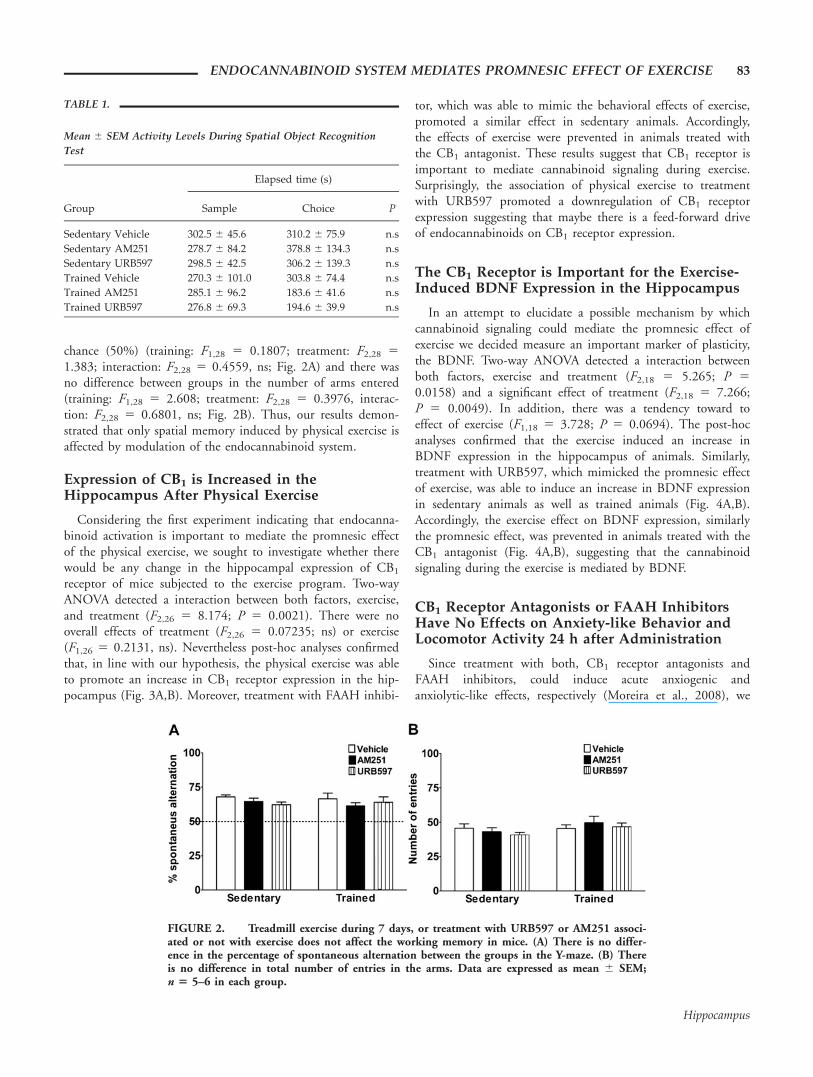
chance (50%) (training: F1,28 5 0.1807; treatment: F2,28 5
1.383; interaction: F2,28 5 0.4559, ns; Fig. 2A) and there wasno difference between groups in the number of arms entered(training: F1,28 5 2.608; treatment: F2,28 5 0.3976, interac-tion: F2,28 5 0.6801, ns; Fig. 2B). Thus, our results demon-strated that only spatial memory induced by physical exercise isaffected by modulation of the endocannabinoid system.
Expression of CB1 is Increased in theHippocampus After Physical Exercise
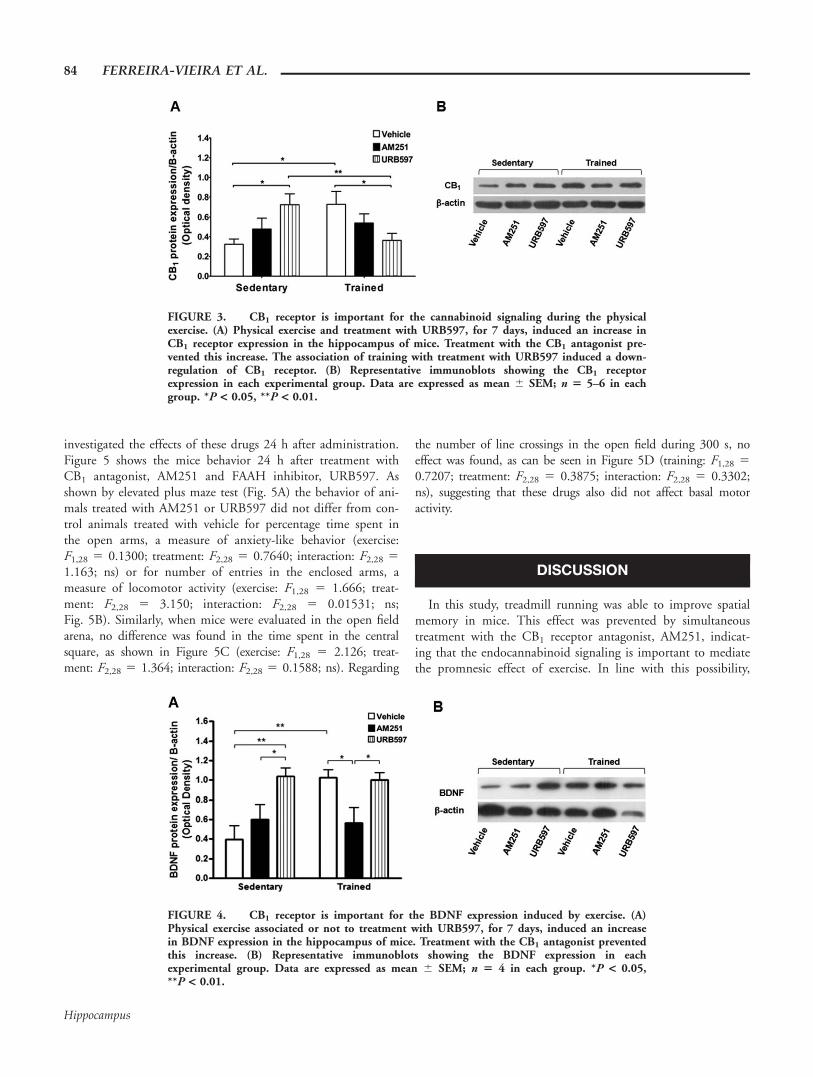
Considering the first experiment indicating that endocanna-binoid activation is important to mediate the promnesic effectof the physical exercise, we sought to investigate whether therewould be any change in the hippocampal expression of CB1
receptor of mice subjected to the exercise program. Two-wayANOVA detected a interaction between both factors, exercise,and treatment (F2,26 5 8.174; P 5 0.0021). There were nooverall effects of treatment (F2,26 5 0.07235; ns) or exercise(F1,26 5 0.2131, ns). Nevertheless post-hoc analyses confirmedthat, in line with our hypothesis, the physical exercise was ableto promote an increase in CB1 receptor expression in the hip-pocampus (Fig. 3A,B). Moreover, treatment with FAAH inhibi-
tor, which was able to mimic the behavioral effects of exercise,promoted a similar effect in sedentary animals. Accordingly,the effects of exercise were prevented in animals treated withthe CB1 antagonist. These results suggest that CB1 receptor isimportant to mediate cannabinoid signaling during exercise.Surprisingly, the association of physical exercise to treatmentwith URB597 promoted a downregulation of CB1 receptorexpression suggesting that maybe there is a feed-forward driveof endocannabinoids on CB1 receptor expression.
The CB1 Receptor is Important for the Exercise-Induced BDNF Expression in the Hippocampus
In an attempt to elucidate a possible mechanism by whichcannabinoid signaling could mediate the promnesic effect ofexercise we decided measure an important marker of plasticity,the BDNF. Two-way ANOVA detected a interaction betweenboth factors, exercise and treatment (F2,18 5 5.265; P 5
0.0158) and a significant effect of treatment (F2,18 5 7.266;P 5 0.0049). In addition, there was a tendency toward toeffect of exercise (F1,18 5 3.728; P 5 0.0694). The post-hocanalyses confirmed that the exercise induced an increase inBDNF expression in the hippocampus of animals. Similarly,treatment with URB597, which mimicked the promnesic effectof exercise, was able to induce an increase in BDNF expressionin sedentary animals as well as trained animals (Fig. 4A,B).Accordingly, the exercise effect on BDNF expression, similarlythe promnesic effect, was prevented in animals treated with theCB1 antagonist (Fig. 4A,B), suggesting that the cannabinoidsignaling during the exercise is mediated by BDNF.
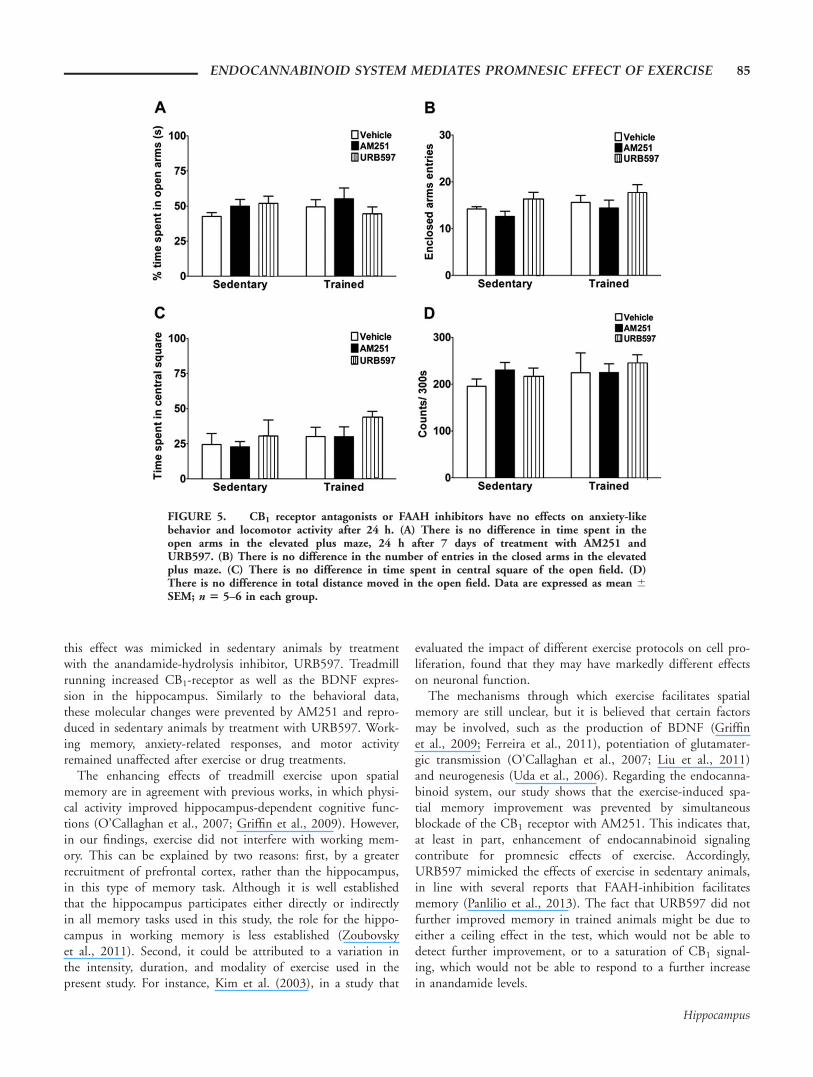
CB1 Receptor Antagonists or FAAH InhibitorsHave No Effects on Anxiety-like Behavior andLocomotor Activity 24 h after Administration
Since treatment with both, CB1 receptor antagonists andFAAH inhibitors, could induce acute anxiogenic andanxiolytic-like effects, respectively (Moreira et al., 2008), we
FIGURE 2. Treadmill exercise during 7 days, or treatment with URB597 or AM251 associ-ated or not with exercise does not affect the working memory in mice. (A) There is no differ-ence in the percentage of spontaneous alternation between the groups in the Y-maze. (B) Thereis no difference in total number of entries in the arms. Data are expressed as mean 6 SEM;n 5 5–6 in each group.
TABLE 1.
Mean 6 SEM Activity Levels During Spatial Object Recognition
Test
Elapsed time (s)
Group Sample Choice P
Sedentary Vehicle 302.5 6 45.6 310.2 6 75.9 n.s
Sedentary AM251 278.7 6 84.2 378.8 6 134.3 n.s
Sedentary URB597 298.5 6 42.5 306.2 6 139.3 n.s
Trained Vehicle 270.3 6 101.0 303.8 6 74.4 n.s
Trained AM251 285.1 6 96.2 183.6 6 41.6 n.s
Trained URB597 276.8 6 69.3 194.6 6 39.9 n.s
ENDOCANNABINOID SYSTEM MEDIATES PROMNESIC EFFECT OF EXERCISE 83
Hippocampus
investigated the effects of these drugs 24 h after administration.Figure 5 shows the mice behavior 24 h after treatment withCB1 antagonist, AM251 and FAAH inhibitor, URB597. Asshown by elevated plus maze test (Fig. 5A) the behavior of ani-mals treated with AM251 or URB597 did not differ from con-trol animals treated with vehicle for percentage time spent inthe open arms, a measure of anxiety-like behavior (exercise:F1,28 5 0.1300; treatment: F2,28 5 0.7640; interaction: F2,28 5
1.163; ns) or for number of entries in the enclosed arms, ameasure of locomotor activity (exercise: F1,28 5 1.666; treat-ment: F2,28 5 3.150; interaction: F2,28 5 0.01531; ns;Fig. 5B). Similarly, when mice were evaluated in the open fieldarena, no difference was found in the time spent in the centralsquare, as shown in Figure 5C (exercise: F1,28 5 2.126; treat-ment: F2,28 5 1.364; interaction: F2,28 5 0.1588; ns). Regarding
the number of line crossings in the open field during 300 s, noeffect was found, as can be seen in Figure 5D (training: F1,28 5
0.7207; treatment: F2,28 5 0.3875; interaction: F2,28 5 0.3302;ns), suggesting that these drugs also did not affect basal motoractivity.
DISCUSSION
In this study, treadmill running was able to improve spatialmemory in mice. This effect was prevented by simultaneoustreatment with the CB1 receptor antagonist, AM251, indicat-ing that the endocannabinoid signaling is important to mediatethe promnesic effect of exercise. In line with this possibility,
FIGURE 3. CB1 receptor is important for the cannabinoid signaling during the physicalexercise. (A) Physical exercise and treatment with URB597, for 7 days, induced an increase inCB1 receptor expression in the hippocampus of mice. Treatment with the CB1 antagonist pre-vented this increase. The association of training with treatment with URB597 induced a down-regulation of CB1 receptor. (B) Representative immunoblots showing the CB1 receptorexpression in each experimental group. Data are expressed as mean 6 SEM; n 5 5–6 in eachgroup. *P < 0.05, **P < 0.01.
FIGURE 4. CB1 receptor is important for the BDNF expression induced by exercise. (A)Physical exercise associated or not to treatment with URB597, for 7 days, induced an increasein BDNF expression in the hippocampus of mice. Treatment with the CB1 antagonist preventedthis increase. (B) Representative immunoblots showing the BDNF expression in eachexperimental group. Data are expressed as mean 6 SEM; n 5 4 in each group. *P < 0.05,**P < 0.01.
84 FERREIRA-VIEIRA ET AL.
Hippocampus
this effect was mimicked in sedentary animals by treatmentwith the anandamide-hydrolysis inhibitor, URB597. Treadmillrunning increased CB1-receptor as well as the BDNF expres-sion in the hippocampus. Similarly to the behavioral data,these molecular changes were prevented by AM251 and repro-duced in sedentary animals by treatment with URB597. Work-ing memory, anxiety-related responses, and motor activityremained unaffected after exercise or drug treatments.
The enhancing effects of treadmill exercise upon spatialmemory are in agreement with previous works, in which physi-cal activity improved hippocampus-dependent cognitive func-tions (O’Callaghan et al., 2007; Griffin et al., 2009). However,in our findings, exercise did not interfere with working mem-ory. This can be explained by two reasons: first, by a greaterrecruitment of prefrontal cortex, rather than the hippocampus,in this type of memory task. Although it is well establishedthat the hippocampus participates either directly or indirectlyin all memory tasks used in this study, the role for the hippo-campus in working memory is less established (Zoubovskyet al., 2011). Second, it could be attributed to a variation inthe intensity, duration, and modality of exercise used in thepresent study. For instance, Kim et al. (2003), in a study that
evaluated the impact of different exercise protocols on cell pro-liferation, found that they may have markedly different effectson neuronal function.
The mechanisms through which exercise facilitates spatialmemory are still unclear, but it is believed that certain factorsmay be involved, such as the production of BDNF (Griffinet al., 2009; Ferreira et al., 2011), potentiation of glutamater-gic transmission (O’Callaghan et al., 2007; Liu et al., 2011)and neurogenesis (Uda et al., 2006). Regarding the endocanna-binoid system, our study shows that the exercise-induced spa-tial memory improvement was prevented by simultaneousblockade of the CB1 receptor with AM251. This indicates that,at least in part, enhancement of endocannabinoid signalingcontribute for promnesic effects of exercise. Accordingly,URB597 mimicked the effects of exercise in sedentary animals,in line with several reports that FAAH-inhibition facilitatesmemory (Panlilio et al., 2013). The fact that URB597 did notfurther improved memory in trained animals might be due toeither a ceiling effect in the test, which would not be able todetect further improvement, or to a saturation of CB1 signal-ing, which would not be able to respond to a further increasein anandamide levels.
FIGURE 5. CB1 receptor antagonists or FAAH inhibitors have no effects on anxiety-likebehavior and locomotor activity after 24 h. (A) There is no difference in time spent in theopen arms in the elevated plus maze, 24 h after 7 days of treatment with AM251 andURB597. (B) There is no difference in the number of entries in the closed arms in the elevatedplus maze. (C) There is no difference in time spent in central square of the open field. (D)There is no difference in total distance moved in the open field. Data are expressed as mean 6SEM; n 5 5–6 in each group.
ENDOCANNABINOID SYSTEM MEDIATES PROMNESIC EFFECT OF EXERCISE 85
Hippocampus

Our results can be supported by several studies showing thatexercise induces an increase in circulating levels of anandamide(Sparling et al., 2003; Raichlen et al., 2012, 2013), in additionto activate cannabinoid signaling (Hill et al., 2010). The endo-cannabinoid signaling seems to be essential to a number ofmolecular and cellular events important for learning and mem-ory, such as LTP formation (Lin et al., 2011; Madronal et al.,2012), the production of neurotrophic factors (Butovsky et al.,2005; Aso et al., 2008) and cellular proliferation/neurogenesis(Aguado et al., 2005; Aguado et al., 2007; Hill et al., 2010).Regarding the facilitation of the endocannabinoid transmission,other works also found positive results in relation to cognitiveperformance in animals not submitted to exercise. For instance,elevating brain levels of anandamide, through either genetic dele-tion or pharmacological inhibition of FAAH, may result in facili-tating extinction processes in the Morris water maze task (Varvelet al., 2007). Similar results were found in relation to memoryacquisition in passive avoidance learning (Mazzola et al., 2009).
In order to strengthen the present results, we sought to findmolecular changes that might be related to the behavioraleffects of exercise, by evaluating the expression of CB1 receptorand BDNF, a marker of synaptic plasticity in the hippocampus.Both were increased by treadmill running, which was preventedby simultaneous CB1 receptor blockade. In addition, theseeffects could be mimicked by URB597 treatment in sedentaryanimals. As expected, the physical exercise associated to treat-ment with URB597 resulted in down-regulation of CB1 recep-tor, in line with our hypothesis of CB1 signaling saturation.The possibility that these effects would result from confound-ing factors related to anxiety-like behavior and locomotion wasdiscarded by testing the animals in the appropriate essays.Thus, these results expand previous investigation in this fieldand show that facilitation of endocannabinoid signaling is nec-essary for exercise to act as a cognitive enhancer.
Previous work has also suggested an increased expression ofCB1 after exercise. Animals exposed to both environmentalenrichment and voluntary wheel show an increase in CB1-mRNA in the hippocampus (Wolf et al., 2010). Similarly, Hillet al. (2010) also showed that voluntary exercise increases theCB1 receptor binding in hippocampus of mice (Hill et al.,2010). Although the mechanisms have remained unclear, onepossibility would be the existence of a feed-forward drive ofendocannabinoids on CB1 receptor expression. In this way,stimulus that increase anandamide levels would lead to anincrease in the expression of CB1, whereas a further increase(association of exercise with treatment with URB597) wouldlead to the opposite effects, by triggering an over-activation ofCB1 receptor with subsequent down-regulation. Consistentlywith this, repeated administration of URB597 decreases thecannabinoid CB1 receptor binding in hippocampus of mice(Marco et al., 2009). Similarly, reduction in CB1 receptorexpression also occurs in the hippocampus of animals thatunderwent a long aerobic physical training program (Gomes daSilva et al., 2010). Others studies also show a decrement inCB1 receptor binding, following chronic cannabinoid agonistadministration (Romero et al., 1998; Sim-Selley et al., 2006).
Our study also shows that the activation of CB1 receptorduring exercise seems important to trigger the production ofBDNF and consequently the pro-cognitive effects. As BDNF isa neurotrophin capable of mediating several processes in themammalian brain (Park and Poo, 2013), it is possible that itwas responsible for mediating promnesic effect of exercisethrough CB1 activation. In line with this, the treatment withAM251 was able to block both, the promnesic effect andBDNF expression. This appears possible since BDNF has beendescribed as capable of mediating several cellular events includ-ing neuronal differentiation and growth, synapse formationand plasticity, and higher cognitive functions (Park and Poo,2013). Other works also observed that CB1 absence leads toemotional impairment and decreases BDNF levels in the hip-pocampus (Aso et al., 2008), suggesting an important linkbetween the activation of the CB1 receptor and the crucial roleplayed by BDNF on the emotional behavior. Evidence has alsoemerged from studies with humans subjects, showing thatBDNF expression might be a mechanism by which the endo-cannabinoid influences the neuroplastic and antidepressanteffects of exercise (Heyman et al., 2012). Hill et al. (2010) in astudy that evaluated the role of cannabinoid system in media-ting the hippocampal cell proliferation induced by voluntaryexercise, showed that endogenous cannabinoid signaling associ-ated with activation of the CB1 receptor during exercise is cru-cial for the occurrence of this phenomenon (Hill et al., 2010),sustaining the idea that BDNF may be a mediator of cannabi-noid signaling.
While exogenous substances that direct activates CB1 recep-tors impair hippocampal function (Hoffman et al., 2007) andlearning (Lichtman et al., 1995), this study revealed that facili-tation of endocannabinoid signaling for seven days, throughphysical exercise or FAAH-inhibition, improves short-term spa-tial memory in animals. Our findings also provide evidencethat an increase in BDNF expression may be one of the mech-anisms involved.
Acknowledgments
A.R.M., F.A.M. and G.S.P. would like to thank CNPq for aproductive research fellowship (level 2) and FAPEMIG for thegrants APQ-01038-11 (FAM), CBB - APQ-02238-12 (ARM)and PRONEM CBB - APQ-04721-10(ARM, GSP).
REFERENCES
Aguado T, Monory K, Palazuelos J, Stella N, Cravatt B, Lutz B,Marsicano G, Kokaia Z, Guzman M, Galve-Roperh I. 2005. Theendocannabinoid system drives neural progenitor proliferation.FASEB J 19:1704–1706.
Aguado T, Romero E, Monory K, Palazuelos J, Sendtner M,Marsicano G, Lutz B, Guzman M, Galve-Roperh I. 2007. TheCB1 cannabinoid receptor mediates excitotoxicity-induced neuralprogenitor proliferation and neurogenesis. J Biol Chem 282:23892–23898.
86 FERREIRA-VIEIRA ET AL.
Hippocampus

Aso E, Ozaita A, Valdizan EM, Ledent C, Pazos A, Maldonado R,Valverde O. 2008. BDNF impairment in the hippocampus isrelated to enhanced despair behavior in CB1 knockout mice.J Neurochem 105:565–572.
Baker KB, Kim JJ. 2002. Effects of stress and hippocampal NMDA recep-tor antagonism on recognition memory in rats. Learn Mem 9:58–65.
Bekinschtein P, Oomen CA, Saksida LM, Bussey TJ. 2011. Effects ofenvironmental enrichment and voluntary exercise on neurogenesis,learning and memory, and pattern separation: BDNF as a criticalvariable? Semin Cell Dev Biol 22:536–542.
Bisogno T, Ligresti A, Di Marzo V. 2005. The endocannabinoid signallingsystem: Biochemical aspects. Pharmacol Biochem Behav 81:224–238.
Bradford MM. 1976. A rapid and sensitive method for the quantita-tion of microgram quantities of protein utilizing the principle ofprotein-dye binding. Anal Biochem 72:248–254.
Broadbent NJ, Squire LR, Clark RE. 2004. Spatial memory, recogni-tion memory, and the hippocampus. Proc Natl Acad Sci USA 101:14515–14520.
Butovsky E, Juknat A, Goncharov I, Elbaz J, Eilam R, Zangen A,Vogel Z. 2005. In vivo up-regulation of brain-derived neurotrophicfactor in specific brain areas by chronic exposure to Delta-tetrahy-drocannabinol. J Neurochem 93:802–811.
Cabral GA, Marciano-Cabral F. 2005. Cannabinoid receptors inmicroglia of the central nervous system: Immune functional rele-vance. J Leukoc Biol 78:1192–1197.
Chaouloff F, Dubreucq S, Bellocchio L, Marsicano G. 2011. Endo-cannabinoids and motor behavior: CB1 receptors also control run-ning activity. Physiology (Bethesda) 26:76–77; author reply 78.
Clarkson-Smith L, Hartley AA. 1989. Relationships between physicalexercise and cognitive abilities in older adults. Psychol Aging 4:183–189.
Davies SN, Pertwee RG, Riedel G. 2002. Functions of cannabinoidreceptors in the hippocampus. Neuropharmacology 42:993–1007.
Devane WA, Hanus L, Breuer A, Pertwee RG, Stevenson LA, GriffinG, Gibson D, Mandelbaum A, Etinger A, Mechoulam R. 1992.Isolation and structure of a brain constituent that binds to the can-nabinoid receptor. Science 258:1946–1949.
Di Marzo V. 2008. Targeting the endocannabinoid system: Toenhance or reduce? Nat Rev Drug Discov 7:438–455.
Di Marzo V. 2009. The endocannabinoid system: Its general strategyof action, tools for its pharmacological manipulation and potentialtherapeutic exploitation. Pharmacol Res 60:77–84.
Di Marzo V, Bifulco M, De Petrocellis L. 2004. The endocannabinoidsystem and its therapeutic exploitation. Nat Rev Drug Discov 3:771–784.
Drumond LE, Mourao FA, Leite HR, Abreu RV, Reis HJ, MoraesMF, Pereira GS, Massensini AR. 2012. Differential effects of swim-ming training on neuronal calcium sensor-1 expression in rat hip-pocampus/cortex and in object recognition memory tasks. BrainRes Bull 88:385–391.
Dubreucq S, Koehl M, Abrous DN, Marsicano G, Chaouloff F. 2010.CB1 receptor deficiency decreases wheel-running activity: Conse-quences on emotional behaviours and hippocampal neurogenesis.Exp Neurol 224:106–113.
Egashira N, Mishima K, Iwasaki K, Fujiwara M. 2002. Intracerebralmicroinjections of delta 9-tetrahydrocannabinol: Search for theimpairment of spatial memory in the eight-arm radial maze in rats.Brain Res 952:239–245.
Ferreira AF, Real CC, Rodrigues AC, Alves AS, Britto LR. 2011. Short-term, moderate exercise is capable of inducing structural, BDNF-independent hippocampal plasticity. Brain Res 1425:111–122.
Fuchs E, Flugge G. 1998. Stress, glucocorticoids and structural plastic-ity of the hippocampus. Neurosci Biobehav Rev 23:295–300.
Galani R, Weiss I, Cassel JC, Kelche C. 1998. Spatial memory, habit-uation, and reactions to spatial and nonspatial changes in rats withselective lesions of the hippocampus, the entorhinal cortex or thesubiculum. Behav Brain Res 96:1–12.
Galiegue S, Mary S, Marchand J, Dussossoy D, Carriere D, CarayonP, Bouaboula M, Shire D, Le Fur G, Casellas P. 1995. Expressionof central and peripheral cannabinoid receptors in human immunetissues and leukocyte subpopulations. Eur J Biochem 232:54–61.
Gomes da Silva S, Araujo BH, Cossa AC, Scorza FA, Cavalheiro EA,Naffah-Mazzacoratti Mda G, Arida RM. 2010. Physical exercise inadolescence changes CB1 cannabinoid receptor expression in therat brain. Neurochem Int 57:492–496.
Gresack JE, Frick KM. 2006. Effects of continuous and intermittentestrogen treatments on memory in aging female mice. Brain Res1115:135–147.
Griffin EW, Bechara RG, Birch AM, Kelly AM. 2009. Exercise enhan-ces hippocampal-dependent learning in the rat: Evidence for aBDNF-related mechanism. Hippocampus 19:973–980.
Heyman E, Gamelin FX, Goekint M, Piscitelli F, Roelands B, LeclairE, Di Marzo V, Meeusen R. 2012. Intense exercise increases circu-lating endocannabinoid and BDNF levels in humans--possibleimplications for reward and depression. Psychoneuroendocrinology37:844–851.
Hill MN, Titterness AK, Morrish AC, Carrier EJ, Lee TT,Gil-Mohapel J, Gorzalka BB, Hillard CJ, Christie BR. 2010.Endogenous cannabinoid signaling is required for voluntaryexercise-induced enhancement of progenitor cell proliferation inthe hippocampus. Hippocampus 20:513–523.
Hillman CH, Erickson KI, Kramer AF. 2008. Be smart, exercise yourheart: Exercise effects on brain and cognition. Nat Rev Neurosci 9:58–65.
Hoffman AF, Oz M, Yang R, Lichtman AH, Lupica CR. 2007.Opposing actions of chronic Delta9-tetrahydrocannabinol and can-nabinoid antagonists on hippocampal long-term potentiation.Learn Mem 14:63–74.
Johnson RA, Rhodes JS, Jeffrey SL, Garland T, Jr, Mitchell GS. 2003.Hippocampal brain-derived neurotrophic factor but notneurotrophin-3 increases more in mice selected for increased vol-untary wheel running. Neuroscience 121:1–7.
Kim YP, Kim HB, Jang MH, Lim BV, Kim YJ, Kim H, Kim SS, KimEH, Kim CJ. 2003. Magnitude- and time-dependence of the effectof treadmill exercise on cell proliferation in the dentate gyrus ofrats. Int J Sports Med 24:114–117.
Lazaroni TL, Raslan AC, Fontes WR, de Oliveira ML, Bader M,Alenina N, Moraes MF, Dos Santos RA, Pereira GS. 2012. Angio-tensin-(1-7)/Mas axis integrity is required for the expression ofobject recognition memory. Neurobiol Learn Mem 97:113–123.
Lichtman AH, Dimen KR, Martin BR. 1995. Systemic or intrahippo-campal cannabinoid administration impairs spatial memory in rats.Psychopharmacology (Berl) 119:282–290.
Lin QS, Yang Q, Liu DD, Sun Z, Dang H, Liang J, Wang YX, ChenJ, Li ST. 2011. Hippocampal endocannabinoids play an importantrole in induction of long-term potentiation and regulation of con-textual fear memory formation. Brain Res Bull 86:139–145.
Lister RG. 1987. The use of a plus-maze to measure anxiety in themouse. Psychopharmacology (Berl) 92:180–185.
Liu HL, Zhao G, Cai K, Zhao HH, Shi LD. 2011. Treadmill exerciseprevents decline in spatial learning and memory in APP/PS1 trans-genic mice through improvement of hippocampal long-term poten-tiation. Behav Brain Res 218:308–314.
Madronal N, Gruart A, Valverde O, Espadas I, Moratalla R, Delgado-Garcia JM. 2012. Involvement of cannabinoid CB1 receptor inassociative learning and in hippocampal CA3-CA1 synaptic plastic-ity. Cereb Cortex 22:550–566.
Marco EM, Rubino T, Adriani W, Viveros MP, Parolaro D, LaviolaG. 2009. Long-term consequences of URB597 administration dur-ing adolescence on cannabinoid CB1 receptor binding in brainareas. Brain Res 1257:25–31.
Matsuda LA, Lolait SJ, Brownstein MJ, Young AC, Bonner TI. 1990.Structure of a cannabinoid receptor and functional expression ofthe cloned cDNA. Nature 346:561–564.
ENDOCANNABINOID SYSTEM MEDIATES PROMNESIC EFFECT OF EXERCISE 87
Hippocampus

Mazzola C, Medalie J, Scherma M, Panlilio LV, Solinas M, Tanda G,Drago F, Cadet JL, Goldberg SR, Yasar S. 2009. Fatty acid amidehydrolase (FAAH) inhibition enhances memory acquisitionthrough activation of PPAR-alpha nuclear receptors. Learn Mem16:332–337.
Mechoulam R, Ben-Shabat S, Hanus L, Ligumsky M, Kaminski NE,Schatz AR, Gopher A, Almog S, Martin BR, Compton DR,Pertwee RG, Griffin G, Bayewitch M, Barg J, Vogel Z0. 1995.Identification of an endogenous 2-monoglyceride, present in caninegut, that binds to cannabinoid receptors. Biochem Pharmacol 50:83–90.
Mitra R, Sundlass K, Parker KJ, Schatzberg AF, Lyons DM. 2006.Social stress-related behavior affects hippocampal cell proliferationin mice. Physiol Behav 89:123–127.
Moreira FA, Kaiser N, Monory K, Lutz B. 2008. Reduced anxiety-likebehaviour induced by genetic and pharmacological inhibition ofthe endocannabinoid-degrading enzyme fatty acid amide hydrolase(FAAH) is mediated by CB1 receptors. Neuropharmacology 54:141–150.
Munro S, Thomas KL, Abu-Shaar M. 1993. Molecular characteriza-tion of a peripheral receptor for cannabinoids. Nature 365:61–65.
Neeper SA, Gomez-Pinilla F, Choi J, Cotman CW. 1996. Physicalactivity increases mRNA for brain-derived neurotrophic factor andnerve growth factor in rat brain. Brain Res 726:49–56.
O’Callaghan RM, Ohle R, Kelly AM. 2007. The effects of forcedexercise on hippocampal plasticity in the rat: A comparison ofLTP, spatial- and non-spatial learning. Behav Brain Res 176:362–366.
Olson AK, Eadie BD, Ernst C, Christie BR. 2006. Environmentalenrichment and voluntary exercise massively increase neurogenesisin the adult hippocampus via dissociable pathways. Hippocampus16:250–260.
Panlilio LV, Justinova Z, Goldberg SR. 2013. Inhibition of FAAHand activation of PPAR: New approaches to the treatment of cog-nitive dysfunction and drug addiction. Pharmacol Ther 138:84–102.
Park H, Poo MM. 2013. Neurotrophin regulation of neural circuitdevelopment and function. Nat Rev Neurosci 14:7–23.
Patel S, Hillard CJ. 2006. Pharmacological evaluation of cannabinoidreceptor ligands in a mouse model of anxiety: Further evidence foran anxiolytic role for endogenous cannabinoid signaling. J Pharma-col Exp Ther 318:304–311.
Piomelli D. 2003. The molecular logic of endocannabinoid signalling.Nat Rev Neurosci 4:873–884.
Raichlen DA, Foster AD, Gerdeman GL, Seillier A, Giuffrida A.2012. Wired to run: Exercise-induced endocannabinoid signalingin humans and cursorial mammals with implications for the ‘run-ner’s high’. J Exp Biol 215:1331–1336.
Raichlen DA, Foster AD, Seillier A, Giuffrida A, Gerdeman GL.2013. Exercise-induced endocannabinoid signaling is modulated byintensity. Eur J Appl Physiol 113:869–875.
Romero J, Berrendero F, Manzanares J, Perez A, Corchero J, FuentesJA, Fernandez-Ruiz JJ, Ramos JA. 1998. Time-course of the canna-binoid receptor down-regulation in the adult rat brain caused byrepeated exposure to delta9-tetrahydrocannabinol. Synapse 30:298–308.
Sim-Selley LJ, Schechter NS, Rorrer WK, Dalton GD, Hernandez J,Martin BR, Selley DE. 2006. Prolonged recovery rate of CB1
receptor adaptation after cessation of long-term cannabinoidadministration. Mol Pharmacol 70:986–996.
Sousa N, Almeida OF, Wotjak CT. 2006. A hitchhiker’s guide tobehavioral analysis in laboratory rodents. Genes Brain Behav 5:5–24.
Soya H, Nakamura T, Deocaris CC, Kimpara A, Iimura M, FujikawaT, Chang H, McEwen BS, Nishijima T. 2007. BDNF inductionwith mild exercise in the rat hippocampus. Biochem Biophys ResCommun 358:961–967.
Sparling PB, Giuffrida A, Piomelli D, Rosskopf L, Dietrich A. 2003.Exercise activates the endocannabinoid system. Neuroreport 14:2209–2211.
Suarez LM, Cid E, Gal B, Inostroza M, Brotons-Mas JR, Gomez-Dominguez D, de la Prida LM, Solis JM. 2012. Systemic injectionof kainic acid differently affects LTP magnitude depending on itsepileptogenic efficiency. PLoS One 7:e48128.
Suenaga T, Kaku M, Ichitani Y. 2008. Effects of intrahippocampalcannabinoid receptor agonist and antagonist on radial maze andT-maze delayed alternation performance in rats. PharmacolBiochem Behav 91:91–96.
Suijo K, Inoue S, Ohya Y, Odagiri Y, Takamiya T, Ishibashi H, ItohM, Fujieda Y, Shimomitsu T. 2012. Resistance exercise enhancescognitive function in mouse. Int J Sports Med 34:368–375.
Uda M, Ishido M, Kami K, Masuhara M. 2006. Effects of chronictreadmill running on neurogenesis in the dentate gyrus of the hip-pocampus of adult rat. Brain Res 1104:64–72.
van Praag H, Christie BR, Sejnowski TJ, Gage FH. 1999. Runningenhances neurogenesis, learning, and long-term potentiation inmice. Proc Natl Acad Sci USA 96:13427–13431.
van Praag H, Kempermann G, Gage FH. 2000. Neural consequencesof environmental enrichment. Nat Rev Neurosci 1:191–198.
Varvel SA, Wise LE, Niyuhire F, Cravatt BF, Lichtman AH. 2007.Inhibition of fatty-acid amide hydrolase accelerates acquisition andextinction rates in a spatial memory task. Neuropsychopharmacol-ogy 32:1032–1041.
Vaynman S, Gomez-Pinilla F. 2005. License to run: Exercise impactsfunctional plasticity in the intact and injured central nervous sys-tem by using neurotrophins. Neurorehabil Neural Repair 19:283–295.
Wang J, Ueda N. 2009. Biology of endocannabinoid synthesis system.Prostaglandins Other Lipid Mediat 89:112–119.
Wise LE, Harloe JP, Lichtman AH. 2009a. Fatty acid amide hydrolase(FAAH) knockout mice exhibit enhanced acquisition of an aver-sive, but not of an appetitive, Barnes maze task. Neurobiol LearnMem 92:597–601.
Wise LE, Thorpe AJ, Lichtman AH. 2009b. Hippocampal CB(1)receptors mediate the memory impairing effects of Delta(9)-tetra-hydrocannabinol. Neuropsychopharmacology 34:2072–2080.
Wolf SA, Bick-Sander A, Fabel K, Leal-Galicia P, Tauber S, Ramirez-Rodriguez G, Muller A, Melnik A, Waltinger TP, Ullrich O,Kempermann G. 2010. Cannabinoid receptor CB1 mediates base-line and activity-induced survival of new neurons in adult hippo-campal neurogenesis. Cell Commun Signal 8:12.
Zoubovsky SP, Pogorelov VM, Taniguchi Y, Kim SH, Yoon P, NwuliaE, Sawa A, Pletnikov MV, Kamiya A. 2011. Working memory def-icits in neuronal nitric oxide synthase knockout mice: Potentialimpairments in prefrontal cortex mediated cognitive function. Bio-chem Biophys Res Commun 408:707–712.
88 FERREIRA-VIEIRA ET AL.
Hippocampus





























