Embed Size (px)
Citation preview

1,25-Dihydroxyvitamin D3 Protects RINm5F and HumanIslet Cells against Cytokine-Induced Apoptosis:Implication of the Antiapoptotic Protein A20
RITA RIACHY, BRIGITTE VANDEWALLE, JULIE KERR CONTE, ERICKA MOERMAN,PAOLA SACCHETTI, BRUNO LUKOWIAK, VALERY GMYR, THOMAS BOUCKENOOGHE,MATHILDE DUBOIS, AND FRANCOIS PATTOU
Cellular Therapy of Diabetes, Institut National de la Sante et de la Recherche Medicale, Equipe de Recherche etd’Innovation Methodologique 0106 (R.R., B.V., J.K.C., B.L., V.G., T.B., M.D., F.P.), and Institut National de la Sante et dela Recherche Medicale Unite 459 (E.M., P.S.), Faculte de Medecine, 59045 Lille, France
Transplantation of islets of Langerhans is a potential cure fortype 1 diabetes, but its success is hampered by destruction ofthe islets. The data presented herein suggest that the activemetabolite of vitamin D3 [1,25-(OH)2D3] may promote islet cellsurvival by modulating the effects of inflammatory cytokines,which contribute to �-cell demise. We investigated some of themechanisms triggering the apoptotic machinery in rat insu-linoma RINm5F cells and human islets treated with IL-1� plusinterferon-� plus TNF� and assessed the effects of 1,25-(OH)2D3 in these processes. Mitochondrial transmembranepermeability and apoptotic features, determined by percent-age of sub-G1 cells, quantitation of DNA strand breaks, andHoechst staining, were significantly increased by cytokinesand reverted toward control values by 1,25-(OH)2D3 cotreat-
ment. The cytoprotection of cells correlated with the abroga-tion of cytokine-induced nitric oxide production. The activa-tion of nuclear factor-�B plays a key role in the differentpathways implicated in nitric oxide generation. We demon-strated for the first time, in both RINm5F cells and humanislets, that 1,25-(OH)2D3 was able to induce and maintain highlevels of A20, an antiapoptotic protein known to block nuclearfactor-�B activation. Our study showed a clear efficiency of1,25-(OH)2D3 on the apoptotic machinery triggered by cyto-kines in �-cells and suggests that 1,25-(OH)2D3 could help over-come a major obstacle encountered in the cellular therapy ofdiabetes, such as nonfunction in the immediate posttrans-plantation period. (Endocrinology 143: 4809–4819, 2002)
TRANSPLANTATION OF PANCREATIC islets of Lang-erhans is a promising therapy for type 1 diabetes, po-
tentially restoring physiologic insulin secretion. In experi-mental models of islet transplantation, a loss of grafted �-cellmass has been observed and attributed, at least in part, to�-cell apoptosis (1). Indeed, apoptotic activity within isletcells has been shown to increase in the immediate period postovulation (2, 3). In addition, the inflammatory response ac-tivates cell death within the islet and adversely influencesengraftment. Impaired function and destruction of �-cellsresult from direct contact with islet-infiltrating macrophagesand T lymphocytes and/or exposure to inflammatory prod-ucts of the islet-infiltrating cells such as free radicals andcytokines.
Inflammatory cytokines IL-1�, interferon (IFN)-�, andTNF� are cytotoxic to �-cells in vitro. They have been shownto induce apoptosis in purified primary human (4), rat (5),and mouse (6) �-cells, possibly by stimulating the synthesisof inducible nitric oxide synthase (7) and nitric oxide (NO)(8). NO is highly efficient in inducing mitochondrial perme-ability transition, thereby causing the liberation of apopto-genic factors from mitochondria, which can induce DNAfragmentation in pancreatic �-cells (9).
The induction of inducible nitric oxide synthase in �-cellsis predominantly regulated by the nuclear factor-�B (NF�B)(10). NF�B is normally inactive and sequestered in the cy-toplasm as a heterodimer comprising two polypeptides of 50and 65 kDa noncovalently complexed with a cytoplasmicinhibitory protein, inhibitor of �B (I�B) (11). In response toa variety of stimuli such as cytokines, I�B proteins are rapidlyphosphorylated by the I�B kinase (IKK). This results in theliberation of NF�B from I�B and subsequent translocation ofNF�B to the nucleus in which it regulates gene transcription.The antiapoptotic A20 gene blocks the activation of NF�Bupstream from the kinase cascade leading to I�B degradation(12). Consequently, A20 is a candidate gene potentially ableto down-regulate the synthesis of cytotoxic mediators.
Frequent suboptimal primary islet function on transplan-tation has drawn attention to the factors promoting isletengraftment and minimizing graft cell injury. The activemetabolite of vitamin D3, 1,25-dihydroxyvitamin D3 [1,25-(OH)2D3], is a secosteroid hormone that not only regulatesbone and calcium/phosphate metabolism but also exerts anumber of other biological activities including immuno-modulation (13) and insulinotropic effect in pancreatic�-cells (14) and vitamin D-deficient rats (15). 1,25-(OH)2D3
and its analogs also have been shown to reduce the incidenceof diabetes (16) and prolong graft survival in spontaneouslydiabetic nonobese diabetic mice (17). We have previouslyshown that 1,25-(OH)2D3 might reduce the vulnerability ofthe human pancreatic islets cells to in vitro cytotoxic cytokine
Abbreviations: IFN, Interferon; I�B, inhibitor of �B; IKK, inhibitor of�B kinase; ��m, mitochondrial transmembrane potential; NF�B, nuclearfactor-�B; NO, nitric oxide; NOi, NO synthase inhibitor NG-methyl-l-arginine; 1,25-(OH)2D3, active metabolite of vitamin D3, 1,25-dihy-droxyvitamin D3; TMRM, tetramethyl rhodamine methyl ester.
0013-7227/02/$15.00/0 Endocrinology 143(12):4809–4819Printed in U.S.A. Copyright © 2002 by The Endocrine Society
doi: 10.1210/en.2002-220449
4809
challenge by decreasing major histocompatibility complexclass I induction, IL-6 production, and nitrite release, a re-flection of NO production (18).
In the present study, we investigated the early mecha-nisms implied in �-cell death by first focusing on the ratinsulin-producing RINm5F �-cell line, a model currentlyused for the study of pancreatic cell death (10, 19–24). Weparticularly studied the disruption of the mitochondrialtransmembrane potential (��m) and apoptotic features toascertain the counteracting action of 1,25-(OH)2D3 on themechanistic targets implied in the death processes such as
NF�B activation and NO production. We then focused on theeffect of 1,25-(OH)2D3 on the expression of the antiapoptoticA20 gene and its induction by cytokines in RINm5F cells, andwe subsequently extended these experiments to human pan-creatic islet cells.
Materials and MethodsCell culture
The rat insulin-producing �-cells RINm5F obtained from ATCC (Ma-nassas, VA) were trypsinized every 5 d and subcultured (1 � 105cells/cm2) at 37 C in 5% CO2 and maintained for 24 h in RPMI 1640 mediumsupplemented with 10% fetal calf serum and antibiotics. Cell processingconsisted of treatments for 48 h in different media including cytokine-treated medium: basal medium plus IL-1� (50 IU/ml), IFN-� (1000IU/ml), and TNF� (1000 IU/ml; cytokines from Valbiotech, AbCys,Paris, France) and cytokine-treated medium containing 1,25-(OH)2D3dissolved in ethanol and used at physiological (10�8 m) and pharma-cological (10�6 m) concentrations. Alcohol concentration did not exceed0.01% in the culture media and was without measurable effect on cellcultures. 1,25-(OH)2D3 was a generous gift from Hoffmann la Roche AG(Basel, Switzerland).
In some experiments, cells were incubated with the NO synthaseinhibitor NG-methyl-l-arginine (NOi, 5 mm; Sigma-Aldrich Chemicals,Saint Quentin Fallavier, France), added for 1 h before stimulation withcytokines. After incubation, analyses were performed on free-floatingcells pooled with the cells detached by mild trypsinization.
Human islet processing
Human pancreases (mean age 38 � 6 yr, n � 5) were harvested fromadult brain-dead donors in accord with French regulations and the localInstitutional Ethical Committee. Pancreatic islets were isolated afterductal distension of the pancreas and digestion of the tissue with liberase(Roche Molecular Biochemicals, Mannheim, Germany). Semipurifica-tion was achieved with Euro-Ficoll discontinuous density gradient. Isletnumber was determined on samples of each preparation after dithizonestaining and expressed as equivalent number of islets. Preparations usedin this study exhibited a 75 � 3% purity and an average yield of 4091 �1127 equivalent number of islets per gram pancreas. Semipurified isletswere cultured for 24 h in CMRL-1066 medium with 2% Ultroser G (LifeTechnologies, Inc., Cergy Pontoise, France). Human �-cell processingconsisted of a 48-h treatment at 37 C with cytokines added or not with1,25-(OH)2D3 used in the same concentrations as for RINm5F cells.
Estimation of metabolic activity of cells
RINm5F cells were treated for 48 h with cytokines with or without1,25-(OH)2D3. Insulin released in the culture media by cells was storedat �80 C before we performed the insulin assay using the RIA kit CTfrom CIS-Bio International (Gif-sur-Yvette, France).
As described previously (25), we used the fluorescent and nontoxicREDOX indicator Alamar blue (Biosource Technologies, Inc., Clini-sciences Montrouge, France) for evaluation of cell viability. It was di-
TABLE 1. Effect of 1,25-(OH)2D3 on nitrite production in cytokine-treated RINm5F cells preincubated or not with NO synthase inhibitorNOi (5 mM)
Nitrite (nmol/106 cells—48 h)
Treatment None(Control)
1,25-(OH)2D3Cytokines
Cytokines �1,25-(OH)2D3
10�8 M
Cytokines �1,25-(OH)2D3
10�6 M10�8 M 10�6 M
Standard (6) 4.4 � 0.7 3.2 � 0.9 2.7 � 0.3 94.8 � 17.5a 71.1 � 9.0b 58.1 � 12.1b
� NOi (3) 2.4 � 0.7 1.4 � 0.8 2.5 � 1.8 3.9 � 1.3 3.4 � 0.6 3.7 � 0.9
RINm5F cells were cultured (1 � 105 cells/10-mm well) in 1.5 ml medium and exposed for 48 h to IL-1� (50 IU/ml), IFN-� (1000 IU/ml), andTNF-� (1000 IU/ml) in the absence or presence of 10�8 and 10�6 M 1,25-(OH)2D3. As an additional control, cells were also exposed to 1,25-(OH)2D3alone. Nitrite accumulation in the media was determined as described in Materials and Methods. Values are means � SEM with the numberof observations within parentheses.
a P � 0.0001, significantly different than the control cells.b P � 0.05, significantly different than cytokine-treated cells.
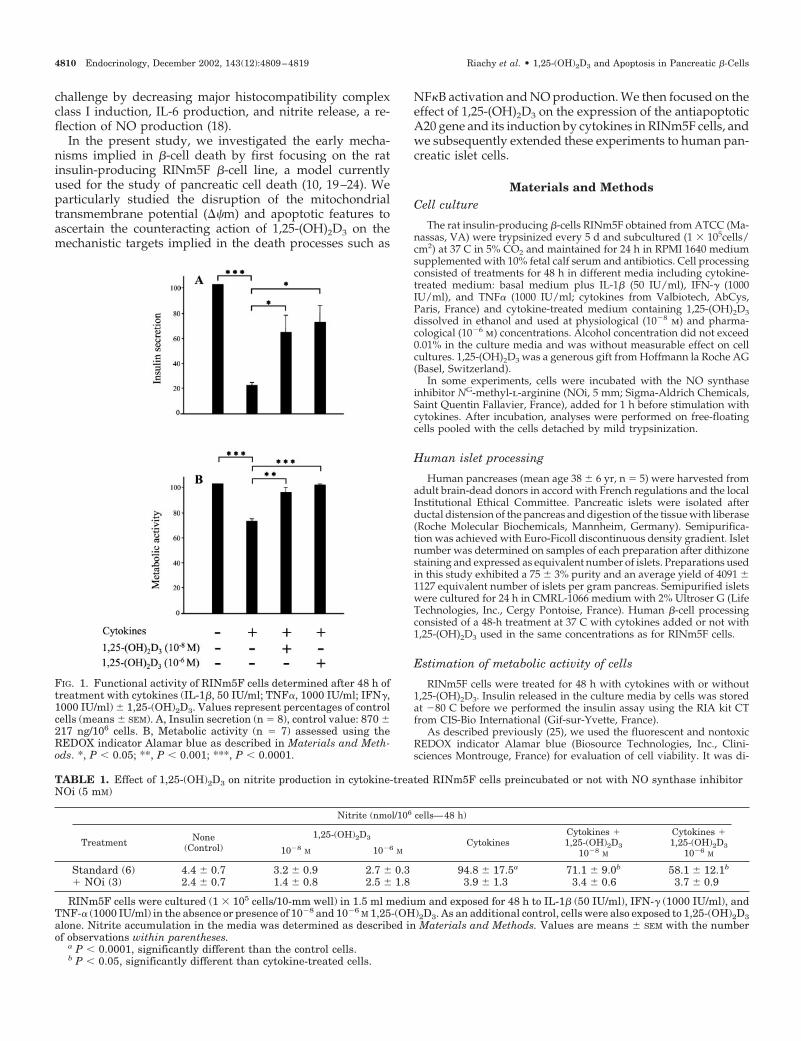
FIG. 1. Functional activity of RINm5F cells determined after 48 h oftreatment with cytokines (IL-1�, 50 IU/ml; TNF�, 1000 IU/ml; IFN�,1000 IU/ml) � 1,25-(OH)2D3. Values represent percentages of controlcells (means � SEM). A, Insulin secretion (n � 8), control value: 870 �217 ng/106 cells. B, Metabolic activity (n � 7) assessed using theREDOX indicator Alamar blue as described in Materials and Meth-ods. *, P � 0.05; **, P � 0.001; ***, P � 0.0001.
4810 Endocrinology, December 2002, 143(12):4809–4819 Riachy et al. • 1,25-(OH)2D3 and Apoptosis in Pancreatic �-Cells
rectly added (10% vol/vol) to cell culture medium for 2 h at 37 C in anatmosphere of air: CO2 (95:5). The fluorescence was read at 544 nm(excitation wavelength) and 590 nm (emission wavelength).
After exposure of cells for 48 h to the different experimental condi-tions, duplicate samples (2 � 120 �l) of media were taken for nitritedetermination. Nitrite assay is based on the reaction of nitrite with2,3-diaminonaphtalene (Sigma-Aldrich Chemicals) to form the fluores-cent product 1-(H)-naphtotriazole, as previously described (25).
Detection of ��m
The ��m was measured by incubating RINm5F cells (5 � 105/ml)during 1 h at 37 C with 0.1 �m lipophilic cationic fluorochrome tetra-methyl rhodamine methyl ester (TMRM; Molecular Probes, Inc. Europe,Leiden, The Netherlands), which accumulates in the mitochondrial ma-trix. Cells were analyzed by flow cytometry (Epics XL-MCL Coulter flowcytometer, Beckman, Margency, France) at 575 nm. A reduction in flu-
TABLE 2. Effect of 1,25-(OH)2D3 on mitochondrial transmembrane potential (��m) in cytokine-treated RINm5F cells preincubated or notwith NO synthase inhibitor NOi (5 mM)
Percentage of cells with low intensity fluorescence
Treatment None(Control)
1,25-(OH)2D3Cytokines
Cytokines �1,25-(OH)2D3
10�8 M
Cytokines �1,25-(OH)2D3
10�6 M10�8 M 10�6 M
Standard (9) 31 � 6 29 � 11 35 � 11 72 � 8a 51 � 7b 55 � 8b
� NOi (3) 17 � 3 16 � 1 20 � 3 27 � 10 16 � 3 16 � 5
RINm5F cells were cultured (1 � 105 cells/10-mm well) in 1.5 ml medium and exposed for 48 h to IL-1� (50 IU/ml), IFN-� (1000 IU/ml), andTNF-� (1000 IU/ml) in the absence or presence of 10�8 and 10�6 M 1,25-(OH)2D3. As an additional control, cells were also exposed to 1,25-(OH)2D3alone. Cells were incubated with 0.1 mM TMRM during 1 h at 37 C before immediately analyzed by flow cytometry. Values (means � SEM)represent the percentages of low fluorescent cells with depolarized mitochondria stained with TMRM with the number of observations withinparentheses.
a P � 0.005, significantly different than the control cells.b P � 0.05, significantly different than cytokine-treated cells.
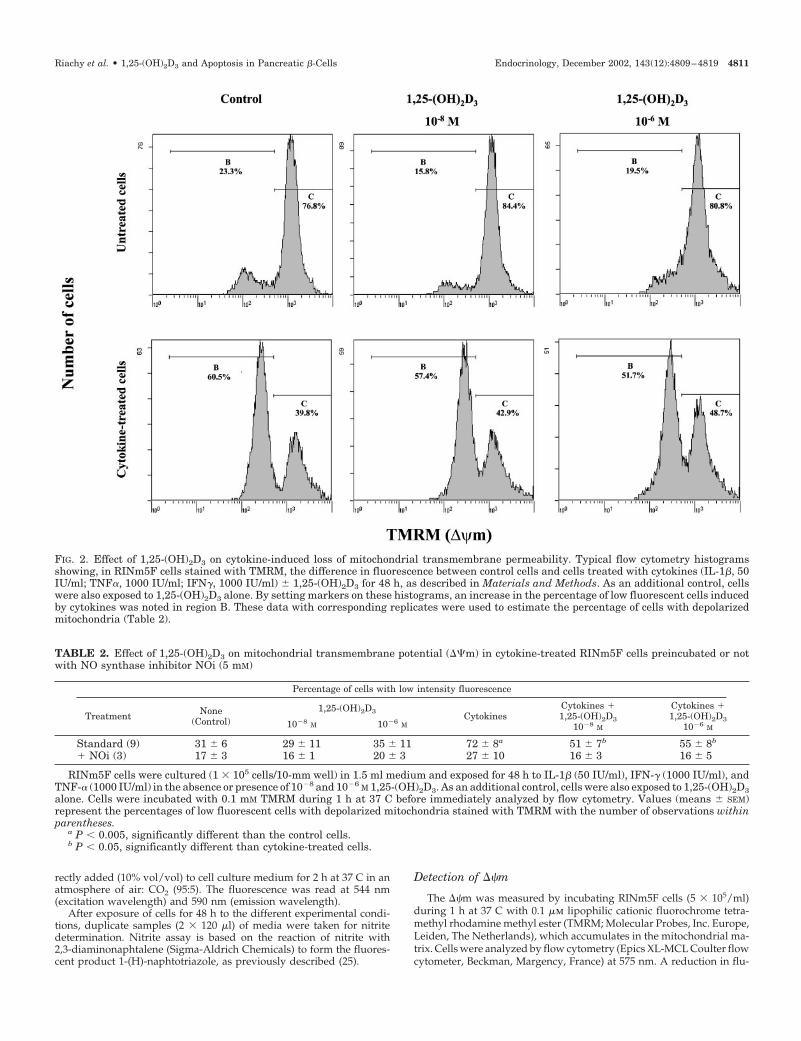
FIG. 2. Effect of 1,25-(OH)2D3 on cytokine-induced loss of mitochondrial transmembrane permeability. Typical flow cytometry histogramsshowing, in RINm5F cells stained with TMRM, the difference in fluorescence between control cells and cells treated with cytokines (IL-1�, 50IU/ml; TNF�, 1000 IU/ml; IFN�, 1000 IU/ml) � 1,25-(OH)2D3 for 48 h, as described in Materials and Methods. As an additional control, cellswere also exposed to 1,25-(OH)2D3 alone. By setting markers on these histograms, an increase in the percentage of low fluorescent cells inducedby cytokines was noted in region B. These data with corresponding replicates were used to estimate the percentage of cells with depolarizedmitochondria (Table 2).
Riachy et al. • 1,25-(OH)2D3 and Apoptosis in Pancreatic �-Cells Endocrinology, December 2002, 143(12):4809–4819 4811
orescence intensity as measured by cytofluorometry is interpreted as anincrease of mitochondrial permeability transition and a dissipation of��m.
Cell cycle analysis
RINm5F cells were fixed with 70% cold methanol and were treatedwith RNase (10 �g/ml, 30 min, 4 C; Sigma-Aldrich Chemicals). DNAcontent was measured by staining cells with the intercalating DNA dyepropidium iodide (50 �g/ml, 15 min at room temperature; Sigma-Aldrich Chemicals). Apoptotic cells show a low DNA stainability
resulting in a distinct, quantifiable region below the Go/G1 peak andanalyzed by flow cytometry at 620 nm.
Determination of DNA fragmentation
RINm5F cells (1.0 � 106) were trypsinized and collected by centrif-ugation at 200 g for 5 min. After cell lysis, the lysates were centrifugedat 20,000 � g for 10 min. The cytoplasmic fractions were prediluted 1:10with incubation buffer and tested for nucleosomes by Cell Death De-tection ELISA Kit from Roche Molecular Biochemicals. The kit is basedon a quantitative sandwich enzyme-immunoassay using mouse mono-
TABLE 3. Effect of 1,25-(OH)2D3 on DNA content below the G0/G1 peak in cytokine-treated RINm5F cells preincubated or not with NOsynthase inhibitor NOi (5 mM)
Percentage of cells below the G0/G1 peak (Sub G1)
Treatment None(Control)
1,25-(OH)2D3Cytokines
Cytokines �1,25-(OH)2D3
10�8 M
Cytokines �1,25-(OH)2D3
10�6 M10�8 M 10�6 M
Standard (10) 8.9 � 1.9 6.7 � 1.7 8.5 � 2.9 22.8 � 4.7a 13.7 � 2.7b 12.7 � 3.6b
� NOi (3) 7 � 2.3 3.7 � 1.3 2.1 � 0.1 9.1 � 4 4.7 � 1.2 3.9 � 1.3
RINm5F cells were cultured (1 � 105 cells/10-mm well) in 1.5 ml medium and exposed for 48 h to IL-1� (50 IU/ml), IFN-� (1000 IU/ml), andTNF-� (1000 IU/ml) in the absence or presence of 10�8 and 10�6 M 1,25-(OH)2D3. As an additional control, cells were also exposed to 1,25-(OH)2D3alone. Cells were pretreated with RNase and stained with propidium iodide as described in Materials and Methods. Percentages of sub-G1 cells(means � SEM), with the number of observations within parentheses, were determined on DNA frequency histogram.
a P � 0.01, significantly different than the control cells.b P � 0.05, significantly different than cytokine-treated cells.
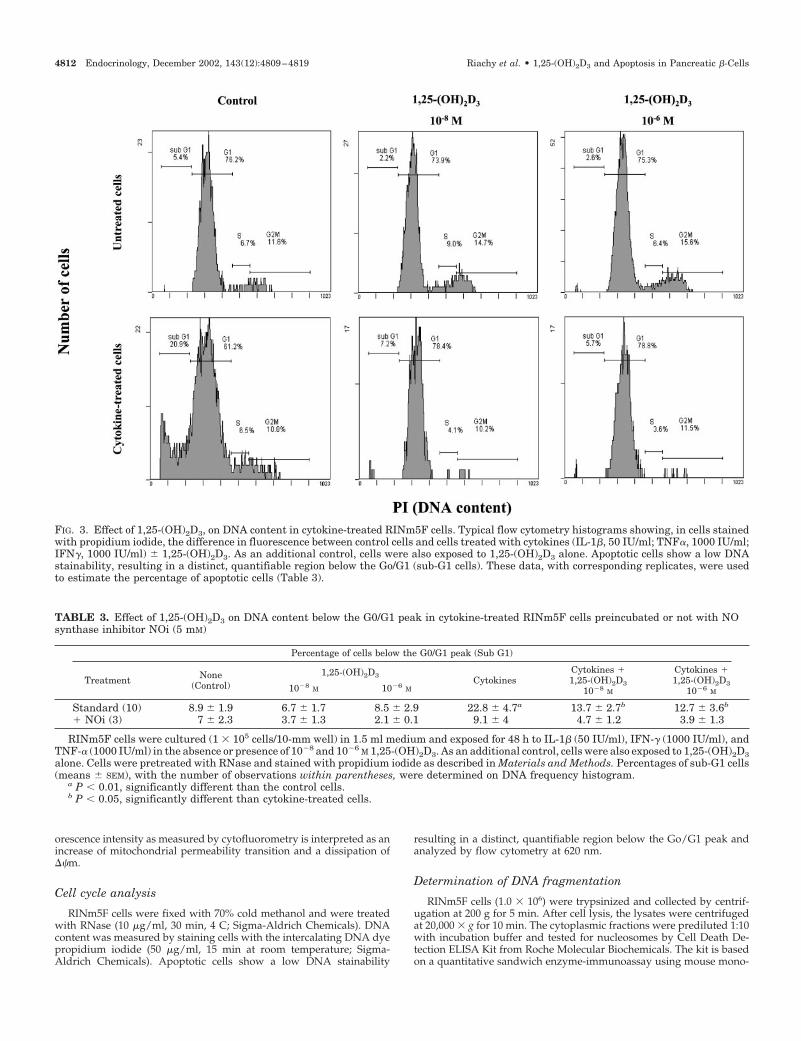
FIG. 3. Effect of 1,25-(OH)2D3, on DNA content in cytokine-treated RINm5F cells. Typical flow cytometry histograms showing, in cells stainedwith propidium iodide, the difference in fluorescence between control cells and cells treated with cytokines (IL-1�, 50 IU/ml; TNF�, 1000 IU/ml;IFN�, 1000 IU/ml) � 1,25-(OH)2D3. As an additional control, cells were also exposed to 1,25-(OH)2D3 alone. Apoptotic cells show a low DNAstainability, resulting in a distinct, quantifiable region below the Go/G1 (sub-G1 cells). These data, with corresponding replicates, were usedto estimate the percentage of apoptotic cells (Table 3).
4812 Endocrinology, December 2002, 143(12):4809–4819 Riachy et al. • 1,25-(OH)2D3 and Apoptosis in Pancreatic �-Cells
clonal antibodies directed against DNA and histones. This allows thespecific determination of mono- and oligonucleosomes in the cytoplas-mic fraction of cell lysates.
Nuclear chromatin staining
Bisbenzimide (Hoechst 33342, 10 �g/ml; France Biochem, Meu-don, France) , which enters cells with intact or damaged membranesand stains DNA, was used to detect differences between normal andapoptotic nuclei in both RINm5F and human islet cells. After stim-ulation with cytokines � 1,25-(OH)2D3, RINm5F cells grown on glasscoverslips were directly stained for 10 min at 37 C, and human isletswere dissociated with trypsin-EDTA, as described (26), and cytocen-trifuged at 700 rpm before being stained. Cells were examined byfluorescence microscopy (excitation 340 –380 nm). Apoptotic cells,identified by the presence of condensed or fragmented nuclei, wereestimated by differential counting of 300 – 400 cells in each experi-mental condition.
Immunohistochemical characterization of IKK�/�
RINm5F and human islet cells were incubated for 48 h with 1,25-(OH)2D3 and then treated with cytokines for 5 min. After stimulation,immunohistolabeling was performed on RINm5F cell cytospins fixedin acetone for 5 min or on fixed, paraffin-embedded human islets.After washings in PBS, fixed cells or deparaffinized sections werepermeabilized for 10 min with 0.1% saponin, and nonspecific siteswere blocked with 10% normal goat serum in PBS. Incubation for 90min at room temperature with the primary antibody (rabbit poly-clonal antibody to IKK�/�, 1:50 dilution; Santa Cruz Biotechnology,Inc., Santa Cruz, CA) was followed by a second incubation withEnVision system, using goat antirabbit immunoglobulin conjugatedto alkaline phosphatase labeled-dextran polymer (DAKO Corp.,Trappes, France) revealed with PhtaloRed from Kirkegaard & Perry(Gaithersburg, MD). Controls included replacing the primary anti-body with PBS containing 1% BSA. Nuclei were counterstained withCarazzi’s hematoxylin.
Nuclear extract preparation and EMSA
RINm5F cells (7.106/condition) were incubated for 48 h with 1,25-(OH)2D3 and then treated with cytokines for 1 h. After stimulation,cells were washed with PBS and nuclear protein fraction extracted,as described (27). Protein concentration was determined using the
Bradford reagent. For the EMSA, a double-stranded oligonucleotidecontaining the �B-binding site: 5-CTTCAGAGGGGACTTTC-CGAGA, was end labeled with [�32P]dATP (3000 Ci-mmol) and T4polynucleotide kinase and used as a probe. A 50-fold molar excess ofnonlabeled oligonucleotide was used as a negative control. The nu-clear fractions (10 �g) were allowed to react with the probe for 30 minon ice in a binding buffer containing 10 mm Tris (pH 8.0), 50 mm KCl,1 mm EDTA, 1 mm dithiothreitol, 10% glycerol, 1 �g polydeoxyi-nosinic acid, and 0.1 ng DNA (15,000 cpm). The samples were thenseparated at 4 C on 5% nondenaturing polyacrylamide gels in 0.25�Tris-borate EDTA. After electrophoresis, gels were dried for autora-diography or PhosphorImager (Molecular Dynamics, Inc., Sunny-vale, CA) quantification.
Cell lysis and Western blotting
After being stimulated, human islet cells were lysed for 30 min on icein 50 �l lysis buffer (20 mm Tris acetate, pH 7.0; 0.27 m sucrose; 1 mmEDTA; 1 mm EGTA; 1 mm sodium vanadate; 50 mm sodium fluoride;1% Triton X-100; 5 mm sodium pyrophosphate; 10 mm �-glycerophos-phate; 1 mm dithiothreitol; 1 mm benzamidine; and 4 �g/ml leupeptin;all reagents were from Fluka, Sigma). The detergent-insoluble materialwas pelleted by centrifugation at 15,000 rpm for 5 min at 4 C. Thesupernatants containing whole-cell lysate were either immediately usedfor Western blotting or stored at �80 C.
For Western blotting, 20 �l (50 �g) protein were added to 20 �l SDSsample buffer (125 mm Tris-HCl, pH 6.8; 25% glycerol; 5% �-mercaptoethanol; and 0.02% bromophenol blue) and boiled for 5 min.SDS-PAGE (10%) was performed and Western blotting carried outaccording to standard protocols (28, 29). Goat polyclonal antibody toA20, 1:200 dilution, (Santa Cruz Biotechnology, Inc.) was used. West-ern blotting detection was achieved using the enhanced chemolumi-nescence plus reagent (ECLplus, Amersham Pharmacia Biotech,Orsay, France).
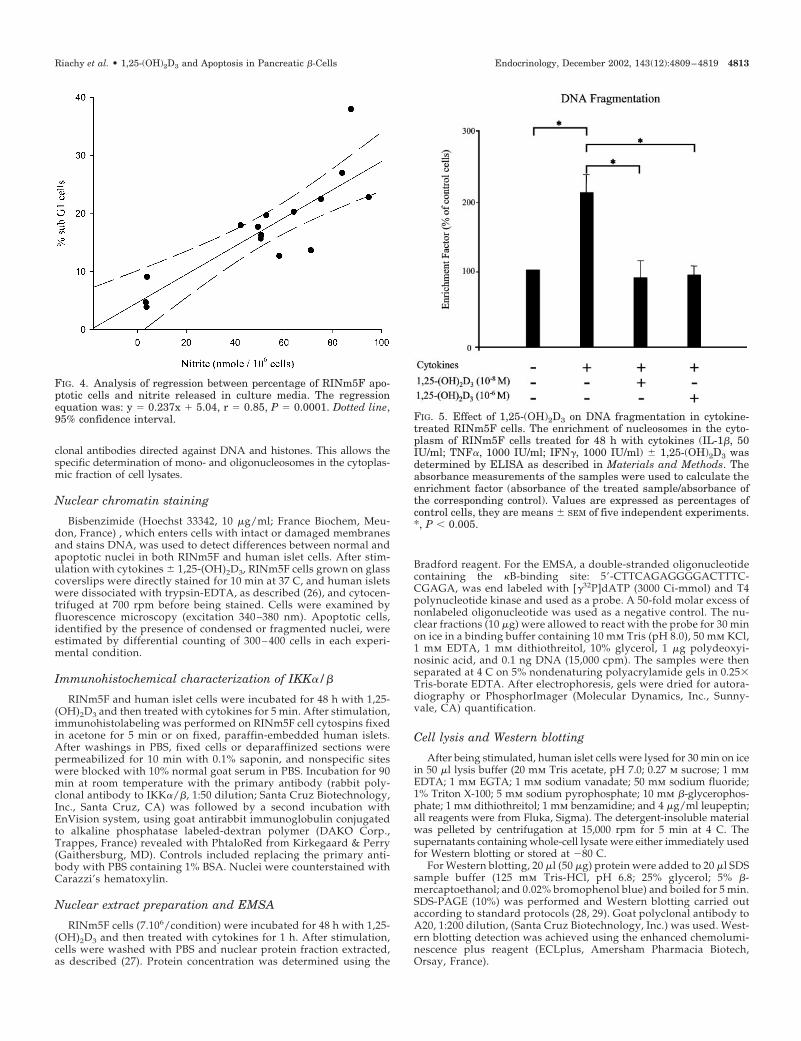
FIG. 4. Analysis of regression between percentage of RINm5F apo-ptotic cells and nitrite released in culture media. The regressionequation was: y � 0.237x � 5.04, r � 0.85, P � 0.0001. Dotted line,95% confidence interval. FIG. 5. Effect of 1,25-(OH)2D3 on DNA fragmentation in cytokine-
treated RINm5F cells. The enrichment of nucleosomes in the cyto-plasm of RINm5F cells treated for 48 h with cytokines (IL-1�, 50IU/ml; TNF�, 1000 IU/ml; IFN�, 1000 IU/ml) � 1,25-(OH)2D3 wasdetermined by ELISA as described in Materials and Methods. Theabsorbance measurements of the samples were used to calculate theenrichment factor (absorbance of the treated sample/absorbance ofthe corresponding control). Values are expressed as percentages ofcontrol cells, they are means � SEM of five independent experiments.*, P � 0.005.
Riachy et al. • 1,25-(OH)2D3 and Apoptosis in Pancreatic �-Cells Endocrinology, December 2002, 143(12):4809–4819 4813
Analysis of A20 mRNA expression
After being treated with 1,25-(OH)2D3 alone, cytokines alone or acombination of both for different time periods, RINm5F cells, and isletcells were lysed in a 1% �-mercaptoethanol containing buffer obtainedfrom an RNA extraction kit (Macherey Nagel, Hoerdt, France). RNA wasextracted as described by the manufacturer and cDNA synthesizedusing random hexamers (Superscript preamplification system for first-strand cDNA synthesis, Life Technologies, Inc.). Semiquantitative,noncompetitive RT-PCR was performed with AmpliTaqGold (Perkin-Elmer Applied Biosystems, Courtaboeuf, France), which allowed simul-taneous coamplification of the housekeeping gene (�-actin) and thetarget gene.
Primer set for A20 was sense: 5-TTTGAGCAATATGCGGAAAGC-3, and antisense: 5-AGTTGTCCCATTCGTCATTCC-3 (Perkin Elmer-Applied Biosystems) resulting in a 479-bp product. PCR was performedwith 1 �l cDNA from each sample in the presence of 200 �m deoxynucle-otide triphosphate, 1.5 mm MgCl2, 5 U AmpliTaq DNA polymerase, and25 pm of each primer. PCR was carried out in a thermal cycler (2400Perkin Elmer-Applied Biosystems) with each cycle consisting of dena-turation at 94 C for 60 sec, annealing at 54 C for 60 sec, and polymer-ization at 72 C for 30 sec. The last PCR step was a final extension at 75C for 7 min. The PCR products were electrophoresed in 2% agarose geland bands were scanned with an integration camera CDD (COHU 4912,
Clara Vision, Orsay, France) and analyzed with GelAnalysts 3.01 FRsoftware (GreyStone-Iconix, MEB Electronique, Paris, France). Band in-tensity was expressed in arbitrary units and A20 expression indexed to�-actin mRNA expression.
Statistical analysis
Data were presented as means � sem. The statistical differencesbetween the groups were determined by ANOVA followed by multiplet tests using the Fisher’s least significant difference test.
ResultsEffect of 1,25-(OH)2D3 on altered metabolic functions incytokine-treated RINm5F cells
As expected, insulin secretion and cell viability were sig-nificantly decreased by cytokines in RINm5F cells. The ad-dition of 1,25-(OH)2D3 resulted in significantly higher valuesdemonstrating a return toward control values (Fig. 1).
Conversely, nitrite released in the culture media wasmarkedly increased by cytokine treatment, and significant
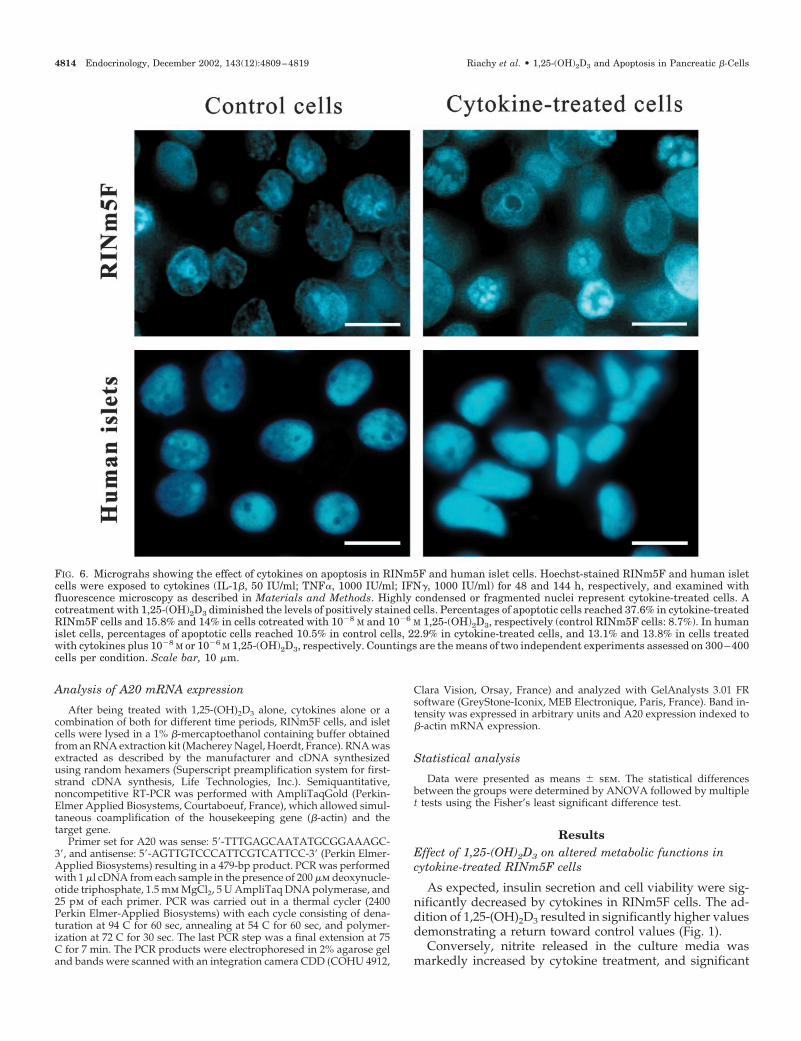
FIG. 6. Micrograhs showing the effect of cytokines on apoptosis in RINm5F and human islet cells. Hoechst-stained RINm5F and human isletcells were exposed to cytokines (IL-1�, 50 IU/ml; TNF�, 1000 IU/ml; IFN�, 1000 IU/ml) for 48 and 144 h, respectively, and examined withfluorescence microscopy as described in Materials and Methods. Highly condensed or fragmented nuclei represent cytokine-treated cells. Acotreatment with 1,25-(OH)2D3 diminished the levels of positively stained cells. Percentages of apoptotic cells reached 37.6% in cytokine-treatedRINm5F cells and 15.8% and 14% in cells cotreated with 10�8 M and 10�6 M 1,25-(OH)2D3, respectively (control RINm5F cells: 8.7%). In humanislet cells, percentages of apoptotic cells reached 10.5% in control cells, 22.9% in cytokine-treated cells, and 13.1% and 13.8% in cells treatedwith cytokines plus 10�8 M or 10�6 M 1,25-(OH)2D3, respectively. Countings are the means of two independent experiments assessed on 300–400cells per condition. Scale bar, 10 �m.
4814 Endocrinology, December 2002, 143(12):4809–4819 Riachy et al. • 1,25-(OH)2D3 and Apoptosis in Pancreatic �-Cells
counteracting effects of 1,25-(OH)2D3 were noted. The ad-dition of NOi during 1 h before stimulation with cytokinesresulted in a near complete abolition of nitrite overproduc-tion (Table 1).
Effect of 1,25-(OH)2D3 on cytokine-induced loss ofmitochondrial transmembrane permeability inRINm5F cells
TMRM is a cationic green fluorochrome that, as a conse-quence of the transmembrane potential of the mitochondria(active proton pump), is taken up by these organelles. Livenonapoptotic cells with an intact plasma membrane andactive (charged) mitochondria concentrate TMRM and ex-hibit strong fluorescence. As cells undergo apoptosis, theirplasma membrane integrity is preserved for some period oftime. However, they lose their ��m and hence the ability tostain with TMRM.
As shown in Fig. 2, a clear increase in weakly fluorescentcells vs. control cells was noted after cytokine treatment.Significant restoration of fluorescence intensity was achieved
by 1,25-(OH)2D3 treatment. In the presence of NOi, the effectof cytokine treatment was less marked and no longer reachedstatistical significance (Table 2).
Effect of 1,25-(OH)2D3 on DNA fragmentation andchromatin condensation in cytokine-treated RINm5F andhuman islet cells
Single-parameter flow cytometric measurement of apo-ptosis was determined on DNA frequency histograms ofRINm5F cells, in which apoptotic cells, containing lessDNA, stain less intensely with the DNA fluorochrome.Apoptotic cells are therefore located to the left of the G1peak and constitute the sub-G1 cells. Figure 3 shows thata significant increase of sub-G1 cells was induced by cy-tokines but significantly reduced by 1,25-(OH)2D3 cotreat-ment (Table 3). The addition of NOi almost completelyabolished the efficiency of cytokines to induce apoptosis,whereas the addition of 1,25-(OH)2D3 seemed to enhancethe process. Interestingly, when we compared the cellsstimulated in these various conditions, a clear correlation
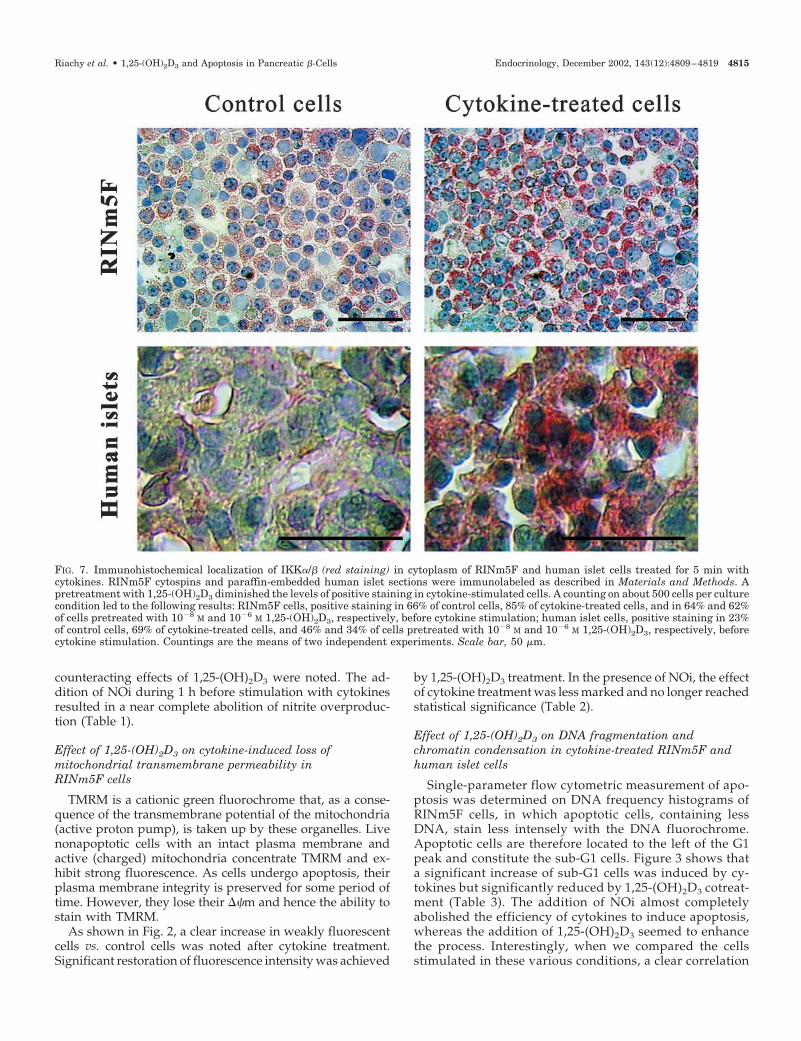
FIG. 7. Immunohistochemical localization of IKK�/� (red staining) in cytoplasm of RINm5F and human islet cells treated for 5 min withcytokines. RINm5F cytospins and paraffin-embedded human islet sections were immunolabeled as described in Materials and Methods. Apretreatment with 1,25-(OH)2D3 diminished the levels of positive staining in cytokine-stimulated cells. A counting on about 500 cells per culturecondition led to the following results: RINm5F cells, positive staining in 66% of control cells, 85% of cytokine-treated cells, and in 64% and 62%of cells pretreated with 10�8 M and 10�6 M 1,25-(OH)2D3, respectively, before cytokine stimulation; human islet cells, positive staining in 23%of control cells, 69% of cytokine-treated cells, and 46% and 34% of cells pretreated with 10�8 M and 10�6 M 1,25-(OH)2D3, respectively, beforecytokine stimulation. Countings are the means of two independent experiments. Scale bar, 50 �m.
Riachy et al. • 1,25-(OH)2D3 and Apoptosis in Pancreatic �-Cells Endocrinology, December 2002, 143(12):4809–4819 4815
(r � 0.85, P � 0.0001) was found between the percentagesof sub-G1 cells and nitrite release in the culture media(Fig. 4).
Cytoplasmic oligonucleosome enrichment, as determinedby ELISA, was also found to be significantly higher incytokine-treated RINm5F cells than in control cells. Resultsobtained with additional 1,25-(OH)2D3 confirmed the returntoward control values as previously measured by flow cy-tometric DNA analyses (Fig. 5).
Cells stained with Hoechst dye were analyzed in bothRINm5F and human islet cells. Apoptotic features such aschromatin condensation, which often aggregates peripher-ally under the nuclear membrane, and extensive surfaceblebbing followed by formation of characteristic apoptoticbodies were particularly obvious (2- to 3-fold increase) after48 h of cytokine treatment for RINm5F and 144 h for humanislet cells. In both cases, cotreatment with 1,25-(OH)2D3 re-sulted in a return toward control levels (Fig. 6).
Inhibition of cytokine-induced increase of I�B kinaseactivity and NF�B translocation by 1,25-(OH)2D3
As shown in Fig. 7, cytokine treatment increased intensityand number of IKK�/�-stained cells in both RINm5F andhuman islets. A pretreatment of cells with 1,25-(OH)2D3
reduced the stimulating effect of cytokines. To furtherdemonstrate the action of 1,25-(OH)2D3 on the cytokine-stimulated NF�B pathway, we examined the translocationof NF�B from the cytosol to the nucleus by EMSA. Asshown in Fig. 8, nuclear extracts from cytokine-treatedRINm5F cells contained higher NF�B-binding activitythan extracts from cells that had been treated for 48 h by1,25-(OH)2D3 before cytokine stimulation.
Modulation of A20 mRNA and protein expression by 1,25-(OH)2D3 in RINm5F and human islet cells
In RINm5F cells, we found, and our results corroboratethose reported by Grey et al. (12), that cytokines induced arapid (15–60 min) expression of the antiapoptotic A20 gene.This expression was, however, transient and gradually de-clined. 1,25-(OH)2D3 alone induced a more belated expres-
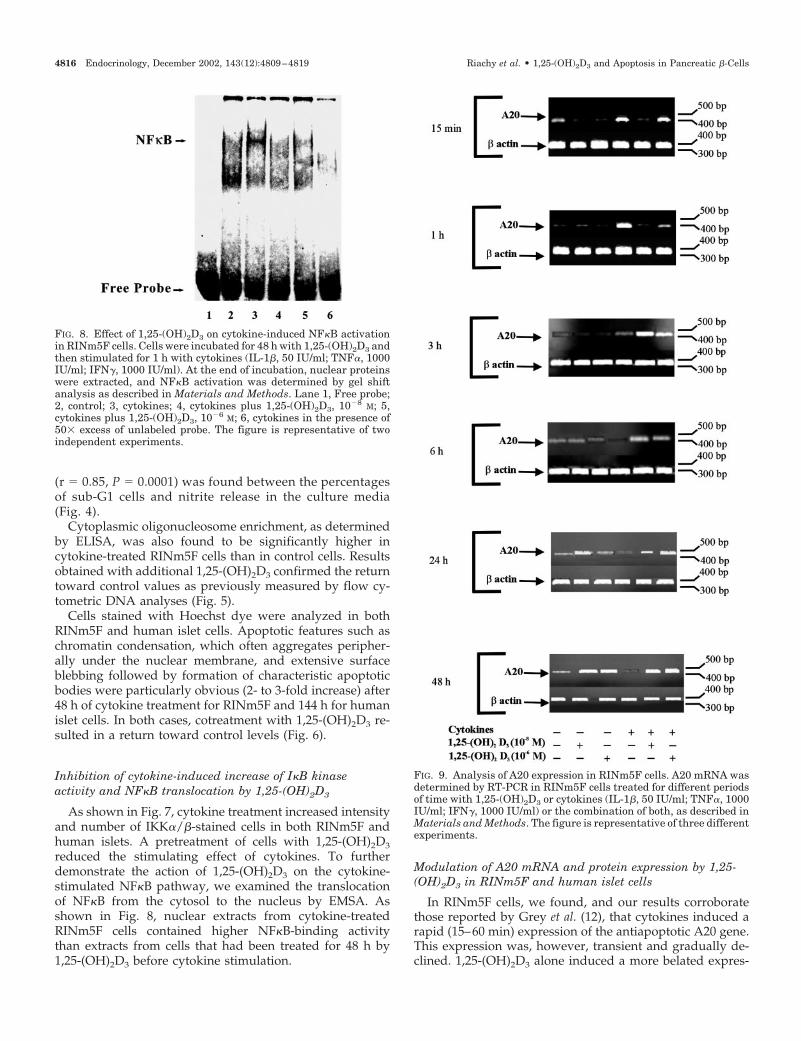
FIG. 8. Effect of 1,25-(OH)2D3 on cytokine-induced NF�B activationin RINm5F cells. Cells were incubated for 48 h with 1,25-(OH)2D3 andthen stimulated for 1 h with cytokines (IL-1�, 50 IU/ml; TNF�, 1000IU/ml; IFN�, 1000 IU/ml). At the end of incubation, nuclear proteinswere extracted, and NF�B activation was determined by gel shiftanalysis as described in Materials and Methods. Lane 1, Free probe;2, control; 3, cytokines; 4, cytokines plus 1,25-(OH)2D3, 10�8 M; 5,cytokines plus 1,25-(OH)2D3, 10�6 M; 6, cytokines in the presence of50� excess of unlabeled probe. The figure is representative of twoindependent experiments.
FIG. 9. Analysis of A20 expression in RINm5F cells. A20 mRNA wasdetermined by RT-PCR in RINm5F cells treated for different periodsof time with 1,25-(OH)2D3 or cytokines (IL-1�, 50 IU/ml; TNF�, 1000IU/ml; IFN�, 1000 IU/ml) or the combination of both, as described inMaterials and Methods. The figure is representative of three differentexperiments.
4816 Endocrinology, December 2002, 143(12):4809–4819 Riachy et al. • 1,25-(OH)2D3 and Apoptosis in Pancreatic �-Cells

sion of A20 because it was obvious after 24 h, when cytokine-stimulated A20 expression had vanished and was evenweaker than in controls. The addition of 1,25-(OH)2D3 tocytokines maintained high levels of A20 (Fig. 9).
In human islet cells, we tested the expression of A20 levelsafter treatment for 48 h with 1,25-(OH)2D3 alone, cytokines,or the combination of both. As observed in RINm5F cells, ourdata confirmed that 1,25-(OH)2D3 up-regulates A20 mRNAand maintains high levels of A20 in cotreated cells, comparedwith cytokine-treated cells. Identical up-regulations werealso observed on the protein level by Western blot (Fig. 10).
Discussion
Islet graft injury by cytokines released from inflammatorycells that infiltrate the transplant is an important mechanismof early islet transplant dysfunction. The emphasis on pres-ervation of islet function has drawn attention to the factorspromoting islet engraftment or minimizing graft cell injury.We have previously shown that 1,25-(OH)2D3 contributed torestore some of the metabolic functions altered by cytokinesin human pancreatic islets in vitro (18). In this study, weanalyzed the effect of 1,25-(OH)2D3 on cytokine-inducedpancreatic �-cell apoptosis. In an attempt to determinethe finer mechanisms implied in the protective effect of1,25-(OH)2D3, we started by focusing on insulin-produc-ing cells to obviate the mechanisms possibly induced byother cells enclosed in the islets. Our data, obtained in theRINm5F cells, a �-cell model currently used for the study ofpancreatic cell death, reproduced the beneficial effects of1,25-(OH)2D3 on cytokine-induced insulin decrease and NOoverproduction, which we previously described in human
islet cells (18), establishing RINm5F as a suitable model. Inthese latter cells, we tested, as we did in human �-cells, twoconcentrations of 1,25-(OH)2D3, a physiological (10�8 m) anda pharmacological one (10�6 m). No significant additionaleffects were observed using the higher concentration, sug-gesting a conceivable action of 1,25-(OH)2D3 via its specifichigh-affinity receptors (30–32). First we studied the possiblealterations of the mitochondrial transmembrane permeabil-ity in RINm5F cells. Our results showed that cytokinesclearly increased the proportion of cells having lost their��m, whereas 1,25-(OH)2D3 exhibited an efficient counter-acting effect at this level. Changes in the permeability of themitochondrial membrane lets certain proteins escape suchas cytochrome C, which activates caspase-3, or other pro-teins like apoptosis-inducing factor implicated in apoptosis(33, 34).
Apoptotic RINm5F and human islet cells were countedbased on nuclear apoptotic features in Hoechst-stained cells.Apoptotic features induced by cytokines were obvious after2 d of treatment in RINm5F cells and 6 d in human islet cells.In both cases, cotreatment with 1,25-(OH)2D3 reducedapoptosis.
These results were firmly confirmed in RINm5F cells eitherby quantitation of DNA strand breaks or measurement ofsub-G1 cell population by flow cytometry. They revealed a 2-to-3-fold increase of apoptosis in cytokine-induced cells. Apo-ptosis was significantly obviated by the addition of 1,25-(OH)2D3. A clear implication of NO in these apoptotic processeswas highlighted by the fact that coincubation of cells with anNO synthase inhibitor reduced the effect of cytokines on theinduction of sub-G1 cells and a clear correlation between thepercentages of sub-G1 cells and nitrite levels was noted.
NO is known to mediate physiological processes via areaction with superoxide or by complexing with a metal atthe active sites of enzymes, and particularly on the Krebscycle, ultimately leading to a severe decrease in glucosemetabolism and ATP production (35). Cytokine-induced NOformation may also cause �-cell toxicity by provoking DNAstrand breaks, thereby activating DNA repair mechanisms,which can further cause cell death through depletion of cel-lular nicotinamide adenine dinucleotide.
The processes leading to the inhibiting effect of 1,25-(OH)2D3 on nitrite production are worth being investigated.Cytokine-induced NO production has been reported to bemediated by NF�B (36). Our study indicated an incidenceof 1,25-(OH)2D3 on activation of this transcription factorby cytokines: When analyzing the NF�B-binding activity incytokine-treated RINm5F cells, we noted, as expected, a clearincrease in the specific band intensity in contrast to the at-tenuation noted when cells had been pretreated with 1,25-(OH)2D3 for 48 h. IKK�/�, the kinases that phosphorylateI�B on the sites that trigger its degradation, appear to becritical for NF�B activation in response to proinflammatorycytokines. Treatment of cells with 1,25-(OH)2D3 before cy-tokine stimulation resulted in an attenuated levels ofIKK�/�, suggesting that 1,25-(OH)2D3 may act upstreamfrom NF�B activation. Such an effect has been demonstratedfor the A20 antiapoptotic protein, reported to exert a cyto-protective effect that was dependent on the abrogation ofcytokine-induced NO production (12). The inhibitory effect
FIG. 10. Analysis of A20 expression in human islet cells. A, Typicalexample of A20 mRNA expression. A20 mRNA was determined byRT-PCR in human islet cells treated for 48 h with 1,25-(OH)2D3 orcytokines (IL-1�, 50 IU/ml; TNF�, 1000 IU/ml; IFN�, 1000 IU/ml) orthe combination of both, as described in Materials and Methods. Thefigure is representative of five independent experiments. B, Typicalexample of Western blot analysis performed on islet cell lysates fromcells treated for 48 h with 1,25-(OH)2D3 or cytokines (IL-1�, 50 IU/ml;TNF�, 1000 IU/ml; IFN�, 1000 IU/ml) or the combination of both, asdescribed in Materials and Methods. A20 appears as an 86-kDa pro-tein band. The figure is representative of two independent experi-ments.
Riachy et al. • 1,25-(OH)2D3 and Apoptosis in Pancreatic �-Cells Endocrinology, December 2002, 143(12):4809–4819 4817

of A20 on cytokine-stimulated NO production was moreovercaused by transcriptional blockade of NO synthase by inhi-bition of the activation of NF�B (37, 38). We therefore in-vestigated A20 induction and confirmed the rapid increaseof its expression after stimulation of RINm5F cells with cy-tokines. Although this effect vanished with time, that ofcotreatment with 1,25-(OH)2D3 maintained high levels ofA20 expression in cytokine-treated cells. 1,25-(OH)2D3 alonewas able to stimulate A20 expression, the effects being no-ticeable after 24 h. In human islet cells, the study of A20expression after 48 h of stimulation with cytokines withand without 1,25-(OH)2D3 clearly demonstrated that 1,25-(OH)2D3 up-regulated both the expression of A20 gene andits protein.
Overexpression of the A20 antiapoptotic gene by means ofadenovirus-mediated gene transfer has recently been shownto protect human and rat islets against cytokine-inducedapoptosis (12). Likewise, inhibition of NF�B activity and NOproduction by a dominant negative inhibitor of NF�B (39) oroverexpression of I�B, an inhibitor of NF�B activity (40) wasdemonstrated to be cytoprotective for cytokine-induced celldeath in the MIN6 �-cell line and human islets, respectively.These studies indicate that cytokine-induced cell death in�-cells involves mechanisms that are largely NF�B and NOdependent. NO reduction achieved by 1,25-(OH)2D3 in ourcytokine-treated cells may be, at least in part, attributable tothe inhibition of NF�B via expression and/or stabilization ofthe antiapoptotic gene A20.
Globally our results clearly indicated a protective effect of1,25-(OH)2D3 on the apoptotic machinery triggered by proin-flammatory cytokines in �-cells. These cytokines released byactivated mononuclear cells, activate �-cells to up-regulateNO production that in turn mediates toxic mechanisms re-sponsible for �-cell dysfunction and apoptosis. In addition toapoptosis mediated by these soluble mediators, NO is knownto induce Fas expression on �-cells, priming them to T-lymphocyte-mediated killing through direct interactions us-ing Fas/Fas ligand systems (41–43), a process we did not dealwith in this study. Induction of Fas gene expression hasrecently been shown to require NF�B in RINm5F cells (20).1,25-(OH)2D3 might therefore exert an additional antiapo-ptotic action by inhibiting Fas via NF�B repression whetherrelated to NO reduction or not (20, 44).
Altogether, treatment of grafts with 1,25-(OH)2D3, as wellas eventually the recipients, provided hypercalcemia isavoided by regional delivery (45) or use of nonhypercalcemicderivatives of 1,25-(OH)2D3 (46), could be beneficial at sev-eral levels: first, because 1,25-(OH)2D3 has known effectson the immunoreactivity of the recipients by decreasingalloreactive T lymphocytes (CD8�) and down-regulatingcytokine-secreting cells (macrophages and monocytes) (47,48); second, because it reduces the vulnerability of graft cellsby decreasing cytokine-induced major histocompatibilitycomplex class I overexpression and IL-6 production, as dem-onstrated earlier in rat and human islets (18, 48); and third,as shown herein, because 1,25-(OH)2D3 may protect �-cellsfrom cytokine-induced death via its antiapoptotic effectmainly mediated by NO reduction and attributable, at leastin part, to induction of the antiapoptotic gene A20.
Acknowledgments
The authors are most grateful to Dr. T. Idziorek (Institut National dela Sante et de la Recherche Medicale Unite 459) for fruitful discussionand Dr. M. d’Herbometz for insulin assays. We also acknowledge S.Belaıch, N. Jouy, and V. Thery for technical assistance and L. Touzet forproofreading. Part of this work was possible thanks to equipment pro-vided by the Institut Federatif de Recherche (IFR 114, Institut Nationalde la Sante et de la Recherche Medicale).
Received April 25, 2002. Accepted August 9, 2002.Address all correspondence and requests for reprints to: B. Vande-
walle, Therapie Cellulaire du Diabete, Institut National de la Sante et dela Recherche Medicale, Equipe de Recherche et d’Innovation Method-ologique 0106, Faculte de Medecine, Place de Verdun, 59045 Lille,France. E-mail: [email protected].
This work was supported by grants from Institut National de la Santeet de la Recherche Medicale, Juvenile Diabetes Foundation, and Etab-lissement Francais des Greffes.
References
1. Davalli AM, Scaglia L, Zangen DH, Hollister J, Bonner-Weir S, Weir GC 1996Vulnerability of islets in the immediate posttransplantation period. Dynamicchanges in structure and function. Diabetes 45:1161–1167
2. Paraskevas S, Aikin R, Maysinger D, Lakey JRT, Cavanagh TJ, Agapitos D,Wang RN, Rosenberg L 2001 Modulation of JNK and p38 stress activatedprotein kinases in isolated islets of Langerhans—insulin as an autocrine sur-vival signal. Ann Surg 233:124–133
3. Berney T, Molano RD, Cattan P, Pileggi A, Vizzardelli C, Oliver R, RicordiC, Inverardi L 2001 Endotoxin-mediated delayed islet graft function is asso-ciated with increased intra-islet cytokine production and islet cell apoptosis.Transplantation 71:125–132
4. Delaney CA, Pavlovic D, Hoorens A, Pipeleers DG, Eizirik DL 1997 Cyto-kines induce deoxyribonucleic acid strand breaks and apoptosis in humanpancreatic islet cells. Endocrinology 138:2610–2614
5. Hoorens A, Pipeleers D 1999 Nicotinamide protects human � cells againstchemically induced necrosis, but not against cytokine-induced apoptosis. Dia-betologia 42:55–59
6. Pavlovic D, Chen MC, Gysemans CA, Mathieu C, Eizirik DL 1999 The roleof interferon regulatory factor-1 in cytokine-induced mRNA expression andcell death in murine pancreatic �-cells. Eur Cytokine Netw 10:403–411
7. Arnush M, Scarim AL, Heitmeier MR, Kelly CB, Corbett JA 1998 Potentialrole of resident islet macrophage activation in the initiation of autoimmunediabetes. J Immunol 160:2684–2691
8. Heitmeier MR, Scarim AL, Corbett JA 1997 Interferon-� increases the sensi-tivity of islets of Langerhans for inducible nitric-oxide synthase expressioninduced by interleukin 1. J Biol Chem 272:13697–13704
9. Carpenter L, Cordery D, Biden TJ 2002 Inhibition of protein kinase C deltaprotects rat INS-1 cells against interleukin-1� and streptozotocin-induced ap-optosis. Diabetes 51:317–324
10. Kwon G, Corbett JA, Rodi CP, Sullivan P, McDaniel ML 1995 Interleukin-1�-induced nitric oxide synthase expression by rat pancreatic �-cells: evidencefor the involvement of nuclear factor �B in the signaling mechanism. Endo-crinology 136:4790–4795
11. You M, Flick LM, Yu D, Feng GS 2001 Modulation of the nuclear factor �Bpathway by Shp-2 tyrosine phosphatase in mediating the induction of inter-leukin (IL)-6 by IL-1 or tumor necrosis factor. J Exp Med 193:101–110
12. Grey ST, Arvelo MB, Hasenkamp W, Bach FH, Ferran C 1999 A20 inhibitscytokine-induced apoptosis and nuclear factor �B-dependent gene activationin islets. J Exp Med 190:1135–1146
13. Penna G, Adorini L 2001 Inhibition of costimulatory pathways for T-cellactivation by 1,25-dihydroxyvitamin D(3). Transplant Proc 33:2083–2084
14. Kajikawa M, Ishida H, Fujimoto S, Mukai E, Nishimura M, Fujita J, TsuuraY, Okamoto Y, Norman AW, Seino Y 1999 An insulinotropic effect of vitaminD analog with increasing intracellular Ca2� concentration in pancreatic �-cellsthrough nongenomic signal transduction. Endocrinology 140:4706–4712
15. Cade C, Norman AW 1987 Rapid normalization/stimulation by 1,25-dihy-droxyvitamin D3 of insulin secretion and glucose tolerance in the vitaminD-deficient rat. Endocrinology 120:1490–1497
16. Overbergh L, Decallonne B, Waer M, Rutgeerts O, Valckx D, Casteels KM,Laureys J, Bouillon R, Mathieu C 2000 1�, 25-dihydroxyvitamin D3 inducesan autoantigen-specific T-helper 1/T-helper 2 immune shift in NOD miceimmunized with GAD65 (p524–543). Diabetes 49:1301–1307
17. Van Etten E, Gysemans C, Verstuyf A, Bouillon R, Mathieu C 2001 Immu-nomodulatory properties of a 1, 25(OH)(2) vitamin D(3) analog combined withIFN� in an animal model of syngeneic islet transplantation. Transplant Proc33:2319
18. Riachy R, Vandewalle B, Belaich S, Kerr-Conte J, Gmyr V, Zerimech F,
4818 Endocrinology, December 2002, 143(12):4809–4819 Riachy et al. • 1,25-(OH)2D3 and Apoptosis in Pancreatic �-Cells

D’Herbomez M, Lefebvre J, Pattou F 2001 Beneficial effect of 1, 25 dihy-droxyvitamin D3 on cytokine-treated human pancreatic islets. J Endocrinol169:161–168
19. Mizuno N, Yoshitomi H, Ishida H, Kuromi H, Kawaki J, Seino Y, Seino S1998 Altered bcl-2 and bax expression and intracellular Ca2� signaling inapoptosis of pancreatic cells and the impairment of glucose-induced insulinsecretion. Endocrinology 139:1429–1439
20. Darville MI, Eizirik DL 2001 Cytokine induction of fas gene expression ininsulin-producing cells requires the transcription factors NF-�B and C/EBP.Diabetes 50:1741–1748
21. Tejedo J, Bernabe JC, Ramirez R, Sopbrino F, Bedoya FJ 1999 NO induces acGMP-independent release of cytochrome c from mitochondria which pre-cedes caspase 3 activation in insulin producing RINm5F cells. FEBS Lett 459:238–243
22. Bernabe JC, Tejedo JR, Rincon P, Cahuana GM, Ramirez R, Sobrino F,Bedoya FJ 2001 Sodium nitroprusside-induced mitochondrial apoptotic eventsin insulin-secreting RINm5F cells are associated with MAP kinases activation.Exp Cell Res 269:222–229
23. Barbu A, Welsh N, Saldeen J 2002 Cytokine-induced apoptosis and necrosisare preceded by disruption of the mitochondrial membrane potential (Delta-psi(m)) in pancreatic RINm5F cells: prevention by Bcl-2. Mol Cell Endocrinol190:75–82
24. Elliott J, Scarpello JH, Morgan NG 2002 Differential effects of genistein onapoptosis induced by fluoride and pertussis toxin in human and rat pancreaticislets and RINm5F cells. J Endocrinol 172:137–143
25. Vandewalle B, Douillard C, Kerr-Conte J, Gmyr V, Riachi R, D’HerbomezM, Pattou F, Lefebvre J 1999 Human pancreatic islet quality control: easyassessment of metabolic functions. Exp Clin Endocrinol Diabetes 107:214–219
26. Lukowiak B, Vandewalle B, Riachy R, Kerr-Conte J, Gmyr V, Belaich S,Lefebvre J, Pattou F 2001 Identification and purification of functional human�-cells by a new specific zinc-fluorescent probe. J Histochem Cytochem 49:519–528
27. Dignam JD 1990 Preparation of extracts from higher eukaryotes. MethodsEnzymol 182:194–203
28. Laemmli UK 1970 Cleavage of structural proteins during the assembly of thehead of bacteriophage T4. Nature 227:680–685
29. Seedorf K, Kostka G, Lammers R, Bashkin P, Daly R, Burgess WH, van derBliek AM, Schlessinger J, Ullrich A 1994 Dynamin binds to SH3 domains ofphospholipase C � and GRB-2. J Biol Chem 269:16009–16014
30. Ishida H, Norman AW 1988 Demonstration of a high affinity receptor for 1,25-dihydroxyvitamin D3 in rat pancreas. Mol Cell Endocrinol 60:109–117
31. Lee S, Clark SA, Gill RK, Christakos S 1994 1, 25-Dihydroxyvitamin-D(3) andpancreatic �-cell function—vitamin-D receptors, gene expression, and insulinsecretion. Endocrinology 134:1602–1610
32. Clark SA, Stumpf WE, Sar M, Deluca HF, Tanaka Y 1980 Target cells for 1,25 dihydroxyvitamin D3 in the pancreas. Cell Tissue Res 209:515–520
33. Daugas E, Susin SA, Zamzami N, Ferri KF, Irinopoulou T, Larochette N,Prevost MC, Leber B, Andrews D, Penninger J, Kroemer G 2000 Mitochon-drio-nuclear translocation of AIF in apoptosis and necrosis. FASEB J 14:729–739
34. Joza N, Susin SA, Daugas E, Stanford WL, Cho SK, Li CYJ, Sasaki T, EliaAJ, Cheng HYM, Ravagnan L, Ferri KF, Zamzami N, Wakeham A, HakemR, Yoshida H, Kong YY, Mak TW, Zuniga P, Flucker JC, Kroemer G, Pen-ninger JM 2001 Essential role of the mitochondrial apoptosis-inducing factorin programmed cell death. Nature 410:549–554
35. Welsh N, Eizirik DL, Bendtzen K, Sandler S 1991 Interleukin-1�-inducednitric oxide production in isolated pancreatic islets requires gene transcriptionand may lead to inhibition of the Krebs cycle enzyme aconitase. Endocrinology129:3167–3173
36. Flodstrom M, Welsh N, Eizirik DL 1996 Cytokines activate the nuclear factor�B (NF-�B) and induce nitric oxide production in human pancreatic islets.FEBS Lett 385:4–6
37. Ferran C, Stroka DM, Badrichani AZ, Cooper JT, Wrighton CJ, Soares M,Grey ST, Bach FH 1998 A20 inhibits NF-�B activation in endothelial cellswithout sensitizing to tumor necrosis factor-mediated apoptosis. Blood 91:2249–2258
38. Heyninck K, Beyaert R 1999 The cytokine-inducible zinc finger protein A20inhibits IL-1-induced NF-�B activation at the level of TRAF6. FEBS Lett 442:147–150
39. Baker MS, Chen XJ, Cao XC, Kaufman DB 2001 Expression of a dominantnegative inhibitor of NF-�B protects MIN6 �-cells from cytokine-inducedapoptosis. J Surg Res 97:117–122
40. Giannoukakis N, Rudert WA, Trucco M, Robbins PD 2000 Protection ofhuman islets from the effects of interleukin-1� by adenoviral gene transfer ofan I�B repressor. J Biol Chem 275:36509–36513
41. Chervonsky AV, Wang Y, Wong FS, Visintin I, Flavell RA, Janeway Jr CA,Matis LA 1997 The role of Fas in autoimmune diabetes. Cell 89:17–24
42. Stassi G, Todaro M, Richiusa P, Giordano M, Mattina A, Sbriglia MS, LoMonte A, Buscemi G, Galluzzo A, Giordano C 1995 Expression of apoptosis-inducing CD95 (Fas/Apo-1) on human B-cells sorted flow-cytometry andcultured in vitro. Transplant Proc 27:3271–3275
43. Stassi G, Demaria R, Trucco G, Rudert W, Testi R, Galluzzo A, Giordano C,Trucco M 1997 Nitric oxide primes pancreatic � cells for fas-mediated de-struction in insulin-dependent diabetes mellitus. J Exp Med 186:1193–1200
44. Zumsteg U, Frigerio S, Hollander GA 2000 Nitric oxide production and Fassurface expression mediate two independent pathways of cytokine-inducedmurine �-cell damage. Diabetes 49:39–47
45. Pourgholami MH, Akhter J, Finlay IG, Morris DL 2000 1, 25-dihydroxyvi-tamin D3 dissolved in lipiodol produces a sustained antiproliferative effect inthe human hepatoblastoma cell line HepG2. Anticancer Res 20:723–727
46. Verstuyf A, Segaert S, Verlinden L, Bouillon R, Mathieu C 2000 Recentdevelopments in the use of vitamin D analogues. Expert Opin Investig Drugs9:443–455
47. Casteels KM, Gysemans CA, Waer M, Bouillon R, Laureys JM, Depovere J,Mathieu C 1998 Sex difference in resistance to dexamethasone-induced apo-ptosis in nod mice: treatment with 1, 25(OH)(2)D-3 restores defect. Diabetes47:1033–1037
48. Van Etten E, Branisteanu DD, Verstuyf A, Waer M, Bouillon R, Mathieu C2000 Analogs of 1, 25-dihydroxyvitamin D3 as dose-reducing agents for clas-sical immunosuppressants. Transplantation 69:1932–1942
Riachy et al. • 1,25-(OH)2D3 and Apoptosis in Pancreatic �-Cells Endocrinology, December 2002, 143(12):4809–4819 4819





























