Embed Size (px)
Citation preview

of January 11, 2022.This information is current as
EditingRecombination Accessibility and ReceptorJoining in C57BL/6 Mice: Implications for
J− Repertoire and VκSkewed Primary Ig
Schork and David NemazeeMiyo Aoki-Ota, Ali Torkamani, Takayuki Ota, Nicholas
http://www.jimmunol.org/content/188/5/2305doi: 10.4049/jimmunol.1103484January 2012;
2012; 188:2305-2315; Prepublished online 27J Immunol
MaterialSupplementary
4.DC1http://www.jimmunol.org/content/suppl/2012/01/27/jimmunol.110348
Referenceshttp://www.jimmunol.org/content/188/5/2305.full#ref-list-1
, 31 of which you can access for free at: cites 67 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2012 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on January 11, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on January 11, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on January 11, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on January 11, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

The Journal of Immunology
Skewed Primary Igk Repertoire and V–J Joining in C57BL/6Mice: Implications for Recombination Accessibility andReceptor Editing
Miyo Aoki-Ota,* Ali Torkamani,† Takayuki Ota,* Nicholas Schork,† and David Nemazee*
Previous estimates of the diversity of the mouse Ab repertoire have been based on fragmentary data as a result of many technical
limitations, in particular, the many samples necessary to provide adequate coverage. In this study, we used 59-coding end
amplification of Igk mRNAs from bone marrow, splenic, and lymph node B cells of C57BL/6 mice combined with amplicon
pyrosequencing to assess the functional and nonfunctional Vk repertoire. To evaluate the potential effects of receptor editing, we
also compared V/J associations and usage in bone marrows of mouse mutants under constitutive negative selection or an altered
ability to undergo secondary recombination. To focus on preimmune B cells, our cell sorting strategy excluded memory B cells and
plasma cells. Analysis of ∼90 Mbp, representing >250,000 individual transcripts from 59 mice, revealed that 101 distinct func-
tional Vk genes are used but at frequencies ranging from ∼0.001 to ∼10%. Usage of seven Vk genes made up >40% of the
repertoire. A small class of transcripts from apparently nonfunctional Vk genes was found, as were occasional transcripts from
several apparently functional genes that carry aberrant recombination signals. Of 404 potential V–J combinations (101 Vks 3 4
Jks), 398 (98.5%) were found at least once in our sample. For most Vk transcripts, all Jks were used, but V–J association biases
were common. Usage patterns were remarkably stable in different selective conditions. Overall, the primary k repertoire is highly
skewed by preferred rearrangements, limiting Ab diversity, but potentially facilitating receptor editing. The Journal of Immu-
nology, 2012, 188: 2305–2315.
Immunoglobulin genes encode Abs vital to adaptive immunity.In B cell development, Ab H and L chain genes are assembledindependently at sequential developmental stages by recom-
bination of the Igh followed by Igk (or Igl) gene loci, respec-tively. Igk and Igl loci are encoded on different chromosomes invertebrates and recombine in cis to assemble V and J gene seg-ments. In the C57BL/6 (B6) strain mouse, the genomic sequencesof the Ab loci have been assembled and analyzed (1–12).The Igklocus occupies .3 megabase of DNA, carrying over 100 poten-tially functional V elements and 4 functional J elements (desig-nated throughout as Vk and Jk, respectively) (1–6, 8, 13–16).Although it is widely assumed that V–J association is relatively
random, consistent with a role in diversification of the Ab reper-
toire (17), the overall biases in Vk association with various Jks arenot known. Moreover, the k locus plays an important role in re-ceptor selection. Vk usage and joining to Jk elements are influ-enced by a feedback process that is regulated by the nature of theL chain generated. As a consequence, rearrangements resultingin out-of-frame, underexpressed, or autoreactive receptors fail toterminate recombination, frequently leading to secondary rear-rangements or receptor editing (reviewed in Ref. 18). The murineIgk locus favors primary rearrangements to upstream Jks (Jk1 andJk2) and secondary rearrangements to downstream Jk4 and Jk5(Jk3 is a pseudogene) (13, 19–22). Adding to the complexity is thefact that over half of Vks are in the opposite transcriptional ori-entation to the Jk-Ck cluster (Ref. 8, in this study, we find 64%).As a result, some primary V–J rearrangements lead to deletion ofintervening DNA, preventing the subsequent use of potentiallyfunctional elements in between, and limiting the repertoire avail-able for secondary recombination, whereas other rearrangementslead to inversion, retaining the Vk repertoire for secondary use.Thus, the pattern of Vk-Jk associations may provide clues as to therules and regulation of receptor editing.The Igk locus also carries an element called the k deleting ele-
ment recombining sequence (RS) in mice, located ∼25 kb down-stream of Ck, which can undergo recombination in cis with Vks orheptamer sites in the Jk-Ck intron, leading to silencing of previ-ously functional Vk-Jk rearrangements (23–27). RS recombina-tion occurs in most cells that go on to l L chain expression (28)and on the silent Igk allele in ∼10% of k B cells (29). Mutation ofthe recombination signal of RS in the mouse germline suppressesreceptor editing in a subset of cells and indirectly reduces theproduction of Igl-expressing cells (30), consistent with a role inrescuing autoreactive B cells that have “run out” of editing pos-sibilities on a particular Igk allele (25).Expression of the Igk repertoire has been analyzed over the
years, first with plasmacytoma panels and protein sequencing,
*Department of Immunology and Microbial Science, The Scripps Research Institute,La Jolla, CA 92037; and †Translational Sciences Institute, The Scripps Research Insti-tute, La Jolla, CA 92037
Received for publication December 5, 2011. Accepted for publication December 28,2011.
This work was supported by the National Institutes of Health Grants RO1AI059714and R01DE017568. A.T. and N.S. are supported in part by the National Institutes ofHealth/National Center for Research Resources Grant UL1 RR025774.
The sequences presented in this article have been submitted to the National Centerfor Biotechnology Information (http://www.ncbi.nlm.nih.gov/sra/?term=SRX113094)under accession number SRX113094.
Address correspondence and reprint requests to Dr. David Nemazee, The ScrippsResearch Institute, 10550 North Torrey Pines Road, La Jolla, CA 92037. E-mailaddress: [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: B6, C57BL/6; BM, bone marrow; IMGT, In-ternational Immunogenetics Information System; LN, lymph node; MID, multipleidentifier; NCBI, National Center for Biotechnology Information; pUlik, designationof mouse strain expressing Igk superantigen; RIC, recombination signal informationcontent; RS, recombining sequence/murine k deleting element; SP, spleen; TSRI, TheScripps Research Institute.
Copyright� 2012 by The American Association of Immunologists, Inc. 0022-1767/12/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1103484
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

subsequently by cDNA cloning and sequencing, then later by di-rect sequencing and RT-PCR approaches, often using hybridomapanels (31–33). However, the sheer size, diversity, and polymor-phism of the Igk repertoire have hindered a comprehensive viewof Ab L chain usage. Analysis of publicly available sequencecompendia such as the National Center for Biotechnology Infor-mation (NCBI) (http://www.ncbi.nlm.nih.gov/), Kabat (34), andthe International Immunogenetics Information System (IMGT)(16) databases have been important tools, but they combine datacollected in different ways for varying purposes and from multiplemouse strains. With high-throughput sequencing, it is now pos-sible to obtain sufficient data from 59-RACE amplicons to obtaina better picture of the overall baseline usage. In addition to ananalysis of functional transcripts from normal mice, we were ableto obtain a large number of nonfunctional VJ sequences from bonemarrow (BM) B cells of normal mice and functional sequencesfrom BM B cells of mice under uniform negative selection, thusproviding information on rearrangement preferences.
Materials and MethodsMice
All micewere bred andmaintained in The Scripps Research Institute (TSRI)Animal Resources facility according to the Institutional Animal Care andUse guidelines. C57BL/6J mice were purchased from The Jackson Labo-ratory and bred in the TSRI custom breeding colony. pUlik (35), JkCk-deleted (JCk2/2) (36), and RS2/2 (30) mice have been described previ-ously. All mice used in this study had been extensively backcrossed to a B6background.
B cell isolation from spleen and lymph nodes
B6 B cells were isolated from four 3-mo-old mice, which included twofemales and two males. Spleen (SP) and lymph node (LN) cells wereharvested and then depleted of erythrocytes using ammonium chloride lysisprior to B cell isolation. For B cell isolation, 50 million SP cells or 40million LN cells from each gender were combined and B cells were isolatedusing the “no touch” B cell isolation kit (Miltenyi Biotec).
BM B cell isolation
CD4/82B220+IgD2CD1382 BM B cells were isolated from 2- to 3-mo-oldC57BL/6J male mice (n = 13), JCk+/2mice (n = 9), RS2/2 (n = 11), or pUlikmice (n = 11) as follows. BM cells were harvested from femurs, pelvis, andhumerus and then depleted of erythrocytes. Prior to FACS sorting, B220+
B cells were enriched by using CD45R (B220) MicroBeads (Miltenyi Bio-tec). Briefly, 400–500 million cells were incubated with 400–500 ml B220MicroBeads, and B220+ cells were magnetically separated. Then, cell sur-face marker stains were performed using a standard protocol. All of thefollowing Abs were used at 1:200 dilution in FACS buffer (HBSS containing0.5% BSA, 1 mM EDTA, and 0.1% NaN3) with 3 3 106 cells/stain: FITCanti-B220 (RA3-6B2; BD), PE anti-CD138 (281-2; BD), alexa647 anti-IgD(JA12.5), PerCP-Cy5.5 anti-CD4 (GK1.5; BD), and PerCP-Cy5.5 anti-CD8(53-6.7; BD). After staining, CD4/82B220+IgD2CD1382 BM B cells wereisolated by sorting using a FACSAria (BD) machine.
454 sequencing
Total RNA was obtained from purified B cells using TRIzol reagents(Invitrogen). k-chain cDNAs from each sample were synthesized usinga 59-RACE kit (Ambion), according to the manufacturer’s protocol. A totalof 0.5 mg total RNA/sample were used. For reverse transcriptase or PCR,Transcriptor High Fidelity (Roche) and Phusion Hot Start (New EnglandBiolabs) was used, respectively. For SP and LN B cells, a bar-code strategywas used to distinguish samples in later analysis. The following oligonu-cleotides were used to amplify k-chain sequences: K-R1 (male), 59-TTGA-CTGCTCACTGGATGGTGGGAAGATGG-39; K-R2 (female), 59-TTATCT-GCTCACTGGATGGTGGGAAGATGG-39; RACE-1 (male), 59-TTGACG-CGGATCCGAACACTGCGTTTGCTGGCTTTGATG-39; and RACE-2 (fe-male), 59-TTGTCGCGGATCCGAACACTGCGTTTGCTGGCTTTGATG-39.
PCR products were purified on agarose gels, and 2.5 mg each of femaleand male-derived amplicons was combined and used for 454 sequenceanalysis. For BM B cells, the “A-key” sequence was appended to the 39-primer, and the B-key was added to the 59-primer to allow selective se-quencing from the 39-end, which included the V–J junctions. Sequencing
was performed using GS FLX Titanium emPCR Kits (Lib-L; Roche).Briefly, the beads included in the kit capture only nucleotides carrying theB-key and yield sequences that start from the A-key primer site. A bar-code strategy with multiple identifier (MID) sequences was also used todistinguish each strain. The following oligonucleotides were used to am-plify k-chain sequences: A-key+MID1+KR 59-CCATCTCATCCCTGCGT-GTCTCCGACTCAGACGAGTGCGTCTGCTCACTGGATGGTGGGAA-GATGG-39, A-key+MID2+KR 59-CCATCTCATCCCTGCGTGTCTCCG-ACTCAGACGCTCGACACTGCTCACTGGATGGTGGGAAGATGG-39,A-key+MID3+KR 59-CCATCTCATCCCTGCGTGTCTCCGACTCAGAG-ACGCACTCCTGCTCACTGGATGGTGGGAAGATGG-39, A-key+MID4+KR 59-CCATCTCATCCCTGCGTGTCTCCGACTCAGAGCACTGTAG-CTGCTCACTGGATGGTGGGAAGATGG-39, B-key+RACE 59-CCTAT-CCCCTGTGTGCCTTGGCAGTCTCAGCGCGGATCCGAACACTGCG-TTTGCTGGCTTTGATG-39.
Each PCR product was purified on agarose gel, and 100 ng of eachamplicon library was combined and used for 454 sequencing analysis. GSFLX Titanium sequencing kit XLR70 (Roche) was used for samplepreparation, and a four-region gasket setup was used for processing. Datawere collected at the TSRI Next Generation Sequencing core and at theUniversity of California Los Angeles Sequencing & Genotyping Core. Datawere analyzed using stand-alone basic local alignment search tool program(37) downloaded from NCBI. Reference Vk and Jk sequences were ob-tained from the IMGT Web site (http://www.imgt.org) (16).We analyzedclones without unread bases using the Megablast algorithm as describedpreviously (38). Sequencing data have been submitted to the NCBI data-base under accession number SRX113094 (http://www.ncbi.nlm.nih.gov/sra/?term=SRX113094).
ResultsIdentification of 101 functional Vk genes and ∼400 Vk/Jkcombinations
To evaluate the Igk repertoire of B6 mice, we analyzed Igk tran-scripts in BM, SP, and LN. BM samples were sorted on B220+
IgD2CD1382 cells to exclude from analysis mature B cells andplasma cells. Splenic and LN samples were sorted similarly. Asdescribed below, we also analyzed BM B cells of selected B6congenic mutants of interest, including heterozygous JkCk-deleted(JCk+/2), murine k deleting element knockouts (RS2/2), and micetransgenic for a ubiquitously expressed k superantigen (pUIik).Amplicons were generated from k mRNA using a 59-RACE PCRapproach with two invariant primers, one in Ck and the second ina universal 59-linker; we then carried out 454 sequencing andanalysis as described previously (38).These methods were designedto sample the preimmune repertoire and to avoid biases caused bypreferential PCR amplification. Table I summarizes the data set.We could obtain for each sample at least 11,000 reads with iden-tifiable Vks (mean, 17,222 6 5,850). Most of these carried iden-tifiable VJ combinations (mean, 14,067 6 4,833). A total of 101functional Vk genes were found (Supplemental Table I), including10 defined as open reading frames of unclear functionality byIMGT (16) and the expected 4 Jks. Theoretically, there should beas many as 404 VJ combinations represented. We found from 327to 382 combinations in combined samples of the various tissues ofB6 mice and somewhat higher numbers of combinations in B6congenic BM with defined mutations. Combining data from allsamples, ∼98.5% of possible VJ combinations were seen at leastonce. In addition to functional sequences, in our Vk usage analysis,we also captured nonproductive sequences that provide informa-tion on the underlying probabilities of rearrangement.
Skewed Igk rearrangements in immature B cells in BM
To evaluate the Igk repertoire in the context of minimal selection,we analyzed Vk usage in B220+IgD2CD1382CD42CD82 B cellsof the BM, which include pre-B and newly formed sIgM+ andsIgM2 B cells (the latter cells have often downregulated theirreceptors (38)). These samples will be called simply “BM” Bcells. Previous studies suggested that chromosome contractionmight equalize the chances of rearrangement of distal and proxi-
2306 SKEWED Igk V GENE USAGE AND V–J ASSOCIATIONS IN B6 MICE
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
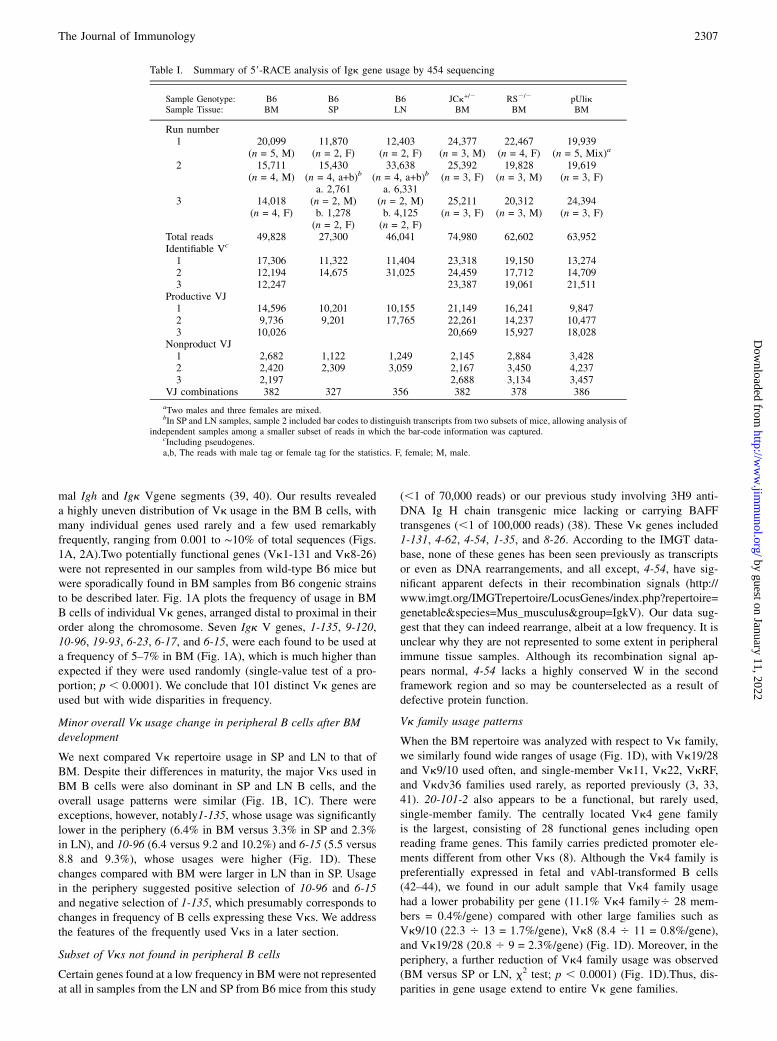
mal Igh and Igk Vgene segments (39, 40). Our results revealeda highly uneven distribution of Vk usage in the BM B cells, withmany individual genes used rarely and a few used remarkablyfrequently, ranging from 0.001 to ∼10% of total sequences (Figs.1A, 2A).Two potentially functional genes (Vk1-131 and Vk8-26)were not represented in our samples from wild-type B6 mice butwere sporadically found in BM samples from B6 congenic strainsto be described later. Fig. 1A plots the frequency of usage in BMB cells of individual Vk genes, arranged distal to proximal in theirorder along the chromosome. Seven Igk V genes, 1-135, 9-120,10-96, 19-93, 6-23, 6-17, and 6-15, were each found to be used ata frequency of 5–7% in BM (Fig. 1A), which is much higher thanexpected if they were used randomly (single-value test of a pro-portion; p , 0.0001). We conclude that 101 distinct Vk genes areused but with wide disparities in frequency.
Minor overall Vk usage change in peripheral B cells after BMdevelopment
We next compared Vk repertoire usage in SP and LN to that ofBM. Despite their differences in maturity, the major Vks used inBM B cells were also dominant in SP and LN B cells, and theoverall usage patterns were similar (Fig. 1B, 1C). There wereexceptions, however, notably1-135, whose usage was significantlylower in the periphery (6.4% in BM versus 3.3% in SP and 2.3%in LN), and 10-96 (6.4 versus 9.2 and 10.2%) and 6-15 (5.5 versus8.8 and 9.3%), whose usages were higher (Fig. 1D). Thesechanges compared with BM were larger in LN than in SP. Usagein the periphery suggested positive selection of 10-96 and 6-15and negative selection of 1-135, which presumably corresponds tochanges in frequency of B cells expressing these Vks. We addressthe features of the frequently used Vks in a later section.
Subset of Vks not found in peripheral B cells
Certain genes found at a low frequency in BMwere not representedat all in samples from the LN and SP from B6 mice from this study
(,1 of 70,000 reads) or our previous study involving 3H9 anti-DNA Ig H chain transgenic mice lacking or carrying BAFFtransgenes (,1 of 100,000 reads) (38). These Vk genes included1-131, 4-62, 4-54, 1-35, and 8-26. According to the IMGT data-base, none of these genes has been seen previously as transcriptsor even as DNA rearrangements, and all except, 4-54, have sig-nificant apparent defects in their recombination signals (http://www.imgt.org/IMGTrepertoire/LocusGenes/index.php?repertoire=genetable&species=Mus_musculus&group=IgkV). Our data sug-gest that they can indeed rearrange, albeit at a low frequency. It isunclear why they are not represented to some extent in peripheralimmune tissue samples. Although its recombination signal ap-pears normal, 4-54 lacks a highly conserved W in the secondframework region and so may be counterselected as a result ofdefective protein function.
Vk family usage patterns
When the BM repertoire was analyzed with respect to Vk family,we similarly found wide ranges of usage (Fig. 1D), with Vk19/28and Vk9/10 used often, and single-member Vk11, Vk22, VkRF,and Vkdv36 families used rarely, as reported previously (3, 33,41). 20-101-2 also appears to be a functional, but rarely used,single-member family. The centrally located Vk4 gene familyis the largest, consisting of 28 functional genes including openreading frame genes. This family carries predicted promoter ele-ments different from other Vks (8). Although the Vk4 family ispreferentially expressed in fetal and vAbl-transformed B cells(42–44), we found in our adult sample that Vk4 family usagehad a lower probability per gene (11.1% Vk4 family4 28 mem-bers = 0.4%/gene) compared with other large families such asVk9/10 (22.3 4 13 = 1.7%/gene), Vk8 (8.4 4 11 = 0.8%/gene),and Vk19/28 (20.8 4 9 = 2.3%/gene) (Fig. 1D). Moreover, in theperiphery, a further reduction of Vk4 family usage was observed(BM versus SP or LN, x2 test; p , 0.0001) (Fig. 1D).Thus, dis-parities in gene usage extend to entire Vk gene families.
Table I. Summary of 59-RACE analysis of Igk gene usage by 454 sequencing
Sample Genotype: B6 B6 B6 JCk+/2 RS2/2 pUlikSample Tissue: BM SP LN BM BM BM
Run number1 20,099 11,870 12,403 24,377 22,467 19,939
(n = 5, M) (n = 2, F) (n = 2, F) (n = 3, M) (n = 4, F) (n = 5, Mix)a
2 15,711 15,430 33,638 25,392 19,828 19,619(n = 4, M) (n = 4, a+b)b (n = 4, a+b)b (n = 3, F) (n = 3, M) (n = 3, F)
a. 2,761 a. 6,3313 14,018 (n = 2, M) (n = 2, M) 25,211 20,312 24,394
(n = 4, F) b. 1,278 b. 4,125 (n = 3, F) (n = 3, M) (n = 3, F)(n = 2, F) (n = 2, F)
Total reads 49,828 27,300 46,041 74,980 62,602 63,952Identifiable Vc
1 17,306 11,322 11,404 23,318 19,150 13,2742 12,194 14,675 31,025 24,459 17,712 14,7093 12,247 23,387 19,061 21,511
Productive VJ1 14,596 10,201 10,155 21,149 16,241 9,8472 9,736 9,201 17,765 22,261 14,237 10,4773 10,026 20,669 15,927 18,028
Nonproduct VJ1 2,682 1,122 1,249 2,145 2,884 3,4282 2,420 2,309 3,059 2,167 3,450 4,2373 2,197 2,688 3,134 3,457
VJ combinations 382 327 356 382 378 386
aTwo males and three females are mixed.bIn SP and LN samples, sample 2 included bar codes to distinguish transcripts from two subsets of mice, allowing analysis of
independent samples among a smaller subset of reads in which the bar-code information was captured.cIncluding pseudogenes.a,b, The reads with male tag or female tag for the statistics. F, female; M, male.
The Journal of Immunology 2307
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
Jk usage in B6 mice
Jk usage in BM, SP, and LN was analyzed among productivesequences, revealing a stable hierarchy (Fig. 1E) with Jk1 used
most frequently (34, 35, and 33% in BM, SP, and LN, respec-
tively; p , 0.0001 compared with other Jks), followed by Jk5 (29,
29, and 31%), Jk2 (20, 22, and 21%), and Jk4, which was used
significantly less than other Jks (16, 15, and 14%; p , 0.0001
compared with other Jks). In LN, Jk5 usage was significantly
elevated compared with BM (x2 test; p , 0.0001). Jk2 has
a nonamer and a heptamer that are noncanonical: AGTTTTTGT
instead of GGTTTTTGT and CAGTGTG instead of CACTGTG,
respectively. Heptamers and nonamers with such sequences are
rare among Vk and Jk gene segments (Supplemental Tables I, II).
Jk4 has a noncanonical spacer, which is 1 base longer than normal
(24 instead of 23 bases; Supplemental Table II). A comparable
increase in spacer length of recombination substrates leads to
a 15–40% decrease in recombination efficiency in transient trans-fection assays (45). These differences may explain in part therelative underrepresentation of Jk2 and Jk4.
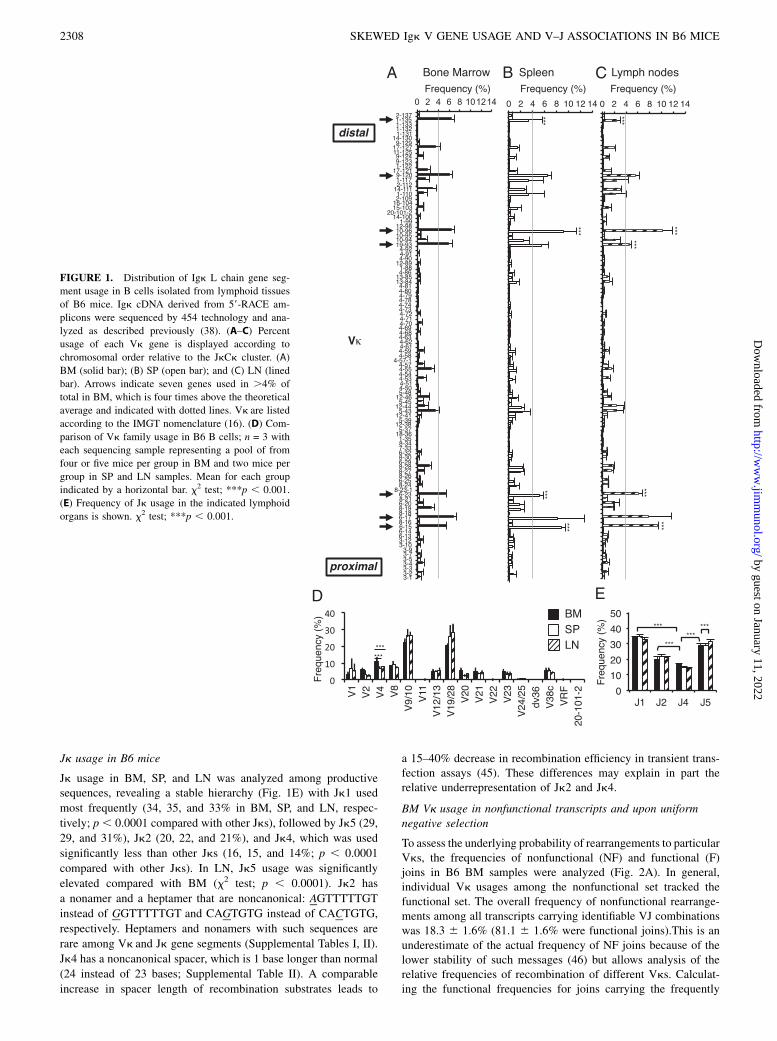
BM Vk usage in nonfunctional transcripts and upon uniformnegative selection
To assess the underlying probability of rearrangements to particularVks, the frequencies of nonfunctional (NF) and functional (F)joins in B6 BM samples were analyzed (Fig. 2A). In general,individual Vk usages among the nonfunctional set tracked thefunctional set. The overall frequency of nonfunctional rearrange-ments among all transcripts carrying identifiable VJ combinationswas 18.3 6 1.6% (81.1 6 1.6% were functional joins).This is anunderestimate of the actual frequency of NF joins because of thelower stability of such messages (46) but allows analysis of therelative frequencies of recombination of different Vks. Calculat-ing the functional frequencies for joins carrying the frequently
FIGURE 1. Distribution of Igk L chain gene seg-
ment usage in B cells isolated from lymphoid tissues
of B6 mice. Igk cDNA derived from 59-RACE am-
plicons were sequenced by 454 technology and ana-
lyzed as described previously (38). (A–C) Percent
usage of each Vk gene is displayed according to
chromosomal order relative to the JkCk cluster. (A)
BM (solid bar); (B) SP (open bar); and (C) LN (lined
bar). Arrows indicate seven genes used in .4% of
total in BM, which is four times above the theoretical
average and indicated with dotted lines. Vk are listed
according to the IMGT nomenclature (16). (D) Com-
parison of Vk family usage in B6 B cells; n = 3 with
each sequencing sample representing a pool of from
four or five mice per group in BM and two mice per
group in SP and LN samples. Mean for each group
indicated by a horizontal bar. x2 test; ***p , 0.001.
(E) Frequency of Jk usage in the indicated lymphoid
organs is shown. x2 test; ***p , 0.001.
2308 SKEWED Igk V GENE USAGE AND V–J ASSOCIATIONS IN B6 MICE
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
used Vks revealed a similar range of frequencies (81.7–85.5%);however, 1-135 had a lower frequency (70.6 6 1.9%, x2test; p ,0.0001) (Fig. 2C). Because of its high level of usage and distallocation, 1-135 likely represents a special case (see Discussion).The patterns of Jk usage among functional and nonfunctionalsequences were similar overall and for individual highly usedgenes (Fig. 2D, 2E).We then compared Vk usage of nonfunctional B6 sequences
with functional sequences found in BM of B6 congenic micecarrying a ubiquitously expressed k superantigen transgene(pUIik). The superantigen negatively selects all k B cells andstimulates receptor editing, leading to a massive increase in lB cell production (35). Remarkably, the pattern of Vk usage wasnearly identical with that of nonfunctional B6 BM (Fig. 2B).Again, Vk genes dominantly seen among functional B6 BMsamples were most frequent. Thus, functional Vk usage in im-mature BM B cells under conditions of uniform negative selectionwas similar to Vk usage among nonfunctional transcripts. Weconclude that the dominance of certain Vk gene usage is a resultof intrinsic rearrangement (and perhaps expression) preferencesrather than subsequent selection.
Analysis of the features of frequently used Vks
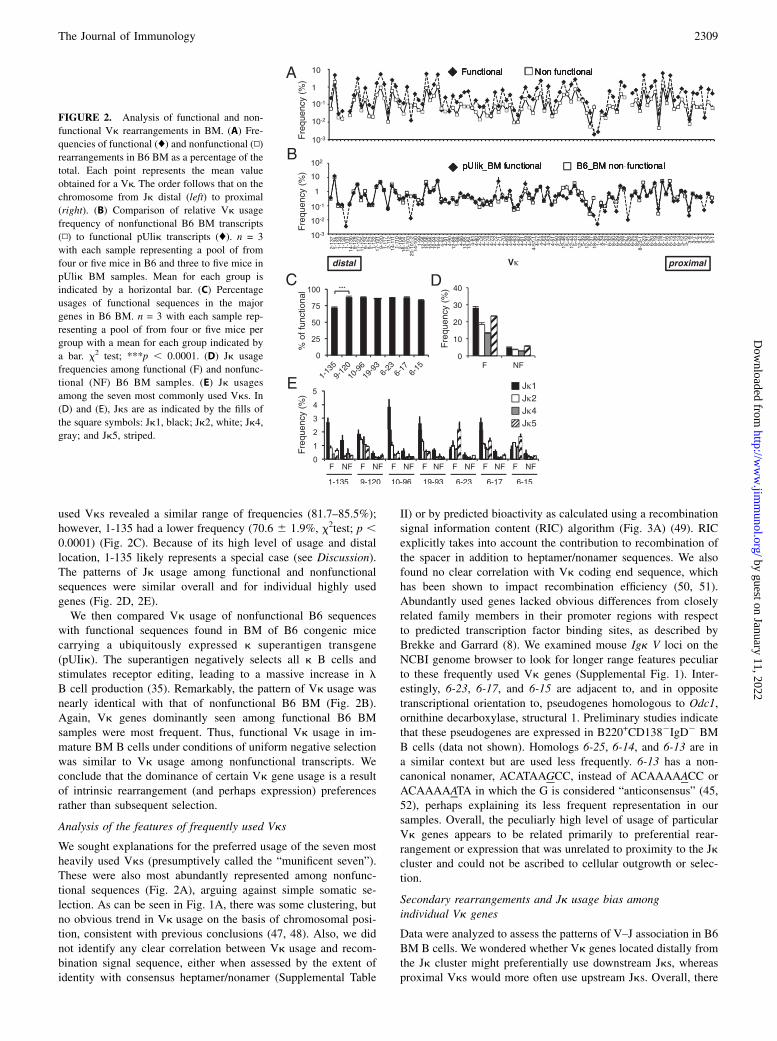
We sought explanations for the preferred usage of the seven mostheavily used Vks (presumptively called the “munificent seven”).These were also most abundantly represented among nonfunc-tional sequences (Fig. 2A), arguing against simple somatic se-lection. As can be seen in Fig. 1A, there was some clustering, butno obvious trend in Vk usage on the basis of chromosomal posi-tion, consistent with previous conclusions (47, 48). Also, we didnot identify any clear correlation between Vk usage and recom-bination signal sequence, either when assessed by the extent ofidentity with consensus heptamer/nonamer (Supplemental Table
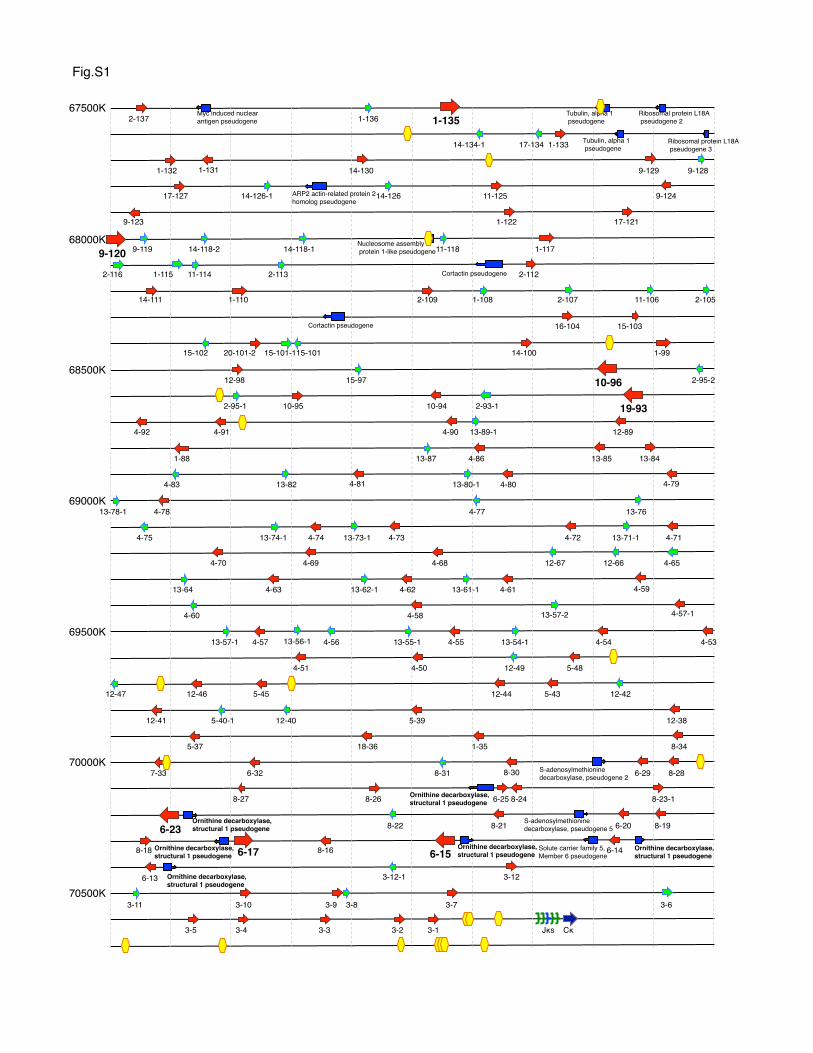
II) or by predicted bioactivity as calculated using a recombinationsignal information content (RIC) algorithm (Fig. 3A) (49). RICexplicitly takes into account the contribution to recombination ofthe spacer in addition to heptamer/nonamer sequences. We alsofound no clear correlation with Vk coding end sequence, whichhas been shown to impact recombination efficiency (50, 51).Abundantly used genes lacked obvious differences from closelyrelated family members in their promoter regions with respectto predicted transcription factor binding sites, as described byBrekke and Garrard (8). We examined mouse Igk V loci on theNCBI genome browser to look for longer range features peculiarto these frequently used Vk genes (Supplemental Fig. 1). Inter-estingly, 6-23, 6-17, and 6-15 are adjacent to, and in oppositetranscriptional orientation to, pseudogenes homologous to Odc1,ornithine decarboxylase, structural 1. Preliminary studies indicatethat these pseudogenes are expressed in B220+CD1382IgD2 BMB cells (data not shown). Homologs 6-25, 6-14, and 6-13 are ina similar context but are used less frequently. 6-13 has a non-canonical nonamer, ACATAAGCC, instead of ACAAAAACC orACAAAAATA in which the G is considered “anticonsensus” (45,52), perhaps explaining its less frequent representation in oursamples. Overall, the peculiarly high level of usage of particularVk genes appears to be related primarily to preferential rear-rangement or expression that was unrelated to proximity to the Jkcluster and could not be ascribed to cellular outgrowth or selec-tion.
Secondary rearrangements and Jk usage bias amongindividual Vk genes
Data were analyzed to assess the patterns of V–J association in B6BM B cells. We wondered whether Vk genes located distally fromthe Jk cluster might preferentially use downstream Jks, whereasproximal Vks would more often use upstream Jks. Overall, there
FIGURE 2. Analysis of functional and non-
functional Vk rearrangements in BM. (A) Fre-
quencies of functional (♦) and nonfunctional (N)
rearrangements in B6 BM as a percentage of the
total. Each point represents the mean value
obtained for a Vk. The order follows that on the
chromosome from Jk distal (left) to proximal
(right). (B) Comparison of relative Vk usage
frequency of nonfunctional B6 BM transcripts
(N) to functional pUlik transcripts (♦). n = 3
with each sample representing a pool of from
four or five mice in B6 and three to five mice in
pUlik BM samples. Mean for each group is
indicated by a horizontal bar. (C) Percentage
usages of functional sequences in the major
genes in B6 BM. n = 3 with each sample rep-
resenting a pool of from four or five mice per
group with a mean for each group indicated by
a bar. x2 test; ***p , 0.0001. (D) Jk usage
frequencies among functional (F) and nonfunc-
tional (NF) B6 BM samples. (E) Jk usages
among the seven most commonly used Vks. In
(D) and (E), Jks are as indicated by the fills of
the square symbols: Jk1, black; Jk2, white; Jk4,
gray; and Jk5, striped.
The Journal of Immunology 2309
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
was no such correlation, with several distal genes (1-135, 10-96,and 19-93) preferentially rearranged to Jk1, whereas many prox-imal and intermediately positioned Vks (6-23, 6-15, and multiplemembers of the Vk4 family) preferentially rearranged to Jk5.These usage data are shown in Fig. 3B, which for clarity onlyshows V–J association data for Jk1 and Jk5. Preferential associ-ation of Vk4 members with Jk5 has been noted previously (44,53). We also asked whether genes that were frequently used mightbe associated preferentially with Jk1, whereas rarely used Vkswould be limited to downstream J usage. Among the seven mostfrequently used Vks, five of seven were associated more oftenwith Jk1 than with Jk5; however, 6-15 and 6-23 were preferen-tially associated with Jk5 (Fig. 3B). The 20 most frequently usedVks made up 72% of the sample and had a Jk usage distributionsimilar to the overall averages (Jk1, 35%; Jk2, 21%; Jk4, 16%;and Jk5, 27%). In contrast, among the 20 least-used Vks, whichaccounted for 0.62% of the sample, 49% were joined to Jk5,whereas only 14% used Jk1, indicating that secondary recombi-nations were indeed important for their representation.
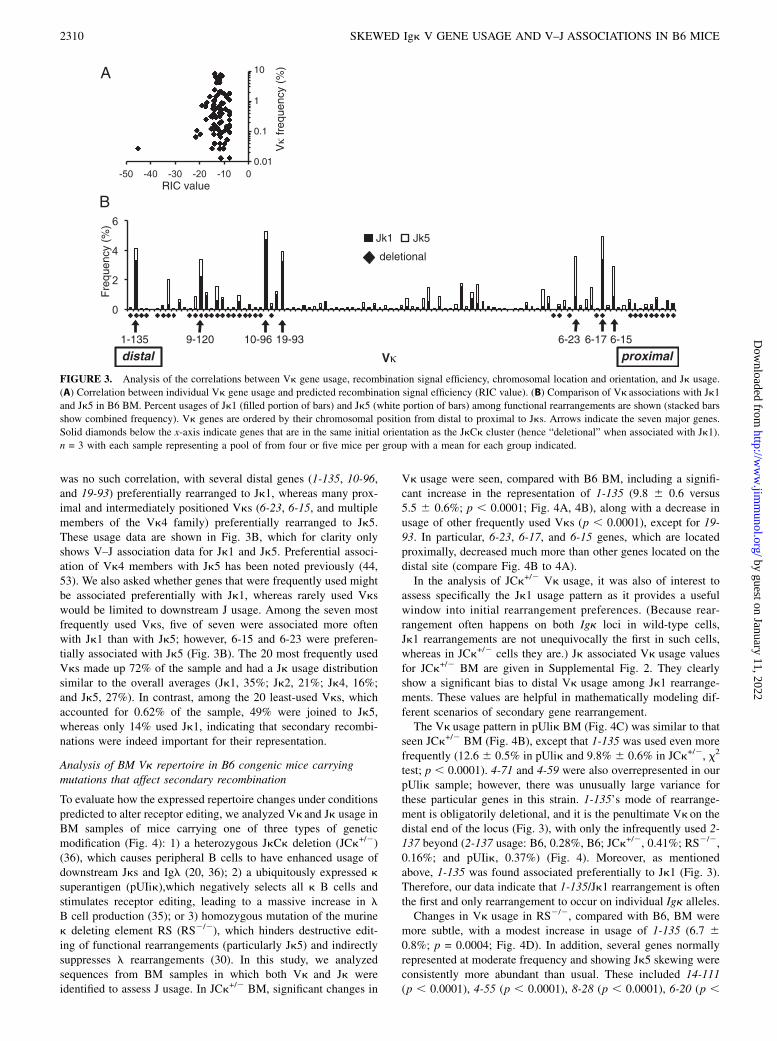
Analysis of BM Vk repertoire in B6 congenic mice carryingmutations that affect secondary recombination
To evaluate how the expressed repertoire changes under conditionspredicted to alter receptor editing, we analyzed Vk and Jk usage inBM samples of mice carrying one of three types of geneticmodification (Fig. 4): 1) a heterozygous JkCk deletion (JCk+/2)(36), which causes peripheral B cells to have enhanced usage ofdownstream Jks and Igl (20, 36); 2) a ubiquitously expressed ksuperantigen (pUIik),which negatively selects all k B cells andstimulates receptor editing, leading to a massive increase in lB cell production (35); or 3) homozygous mutation of the murinek deleting element RS (RS2/2), which hinders destructive edit-ing of functional rearrangements (particularly Jk5) and indirectlysuppresses l rearrangements (30). In this study, we analyzedsequences from BM samples in which both Vk and Jk wereidentified to assess J usage. In JCk+/2 BM, significant changes in
Vk usage were seen, compared with B6 BM, including a signifi-cant increase in the representation of 1-135 (9.8 6 0.6 versus5.5 6 0.6%; p , 0.0001; Fig. 4A, 4B), along with a decrease inusage of other frequently used Vks (p , 0.0001), except for 19-93. In particular, 6-23, 6-17, and 6-15 genes, which are locatedproximally, decreased much more than other genes located on thedistal site (compare Fig. 4B to 4A).In the analysis of JCk+/2 Vk usage, it was also of interest to
assess specifically the Jk1 usage pattern as it provides a usefulwindow into initial rearrangement preferences. (Because rear-rangement often happens on both Igk loci in wild-type cells,Jk1 rearrangements are not unequivocally the first in such cells,whereas in JCk+/2 cells they are.) Jk associated Vk usage valuesfor JCk+/2 BM are given in Supplemental Fig. 2. They clearlyshow a significant bias to distal Vk usage among Jk1 rearrange-ments. These values are helpful in mathematically modeling dif-ferent scenarios of secondary gene rearrangement.The Vk usage pattern in pUlik BM (Fig. 4C) was similar to that
seen JCk+/2 BM (Fig. 4B), except that 1-135 was used even morefrequently (12.6 6 0.5% in pUlik and 9.8% 6 0.6% in JCk+/2, x2
test; p , 0.0001). 4-71 and 4-59 were also overrepresented in ourpUlik sample; however, there was unusually large variance forthese particular genes in this strain. 1-135’s mode of rearrange-ment is obligatorily deletional, and it is the penultimate Vkon thedistal end of the locus (Fig. 3), with only the infrequently used 2-137 beyond (2-137 usage: B6, 0.28%, B6; JCk+/2, 0.41%; RS2/2,0.16%; and pUIik, 0.37%) (Fig. 4). Moreover, as mentionedabove, 1-135 was found associated preferentially to Jk1 (Fig. 3).Therefore, our data indicate that 1-135/Jk1 rearrangement is oftenthe first and only rearrangement to occur on individual Igk alleles.Changes in Vk usage in RS2/2, compared with B6, BM were
more subtle, with a modest increase in usage of 1-135 (6.7 60.8%; p = 0.0004; Fig. 4D). In addition, several genes normallyrepresented at moderate frequency and showing Jk5 skewing wereconsistently more abundant than usual. These included 14-111(p , 0.0001), 4-55 (p , 0.0001), 8-28 (p , 0.0001), 6-20 (p ,
FIGURE 3. Analysis of the correlations between Vk gene usage, recombination signal efficiency, chromosomal location and orientation, and Jk usage.
(A) Correlation between individual Vk gene usage and predicted recombination signal efficiency (RIC value). (B) Comparison of Vk associations with Jk1
and Jk5 in B6 BM. Percent usages of Jk1 (filled portion of bars) and Jk5 (white portion of bars) among functional rearrangements are shown (stacked bars
show combined frequency). Vk genes are ordered by their chromosomal position from distal to proximal to Jks. Arrows indicate the seven major genes.
Solid diamonds below the x-axis indicate genes that are in the same initial orientation as the JkCk cluster (hence “deletional” when associated with Jk1).
n = 3 with each sample representing a pool of from four or five mice per group with a mean for each group indicated.
2310 SKEWED Igk V GENE USAGE AND V–J ASSOCIATIONS IN B6 MICE
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

0.0001), and 8-19 (p = 0.0008) (Fig. 5A). The results indicatedthat under normal conditions RS recombination sometimes si-lenced these genes, presumably because their properties promotedediting under some circumstances.
Jk usages in BMs of genetically modified strains
Although overall Jk usage preference was not radically changed inmutant strains in that Jk1 and Jk5 continued to be used mostfrequently, significant differences were seen (Fig. 5B). JCk+/2 andpUlik BM had similar reductions in usage of Jk1 (B6 versusJCk+/2; p , 0.0001, B6 versus pUlik; p , 0.0001) and increasesin usage of Jk5 (B6 versus JCk+/2; p , 0.0001, B6 versus pUlik;p , 0.0001). In addition, RS2/2 BM B cells had significantlyhigher Jk5 usage compared with all other strains (p , 0.0001).Surprisingly, however, when VJ rearrangements in the seven mostfrequently used Vks were compared between B6 and the geneti-cally modified mice, no major changes in their Jk associationswere observed. For each Vk, either Jk1 or Jk5 usage predomi-nated, with 1-135, 9-120, 10-96, 19-93, and 6-17 genes prefer-entially rearranged to Jk1 (Fig. 6, A–D, 6F) and 6-23 and 6-15preferentially rearranged to Jk5 (Fig. 6E, 6G).
CDR3 analysis
Our VJ analysis suggests that Igk rearrangements converge oncertain major usages in many situations with predicted limitationson diversity. However, this analysis does not take into accountjunctional diversity. To address this, we analyzed predicted CDR3aa diversity generated by individual Vk gene association withparticular Jks (Fig. 7). In the case of 1-135/J1, there were 38
FIGURE 4. Igk repertoire in BM from B6 and genetically modified
mouse strains. Usage frequencies of Vk genes among functional rear-
rangements in B cells from the following strains: B6 (A); JCk+/2 (B);
pUlik (C); and RS2/2 (D). The seven major genes indicated with unfilled
bars that were found in .4% of sequences from B6 BM. Vk genes are
ordered by their chromosomal position from distal to proximal to Jks.
n = 3 with each sample representing a pool of from three to five mice per
group in each strain with mean for each group indicated by a horizontal
bar.
0
15
30
45
J1 J2 J4 J5
0
2
4
6
8
B6_BMJC +/-_BM
RS-/-_BMpUli _BM
***
******
******
***
Fre
quen
cy (
%)
A
B
Fre
quen
cy (
%)
***
******
***
***
***
FIGURE 5. Comparative usage analysis in BM of B6 and genetically
modified strains showing Vks with elevated usage in RS2/2 and overall Jk
usage distributions. (A) Frequencies of six genes with increased repre-
sentation in RS2/2 samples are shown. (B) BM Jk usages of the indicated
strains are shown. B6, solid bar; JCk+/2, open bar; RS2/2, gray bar; and
pUlik, lined bar. n = 3 with each sample representing a pool of from three
to five mice per group in each strain. x2 test; ***p , 0.001.
FIGURE 6. Jk usages associated with the major Vk genes in BM of B6,
JCk+/2, RS2/2, and pUIik mice. B6, solid bar; JCk+/2, open bar; RS2/2,
gray bar; and pUlik, lined bar. n = 3 with each sample representing a pool
of from three to five mice per group in each strain with a mean for each
group indicated by a horizontal bar. Individual panels show frequencies
of the indicated Vk genes: (A) 1-135; (B) 9-120; (C) 10-96; (D) 19-93; (E)
6-23; (F) 6-17; (G) 6-15.
The Journal of Immunology 2311
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
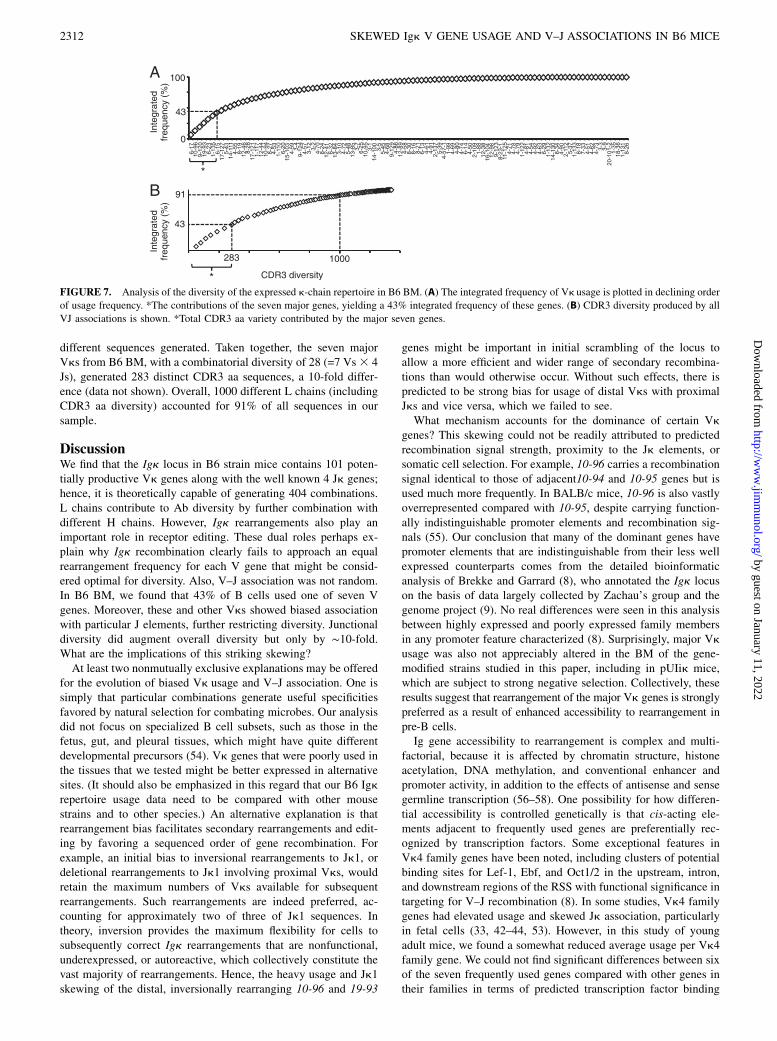
different sequences generated. Taken together, the seven majorVks from B6 BM, with a combinatorial diversity of 28 (=7 Vs3 4Js), generated 283 distinct CDR3 aa sequences, a 10-fold differ-ence (data not shown). Overall, 1000 different L chains (includingCDR3 aa diversity) accounted for 91% of all sequences in oursample.
DiscussionWe find that the Igk locus in B6 strain mice contains 101 poten-tially productive Vk genes along with the well known 4 Jk genes;hence, it is theoretically capable of generating 404 combinations.L chains contribute to Ab diversity by further combination withdifferent H chains. However, Igk rearrangements also play animportant role in receptor editing. These dual roles perhaps ex-plain why Igk recombination clearly fails to approach an equalrearrangement frequency for each V gene that might be consid-ered optimal for diversity. Also, V–J association was not random.In B6 BM, we found that 43% of B cells used one of seven Vgenes. Moreover, these and other Vks showed biased associationwith particular J elements, further restricting diversity. Junctionaldiversity did augment overall diversity but only by ∼10-fold.What are the implications of this striking skewing?At least two nonmutually exclusive explanations may be offered
for the evolution of biased Vk usage and V–J association. One issimply that particular combinations generate useful specificitiesfavored by natural selection for combating microbes. Our analysisdid not focus on specialized B cell subsets, such as those in thefetus, gut, and pleural tissues, which might have quite differentdevelopmental precursors (54). Vk genes that were poorly used inthe tissues that we tested might be better expressed in alternativesites. (It should also be emphasized in this regard that our B6 Igkrepertoire usage data need to be compared with other mousestrains and to other species.) An alternative explanation is thatrearrangement bias facilitates secondary rearrangements and edit-ing by favoring a sequenced order of gene recombination. Forexample, an initial bias to inversional rearrangements to Jk1, ordeletional rearrangements to Jk1 involving proximal Vks, wouldretain the maximum numbers of Vks available for subsequentrearrangements. Such rearrangements are indeed preferred, ac-counting for approximately two of three of Jk1 sequences. Intheory, inversion provides the maximum flexibility for cells tosubsequently correct Igk rearrangements that are nonfunctional,underexpressed, or autoreactive, which collectively constitute thevast majority of rearrangements. Hence, the heavy usage and Jk1skewing of the distal, inversionally rearranging 10-96 and 19-93
genes might be important in initial scrambling of the locus toallow a more efficient and wider range of secondary recombina-tions than would otherwise occur. Without such effects, there ispredicted to be strong bias for usage of distal Vks with proximalJks and vice versa, which we failed to see.What mechanism accounts for the dominance of certain Vk
genes? This skewing could not be readily attributed to predictedrecombination signal strength, proximity to the Jk elements, orsomatic cell selection. For example, 10-96 carries a recombinationsignal identical to those of adjacent10-94 and 10-95 genes but isused much more frequently. In BALB/c mice, 10-96 is also vastlyoverrepresented compared with 10-95, despite carrying function-ally indistinguishable promoter elements and recombination sig-nals (55). Our conclusion that many of the dominant genes havepromoter elements that are indistinguishable from their less wellexpressed counterparts comes from the detailed bioinformaticanalysis of Brekke and Garrard (8), who annotated the Igk locuson the basis of data largely collected by Zachau’s group and thegenome project (9). No real differences were seen in this analysisbetween highly expressed and poorly expressed family membersin any promoter feature characterized (8). Surprisingly, major Vkusage was also not appreciably altered in the BM of the gene-modified strains studied in this paper, including in pUIik mice,which are subject to strong negative selection. Collectively, theseresults suggest that rearrangement of the major Vk genes is stronglypreferred as a result of enhanced accessibility to rearrangement inpre-B cells.Ig gene accessibility to rearrangement is complex and multi-
factorial, because it is affected by chromatin structure, histoneacetylation, DNA methylation, and conventional enhancer andpromoter activity, in addition to the effects of antisense and sensegermline transcription (56–58). One possibility for how differen-tial accessibility is controlled genetically is that cis-acting ele-ments adjacent to frequently used genes are preferentially rec-ognized by transcription factors. Some exceptional features inVk4 family genes have been noted, including clusters of potentialbinding sites for Lef-1, Ebf, and Oct1/2 in the upstream, intron,and downstream regions of the RSS with functional significance intargeting for V–J recombination (8). In some studies, Vk4 familygenes had elevated usage and skewed Jk association, particularlyin fetal cells (33, 42–44, 53). However, in this study of youngadult mice, we found a somewhat reduced average usage per Vk4family gene. We could not find significant differences between sixof the seven frequently used genes compared with other genes intheir families in terms of predicted transcription factor binding
FIGURE 7. Analysis of the diversity of the expressed k-chain repertoire in B6 BM. (A) The integrated frequency of Vk usage is plotted in declining order
of usage frequency. *The contributions of the seven major genes, yielding a 43% integrated frequency of these genes. (B) CDR3 diversity produced by all
VJ associations is shown. *Total CDR3 aa variety contributed by the major seven genes.
2312 SKEWED Igk V GENE USAGE AND V–J ASSOCIATIONS IN B6 MICE
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

sites or recombination signals. The seventh gene, 19-93, is uniquein that it is a member of a single gene family. 19-93 has an extraoctamer (Oct-binding) site upstream of the conserved one close tothe TATA box, possibly explaining its elevated usage. Other no-table features of 19-93 were its preferred association with Jk1 andthat its usage was not diminished in abundance or skewed much inits Jk association in cells predicted to undergo more editing, suchas in pUIik BM B cells. The explanation for this is not clear butcould come about from an idiosyncrasy of the locus organization:19-93 rearranges to Jk1 by inversion, which then puts the un-derused Vk4 family proximal to the JkCk cluster and frequentlyused Vk6 genes distal, potentially lowering the chances of sec-ondary recombinations editing away the 19-93/Jk1 join.The frequently used Vk19/28 family genes 6-23, 6-17, and 6-15
are adjacent to Odc1-like pseudogenes in an opposite transcrip-tional orientation. Nearby 6-14 and 6-13 genes have the samefeatures, but are used less frequently. Other non-Ig pseudogenesare also near 1-135, and several other less well used genes. Thesefeatures raise the possibility of a regulatory function of noncodingRNAs originating in adjacent pseudogenes, be they Vk or non-Ig.Although sterile transcription of germline V coding gene segmentswas shown many years ago to correlate with accessibility to V(D)Jrecombination (59), noncoding RNA has recently been implicatedin actively promoting accessibility. Bolland et al. (60) identified inearly B cells genic and intergenic antisense transcripts through-out the IGHV region that were strictly upregulated at the transi-tion from DH-to-JH to VH-to-DJH recombination. Abarrategui andKrangel (61) showed that transcriptional elongation regulates theprogrammed accessibility of TCR Ja genes. Such transcripts maypromote chromatin accessibility and allow RAG binding.An additional possibility for how preferential Vk usage is reg-
ulated relates to chromatin remodeling mediated by longer rangelooping interactions by CTCF, cohesins, and associated proteins(40, 58, 62–65). Locus contraction is believed to facilitate Ig generearrangements (39, 66). Locus looping might promote accessi-bility or bring separated gene segments closer together, facilitatingrearrangements. Alternatively, loop tethers may serve as insulatorsthat delimit Vk groups, which might compete with one anotherfor rearrangement. In the Igh locus, CTCF binding has been im-plicated in both limiting and facilitating rearrangements to VH
through downregulation of transcriptional read-through and re-tention of DNA associate with heterochromatin, and by facilitat-ing locus contraction and element colocalization (62–64). CTCFbinding sites within the Igh and Igk loci clearly correlate withproximity to VH elements but not to Vk elements (40, 58, 62).Also, CTCF sites are not particularly close to frequently usedVks (Supplemental Fig. 1). One CTCF binding site is associatedwith the recombination silencer Sis (67) located in the Vk-Jkintron (40, 65). Regulation involving CTCF might involve insu-lator function, as proposed by Hendriks and colleagues (40), whoshowed dramatic changes in Vk usage in B cells that were con-ditionally deficient in CTCF, leading to an increase in proximalVk usage at the expense of distal usage. For example, use of 1-135might be enhanced because it is literally insulated from compe-tition for recombination by looping anchored by a CTCF bindingsite ∼35 kb downstream (Fig. 8), which in turn might be tetherednear the intronic CTCF. (Alternatively or in addition, the distalend of the locus may carry an enhancer or LCR element providingan additional tether.) A similar argument could potentially explainthe usage pattern of other genes. Two strong CTCF binding sitesflank a total of 30 Vk elements that includes all of the Vk4 familymembers. If the two sites form the base of a large loop with accessto the JCk cluster, Vk4 family genes may compete with each otherwithin the loop, possibly explaining the relatively low frequency
of Vk4 family usage per gene (Fig. 8). However, there appear tobe many additional lower occupancy CTCF sites among Vk4family genes as well (41).The 1-135 gene had many unexpected features, including high-
frequency usage of 1-135/Jk1 in BM, increased frequency in theBM B cells of JCk+/2 and pUlik mice, counterselection from BMto SP, and elevated proportion of nonfunctional sequences in BM.In particular, out-of-frame rearrangements to Jk1 should be rap-idly followed by secondary rearrangements. We ascribe thesespecial features of 1-135 to its chromosomal position rather thanto its protein sequence. Because1-135 is located in the penultimateposition on the distal end of the locus and undergoes deletionalrearrangement, subsequent rearrangement or editing on that alleleis limited just to the rarely used Vk gene 2-137. Apoptosis ofautoreactive B cells is a slow process preceded by a time windowin which editing takes place, so autoreactive 1-135–expressing“dead-end” cells unable to edit further should tend to build up inBM but be barred from release to the peripheral immune system.A striking finding from this study was the overall similarities
in k gene segment usage in BM B cells from pUIik, RS2/2, andJCk+/2 mice compared with wild-type BM B cells. Although somesignificant differences were documented, they were relativelysubtle. We had expected the overall Jk usage and V–J associationsto be more strikingly skewed, particularly in pUIik mice, in whichediting is massively induced (35). One explanation is that sec-ondary rearrangements are already extensive in wild-type cells asa result of the high proportion of nonfunctional rearrangements(more than two of three) and additional editing of functional Lchains as a result of autoreactivity, nonassociation with H chain,and suboptimal expression. Hence, additional editing may havea relatively subtle overall impact over this high background.When a B cell’s Ig H chain is fixed by transgenesis, L chain se-lection is strikingly revealed. For example, in anti-DNA 3H9 Hchain transgenic mice, several of the normally dominant Vks arestrongly underrepresented in peripheral B cells, and major V–Jassociations are also skewed (22, 38, 68). In mice with a diverserepertoire of H chains, these skewings may balance out, ultimately
FIGURE 8. Potential looping of Igk locus on the basis of the reported
CTCF binding sites. Cartoon depicts Igk locus assuming clustering of
major CTCF sites, highlighting distribution of major Vks and Vk4 family
on loops. CTCF sites, blue circles; major Vk genes, yellow arrows; Vk
genes used at 1–3.9% of total, medium sized green arrows; and infre-
quently used Vk genes (,1% of total), small red arrows. Jks and Ck are
indicated by pink symbols and blue arrow, respectively. RS, blue solid
diamond.
The Journal of Immunology 2313
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

reflecting underlying rearrangement preferences. A second ex-planation is that autoreactive B cells in BM may be actually over-represented because they transit more slowly through that com-partment (35) and are especially enriched among BM B cells thathave downregulated sIgM (69).
AcknowledgmentsWe thank Eline Luning Prak (University of Pennsylvania, Philadelphia,
PA), Kees Murre (University of California at San Diego, La Jolla, CA),
and Ann Feeney (The Scripps Research Institute, La Jolla, CA) for helpful
suggestions and Ann Feeney, Rudi Hendriks (Erasmus University, Rotter-
dam, The Netherlands), Kazuko Miyazaki (University of California at San
Diego), and Kees Murre for data on CTCF binding sites. We also thank
Steven Head of the Sequencing Core (The Scripps Research Institute).
DisclosuresThe authors have no financial conflicts of interest.
References1. Apel, T. W., A. Scherer, T. Adachi, D. Auch, M. Ayane, and M. Reth. 1995. The
ribose 5-phosphate isomerase-encoding gene is located immediately downstreamfrom that encoding murine immunoglobulin k. Gene 156: 191–197.
2. George, J. B., S. Li, and W. T. Garrard. 1995. Yeast artificial chromosomecontigs reveal that distal variable-region genes reside at least 3 megabases fromthe joining regions in the murine immunoglobulin k locus. Proc. Natl. Acad. Sci.USA 92: 12421–12425.
3. Schable, K. F., R. Thiebe, A. Bensch, J. Brensing-Kuppers, V. Heim,T. Kirschbaum, R. Lamm, M. Ohnrich, S. Pourrajabi, F. Roschenthaler, et al.1999. Characteristics of the immunoglobulin Vk genes, pseudogenes, relics andorphons in the mouse genome. Eur. J. Immunol. 29: 2082–2086.
4. Thiebe, R., K. F. Schable, A. Bensch, J. Brensing-Kuppers, V. Heim,T. Kirschbaum, H. Mitlohner, M. Ohnrich, S. Pourrajabi, F. Roschenthaler, et al.1999. The variable genes and gene families of the mouse immunoglobulin klocus. Eur. J. Immunol. 29: 2072–2081.
5. Roschenthaler, F., T. Kirschbaum, V. Heim, V. Kirschbaum, K. F. Schable,J. Schwendinger, I. Zocher, and H. G. Zachau. 1999. The 59 part of the mouseimmunoglobulin k locus. Eur. J. Immunol. 29: 2065–2071.
6. Kirschbaum, T., F. Roschenthaler, A. Bensch, B. Holscher, A. Lautner-Rieske,M. Ohnrich, S. Pourrajabi, J. Schwendinger, I. Zocher, and H. G. Zachau. 1999.The central part of the mouse immunoglobulin k locus. Eur. J. Immunol. 29:2057–2064.
7. Gerdes, T., and M. Wabl. 2002. Physical map of the mouse l light chain andrelated loci. Immunogenetics 54: 62–65.
8. Brekke, K. M., and W. T. Garrard. 2004. Assembly and analysis of the mouseimmunoglobulin k gene sequence. Immunogenetics 56: 490–505.
9. Johnston, C. M., A. L. Wood, D. J. Bolland, and A. E. Corcoran. 2006. Completesequence assembly and characterization of the C57BL/6 mouse Ig heavy chain Vregion. J. Immunol. 176: 4221–4234.
10. Retter, I., C. Chevillard, M. Scharfe, A. Conrad, M. Hafner, T. H. Im,M. Ludewig, G. Nordsiek, S. Severitt, S. Thies, et al. 2007. Sequence andcharacterization of the Ig heavy chain constant and partial variable region of themouse strain 129S1. J. Immunol. 179: 2419–2427.
11. Martinez-Jean, C., G. Folch, and M. P. Lefranc. 2001. Nomenclature and over-view of the mouse (Mus musculus and Mus sp.) immunoglobulin k (IGK) genes.Exp. Clin. Immunogenet. 18: 255–279.
12. de Bono, B., M. Madera, and C. Chothia. 2004. VH gene segments in the mouseand human genomes. J. Mol. Biol. 342: 131–143.
13. Max, E. E., J. G. Seidman, and P. Leder. 1979. Sequences of five potential re-combination sites encoded close to an immunoglobulin k constant region gene.Proc. Natl. Acad. Sci. USA 76: 3450–3454.
14. Brodeur, P. H., and R. Riblet. 1984. The immunoglobulin heavy chain variableregion (Igh-V) locus in the mouse. I. One hundred Igh-V genes comprise sevenfamilies of homologous genes. Eur. J. Immunol. 14: 922–930.
15. Strohal, R., A. Helmberg, G. Kroemer, and R. Kofler. 1989. Mouse Vk geneclassification by nucleic acid sequence similarity. Immunogenetics 30: 475–493.
16. Lefranc, M. P., V. Giudicelli, C. Ginestoux, J. Jabado-Michaloud, G. Folch,F. Bellahcene, Y. Wu, E. Gemrot, X. Brochet, J. Lane, et al. 2009. IMGT, theinternational ImMunoGeneTics information system. Nucleic Acids Res. 37(Da-tabase issue): D1006–D1012.
17. Tonegawa, S. 1983. Somatic generation of antibody diversity. Nature 302: 575–581.
18. Nemazee, D. 2006. Receptor editing in lymphocyte development and centraltolerance. Nat. Rev. Immunol. 6: 728–740.
19. Wood, D. L., and C. Coleclough. 1984. Different joining region J elements of themurine k immunoglobulin light chain locus are used at markedly different fre-quencies. Proc. Natl. Acad. Sci. USA 81: 4756–4760.
20. Prak, E. L., M. Trounstine, D. Huszar, and M. Weigert. 1994. Light chain editingin k-deficient animals: a potential mechanism of B cell tolerance. J. Exp. Med.180: 1805–1815.
21. Constantinescu, A., and M. S. Schlissel. 1997. Changes in locus-specific V(D)Jrecombinase activity induced by immunoglobulin gene products during B celldevelopment. J. Exp. Med. 185: 609–620.
22. Radic, M. Z., J. Erikson, S. Litwin, and M. Weigert. 1993. B lymphocytes mayescape tolerance by revising their antigen receptors. J. Exp. Med. 177: 1165–1173.
23. Durdik, J., M. W. Moore, and E. Selsing. 1984. Novel k light-chain gene rear-rangements in mouse l light chain-producing B lymphocytes. Nature 307: 749–752.
24. Siminovitch, K. A., M. W. Moore, J. Durdik, and E. Selsing. 1987. The human kdeleting element and the mouse recombining segment share DNA sequencehomology. Nucleic Acids Res. 15: 2699–2705.
25. Retter, M. W., and D. Nemazee. 1998. Receptor editing occurs frequently duringnormal B cell development. J. Exp. Med. 188: 1231–1238.
26. Harada, K., and H. Yamagishi. 1991. Lack of feedback inhibition of Vk generearrangement by productively rearranged alleles. J. Exp. Med. 173: 409–415.
27. Brauninger, A., T. Goossens, K. Rajewsky, and R. Kuppers. 2001. Regulation ofimmunoglobulin light chain gene rearrangements during early B cell develop-ment in the human. Eur. J. Immunol. 31: 3631–3637.
28. Selsing, E., and L. E. Daitch. 1995. Immunoglobulin l genes. In ImmunoglubulinGenes. T. Honjo, and F. Alt, eds. Acadenic Press Limited, London, p. 194–203.
29. Dunda, O., and D. Corcos. 1997. Recombining sequence recombination innormal k-chain–expressing B cells. J. Immunol. 159: 4362–4366.
30. Vela, J. L., D. Aıt-Azzouzene, B. H. Duong, T. Ota, and D. Nemazee. 2008.Rearrangement of mouse immunoglobulin k deleting element recombining se-quence promotes immune tolerance and l B cell production. Immunity 28: 161–170.
31. Wu, T. T., and E. A. Kabat. 1970. An analysis of the sequences of the variableregions of Bence Jones proteins and myeloma light chains and their implicationsfor antibody complementarity. J. Exp. Med. 132: 211–250.
32. Kofler, R., S. Geley, H. Kofler, and A. Helmberg. 1992. Mouse variable-regiongene families: complexity, polymorphism and use in non-autoimmune responses.Immunol. Rev. 128: 5–21.
33. Kalled, S. L., and P. H. Brodeur. 1991. Utilization of Vk families and Vk exons:implications for the available B cell repertoire. J. Immunol. 147: 3194–3200.
34. Kabat, E. A., T. T. Wu, H. M. Perry, K. S. Gottesman, and C. Foeller. 1991.Sequences of proteins of Immunological Interest. U.S. Department of Health andHuman Services, Bethesda, MD.
35. Aıt-Azzouzene, D., L. Verkoczy, J. Peters, A. Gavin, P. Skog, J. L. Vela, andD. Nemazee. 2005. An immunoglobulin Ck-reactive single chain antibody fu-sion protein induces tolerance through receptor editing in a normal polyclonalimmune system. J. Exp. Med. 201: 817–828.
36. Chen, J., M. Trounstine, C. Kurahara, F. Young, C. C. Kuo, Y. Xu, J. F. Loring,F. W. Alt, and D. Huszar. 1993. B cell development in mice that lack one or bothimmunoglobulin k light chain genes. EMBO J. 12: 821–830.
37. Altschul, S. F., W. Gish, W. Miller, E. W. Myers, and D. J. Lipman. 1990. Basiclocal alignment search tool. J. Mol. Biol. 215: 403–410.
38. Ota, M., B. H. Duong, A. Torkamani, C. M. Doyle, A. L. Gavin, T. Ota, andD. Nemazee. 2010. Regulation of the B cell receptor repertoire and self-reactivity by BAFF. J. Immunol. 185: 4128–4136.
39. Jhunjhunwala, S., M. C. van Zelm, M. M. Peak, S. Cutchin, R. Riblet, J. J. vanDongen, F. G. Grosveld, T. A. Knoch, and C. Murre. 2008. The 3D structure ofthe immunoglobulin heavy-chain locus: implications for long-range genomicinteractions. Cell 133: 265–279.
40. Ribeiro de Almeida, C., R. Stadhouders, M. J. de Bruijn, I. M. Bergen,S. Thongjuea, B. Lenhard, W. van Ijcken, F. Grosveld, N. Galjart, E. Soler, andR. W. Hendriks. 2011. The DNA-binding protein CTCF limits proximal Vkrecombination and restricts k enhancer interactions to the immunoglobulin klight chain locus. Immunity 35: 501–513.
41. Whitcomb, E. A., and P. H. Brodeur. 1998. Rearrangement and selection in thedeveloping Vk repertoire of the mouse: an analysis of the usage of two Vk genesegments. J. Immunol. 160: 4904–4913.
42. Kalled, S. L., and P. H. Brodeur. 1990. Preferential rearrangement of Vk4 genesegments in pre-B cell lines. J. Exp. Med. 172: 559–566.
43. Medina, C. A., and J. M. Teale. 1993. Restricted k chain expression in earlyontogeny: biased utilization of Vk exons and preferential Vk-Jk recombinations.J. Exp. Med. 177: 1317–1330.
44. Ramsden, D. A., C. J. Paige, and G. E. Wu. 1994. k Light chain rearrangement inmouse fetal liver. J. Immunol. 153: 1150–1160.
45. Hesse, J. E., M. R. Lieber, K. Mizuuchi, and M. Gellert. 1989. V(D)J recom-bination: a functional definition of the joining signals. Genes Dev. 3: 1053–1061.
46. Chemin, G., A. Tinguely, C. Sirac, F. Lechouane, S. Duchez, M. Cogne, andL. Delpy. 2010. Multiple RNA surveillance mechanisms cooperate to reduce theamount of nonfunctional Ig k transcripts. J. Immunol. 184: 5009–5017.
47. Kaushik, A., D. H. Schulze, C. Bona, and G. Kelsoe. 1989. Murine Vk geneexpression does not follow the VH paradigm. J. Exp. Med. 169: 1859–1864.
48. Lawler, A. M., J. F. Kearney, M. Kuehl, and P. J. Gearhart. 1989. Early rear-rangements of genes encoding murine immunoglobulin k chains, unlike genesencoding heavy chains, use variable gene segments dispersed throughout thelocus. Proc. Natl. Acad. Sci. USA 86: 6744–6747.
49. Cowell, L. G., M. Davila, K. Yang, T. B. Kepler, and G. Kelsoe. 2003. Pro-spective estimation of recombination signal efficiency and identification offunctional cryptic signals in the genome by statistical modeling. J. Exp. Med.197: 207–220.
50. Gerstein, R. M., and M. R. Lieber. 1993. Coding end sequence can markedlyaffect the initiation of V(D)J recombination. Genes Dev. 7(7B): 1459–1469.
2314 SKEWED Igk V GENE USAGE AND V–J ASSOCIATIONS IN B6 MICE
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

51. Yu, K., and M. R. Lieber. 1999. Mechanistic basis for coding end sequenceeffects in the initiation of V(D)J recombination. Mol. Cell. Biol. 19: 8094–8102.
52. Lee, A. I., S. D. Fugmann, L. G. Cowell, L. M. Ptaszek, G. Kelsoe, andD. G. Schatz. 2003. A functional analysis of the spacer of V(D)J recombinationsignal sequences. PLoS Biol. 1: E1.
53. Milstein, C., J. Even, J. M. Jarvis, A. Gonzalez-Fernandez, and E. Gherardi.1992. Non-random features of the repertoire expressed by the members of oneVk gene family and of the V-J recombination. [Published erratum appears in1992 Eur. J. Immunol. 22:1958.] Eur. J. Immunol. 22: 1627–1634.
54. Hardy, R. R., P. W. Kincade, and K. Dorshkind. 2007. The protean nature of cellsin the B lymphocyte lineage. Immunity 26: 703–714.
55. Fitzsimmons, S. P., B. T. Rotz, and M. A. Shapiro. 1998. Asymmetric contri-bution to Ig repertoire diversity by Vk exons: differences in the utilization ofVk10 exons. J. Immunol. 161: 2290–2300.
56. Schatz, D. G., and Y. Ji. 2011. Recombination centres and the orchestration ofV(D)J recombination. Nat. Rev. Immunol. 11: 251–263.
57. Corcoran, A. E. 2010. The epigenetic role of non-coding RNA transcription andnuclear organization in immunoglobulin repertoire generation. Semin. Immunol.22: 353–361.
58. Degner-Leisso, S. C., and A. J. Feeney. 2010. Epigenetic and 3-dimensionalregulation of V(D)J rearrangement of immunoglobulin genes. Semin. Immunol.22: 346–352.
59. Yancopoulos, G. D., and F. W. Alt. 1985. Developmentally controlled and tissue-specific expression of unrearranged VH gene segments. Cell 40: 271–281.
60. Bolland, D. J., A. L. Wood, C. M. Johnston, S. F. Bunting, G. Morgan,L. Chakalova, P. J. Fraser, and A. E. Corcoran. 2004. Antisense intergenictranscription in V(D)J recombination. Nat. Immunol. 5: 630–637.
61. Abarrategui, I., and M. S. Krangel. 2006. Regulation of T cell receptor-a generecombination by transcription. Nat. Immunol. 7: 1109–1115.
62. Lucas, J. S., C. Bossen, and C. Murre. 2011. Transcription and recombinationfactories: common features? Curr. Opin. Cell Biol. 23: 318–324.
63. Guo, C., H. S. Yoon, A. Franklin, S. Jain, A. Ebert, H. L. Cheng, E. Hansen,O. Despo, C. Bossen, C. Vettermann, et al. 2011. CTCF-binding elements me-diate control of V(D)J recombination. Nature 477: 424–430.
64. Degner, S. C., J. Verma-Gaur, T. P. Wong, C. Bossen, G. M. Iverson,A. Torkamani, C. Vettermann, Y. C. Lin, Z. Ju, D. Schulz, et al. 2011. CCCTC-binding factor (CTCF) and cohesin influence the genomic architecture of the Ighlocus and antisense transcription in pro-B cells. Proc. Natl. Acad. Sci. USA 108:9566–9571.
65. Degner, S. C., T. P. Wong, G. Jankevicius, and A. J. Feeney. 2009. Cutting edge:developmental stage-specific recruitment of cohesin to CTCF sites throughoutimmunoglobulin loci during B lymphocyte development. J. Immunol. 182: 44–48.
66. Roldan, E., M. Fuxa, W. Chong, D. Martinez, M. Novatchkova, M. Busslinger,and J. A. Skok. 2005. Locus “decontraction” and centromeric recruitment con-tribute to allelic exclusion of the immunoglobulin heavy-chain gene. Nat.Immunol. 6: 31–41.
67. Liu, Z., P. Widlak, Y. Zou, F. Xiao, M. Oh, S. Li, M. Y. Chang, J. W. Shay, andW. T. Garrard. 2006. A recombination silencer that specifies heterochromatinpositioning and ikaros association in the immunoglobulin k locus. Immunity 24:405–415.
68. Ibrahim, S. M., M. Weigert, C. Basu, J. Erikson, and M. Z. Radic. 1995. Lightchain contribution to specificity in anti-DNA antibodies. J. Immunol. 155: 3223–3233.
69. Duong, B. H., T. Ota, M. Aoki-Ota, A. B. Cooper, D. Ait-Azzouzene, J. L. Vela,A. L. Gavin, and D. Nemazee. 2011. Negative selection by IgM superantigendefines a B cell central tolerance compartment and reveals mutations allowingescape. J. Immunol. 187: 5596–5605.
The Journal of Immunology 2315
by guest on January 11, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
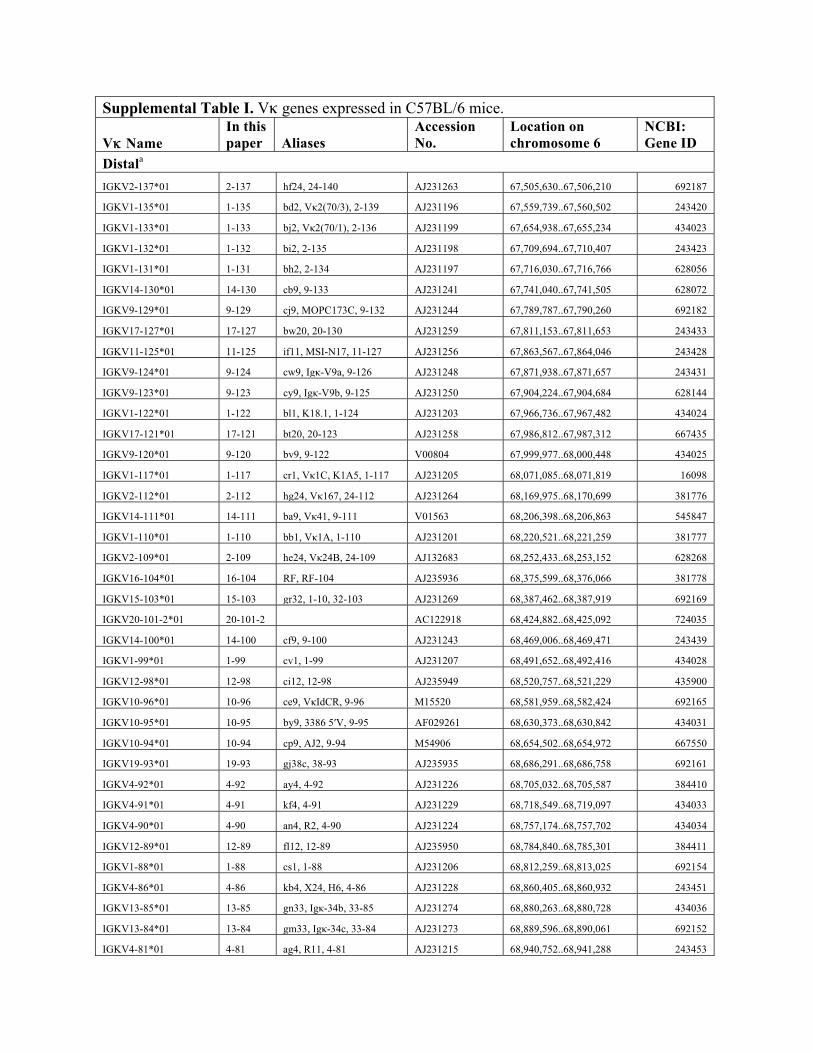
Supplemental Table I. Vκ genes expressed in C57BL/6 mice.
Vκκ Name In this paper Aliases
Accession No.
Location on chromosome 6
NCBI: Gene ID
Distala
IGKV2-137*01 2-137 hf24, 24-140 AJ231263 67,505,630..67,506,210 692187
IGKV1-135*01 1-135 bd2, Vκ2(70/3), 2-139 AJ231196 67,559,739..67,560,502 243420
IGKV1-133*01 1-133 bj2, Vκ2(70/1), 2-136 AJ231199 67,654,938..67,655,234 434023
IGKV1-132*01 1-132 bi2, 2-135 AJ231198 67,709,694..67,710,407 243423
IGKV1-131*01 1-131 bh2, 2-134 AJ231197 67,716,030..67,716,766 628056
IGKV14-130*01 14-130 cb9, 9-133 AJ231241 67,741,040..67,741,505 628072
IGKV9-129*01 9-129 cj9, MOPC173C, 9-132 AJ231244 67,789,787..67,790,260 692182
IGKV17-127*01 17-127 bw20, 20-130 AJ231259 67,811,153..67,811,653 243433
IGKV11-125*01 11-125 if11, MSI-N17, 11-127 AJ231256 67,863,567..67,864,046 243428
IGKV9-124*01 9-124 cw9, Igκ-V9a, 9-126 AJ231248 67,871,938..67,871,657 243431
IGKV9-123*01 9-123 cy9, Igκ-V9b, 9-125 AJ231250 67,904,224..67,904,684 628144
IGKV1-122*01 1-122 bl1, K18.1, 1-124 AJ231203 67,966,736..67,967,482 434024
IGKV17-121*01 17-121 bt20, 20-123 AJ231258 67,986,812..67,987,312 667435
IGKV9-120*01 9-120 bv9, 9-122 V00804 67,999,977..68,000,448 434025
IGKV1-117*01 1-117 cr1, Vκ1C, K1A5, 1-117 AJ231205 68,071,085..68,071,819 16098
IGKV2-112*01 2-112 hg24, Vκ167, 24-112 AJ231264 68,169,975..68,170,699 381776
IGKV14-111*01 14-111 ba9, Vκ41, 9-111 V01563 68,206,398..68,206,863 545847
IGKV1-110*01 1-110 bb1, Vκ1A, 1-110 AJ231201 68,220,521..68,221,259 381777
IGKV2-109*01 2-109 he24, Vκ24B, 24-109 AJ132683 68,252,433..68,253,152 628268
IGKV16-104*01 16-104 RF, RF-104 AJ235936 68,375,599..68,376,066 381778
IGKV15-103*01 15-103 gr32, 1-10, 32-103 AJ231269 68,387,462..68,387,919 692169
IGKV20-101-2*01 20-101-2 AC122918 68,424,882..68,425,092 724035
IGKV14-100*01 14-100 cf9, 9-100 AJ231243 68,469,006..68,469,471 243439
IGKV1-99*01 1-99 cv1, 1-99 AJ231207 68,491,652..68,492,416 434028
IGKV12-98*01 12-98 ci12, 12-98 AJ235949 68,520,757..68,521,229 435900
IGKV10-96*01 10-96 ce9, VκIdCR, 9-96 M15520 68,581,959..68,582,424 692165
IGKV10-95*01 10-95 by9, 3386 5′V, 9-95 AF029261 68,630,373..68,630,842 434031
IGKV10-94*01 10-94 cp9, AJ2, 9-94 M54906 68,654,502..68,654,972 667550
IGKV19-93*01 19-93 gj38c, 38-93 AJ235935 68,686,291..68,686,758 692161
IGKV4-92*01 4-92 ay4, 4-92 AJ231226 68,705,032..68,705,587 384410
IGKV4-91*01 4-91 kf4, 4-91 AJ231229 68,718,549..68,719,097 434033
IGKV4-90*01 4-90 an4, R2, 4-90 AJ231224 68,757,174..68,757,702 434034
IGKV12-89*01 12-89 fl12, 12-89 AJ235950 68,784,840..68,785,301 384411
IGKV1-88*01 1-88 cs1, 1-88 AJ231206 68,812,259..68,813,025 692154
IGKV4-86*01 4-86 kb4, X24, H6, 4-86 AJ231228 68,860,405..68,860,932 243451
IGKV13-85*01 13-85 gn33, Igκ-34b, 33-85 AJ231274 68,880,263..68,880,728 434036
IGKV13-84*01 13-84 gm33, Igκ-34c, 33-84 AJ231273 68,889,596..68,890,061 692152
IGKV4-81*01 4-81 ag4, R11, 4-81 AJ231215 68,940,752..68,941,288 243453
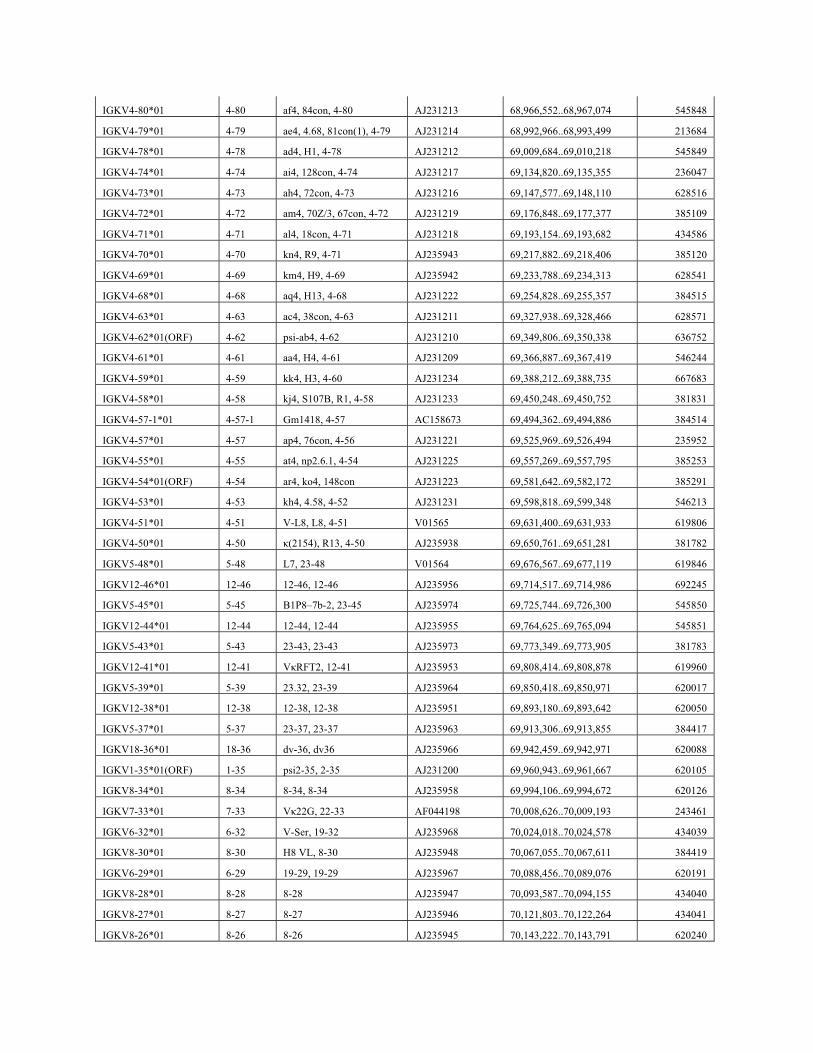
IGKV4-80*01 4-80 af4, 84con, 4-80 AJ231213 68,966,552..68,967,074 545848
IGKV4-79*01 4-79 ae4, 4.68, 81con(1), 4-79 AJ231214 68,992,966..68,993,499 213684
IGKV4-78*01 4-78 ad4, H1, 4-78 AJ231212 69,009,684..69,010,218 545849
IGKV4-74*01 4-74 ai4, 128con, 4-74 AJ231217 69,134,820..69,135,355 236047
IGKV4-73*01 4-73 ah4, 72con, 4-73 AJ231216 69,147,577..69,148,110 628516
IGKV4-72*01 4-72 am4, 70Z/3, 67con, 4-72 AJ231219 69,176,848..69,177,377 385109
IGKV4-71*01 4-71 al4, 18con, 4-71 AJ231218 69,193,154..69,193,682 434586
IGKV4-70*01 4-70 kn4, R9, 4-71 AJ235943 69,217,882..69,218,406 385120
IGKV4-69*01 4-69 km4, H9, 4-69 AJ235942 69,233,788..69,234,313 628541
IGKV4-68*01 4-68 aq4, H13, 4-68 AJ231222 69,254,828..69,255,357 384515
IGKV4-63*01 4-63 ac4, 38con, 4-63 AJ231211 69,327,938..69,328,466 628571
IGKV4-62*01(ORF) 4-62 psi-ab4, 4-62 AJ231210 69,349,806..69,350,338 636752
IGKV4-61*01 4-61 aa4, H4, 4-61 AJ231209 69,366,887..69,367,419 546244
IGKV4-59*01 4-59 kk4, H3, 4-60 AJ231234 69,388,212..69,388,735 667683
IGKV4-58*01 4-58 kj4, S107B, R1, 4-58 AJ231233 69,450,248..69,450,752 381831
IGKV4-57-1*01 4-57-1 Gm1418, 4-57 AC158673 69,494,362..69,494,886 384514
IGKV4-57*01 4-57 ap4, 76con, 4-56 AJ231221 69,525,969..69,526,494 235952
IGKV4-55*01 4-55 at4, np2.6.1, 4-54 AJ231225 69,557,269..69,557,795 385253
IGKV4-54*01(ORF) 4-54 ar4, ko4, 148con AJ231223 69,581,642..69,582,172 385291
IGKV4-53*01 4-53 kh4, 4.58, 4-52 AJ231231 69,598,818..69,599,348 546213
IGKV4-51*01 4-51 V-L8, L8, 4-51 V01565 69,631,400..69,631,933 619806
IGKV4-50*01 4-50 κ(2154), R13, 4-50 AJ235938 69,650,761..69,651,281 381782
IGKV5-48*01 5-48 L7, 23-48 V01564 69,676,567..69,677,119 619846
IGKV12-46*01 12-46 12-46, 12-46 AJ235956 69,714,517..69,714,986 692245
IGKV5-45*01 5-45 B1P8–7b-2, 23-45 AJ235974 69,725,744..69,726,300 545850
IGKV12-44*01 12-44 12-44, 12-44 AJ235955 69,764,625..69,765,094 545851
IGKV5-43*01 5-43 23-43, 23-43 AJ235973 69,773,349..69,773,905 381783
IGKV12-41*01 12-41 VκRFT2, 12-41 AJ235953 69,808,414..69,808,878 619960
IGKV5-39*01 5-39 23.32, 23-39 AJ235964 69,850,418..69,850,971 620017
IGKV12-38*01 12-38 12-38, 12-38 AJ235951 69,893,180..69,893,642 620050
IGKV5-37*01 5-37 23-37, 23-37 AJ235963 69,913,306..69,913,855 384417
IGKV18-36*01 18-36 dv-36, dv36 AJ235966 69,942,459..69,942,971 620088
IGKV1-35*01(ORF) 1-35 psi2-35, 2-35 AJ231200 69,960,943..69,961,667 620105
IGKV8-34*01 8-34 8-34, 8-34 AJ235958 69,994,106..69,994,672 620126
IGKV7-33*01 7-33 Vκ22G, 22-33 AF044198 70,008,626..70,009,193 243461
IGKV6-32*01 6-32 V-Ser, 19-32 AJ235968 70,024,018..70,024,578 434039
IGKV8-30*01 8-30 H8 VL, 8-30 AJ235948 70,067,055..70,067,611 384419
IGKV6-29*01 6-29 19-29, 19-29 AJ235967 70,088,456..70,089,076 620191
IGKV8-28*01 8-28 8-28 AJ235947 70,093,587..70,094,155 434040
IGKV8-27*01 8-27 8-27 AJ235946 70,121,803..70,122,264 434041
IGKV8-26*01 8-26 8-26 AJ235945 70,143,222..70,143,791 620240
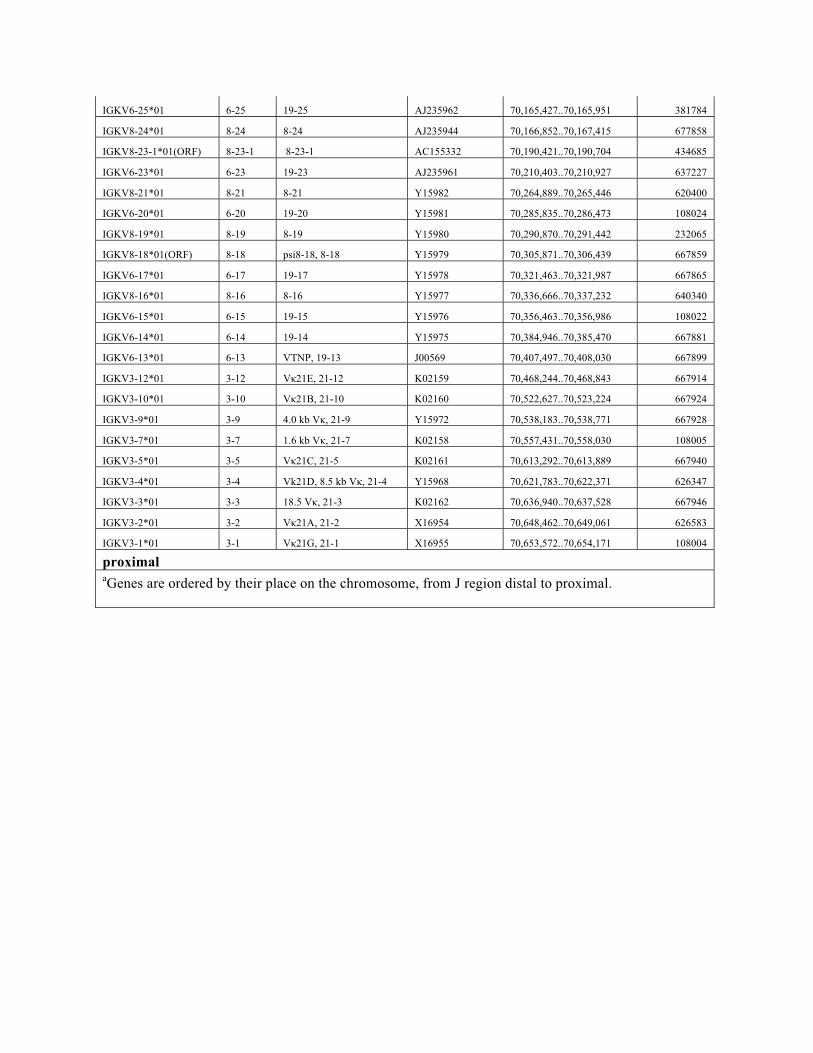
IGKV6-25*01 6-25 19-25 AJ235962 70,165,427..70,165,951 381784
IGKV8-24*01 8-24 8-24 AJ235944 70,166,852..70,167,415 677858
IGKV8-23-1*01(ORF) 8-23-1 8-23-1 AC155332 70,190,421..70,190,704 434685
IGKV6-23*01 6-23 19-23 AJ235961 70,210,403..70,210,927 637227
IGKV8-21*01 8-21 8-21 Y15982 70,264,889..70,265,446 620400
IGKV6-20*01 6-20 19-20 Y15981 70,285,835..70,286,473 108024
IGKV8-19*01 8-19 8-19 Y15980 70,290,870..70,291,442 232065
IGKV8-18*01(ORF) 8-18 psi8-18, 8-18 Y15979 70,305,871..70,306,439 667859
IGKV6-17*01 6-17 19-17 Y15978 70,321,463..70,321,987 667865
IGKV8-16*01 8-16 8-16 Y15977 70,336,666..70,337,232 640340
IGKV6-15*01 6-15 19-15 Y15976 70,356,463..70,356,986 108022
IGKV6-14*01 6-14 19-14 Y15975 70,384,946..70,385,470 667881
IGKV6-13*01 6-13 VTNP, 19-13 J00569 70,407,497..70,408,030 667899
IGKV3-12*01 3-12 Vκ21E, 21-12 K02159 70,468,244..70,468,843 667914
IGKV3-10*01 3-10 Vκ21B, 21-10 K02160 70,522,627..70,523,224 667924
IGKV3-9*01 3-9 4.0 kb Vκ, 21-9 Y15972 70,538,183..70,538,771 667928
IGKV3-7*01 3-7 1.6 kb Vκ, 21-7 K02158 70,557,431..70,558,030 108005
IGKV3-5*01 3-5 Vκ21C, 21-5 K02161 70,613,292..70,613,889 667940
IGKV3-4*01 3-4 Vk21D, 8.5 kb Vκ, 21-4 Y15968 70,621,783..70,622,371 626347
IGKV3-3*01 3-3 18.5 Vκ, 21-3 K02162 70,636,940..70,637,528 667946
IGKV3-2*01 3-2 Vκ21A, 21-2 X16954 70,648,462..70,649,061 626583
IGKV3-1*01 3-1 Vκ21G, 21-1 X16955 70,653,572..70,654,171 108004
proximal aGenes are ordered by their place on the chromosome, from J region distal to proximal.
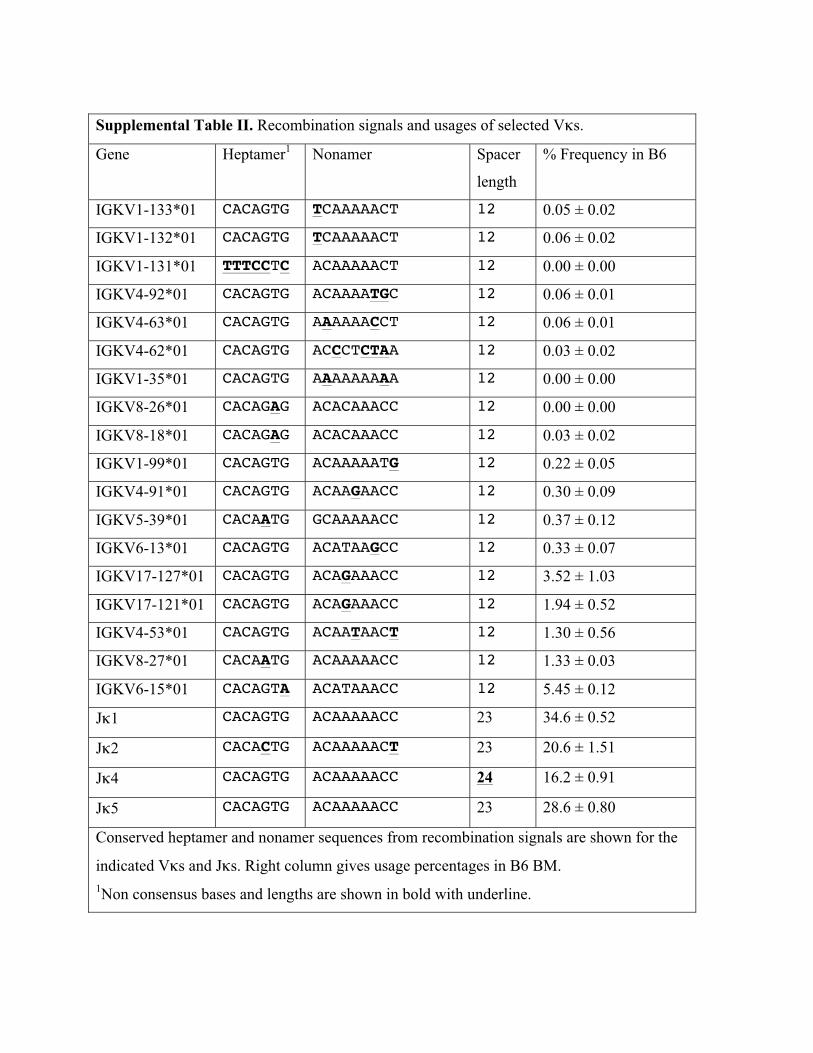
Supplemental Table II. Recombination signals and usages of selected Vκs.
Gene Heptamer1 Nonamer Spacer
length
% Frequency in B6
IGKV1-133*01 CACAGTG TCAAAAACT 12 0.05 ± 0.02
IGKV1-132*01 CACAGTG TCAAAAACT 12 0.06 ± 0.02
IGKV1-131*01 TTTCCTC ACAAAAACT 12 0.00 ± 0.00
IGKV4-92*01 CACAGTG ACAAAATGC 12 0.06 ± 0.01
IGKV4-63*01 CACAGTG AAAAAACCT 12 0.06 ± 0.01
IGKV4-62*01 CACAGTG ACCCTCTAA 12 0.03 ± 0.02
IGKV1-35*01 CACAGTG AAAAAAAAA 12 0.00 ± 0.00
IGKV8-26*01 CACAGAG ACACAAACC 12 0.00 ± 0.00
IGKV8-18*01 CACAGAG ACACAAACC 12 0.03 ± 0.02
IGKV1-99*01 CACAGTG ACAAAAATG 12 0.22 ± 0.05
IGKV4-91*01 CACAGTG ACAAGAACC 12 0.30 ± 0.09
IGKV5-39*01 CACAATG GCAAAAACC 12 0.37 ± 0.12
IGKV6-13*01 CACAGTG ACATAAGCC 12 0.33 ± 0.07
IGKV17-127*01 CACAGTG ACAGAAACC 12 3.52 ± 1.03
IGKV17-121*01 CACAGTG ACAGAAACC 12 1.94 ± 0.52
IGKV4-53*01 CACAGTG ACAATAACT 12 1.30 ± 0.56
IGKV8-27*01 CACAATG ACAAAAACC 12 1.33 ± 0.03
IGKV6-15*01 CACAGTA ACATAAACC 12 5.45 ± 0.12
Jκ1 CACAGTG ACAAAAACC 23 34.6 ± 0.52
Jκ2 CACACTG ACAAAAACT 23 20.6 ± 1.51
Jκ4 CACAGTG ACAAAAACC 24 16.2 ± 0.91
Jκ5 CACAGTG ACAAAAACC 23 28.6 ± 0.80
Conserved heptamer and nonamer sequences from recombination signals are shown for the
indicated Vκs and Jκs. Right column gives usage percentages in B6 BM. 1Non consensus bases and lengths are shown in bold with underline.

Supplemental Material Legends
Supplemental Figure 1. Mouse immunoglobulin κ locus representation. All reported genes on
the mouse κ locus are mapped based on NCBI database. Functional or ORF Vκs; red arrow, Vκ
pseudogenes; light blue arrow, other pseudogenes; blue box with allow, reported CTCF binding
site; yellow hexagon, Jκs; green arrow (Jκ3 is indicated by blue), Cκ; blue arrow. Each arrow
indicates their direction on the locus. Numbers on left indicate nucleotide position on
Chromosome 6.
Supplemental Figure 2. Frequency analysis of individual V-J associations in JCκ+/- BM. Vκ
genes are ordered by their chromosomal order from distal (top) to proximal (bottom). Jκ1, blue;
Jκ2, yellow; Jκ4, green and Jκ5, red. n=3 samples with 4 or 5 mice per sample. Mean for each
group indicated by a horizontal bar.
67500K�
68000K�
68500K�
69000K�
69500K�
70000K�
70500K�
2-137�Myc induced nuclear �antigen pseudogene� 1-136� 1-135�
Tubulin, alpha 1� pseudogene�
Ribosomal protein L18A� pseudogene 2�
14-134-1� 17-134�1-133�
1-132�
Tubulin, alpha 1� pseudogene�
Nucleosome assembly� protein 1-like pseudogene�
1-131� 14-130� 9-129� 9-128�
17-127� 14-126-1� ARP2 actin-related protein 2�homolog pseudogene�
14-126� 11-125� 9-124�
9-123� 1-122� 17-121�
9-120� 9-119� 14-118-2� 14-118-1� 11-118� 1-117�
2-116� 1-115� 11-114� 2-113� Cortactin pseudogene� 2-112�
14-111� 1-110� 2-109� 1-108� 2-107� 11-106� 2-105�
Cortactin pseudogene� 16-104� 15-103�
15-102� 20-101-2� 15-101-1�15-101� 14-100� 1-99�
12-98� 15-97� 10-96� 2-95-2�
2-95-1� 10-95� 10-94� 2-93-1� 19-93�
4-92� 4-91� 4-90� 13-89-1� 12-89�
1-88� 13-87� 4-86� 13-85� 13-84�
4-83� 13-82� 4-81� 13-80-1� 4-80� 4-79�
13-78-1� 4-78� 4-77� 13-76�
4-75� 13-74-1� 4-74� 13-73-1� 4-73� 4-72� 13-71-1� 4-71�
4-70� 4-69� 4-68� 12-67� 12-66� 4-65�
13-64� 4-63� 13-62-1� 4-62� 13-61-1� 4-61� 4-59�
4-60� 4-58� 4-57-1�13-57-2�
13-57-1� 13-56-1�4-57� 4-56� 13-55-1� 4-55� 13-54-1� 4-54� 4-53�
4-51� 4-50� 12-49� 5-48�
12-47� 12-46� 5-45� 12-44� 5-43� 12-42�
12-41� 5-40-1� 12-40� 5-39� 12-38�
5-37� 18-36� 1-35� 8-34�
7-33� 6-32� 8-31� 8-30� 6-29� 8-28�S-adenosylmethionine �decarboxylase, pseudogene 2�
8-27� 8-26� 6-25�8-24� 8-23-1�Ornithine decarboxylase, �structural 1 pseudogene�
6-23�Ornithine decarboxylase, �structural 1 pseudogene� 8-21�8-22�
S-adenosylmethionine �decarboxylase, pseudogene 5�6-20� 8-19�
8-18�Ornithine decarboxylase, �structural 1 pseudogene� 6-17� 8-16� Ornithine decarboxylase, �
structural 1 pseudogene�6-15�Solute carrier family 5,�Member 6 pseudogene�
6-14� Ornithine decarboxylase, �structural 1 pseudogene�
6-13� Ornithine decarboxylase, �structural 1 pseudogene�
3-12-1� 3-12�
3-11� 3-10� 3-9� 3-8� 3-7� 3-6�
3-5� 3-4� 3-3� 3-2� 3-1� Jκs� Cκ�
Ribosomal protein L18A� pseudogene 3�
Fig.S1�
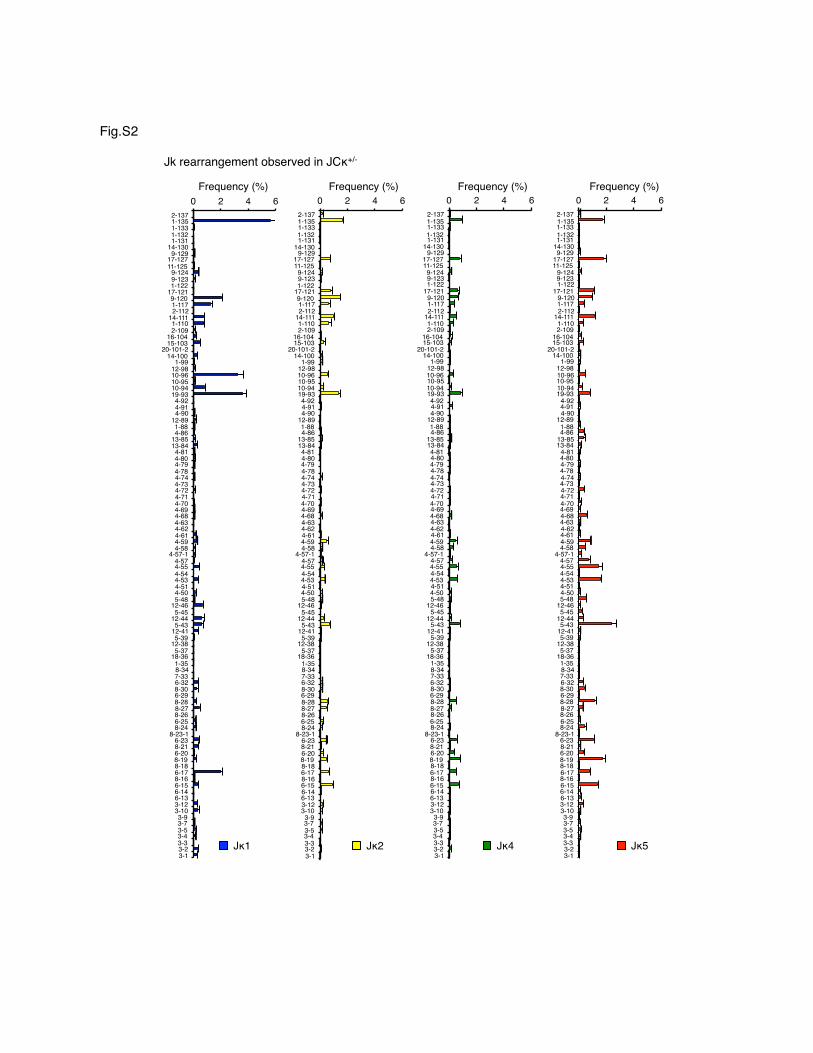
Fig.S2�
Frequency (%)�
Jκ1 � Jκ2 � Jκ4 � Jκ5 �
Jk rearrangement observed in JCκ+/-�
Frequency (%)� Frequency (%)� Frequency (%)�0� 2� 4� 6�
2-137�
1-133�
1-131�
9-129�
11-125�
9-123�
17-121�
1-117�
14-111�
2-109�
15-103�
14-100�
12-98�
10-95�
19-93�
4-91�
12-89�
4-86�
13-84�
4-80�
4-78�
4-73�
4-71�
4-69�
4-63�
4-61�
4-58�
4-57�
4-54�
4-51�
5-48�
5-45�
5-43�
5-39�
5-37�
1-35�
7-33�
8-30�
8-28�
8-26�
8-24�
6-23�
6-20�
8-18�
8-16�
6-14�
3-12�
3-9�
3-5�
3-3�
3-1�
1-135�
1-132�
14-130�
17-127�
9-124�
1-122�
9-120�
2-112�
16-104�
1-110�
20-101-2�
1-99�
10-96�
10-94�
4-92�
1-88�
4-90�
13-85�
4-81�
4-79�
4-74�
4-72�
4-68�
4-70�
4-62�
4-59�
4-57-1�
4-55�
4-53�
12-46�
4-50�
12-44�
12-41�
12-38�
18-36�
8-34�
6-29�
6-32�
8-27�
6-25�
8-23-1�
8-21�
8-19�
6-15�
6-17�
6-13�
3-10�
3-7�
3-4�
3-2�
0� 2� 4� 6� 2-137�
1-133�
1-131�
9-129�
11-125�
9-123�
17-121�
1-117�
14-111�
2-109�
15-103�
14-100�
12-98�
10-95�
19-93�
4-91�
12-89�
4-86�
13-84�
4-80�
4-78�
4-73�
4-71�
4-69�
4-63�
4-61�
4-58�
4-57�
4-54�
4-51�
5-48�
5-45�
5-43�
5-39�
5-37�
1-35�
7-33�
8-30�
8-28�
8-26�
8-24�
6-23�
6-20�
8-18�
8-16�
6-14�
3-12�
3-9�
3-5�
3-3�
3-1�
1-135�
1-132�
14-130�
17-127�
9-124�
1-122�
9-120�
2-112�
16-104�
1-110�
20-101-2�
1-99�
10-96�
10-94�
4-92�
1-88�
4-90�
13-85�
4-81�
4-79�
4-74�
4-72�
4-68�
4-70�
4-62�
4-59�
4-57-1�
4-55�
4-53�
12-46�
4-50�
12-44�
12-41�
12-38�
18-36�
8-34�
6-29�
6-32�
8-27�
6-25�
8-23-1�
8-21�
8-19�
6-15�
6-17�
6-13�
3-10�
3-7�
3-4�
3-2�
0� 2� 4� 6� 2-137�
1-133�
1-131�
9-129�
11-125�
9-123�
17-121�
1-117�
14-111�
2-109�
15-103�
14-100�
12-98�
10-95�
19-93�
4-91�
12-89�
4-86�
13-84�
4-80�
4-78�
4-73�
4-71�
4-69�
4-63�
4-61�
4-58�
4-57�
4-54�
4-51�
5-48�
5-45�
5-43�
5-39�
5-37�
1-35�
7-33�
8-30�
8-28�
8-26�
8-24�
6-23�
6-20�
8-18�
8-16�
6-14�
3-12�
3-9�
3-5�
3-3�
3-1�
1-135�
1-132�
14-130�
17-127�
9-124�
1-122�
9-120�
2-112�
16-104�
1-110�
20-101-2�
1-99�
10-96�
10-94�
4-92�
1-88�
4-90�
13-85�
4-81�
4-79�
4-74�
4-72�
4-68�
4-70�
4-62�
4-59�
4-57-1�
4-55�
4-53�
12-46�
4-50�
12-44�
12-41�
12-38�
18-36�
8-34�
6-29�
6-32�
8-27�
6-25�
8-23-1�
8-21�
8-19�
6-15�
6-17�
6-13�
3-10�
3-7�
3-4�
3-2�
0� 2� 4� 6� 2-137�
1-133�
1-131�
9-129�
11-125�
9-123�
17-121�
1-117�
14-111�
2-109�
15-103�
14-100�
12-98�
10-95�
19-93�
4-91�
12-89�
4-86�
13-84�
4-80�
4-78�
4-73�
4-71�
4-69�
4-63�
4-61�
4-58�
4-57�
4-54�
4-51�
5-48�
5-45�
5-43�
5-39�
5-37�
1-35�
7-33�
8-30�
8-28�
8-26�
8-24�
6-23�
6-20�
8-18�
8-16�
6-14�
3-12�
3-9�
3-5�
3-3�
3-1�
1-135�
1-132�
14-130�
17-127�
9-124�
1-122�
9-120�
2-112�
16-104�
1-110�
20-101-2�
1-99�
10-96�
10-94�
4-92�
1-88�
4-90�
13-85�
4-81�
4-79�
4-74�
4-72�
4-68�
4-70�
4-62�
4-59�
4-57-1�
4-55�
4-53�
12-46�
4-50�
12-44�
12-41�
12-38�
18-36�
8-34�
6-29�
6-32�
8-27�
6-25�
8-23-1�
8-21�
8-19�
6-15�
6-17�
6-13�
3-10�
3-7�
3-4�
3-2�








![[Immunology of celiac disease]](https://img.dokumen.tips/doc/110x75/635861503cd558f04e054612/immunology-of-celiac-disease.jpg)




















