Embed Size (px)
Citation preview

A NEW PIPINE FROG FROM AN EOCENE CRATER
LAKE IN NORTH-CENTRAL TANZANIA
by ANA MARIA BAEZ* and TERRY HARRISON�
*Department of Geology, Facultad de Ciencias Exactas, Universidad de Buenos Aires, Pabellon 2, Ciudad Universitaria, 1428 Buenos Aires, Argentina;
e-mail: [email protected]�Department of Anthropology, New York University, 25 Waverly Place, New York, NY 10003, USA
Typescript received 23 January 2004; accepted in revised form 22 April 2004
Abstract: The fossiliferous site of Mahenge, Tanzania, inter-
preted as a small lake that formed in a kimberlite intrusion,
has yielded a vertebrate assemblage that includes scarce frog
remains. Radiometric dating of the pipe indicates that the
volcanic activity took place at 45Æ83 ± 0Æ17 Ma, whereas infil-
ling of the crater was completed in 0Æ2–1Æ0 myr after
emplacement. The frogs, described herein, are preserved as
partially articulated skeletons that represent a new pipine
taxon, Singidella latecostata gen. et sp. nov. A parsimony
analysis of fossil and extant pipoid frogs indicates that
Singidella is more closely related to African Hymenochirus
and Pseudhymenochirus than to South American Pipa. It also
provides weak support for the sister-group relationship of
Singidella and the bizarre Late Cretaceous Pachycentrata ta-
queti (replacement name for Pachybatrachus taqueti) from
Niger, although the incomplete preservation of Pachycentrata
precludes thorough comparisons. This record indicates that
prior to the rifting in the Miocene the distribution of pipines
extended into eastern Africa where they do not occur today.
Key words: Eocene, Africa, Pipidae, Pipinae, Hymeno-
chirini, phylogeny, Singidella gen. nov.
Pipid frogs are a characteristic component of the present
African fauna. Pipids have a distinctive morphology that
has long been interpreted as a character suite for an aquatic
mode of life (Noble 1954; Cannatella and Trueb 1988a;
Trueb 1996), although their phylogenetic relationships
within Anura are still controversial (Cannatella and Trueb
1988a; Garcıa Parıs et al. 2003; Haas 2003; Pugener et al.
2003). Both clades that compose this monophyletic group,
Xenopodinae and Pipinae, are represented in Africa today.
However, in contrast to xenopodines, which are restricted
to Africa, pipines have a disjunct distribution, with
hymenochirines (Hymenochirus and Pseudhymenochirus)
occurring in equatorial Africa and Pipa inhabiting tropical
South America east of the Andes and Panama (Duellman
and Trueb 1986). Available palaeontological data (Baez and
Rage 1998) indicate that the presence of the hymenochirine
lineage in Africa dates back at least to the Late Cretaceous.
By contrast, no undisputed Mesozoic or Tertiary pipine has
yet been reported from South America (Baez 1996; Baez
et al. 2000). However, if Pipa and hymenochirines are sister
groups, according to our present understanding of pipid
phylogeny (Cannatella and Trueb 1988a; Ford and Canna-
tella 1993; Baez and Trueb 1997), the lineage represented
by Pipa today had already diverged from other pipines by
the Late Cretaceous.
Herein we describe pipid remains from the Eocene site
of Mahenge, located near the locality of Singida, Tanzania
(Text-fig. 1). The specimens consist of partially articulated
skeletons and were identified as representing a new pipine
taxon (Baez 1999; Harrison et al. 2001). This preliminary
taxonomic placement is corroborated by a parsimony
analysis of fossil and extant pipoid frogs. Moreover, this
analysis strongly supports a relationship between the new
taxon and hymenochirines, and suggests a sister-group
relationship with the bizarre, hyperossified, but partially
preserved, Pachycentrata (generic name proposed by Baez
and Rage 2004 to replace Pachybatrachus because of
homonymy) from the Upper Cretaceous of Niger (Baez
and Rage 1998). This record sheds new light on the evo-
lution of several unique features of the still poorly known
African pipines.
GEOLOGICAL CONTEXT AND AGE
The site of Mahenge (4� 75¢ 38¢¢ S, 34� 15¢ 28¢¢ E), located
63 km west of the town of Singida in north-central
Tanzania (Text-fig. 1), is unique in being the only early
Tertiary site in East Africa to have yielded the remains of
fossil vertebrates. A detailed review of the regional and
local geology is presented in Harrison et al. (2001). The
fossiliferous lacustrine sediments overlie one of the largest
kimberlite pipes in the region (Mannard 1962; Edwards
and Howkins 1966; Mitchell 1986; Harrison et al. 2001).
[Palaeontology, Vol. 48, Part 4, 2005, pp. 723–737]
ª The Palaeontological Association 723

During the Cretaceous and Palaeogene, the basement
rocks in northern Tanzania were disrupted by the intru-
sion of a swarm of kimberlite pipes (Williams 1939; Man-
nard 1962; Dawson 1970; Mitchell 1986). A Pb-U date for
the intrusive event at Mahenge has yielded an age of
45Æ83 ± 0Æ17 Ma (Harrison et al. 2001), correlating with
the early Middle Eocene (Lutetian). The kimberlite pipe
at Mahenge formed a small crater bounded by a steep-
sided wall of basement rock. Subsequent eruptions of pyr-
oclastic kimberlite produced a series of well-stratified,
fine-grained tuffs that formed a low ejecta cone around
the rim of the crater (Mannard 1962; Hawthorne 1975).
After the volcanic activity ceased, the crater filled with
water to produce a shallow, circular crater-lake, with a
diameter of c. 370 m (Mannard 1962; Harrison et al.
2001). The inner slope of the tuff cone was probably rel-
atively steep, forming a narrow shoreline (Edwards and
Howkins 1966; Smith 1986). Material eroding from the
tuff cone was washed into the lake to produce a series of
sedimentary tuffaceous beds (Mannard 1962; Smith 1986;
Harrison et al. 2001). Based on conservative estimates of
sedimentation rate and compaction, it is thought that the
crater at Mahenge would have been filled in 0Æ2–1Æ0 myr
(Harrison et al. 2001). This provides an estimated date
for the fossiliferous horizons of c. 46–47 Ma.
The bottom of the section consists of massive, yellow-
grey clayey sandstone (Text-fig. 2). The sediments are
relatively soft and friable, with only weak horizontal bed-
ding. Overlying the sandstone is an alternating series of
well-stratified calcareous mudstones and soft friable shales
containing well-preserved fossils. Although the average
thickness of the mudstones is 0Æ35 m, they can form
prominent, pavement-like layers up to 0Æ6 m thick. The
thickest and most easily definable of the mudstones have
been designated as marker beds; these are numbered Lev-
els 1–8 from the top of the sequence down. The sedi-
ments between each marker bed are identified as
sublevels; thus, Level 1 is separated from Level 2 by Sub-
level 1. Above the fossiliferous beds is a layer of slumped
blocks of consolidated mudstones in a matrix of friable
yellowish grey shale. The uppermost layer consists of a
recently formed colluvium of dark grey clay and sandy
loams containing numerous detrital fragments of granite
and quartz, and aggregations of decomposed kimberlite.
In addition to the fossil frogs described here, the fauna
and flora include a microchiropteran bat (Tanzanycteris
mannardi) (Gunnell et al. 2003), a rich assemblage of fos-
sil fishes (Palaeodenticeps tanganikae, Singida jacksonoides,
Mahengechromis spp., Mahengecharax carrolli, Eocitharinus
macrognathus, Chrysichthys mahengeensis, and an
unnamed osteoglossomorph) (Greenwood 1960; Green-
wood and Patterson 1967; Murray 2000, 2001, 2003a, b;
Murray and Budney 2003), several undescribed insects
(Harrison et al. 2001) and well-preserved plant remains
(Herendeen and Jacobs 2000). Layers 7 and 8 (Text-
fig. 2) are the most productive fossil horizons, and these
alone have yielded 57 per cent of the fishes, 56 per cent
of the leaves and all of the insects. The fossil frogs were
recovered from Sublevel 7 (WM 400 ⁄ 96 and 401 ⁄ 96) and
Level 8 (WM 502 ⁄ 96 and 503 ⁄ 96). The remarkably
complete preservation of the fossils, and the absence of
any evidence of damage or dispersal by benthic scaven-
gers or bioturbation, suggest stratification of the water
column in the lake in which the deeper, cooler and
anoxic waters would have been a hostile environment for
most organisms (Beadle 1981; Smith 1986; Harrison et al.
2001).
MATERIAL AND METHODS
The material consists of four partially articulated, incom-
plete skeletons. The bones are exposed on slabs preserved
as part and counterpart. Most of the bones are broken
into innumerable fragments and, thus, little detailed ana-
tomical information could be obtained from them. Bone
was removed completely from one of the specimens
(400a, b) and moulds of silicone rubber were prepared
from the sharp impression that remained. The specimens
represent four individuals, two of which (WM400, 502)
are adults, whereas the other two (WM503, 401) corres-
pond to juvenile individuals. Descriptions and illustra-
tions were prepared with the aid of a stereomicroscope
equipped with a camera lucida. Materials of living and
extinct taxa used for comparisons are listed in the Appen-
dix.TEXT -F IG . 1 . Map of Tanzania showing the location of the
fossil site.
724 P A L A E O N T O L O G Y , V O L U M E 4 8

A cladistic analysis based on 56 equally weighted, adult
osteological characters, listed in the Appendix, was per-
formed to determine the phylogenetic position within
Pipoidea of the taxon represented by the materials des-
cribed herein. The characters and character-states used in
this analysis are listed in the Appendix; numbers in par-
entheses in the discussion of the results refer to this list.
Most of these characters are from Baez and Pugener
(2003), with the addition of several characters, including
a few (Characters 33 and 34) from the analysis of Baez
and Rage (1998). New character coding for fossil taxa
other than the one described herein is from the literature
and ⁄or personal observations (AMB), as follows: Chel-
omophrynus (Henrici 1991), Cordicephalus (pers. obs.),
Eoxenopoides (Estes 1977; pers. obs.), Llankibatrachus
(Baez and Pugener 2003), Pachycentrata (Baez and Rage
1998), Palaeobatrachus (Spinar 1972), Saltenia (Baez
1981), Shelania pascuali (Baez and Trueb 1997), Shelania
laurenti (Baez and Pugener 1998), ‘Xenopus’ romeri (Estes
1975). The data matrix is presented in the Appendix. All
multistate characters were treated as unordered except
Character 17, which was considered to be of ordered type.
The data matrix was analysed using the branch-and-
bound search of the program PAUP (Phylogenetic Analy-
sis Using Parsimony), version 4.0b.10 (Swofford 2002) for
Macintosh. Trees were rooted using Ascaphus and Discog-
lossus. The strength of each node in the tree was estima-
ted using the Bremer Decay Index, calculated with
TreeRot (Sorenson 1999).
SYSTEMATIC PALAEONTOLOGY
Institutional abbreviations. KU, Natural History Museum, Univer-
sity of Kansas, Lawrence; MCZ, Museum of Comparative Zoology,
Harvard University, Cambridge; USNM, United States National
Museum, Washington DC, all USA.
Class AMPHIBIA Linnaeus, 1758
Order ANURA, Rafinesque, 1815
Family PIPIDAE Gray, 1825
Subfamily PIPINAE Gray, 1825
Genus SINGIDELLA gen. nov.
Derivation of name. After Singida, the town in Tanzania near
where the fossil material was collected.
Type species. Singidella latecostata sp. nov.
Diagnosis. As for the only known species.
Singidella latecostata sp. nov.
Text-figures 3A–B, 4A–C, 5
Derivation of name. Latin late, wide, and costa, rib or flank, with
reference to the widely expanded transverse processes of anterior
presacral vertebrae.
Recent colluvium with poorly-sorted fragments of granite and quartz, and aggregations of decomposed kimberlite.
Shales with blocks of silicified mudstone.
Alternating series of mudstones and friable shales with variable degrees of silification.
Main fossiliferous horizons.
Massive clayey sandstone with weak horizontal bedding.
Silicified mudstone.Shales with blocks of consolidated mudstone.
compact mudstone
friable shalespartially silicified mudstonesclayey sandstone
TEXT -F IG . 2 . Geological section of the succession at Mahenge, indicating position of the frog-bearing horizons. The numbers refer
to marker horizons that comprise compact and easily identifiable mudstones in the fossil-bearing beds (after Harrison et al. 2001).
B A E Z A N D H A R R I S O N : E O C E N E F R O G F R O M T A N Z A N I A 725

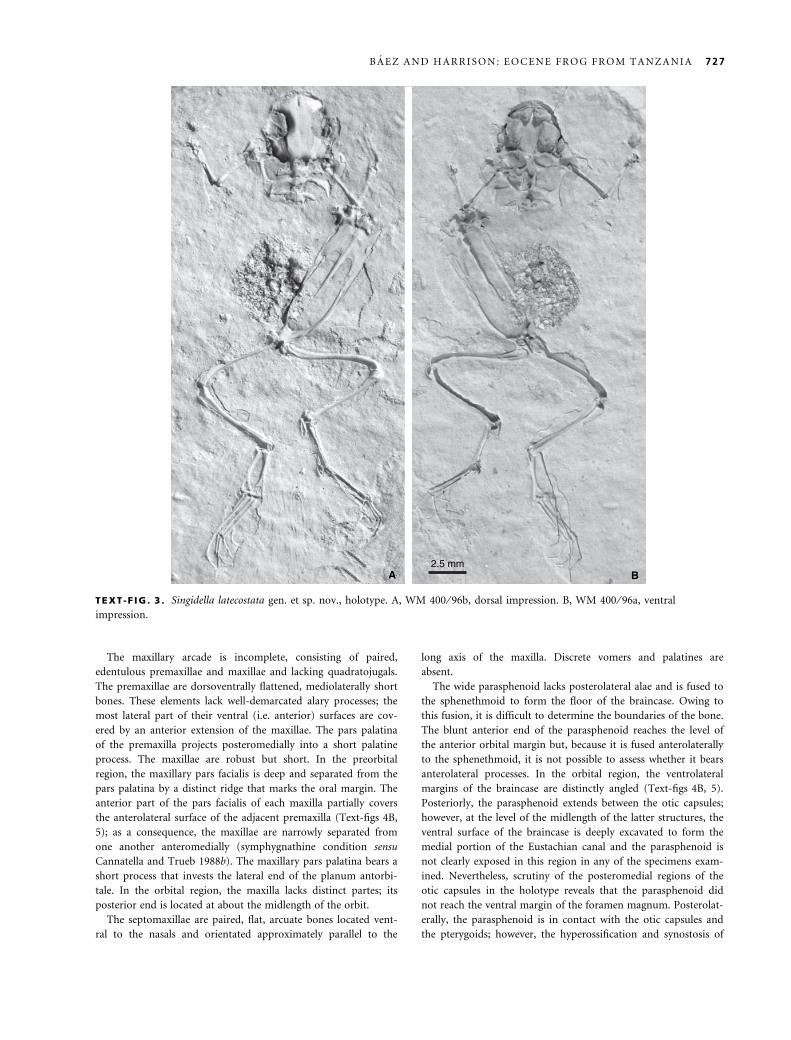
Holotype. WM 400 ⁄ 96a, b; ventral (a) and dorsal (b) impres-
sions of a partially articulated skeleton; Permanent Collection,
National Museums of Tanzania, Dar es Salaam.
Locality and horizon. Mahenge, Singida Plateau, about 53 km
west of the town of Singida, Tanzania; Level 7 of section at
Mahenge, Eocene (Harrison et al. 2001).
Referred material. Wembere-Manonga Paleontological Exped-
ition numbers 401, 502 and 503a, b.
Diagnosis. Pipine frog that resembles Hymenochirus and
Pseudhymenochirus, and differs from Pipa, in the presence
of symphygnathine maxillary arcade, extensive articulation
between parasphenoid and pterygoid, six discrete presac-
ral vertebrae, ventrally deflected articulating surfaces of
postzygapophyses, and fused clavicle and scapula. Singid-
ella differs from the hymenochirines and resembles Pachy-
centrata in having lateral expansions of the frontoparietal
that are grooved for the occipital arteries. It further differs
from the hymenochirines in having a pterygoid bearing an
anterior ramus and widely expanded transverse processes
on the anterior presacrals, and is unlike Pachycentrata in
lacking a modified occiput for the reception of the atlantal
odontoid process and a hyperossified vertebral column.
Description
The adult snout-vent length of this frog is estimated to have been
about 41–45 mm. Both the skull and the postcranial skeleton are
well ossified. However, only the frontoparietal bears ornamenta-
tion, which consists of faint, irregular ribs. The skull is about as
long as wide and distinctly wedge-shaped in lateral profile. The
nasal region is relatively short; its anteroposterior length is
between one-quarter and one-fifth the total length of the skull.
Most of the endochondral bones are intensively ossified and fused
to dermal elements; this obscures their extent and relationships.
Cranial skeleton. The nasals are extensive and roofed the nasal
capsules completely. Their anterior margins are slightly convex
and lack medial rostral processes; anteriorly, they overlap the
maxillary arch. Posterolaterally, these bones rest on the ossified
posterior walls of the olfactory capsules. Although the nasals
appear not to have been fused to one another medially (WM
400 ⁄ 96), each nasal seems to have been fused to the posteriorly
adjacent frontoparietal, as evidenced by the absence of a suture
between the nasal and frontoparietal in mature individuals.
The frontoparietal is a prominent dorsal roofing bone that lacks
a medial suture between left and right halves (Text-figs 3A, 4).
Anteriorly, the bone is synostotically fused to the nasals and to the
underlying sphenethmoid in mature individuals (e.g. WM
502 ⁄ 96); consequently, it is impossible to assess the shape of its
anterior margin, as well as the presence or absence of anterolateral
alae. In less ossified examples (WM 401, 503a), however, it is poss-
ible to discern a short, medial rostral projection that is in contact
with the posteromedial margins of the nasals. At the level of the
anterior margin of the orbits, the dorsal surface of the frontopari-
etal has a pair of shallow, but distinct, depressions that are deeply
lunate in dorsal shape; the pineal foramen is visible slightly in
front of these depressions (Text-fig. 4A). In the orbital region, the
lateral margins of the frontoparietal are relatively straight and
form narrow supraorbital flanges. At the level of the otic capsules,
the frontoparietal has lateral extensions that overlap the medial
region of the roof of each otic capsule. Posteriorly, an open groove
that probably housed the occipital artery is visible between the
dorsal table and each posterolateral extension of the frontopari-
etal; the anterior two-thirds of these grooves, however, become
bridged over by frontoparietal ossification.
The well-ossified sphenethmoid is synostotically united to sur-
rounding dermal elements. Thus, it is indistinguishably fused to
the perpendicular laminae of the frontoparietal at both sides of
the braincase, and to the parasphenoid, ventrally. Also, in the
orbital region the sphenethmoid is completely fused to the pro-
otics. Anteriorly, the sphenethmoid is continuous with the pos-
terior and lateral walls of each olfactory capsule. The
sphenethmoidal ossification extends into the medial septum that
separates the olfactory capsules and forms a bony partition up
to the tip of the snout (Text-figs 4B, 5). On each side of the sep-
tum nasi, the channel for the olfactory nerve pierces the poster-
ior wall of the olfactory capsule. Anterolaterally, the antorbital
plane is ossified up to the maxillary arcade; thus, the sphe-
nethmoidal ossification must have enclosed the orbitonasal fora-
men. Dorsally, the sphenethmoid appears not to have bounded a
dorsal fenestra anteriorly; the great anterior extent of the impres-
sions of the cerebral hemispheres on the ventral surface of pre-
served pieces of the frontoparietal (WM 500 ⁄ 96) suggests that
the sphenethmoid did not participate in the formation of the
roof of the braincase in the anterior orbital region. At about the
level of the posterior third of the orbits, a small foramen,
presumably for the exit of the optic nerve, is visible on both
lateral walls of the braincase. Posterior to this foramen,
a larger opening, possibly the prootic foramen, lies between
the orbital wall of the braincase and the anterior part of the
prootic.
The prootics and the exoccipitals are synostotically fused to
one another to form the otic capsules, but the nature of the
medial association of these latter structures is unknown. The
otic capsules are relatively narrow owing to the lack of exten-
sively ossified crista parotica. Although crests are absent on the
dorsal surface of the otic capsule, the medial region of the roof
of each capsule bears weak ornamentation similar to that of
the frontoparietal; these sculptured portions seem to corres-
pond to posterolateral extensions of the frontoparietal that
overlap and fuse to the prootics, as described above. Ventrally,
a deep groove that must have housed the Eustachian tube
extends across the ventral surface of each otic capsule (Text-
fig. 5). Posterior to the Eustachian canal, the ventral surface of
the prootic corresponding to the inner-ear regions are flat,
lacking crests along the posterior margins of the canals. More
posteriorly, the exoccipitals form the occipital condyles, which
are stalked and have articular surfaces orientated lateroventrally.
Lateral, but near, to the condyle one small foramen is visible
in the holotype (WM 400 ⁄ 96a).
726 P A L A E O N T O L O G Y , V O L U M E 4 8
The maxillary arcade is incomplete, consisting of paired,
edentulous premaxillae and maxillae and lacking quadratojugals.
The premaxillae are dorsoventrally flattened, mediolaterally short
bones. These elements lack well-demarcated alary processes; the
most lateral part of their ventral (i.e. anterior) surfaces are cov-
ered by an anterior extension of the maxillae. The pars palatina
of the premaxilla projects posteromedially into a short palatine
process. The maxillae are robust but short. In the preorbital
region, the maxillary pars facialis is deep and separated from the
pars palatina by a distinct ridge that marks the oral margin. The
anterior part of the pars facialis of each maxilla partially covers
the anterolateral surface of the adjacent premaxilla (Text-figs 4B,
5); as a consequence, the maxillae are narrowly separated from
one another anteromedially (symphygnathine condition sensu
Cannatella and Trueb 1988b). The maxillary pars palatina bears a
short process that invests the lateral end of the planum antorbi-
tale. In the orbital region, the maxilla lacks distinct partes; its
posterior end is located at about the midlength of the orbit.
The septomaxillae are paired, flat, arcuate bones located vent-
ral to the nasals and orientated approximately parallel to the
long axis of the maxilla. Discrete vomers and palatines are
absent.
The wide parasphenoid lacks posterolateral alae and is fused to
the sphenethmoid to form the floor of the braincase. Owing to
this fusion, it is difficult to determine the boundaries of the bone.
The blunt anterior end of the parasphenoid reaches the level of
the anterior orbital margin but, because it is fused anterolaterally
to the sphenethmoid, it is not possible to assess whether it bears
anterolateral processes. In the orbital region, the ventrolateral
margins of the braincase are distinctly angled (Text-figs 4B, 5).
Posteriorly, the parasphenoid extends between the otic capsules;
however, at the level of the midlength of the latter structures, the
ventral surface of the braincase is deeply excavated to form the
medial portion of the Eustachian canal and the parasphenoid is
not clearly exposed in this region in any of the specimens exam-
ined. Nevertheless, scrutiny of the posteromedial regions of the
otic capsules in the holotype reveals that the parasphenoid did
not reach the ventral margin of the foramen magnum. Posterolat-
erally, the parasphenoid is in contact with the otic capsules and
the pterygoids; however, the hyperossification and synostosis of
A2.5 mm
B
TEXT -F IG . 3 . Singidella latecostata gen. et sp. nov., holotype. A, WM 400 ⁄ 96b, dorsal impression. B, WM 400 ⁄ 96a, ventral
impression.
B A E Z A N D H A R R I S O N : E O C E N E F R O G F R O M T A N Z A N I A 727

the elements in the suspensorial region obscure their shapes and
relationships with one another. Three foramina lie on each side,
near the union of the parasphenoid-sphenethmoid complex with
the otic capsules (Text-figs 4B, 5). Two of these three foramina
are located in a short groove; the most anterior of them might
correspond to the foramen for the ramus palatinus of the facial
nerve. If this identification is correct, an osseous bridge extends
between the ventrolateral corner of the neurocranium and the
floor of the otic capsule in front of the palatine foramen. The
third foramen, the identity of which is uncertain, is located lateral
to the palatine foramen.
The squamosal is conch-shaped, probably owing to the inva-
sion of the squamosal ossification into the funnel-shaped portion
of the tympanic annulus. The poor preservation of this bone in
all specimens examined precludes a detailed description; however,
available evidence suggests that the zygomatic ramus was either
absent or greatly reduced. The plectral apparatus is represented
by the well-ossified pars media plectri. This element is a slightly
curved, rod-like bone, the proximal end of which is expanded
and probably fitted into the fenestra ovalis. At about its mid-
length, the stapes bears one short process that might have been in
contact with the posterior margin of the squamosal.
On each side of the skull, a long strut of bone extends from the
anterolateral part of the otic capsule to brace the maxillary arcade
and articulate with the dorsal surface of the maxilla at the level of
the anterior part of the orbit (WM 400, 401). These bones are
thought to be the anterior rami of the pterygoids. The medial and
lateral rami of each pterygoid seem to have formed an elongated
otic plate that floored at least the most lateral part of the Eusta-
chian canal (Text-fig. 4B); however, this plate is incomplete in all
of the specimens examined and, thus, it is not possible to deter-
mine its medial extent. Anteriorly, the otic plate of the pterygoid
is fused to the overlying otic capsule; a faint line indicates that
it has a broad contact with the parasphenoid-sphenethmoid
complex. However, the lack of scars on the venter of the otic cap-
sule suggests that the posterior margin of the otic plate remained
unfused to this latter structure. Laterally, the otic plate extends to
the ventral part of the squamosal. The ossified pars articularis of
the palatoquadrate cartilage, distinctly located at midlength of the
otic capsule, projects ventrally between this part of the squamosal
and the pterygoid.
The lower jaw is not well preserved in any of the specimens
examined. In WM 401 ⁄ 96, the shape of the longitudinal section
of the angulosplenial suggests the presence of a well-developed
coronoid process.
Hyobranchial skeleton. The only portions of this structure that
are preserved are the ossified posteromedial processes. These
rodike bones have slightly expanded posterior ends and are
about one-quarter the length of the skull.
C
4 mm
B
A
TEXT -F IG . 4 . Singidella latecostata gen. et sp. nov. A, restoration of the skeleton in dorsal view, pectoral girdle shown on right side
only, cleithrum and hyobranchial skeleton not shown. B, partial restoration of the skull in ventral view. C, restoration of pectoral
girdle in ventral view, with cleithra deflected ventrally; cartilages not shown.
728 P A L A E O N T O L O G Y , V O L U M E 4 8

Postcranial skeleton. The vertebral column is composed of six
discrete opisthocoelous presacral vertebrae, the sacrum and the
urostyle (Text-fig. 4A). Well-developed spinous processes on
the neural arches are not evident; moreover, the posterior mar-
gins of the neural arches of most vertebrae are slightly concave.
However, the dorsal exposure of the medulla in life must have
been prevented by the long anterior extension of the neural
arches between both prezygapophyses. The articular facets of
the prezygapophyses are rounded and flat, whereas those of the
postzygapophyses are curved ventrally to form a groove where
the corresponding prezygapophysis of the succeeding vertebra
fitted when the vertebrae were in articulation. Accessory articu-
lations between adjacent pedicles are visible on both sides
of the vertebral centra. The centra are extremely shallow
(Text-fig. 5), suggesting that they were probably epichordal in
development.
The first presacral vertebra has minute spinal nerve foramina
in the neural arch, anterior to the transverse processes. This
indicates that the first vertebra includes fused Vertebrae I and II.
This compound element (herein called atlas) bears short trans-
verse processes that are laterally orientated and broadly expan-
ded distally (e.g. WM 400 ⁄ 96). This marked distal expansion is
not evident in young individuals (WM 401 ⁄ 96). The anterior
margin of the atlantal neural arch is straight; this prevented the
dorsal exposure of the medulla when the vertebral column was
articulated with the skull. The atlantal neural arch is wider than
long, and, in common with the neural arches of the rest of the
presacral vertebrae, lacks well-developed neural spines and orna-
mentation. Ventrally, the anterior margin of the atlas is acumi-
nate. The atlantal cotyles are elongated and orientated
lateroventrally, but no notch occurs in the intercotylar area.
The second presacral (Vertebra III) has long, distally expan-
ded transverse processes, which are orientated posterolaterally.
The distal margins of these processes bear grooves that may
indicate that plates of cartilage extended from them. The
neural arch of this vertebra is somewhat narrower and longer
than the neural arches of the remaining presacrals. As with the
atlantal transverse processes, the transverse processes of this
vertebra are less distinctly expanded in young individuals (WM
401 ⁄ 96). The third presacral (Vertebra IV) bears laterally orien-
tated transverse processes, which are not well preserved in any
of the specimens examined. In WM 400 ⁄ 96 and 401 ⁄ 96 these
transverse processes are about as long as those on the first ver-
tebra; their irregular distal ends suggest that they are broken
off or, more likely, were terminated in cartilage. The number
of posterior presacrals seems to vary. Two additional presacral
vertebrae (V and VI) are present in the articulated column of
specimen 401 ⁄ 96. Three other vertebrae occur in specimen
400 ⁄ 96; two of these are disarticulated from the rest of the
vertebral column and the other, the last presacral, is still in
articulation with the sacrum. These posterior presacral verteb-
rae bear short, pointed transverse processes, with a marked an-
terolateral orientation.
The sacrum is fused to the urostyle and is probably formed
by more than one vertebra, as suggested by the presence of two
pairs of spinal nerve foramina on its ventral surface. The sacral
diapophyses are greatly expanded (Text-fig. 4A). The distal
expansion varies in the available specimens, being particularly
conspicuous in WM 400 ⁄ 96, which is equal to almost two-thirds
of the preacetabular ilial length. No spinous process occurs on
the sacrum, although a low, rounded ridge is present along its
midline. The urostyle is relatively short, its length being nearly
equivalent to the length of the first five presacral vertebrae. In
WM 400 ⁄ 96a, scars on the ventral surface of the sacral centra
mark the former fusion of the flat anterior head of the hypo-
chord to the sacrum. According to this evidence, the anterior
end of the hypochord reached between the eighth and ninth ver-
tebral elements.
Pectoral girdle. The clavicles are robust and deeply bowed anteri-
orly (Text-fig. 4C). The medial end of each clavicle is slightly
expanded and its lateral end is fused to the pars acromialis of
the scapula. The scapulae are bicapitate; a shallow, but distinct,
notch divides the medial end of each scapula into partes acromi-
alis and glenoidalis. The diameter of the glenoidal part of the
scapula is about one-third of the total length of the bone. Its
leading edge is slightly concave and distinctly thinner than the
rest of the shaft. Both ends of each coracoid are expanded, espe-
cially the sternal end, which forms a broad, fan-shaped plate.
This expansion of the sternal end of the coracoid equals about
three-quarters of the length of the bone in adults; the expansion
is less marked in juveniles (WM 503 ⁄ 96). The cleithrum is parti-
ally preserved in all specimens examined but, based on WM
502 ⁄ 96, it is possible to ascertain that these bones were extensive
and each had a short anterior prong separated by a small notch
from a posterior laminar portion. No indication of the cartila-
ginous parts of the pectoral girdle is evident on the examined
specimens.
Forelimb. The humerus is a slender, relatively long bone. The
deltoid crest is moderately developed and extends along the
proximal third of the diaphysis. A low accessory crest lies medial
to the deltoid crest. At the distal end, the well-ossified eminentia
capitata is clearly visible between both epicondyles. A distinct
triangular concavity (fossa cubitalis ventralis) is present anterior
to the eminentia capitata. The radioulna is flattened and expan-
ded distally; it is relatively long, its length being about three-
quarters of the length of the humerus. A large, flat bone occurs
at the humeral-radioulnar joint; this heterotopic bone corres-
ponds to the patella ulnaris on the distal anconaeus tendon
(Sokol 1969). The carpus is not well preserved in any of the
specimens. However, in the holotype it is possible to ascertain
the presence of a large carpal element at the ulnar side of the
distal end of the radioulna (WM 400 ⁄ 96a), which probably cor-
responds to the ulnare. Distal to this carpal, an elongate bone
extends to the base of Metacarpal V. This elongate element may
correspond to Distal Carpal 5, but it is unclear whether it is
fused to the ulnare. Two other carpals occur on the preaxial side
of the latter elongate bone: one at the base of Metacarpal IV and
another at the base of Metacarpals II and III. Although the iden-
tification of most of these bones remains equivocal, the presence
of a few, but large, elements in the carpus is evident. Two meta-
carpals are present on WM 502 ⁄ 96, the identities of which are
uncertain, but the faint impressions of the four metacarpals are
visible in the holotype. Metacarpal II is distinctly shorter than
the other metacarpals. The longest metacarpals are about
B A E Z A N D H A R R I S O N : E O C E N E F R O G F R O M T A N Z A N I A 729

two-thirds of the length of the radioulna. The digits are not pre-
served.
Pelvic girdle. The articulated pelvis is visible in WM 400 ⁄ 96,
401 ⁄ 96 and 502 ⁄ 96. The ilia have long shafts, which bear lat-
eral crests along their distal halves. Posteriorly, these bones
form a strong symphysis and are fused to the ischia. In the
only specimen in which the pelvis is exposed ventrally (WM
400 ⁄ 96), one unossified area, which might have remained carti-
laginous, occurs in each of the otherwise bony pubes. The
pubes formed a distinct transverse ridge on the ventral surface
of the pelvis. The putative cartilaginous areas might have been
involved in the support of a prepubis, a small plate of cartilage
synchondrotically united with the pubis (Duellman and Trueb
1986).
Hind limb. The femur is distinctly sigmoid and has a rounded,
well-developed head. Its length is about 46 per cent of the
snout-vent length. The tibiofibula is slightly shorter than the
femur; a distinct sulcus intermedius marks the former fusion
between tibia and fibula. Fine crests along the length of the tib-
iofibulae are evident, particularly at both ends. The slender tibi-
ale and fibulare are fused at their expanded anterior and
posterior ends. The length of the compound bone is about 65
per cent of the tibiofibular length. No flanges are evident on this
element. Two large bones at the joint between tibiofibula and
tarsus are visible on the hindlimbs in WM 400 ⁄ 96; these ele-
ments probably correspond to the proximal and distal ossa sesa-
moidia tarsalia (Nussbaum 1982). One distal tarsal occurs at the
base of Metatarsal III. Three large bones are present on the plan-
tar surface between the tibiofibulare and the bases of the meta-
tarsals of the left limb of the same specimen. The largest of these
elements is a rectangular, relatively flat bone that lies superfi-
cially at the base of Metatarsal III; this bone is thought to be a
sesamoid. On the postaxial side and partially underlying this ses-
amoid there is another bone. In addition, a further large bone is
located between the bases of Metatarsals V and IV. This series of
bones is interpreted as part of the heterotopic bone series that
may form in the cartilago plantares (Olson 1998).
The elongated metatarsals are almost as long as the tibiofibul-
are. Their length decreases in the following order: 3 > 5 ¼4 > 2 > 1. Metatarsal III is distinctly more robust and more
widely expanded at the proximal and distal ends than the rest of
the metatarsals. The phalangeal formula is 2-2-3-4-3. The distal
phalanges are delicate and pointed. There is no evidence that a
prehallux was present.
COMPARISONS AND PHYLOGENETICRELATIONSHIPS
Recent phylogenetic analyses of pipoid frogs, including
fossil as well as extant taxa, have corroborated the mo-
nophyly of this group (Cannatella and Trueb 1988a; Baez
and Trueb 1997; Baez and Pugener 1998, 2003; Henrici
1998). As inferred from these analyses, the following char-
acter states are pipid synapomorphies: optic foramina
bound in sphenethmoidal ossification, large, arcuate
septomaxillae, parasphenoid forming part of the braincase
pmx
snmx
pt
ps
op
Ec
3 mm co
v
h
cl
tp IIIIV
TEXT -F IG . 5 . Singidella latecostata gen. et sp. nov. Photograph of cast of WM 400 ⁄ 96a (holotype) showing details of the ventral
view of the skull and anterior part of the postcranial skeleton. co, coracoid; cl, clavicle; Ec, Eustachian canal; h, posteromedial
processes of the hyoid; mx, maxilla; op, otic plate; pmx, premaxilla; ps, parasphenoid; pt, pterygoid; sn, nasal septum; tp III,
transverse process of Vertebra III; v IV, Vertebra IV.
730 P A L A E O N T O L O G Y , V O L U M E 4 8

floor; conch-shaped squamosal, medial and lateral ramus
of the pterygoid expanded to form an otic plate, prootics
deeply excavated to house the Eustachian tubes; partes of
the maxilla not distinct in the orbital region; opisthocoe-
lous vertebral centra, fused sacrum and urostyle. Their
presence in Singidella latecostata indicates that this taxon is
a member of the Pipidae. Our parsimony analysis resulted
in three most-parsimonious trees of 104 steps, a consis-
tency index (CI) of 0Æ683, a retention index (RI) of 0Æ847
and a rescaled consistency index (RC) of 0Æ578. These trees
differ only in the interrelationships of three Palaeogene
South American xenopodinomorph species (Text-fig. 6).
Singidella is clearly a pipine; this conclusion is supported
by two unique synapomorphies for this group: wedge-
shaped skull (1) and presence of ossa sesamoidia tarsalia
(56), and, possibly, another uniquely derived condition:
parasagittal processes on neural arches (44). It also posses-
ses several pipine-derived features that occur convergently
in other taxa, such as lack of discrete vomers (22) and
reduced zygomatic process of the squamosal (28).
Within Pipinae, the presence of maxillae that extend
over the anterior parts of the premaxillae (17), an anteri-
orly blunt parasphenoid (25), and presacral vertebrae
bearing postzygapophyses whose articular surfaces are
deflected ventrally (43) indicate that Singidella is more
closely related to Hymenochirus and Pseudhymenochirus
than it is to Pipa. In addition to these features unique to
hymenochirines, Singidella shares other, presumably
derived, conditions with this group, such as reduction of
the number of presacral vertebrae to six discrete elements.
However, Singidella has the plesiomorphic state of several
hymenochirine synapomorphies and, thus, it seems to lie
outside the node that unites Pseudymenochirus and
Hymenochirus. These plesiomorphic character states are:
premaxilla not reduced and lacking a dagger-like palatine
process (15), and pterygoid bearing an anterior ramus
(30). In addition, the lack of any evidence of the large
bony ceratohyals, which ossify before the end of meta-
morphosis, strongly suggests that these elements were
either cartilaginous or absent (40). Also, the premaxillae
are only partially covered by the anterior ends of the
maxillae (17) and the relative expansion of the sternal
margin of the coracoid is slightly less than in the extant
hymenochirines.
Rhinophrynus
Chelomophrynus
Palaeobatrachus
Xenopus
Silurana
Shelania pascuali
‘Shelania’ laurenti
‘Xenopus’ romeri
Saltenia
Eoxenopoides
Pipa
Cordicephalus
Llankibatrachus
Hymenochirus
Singidella
Pachycentrata
HYMENOCHIRINI
PIPINAE
PIPOIDEA
PIPIDAE
XENOPODINOMORPHA
PIPINOMORPHA
Pseudhymenochirus
XENOPODINAE
2
Ascaphus
Discoglossus
4
1
4
7
2
4
1
2
10
1
4
1
1
1
TEXT -F IG . 6 . Strict consensus of three most-parsimonious trees (104 steps) obtained in the analysis of the data used in this study
(Appendix). Numbers correspond to decay indices (Bremer 1994).
B A E Z A N D H A R R I S O N : E O C E N E F R O G F R O M T A N Z A N I A 731

Singidella shares with Pachycentrata the frontoparietal
bearing lateral extensions that partially overlap the dorsal
surface of the otic capsules and elaborate canals for the
occipital arteries (6). In both taxa, the pterygoid otic
plates remained unfused to the otic capsules, a plesiomor-
phic condition. However, in Pachycentrata, there are
crests along the posterolateral margins of the parasphe-
noid-sphenethmoid complex and the posterior margin of
the Eustachian canal that mark the extent of the plates
(Baez and Rage 1998). In Pipa (P. carvalhoi KU 194400;
P. myersi KU 113663; P. parva USNM 115775), the pos-
terior margins of the otic plates articulate with distinct
crests on the prootics. These crests are absent in Hymeno-
chirus (H. curtipes KU 204133; H. boettgeri KU 209543),
as well as in Singidella. In Pseudhymenochirus the otic
plate is indistinguishably fused to the otic capsules poste-
riorly.
The taxonomic placement of Singidella proposed herein
(Text-fig. 6) requires the reinterpretation of the evolution
of several putative pipine synapomorphies, such as the
presence of osseous crests on the tibiale, fibulare and met-
atarsals (Baez 1981; Cannatella and Trueb 1988a). The
crests on the hind limbs may have appeared independ-
ently in hymenochirines and Pipa or its absence in Singid-
ella is a reversal.
The fused nasals and frontoparietal of Singidella also
occurs in Pseudhymenochirus, but this fusion may be
observed occasionally in mature specimens of other
pipid genera (Pipa, Xenopus) as well. As in Singidella, a
completely ossified septum is present in Pachycentrata,
Hymenochirus and Pseudhymenochirus, and primitively in
Pipa (Cannatella and Trueb 1988a); this condition was
considered a pipine synapomorphy (Cannatella and Trueb
1988b). In this regard, it is interesting to note that in
Hymenochirus curtipes the septum nasi has an independent
centre of ossification at its midlength; subsequently, this
ossification extends in an anterior direction (KU 204237,
204130) and the most posterior portion of the septum
may long remain cartilaginous. Examination of a juvenile
Pipa carvalhoi (KU 194400) in which the posterior portion
of the septum is ossified whereas the anterior half is still
cartilaginous points to a different pattern of ossification,
as suggested by Cannatella and Trueb (1988b). Similarly,
the plana anteorbitalae have independent centres of ossifi-
cation in Hymenochirus, unlike Pipa carvalhoi.
The braincase of Singidella is relatively wider than that
of living hymenochirines and differs from the braincase
of Pachycentrata in being parallel-sided, lacking the con-
striction anteriad of the otic capsules of the latter taxon.
In both Singidella and Pachycentrata, the pars articularis
of the palatoquadrate is located more posteriorly than it
is in hymenochirines, although this resemblance might be
plesiomorphic. Because the suspensorial region migrates
backward during metamorphosis, the markedly anterior
position of the quadrate in Hymenochirus was considered
as evidence of paedomorphosis (Paterson 1945, 1955). As
in other pipines, in Singidella the parasphenoid forms the
greater part of the ventral surface of the braincase, but in
contrast to the distinctly convex parasphenoid of Pachy-
batrachus this bone is flat in Singidella.
Singidella has anterior vertebrae bearing transverse pro-
cesses (presumably fused to ribs as in other pipids) that
are distally expanded in adults; in contrast, in living
hymenochirines the anterior presacrals bear long, but unex-
panded, processes. In this regard, the vertebral column of
Singidella resembles that of Pipa in the presence of expan-
ded transverse processes on the first presacral vertebra.
Expanded processes on the following two vertebrae occur
in Pipa myersi and P. parva, the more basal species of Pipa,
according to a well-corroborated phylogeny of the species
of this genus (Trueb and Cannatella 1986). The shortness
of the transverse processes of the third discrete vertebra
(Vertebra IV) in the examined specimens of Singidella is
puzzling because the transverse processes of this vertebra
extend beyond the lateral margins of the sacral diapophyses
in pipids. Moreover, the skeleton, especially that of the
holotype, is rather well preserved (Text-fig. 3). In addition,
there is no clear indication that the preceding presacral
includes fused Vertebrae III and IV and, thus, that the third
vertebra is actually Vertebra V. Therefore, we tentatively
assume that the distal ends of the transverse processes of
Vertebra IV might have extended in cartilage.
The fusion of the sacral vertebra and the urostyle is a
derived feature that characterizes all pipids (Cannatella and
Trueb 1988a; Baez and Trueb 1997), although this condi-
tion evolved independently several times within Anura. The
fused joint, however, arises through ontogenetic rearrange-
ment of the same elements that are present in taxa with the
more widespread bicondylar articulation between sacrum
and urostyle (Wiens 1989). Seemingly, in Singidella the
anterior end of the hypochord fused to Vertebra IX, as it
does in Xenopus laevis and Pipa carvalhoi (pers. obs.). How-
ever, in Singidella, this condition persisted despite the
incorporation of at least one additional posterior presacral
vertebra (Vertebra VIII) into the sacrum, a condition that
does not occur in the other two taxa. This shortening of the
presacral part of the vertebral column also occurs in living
hymenochirines although no data are available to us
regarding the position of the hypochord and the formation
of the sacro-urostylar joint in this group.
The possible presence of an epipubis in Singidella
would lend support to the hypothesis that absence of this
element in living Hymenochirus and Pipa results from
independent losses (Sokol 1977; Cannatella and Trueb
1988b).
Recently, Olson (1998) found an unusually high num-
ber of heterotopic bones in the appendicular skeleton of
Hymenochirus boettgeri, most of them having a consistent
732 P A L A E O N T O L O G Y , V O L U M E 4 8

morphology and location. Several of these elements, such
as the patella ulnaris (Sokol 1969) at the elbow joint, and
the posterior lunula and fabella, both at the knee joint,
were considered autapomorphies of Hymenochirus (Olson
1998). The presence of the patella ulnaris, however, was
also documented in Pipa pipa by Trueb et al. (2000) and
we observed this bone and the distal os sesamoides tarsale
in P. carvalhoi, the only species of Pipa available to us to
assess these traits. In addition, our examination of Pseud-
hymenochirus revealed that most of these elements, such
as the patella ulnaris, posterior lunula, fabella and prox-
imal os sesamoides tarsale, are not unique to Hymenochi-
rus but occur in Pseudhymenochirus as well. In addition,
we ascertained the presence of at least the patella ulnaris,
and proximal and distal ossa sesamoidia tarsalia in Sin-
gidella. This evidence suggests that at least the patella uln-
aris and distal os sesamoides tarsale characterize a more
inclusive set of taxa (Pipinae?).
DISCUSSION
The closest extant relatives of Singidella are the hymeno-
chirines, which are currently found in rivers, pools and
swamps of sub-Saharan Africa, with some species restric-
ted to forested regions and others to more open environ-
ments (Noble 1924). Living hymenochirines are
characterized by a number of distinctive features, many of
which are related with hyperossification, such as ossifica-
tion of the antorbital plane and ceratohyals, and fusion of
the parasphenoid, squamosal and pterygoid to the brain-
case (Cannatella and Trueb 1988b). Some of these fea-
tures are already present in Singidella, the size of which
falls within the range of living hymenochirines. However,
the new taxon partially fills the morphological gap that
exists between hymenochirines and the basal members of
the Pipa clade.
Extant hymenochirines, fully aquatic as other pipids,
are the only anurans that evolved suction-feeding in
adults as an adaptation to feeding under water (Sokol
1969; O’Reilly et al. 2002). Rapid pressure reduction in
the buccopharyngeal cavity by protraction of the hyola-
ryngeal apparatus and flattening of the mouth’s floor in
these frogs causes a flow of water that draws the prey to
the mouth (Sokol 1969). The probable different configu-
rations of the hyobranchial apparatus and suspensorial
region, with the more posterior position of the jaw arti-
culation and presence of the anterior ramus of the ptery-
goid, with respect to its living relatives suggest that this
mechanism of prey capture might have not been present
in adult Singidella
The occurrence of Singidella in eastern Africa implies
that in the Palaeogene, African pipines had a distribution
different from that of their extant representatives. Living
hymenochirines are confined to lowland habitats below
800 m in West Africa and the Zaire Basin, but are not
recorded east of the Eastern Rift escarpments (Frost 1985;
Poynton 1999). Moreover, they are endemic to the West
Equatorial Biogeographical Region, covered mostly by
rainforest and having marked seasonal rainfall (Poynton
1999). At present, the Central African Highlands consti-
tute an important watershed, separating the lowland for-
ests of the Zaire Basin from the savannas of eastern
Africa. Thus, these highlands represent a physiographical
feature of biogeographical significance, especially for
freshwater animals. However, in Palaeogene times most of
the eastern region was drained towards the west by rivers
that could escape to the Zaire (Congo) system (Grove
1986). Thus, pipines may have been more widely distri-
buted across equatorial Africa prior to the change of
topography and drainage pattern caused by rifting in the
Miocene. Despite its low palaeolatitude (c. 15�S), the cra-
ter lake at Mahenge was located in a warm region of
markedly seasonal or relatively low mean annual precipi-
tation and surrounded by an open plant community
(Harrison et al. 2001). These environmental conditions
are not unlike those that prevail in the regions where
some of the South American pipines now live (e.g. Rivero
1961). Some evidence of seasonal rainfall also occurs at
the site from which remains of the Late Cretaceous
hyperossified Pachycentrata were collected (Baez and Rage
1998). However, little is known of how particular climatic
factors and vegetation types affect amphibian distribution,
including the availability of appropriate sites during the
breeding season (Noble 1924; Poynton 1999).
Acknowledgements. We thank Charles Msuya for co-directing
field operations at Mahenge, Bill Sanders for casting and photo-
graphy, John Van Couvering for help with the sedimentology and
stratigraphy, and the staff of the National Museums of Tanzania
for curatorial and logistical assistance. We extend our thanks to
Linda Trueb, University of Kansas, for the loan of specimens for
comparisons and comments on the manuscript. Permission to
conduct research in Tanzania was granted by the Tanzania Com-
mission for Science and Technology, the Unit of Antiquities, and
the Regional and District Administrative Offices in Singida. This
research was supported by grants (to TH) from the National
Geographic Society, the Boise Fund of Oxford University, and the
New York University Research Challenge Fund and (to AMB)
UBACyT TX 02 from the University of Buenos Aires
REFERENCES
B A E Z , A. M. 1981. Redescription and relationships of Saltenia
ibanezi, a late Cretaceous pipid frog from northwestern Argen-
tina. Ameghiniana, 18, 127–154.
—— 1996. The fossil record of the Pipidae. 329–347. In T I N S -
L E Y , R. C. and KO B E L , H. R. (eds). The biology of Xenopus.
B A E Z A N D H A R R I S O N : E O C E N E F R O G F R O M T A N Z A N I A 733

Symposia of the Zoological Society of London, 68. Clarendon
Press, Oxford, 440 pp.
—— 1999. An Eocene pipine frog from north central Tanzania.
Journal of Vertebrate Paleontology, 19 (Supplement to No. 3),
28A.
—— and P UG E N E R , A. L. 1998. A new Paleogene pipid frog
from northwestern Patagonia. Journal of Vertebrate Paleontology,
18, 511–524.
—— —— 2003. Ontogeny of a new Palaeogene pipid frog from
southern South America and xenopodinomorph evolution.
Zoological Journal of the Linnean Society, 139, 439–476.
—— and RA GE , J.-C. 1998. Pipid frogs from the Upper Creta-
ceous of In Beceten, Niger. Palaeontology, 41, 669–691.
—— —— 2004. Pachycentrata, a replacement name for Pachy-
batrachus taqueti (Amphibia, Anura). Ameghiniana, 41, 346.
—— and TR U E B , L. 1997. Redescription of the Paleogene
Shelania pascuali from Patagonia and its bearing on the rela-
tionships of fossil and Recent pipoid frogs. Natural History
Museum, the University of Kansas, Scientific Paper, 4, 1–41.
—— —— and CA L V O , J. O. 2000. The earliest known pipoid
frog from South America: a new genus from the middle Cre-
taceous of Argentina. Journal of Vertebrate Paleontology, 20,
490–500.
B E A DL E , L. C. 1981. The inland waters of tropical Africa. An
introduction to tropical limnology. Second edition. Longman,
New York, 365 pp.
B R E M E R , K. 1994. Branch support and tree stability. Cladis-
tics, 10, 295–304.
C A N N A T E L L A , D. C. and T R UE B , L. 1988a. Evolution of
pipoid frogs: intergeneric relationships of the aquatic frog fam-
ily Pipidae. Zoological Journal of the Linnean Society, 94, 1–38.
—— —— 1988b. Evolution of pipoid frogs: morphology and
phylogenetic relationships of Pseudhymenochirus. Journal of
Herpetology, 22, 439–456.
D A W S O N , J. B. 1970. The structural setting of African kim-
berlite magmatism. 321–335. In C L I F F O R D , T. N. and
G A S S , I. G. (eds). African magmatism and tectonics. Hafner,
Darien, Connecticut, 319 pp.
D UE L L M A N , E. and T R U E B, L. 1986. Biology of amphibians.
McGraw Hill, New York, 670 pp.
E D W A R DS , C. B. and H O W K I N S , J. B. 1966. Kimberlites in
Tanganyika with special reference to the Mwadui occurrence.
Economic Geology, 6, 537–554.
E S T E S , R. 1975. Fossil Xenopus from the Paleocene of South
America and the zoogeography of pipid frogs. Herpetologica,
31, 263–278.
—— 1977. Relationships of the South African fossil frog Eoxe-
nopoides reuningi (Anura, Pipidae). Annals of the South African
Museum, 73, 49–80.
F O R D, L. S. and C A N N A T E L L A , D. C. 1993. The major
clades of frogs. Herpetological Monographs, 7, 94–117.
F R O S T , D. (ed.). 1985. Amphibian species of the world. Associ-
ation of Systematic Collections and Allen Press, Lawrence,
Kansas, 732 pp.
G A R C I A P A R I S , M., B UC H HO L Z , D. R. and PA R R A
OL E A , G. 2003. Phylogenetic relationships of Pelobatoidea
re-examined using mtDNA. Molecular Phylogenetics and Evolu-
tion, 28, 12–23.
G R A Y , J. E. 1825. A synopsis of the genera of reptiles and
amphibians, with a description of some new species. Annals of
Philosophy, 10, 193–217.
G R E E N W O OD , P. H. 1960. Fossil denticipitid fishes from
East Africa. Bulletin of the British Museum (Natural History),
Geology, 5, 1–11.
—— and PA T T E R S O N , C. 1967. A fossil osteoglossoid fish
from Tanzania (E. Africa). Journal of the Linnean Society
(Zoology), 47, 211–223.
G R O V E , A. T. 1986. Geomorphology of the African Rift System.
9–16. In FR O S T I C K , L. E., R E N A UT , R. W., R E I D, I. and
T I E R C E L I N , J.-J. (eds). Sedimentation in the African rifts.
Geological Society, London, Special Publication, 25, 382 pp.
G UN N E L L , G. F., J A CO B S , B. F., H E R E N D E E N , P. S.,
HE A D, J. J., KO W A L S K I , E., M S U Y A , C. P.,
M I Z A M B W A , F. A., H A R R I S O N , T., H A B E R S E T Z E R ,
J. and S T OR C H , G. 2003. Oldest placental mammal from
sub-Saharan Africa: Eocene microbat from Tanzania – evi-
dence for early evolution of sophisticated echolocation. Palae-
ontologia Electronica, 5, 1–10.
H A A S , A. 2003. Phylogeny of frogs as inferred from primarily
larval characters (Amphibia: Anura). Cladistics, 19, 23–89.
H A R R I S O N , T., M S UY A , C. P., M UR R A Y , A. M.,
J A C OB S , B. F., B A E Z , A. M., M UN D I L , R. and
L UD W I G , K. R. 2001. Paleontological investigations at the
Eocene locality of Mahenge in north-central Tanzania, East
Africa. 39–74. In G U N N E L L , G. F. (ed.). Eocene biodiversity:
unusual occurrences and rarely sampled habitats. Kluwer Aca-
demic ⁄ Plenum Publishers, New York, 442 pp.
H A W T H OR N E , J. B. 1975. Model of a kimberlite pipe. Phys-
ics and Chemistry of the Earth, 9, 1–15.
H E N R I C I , A. 1991. Chelomophrynus bayi (Amphibia, Anura,
Rhinophrynidae), a new genus and species from the middle
Eocene of Wyoming: ontogeny and relationships. Annals of
the Carnegie Museum, 60, 97–144.
—— 1998. A new pipoid anuran from the late Jurassic Morrison
Formation at Dinosaur National Monument, Utah. Journal of
Vertebrate Paleontology, 18, 321–332.
H E R E N DE E N , P. S. and J A C OB S , B. F. 2000. Fossil legumes
from the middle Eocene (46.0 Ma) Mahenge flora of Singida,
Tanzania. American Journal of Botany, 8, 1358–1366.
L I N N A E US , C. 1758. Systema Naturae. Tenth edition. L. Salv-
ii, Stockholm, 824 pp.
M A N N A R D, G. W. 1962. The geology of the Singida Kimber-
lite Pipes, Tanganyika. Unpublished PhD thesis, McGill
University, Montreal, 348 pp.
M I T CH E L L , R. H. 1986. Kimberlite: mineralogy, geochemistry,
and petrology. Plenum Press, New York, 442 pp.
M UR R A Y , A. M. 2000. Eocene cichlid fishes from Tanzania,
East Africa. Journal of Vertebrate Paleontology, 20, 65–664.
—— 2001. The oldest fossil cichlids (Teleost: Perciformes): indi-
cation of a 45 million-year-old species flock. Proceedings of the
Royal Society of London B, 268, 679–684.
—— 2003a. A new characiform fish (Teleosti: Ostariophysi)
from the Eocene of Tanzania. Canadian Journal of Earth Sci-
ences, 40, 473–481.
734 P A L A E O N T O L O G Y , V O L U M E 4 8

—— 2003b. A new Eocene citharinoid fish (Ostariophysi: Char-
aciformes) from Tanzania. Journal of Vertebrate Paleontology,
23, 501–507.
—— and BU DN E Y , L. A. 2003. A new species of catfish (Cla-
roteidae, Chrysichthys) from an Eocene crater-lake in East
Africa. Canadian Journal of Earth Sciences, 40, 983–993.
N O B L E , G. K. 1924. Herpetology of the Belgian Congo. Bul-
letin of the American Museum of Natural History, 49, 147–347.
—— 1954. The biology of the Amphibia. Second edition. Dover
Publications, New York, 577 pp.
N U S S B A UM , R. A. 1982. Heterotopic bones in the hindlimbs
of frogs in the families Pipidae, Ranidae, and Sooglossidae.
Herpetologica, 38, 312–320.
O L S O N , W. M. 1998. Evolutionary and developmental mor-
phology of the dwarf African clawed frog Hymenochirus Boett-
geri (Amphibia: Anura: Pipidae). Unpublished PhD thesis,
University of California, Berkeley, 155 pp.
O ’ R E I L L Y , J. C., DE B A N , S. M. and N I S H I K A W A , K. C.
2002. Derived life history characteristics constrain the evolu-
tion of aquatic feeding behavior in adult amphibians. 153–
190. In A E R T S , P., D ’ A O U T , K. and VAN D A M M E , R.
(eds). Topics in functional and ecological vertebrate morphology.
Shaker Publishing, Maastricht, 359 pp.
P A T E R S O N , N. F. 1945. The skull of Hymenochirus curtipes.
Proceedings of the Zoological Society of London, 115, 327–
354.
—— 1955. The skull of the toad of Hemipipa carvalhoi. Proceed-
ings of the Zoological Society of London, 125, 223–252.
P OY N T ON , J. C. 1999. Distribution of amphibians in Sub-
Saharan Africa, Madagascar, and Seychelles. 483–539. In
DU E L L M A N , W. E. (ed.). Patterns of distribution of amphib-
ians. A global perspective. The Johns Hopkins University Press,
Baltimore, 597 pp.
P UG E N E R , L. A., M A G L I A , A. M. and T R UE B , L. 2003.
Revisiting the contribution of larval characters to an analysis
of phylogenetic relationships of basal anurans. Zoological Jour-
nal of the Linnean Society, 139, 129–155.
R A F I N E S QU E , C. S. 1815. Analyse de la nature ou tableau de
l’univers et des corps organises. J. Barravecchia, Palermo, 224
pp.
R I V E R O, J. A. 1961. Salientia of Venezuela. Bulletin of the
Museum of Comparative Zoology, 126, 1–207.
S M I T H , R. M. H. 1986. Sedimentation and palaeoenviron-
ments of Late Cretaceous crater-lake deposits in Bushmanland,
South Africa. Sedimentology, 33, 369–386.
S O K OL , O. M. 1969. Feeding in the pipid frog Hymenochirus
boettgeri (Tornier). Herpetologica, 25, 9–24.
—— 1977. The free swimming Pipa larvae, with a review of
pipid phylogeny (Anura: Pipidae). Journal of Morphology, 154,
357–183.
S O R E N S O N , M. D. 1999. TreeRot. Version 2. University of
Michigan, Ann Arbor.
S P I N A R , Z. 1972. Tertiary frogs from Central Europe. Dr W.
Junk, The Hague, 286 pp.
S W OF F OR D , D. L. 2002. PAUP–Phylogenetic Analysis Using
Parsimony. Version 4.0b10 (Apple Macintosh Compatible).
Sinauer Associates, Sunderland, Massachusetts.
T R U E B, L. 1996. Historical constraints and morphological nov-
elties in the evolution of the skeletal system of pipid frogs.
349–376. In T I N S L E Y , R. C. and KO B E L , H. R. (eds). The
biology of Xenopus. Symposia of the Zoological Society of
London, 68. Clarendon Press, Oxford, 440 pp.
—— and CA N N A T E L L A , D. C. 1986. Systematics, morphol-
ogy, and phylogeny of the genus Pipa (Anura: Pipidae).
Herpetologica, 42, 412–449.
—— P U G E N E R , L. A. and M A GL I A , A. M. 2000. Ontogeny
of the bizarre: an osteological description of Pipa pipa, with
an account of skeletal development in the species. Journal of
Morphology, 243, 75–104.
W I E N S , J. J. 1989. Ontogeny of the skeleton of Spea bombifrons
(Anura: Pelobatidae). Journal of Morphology, 202, 29–51.
W I L L I A M S , G. J. 1939. The Kimberlite province and associ-
ated diamond deposits of Tanganyika Territory. Bulletin of the
Geological Survey of Tanganyika, 12, 1–38.
APPENDIX
Material used for comparisons
Hymenochirus boettgeri Tornier, KU 209543, male
Hymenochirus curtipes Noble, KU 204133, female; KU 204135,
female, s-v l: 33Æ4 mm; KU 204127, female, s-v l: 32Æ9 mm; KU
204126, female, s-v l: 32Æ5 mm; KU 204131, female, s-v l:
24Æ8 mm; KU 204132, male, s-v l: 24Æ6 mm; KU 204130, female,
21Æ5 mm; KU 204134, s-v l: 16Æ5. mm; KU 204137, s-v l:
18Æ7 mm.
Pipa arrabali Izecksohn, KU 167437, female
Pipa carvalhoi (Miranda Ribeiro), KU 128761, female, s-v l:
55Æ5 mm; KU 194400, female, s-v l: 29Æ75 mm; KU 92370,
female
Pipa myersi Trueb, KU 113663, female
Pipa parva Ruthven and Gage, USNM 115775, female, s-v l:
35Æ0 mm
Pipa pipa (Linneo), KU 288854, s-v l: c. 156Æ0 mm
Pipa snethlageae Muller, MCZ 85572, no data, sv: no data, MCZ
85573, no data, sv: no data.
Pseudhymenochirus merlini Chabanaud, KU 206875, male, s-v l:
29Æ9 mm.
Character list
The 55 characters and their character states of the adult osteol-
ogy used in the parsimony analysis are listed below, followed by
the data matrix. 1–36, cranial characters; 37–40, characters of
the hyobranchial skeleton; 41–56, postcranial characters.
1. skull shape in lateral profile: 0, rounded; 1, wedge-shaped.
2. nasal region: 0, one-third, or more, of the skull length; 1,
one-quarter, or less, of the skull length.
3. nasals: 0, paired; 1, fused.
B A E Z A N D H A R R I S O N : E O C E N E F R O G F R O M T A N Z A N I A 735

4. frontoparietals ⁄ nasals: 0, not overlapping; 1, overlapping.
5. frontoparietals: 0, paired; 1, fused.
6. frontoparietal posterolateral extensions: 0, absent; 1, present.
7. anterior margin of the frontoparietal fenestra: 0, sphe-
nethmoidal ossification surrounds the frontoparietal fenestra
anteriorly; 1, sphenethmoidal ossification does not surround
the fenestra anteriorly.
8. olfactory foramina: 0, bound completely or partially in bone;
1, bound in cartilage.
9. antorbital plane of the skull: 0, partially or completely carti-
laginous; 1, ossified up to the maxillary arcade.
10. margin of the optic foramina: 0, not completely bound in
sphenethmoidal ossification; 1, bound in sphenethmoidal
ossification.
11. floor of the braincase in the orbital region: 0, rounded; 1,
distinctly angled.
12. Eustachian canal: 0, absent; 1, present, prootic with deep
furrow; 2, present, prootic with shallow anterior depression.
13. inferior perilymphatic foramina: 0, present, not ventral to
jugular foramina; 1, present, ventral to jugular foramina; 2,
absent.
14. superior perilymphatic foramina: 0, present; 1, absent.
15. premaxillary palatine process: 0: not dagger-like; 1: dagger-
like
16. anterior end of the maxilla: 0, lacking pointed process that
overlaps premaxilla; 1, having pointed process that overlaps
premaxilla.
17. maxillary pars facialis: 0, not extending over premaxilla an-
teriorly; 1, partially extending over premaxilla anteriorly; 2,
nearly covering premaxilla anteriorly.
18. maxillary antorbital process: 0, absent; 1, present
19. partes of the maxilla in the orbital region: 0, distinct; 1, not
distinct.
20. maxillary arcade: 0, complete; 1, incomplete.
21. septomaxillae: 0, small and complex; 1, large and arcuate.
22. vomers: 0, medial to the choanae; 1, posterior to the cho-
anae; 2, absent.
23. parasphenoid ⁄ braincase: 0, not fused; 1, partially or com-
pletely fused.
24. alae of the parasphenoid: 0, present; 1, absent.
25. anterior extent of the parasphenoid: 0, not reaching maxil-
lary arcade; 1, reaching maxillary arcade.
26. posteromedial extent of the parasphenoid: 0, extending near
the ventral margin of the foramen magnum; 1, ending well
anterior to the ventral margin of the foramen magnum.
27. squamosal shape: 0, T-shaped without stapedial process; 1,
T-shaped with stapedial process; 2, conch-shaped.
28. zygomatic ramus of the squamosal: 0, well developed; 1,
reduced or absent.
29. pterygoid knob: 0, absent; 1, present.
30. position of the anterior ramus of the pterygoid: 0, medial to
maxilla; 1, abuts maxilla; 2, dorsal to maxilla; 3, anterior
ramus absent.
31. medial ramus of the pterygoid: 0, present, lacking indenta-
tion; 1, present, with indentation; 2, absent.
32. configuration of the pterygoid in the otic region: 0, not
expanded; 1, expanded to form an otic plate.
33. pterygoid medial ramus-parasphenoid contact: 0, limited or
no contact; 1, extensive contact.
34. fusion of pterygoid and otic capsule: 0, not fused; 1, fused
35. coronoid process of the angulosplenial: 0, poorly developed;
1, blade-like.
36. relative position of the lower jaw articulation: 0, lateral to
the otic capsule; 1, at the anterior margin of the otic cap-
sule.
37. length of the posteromedial process of the hyoid: 0, length
less than half the anteroposterior length of the lower jaw; 1,
length more than half the anteroposterior length of the
lower jaw.
38. anterior end of the posteromedial process of the hyoid: 0,
wider than posterior end; 1, narrower than posterior end.
39. parahyoid bone: 0, present; 1, absent.
40. ceratohyal: 0, not ossified; 1, ossified.
41. shape of the vertebral centra: 0, cylindrical; 1, depressed.
42. articulation facets of the vertebral centra: 0, notochordal; 1,
opisthocoelous; 2, procoelous.
43. articulation facets of the postzygapophyses: 0, flat; 1, with
grooves and ridges; 2, curved ventrally.
44. neural spines of the presacral vertebrae: 0, sagittal; 1, para-
sagittal.
45. Vertebrae I and II: 0, separate, weak or no imbrication; 1,
separate, broad imbrication; 2, fused.
46. ribs: 0, free ribs present in larvae and adults; 1, free ribs pre-
sent in larvae and fused to transverse processes in adults; 2,
ribs absent in larvae and adults.
47. sacrum and urostyle: 0, not fused; 1, fused.
48. clavicle ⁄ scapula: 0, lateral end contacts medial edge of pars
acromialis; 1, lateral end overlaps anterior edge of scapula;
2, lateral end is fused to scapula.
49. medial end of the clavicle: 0, not expanded; 1, expanded.
50. proportions of the scapula: 0, glenoid area one-third of the
total length of the scapula; 1, glenoid area more than one-
third of the total length of the scapula.
51. cleithrum: 0, not covering the posterior edge of the supra-
scapular cartilage; 1, covering part of the posterior edge of
the suprascapular cartilage.
52. sternal expansion of the coracoid relative to its length: 0,
sternal expansion less than half the length of the coracoid; 1,
sternal expansion nearly half the length of the coracoid; 2,
sternal expansion nearly the length of the coracoid.
53. ilial supra-acetabular expansion: 0, present; 1, absent.
54. ilial preacetabular expansion: 0, present; 1, absent.
55. pubis: 0, cartilaginous; 1, ossified.
56. distal os sesamoides tarsale: 0, absent; 1, present.
736 P A L A E O N T O L O G Y , V O L U M E 4 8
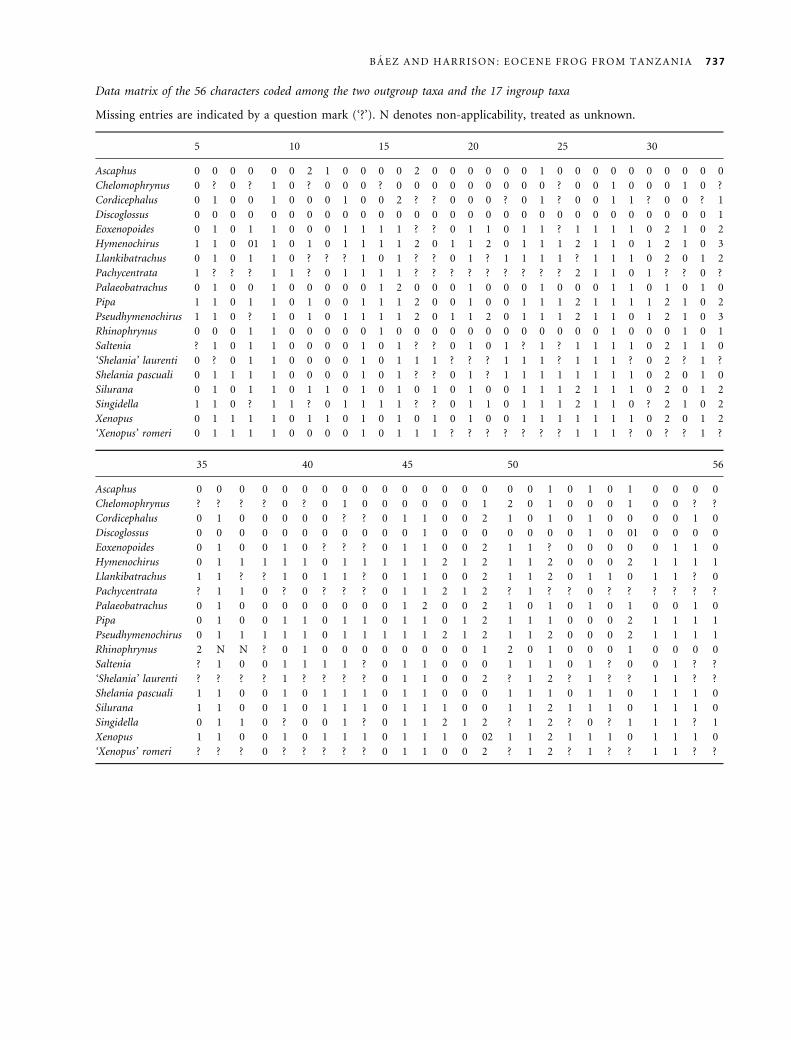
Data matrix of the 56 characters coded among the two outgroup taxa and the 17 ingroup taxa
Missing entries are indicated by a question mark (‘?’). N denotes non-applicability, treated as unknown.
5 10 15 20 25 30
Ascaphus 0 0 0 0 0 0 2 1 0 0 0 0 2 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0
Chelomophrynus 0 ? 0 ? 1 0 ? 0 0 0 ? 0 0 0 0 0 0 0 0 0 ? 0 0 1 0 0 0 1 0 ?
Cordicephalus 0 1 0 0 1 0 0 0 1 0 0 2 ? ? 0 0 0 ? 0 1 ? 0 0 1 1 ? 0 0 ? 1
Discoglossus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1
Eoxenopoides 0 1 0 1 1 0 0 0 1 1 1 1 ? ? 0 1 1 0 1 1 ? 1 1 1 1 0 2 1 0 2
Hymenochirus 1 1 0 01 1 0 1 0 1 1 1 1 2 0 1 1 2 0 1 1 1 2 1 1 0 1 2 1 0 3
Llankibatrachus 0 1 0 1 1 0 ? ? ? 1 0 1 ? ? 0 1 ? 1 1 1 1 ? 1 1 1 0 2 0 1 2
Pachycentrata 1 ? ? ? 1 1 ? 0 1 1 1 1 ? ? ? ? ? ? ? ? ? 2 1 1 0 1 ? ? 0 ?
Palaeobatrachus 0 1 0 0 1 0 0 0 0 0 1 2 0 0 0 1 0 0 0 1 0 0 0 1 1 0 1 0 1 0
Pipa 1 1 0 1 1 0 1 0 0 1 1 1 2 0 0 1 0 0 1 1 1 2 1 1 1 1 2 1 0 2
Pseudhymenochirus 1 1 0 ? 1 0 1 0 1 1 1 1 2 0 1 1 2 0 1 1 1 2 1 1 0 1 2 1 0 3
Rhinophrynus 0 0 0 1 1 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 1 0 1
Saltenia ? 1 0 1 1 0 0 0 0 1 0 1 ? ? 0 1 0 1 ? 1 ? 1 1 1 1 0 2 1 1 0
‘Shelania’ laurenti 0 ? 0 1 1 0 0 0 0 1 0 1 1 1 ? ? ? 1 1 1 ? 1 1 1 ? 0 2 ? 1 ?
Shelania pascuali 0 1 1 1 1 0 0 0 0 1 0 1 ? ? 0 1 ? 1 1 1 1 1 1 1 1 0 2 0 1 0
Silurana 0 1 0 1 1 0 1 1 0 1 0 1 0 1 0 1 0 0 1 1 1 2 1 1 1 0 2 0 1 2
Singidella 1 1 0 ? 1 1 ? 0 1 1 1 1 ? ? 0 1 1 0 1 1 1 2 1 1 0 ? 2 1 0 2
Xenopus 0 1 1 1 1 0 1 1 0 1 0 1 0 1 0 1 0 0 1 1 1 1 1 1 1 0 2 0 1 2
‘Xenopus’ romeri 0 1 1 1 1 0 0 0 0 1 0 1 1 1 ? ? ? ? ? ? ? 1 1 1 ? 0 ? ? 1 ?
35 40 45 50 56
Ascaphus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 1 0 0 0 0
Chelomophrynus ? ? ? ? 0 ? 0 1 0 0 0 0 0 0 1 2 0 1 0 0 0 1 0 0 ? ?
Cordicephalus 0 1 0 0 0 0 0 ? ? 0 1 1 0 0 2 1 0 1 0 1 0 0 0 0 1 0
Discoglossus 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 1 0 01 0 0 0 0
Eoxenopoides 0 1 0 0 1 0 ? ? ? 0 1 1 0 0 2 1 1 ? 0 0 0 0 0 1 1 0
Hymenochirus 0 1 1 1 1 1 0 1 1 1 1 1 2 1 2 1 1 2 0 0 0 2 1 1 1 1
Llankibatrachus 1 1 ? ? 1 0 1 1 ? 0 1 1 0 0 2 1 1 2 0 1 1 0 1 1 ? 0
Pachycentrata ? 1 1 0 ? 0 ? ? ? 0 1 1 2 1 2 ? 1 ? ? 0 ? ? ? ? ? ?
Palaeobatrachus 0 1 0 0 0 0 0 0 0 0 1 2 0 0 2 1 0 1 0 1 0 1 0 0 1 0
Pipa 0 1 0 0 1 1 0 1 1 0 1 1 0 1 2 1 1 1 0 0 0 2 1 1 1 1
Pseudhymenochirus 0 1 1 1 1 1 0 1 1 1 1 1 2 1 2 1 1 2 0 0 0 2 1 1 1 1
Rhinophrynus 2 N N ? 0 1 0 0 0 0 0 0 0 0 1 2 0 1 0 0 0 1 0 0 0 0
Saltenia ? 1 0 0 1 1 1 1 ? 0 1 1 0 0 0 1 1 1 0 1 ? 0 0 1 ? ?
‘Shelania’ laurenti ? ? ? ? 1 ? ? ? ? 0 1 1 0 0 2 ? 1 2 ? 1 ? ? 1 1 ? ?
Shelania pascuali 1 1 0 0 1 0 1 1 1 0 1 1 0 0 0 1 1 1 0 1 1 0 1 1 1 0
Silurana 1 1 0 0 1 0 1 1 1 0 1 1 1 0 0 1 1 2 1 1 1 0 1 1 1 0
Singidella 0 1 1 0 ? 0 0 1 ? 0 1 1 2 1 2 ? 1 2 ? 0 ? 1 1 1 ? 1
Xenopus 1 1 0 0 1 0 1 1 1 0 1 1 1 0 02 1 1 2 1 1 1 0 1 1 1 0
‘Xenopus’ romeri ? ? ? 0 ? ? ? ? ? 0 1 1 0 0 2 ? 1 2 ? 1 ? ? 1 1 ? ?
B A E Z A N D H A R R I S O N : E O C E N E F R O G F R O M T A N Z A N I A 737





























