Embed Size (px)
Citation preview

Ž .Matrix Biology 18 1999 343]355
b -Integrin]collagen interaction reduces chondrocyte1
apoptosis
Liu Cao, Vivian Lee, Mark E. Adams1, Chris Kiani, Yaou Zhang, Wendy Hu,Burton B. Yang2,U
Sunnybrook and Women’s College Health Science Centre, and Department of Laboratory Medicine and Pathobiology, Uni ersity ofToronto, Toronto, M4N 3M5 Canada
Received 31 July 1998; accepted 24 March 1999
Abstract
We have observed that the spent culture media in suspended chondrocyte cultures is essential for the survival of the cells,Ž .since complete change of the spent media induces severe programmed cell death apoptosis . Moreover, we showed that
Ž .extracellular matrix ECM molecules in the culture media provide vital chondrocyte]matrix interactions; when media arechanged, cells are deprived of matrix molecules and undergo apoptosis. In this paper we report that interaction with collagen,a ubiquitous extracellular matrix molecule, is essential for chondrocyte survival. Such an interaction causes chondrocyteaggregation and reduces the level of chondrocyte apoptosis. Hyaluronan, an abundant ECM molecule, can influence theeffects of collagen by preventing chondrocyte aggregation. Degradation of hyaluronan with hyaluronidase results in chondro-cyte aggregation, and this reduces the level of chondrocyte apoptosis. Experiments with an antibody to integrin b suggest that1the collagen]chondrocyte interactions are mediated through integrin b , and these interactions may protect chondrocytes1from apoptosis. We hypothesize that hyaluronan binds aggrecan and link protein, forming stable ternary complexes, whichinteract with the chondrocyte surface, perhaps via CD44, and thus maintains a stable chondrocyte]matrix network. Q 1999Elsevier Science B.V.rInternational Society of Matrix Biology. All rights reserved.
Keywords: Collagen; Hyaluronan; Chondrocyte; Adhesion; Apoptosis
1. Introduction
The central event in the pathogenesis of joint dis-eases such as osteoarthritis is the progressive deteri-
Žoration and destruction of cartilage Lohmander et
Abbre¨iations: HA, hyaluronan; HAse, hyaluronidase; HAR,hyaluronan receptor; LP, link protein; ECM, extracellular matrix;DMEM, Dulbecco’s modified Eagle’s medium; FBS, fetal bovineserum; PAGE, polyacrylamide gel electrophoresis
U Corresponding author. q1-416-480-5874; fax: q1-416-480-5737.Ž .E-mail address: [email protected]. B.B. Yang
1On sabbatical from the University of Calgary.2Scholar of the Arthritis Society of Canada.
.al., 1993; Poole et al., 1994; Lark et al., 1997 . Incartilage, the most abundant macromolecules are theextracellular matrix molecules such as collagen,hyaluronan and aggrecan. It has been observed thatexpression of collagen and aggrecan increases in the
Žearly stages of osteoarthritis Matyas et al., 1995,.1997 . However, the increase in expression of these
two molecules is disproportionate. As a result, theconcentration of aggrecan is relatively higher thanthat of collagen in the osteoarthritic cartilage. Thisconcentration disequilibrium may be a contributingfactor to this disease pathogenesis.
An in vitro model of the three-dimensional cultureof chondrocytes is essential to study the metabolism
0945-053Xr99r$ - see front matter Q 1999 Elsevier Science B.V.rInternational Society of Matrix Biology. All rights reserved.Ž .PII: S 0 9 4 5 - 0 5 3 X 9 9 0 0 0 2 7 - X

( )L. Cao et al. r Matrix Biology 18 1999 343]355344
of the extracellular matrix molecules in detail. Ini-tially, we have found that chondrocytes always adhereto the tissue culture plates and eventually exhibit afibroblast-like phenotype. Chondrocytes in monolayerexpress low levels of extracellular matrix moleculesand hence their environment is completely differentfrom the in vivo environment. To simulate the in vivoenvironment more accurately, we grew chondrocytesin an alginate gel so as to obtain a three-dimensionalculture. Although the cells expressed increasedamounts of matrix molecules, they were immobilized,making it difficult to study their activities. We there-fore developed a method to grow chondrocytes as asuspension culture. Cells in such cultures expressedhigh levels of matrix molecules and thus, at least inthis respect, mimic the conditions found in cartilagetissue. Using this model, we observed that complexesincluding aggrecan, hyaluronan and link protein playan important role in chondrocyte]matrix interactionŽ .Yang et al., 1998 . Moreover, we also observed that,after the culture media were changed, some chondro-cytes looked abnormal and appeared to have under-gone programmed cell death, or apoptosis.
Chondrocyte apoptosis may be involved in a num-ber of disease states. It has been observed in rheuma-
Ž .toid arthritis synovium Firestein et al., 1995 . Chon-drocyte number and DNA concentration are reduced
Žin osteoarthritic cartilage Vignon et al., 1983; Matyas.et al., 1995 , perhaps due to programmed death of
chondrocytes. Studies of transgenic mice lacking typeII collagen have provided direct evidence that chon-drocytes undergo apoptosis in the absence of collagenŽ .Yang et al., 1997 . In general, in vivo studies ofchondrocyte apoptosis are difficult since the degener-ation of cartilage is slow, occurring over a long periodof time. Cells in culture are a much more convenientmodel to study apoptosis and a number of studieshave been done in this context. For example, chon-drocyte apoptosis can be induced by nitric oxideŽ .Blanco et al., 1995 , FasrFas ligand expressionŽ .Hashimoto et al., 1997 and by terminal differentia-
Ž .tion Gibson et al., 1997 . These studies have sug-gested that chondrocyte apoptosis may be involved incartilage degradation.
Our observation that media change resulted inchondrocyte apoptosis may be due to a disruption ofthe structure of extracellular matrix and the interac-tion of chondrocyte with matrix. If so, it would pro-vide an appropriate model to investigate the role ofextracellular matrix molecules and how they modulatechondrocyte apoptosis, which may be an essentialcellular event in osteoarthritic cartilage development.To test this, we investigated the roles of collagen andthe aggrecan]hyaluronan-link protein ternary com-plexes, since they are the major structural moleculesin cartilage. We report here that the removal of
collagen induced apoptosis of chondrocytes in suspen-sion cultures; cell death was prevented by re-additionof collagen. Hyaluronidase treatment led to chondro-cyte attachment and aggregation, which inhibitedchondrocyte apoptosis. An anti-integrin b monoclo-1nal antibody abolished the ability of collagen to medi-ate chondrocyte aggregation, presumably by hinderingessential collagen]integrin interaction.
2. Methods
2.1. Chondrocyte isolation and culture
Chondrocytes were isolated from the sterna of 18-day chicken embryos. The sterna were removed usingsterile scissors and rinsed with PBS. Forty-eightchicken embryos were used each time. The caudal
Ž .portion 1r3 of each sterna was removed with asterile surgical knife and incubated in dissociation
Ž .medium 0.3% collagenase dissolved in HBSS at 378Cfor 30 min. The dissociation medium was changed andincubation took place for an additional 1 h. At thispoint an equal volume of growth medium was addedto stop the enzymatic reaction. The mixture was vor-texed and the cells were collected by passing themixture through a filter and centrifugation at 1100rev.rmin. The cells were resuspended in growth
Ž .medium DMEM containing 10% FBS, Gibco . Thesizes and morphology of the cells were uniform.
The newly-isolated chondrocytes can be maintainedas a suspension culture in tissue culture plates. Afterseveral days, some chondrocytes adhered to the tissueculture plates. Gradually, the chondrocytes could becultured as two pools: an adherent culture and asuspension culture. A small portion of each passageof suspended chondrocytes always attached to theculture plates. Nevertheless, a pool of chondrocytesgrowing in suspension could be maintained by passingonly the suspended chondrocytes to a new cultureeach time. The adherent chondrocytes exhibited afibroblast-like morphology and could be maintainedfor many months. However, chondrocytes growing insuspension could only be maintained for approxi-mately 1 month before they lost the suspensionproperty and adhered to the tissue culture plates. Allexperiments were performed in the early stages of
Ž .cultures 2 weeks after cell isolation while cells wereactively proliferating and matrix production was suf-ficient to maintain the cells as suspension cultures.The chondrocytes in these stages were uniform in sizeand morphology. They were hypertrophic chondro-cytes and were matrix-dependent for survival.
We also maintained suspension cultures on bacte-rial petri dishes and agarose-coated plates. Thesetechniques allowed chondrocytes to be maintained

( )L. Cao et al. r Matrix Biology 18 1999 343]355 345
longer as suspension cultures. As well, to reduce theamount of apoptosis, we changed only half of spentculture medium with fresh medium. In this way, therewas always a sufficient amount of extracellular matrixmolecules surrounding the cells, and this method hasallowed us to maintain the suspension cultures formany weeks. However, the morphology of the cellschanged eventually, and the sizes of some cells be-came much larger. We stopped growing such culturesat this stage.
2.2. Chondrocyte apoptosis analyzed by FACS and DNAfragmentation
Apoptosis was analyzed by flow cytometry of pro-Ž .pidium iodide PI -labeled DNA, as described byŽ . 5Nicoletti et al. 1991 . Briefly, 5=10 chondrocytes
were grown in 100-mm bacterial petri dishes withappropriate treatment. For the apoptosis assay, thecells were pelleted and resuspended in 1.2 ml of
Ž .hypotonic PI solution 50 mgrml dissolved in 0.1%Ž .sodium citrate plus 0.1% Triton x-100 Sigma .
DNA fragmentation of apoptotic chondrocytes wasŽ .assessed as described by Smith et al. 1989 . Chondro-
cytes were pooled together and resuspended in l ml ofŽlysis solution 10 mM Tris]Cl, pH 8.0, 100 mM NaCl,
.25 mM EDTA and 1% SDS . The lysis mixture wasincubated at 378C for 1 h with gentle agitation fol-
Ž .lowed by extraction with phenolrchloroform 2= .The DNA in the upper phase was precipitated with2.5 volumes of ethanol and resuspended in TE bufferŽ .10 mM Tris]Cl, pH 8.0, 1 mM EDTA at a concen-tration of 100 mgrml. The DNA samples were treatedwith RNase-A, subjected to 2% agarose gel elec-trophoresis, stained with ethidium bromide and pho-tographed.
2.3. Chondrocyte apoptosis and interaction affected bycollagen
To test the effect of collagen on chondrocyte apop-Žtosis, collagenase Sigma, Cat. No. C1913, dissolved in.HBSS, 2000 Urmg , was added to the chondrocyte
cultures at different concentrations as indicated inthe figure legends. The conditions for maintaining thecultures were also provided in detail in each figure.Chondrocyte apoptosis was analyzed by flow cytome-try.
In collagen-add-back experiments, chondrocyte cul-tures were digested with collagenase at a concentra-tion of 2 mgrml at 378C for 30 min. Chondrocyteswere collected by centrifugation and resuspended inDMEM supplemented with 10% FBS, to which type I
Žcollagen named as type II, Sigma, Cat. No. C 8886,.from bovine achilles tendon was added at a final
concentration of 50 mgrml. The cultures were main-tained at 378C in a tissue culture incubator for 24 hand chondrocyte apoptosis was analyzed. As the typeI collagen was difficult to dissolve in water nor ingrowth medium, we used two methods to make itdissolve. Type I collagen was briefly digested with
Ž .collagenase 0.05% at 378C for 1 h to reduce vis-cosity. The collagenase was then heat-inactivated andthe collagen solution could be used in this way. Alter-natively, we also used a mortar and pestle to groundthe collagen to increase solubility and reduce vis-cosity. After collagen dissolved, the solution was fil-tered before use. As a control, a different product of
Žtype II collagen Sigma, Cat. No. C 9301, from chicken.sternal cartilage was used to test its effect on inhibit-
ing chondrocyte apoptosis.To examine the role of collagen in chondrocyte
aggregation, adherent chondrocytes were harvestedand resuspended in regular culture media with orwithout exogenous addition of type I collagen at aconcentration of 50 mgrml. Chondrocyte aggregationwas examined using a light microscope.
2.4. Chondrocyte interaction and apoptosis affected byhyaluronan
In cell attachment assay, 24-well tissue cultureplates were coated with chicken type II collagen at aconcentration of 0.5 mgrml. The following day, themixture was removed and the plates were dried brieflyin a tissue culture incubator. Hyaluronidase or colla-genase-treated suspended chondrocytes were seededon these plates at a concentration of 5 = 104
cellsrwell and maintained at 378C for 3 h. Growthmedium and unattached chondrocytes were then re-moved by aspiration and fresh growth medium wasadded. The attachment of chondrocytes to tissue cul-ture plates was examined using light microscopy.These attached chondrocytes were harvested with abrief incubation with 10 mM EDTA and the celldensity was determined with a cytometer.
In cell-spreading assay, chondrocyte cultures grownon the collagen-coated plates were also incubated
Žwith hyaluronidase Sigma, from bovine testes, Cat..No. H 3506 at a final concentration of 200 Urml.
The cultures were incubated at 378C in a tissue cul-ture incubator for 48 h. The spreading of chondro-cytes to the plates was examined after 48 h of incuba-tion.
In a cell aggregation assay, hyaluronidase was alsoadded to the suspended chondrocyte cultures at aconcentration of 200 Urml and the cells were in-cubated at 378C for 3 h. Chondrocyte aggregation wasexamined using a light microscope.

( )L. Cao et al. r Matrix Biology 18 1999 343]355346
2.5. Detection of integrin b by FACS and Western blot1
The suspended chondrocytes were treated with orŽ . Žwithout collagenase 0.2% or hyaluronidase 200
.Urml for 30 min. The cells were then labeled withŽanti-integrin b monoclonal antibody Sigma, Cat. No.1
.I 8638 . Unlabeled cells were used as a negativecontrol. All samples were then labeled with FITC-conjugated goat-anti-mouse IgG and analyzed on a
Ž .FACScan Becton Dickinson .Protein samples collected from the same number of
Ž .cells were subjected to SDS-PAGE Laemmli, 1970 .ŽThe buffer system is 1= TG Tris-glycine buffer,
.Amresco containing 1% SDS. Proteins separated onSDS-PAGE were transblotted onto a nitrocellulose
Ž .membrane Bio-Rad in 1= TG buffer containing20% methanol. The membrane was blocked in TBSTŽ10 mM Tris]Cl pH 8.0, 150 mM NaCl, 0.05% Tween. Ž .20 containing 10% non-fat dry milk powder TBSTM
for 1 h at room temperature, and then incubated at48C overnight with primary antibody in TBSTM. The
Ž .membranes were washed with TBST 3=30 min andthen incubated for 1 h with horseradish peroxidaseŽ .HRP -conjugated secondary antibody in TBSTM. Af-ter washing as above, the bound antibody was visual-ized with an ECL kit according to the manufacturer’s
Ž .instructions Amersham .
3. Results
3.1. Loss of collagen induces chondrocyte apoptosis
To study the functions of extracellular matrixmolecules of cartilage, we maintained chondrocytes asa suspension culture and an adherent culture. Weobserved that the adherent culture was relatively easyto maintain. It developed a fibroblast-like morphologyand we were able to grow this adhesion culture in away similar to that for fibroblasts. However, the sus-pension culture was difficult to maintain. This culturetended to lose its ability to grow in suspension, andadhered to the tissue culture plates. To maintain theculture in suspension, it was essential to use onlysuspended cells at each passage. However, weobserved that after changing media, it took 2]3 daysfor the chondrocytes to recover. Some chondrocyteswere broken down and the cell debris could be easily
Ž .detected using a light microscope Fig. 1A . Initially,the suspension cultures were maintained either inpetri dishes or in agarose-coated tissue culture plates.These two culture methods are good for short-termassays. We performed most of our suspension culturesin agarose-coated plates. Those assays performed inpetri dishes were indicated in the legend of each
Ž .figure Fig. 1 .
We suspected that the chondrocytes underwentapoptosis. To confirm this, chondrocytes were har-vested before and after media change for DNA end-labeling to analyze in a FACScan. It showed that 30%of chondrocytes underwent apoptosis after the mediawere changed as compared to the 11% which were
Ž .apoptotic prior to the change of media Fig. 1b . Thisexperiment was repeated three times and the resultswere analyzed statistically. A significant difference
Ž .was obtained data not shown . Total DNA was pre-pared from chondrocytes before and after mediachange, and analyzed by agarose gel electrophoresis.The smear resulting from DNA fragmentation furtherconfirms severe chondrocyte apoptosis in chondro-
Ž .cytes harvested after media change Fig. 1c .Chondrocyte apoptosis was monitored in both sus-
pension culture and adherent culture after mediawere changed. Of the two, only the suspension cultureunderwent significant apoptosis. As the chondrocytesin the suspension cultures recovered and started togrow 1 day after inoculation, the total live chondro-cytes increased, and the proportion of apoptotic cells
Ž .declined each day Fig. 1D . To study the dose-de-pendency of media removal on chondrocyte apoptosis,different proportions of culture media were ex-changed in suspension culture and adherent culture.As seen in Fig. 1E, apoptosis in the suspension cul-ture increased as greater amounts of new media wereadded, reaching a rate of nearly 30% when all thespent media was replaced with fresh media. The ef-fect on adherent chondrocytes was negligible.
We have previously demonstrated that the sus-pended chondrocytes are surrounded by the extracel-lular matrix molecules such as hyaluronan, aggrecanand link protein and that expression of the G1 do-main of aggrecan induces chondrocyte apoptosisŽ .Yang et al., 1998; Cao and Yang, 1999 . The mostabundant matrix molecule in cartilage and chondro-cyte cultures is collagen. When the media is changed,large amounts of collagen would then be removedfrom the chondrocyte cultures. Loss of collagen maycontribute to chondrocyte apoptosis. To test this,chondrocyte cultures were incubated with or withoutcollagenase. After collagenase treatment, cultures ex-hibited significant apoptosis, as gauged by the amountof fragmented DNA, as compared with the chondro-
Ž .cyte culture without collagenase treatment Fig. 2a .Addition of type I collagen to the collagenase-treated
Ž .cultures inhibited chondrocyte apoptosis Fig. 2a .These experiments were repeated three times and
Ž .similar results were obtained each time Fig. 2b .Collagenase was introduced into the suspended
chondrocyte cultures and the adherent chondrocytecultures at different concentrations, and the cultureswere maintained at 378C for 24 h. Chondrocyte apop-tosis in the suspended culture was promoted by colla-
( )L. Cao et al. r Matrix Biology 18 1999 343]355 347
Žgenase treatment in a dose-dependent manner Fig..2c . Collagenase had little effect on the apoptosis of
the adherent chondrocytes, further suggesting thatdegradation of collagen was involved in the apoptosisof the suspended chondrocytes, rather than the effectof any protease contamination in the collagenasepreparation. Finally, when exogenous type I collagenwas added into collagenase-treated chondrocyte cul-tures and the cultures were maintained at 378C for 24
h, collagen inhibited the apoptosis in suspended chon-drocytes in a dose-dependent manner, but had little
Ž .effect on adherent chondrocytes Fig. 2d .
3.2. The roles of HA and collagen in chondrocyteinteraction and apoptosis
To study the function of collagen in chondrocyte]matrix interactions, adherent chondrocytes were har-
See overleaf for legend to Fig. 1

( )L. Cao et al. r Matrix Biology 18 1999 343]355348
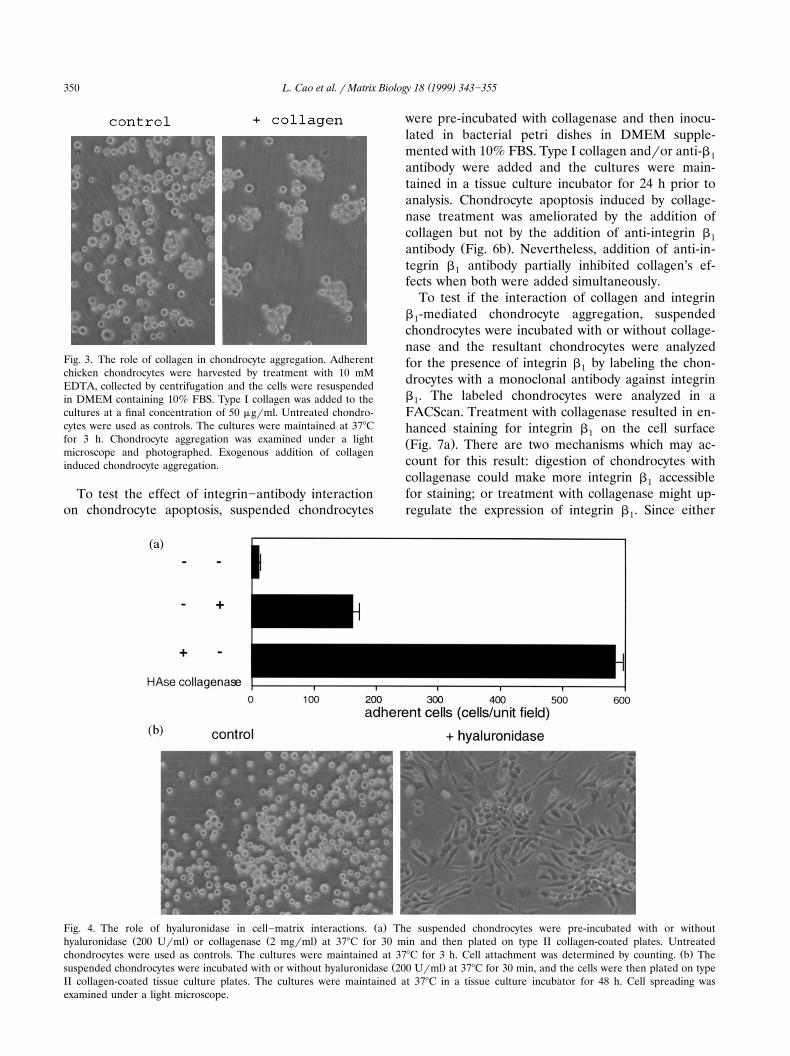
vested and incubated with or without exogenous addi-tion of type I collagen and collagenase. Normally,these chondrocytes started to aggregate 3 h after
Ž .being seeded in petri dishes Fig. 3 . Exogenous addi-tion of collagen enhanced the aggregation by forming
Ž .more and larger conglomerates Fig. 3 . Degradationof endogenous collagen by addition of collagenase
Žinhibited the aggregation of chondrocytes data not.shown .
To further test the effect of hyaluronidase andcollagenase on the interaction of chondrocytes withcollagen, the suspended chondrocytes were pre-in-cubated with or without hyaluronidase or collagenase,and then inoculated in type II collagen-coated plates.Chondrocyte attachment was analyzed. Without treat-ment, few cells adhered to the collagen-coated platesŽ .Fig. 4a . Collagenase treatment caused a small in-crease in the number of cells adhering, while
Ž .hyaluronidase HAse digestion resulted in a largeŽ .more than 20-fold increase in the number of adher-ent cells.
Hyaluronidase-digested and untreated chondro-cytes were plated on type II collagen-coated platesand maintained in a tissue culture incubator for 2days. The hyaluronidase-treated chondrocytes exhib-ited a fibroblast-like morphology and spread on the
Ž .collagen-coated plates Fig. 4b . The untreated chon-Ž .drocytes still grew as a suspension culture Fig. 4b .
Another major component of cartilage is hyaluro-nan. To test the effects of this molecule, suspendedchondrocytes were pre-incubated with collagenase andthen treated with hyaluronidase. The addition of
Žhyaluronidase inhibited chondrocyte apoptosis Fig..5a . At the concentrations used for suspended chon-
drocytes, hyaluronidase was also incubated with ad-herent chondrocytes, and they had little effect on
Ž .chondrocyte apoptosis data not shown . This indi-cated that the effect of any possible protease contami-
nation in the hyaluronidase preparations on chondro-cyte apoptosis was negligible. The suspended chon-drocytes were pre-digested with collagenase and thenthe cells were harvested and incubated in type Icollagen. Chondrocyte apoptosis was analyzed withFACScan. The results showed that chondrocyte apop-tosis induced by collagenase digestion was signifi-
Ž .cantly inhibited by addition of collagen Fig. 5b . Totest the effects of collagenase in chondrocyte apopto-sis, collagenase was introduced into the suspendedchondrocytes. We found that collagenase treatment
Ž .caused significant apoptosis Fig. 5c .The effects of type I and type II collagen on chon-
drocyte apoptosis were then tested by addition ofcollagen solution to suspended chondrocytes that hadbeen treated with collagenase for 30 min. The cul-tures were maintained in an incubator for 24 h fol-lowed by analysis of cell apoptosis. Chondrocytes thathad been treated with collagenase but were not in-cubated with collagen were used as controls. As ex-pected, the effect of type II collagen was similar totype I collagen on inhibiting apoptosis of chondro-
Ž .cytes Fig. 5d .
3.3. HA moderated the effect of collagen and integrin onaggregation and apoptosis
Integrin b , a cell surface molecule expressed by1Žchondrocytes, binds to collagen Enomoto et al., 1993;
.Loeser et al., 1995 . Thus, we reasoned that integrinb may be involved in collagen’s mediation of chon-1drocyte aggregation. To test this, suspended chondro-cytes were incubated with hyaluronidase, withhyaluronidase and collagenase, or hyaluronidase andanti-integrin b antibody. Chondrocyte aggregation1was examined with a light microscope. Hyaluronidasedigestion induced aggregation of the suspended chon-drocytes as compared with the untreated chondro-
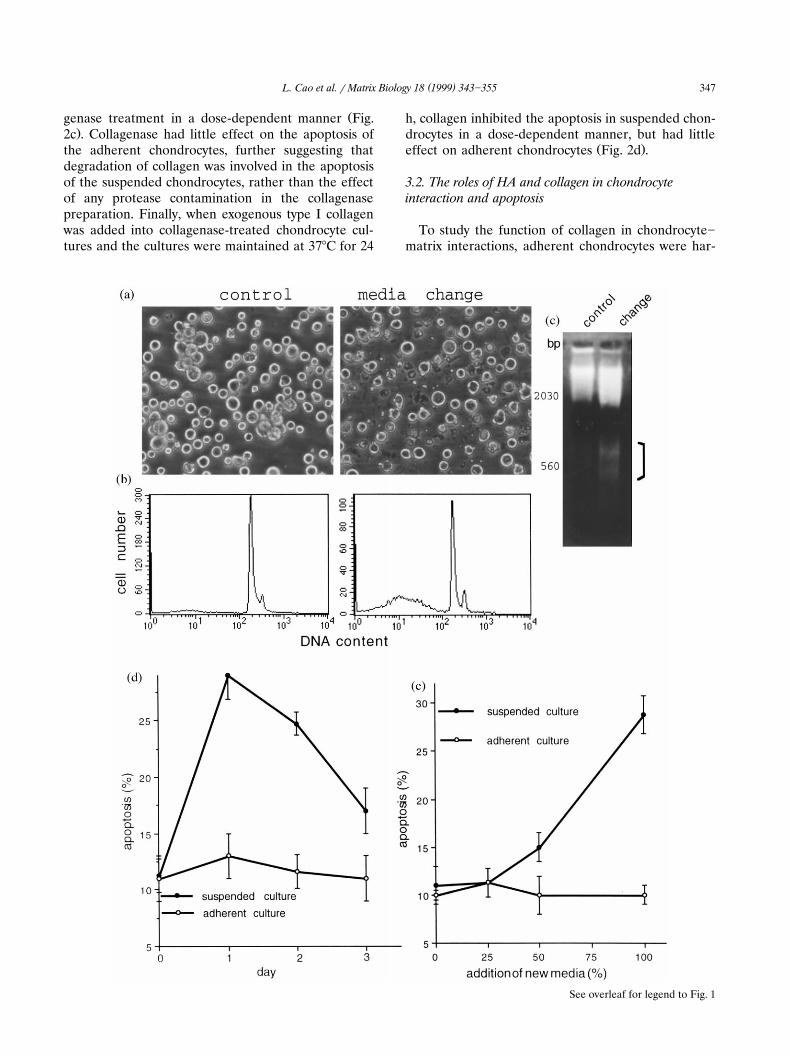
Ž .Fig. 1. The effect of media-change on chondrocyte apoptosis. a The suspended chicken chondrocytes were inoculated in 100-mm bacterial6 Ž .petri dishes at a concentration of 1=10 cellsrml subconfluence , the cultures were divided into two groups. In one group, culture media
Ž . Ž .was removed by aspiration and replaced with fresh media media changed . The other group was maintained in the same conditions control ,without the media change. The cultures were incubated at 378C in a tissue culture incubator for 24 h. The cultures were examined under alight microscope. A large amount of chondrocyte debris was detected in the cultures which were maintained in the fresh media, indicating
Ž .increased cell death. b Apoptosis was tested in chondrocytes cultured in petri dishes with or without media changed. Chondrocytes werecollected from the culture by centrifugation, stained with PI and analyzed in a FACScan. The media-changed chondrocytes exhibited a twofold
Ž . Ž .increase in apoptosis as compared with the controls 30% vs. 10.8% apoptosis . c Chondrocytes cultured in petri dishes were harvested 24 hafter culture media was changed. Cultures not subjected to media change were used as a control. Total DNA isolated from each sample wassubjected to 2% agarose gel electrophoresis and the gel was stained with ethidium bromide to visualize the DNA. DNA from cells collected
Ž . Ž .after media change was fragmented bracket area , a characteristic property of apoptosis. d Suspended chondrocytes were maintained inpetri dishes, and adherent chondrocytes were cultured in tissue culture plates. At day 0, all media in both types of treatments were carefullyremoved, and fresh media were introduced into each dish. The cultures were maintained at 378C in an incubator. Chondrocyte apoptosis wasanalyzed after media change in suspended and adherent chondrocyte cultures. In suspended cultures, apoptosis increased significantly within
Ž .24 h and then declined. The adherent chondrocytes showed little apoptosis. e The culture media were changed at different proportions inŽ . Ž .suspended cultures in petri dishes and adherent cultures in tissue culture plates . The cultures were maintained at 378C for 1 day and
chondrocyte apoptosis was analyzed by FACScan. Apoptosis in suspended chondrocytes was dose-dependent. The change of culture media inadherent chondrocytes had no effect on apoptosis.
( )L. Cao et al. r Matrix Biology 18 1999 343]355 349
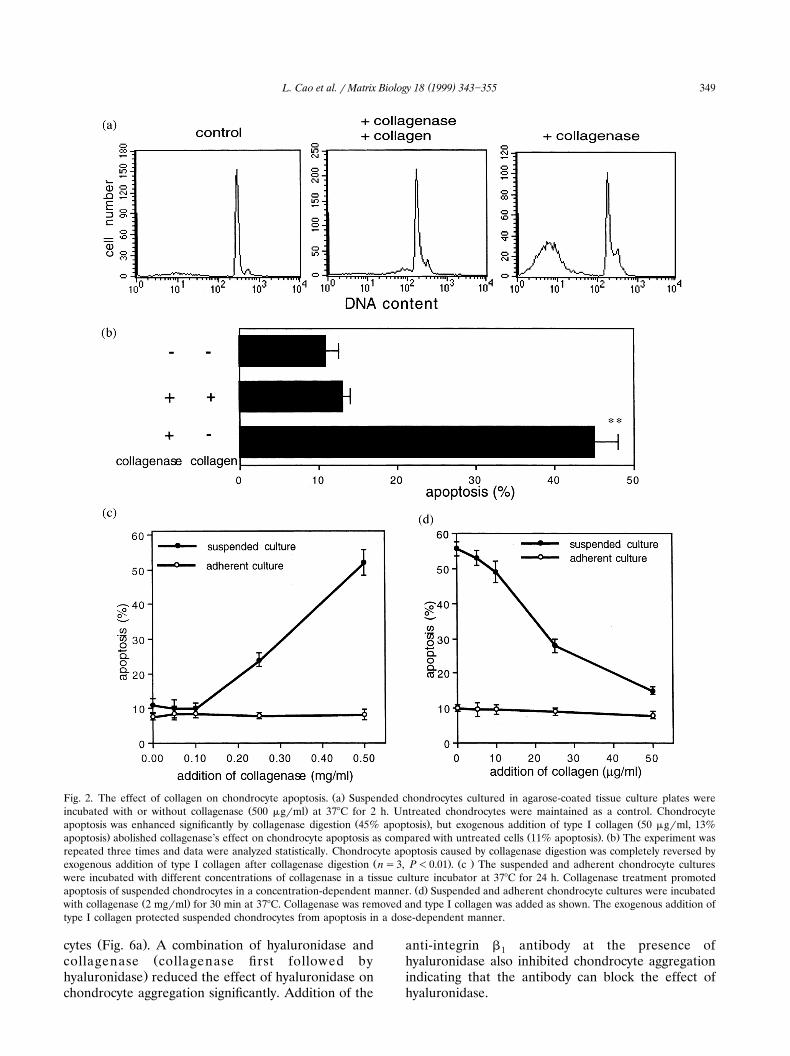
Ž .Fig. 2. The effect of collagen on chondrocyte apoptosis. a Suspended chondrocytes cultured in agarose-coated tissue culture plates wereŽ .incubated with or without collagenase 500 mgrml at 378C for 2 h. Untreated chondrocytes were maintained as a control. Chondrocyte
Ž . Žapoptosis was enhanced significantly by collagenase digestion 45% apoptosis , but exogenous addition of type I collagen 50 mgrml, 13%. Ž . Ž .apoptosis abolished collagenase’s effect on chondrocyte apoptosis as compared with untreated cells 11% apoptosis . b The experiment was
repeated three times and data were analyzed statistically. Chondrocyte apoptosis caused by collagenase digestion was completely reversed byŽ . Ž .exogenous addition of type I collagen after collagenase digestion ns3, P-0.01 . c The suspended and adherent chondrocyte cultures
were incubated with different concentrations of collagenase in a tissue culture incubator at 378C for 24 h. Collagenase treatment promotedŽ .apoptosis of suspended chondrocytes in a concentration-dependent manner. d Suspended and adherent chondrocyte cultures were incubated
Ž .with collagenase 2 mgrml for 30 min at 378C. Collagenase was removed and type I collagen was added as shown. The exogenous addition oftype I collagen protected suspended chondrocytes from apoptosis in a dose-dependent manner.
Ž .cytes Fig. 6a . A combination of hyaluronidase andŽcollagenase collagenase first followed by
.hyaluronidase reduced the effect of hyaluronidase onchondrocyte aggregation significantly. Addition of the
anti-integrin b antibody at the presence of1hyaluronidase also inhibited chondrocyte aggregationindicating that the antibody can block the effect ofhyaluronidase.
( )L. Cao et al. r Matrix Biology 18 1999 343]355350
Fig. 3. The role of collagen in chondrocyte aggregation. Adherentchicken chondrocytes were harvested by treatment with 10 mMEDTA, collected by centrifugation and the cells were resuspendedin DMEM containing 10% FBS. Type I collagen was added to thecultures at a final concentration of 50 mgrml. Untreated chondro-cytes were used as controls. The cultures were maintained at 378Cfor 3 h. Chondrocyte aggregation was examined under a lightmicroscope and photographed. Exogenous addition of collageninduced chondrocyte aggregation.
To test the effect of integrin]antibody interactionon chondrocyte apoptosis, suspended chondrocytes
were pre-incubated with collagenase and then inocu-lated in bacterial petri dishes in DMEM supple-mented with 10% FBS. Type I collagen andror anti-b1antibody were added and the cultures were main-tained in a tissue culture incubator for 24 h prior toanalysis. Chondrocyte apoptosis induced by collage-nase treatment was ameliorated by the addition ofcollagen but not by the addition of anti-integrin b1
Ž .antibody Fig. 6b . Nevertheless, addition of anti-in-tegrin b antibody partially inhibited collagen’s ef-1fects when both were added simultaneously.
To test if the interaction of collagen and integrinb -mediated chondrocyte aggregation, suspended1chondrocytes were incubated with or without collage-nase and the resultant chondrocytes were analyzedfor the presence of integrin b by labeling the chon-1drocytes with a monoclonal antibody against integrinb . The labeled chondrocytes were analyzed in a1FACScan. Treatment with collagenase resulted in en-hanced staining for integrin b on the cell surface1Ž .Fig. 7a . There are two mechanisms which may ac-count for this result: digestion of chondrocytes withcollagenase could make more integrin b accessible1for staining; or treatment with collagenase might up-regulate the expression of integrin b . Since either1
Ž .Fig. 4. The role of hyaluronidase in cell]matrix interactions. a The suspended chondrocytes were pre-incubated with or withoutŽ . Ž .hyaluronidase 200 Urml or collagenase 2 mgrml at 378C for 30 min and then plated on type II collagen-coated plates. Untreated
Ž .chondrocytes were used as controls. The cultures were maintained at 378C for 3 h. Cell attachment was determined by counting. b TheŽ .suspended chondrocytes were incubated with or without hyaluronidase 200 Urml at 378C for 30 min, and the cells were then plated on type
II collagen-coated tissue culture plates. The cultures were maintained at 378C in a tissue culture incubator for 48 h. Cell spreading wasexamined under a light microscope.

( )L. Cao et al. r Matrix Biology 18 1999 343]355 351
Fig. 5. The effect of hyaluronidase and collagenase on apoptosis.Ž . Ža Suspended chondrocytes were treated with collagenase 2
.mgrml for 30 min, and then incubated in culture media with orŽ .without hyaluronidase 200 Urml at 378C in a tissue incubator for
24 h. Chondrocyte apoptosis was analyzed by FACS. Without theaddition of hyaluronidase, chondrocytes underwent significantapoptosis due to collagenase digestion. Addition of hyaluronidase
Ž .reduced the apoptosis. b Suspended chondrocytes were pre-treatedŽ .with collagenase 2 mgrml at 378C for 30 min and then the treated
chondrocytes were harvested by centrifugation and incubated withŽ .type I collagen 50 mgrml . The cultures were maintained at 378C
for 24 h and chondrocyte apoptosis was analyzed by FACS. Apopto-sis induced by collagenase treatment was reduced significantly by
Ž .exogenous addition of type I collagen. c Suspended chondrocytesŽ .were treated with collagenase 2 mgrml at 378C for 24 h. Un-
treated chondrocytes were used as controls. Chondrocyte apoptosiswas determined. Treatment with collagenase induced chondrocyte
Ž .apoptosis significantly. d To compare the protecting effects oftype I and type II collagen on chondrocyte apoptosis, suspensionchondrocytes were pre-treated with collagenase and cells harvestedas described above. Type I and type II collagen were added to thechondrocytes at a concentration of 50 mgrml, respectively. Collage-nase-treated chondrocytes without addition of collagen were usedas controls. Apoptosis of chondrocytes was analyzed as above.Addition of type II collagen, similar to type I collagen, inhibitedchondrocyte apoptosis as compared with the controls.
one could result in chondrocyte aggregation, it wasnecessary to test which mechanism was responsible.Cell lysate was prepared from the collagenase-treatedchondrocytes and the untreated chondrocytes, andanalyzed for integrin b expression in Western blot1stained with the same antibody as above. Since littlechange in the level of b expression could be detected1
Žin cultures treated with or without collagenase Fig..7b and this result was confirmed by repeating the
experiment, it seems unlikely that increased b ex-1pression is responsible for the increased b staining1observed in collagenase-treated cultures. Under nor-mal conditions, collagen probably binds to integrin b1in a way that obstructs the interaction of anti-integrinb antibody with b . Collagenase treatment would1 1thus remove this obstruction and result in enhancedb staining.1
Another major component surrounding chondro-cytes in cartilage is hyaluronan. To test the possibilitythat hyaluronan may also block the availability ofintegrin b , the suspended chondrocytes were in-1cubated with or without hyaluronidase and the resul-tant chondrocytes were stained for accessible integrinb as described above. The result showed that1hyaluronidase digestion also enhanced the availabilityof integrin b stained by the anti-integrin b antibody1 1Ž .Fig. 7c . However, hyaluronidase treatment did notenhance the expression of integrin b , as analyzed by1
Ž .Western blot Fig. 7d . This experiment was repeatedthree times and similar result was obtained. To assurethat collagenase and hyaluronidase had no effect onintegrin b expression, adherent chondrocytes were1incubated with collagenase and hyaluronidase for 3days. Expression of integrin b did not change in the1adherent chondrocytes analyzed with Western blot
Ž .assays data not shown .The results described above suggest that the inter-
action of collagen and integrin can modulate chon-drocyte aggregation and apoptosis. Hyaluronan in-hibits chondrocyte aggregation. We present a model
Ž .to explain such relationships Fig. 8 . In this model,hyaluronan binds to aggrecan and link protein toform stable ternary complexes. These complexes me-diate chondrocyte]matrix interactions by binding tochondrocyte surface through hyaluronan-binding re-ceptors and inhibit chondrocyte aggregation. Degra-dation of hyaluronan will destroy the complexes andinduce chondrocyte aggregation. As our experimentsusing the anti-integrin b antibody suggest, aggrega-1tion seems to be mediated by an interaction betweenintegrin, an integral membrane protein found on thesurface of the plasma membrane, and collagen, aubiquitous extracellular matrix molecule.
4. Discussion
Our studies stemmed from our observation thatchondrocytes maintained as a suspension culture suf-fered from significant levels of cell death every timeculture media were changed. Chondrocytes secretelarge amounts of extracellular matrix molecules, andthese spent culture media contained collagen,
( )L. Cao et al. r Matrix Biology 18 1999 343]355352
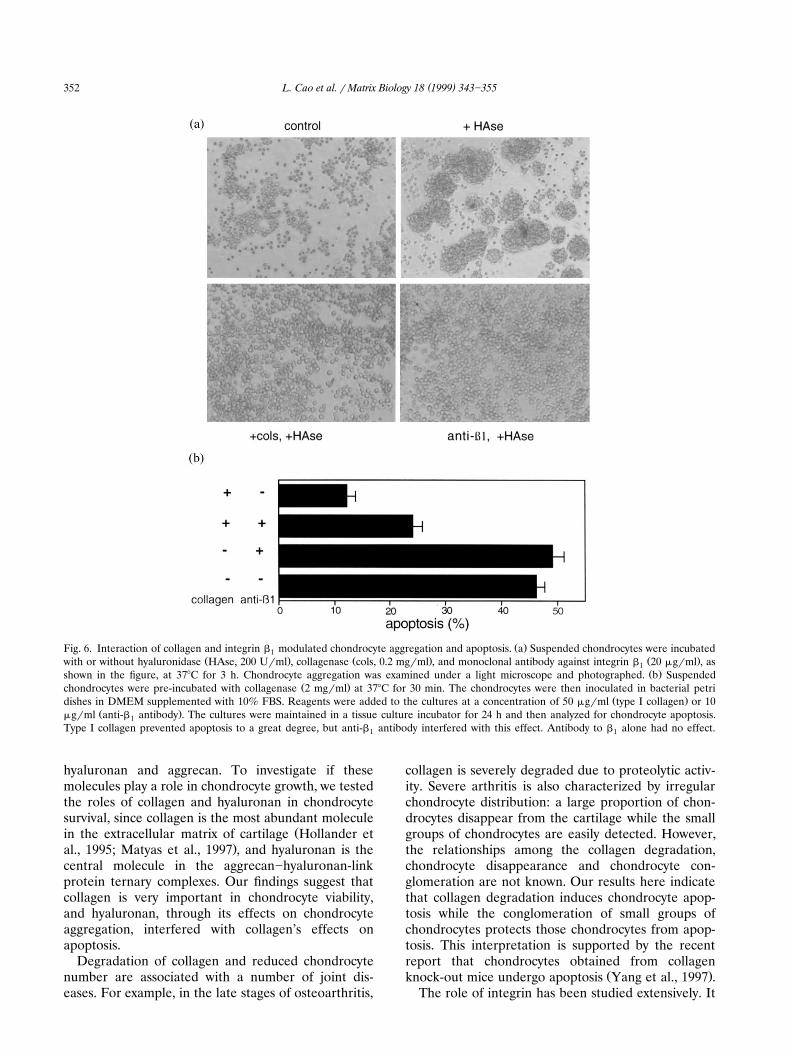
Ž .Fig. 6. Interaction of collagen and integrin b modulated chondrocyte aggregation and apoptosis. a Suspended chondrocytes were incubated1Ž . Ž . Ž .with or without hyaluronidase HAse, 200 Urml , collagenase cols, 0.2 mgrml , and monoclonal antibody against integrin b 20 mgrml , as1
Ž .shown in the figure, at 378C for 3 h. Chondrocyte aggregation was examined under a light microscope and photographed. b SuspendedŽ .chondrocytes were pre-incubated with collagenase 2 mgrml at 378C for 30 min. The chondrocytes were then inoculated in bacterial petri
Ž .dishes in DMEM supplemented with 10% FBS. Reagents were added to the cultures at a concentration of 50 mgrml type I collagen or 10Ž .mgrml anti-b antibody . The cultures were maintained in a tissue culture incubator for 24 h and then analyzed for chondrocyte apoptosis.1
Type I collagen prevented apoptosis to a great degree, but anti-b antibody interfered with this effect. Antibody to b alone had no effect.1 1
hyaluronan and aggrecan. To investigate if thesemolecules play a role in chondrocyte growth, we testedthe roles of collagen and hyaluronan in chondrocytesurvival, since collagen is the most abundant molecule
Žin the extracellular matrix of cartilage Hollander et.al., 1995; Matyas et al., 1997 , and hyaluronan is the
central molecule in the aggrecan]hyaluronan-linkprotein ternary complexes. Our findings suggest thatcollagen is very important in chondrocyte viability,and hyaluronan, through its effects on chondrocyteaggregation, interfered with collagen’s effects onapoptosis.
Degradation of collagen and reduced chondrocytenumber are associated with a number of joint dis-eases. For example, in the late stages of osteoarthritis,
collagen is severely degraded due to proteolytic activ-ity. Severe arthritis is also characterized by irregularchondrocyte distribution: a large proportion of chon-drocytes disappear from the cartilage while the smallgroups of chondrocytes are easily detected. However,the relationships among the collagen degradation,chondrocyte disappearance and chondrocyte con-glomeration are not known. Our results here indicatethat collagen degradation induces chondrocyte apop-tosis while the conglomeration of small groups ofchondrocytes protects those chondrocytes from apop-tosis. This interpretation is supported by the recentreport that chondrocytes obtained from collagen
Ž .knock-out mice undergo apoptosis Yang et al., 1997 .The role of integrin has been studied extensively. It
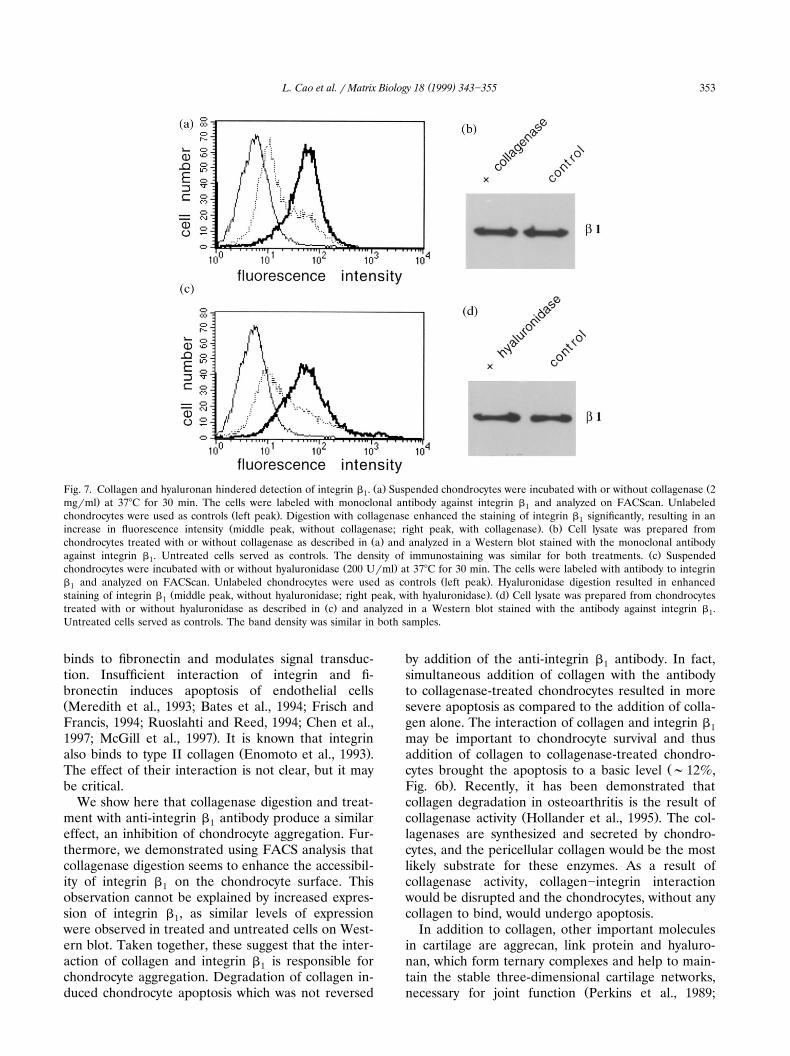
( )L. Cao et al. r Matrix Biology 18 1999 343]355 353
Ž . ŽFig. 7. Collagen and hyaluronan hindered detection of integrin b . a Suspended chondrocytes were incubated with or without collagenase 21.mgrml at 378C for 30 min. The cells were labeled with monoclonal antibody against integrin b and analyzed on FACScan. Unlabeled1
Ž .chondrocytes were used as controls left peak . Digestion with collagenase enhanced the staining of integrin b significantly, resulting in an1Ž . Ž .increase in fluorescence intensity middle peak, without collagenase; right peak, with collagenase . b Cell lysate was prepared from
Ž .chondrocytes treated with or without collagenase as described in a and analyzed in a Western blot stained with the monoclonal antibodyŽ .against integrin b . Untreated cells served as controls. The density of immunostaining was similar for both treatments. c Suspended1
Ž .chondrocytes were incubated with or without hyaluronidase 200 Urml at 378C for 30 min. The cells were labeled with antibody to integrinŽ .b and analyzed on FACScan. Unlabeled chondrocytes were used as controls left peak . Hyaluronidase digestion resulted in enhanced1
Ž . Ž .staining of integrin b middle peak, without hyaluronidase; right peak, with hyaluronidase . d Cell lysate was prepared from chondrocytes1Ž .treated with or without hyaluronidase as described in c and analyzed in a Western blot stained with the antibody against integrin b .1
Untreated cells served as controls. The band density was similar in both samples.
binds to fibronectin and modulates signal transduc-tion. Insufficient interaction of integrin and fi-bronectin induces apoptosis of endothelial cellsŽMeredith et al., 1993; Bates et al., 1994; Frisch andFrancis, 1994; Ruoslahti and Reed, 1994; Chen et al.,
.1997; McGill et al., 1997 . It is known that integrinŽ .also binds to type II collagen Enomoto et al., 1993 .
The effect of their interaction is not clear, but it maybe critical.
We show here that collagenase digestion and treat-ment with anti-integrin b antibody produce a similar1effect, an inhibition of chondrocyte aggregation. Fur-thermore, we demonstrated using FACS analysis thatcollagenase digestion seems to enhance the accessibil-ity of integrin b on the chondrocyte surface. This1observation cannot be explained by increased expres-sion of integrin b , as similar levels of expression1were observed in treated and untreated cells on West-ern blot. Taken together, these suggest that the inter-action of collagen and integrin b is responsible for1chondrocyte aggregation. Degradation of collagen in-duced chondrocyte apoptosis which was not reversed
by addition of the anti-integrin b antibody. In fact,1simultaneous addition of collagen with the antibodyto collagenase-treated chondrocytes resulted in moresevere apoptosis as compared to the addition of colla-gen alone. The interaction of collagen and integrin b1may be important to chondrocyte survival and thusaddition of collagen to collagenase-treated chondro-
Žcytes brought the apoptosis to a basic level ;12%,.Fig. 6b . Recently, it has been demonstrated that
collagen degradation in osteoarthritis is the result ofŽ .collagenase activity Hollander et al., 1995 . The col-
lagenases are synthesized and secreted by chondro-cytes, and the pericellular collagen would be the mostlikely substrate for these enzymes. As a result ofcollagenase activity, collagen]integrin interactionwould be disrupted and the chondrocytes, without anycollagen to bind, would undergo apoptosis.
In addition to collagen, other important moleculesin cartilage are aggrecan, link protein and hyaluro-nan, which form ternary complexes and help to main-tain the stable three-dimensional cartilage networks,
Žnecessary for joint function Perkins et al., 1989;

( )L. Cao et al. r Matrix Biology 18 1999 343]355354
Fig. 8. A model for inhibition of collagen]chondrocyte interactionŽ .by hyaluronan. Hyaluronan HA binds to aggrecan and link pro-
Ž .tein LP forming stable ternary complexes. The complexes bind toŽ .chondrocyte surface through hyaluronan-binding receptor HAR
on the cell surface. These complexes are important in chondro-cyte]matrix interaction and they prevent the aggregation of chon-drocytes. Degradation of hyaluronan destroys the complexes. As aresult, chondrocytes conglomerate through the interaction of colla-gen and integrin.
Barry et al., 1992; Li et al., 1993; Neame and Barry,.1993; Margolis and Margolis, 1994 . We have demon-
strated that the aggrecan]hyaluronan-link proteinternary complexes are important in mediating chon-
Ž .drocyte]matrix interaction Yang et al., 1998 . How-ever, the role of these ternary complexes in chondro-cyte aggregation and apoptosis is complicated in thestudies presented here. Degradation of hyaluronanwith hyaluronidase induces chondrocyte aggregation,and the aggregation inhibits apoptosis.
Hyaluronidase also degraded chondroitin sulfate, amajor component of chondrocyte products, and thisalso enhanced chondrocyte aggregation. The aggrega-tion also reduced the level of chondrocyte apoptosis.It is conceivable that, even in the presence ofhyaluronidase, newly synthesized collagen could medi-ate chondrocyte aggregation and, therefore, inhibitapoptosis. It should be noted that degradation ofhyaluronan inducing cell aggregation might involveother mechanisms. For example, removal of hyaluro-nan will allow other adhesion molecules on the cellsurface to interact with each other.
In the chondrocyte]substratum interaction assay,we sought to directly test the role of hyaluronan inchondrocyte]collagen interaction by introducinghyaluronidase into the suspended chondrocyte cul-tures to degrade hyaluronan. Chondroitin sulfatemight also be degraded and this might result in betterinteractions of chondrocytes with collagen. Indeed,chondrocytes treated with hyaluronidase bound to thecollagen-coated plates strongly. Hyaluronan inter-fered with the interaction of chondrocytes with colla-
gen. The result of FACScan analysis showed thathyaluronidase digestion enhanced immunostaining ofintegrin b in the chondrocyte surface. We also1observed that collagenase digestion enhanced the in-teraction of chondrocytes with the collagen-coatedplates. Removal of collagen from the chondrocytesincreased the accessibility of endogenous integrin b1and enhanced chondrocyte binding to collagen-coatedplates. The result of FACScan analysis also demon-strated that collagen inhibited the immunostaining ofintegrin b .1
In normal chondrocyte cultures, addition ofhyaluronan had no effect on chondrocyte apoptosis,because collagen]chondrocyte interaction plays adominant function in protecting chondrocytes fromapoptosis. In collagenase-treated chondrocyte cul-tures, however, apoptosis is high. Addition ofhyaluronidase also inhibited chondrocyte apoptosisand this might be due to interactions between chon-drocytes and newly synthesized collagen, in theabsence of hyaluronan. The newly synthesized colla-gen-mediated chondrocyte aggregation and small con-glomerates could be seen.
Our results here demonstrate that hyaluronan,which binds to aggrecan and link protein to formstable ternary complexes, plays an important functionin inhibiting chondrocyte aggregation, and collagenplays an important role in protecting chondrocytesfrom apoptosis. Perhaps, degradation and insufficientproduction of the hyaluronan]aggrecan-link proteinternary complexes cause chondrocyte aggregation.Degradation of collagen by proteolytic activities in-duces chondrocyte apoptosis. Since these phenomenaare associated with many kinds of joint diseases, fur-ther studies are required to uncover the detailedmechanisms by which diseases develop and to findways to block such processes.
Acknowledgements
We thank Dr R.S. Kerbel for helpful comments.This work was supported by Grant MT-13730 fromthe Medical Research Council of Canada to BBY
References
Barry, F.P., Gaw, J.U., Young, C.N., Neame, P.J., 1992. Hyaluro-nan-binding region of aggrecan from pig laryngeal cartilage.Amino acid sequence, analysis of N-linked oligosaccharides andlocation of the keratan sulphate. Biochem. J. 286, 761]769.
Bates, R.C., Buret, A., van Helden, D.F., Horton, M.A., Burns,G.F., 1994. Apoptosis induced by inhibition of intercellular con-tact. J. Cell Biol. 125, 403]415.
Blanco, F.J., Ochs, R.L., Schwarz, H., Lotz, M., 1995. Chondrocyteapoptosis induced by nitric oxide. Am. J. Pathol. 146, 75]85.
Cao, L., Yang, B.B., 1999. Chondrocyte apoptosis induced by aggre-

( )L. Cao et al. r Matrix Biology 18 1999 343]355 355
can G1 domain as a result of decreased cell adhesion. Exp. CellRes. 246, 527]537.
Chen, C.S., Marksich, M., Huang, S., Whitesides, G.M., Ingber,D.E., 1997. Geometric control of cell life and death. Science 276,1425]1428.
Enomoto, M., Leboy, P.S., Menko, A.S., Boettiger, D., 1993. ß1Integrins mediate chondrocyte interaction with type I collagen,type II collagen, and fibronectin. Exp. Cell Res. 205, 276]285.
Firestein, G.S., Yeo, M., Zvaifler, N.J., 1995. Apoptosis in rheuma-toid Aarthritis synovium. J. Clin. Invest. 96, 1631]1638.
Frisch, S.M., Francis, H., 1994. Disruption of epithelial cell]matrixinteractions induces apoptosis. J. Cell Biol. 124, 619]626.
Gibson, G., Lin, D.-L., Roque, M., 1997. Apoptosis of terminallydifferentiated chondrocytes in culture. Exp. Cell Res. 233,372]382.
Hashimoto, S., Setareh, M., Ochs, R.L., Lotz, M., 1997. FASrFASligand expression and induction of apoptosis in chondrocytes.Arthritis Rheum. 40, 1749]1755.
Hollander, A.P., Pidoux, I., Reiner, A., Rorabeck, C., Bourne, R.,Poole, A.R., 1995. Damage to type II collagen in aging andosteoarthritis starts at the articular surface, originates aroundchondrocytes, and extends into the cartilage with progressivedegeneration. J. Clin. Invest. 96, 2859]2869.
Laemmli, U.K., 1970. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature 227, 680]685.
Lark, M.W., Bayne, E.K., Flanagan, J., 1997. Aggrecan degradationin human cartilage: Evidence for both matrix metalloproteinaseand aggrecanase activity in normal, osteoarthritic, and rheuma-toid joints. J. Clin. Invest. 100, 93]106.
Li, H., Schwartz, N.B., Vertel, B.M., 1993. cDNA cloning of chickŽ .cartilage chondroitin sulfate aggrecan core protein and identi-
fication of a stop codon in the aggrecan gene associated with thechondrodystrophy, nanomelia. J. Biol. Chem. 268, 23504]23511.
Loeser, R.F., Carlson, C.S., McGee, M.P., 1995. Expression of b1integrins by cultured articular chondrocytes and in osteoarthriticcartilage. Exp. Cell Res. 217, 248]257.
Lohmander, L.S., Neame, P., Sandy, J.D., 1993. The structure ofaggrecan fragments in human synovial fluid: Evidence that ag-grecanase mediates cartilage degradation in inflammatory jointdisease, joint injury and osteoarthritis. Arthritis Rheum. 36,1214]1222.
Margolis, R.U., Margolis, R.K., 1994. Aggrecan-versican-neurocanfamily proteoglycans. Meth. Enz. 245, 105]126.
Matyas, J.R., Adams, M.E., Huang, D., Sandell, L.J., 1995. Discoor-dinate gene expression of aggrecan and Type II collagen inexperimental osteoarthritis. Arthritis Rheum. 38, 420]425.
Matyas, J.R., Adams, M.E., Huang, D., Sandell, L.J., 1997. Majorrole of collagen IIB in the elevation of total Type II procollagenmessenger RNA in the hypertrophic phase of experimentalosteoarthritis. Arthritis Rheum. 40, 1046]1049.
McGill, G., Shimamura, A., Bates, R.C., Savage, R.E., Fisher, D.E.,1997. Loss of matrix adhesion triggers rapid transformation-selective apoptosis in fibroblasts. J. Cell Biol. 138, 901]911.
Meredith Jr, J.E., Fazeli, B., Schwartz, M.A, 1993. The extracellularmatrix as a cell survival factor. Mol. Biol. Cell. 4, 953]961.
Neame, P.J., Barry, F.P., 1993. The link proteins. Experientia 49,393]402.
Nicoletti, I., Migliorati, G., Pagliacci, M.C., Grignani, F., Riccardi,C., 1991. A rapid and simple method for measuring thymocyteapoptosis by propidium iodide staining and flow cytometry. J.Immunol. Methods 139, 271]279.
Perkins, S.J., Nealis, A.S., Dudhia, J., Hardingham, T.E., 1989.Immunoglobulin fold and tandem repeat structures in proteogly-can N-terminal domains and link protein. J. Mol. Biol. 206,737]748.
Poole, A.R., Ionescu, M., Swan, A., Dieppe, P.A., 1994. Changes incartilage metabolism in arthritis are reflected by altered serumand synovial fluid levels of the cartilage proteoglycan aggrecan.J. Clin. Invest. 94, 25]33.
Ruoslahti, E., Reed, J.C., 1994. Anchorage dependence, integrins,and apoptosis. Cell 77, 477]478.
Smith, C.A., Williams, G.T., Kingston, R., Jenkinson, E.J., Owen,J.J.T., 1989. Antibodies to CD3rT-cell receptor complex inducedeath by apoptosis in immature T cells in thymic cultures.Nature 337, 181]184.
Vignon, E., Arlot, M., Hartmann, D.J., Moyen, G., Ville, G., 1983.Hypertrophic repair of articular cartilage in experimental os-teoarthrosis. Ann. Rheum. Dis. 42, 82]88.
Yang, C., Li, S.-W., Helminen, H.J., Khillan, J.S., Bao, Y., Prockop,D.J., 1997. Apoptosis of chondrocytes in transgenic mice lackingcollagen II. Exp. Cell Res. 235, 370]373.
Yang, B., Zhang, Y., Cao, L., Yang, B.L., 1998. Aggrecan and linkprotein affect cell adhesion to culture plates and to type IIcollagen. Matrix Biol. 16, 541]561.





























