Embed Size (px)
Citation preview

1
Variability in Middle Stone Age faunal exploitation and use of thephysical and social landscapes inthe southwestern cape, South Africa
Jessica C. Thompson
15
Abstract. Discussions of the Middle Stone Age (MSA) are often conducted in broad termsthat homogenize behavioral variability during a period that covers an enormous span ofboth time and space. This makes it extremely difficult to examine MSA settlement dynam-ics at a level that is above that of intra-site analysis but still below large-scale compar-isons between, for example, continental subregions. Recent reinvestigations of existingcollections and an expanded empirical record from new projects have shown that morebehavioral variability is represented in MSA lithic technology, artifact manufacture, andsite-use than was previously known. Case studies from two coastal MSA faunal accumu-lations in South Africa, Blombos Cave and Pinnacle Point Cave 13B, show that MSA fau-nal exploitation was equally heterogeneous across the landscape. Subsistence decisionssuch as prey selection and transport were not uniform through either time or space, andwere at least partially determined by the site context.
Résumé. Les discussions sur le Middle Stone Age (MSA) sont souvent posées au senslarge ce qui homogénéise la variabilité comportementale pendant une période qui couvreun horizon énorme de temps et d’espace. Ceci rend extrêmement difficile l’examen de ladynamique de colonisation du MSA à un niveau supérieur à l’analyse sur-site maisencore inférieur aux comparaisons à grande échelle entre, par exemple, les sous-régionscontinentales. Grâce à la reprise récente des recherces sur les collections existantes etaux resultats des travaux empiriques des nouveaux projets, il a été démontré que lavariabilité comportementale est représentée dans la technologie du lithic du MSA, l'objetfabriqué, et l'usage de site précédemment découvert. L’étude de deux accumulations defaune des sites MSA côtières dans l’Afrique du Sud, Blombos Cave et Pinnacle PointCave 13B, montre que cette exploitation de faune du MSA était également inhomogèneà travers le paysage.
IntroductIon
The term ‘Middle Stone Age’ (MSA) refers to both a period of time (ca. 280–30 thousand
years ago [ka]), and to a diagnostic suite of techno-behavioral adaptations in sub-Saharan
Africa that characterize it. Discussions of the latter aspect are often conducted in highly gen-
eralized terms, giving the impression that the MSA represented a large portion of prehistory
characterized by very little variation. MSA faunal exploitation especially has been described
in generalized temporal and spatial terms, with ‘the MSA’ being compared as a whole to

2
thompson
another entity, such as ‘the LSA’ (e.g., Klein 1975, 1976, 1978a, 1987, 2000). This has per-
petuated the impression that human faunal exploitation behavior across the enormous spans
of time and space encompassed by the MSA was rather static and homogeneous, and that
most major technological and subsistence shifts took place at the Middle-Later Stone Age
transition.
Generalizations can be useful for large-scale comparisons between continental subre-
gions (e.g., southern, eastern, or northern Africa, the Horn, etc.). However, because they nec-
essarily gloss over the complex details of local archaeological records, such generalizations
are overly simplistic for investigating subtle changes in MSA settlement dynamics over both
time and space. Such investigations are important because resolving smaller-scale patterning
in the utilization of the social and physical landscapes is essential for understanding larger
changes in human behavior that took place during the MSA. This requires an examination of
the entire behavioral suite, which includes differentiation in site-use, behavior, and adaptive
strategies with regard to faunal exploitation. The greatest obstacles to achieving this goal, and
how they have been addressed, are discussed here. The zooarchaeological record of South
Africa, particularly the Western Cape, is used throughout to illustrate these points, and an
example of an analysis that overcomes most of these obstacles is provided that uses two new
faunal datasets from this region. The locations of the major sites discussed are shown in
figure 1.
theoretIcAl obStAcleS
There are some underlying theoretical issues that have discouraged workersresearchers from
explicitly seeking to describe and interpret the range of variability in MSA behavior. Firstly,
it has only been in the last two decades that the MSA has begun to attract a substantial meas-
ure of interest (Marean and Thompson 2003). This recent upswing in research focus is mainly
owed due to the recognition that the MSA was the critical time period that saw the emergence
of the modern anatomy, genotype, and behavioral repertoire (e.g., Aiello 1993; Henshilwood
and Marean 2003; Klein 1995, 2000; McBrearty and Brooks 2000; Miller et al. 1999,
Relethford 1995). The longest-standing model for the advent of behavioral modernity has
previously been one of an extended period of stasis for most of the MSA during which the
material expression of human behavior was relatively simple and homogenous. This was
generally supported in the artifact record as it was known at the time, which showed little
variation across time and space for lithics, and with the manufacture of artifacts on other
materials such as bone being virtually unknown.
Under this model, the stasis was ended abruptly by a rapid change at ca. 50–40 ka
(Gamble 1994; Klein 1989b 2000, 2003; Mellars 1989; Mellars and Stringer; 1989; Mithen
1999). This ‘cultural revolution’ has been explained by a sudden mutation or change in
human cognition, perhaps mediated by the advent of fully articulate language and other
forms of symbolism. Because the transition to modernity was considered to be an abrupt one
possibly instigated by external factors, the human behavioral suite at and after the MSA-LSA
transition would have been somewhat independent of that which preceded the transition.
Therefore, any variability in MSA behavior during the long period leading up to this critical
change was considered to be largely irrelevant under this model.
In contrast to sudden-change models, more recently developed, process-based models
implicitly acknowledge that behavior was not static during the MSA. These regard the advent

3
Variability in Middle Stone Age faunal exploitation and use of landscapes
of modernity as an accretionary process deeply rooted in the Middle–Upper Pleistocene
(Chase and Dibble 1990; Foley and Lahr 1997; McBrearty and Brooks 2000; Henshilwood
and Marean 2003). Following this, indications of modern human behavior are considered to
have appeared at disparate times over the course of the last 350 ka, stretching back into the
Middle Pleistocene, rather than in a suddenly appearing package. Some behaviors, such as
symbolic use of pigment, may have made their first appearance early but in a very simple
manner, slowly becoming more complex over the course of time (McBrearty and Brooks
2000). Under this model, identifying variability in MSA behavior across time and space
becomes essential for diagnosing changes in the adaptive strategies that ultimately led to the
complete modern behavioral package.
eMpIrIcAl obStAcleS
The historical lack of research focus on the MSA and the persistence of sudden-change mod-
els have both been directly influenced by the state of the empirical record (Henshilwood and
Marean 2003). When compared to contemporaneous, but better studied, regions such as
Western Europe, there is a tiny available sample of well-dated and comprehensively studied
Fig. 1. Map of the study area showing location of sites
discussed in the text.

4
thompson
MSA sites. This is in turn the result of poor site preservation, the lack of a long and well-
established tradition of research, and the logistically challenging nature of working in Africa.
Furthermore, because they are the most commonly preserved archaeological tracematerial,
behavioral inferences have traditionally relied on lithic assemblages. If these appear
immutable through time and across space, it reinforces the impression that most aspects of
the MSA were equally static. An apparent lack of dynamic change in lithics can create a neg-
ative feedback loop overall, in which variability may not be investigated because variability
is not considered to be present.
Despite the limitations imposed by the empirical record, some of the earliest discussions
of variability in MSA behavior were based on observations from lithic assemblages. Clark
(1988) first suggested that regional diversity began during the early Middle Stone Age, and
used evidence from the East African lithic record to support this. However, the regions and
time periods in question were still extremely large. Also, south of the Zambezi the MSA has
traditionally been considered to be typologically and technologically very homogenous, with
most sites dominated by Volman’s (1984) monolithic category of ‘MSA II’ (Thackeray
1993).
Some researchers have argued that the South African MSA witnessed changes in the
logistical exploitation of lithic resources through time (Deacon 1989; Thackeray 1993), and
newer studies have determined that distinct technological differences are detectable within
these basic lithic subdivisions (Wurz 2002). However, relative to later industries the MSA
still represents a long stretch of time in which changes in lithic technology were generally
quite subtle. Two notable exceptions in South Africa are the Still Bay, characterized by finely
worked bifaces, and the Howieson’s Poort (HP), characterized by backed pieces (Thackeray
1993).
Quantities of Still Bay points have now been recovered from the upper layers of Blombos
Cave (BBC), along the southern coast of the Western Cape Province, which have been well
dated using luminescence techniques to ca. 73 ka (Jacobs et al. 2006; Tribolo et al. 2006).
The industry has also been recovered at Sibudu, an inland site overlooking the Tongati River
in the province of KwaZulu-Natal, and the dates for this industry are in general agreement
with those from Blombos (Jacobs et al. 2008; Wadley 2007). Recent dates for the HP using
a variety of techniques have shown that the HP was slightly younger than the Still Bay, likely
dating to between 62–52 ka (Jacobs et al. 2008; Miller et al. 1999; Tribolo 2003; Wadley and
Jacobs 2004, 2006). A series of samples from several HP-bearing sites have been analyzed
using a single consistent method and have resulted in a more precise and surprisingly brief
age estimate of 64.8–59.5 ka (Jacobs et al. 2008).
In addition, recently reported finds from Pinnacle Point Cave 13B (PP13B) include
bladelets crafted on quartzite and silcrete (Marean et al. 2007). Although much attention has
been paid to the benefits of blade technology and its precocious appearance in Africa has
been acknowledged (Bar-Yosef and Kuhn 1999; McBrearty and Brooks 2000), bladelet tech-
nology was previously been thought to be a hallmark of Later Stone Age and Upper
Paleolithic stone tool technology. Moreover, the bladelets at PP13B are of an unprecedented
antiquity, dating to ca. 168 ka (Marean et al. 2007). Recent advances in dating technology
and their application to both previously recovered and recently excavated assemblages have
better secured the place of these tool types in the MSA lithic record: they now appear to be
less eccentricities and more adaptive responses to environmental or social pressures (e.g.,
Ambrose and Lorenz 1990; Deacon 1989; McCall 2007). It is also intriguing that such indus-

5
Variability in Middle Stone Age faunal exploitation and use of landscapes
tries were likely short-lived and separated sufficiently in time that they do not represent a
series of related innovations.
Variability in non-lithic artifact types during the MSA has also become apparent.
At Blombos Cave, evidence of complex symbolic behavior has been unearthed in the form
of abundant worked ochre and ochre incised with cross-hatching (Henshilwood et al. 2002).
While ochre has long been recognized to occur at MSA sites, particularly those postdating
about 100 ka, recent work has shown that it was selected for its red color, rich saturation, and
non-utilitarian qualities as a pigment (Hovers et al. 2003; Watts 1999). In South Africa, ochre
incised with cross-hatching and deep gouges has also been identified from Klein Kliphuis,
showing that the finds from Blombos were not isolated occurrences (Mackay and Welz
2008).
Importantly, the same excavations in South Africa that have demonstrated such variability
in artifact manufacture have also resulted in the recovery of well-preserved remains of less
durable materials such as shell and bone. Some of these assemblages include deliberate
human modifications made to these materials, showing that such behavior was not restricted
to ochre. Examples are incisions on bone from Blombos and Sibudu (Cain 2004; d’Errico
et al. 2001) and engravings on ostrich eggshell at Diepkloof (Poggenpoel et al. 2005). Shell
beads have been recovered from Blombos (Henshilwood et al. 2004), and both formal and
informal bone tools have been recovered at several sites in the South African record.
Although some have previously been known to occur at the Peers Cave and Klasies River
Mouth (KRM) sites, they were isolated finds with uncertain proveniences (Backwell et al.
2008). Now at Sibudu (Backwell et al. 2008) and Blombos (d’Errico et al. 2001; d’Errico and
Henshilwood 2007; Henshilwood and Sealy 1997; Henshilwood et al. 2001a, 2002), such
tools have been dated to ca. 90 ka, ca. 61 ka, and between 70–85 ka, respectively (Jacobs
et al. 2006).
An important point is that not all sites with preserved bone and shell have yielded such
artifacts. The faunal collection from Die Kelders Cave 1 (DK1) has been extensively studied
using microscopic techniques, and yet no bone tools, shell beads, or modified eggshell have
been reported (Marean et al. 2000a, 2000b). Moreover, the lithic assemblage has been
described as one that is relatively generic and with little typological distinction (Thackeray
2000). The deposits at DK1 have been dated by TL, OSL (Feathers and Bush 2000), and ESR
(Schwarcz and Rink 2000) to ca. 70–60 ka, which places them toward the younger end of the
MSA age range. These data indicate that at least in the Western Cape of South Africa the
MSA was characterized by a series of technological and cultural changes rather than a simple
linear increase in material culture complexity over time. It also shows that not all sites dating
to the time periods that included industries such as the Still Bay and HP have evidence of
these industries.
Other MSA sites with preserved fauna also lack reported finds of bone tools. These
include Border Cave (Klein 1977), Boomplaas (BPS; Klein 1978b), and Ysterfontein 1 (YF1;
Halkett et al. 2003; Klein et al. 2004). It is possible that if these assemblages were examined
in greater detail some bone tools would be identified, but it is unlikely that these would be as
formal or as abundant as at sites such as Blombos. This is evidenced by a microscopic study
of over 16,000 well-preserved bone fragments from PP13B, which yielded only a single bro-
ken and informal bone tool (Thompson 2008). Overall, in terms of artifact manufacture it
now seems clear that not all MSA groups in South Africa were doing the same things at the
same times.

6
thompson
The recent addition of important assemblages such as this to the MSA record of South
Africa has partially addressed some of the persistent taphonomic obstacles discussed above,
and from this improved, but still limited, sample two major new pieces of information have
arisen. Firstly, artifact manufacture occurred on a much wider variety of substrates and in
more forms than what can be inferred from only the lithic record. Secondly, there is a great
deal of variability between sites in the production and discard of such materials. The second
point is critical to understanding MSA behavior because when taphonomic obstacles have
been removed it has not so far resulted in a one-to-one increase in the number of bone or shell
artifacts that have been recovered. It seems to be a true pattern that some sites have abundant
traces of certain behaviors that others do not. An important next step is therefore to determine
what other, more subtle indications of hominin behavioral variability may be apparent in the
zooarchaeological assemblages from these sites.
MethodologIcAl obStAcleS
The present increased understanding of variability in the MSA artifact record has been made
possible by revisiting old assemblages, excavating new ones with modern techniques, and
linking the finds to more secure chronologies. However, similar revelations about variability
in subsistence behavior have been slower to materialize. In addition to some of the tapho-
nomic obstacles discussed above, there are several methodological reasons that reliable inter-
pretations and comparisons of human-accumulated faunal assemblages have been extremely
elusive.
Published interpretations of South African MSA faunal assemblages have until recently
relied on data from KRM (Binford 1984; Klein 1976; Milo 1998), Border Cave (Klein 1977),
and Boomplaas (Klein 1978b). With the exception of Milo’s (1998) work on the Klasies
River assemblage, these reports have all neglected to provide a detailed taphonomic assess-
ment of the collections, and instead have focused on taxonomic and skeletal element abun-
dances. The underlying assumption has been that MSA hominins were the primary
accumulators of the faunal assemblages in question, with only an insignificant contribution
by non-human predators and other biotic and abiotic bone collectors. However, without sys-
tematic taphonomic studies to document that this was the case for each assemblage, attempts
to understand variability in MSA faunal exploitation using these datasets can only be specu-
lative.
The converse is equally problematic, in that the small fauna from of these assemblages
(hares, Cape dune mole rats, hyraxes, and tortoises) are often implicitly assumed to have not
been a major component of the hominin diet. It is clear that there is a great deal of variability
in the representation of this component relative to larger mammals. Counts of the Number of
Identified Specimens (NISP) for tortoises are not readily available in the literature, but small
mammal data show that sites such as DK1 (Klein and Cruz-Uribe 2000), Ysterfontein 1
(Halkett et al. 2003; Klein et al. 2004), and Blombos (Henshilwood et al. 2001b) have very
high NISP counts of small fauna while sites such as PP13B (Thompson 2008;
Thompson n.d.) have very low counts (fig. 2). However, as with the larger mammals, until
systematic taphonomic studies have been performed that identify the accumulator(s) of the
small fauna, the question remains open about the role these animals played in MSA subsis-
tence and what this variability actually means.

7
Variability in Middle Stone Age faunal exploitation and use of landscapes
A microscopic examination of bone surfaces has been shown to be the most reliable way
of diagnosing the agent that has modified a bone—especially for hominins and carnivores
(Blumenschine et al. 1996). Milo (1998:102) illustrated this point well when he revisited the
KRM assemblage with microscopic techniques and found that the actual number of hominin-
inflicted marks on bovid remains was more than double what had been previously reported
using only macroscopic techniques (Binford 1984). Unfortunately, even where such tapho-
nomic studies have been published, as is the case for the larger mammals from KRM (Milo
1998), DK1 (Marean et al. 2000b) and Sibudu (Cain 2004; Clark and Plug 2008), the
methods are not described in sufficient detail, are different enough, or are based on incom-
plete assemblages so as to make the results largely incomparable to one another. This is the
case both for data collection and for data presentation, and is a major obstacle for identifying
and interpreting variability in MSA faunal assemblages.
All interpretations from the KRM faunal assemblage suffer from the persistent problem
of selective post-excavation removal of certain components, such as shaft fragments. This
has rendered the assemblage incomplete and biased toward more easily identified fragments
(Bartram and Marean 1999). Such bias has been proposed to be a serious impediment to
accurate assessment of skeletal element abundance and surface modification (e.g., Marean
and Frey 1997; Marean and Kim 1998; Marean et al. 2004; Pickering et al. 2003), although
other researchers have contended that this is not necessarily the case (e.g., Klein et al. 1999;
Stiner 2002).
Analytical selection can have the same effect. At Sibudu (Cain 2006) a large sample that
included shaft fragments was examined microscopically, but the researcher employed an
arbitrary 20 mm size cutoff point. This cutoff basically guarantees that most of the bone
flakes that are detached when a long bone is broken open to obtain marrow will be missed
(Marean and Bertino 1994). Importantly, when these flakes detach they often retain the mark
left by either the hammerstone percussor or the carnivore’s tooth (Capaldo and Blumenschine
1994). This makes their surfaces critical places to look for evidence of such activities, a fact
Fig. 2. relative proportions of small and large mammals at MSA faunal
assemblages that have been reported from the Western cape. All data are
in nISp except sites marked with an asterisk (*), which only have Minimum
number of Individuals (MnI) counts available.

8
thompson
that is supported by recent analyses from PP13B and Blombos, in which 35% and 47% of
percussion marks on shaft fragments respectively were identified from fragments less than
20 mm in their maximum dimension (Thompson 2008). At present, the only published tapho-
nomic analysis that employs a microscopic examination of all specimens with no size cutoff
is that from Layers 10 and 11 at DK1 (Marean et al. 2000b). However, two new analyses from
PP13B and Blombos have been conducted using identical data collection methods and ana-
lytical subsets (Thompson 2008). These promise to provide the first opportunity to compare
reliably MSA faunal assemblages, making it possible for subtle details of variability in MSA
faunal exploitation to be examined.
lAnd uSe And FAunAl exploItAtIon behAVIor: dIetAry coMpoSItIon
The zooarchaeological record is critical for an overall understanding of MSA land use
because many of the decisions regarding group size, mobility, and territoriality can be direct-
ly linked to the availability of key subsistence resources (e.g., Kelly 1995). Several of these
key resources were almost certainly animal-derived. Although the MSA hominin skeletal
record is scant, there are few indications that their anatomies were substantially different
from our own. In particular, we know they were in possession of a large brain that would have
required maintenance with the fats and proteins found most effectively in animal resources
(Aiello and Wheeler 1995; Speth and Spielman 1983). In modern hunter-gatherers these
macronutrients are highly valued, and they are essential for proper growth and nutrition
(Eaton and Konner 1997; Kaplan et al. 2000; Milton 1999). This is particularly true for
infants, juveniles, and nursing mothers who require docosahexaenoic fatty acids (DHA) such
as those recovered from the marrow and brain cavities and from marine resources for optimal
neurologic development and maintenance (Broadhurst et al. 2002; Langdon 2006).
Given their key role in the modern human diet, animal products have the high social value
that would be expected (e.g., Hawkes et al. 1991; Hawkes and Bird 2002). If the metabolic
and nutritive requirements of MSA hominins were not substantially different from what is
seen in modern humans, then animal resources can be taken to have been an important con-
tribution to the diet. In many modern hunter-gatherer groups, collectors contribute to the diet
in the form of roots, nuts, seeds, and other gathered resources. Patches of these resources are
often more reliable and predictable than terrestrial mammal resources (Burger et al. 2005).
Despite this, the contribution of hunters is often considered more valuable and carries a high-
er status than the contribution of the gatherers (Hawkes and Bird 2002). Some authors have
suggested that this is because hunting is a more dangerous way to acquire resources, and thus
hunters are engaging in costly signaling, or ‘showing off’ in order to enhance their reproduc-
tive success (Hawkes and Bird 2002; but see Wood 2006). However, it is important to note
that hunting of large ungulates is not the only way to obtain fats and proteins.
There has been much debate on whether MSA groups obtained large mammal resources
through hunting, scavenging, or some combination of these strategies (Bartram and Marean
1999, Binford 1984, Klein 1989a 1995, Marean and Assefa 1999, Marean et al. 2000b; Milo
1998). The most recent work shows several lines of evidence in support of MSA hominins as
fully capable hunters (e.g., Assefa 2005; Clark and Plug 2008; Faith 2008; Thompson 2008).
Furthermore, obligate scavengers, for which animal resources comprise a major portion of
the diet, have specialized adaptations for ranging widely to find carcasses, locating them,
stripping them of remaining nutrients, and avoiding other predators at the kill-site. These

9
Variability in Middle Stone Age faunal exploitation and use of landscapes
adaptations may be olfactory, locomotor, behavioral, or include biological solutions for
breaking into bones or digesting tough carcass components left by carnivores with earlier
access. Although technological solutions such as hammerstones to break open bones and fire
to cook otherwise indigestible components solve many of these problems for hominins, their
lack of biological adaptations for encountering carcasses on a more than opportunistic basis
renders them an unlikely candidate to be obligate scavengers.
In environments such as that of the southern coast of Africa, where both marine resources
and tortoises are available for collection, MSA hominins did not have to be either hunters or
scavengers of large ungulates in order to obtain animal resources critical for the diet.
However, collection of shellfish and tortoises is more akin to gathering and is an activity in
which all members of a group can participate. This has implications for the social structuring
of the group, as the role and status of juveniles, the elderly, pregnant females, or females with
young will be much different in a society in which they also contribute animal resources than
one in which animal resource procurement falls largely to males in the form of active hunting
(Hawkes and Bird 2002). If fats and proteins were normally obtained through efforts similar
to gathering, then we might expect a relatively equitable social hierarchy and little sex-spe-
cific or even age-specific division of labor in MSA groups (Hawkes 1996).
Under this scenario, scavenging of large mammal resources could feasibly act as a sup-
plement to the MSA diet rather than a critical mode of resource acquisition—as the majority
of animal resources would be obtained in other ways. This has further implications for land
use, because resources such as shellfish tend to occur at targeted points on the landscape,
requiring foreknowledge of when and where to obtain them. Shellfish beds could also be con-
sidered a ‘defensible resource’, and a reliance on them would have necessitated both a sys-
tematic pattern of mobility designed to intercept this resource at times when they were
productive and a series of strategies to ensure that other hominin groups did not do the same.
In contrast, tortoises occur more evenly on the landscape but reproduce slowly. If they com-
prised a major portion of the diet, then their rapid depletion would have required a mobility
strategy that allowed time for their populations to recover before a region was once again
exploited.
It is interesting to note that PP13B is a site where targeted shellfish exploitation has been
documented, and that it is also the only reported MSA site along the South African coast that
does not have abundant representation of tortoises (Marean et al. 2007; Thompson n.d.). A
taphonomic study of the tortoises that do occur at the site shows that they were only oppor-
tunistically exploited by MSA hominins. Differences in faunal composition such as these
could be potentially very informative about not only MSA diet, but also land use. However,
until similar work has been done for other sites, it is difficult to situate this pattern in the larg-
er context of MSA sociality with regards to subsistence.
lAnd uSe And FAunAl exploItAtIon behAVIor: A cASe Study oF lArge MAMMAl trAnSport
overview of sites and samples
The most highly visible and well-studied component of MSA faunal assemblages is the larger
mammals (e.g., Assefa 2005; Clark and Plug 2008; Halkett et al. 2003; Henshilwood et al.
2001b; Klein 1976, 1978; Klein and Cruz-Uribe 2000; Klein et al. 2004; Marean et al.

10
thompson
2000b). The large mammal dataset is capable of addressing a wide range of issues about
hominin behavior, such as: prey selection, transport strategies, butchering methods, nutrient
extraction, food preparation, and the within-site distribution of these activities. Many of the
decisions that underlie such behaviors are related to individual site context, and this is why
reliable comparisons between faunal assemblages are so critical for building models of
greater MSA land-use strategies. One example of the fruitful way in which such comparisons
can be employed is provided here, using new data that were collected and analyzed using
identical methods from two MSA sites: PP13B and Blombos Cave.
The data collection methods and taphonomic details of the two assemblages will only be
summarized here, based on data from Thompson (2008, n.d.). Primary zooarchaeological
data from Pinnacle Point Cave 13B (PP13B) and Blombos Cave were collected at Iziko:
South African Museums of Cape Town. For both sites, all fauna identifiable to element was
cleaned of sediment and adhering matrix with fresh water, although not all matrix could be
removed with water. Fragments were then given individual specimen numbers and each
numbered specimen was entered as an individual record into a Microsoft Access database
designed for this purpose. These specimens include less identifiable long bone fragments
which are not assignable to a specific element but that could be identified at least to this very
general level. The remainder was identified to the greatest degree possible to body size and
skeletal element, with percentages of preserved diagnostic bone landmarks recorded in incre-
ments of 10% (e.g., deltoid tuberosity of the humerus, linea aspera of the femur, medial
condyle of the tibia, etc.). Pieces of spongy bone, bits of cortical bone with no evidence of
having a medullary cavity or having otherwise come from a long bone, and fragments of cor-
tical bone with no facets, diagnostic shape, or muscle markings that could indicate their
approximate location in the skeleton, were not included in either study.
All large mammal data are presented in terms of body size and general taxonomic cate-
gory of the family level or above. Each fragment was placed under a 10–40 x binocular light
microscope with a fiber-optic halogen light shining obliquely across the bone surface. This
is a method that successfully diagnoses cut, tooth and percussion marks 97–100% of the time
with minimal training (Blumenschine et al. 1996), and for which the author has been admin-
istered blind tests to ascertain her own level of accuracy. The percentage of the surface that
was covered by matrix or rodent gnawing was recorded in increments of 10%. Gastric etch-
ing was recorded as presence/absence, weathering stages followed Behrensmeyer (1978),
and post-depositional/geochemical alterations of the bone surface followed Thompson
(2005). The angle and outline of break edges were recoded for long bones following Villa
and Mahieu (1991), and excavation versus sediment or ancient breaks were recorded as being
‘extremely different’, ‘slightly different’, or ‘not different’ from the adjacent bone surface.
Of the larger mammals, ungulates (especially bovids) dominate all the faunal assem-
blages, but there are differences between sites and between major stratigraphic units. At
PP13B overall these ungulates are divided nearly evenly between size classes 1–3. When
broken down into the chronological and spatial groupings, small fauna (size 1 and 2) have
been found to be more abundant during MIS 5 and at the front of the cave during both time
periods. This is particularly true for size class 1 ungulates.
At Blombos, size 1 ungulates comprise the majority of the larger mammal faunal assem-
blage in M2 and M3, with the highest proportion in M2. The situation changes in the upper-
most level, M1, when size 2 ungulate representation is increased at the expense of the size 1
component. This pattern could be explicable in either environmental or behavioral terms. M1

11
Variability in Middle Stone Age faunal exploitation and use of landscapes
saw the introduction of a colder climatic period that would certainly have influenced both
local environments and hominin behavior to some extent. A reverse pattern, but one that was
potentially also climate related, occurred at PP13B. Here, the shift to a higher representation
of small fauna coincides with a general climatic change from MIS 6 to MIS 5, when climatic
conditions improved.
As is typical at most archaeological sites, there has been a substantial degree of density-
mediated destruction at all three sites. This has resulted in a high representation of long bone
midshafts relative to epiphyseal ends and spongy elements. The correlation between long
bone portion representation and density as measured by Computed Tomography (Lam et al.
1998, 1999, 2003) was found to be positive and significant for nearly all body size classes
and all major stratigraphic horizons at PP13B, Blombos, and DK1 (Thompson 2008). For this
reason, the recommendations of Marean and Cleghorn (2003) have been followed here, and
only elements in the high-survival set are compared to one another in this example.
Several lines of evidence indicate that MSA hominins were the main accumulator of the
size 2–4 body size classes at PP13B and Blombos. The sample of size 5 fauna from PP13B
and Blombos is too small to be informative. At both sites size 1 fauna had some hominin
input, but was also modified to varying degrees by other agents such as carnivores. At DK1
Marean et al. (2000b) determined that raptors were a major accumulator of this body size
class, but at PP13B and Blombos raptors are not implicated.
At PP13B a relatively substantial independent carnivore input was seen in the low co-
occurrence of tooth and percussion marks on the same midshaft fragment during MIS 6 only.
During MIS 5 the proportion of size 1 ungulates increases and with it increases the evidence
of hominin exploitation. In contrast, at Blombos the two bottom layers M2 and M3 are the
ones with the highest representation of size 1 ungulates but the highest proportions of per-
cussion marks on midshafts from larger ungulates, suggesting that a larger degree of non-
hominin (likely carnivore) input occurred in the lower two layers, and that this was
predominately on the smallest ungulates.
Despite their abundance at Blombos, size 1 ungulates were not the main prey focus for
hominins. This is in contrast to the situation at PP13B where as relative abundances of size 1
fauna increase, so too does evidence for hominin involvement with this category. The early
part of the Blombos occupation seems most similar to the MIS 6 deposits at PP13B in that
hominins did not put much effort into the capture of less abundant and elusive small antelope
but where the sites still received moderate quantities of these as a combination of both
hominin and carnivore efforts. In MIS 5 at PP13B, which overlaps in time only with the M3
deposits at Blombos, hominins seemed to have begun to exploit these small ungulates delib-
erately and more frequently. Despite being very close in time and despite likely having sim-
ilar surrounding environments during this time, hominins at PP13B were treating size 1
ungulates differently than were hominins at Blombos. These conclusions must be considered
in the following analysis, as data from the larger ungulates (size 3 and 4) are likely to provide
a ‘cleaner’ signature of hominin transport decisions than the smaller (size 1 and 2) ungulates.
Today the two sites are situated on the South African coast of the Western Cape Province,
although changes in global sea level over time would have periodically placed them at vary-
ing distances to the sea. Together, they cover a time range of human occupation that is likely
between ca. 168–73 ka, with deposits at PP13B ending by 92 ka and deposits at Blombos
beginning at around 100 ka. (Jacobs et al. 2003a, 2003b, 2006; Marean et al. 2007; Tribolo

12
thompson
et al. 2006). There was therefore likely a small period between ca. 100–92 ka during which
MSA occupation is recorded from both sites.
The time range covered by the sites includes both the arid and relatively cool Marine
Isotope Stage (MIS) 6 and the relatively warm MIS 5—including the height of the Last
Interglacial (MIS 5e) between about 130–119 ka. The continuous and in some cases overlap-
ping range of time represented by the two sites, along with their proximity to one another
within a 100-km stretch along the South African coast, makes them extremely well situated
for behavioral comparisons as well as understanding how changes in environment and local
ecology may have affected MSA faunal exploitation strategies. Such a relatively constrained
area and time range is also ideal for capturing any subtle variability in faunal acquisition, pro-
cessing, and transport strategies that the limited empirical record has previously rendered
undetectable.
Much of the variability that may be observed in MSA faunal assemblages could be
accounted for by differences in site context. Site context can refer to the behavioral, physical,
and environmental characteristics of the sites themselves and the artifactual assemblages
recovered from them. The behavioral context refers to the material culture traces that have
been recovered from each site. As discussed above, there are substantial differences between
the lithic assemblages from PP13B and Blombos, as well as differences between major strati-
graphic layers within each site (Henshilwood et al. 2001b; Marean et al. 2007). The abun-
dance of artifacts at Blombos that are less commonly recovered in MSA contexts (e.g.,
worked bone, ochre, and shell) indicates that this site was used differently than was PP13B—
or indeed most other MSA cave sites in South Africa—and that early symbolic behavior was
a component of this use. It is also possible that some of this differentiation in site-use could
be evident in the faunal assemblages. However, other attribute differences between the sites
could be invoked to explain differences in the acquisition, butchering, and transport of large
mammals.
Physically, Blombos is a small, isolated crevice inset into a cliff high above sea level
(ca. 35 m). PP13B is a large cave set 15 m above modern sea level, and is in close proximity
to other large openings on the landscape. Although not strictly a part of the physical config-
uration of the sites, sea level would have had an influence on their relative accessibility.
At the height of the Last Interglacial around 128 ka, with a global sea level that rose rapidly
to between approximately 4–6 m higher than today (Rohling et al. 2008), PP13B would have
become nearly inaccessible even if its physical properties did not otherwise change substan-
tially (Chappell and Shackleton 1986). In comparison, Blombos would have remained con-
sistently difficult to access for a different reason: its position on a steep cliff. In general, a
less accessible site would invoke greater transport costs, and decisions regarding which prey
body sizes and skeletal elements to bring to the site may be predicted in terms of these costs.
In contrast to most physical characteristics of the sites, environmental parameters such as
precipitation, temperature, and local biotic communities would have constantly shifted over
time, even on a decade scale. Current local data from paleoenvironmental proxies such as
speleothems, microfaunal assemblages, magnetic data, isotopes from shellfish, etc., are
either not available or not of sufficient resolution to be tied with much detail to the faunal
data presented here. However, broad descriptions of changes in proximity to the ocean, tem-
perature, and the overall ecology of the terrestrial and marine ecosystems of the Western
Cape are available and these provide a general basis for outlining key paleoenvironmental

13
Variability in Middle Stone Age faunal exploitation and use of landscapes
descriptions of the study sites (e.g., Cowling and Procheş 2005; Meadows and Baxter 1999;
Rau et al. 2002; van Andel 1989).
MIS 6 is generally accepted to have been a period of extreme climatic deterioration and
aridity in both the northern and southern hemispheres, followed by an extremely rapid cli-
matic amelioration (Augustin et al. 2004; Blunier et al. 1998; Jouzel et al. 1993; Rohling
et al. 2008; Schefuß et al. 2003). MIS 5 was basically warmer but with some fluctuations,
and MIS 4 represented another decline into cooler conditions. For comparative purposes the
deposits from PP13B and Blombos have been divided into several analytical units that take
these large-scale environmental parameters into account. At PP13B the data are divided into
MIS 6 and MIS 5, and at Blombos they are divided into the three major stratigraphic group-
ings recognized at the sites: M1 (corresponding to MIS 4), M2, and M3 (corresponding to
MIS 5).
transport and the MSA record
Ethnoarchaeological research has provided some valuable observations about patterning that
would be expected at transport sites for skeletal element and taxonomic abundance given dif-
ferent prey body sizes and carcass portions (e.g., Bunn 1988; Monahan 1998; O’Connell
et al. 1988, 1990). However, density-mediated attrition has been shown to provide a good
model for which elements and element portions will be best preserved in sheltered settings,
with higher density usually resulting in better preservation (e.g., Lam et al. 1998; 1999; 2003;
Lyman 1984). Thus, reconstructing carcass transport decisions from archaeological assem-
blages and determining how much of the patterning is attributable to human behavior and
how much to other taphonomic processes has been a topic of much debate (Bartram and
Marean 1999; Cleghorn and Marean 2004; Faith and Behrensmeyer 2006; Faith and Gordon
2007; Klein et al. 1999; Rogers 2000; Marean et al. 2004; Pickering et al. 2003; Stiner 2002).
Marean and Cleghorn (2003) approach the problem by suggesting that researchers restrict
comparisons of relative skeletal element abundances to within a low-survival or a high-sur-
vival set. Elements comprised entirely of low-density spongy bone fall into the low-survival
group and elements with a higher-density portion in addition to spongy portions (such as
mandibles and long bones) fall into the high-survival group. Unfortunately, when attempting
to apply optimal foraging theory to skeletal element abundances, less dense axial elements—
the low-survival set of Marean and Cleghorn (2003)—are also the elements with the highest
caloric return rate (Metcalfe and Jones 1988). They are furthermore the elements that require
more intensive processing to extract these calories, and are thus ethnoarchaeologically doc-
umented to be the elements most frequently transported away from a kill site (Monahan
1998). Finally, once transported they are elements that are potential candidates for purposeful
hominin fragmentation for grease extraction (Church and Lyman 2003). This presents a prob-
lem in which the elements with the highest caloric return will in most cases provide the least
reliable estimates of skeletal element abundance, despite the fact that optimal foraging theory
would predict them to be the most abundant elements at transport sites such as PP13B and
Blombos.
Owing to these taphonomic difficulties, many of the behavioral inferences that can be
made with regard to MSA transport strategies must be based on a constrained sample of ele-
ments, usually within the high-survival set. Marean and Cleghorn (2003:39) have wondered
if there is enough nutritive variability in such a sample to make a substantial difference in

14
thompson
hominin transport decisions, and therefore in a researcher’s ability to detect such decisions.
They suggest that there is, because within the long bones there are substantial differences in
both the gross and the net utility of metapodials (distal limbs) versus the other long bones that
make up the proximal limb (Madrigal and Holt 2002; Metcalfe and Jones 1988).
Marean et al. (2000b) have found suggestive patterning to this effect at the MSA site of
DK1, where there is a lack of metapodials relative to other long bones. Because their analysis
was restricted to high-survival elements from a subset of fragments that were accumulated
by MSA hominins, the authors felt confident in proceeding to an interpretation of these data.
They employed ethnoarchaeological observations of the modern Hadza in East Africa to
build an argument that the relative lack of metapodials at DK1 was attributable to these ele-
ments being the most frequently field-processed long bones, and are therefore the least likely
to be transported away from a kill or butchery site (Monahan 1998). This has been shown to
be particularly the case for larger (size 3 and 4) ungulates such as those that dominate the
hominin-accumulated fauna at DK1.
Under this scenario, DK1 would appear to have been used as an occupation site to which
high-utility elements were transported while heavier or lower-utility elements were purpose-
fully left behind at open points on the landscape. This implies that at the time that MSA
hunters encountered large prey they already had in mind that they would bring high-utility
portions back to DK1. In the case of the Hadza such butchery decisions are made because
other group members have remained behind at a set location on the landscape, and hunters
provision these members (Hawkes and Bird 2002). It is speculative if this also occurred
among MSA hominins, but certainly the implication is that there was a measure of planning
as each hunt took place. Furthermore, it implies that use of the landscape around DK1 was
structured and non-random, such that activities at one locality were mediated by knowledge
of the next locality to be visited and what resources would be available there.
Marean et al. (2000b: 221) have described the pattern at DK1 as “atypical for South
African MSA sites, and cave and rockshelter sites in general”. However, DK1 was at the time
of this statement the only MSA site for which a detailed taphonomic study was conducted
that included complete, unbiased assemblages. This assertion can now be examined in light
of new data from PP13B and Blombos, and an examination of its validity is used here to illus-
trate how variability in strategies for skeletal element transport across the MSA landscape
may be detected in the zooarchaeological record.
the proximal-distal limb transport records from pp13b and blombos
The Minimum Animal Units (MAU) was calculated for each analytical unit using MNE
counts generated by an image-analysis GIS program designed by Marean et al. (2001). Visual
examination of the MAU data shows a tendency at both sites for a dip in the representation
of metapodials relative to other long bones (figs. 3 and 4). It is also useful to look more close-
ly and quantitatively at the representation of individual skeletal elements relative to the gen-
eral pattern. For each set of MAU data the mean and standard deviation was determined. This
was done separately for the high- and low-survival groups. A z-score was then calculated for
each skeletal element so that its individual representation, in terms of standard deviations
from the mean, could be examined relative to other elements in the set. Points falling above
the dashed line are represented more often than the mean and points falling below the dashed
line are represented less often than the mean (figs. 5 and 6).

15
Variability in Middle Stone Age faunal exploitation and use of landscapes
The z-scores do not show an overall tendency for less metapodial representation. Instead,
there is substantial variation at both sites in the relative representation of metacarpals and
metatarsals, with some elements actually represented more often than others. As a further
step toward quantifying the representation of proximal versus distal limb elements, an
expected versus observed ratio can be examined. If complete limbs were transported without
bias against distal portions (metapodials), each forelimb should contain one humerus and
radius for every metacarpal. Similarly, the hindlimb should contain one femur and one tibia
for every metatarsal. The overall ratio between these bones in a complete-limb transport
strategy should therefore be 2:1, and the proximal limb elements of the humerus, radius,
femur, and tibia should account for 67.6% of the total. MAU values for proximal limb ele-
ments were summed separately from distal limb elements for large and small ungulates from
the grouped analytical units that comprise MIS 5 and MIS 6 at PP13B and from each layer
at Blombos. These are then displayed against the expected figure to determine if they truly
do appear to be underrepresented (fig. 7).
This analysis reveals that the pattern observed at DK1 in Layers 10 and 11 is not ubiqui-
tous throughout the MSA. Substantial variability appears to be present in relative distal limb
representation both between sites and between analytical units within sites. Most of this vari-
ability occurs within a lower-than-expected frequency of metapodials, as was originally sug-
gested to be the case at DK1. Much of it also records subtle differences in the representation
of distal limbs between body sizes within the same layer and analytical unit. However, this
is not borne out statistically when Fisher’s Exact Test was used to determine the statistical
robusticity of the observed pattern versus the expected pattern within each analytical unit.
Fig. 3. MAu data from MIS 5 (a) and MIS 6 (b) showing patterns of skeletal element
representation for small (size 1 and 2) and large (size 3, 4 and 5) ungulates.

16
thompson
Expected values were obtained by taking the sum of the proximal and distal limb MAU and
standardizing it by the ratio 2:1 (67.7% and 33.3%, respectively). These tests were conducted
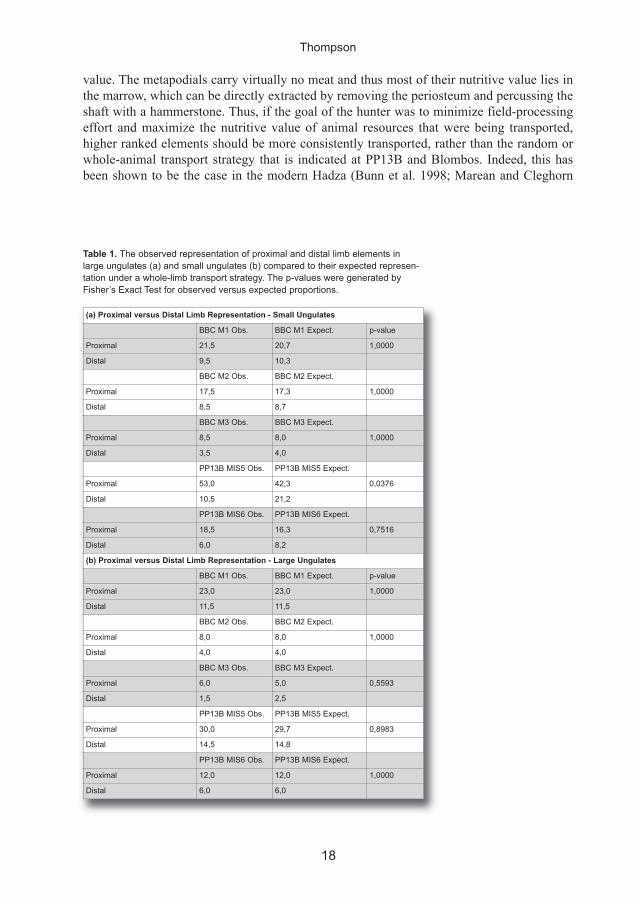
for all layers shown in figure 7 (table 1). The tests found that only one p-value was significant
below the α = 0.05 level, which indicates that although both sites visually appeared to be
impoverished in metapodials, they are in a statistical sense represented in the proportions that
would be expected relative to proximal limbs given a whole-limb transport strategy.
This is initially confusing, as some of the highest ranked long bones are the femur, tibia,
and humerus, which require processing of both meat and marrow to extract their maximum
Fig. 4. MAu data from M1 (a), M2 (b), and M3 (c) showing patterns of skeletal ele-
ment representation for small (size 1 and 2) and large (size 3, 4, and 5) ungulates.

17
Variability in Middle Stone Age faunal exploitation and use of landscapes
Fig. 5. Z-scores for the MAu of the high-survival
set for small (size 1 and 2, black boxes) and
large (size 3, 4, and 5, open circles) ungulates
in MIS 5 (a) and MIS 6 (b) at pp13b.
Fig. 6. Z-scores for the MAu of the high-survival
set for small (size 1 and 2, black boxes) and large
(size 3, 4, and 5, open circles) ungulates in M1 (a),
M2 (b), and M3 (c) at blombos.
Fig. 7. ratio of proximal to distal limb elements (by MAu) in each major stratigraphic
horizon from each site compared to the expected proportion in a whole-limb trans-
port strategy (top). Metapodial representation is less than expected if it falls to the
right of the white bar and more than expected if it falls to the left.
18
thompson
Table 1. the observed representation of proximal and distal limb elements in
large ungulates (a) and small ungulates (b) compared to their expected represen-
tation under a whole-limb transport strategy. the p-values were generated by
Fisher’s exact test for observed versus expected proportions.
(a) Proximal versus Distal Limb Representation - Small Ungulates
bbc M1 obs. bbc M1 expect. p-value
proximal 21,5 20,7 1,0000
distal 9,5 10,3
bbc M2 obs. bbc M2 expect.
proximal 17,5 17,3 1,0000
distal 8,5 8,7
bbc M3 obs. bbc M3 expect.
proximal 8,5 8,0 1,0000
distal 3,5 4,0
pp13b MIS5 obs. pp13b MIS5 expect.
proximal 53,0 42,3 0,0376
distal 10,5 21,2
pp13b MIS6 obs. pp13b MIS6 expect.
proximal 18,5 16,3 0,7516
distal 6,0 8,2
(b) Proximal versus Distal Limb Representation - Large Ungulates
bbc M1 obs. bbc M1 expect. p-value
proximal 23,0 23,0 1,0000
distal 11,5 11,5
bbc M2 obs. bbc M2 expect.
proximal 8,0 8,0 1,0000
distal 4,0 4,0
bbc M3 obs. bbc M3 expect.
proximal 6,0 5,0 0,5593
distal 1,5 2,5
pp13b MIS5 obs. pp13b MIS5 expect.
proximal 30,0 29,7 0,8983
distal 14,5 14,8
pp13b MIS6 obs. pp13b MIS6 expect.
proximal 12,0 12,0 1,0000
distal 6,0 6,0
value. The metapodials carry virtually no meat and thus most of their nutritive value lies in
the marrow, which can be directly extracted by removing the periosteum and percussing the
shaft with a hammerstone. Thus, if the goal of the hunter was to minimize field-processing
effort and maximize the nutritive value of animal resources that were being transported,
higher ranked elements should be more consistently transported, rather than the random or
whole-animal transport strategy that is indicated at PP13B and Blombos. Indeed, this has
been shown to be the case in the modern Hadza (Bunn et al. 1998; Marean and Cleghorn

19
Variability in Middle Stone Age faunal exploitation and use of landscapes
2003; Monahan 1998; O’Connell et al. 1988, 1990). It is further confusing that in general
when there is a tendency for metapodials to be less well represented, as at PP13B during
MIS 5, this is mainly the case for the small ungulates where selective transport is not predict-
ed given that it is relatively easy for even a lone hunter to transport an entire animal with min-
imal field processing (Monahan 1998).
A possible answer to this puzzle may lie within the types of resources that are available
for each element, rather than Standardized Food Utility (SFUI; Binford 1978; Metcalfe and
Jones 1988). Marrow varies in its composition both within the skeleton and in accordance
with the age and nutritional status of the animal from which it is derived. Young animals tend
to have marrow lower in fat content, and animals under nutritional stress deplete the fat
reserves in their marrow in a predictable sequence (Brooks et al. 1977; deCalesta et al. 1977;
Dunham and Murray 1982; Sinclair and Duncan 1972). Moreover, carnivores appear capable
of picking up on these subtle differences and will shift their ‘typical’ sequence of carcass por-
tion consumption to take these variables into account—effectively reorganizing their ranking
in bone choice in a way that is paleontologically invisible except possibly in the case of
neonates (Blumenschine 1986).
Based on interviews with the Nunamiut about their food preferences, Binford (1978) pro-
posed that there is a preference for marrow with higher oleic acid content, and that marrow
with higher proportions of such unsaturated fats can be easily identified using visual cues. In
a recent study, Morin (2007: 77) developed an Unsaturated Marrow Index (UMI), and sug-
gested that this variable is one that might influence long bone transport decisions independ-
ently of the total nutritive value of a skeletal element as measured by SFUI. This model
stands in contrast to that proposed by Jones and Metcalfe (1988), who indicate that simple
marrow volume is an effective measure of long bone ranking with regards to this resource.
Interestingly, distal limb elements such as the metapodials are the most enriched in oleic acid
content. Elements become progressively more impoverished in this unsaturated fat as one
moves proximally through the same bone and even more proximally through the entire limb.
The UMI index therefore predicts a correlation with long bone abundance that is gener-
ally the reverse of what would be predicted by food utility (which places the metapodials at
the lowest ranking). The only reason it is not the exact reverse is that the amount of unsatu-
rated fat is multiplied by the overall marrow volume as emphasized by Jones and Metcalfe
(1988). This has resulted in a weak (although insignificant) positive correlation with food
utility for the major long bones (Spearman’s Rho = 0.4857; p = 0.3287).
Thus, the UMI is convenient for zooarchaeologists because, when combined with the
SFUI index, a ready explanation becomes available for either a pattern of distal limb impov-
erishment or enrichment in an archaeological assemblage. In many ways, this risks failing to
identify subtle strategies of bone preference and transport in the archaeological record that
may not be attributable to either food utility or marrow quality. It also places certain key ele-
ments of the hindlimbs (owing to their high marrow volume) in positions where they can
have an extreme influence over the general pattern of skeletal element representation when
it is plotted versus UMI.
Furthermore, it is not certain what such a preference for unsaturated marrow should mean
in terms of transport. Given the relative ease of processing, the ready access to the marrow
in comparison with other long bones, and the higher content of oleic acid in their marrow, it
seems that the UMI in fact should predict greater emphasis on field processing of metapodi-
als to extract this resource immediately—with the result being lower representation of these

20
thompson
elements at the transport site. This appears to have been the case at PP13B, and particularly
for small ungulates during MIS 5, when the ratio of proximal to distal limbs is significantly
lower than expected.
One seeming inconsistency in the data is that between the z-scores, where certain metapo-
dials are in several cases better represented than some of the proximal elements, and in the
overall ratio of proximal: distal limb elements, where distal limbs are generally not as well
represented as are proximal elements. At PP13B the overall pattern of skeletal element trans-
port in the high-survival set seems to be one in which elements were not transported with
strict accordance to their food utility or even with regard to marrow quality. Thus, it would
seem that when making transport decisions MSA hominins at PP13B did not generally con-
sider the relative utilities of limb units. The significant difference seen in the small ungulates
from MIS 5 may represent a behavior that is not documented in modern hunter-gatherers, or
it may be explicable by a relatively greater contribution to this portion of the assemblage by
non-hominin bone collectors. Another possibility that cannot be ignored is that true patterns
in the data are rendered invisible as the result of a relatively small number of individual prey
acquisition and transport events over the course of a relatively large number of years.
It is also unknown to what extent the underlying social and behavioral factors that struc-
tured MSA food transport decisions are mirrored in the behavior of the Hadza, which com-
prise the major available dataset for ethnoarchaeological reference (Bunn et al. 1988; Marean
and Cleghorn 2003; Monahan 1998; O’Connell et al. 1988, 1990). The Hadza transport data
are based on a scenario in which kills are brought back to a central place, and where there are
varying group sizes available to transport these kills. If the foraging group sizes for MSA
hominins differed dramatically from those in the Hadza, then this model is not as useful for
understanding skeletal element transport as it might initially appear. Furthermore, there may
be a disjoin between the way in which archaeologists commonly perceive of caves as central
places and domestic sites and the way in which they were actually used during the MSA. If
PP13B was not a place to which MSA groups commonly returned immediately after a suc-
cessful hunt, the Hadza model would not be an appropriate analogue for the expected pat-
terning in larger ungulates.
The same general arguments can be made for the patterning at Blombos. The data could
be showing a whole-limb transport strategy that remained basically unchanged over time, or
they could be showing a series of individual events, each of which was defined by its own
set of contingent variables. This in turn implies that group sizes were relatively small and that
site-use may have been somewhat sporadic. In contrast, the data reported from DK1 (Marean
et al. 2000b) suggest that it may have been an MSA approximation of a central place for
hominin foraging, as is seen in many modern hunter-gatherer groups (e.g., Binford 1980;
Kelly 1995; Lupo 2001). Central place foraging is a strategy that is not by any means restrict-
ed to hominins (e.g., Sodhi 1992), and although it occurs in modern humans it is not neces-
sarily a hallmark of the condition of being ‘modern’. However, it is interesting that patterning
suggestive of such a strategy is not apparent at earlier sites such as PP13B or Blombos—and
it becomes tempting to ascribe this overall pattern between sites to more general changes in
MSA foraging group size, transport strategies, and partitioning of faunal resources. As noted
in a previous section, this is not the pattern that has been revealed in either lithic technology
or other artifact manufacture, and it demonstrates that there is much to be learned from the
faunal datasets to complement what we know about variability in MSA behavior overall.

21
Variability in Middle Stone Age faunal exploitation and use of landscapes
concluSIonS
Because carcass processing and transport occurs in several stages, each of which represents
a series of decisions by the butcher, human-modified faunal assemblages can be used to
understand MSA decision-making with regard to subsistence, and how these decisions may
have been modified to suit the particular context of a given site. Recent work indicates that
there was more variability in MSA behavior than has commonly been portrayed, but until the
faunal assemblages that accompany this evidence are subjected to a thorough taphonomic
analysis and examined closely, it will not be clear if such variability can also be detected in
MSA subsistence strategies. The very different site contexts observed between sites such as
Blombos and nearby sites such as PP13B and DK1 indicate that comparisons between their
faunal assemblages are a good place to begin examining the evidence for such variability.
Owing to the presence of marrow all long bones have nutritive value, but that of metapo-
dials is less overall relative to the upper limb portions because they are not also a meat-bear-
ing element (Metcalfe and Jones 1988). Relative representation of metapodials may therefore
be predicted to be higher for small animals because they can be more easily transported
whole (Metcalfe and Barlow 1992), and lower for large animals as these elements can be
quickly processed and discarded off-site. However, when taking into account the relatively
high importance that may be placed on elements with a high content of oleic acid, metapodi-
als may actually have a higher ranking than that implied by a simple food utility index based
on calories. All transport expectations should also be influenced by the relative accessibility
of a site, because there is a greater energetic cost for any element that is brought to a relative-
ly inaccessible location.
At both PP13B and Blombos the ratio of proximal to distal limbs initially seemed to be
biased against metapodials as has been reported from DK1. However, when analyzed statis-
tically the data most closely approximated a whole-limb transport strategy for all body size
classes during most time periods (the exception being small ungulates during MIS 5 at
PP13B). This could be because MSA hominins generally employed a whole-limb transport
strategy that did not change over thousands of years alongside concurrent changes in lithic
technology and other behaviors. This would present an interesting contrast to recent revela-
tions about variability in lithic technology, artifact manufacture, and the socio-symbolic
behaviors that underlie these material traces during a time period that has traditionally been
considered to be rather static and homogenous.
Alternatively, the faunal data may actually be showing a high degree of variability that
reflects different individual transport decisions, but the material traces of these decisions are
simply too affected by time-averaging to be resolved in the zooarchaeological record. This
would be the case if the sites were occupied at a low intensity over an extended period of
time, with each individual event based on so many unknowable variables that very little pat-
terning can remain. If this was the case, then the ‘speleo-centric’ view of caves as living sites
or home bases to which MSA hominins regularly returned with transported resources can be
attributed to these sites acting as preservational loci for human behavioral debris rather than
any actual preference for these sites on the parts of the hominins in question.
The issue of faunal transport is highly relevant to understanding MSA land use and set-
tlement dynamics. Variability in faunal exploitation patterns is almost certainly present in
taxonomic and body size representation, and may also have been important in skeletal ele-
ment transport decisions. It is absolutely critical that workers continue to build the empirical

22
thompson
record with standardized data collection methods, and that they employ analyses that facili-
tate the comparisons between sites. This, combined with better time resolution for these
assemblages, is what is needed to overcome many of the obstacles to understanding variabil-
ity in MSA faunal exploitation that have been discussed here. At that point, subtle details
regarding the structured use of faunal resources on the MSA landscape can finally begin to
be revealed, quantified, and described in terms of other traces of hominin behavior.
AcKnoWledgeMentS
Curtis Marean and Christopher Henshilwood provided access to the Pinnacle Point and
Blombos faunal assemblages, respectively. Sarah Wurz, Royden Yates, and Graham Avery all
assisted with invaluable research space and access to the comparative osteological collec-
tions housed at Iziko: South African Museums of Cape Town. Nick Conard kindly gave the
presentation that formed the basis of this paper on the author’s behalf at the UISPP meeting
in Lisbon. Catherine Haradon translated the abstract into French. Curtis Marean provided
access to the GIS program that was used to produce MNE data. Funding for collection of the
data presented here was provided by the Fulbright Foundation, a National Science
Foundation (NSF) Dissertation Improvement grant # 0620317 to Thompson, NSF grants
# BCS-9912465, BCS-0130713, and BCS-0524087 to Marean, funding from the Huxleys,
the Hyde Family Trust, the Institute for Human Origins, and Arizona State University.
reFerenceS
Aiello, l. c., 1993. the fossil evidence for mod-ern human origins in Africa: a revised view.American Anthropologist 95: 73–96.
Aiello, l. c., and p. Wheeler. 1995. the expen-sive-tissue hypothesis: the brain and thedigestive system in human and primate evolu-tion. Current Anthropology 36: 199–221.
Ambrose, S. h., and K. g. lorenz. 1990. Socialand ecological models for the middle stoneage in southern Africa. In The emergence ofmodern humans: an archaeological perspec-tive, ed. by p. Mellars, pp. 3–33. edinburgh:edinburgh university press.
Assefa, 2006. Faunal remains from porc-epic:paleoecological and zooarchaeological investi-gations from a Middle Stone Age site in south-eastern ethiopia. Journal of Human Evolution51: 50–75.
Augustin. l., c. barbante, p. r. F. barnes, J. M.bamola, M. bigler, e. castellano, o. cattani,J. chappellaz, d. dahl-Jensen, b. delmonte,g. dreyfus, g. durand, S. Falourd, h. Fischer,J. Flückiger, M. e. hansson, p. huybrechts, g.Jugie, S. Johnsen, J. Jouzel, p. Kaufmann, J.Kipfstuhl, F. lambert, V. y. lipenkov, g. c. lit-
tot, A. longinelli, r. lorrain, V. Maggi, V. Mas-son-delmotte, h. Miller, r. Mulvaney, J. oerle-mans, h. oerter, g. orombelli, F. parrenin, d.peel, J.-r. petit, d. raynaud, c. ritz, r. ruth,J. Schwander, u. Siegenthaler, r. Souchez, b.Stauffer, J. p. Steffensen, b. Stenni, t. F.Stocker, I. e. tabacco, r. udisti, r. S. W. vande Wal, M. van den broeke, J. Weiss, F. Wil-helms, J-.g. Winther, e. W. Wolff, and M. Zuc-chelli. 2004. eight glacial cycles from anAntarctic ice core. Nature 429: 623–628.
backwell, l., F. d’errico, and l. Wadley. 2008.Middle Stone Age bone tools from thehowieson’s poort layers, Sibudu cave, SouthAfrica. Journal of Archaeological Science 35:1566–1580.
bartram, l. e., and c. W. Marean. 1999. explain-ing the "Klasies pattern": Kua ethnoarchaeolo-gy, the die Kelders Middle Stone Age archae-ofauna, long bone fragmentation and carni-vore ravaging. Journal of Archaeological Science 26: 9–29.
bar-yosef, o., and S. l. Kuhn. 1999. the big dealabout blades: laminar technologies andhuman evolution. American Anthropologist101: 322–338.

23
Variability in Middle Stone Age faunal exploitation and use of landscapes
behrensmeyer, A. K. 1978. taphonomic and ecologic information from bone weathering. Paleobiology 4: 150–162.
binford, l. r. 1978. Nunamiut Ethnoarchaeology.new york: Academic press.
1980. Willow Smoke and dogs' tails: hunter-gatherer Settlement Systems and Archaeolog-ical Site Formation. American Antiquity 45: 4–20.
1984. The Faunal Remains from Klasies RiverMouth. new york: Academic press.
blumenschine, r. J. 1986. carcass consumptionsequences and the archaeological distinctionof scavenging and hunting. Journal of HumanEvolution 15: 639–659.
blumenschine, r. J., c. W. Marean, and S. d.capaldo. 1996. blind tests of inter-analyst cor-respondence and accuracy in the identificationof cut marks, percussion marks, and carnivoretooth marks on bone surfaces. Journal ofArchaeological Science 23: 493–507.
blunier, r., J. chappell, J. Schwander, A. dällen-bach, b. Stauffer, t. F. Stocker, d. raynaud, J. Jouzel, h. b. clausen, c. u. hammer, andS. J. Johnsen. 1998. Asynchrony of Antarcticand greenland climate change during the lastglacial period. Nature 394: 739–743.
broadhurst, c. l., y. Wang, M. A. crawford, S. c.cunnane, J. e. parkington, and W. F. Schmidt.2002. brain-specific lipids from marine, lacus-trine, or terrestrial food resources: potentialimpact on early African homo sapiens. Com-parative Biochemistry and Physiology Part B131: 653–673.
brooks, p. M., J. hanks, and J. V. ludbrook.1977. bone marrow as an index of condition inAfrican ungulates. South African Journal ofWildlife Research 7: 61–66.
bunn, h. t., l. e. bartram, and e. M. Kroll. 1988.Variability in bone assemblage formation fromhadza hunting, scavenging, and carcass processing. Journal of Anthropological Archaeology 7: 412–57.
burger, o., M. J. hamilton, and r. Walker. 2005.the prey as patch model: optimal handling ofresources with diminishing returns. Journal ofArchaeological Science 32: 1147–1158.
cain, c. r. 2004. notched, flaked and groundbone artefacts from Middle Stone Age andIron Age layers of Sibudu cave, KwaZulu-
natal, South Africa. South African Journal of
Science 100: 195–197.
2006. human activity suggested by the taphon-omy of 60 ka and 50 ka faunal remains fromSibudu cave. Southern African Humanities 18:241–260.
capaldo, S. d., and r. J. blumenschine. 1994. A quantitative diagnosis of notches made byhammerstone percussion and carnivore gnaw-ing on bovid long bones. American Antiquity
59: 724–748.
chappell, J., and n. J. Shackleton. 1986. oxygenisotopes and sea level. Nature 324: 137–140.
chase, p. g., and h. dibble. 1987. Middle palae-olithic symbolism: A review of current evidenceand interpretation. Journal of Anthropological
Archaeology 6: 263–96.
church, r. r., and r. l. lyman 2003. Small fragments make small differences in efficiencywhen rendering grease from fractured artio-dactyl bones by boiling. Journal of Archaeo-
logical Science 30: 1077–1084.
clark, J. d., 1988. the Middle Stone age of eastAfrica and the beginnings of regional identity.Journal of World Prehistory 2: 235–303.
clark, J. l, and I. plug. 2008. Animal exploitationstrategies during the South African MiddleStone Age: howiesons poort and post-howiesons poort fauna from Sibudu cave.Journal of Human Evolution 54: 886–898.
cowling, r. M., and Ş. procheş. 2005. patternsand evolution of plant diversity in the capeFloral region. Biol. Skrifter 55: 273–288.
d'errico, F., c. S. henshilwood, and p. nilssen.2001. An engraved bone fragment from c.70,000-year-old Middle Stone Age levels atblombos cave, South Africa: Implications forthe origin of symbolism and language. Antiquity 75: 309–18.
d'errico, F., and c. S. henshilwood. 2007. Addi-tional evidence for bone technology in theSouthern African Middle Stone Age. Journal
of Human Evolution 52: 142–163.
deacon, h. J. 1989. late pleistocene palaeoecol-ogy and archaeology in the Southern cape,South Africa. In The Human Revolution, ed. by p. A. Mellars, and c. Stringer, pp. 547–564.edinburgh: edinburgh university press.

24
thompson
decalesta, d. S., J. g. nagy, and J. A. bailey.1977. experiments on starvation and recoveryin mule deer does. Journal of Wildlife Manage-ment 41: 81–86.
dunham, K. M, and M. g. Murray. 1982. the fatreserves of impala, Aepyceros melampus.African Journal of Ecology 20: 81–87.
eaton, S. b., and M. J. Konner. 1997. paleolithicnutrition revisited: a twelve year retrospectiveon it s nature and implications. European Journal of Clinical Nutrition 51: 207–16.
Faith, J. t., and A. K. behrensmeyer. 2006.changing patterns of carnivore modification ina landscape bone assemblage, Amboselipark, Kenya. Journal of Archaeological Sci-ence 33: 1718–1733.
Faith, J. t., and A. d. gordon. 2007. Skeletal ele-ment abundances in archaeofaunal assem-blages: economic utility, sample size, andassessment of carcass transport strategies.Journal of Archaeological Science 34: 872–882.
Faith, J. t., 2008. eland, buffalo, and wild pigs:were Middle Stone Age humans ineffectivehunters? Journal of Human Evolution 55: 24–36.
Feathers, J. K., and d. A. bush. 2000. lumines-cence dating of the Middle Stone Age depositsat die Kelders. Journal of Human Evolution38: 91–120.
Foley, r. A., and M. M. lahr. 1997. Mode 3 tech-nologies and the evolution of modern humans.Cambridge Archaeological Journal 1: 3–36.
gamble, c. 1994. Timewalkers. cambridge, Massachusetts: harvard university pres,.
halkett, d., t. hart, r. yates, t. p. Volman, J. parkington, J. orton, r. g. Klein, K. cruz-uribe, and g. Avery. 2003. First excavation ofintact Middle Stone Age layers at ysterfontein,Western cape province, South Africa: implica-tions for Middle Stone Age ecology. Journal ofArchaeological Science 30: 955–973.
hawkes, K., J. o’connell, and n. g. blurtonJones. 1991. hunting income patterns amongthe hadza: big game, common goods, forag-ing goals, and the evolution of the human diet.Philosophical Transactions of the Royal Society, Section b, 334: 243–251.
hawkes, K. 1996. the evolutionary basis of sexvariations in the use of natural resources:
human examples. Population and Environ-ment 18: 161–173.
hawkes, K., and r. bird. 2002. Showing off,handicap signaling, and the evolution of men’swork. Evolutionary Anthropology 11: 58–67.
henshilwood, c. S., and J. Sealy. 1997. boneartefacts from the Middle Stone Age at blom-bos cave, South Africa. Current Anthropology38: 890–895.
henshilwood, c. S., F. d'errico, c. W. Marean, r. g. Milo, and r. yates. 2001a. An earlybone tool industry from the Middle Stone Ageat blombos cave, South Africa: Implicationsfor the origins of modern human behavoiur,symbolism and language. Journal of HumanEvolution 41: 631–78.
henshilwood, c. S., r. yates, K. cruz-uribe, p. goldberg, F. e. grine, r. g. Klein, c. poggenpoel, K. Van niekerk, and I. Watts.2001b. blombos cave, Southern cape, SouthAfrica: preliminary report on the 1992–1999excavations of the Middle Stone Age levels.Journal of Archaeological Science 28: 421–48.
henshilwood, c. S., F. d'errico, r. yates, Z. Jacobs, c. tribolo, g. A. duller, t. Mercier,J. Sealy, h. Valladas, I. Watts, and A. g. Win-tle. 2002. emergence of modern humanbehavior: Middle Stone Age engravings fromSouth Africa. Science 295: 1278–80.
henshilwood, c. S., and c. W. Marean. 2003.the origin of modern human behavior: a cri-tique of the models and their test implications.Current Anthropology 44: 627–651.
henshilwood, c. S., F. d'errico, M. Vanhaeren, K. Van niekerk, and Z. Jacobs. 2004. MiddleStone Age shell beads from South Africa. Science 304: 404.
hovers, e., S. Ilani, o. bar-yosef, and b. Vander-meersch. 2003. An early case of color symbol-ism: ochre use by modern humans in Qafzehcave. Current Anthropology 44: 491–522.
Jacobs, Z., A. g. Wintle, and g. A. t. duller.2003a. optical dating of dune sand fromblombos cave, South Africa: I – multiple graindata. Journal of Human Evolution 44: 599–612.
Jacobs, Z., g. A. t. duller, and A. g. Wintle.2003b. optical dating of dune sand fromblombos cave, South Africa: II – single graindata. Journal of Human Evolution 44: 613–26.

25
Variability in Middle Stone Age faunal exploitation and use of landscapes
Jacobs, Z., g. A. t. duller, A. g. Wintle, c. S.henshilwood. 2006. extending the chronologyof deposits at blombos cave, South Africa,back to 140 ka using optical dating of singleand multiple grains of quartz. Journal ofHuman Evolution 51: 255–273.
Jacobs Z., r. g. roberts, r. F. galbraith, h. J.deacon, r. grün, A. Mackay, p. Mitchell, r.Vogelsang, and l. Wadley. 2008. Ages for theMiddle Stone Age of southern Africa: implica-tions for human behavior and dispersal. Sci-ence 322: 733–735.
Jones, K. t., and d. Metcalfe. 1988. bare bonesarchaeology: bone marrow indices and effi-ciency. Journal of Archaeological Science 15:415–423.
Jouzel, J., n. I. barkov, J. M. barnola, M. bender,J. chappellaz, c. genthon, V. M. Kotlyakov, V. lipenkov, c. lorius, J. r. petit, d. raynaud,g. raisbeck, c. ritz, t. Sowers, M. Stieve-nard, F. yiou, and p. yiou. 1993. extending theVostok ice-core record of palaeoclimate to thepenultimate glacial period. Nature 364: 407–412.
Kaplan, h., K. hill, J. lancaster, and A. Magdale-na-hurtado. 2000. A theory of human life his-tory evolution: diet, intelligence, and longevity.Evolutionary Anthropology 9: 156–185.
Kelly, r. l. 1995. The Foraging Spectrum: Diver-sity in Hunter-Gatherer Lifeways. Washington,d.c.: Smithsonian Institution press.
Klein, r. g. 1975. Middle Stone Age man-animalrelationships in southern Africa: evidence fromdie Kelders and Klasies river Mouth. Science190: 265–67.
1976. the mammalian fauna of the Klasies riv-er Mouth sites, Southern cape province,South Africa. South African ArchaeologicalBulletin 31: 75–98.
1977. the mammalian fauna from the Middleand later Stone Age (later pleistocene) levelsof border cave, natal province, South Africa.South African Archaeological Bulletin 32: 14–28.
1978a. Stone Age predation on large Africanbovids. Journal of Archaeological Science 5:195–217.
1978b. A preliminary report on the larger mam-mals from the boomplaas stone age cave site,cango Valley, oudtshoorn district, South
Africa. South African Archaeological Bulletin33: 66–75.
1987. reconstructing how early people exploit-ed animals: problems and prospects. In TheEvolution of Human Hunting, ed. by M. h. nitecki and d. V. nitacki, pp. 11–45.new york and london: plenum press.
1989a. Why does skeletal part representationdiffer between smaller and larger bovids atKlasies river Mouth and other archeologicalsites? Journal of Archaeological Science 6:363–81.
1989b. biological and behavioural perspectiveson modern human origins in Southern Africa.In The Human Revolution, ed. by p. A. Mellarsand c. Stringer, pp. 529–46. edinburgh: edinburgh university press.
1995. Anatomy, behavior, and modern humanorigins. Journal of World Prehistory 9: 167–98.
2000. Archeology and the evolution of humanbehavior. Evolutionary Anthropology 9: 17–36.
2003. Whither the neanderthals? Science 299:1525–31.
Klein, r. g., K. cruz-uribe, and r. g. Milo. 1999.Skeletal part representation in archaeofaunas:comments on "explaining the 'Klasies pat-tern': Kua ethnoarchaeology, the die KeldersMiddle Stone Age archaeofauna, long bonefragmentation and carnivore ravaging" by bar-tram and Marean. Journal of ArchaeologicalScience 26: 1225–34.
Klein, r. g., and K. cruz-uribe. 2000. Middle andlater Stone Age large mammal and tortoiseremains from die Kelders cave 1, Westerncape province, South Africa. Journal ofHuman Evolution 38: 169–96.
Klein, r. g., g. Avery, K. cruz-uribe, d. halkett,J. e. parkington, t. Steele, t. p. Volman, andr. yates. 2004. the ysterfontein 1 MiddleStone Age site, South Africa, and early humanexploitation of coastal resources. Proceedingsof the National Academy of Science 101:5708–5715.
lam, y. M., x. cehn, c. W. Marean, and c. J.Frey. 1998. bone density and long bone repre-sentation in archaeological faunas: comparingresults from ct and photon densitometry.Journal of Archaeological Science 25: 559–70.
lam, y. M., x. chen, and o. M. pearson. 1999.Intertaxonomic variability in patterns of bone

26
thompson
density and the differential representation ofbovid, cervid, and equid elements in thearchaeological record. American Antiquity 64:343–362.
lam, y. M., o. M. pearson, c. W. Marean, and x. chen. 2003. bone density studies in zooar-chaeology. Journal of Archaeological Science30: 1701–1708.
langdon, J. 2006. has an aquatic diet been nec-essary for hominin brain evolution and func-tional development? British Journal of Nutrition96: 7–17.
lupo, K. d. 2001. Archaeological skeletal partprofiles and differential transport: an ethnoar-chaeological example from hadza boneassemblages. Journal of AnthropologicalArchaeology 20: 361–378.
lyman, r. l. 1984. bone density and differentialsurvivorship of fossil classes. Journal ofAnthropological Archaeology 3: 259–99.
Mackay, A., and A. Welz. 2008. engraved ochrefrom a Middle Stone Age context at Klein Kliphuis in the Western cape of South Africa.Journal of Archaeological Science.
Madrigal, t. c., and J. Z. holt. 2002. White-taileddeer meat and marrow return rates and their application to eastern Woodlandsarchaeology. American Antiquity 67: 745–759.
Marean, c. W., and l. bertino. 1994. Intrasitespatial analysis of bone: Subtracting the effectof secondary carnivore consumers. AmericanAntiquity 59: 748–68.
Marean, c. W., and c. J. Frey. 1997. Animalbones from caves to cities: reverse utilitycurves as methodological artifacts. AmericanAntiquity 62: 698–711.
Marean, c. W., and S. y. Kim. 1998. Mousterianlarge-mammal remains from Kobeh cave:behavioral implications for neanderthals andearly modern humans. Current Anthropology39: S79–S113.
Marean, c. W., and Z. Assefa. 1999. Zooarcheo-logical evidence for the faunal exploitationbehavior of neandertals and early modernhumans. Evolutionary Anthropology 8: 22–37.
Marean, c. W., p. goldberg, g. Avery, F. e. grine,and r. g. Klein. 2000a. Middle Stone Agestratigraphy and excavations at die Kelderscave 1 (Western cape province, South
Africa): the 1992, 1993, and 1995 field sea-sons. Journal of Human Evolution 38: 7–42.
Marean, c. W., y. Abe, c. J. Frey, and r. c. ran-dall. 2000b. Zooarchaeological and taphonom-ic analysis of the die Kelders cave 1 layers10 and 11 Middle Stone Age larger mammalfauna. Journal of Human Evolution 38: 197–233.
Marean, c. W., and J. c. thompson. 2003.research on the origins of modern humanscontinues to dominate paleoanthropology.Evolutionary Anthropology 12: 165–67.
Marean, c. W., and n. cleghorn. 2003. largemammal skeletal element transport: applyingforaging theory in a complex taphonomic sys-tem. Journal of Taphonomy 1: 15–42.
Marean, c. W., M. domínguez-rodrigo, and t. r. pickering. 2004. Skeletal element equifi-nality in zooarchaeology begins with method:the evolution and status of the “shaft critique”.Journal of Taphonomy 2: 69–98.
Marean, c.W., M. bar-Matthews, J. bernatchez,e. Fisher, p. goldberg, A. I. r. herries, Z. Jacobs, A. Jerardino, p. Karkanas, t. Minichillo, p. nilssen, e. thompson, I. Watts, and h. Williams. 2007. early humanuse of marine resources and pigment in SouthAfrica during the Middle pleistocene. Nature449: 905–908.
Mcbrearty, S., and A. S. brooks. 2000. the revo-lution that wasn't: a new interpretation of theorigin of modern human behavior. Journal ofHuman Evolution 39: 453–563.
Mccall, g. 2007 behavioral ecological models oflithic technological change during the laterMiddle Stone Age of South Africa. Journal ofArchaeological Science 34: 1738–1751.
Meadows, M. e., and A. J. baxter. 1999. lateQuaternary palaeoenvironments of the south-western cape, South Africa: a regional synthe-sis. Quaternary International 57/58: 193–206.
Mellars, p. 1989. Major issues in the origins ofmodern humans. Current Anthropology 30:349–85.
Mellars, p., and c. Stringer. 1989. The HumanRevolution. edinburgh: edinburgh universitypress.
Metcalfe, d., and K. t. Jones. 1988. A reconsider-ation of animal body-part utility indices. American Antiquity 53: 486–504.

27
Variability in Middle Stone Age faunal exploitation and use of landscapes
Metcalfe, d., and K. r. barlow. 1992. A model forexploring the optimal trade-off between fieldprocessing and transport. American Anthropologist 94: 340–356.
Miller, g. h., p. b. beaumont, h. J. deacon, A. S. brooks, p. e. hare, and A. J. t. Jull.1999. earliest modern humans in southernAfrica dated by isoleucine epimerization inostrich eggshell. Quaternary Science Reviews 18: 1537–1548.
Milo, r. g. 1998. evidence for hominid predationat Klasies river Mouth, South Africa, and itsimplications for the behavior of early modernhumans. Journal of Archaeological Science25: 99–133.
Milton, K. 1999. A hypothesis to explain the role of meat-eating in human evolution. Evolutionary Anthropology 8: 11–21.
Mithen, S. 1999. Symbolism and the supernatu-ral. In The Evolution of Culture, ed. by r. dunbar, c. Knight, and c. power, pp. 147–72. edinburgh: edinburgh universitypress.
Monahan, c. M. 1998. the hadza carcass trans-port debate revisited and its archaeologicalimplications. Journal of Archaeological Science 25: 405–24.
Morin, e. 2007. Fat composition and nunamiutdecision-making: a new look at the marrowand bone grease indices. Journal of Archaeo-logical Science 34: 69–82.
o'connell, J. F., K. hawkes, and n. g. blurtonJones. 1988. hadza hunting, butchering, andbone transport and their archaeological impli-cations. Journal of Anthropological Research44: 131–55.
1990. reanalysis of large mammal body parttransport among the hadza. Journal ofArchaeological Science 17: 301–16.
pickering, t. r., c. W. Marean, and M. domínguez-rodrigo. 2003. Importance oflimb bone shaft fragments in zooarchaeology:a response to "on in situ attrition and verte-brate body part profiles" (2002), by M. c. Stin-er. Journal of Archaeological Science 30:1469–82.
poggenpoel, c.A., J. rigaud, p. texier, and J. e. parkington. 2005. From tool to symbols:the behavioural context of intentionallymarked ostrich eggshell from diepkloof, West-ern cape. In From Tools to Symbols: From
Early Hominids to Modern Humans, ed. by F. d'errico, and l. backwell, pp. 475–492.Johannesburg: Witswatersrand universitypress.
rau, A. J., J. rogers, J. r. e. lutjeharms, J. giraudeau, J. A. lee-thorp, M.-t. chen,and c. Waelbroeck. 2002. A 450-kyr record ofhydrological conditions on the western Agulhas bank Slope, south of Africa. MarineGeology 180: 183–201.
relethford, J. h. 1995. genetics and modernhuman origins. Evolutionary Anthropology 4:53–63.
rogers, A. r. 2000. on equifinality in faunalassemblages. American Antiquity 65: 709–723.
rohling, e. J., K. grant, ch. hemleben, M. b. Siddall, A. A. hoogakker, M. bolshaw,and M. Kucera. 2008. high rates of sea-levelrise during the last interglacial period. NatureGeoscience 1: 38–42.
Schefuß, e., S. Schouten, J. h. F. Jansen, and J. S. Sinninghe damsté. 2003. African vegeta-tion controlled by tropical sea surface temper-atures in the mid-pleistocene period. Nature422: 418–421.
Schwarcz, h. p., and W.J. rink. 2000. eSr dat-ing of the die Kelders cave 1 site, SouthAfrica. Journal of Human Evolution 38: 121–29.
Sinclair, A. r. e., and p. duncan. 1972. Indices of condition in tropical ruminants. East AfricanWildlife Journal 10: 143–149.
Sodhi, n. S. 1992. central place foraging andprey preparation by a specialist predator, theMerlin. Journal of Field Ornithology 63: 71–76.
Speth, J., and K. Spielman. 1983. energy source,protein metabolism, and hunter gatherer sub-sistence strategies. Journal of AnthropologicalArchaeology 2: 1–31.
Stiner, M. c. 2002. on in situ attrition and vertebrate body part profiles. Journal ofArchaeological Science 29: 979–91.
thackeray, A. I. 1993. the Middle Stone Agesouth of the limpopo river. Journal of WorldPrehistory 6: 385–440.
2000. Middle Stone Age artefacts from the 1993and 1995 excavations at die Kelders cave 1,

28
thompson
South Africa. Journal of Human Evolution 38:147–68.
thompson, J. c. 2005. the impact of post-depo-sitional processes on bone surface modifica-tion frequencies: A corrective strategy and itsapplication to the loiyangalani site, Serengetiplain, tanzania. Journal of Taphonomy 3: 57–80.
2008. Zooarchaeological tests for modernhuman behavior at blombos cave and pinna-cle point cave 13b, southwestern cape,South Africa. unpublished ph.d. dissertation,Arizona State university.
n. d. taphonomic analysis of the Middle StoneAge faunal assemblage from pinnacle pointcave 13b, Western cape, South Africa. Jour-nal of Human Evolution. (under revision)
tribolo, c. 2003. Apport des methods de la lumi-nescence a la chronologie des techno-faciesdu Middle Stone Age associes aux premiershommes modernes du Sud de l’Afrique. ph.d.dissertation, university of bordeaux.
tribolo, c., n. Mercier, M. Selo, J-l. Joron, J-l. reyss, c. henshilwood, J. Sealy, r. yates. 2006. tl dating of burnt lithics fromblombos cave (South Africa): further evidencefor the antiquity of modern human behaviour.Archaeometry 48: 341–357.
van Andel, t. h. 1989. late pleistocene sea lev-els and human exploitation of the shore andshelf of southern South Africa. Journal ofWorld Prehistory 6: 385–439.
Villa, p., and e. Mahieu. 1991. breakage patternsof human long bones. Journal of Human Evolution 21: 27–48.
Volman, t. p. 1984. early prehistory of SouthernAfrica. In Southern African Prehistory andPaleoenvironments. Balkema, ed. by r. g. Klein, pp. 169–220. rotterdam.
Wadley, l. 2007. Announcing a Still bay industryat Sibudu cave, South Africa. Journal ofHuman Evolution 52: 681–689.
Wadley, l., and Z. Jacobs. 2004. Sibudu cave,KwaZulu-natal: background to the excavationsof Middle Stone Age and Iron Age occupa-tions. South Africa Journal of Science 100:145–151.
2006. Sibudu cave: background to the excava-tions, stratigraphy and dating. SouthernAfrican Humanities 18: 1–26.
Watts, I. 1999. the origin of Symbolic culture. In The Evolution of Culture, ed. by r. dunbar,c. Knight, and c. power, pp. 113–146. edinburgh: edinburgh university press.
Wood, b. 2006. prestige or provisioning? A test offoraging goals among the hadza. CurrentAnthropology 47: 383–387.
Wurz, S. 2002. Variability in the Middle Stone Agelithic sequence 115,000–60,000 years ago atKlasies river, South Africa. Journal of Archaeological Science 21: 1001–1015.





























