Embed Size (px)
DESCRIPTION
Citation preview

Viral Genetics
Introduction to Viruses:
A virus is a biological agent that reproduces inside the cells of living hosts. When infected by a virus, a host cell is forced to produce many thousands of identical copies of the original virus, at an extraordinary rate. Unlike most living things, viruses do not have cells that divide; new viruses are assembled in the infected host cell. Over 2,000 species of viruses have been discovered. Notable human diseases caused by viruses include SARS, influenza and hepatitis C
A virus consists of two or three parts: all viruses have genes made from either DNA or RNA, long molecules that carry the genetic information; all have a protein coat that protects these genes; and some have an envelope of fat that surrounds them when they are not within a cell. Viruses vary in shape from the simple helical and icosahedral to more complex structures. Viruses are about 100 times smaller than bacteria, and it would take 30,000 to 750,000 of them, side by side, to stretch to 1 centimeter
Life-cycle of Viruses:
When a virus infects a cell, the virus forces it to make thousands more viruses. It does this by making the cell copy the virus's DNA or RNA, making viral proteins, which all assemble to form new virus particles.
There are six basic, overlapping stages in the life cycle of viruses in living cells.
Attachment : the binding of the virus to specific molecules on the surface of the cell. This specificity restricts the virus to a very limited type of cell. For example, the human immunodeficiency virus (HIV) infects only human T cells, because its surface protein, gp120, can only react with CD4 and other molecules on the T cell's surface. Plant viruses can only attach to plant cells and cannot infect animals. This mechanism has evolved to favor those viruses that only infect cells in which they are capable of reproducing.
Penetration follows attachment; viruses penetrate the host cell by endocytosis or by fusion with the cell.
Uncoating: happens inside the cell when the viral capsid is removed and destroyed by viral enzymes or host enzymes, thereby exposing the viral nucleic acid.
Replication of virus particles is the stage where a cell uses viral messenger RNA in its protein synthesis systems to produce viral proteins. The RNA or DNA synthesis abilities of the cell produce the virus's DNA or RNA.

Assembly takes place in the cell when the newly created viral proteins and nucleic acid combine to form hundreds of new virus particles.
Release occurs when the new viruses escape or are released from the cell. Most viruses achieve this by making the cells burst, a process called lysis. Other viruses such as HIV are released more gently by a process called budding.
Viral Genetics:
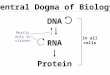
Viral genetics, the study of the genetic mechanisms that operate during the life cycle of viruses, utilizes biophysical, biological, and genetic analyses to study the viral genome and its variation. The virus genome consists of only one type of nucleic acid, which could be a single or double stranded DNA or RNA. Single stranded RNA viruses could contain positive-sense (+RNA), which serves directly as mRNA or negative-sense RNA (-RNA) that must use an RNA polymerase to synthesize a complementary positive strand to serve as mRNA. Viruses are obligate parasites that are completely dependent on the host cell for the replication and transcription of their genomes as well as the translation of the mRNA transcripts into proteins. Viral proteins usually have a structural function, making up a shell around the genome, but may contain some enzymes that are necessary for the virus replication and life cycle in the host cell.
Classification of Viruses:
Viruses are classified in two families depending on whether they have RNA or DNA genomes and whether these genomes are double or single stranded. Further subdivision into types takes into account whether the genome consists of a single RNA molecule or

many molecules as in the case of segmented viruses. Four types of bacteriophages are widely used in biochemical and genetic research. These are the T phages, the temperate phages typified by bacteriophage lambda, the small DNA phages like M13, and the RNA phages. Animal viruses are subdivided in many classes and types. Class I viruses contain a single molecule of double stranded DNA and are exemplified by adenovirus, simian virus 40 (SV40), herpes viruses, and human papillomaviruses. Class II viruses are also called parvoviruses and are made of single stranded DNA that is copied in to double stranded DNA before transcription in the host cell. Class III viruses are double stranded RNA viruses that have segmented genomes which means that they contain 10-12 separate double stranded RNA molecules. The negative strands serve as template for mRNA synthesis. Class IV viruses, typified by poliovirus, have single plus strand genomic RNA that serves as the mRNA. Class V viruses contain a single negative strand RNA which serves as the template for the production of mRNA by specific virus enzymes. Class VI viruses are also known as Retroviruses and contain double stranded RNA genome.
RNA Viruses:An RNA virus is a virus that has RNA (ribonucleic acid) as its genetic material. This nucleic acid is usually single-stranded RNA (ssRNA) but may be double-stranded RNA (dsRNA).
Single stranded RNA viruses:
Single stranded RNA viruses can be further classified according to the sense or polarity of their RNA into negative-sense and positive-sense RNA viruses. Some are positive in that they have a "sense" strand of RNA (coded information about how to build proteins)

as their genetic material. Positive-sense viral RNA is identical to viral mRNA and thus can be immediately translated by the host cell. And other RNA viruses are negative in that they have an "antisense" strand (the paired opposite of the coded information). Negative-sense viral RNA is complementary to mRNA. Negative-strand or antisense-strand RNA viruses are as opposed to positive-strand or sense-strand RNA viruses. As such, purified RNA of a positive-sense virus can directly cause infection though it may be less infectious than the whole virus particle. Purified RNA of a negative-sense virus is not infectious by itself as it needs to be transcribed into positive-sense RNA.
Definition of Negative-strand RNA virus:
Also known as an antisense-strand RNA virus, a virus whose genetic information consists of a single strand of RNA that is the negative or antisense strand which does not encode mRNA (messenger RNA) and is complementary to it. Therefore it must be copied into the complementary plus-sense mRNA before proteins can be made. Thus, besides needing to code for an RNA-dependent RNA-polymerase, these viruses also need to package it (polymerase) in the virion so that they can make mRNAs upon infecting the cell. The positive-sense RNA molecule then acts as viral mRNA, which is translated into proteins by the host ribosome. The resultant protein goes on to direct the synthesis of new virions, such as capsid proteins and RNA replicase, which is used to produce new negative-sense RNA molecules. Examples of negative-strand RNA viruses include influenza virus (orthomyxovirus), measles viruses (paramyxoviruses), and rabies virus (rhabdovirus).
RHABDOVIRUSES (RHABDOVIRIDAE)
Example: Rabies virus. The most intensively studied member is vesicular stomatitis virus. RNA is single stranded, is negative (minus) sense, and codes for 5 proteins. The entire life cycle occurs in the cytoplasm, RNA polymerase and RNA modification enzymes are virally-coded and present in the virion itself. There is no early/late division of gene expression.
Attachment, penetration and uncoating:
The virus adsorbs to cell surface. G (Glycoprotein) is the attachment protein which binds to a receptor on the host cell surface. The attached virus is taken up by endocytosis. The membrane of the virus fuses with the endosome membrane (the acid pH of endosome is important because the G protein needs to be exposed to acidic pH before it can facilitate fusion). As a result of fusion of the viral membrane with the endosome membrane, the nucleocapsid is released into cytoplasm.
Transcription
'Transcription' is used in this context to refer to synthesis of mRNAs. Complete uncoating of the nucleocapsid is not necessary for transcription - the virion RNA polymerase can

copy virion RNA when it is in the nucleocapsid form. This is an advantage in that genomic RNA is therefore somewhat protected from ribonucleases. There is one monocistronic mRNA for each of the five virally coded proteins. The mRNAs are capped, methylated, and polyadenylated. Since this is a cytoplasmic, negative-sense RNA virus, the enzymes for mRNA synthesis and modification are packaged in the virion.
Translation
Messenger RNAs are translated on host ribosomes and all five viral proteins are made at the same time. There is no distinction between early and late functions.
RNA replication
RNA replication is the process by which new copies of genome-length RNAs are made. RNA replication occurs in the cytoplasm and is carried out by the viral RNA polymerase. The full length plus strand is coated with nucleocapsid protein as it is made (mRNAs are not coated with this protein, which would interfere with the host protein translation machinery). The new positive strand is copied into full length minus strand, which is also coated with nucleocapsid protein as it is made.
New negative strands may now:
be used as templates for the synthesis of more full length plus strands. be used as templates for the synthesis of more mRNAs. be packaged into virions. .
Assembly
The newly made RNAs and polymerase complexes are first packed into nucleocapsid containing G protein and then they are modified in envelop by ribosome on rough endoplasmic reticulum.
Definition of Positive-strand RNA virus:
Also known as a sense-strand RNA virus, a virus whose genetic information consists of a single strand of RNA that is the positive (or sense) strand which usually behaves as mRNA (messenger RNA). This mRNA can be translated immediately upon infection of the host cell, producing a single protein which is modified by host and viral proteins to form the various proteins needed for replication... Replication in positive-strand RNA viruses is via a negative-strand intermediate. Examples of positive-strand RNA viruses include poliovirus (picornavirus), Coxsackie virus, and echovirus. Togaviruses, flaviviruses.

PICORNAVIRUSES (PICORNAVIRIDAE)
These are small (28nm), naked icosahedral viruses (pico=very small). The RNA is single-stranded, plus sense, polyadenylated. It functions as mRNA immediately upon infection. The entire life cycle occurs in the cytoplasm and there is no division into early and late gene expression of the genome. Example is poliovirus.
Adsorption and penetration
A viral protein recognizes a receptor on the host cell membrane (this is important in the tropism of virus). It seems that binding to the receptor alters capsid structure in some way, a channel forms across the cell membrane and the RNA is released into cytoplasm. The mRNA is now available for translation.
Synthesis of viral proteins
Poliovirus virion RNA functions as an mRNA but does not have the methylated cap structure typical of eukaryotic mRNAs - it has a "ribosome landing pad" (known as the internal ribosome entry site or IRES) which enables ribosomes to bind without having to recognize a 5' methylated cap structure. Picornaviruses often interfere with host cell methylated cap recognition. Most host cell translation is cap-dependent, so this inhibits a lot of host protein synthesis but not viral protein synthesis - one way in which these viruses can modify the host cell to their advantage. The mRNA is translated into a single polypeptide (polyprotein), which is cleaved. The cleavages occur before translation is complete ( i.e. on the nascent / growing chain) and are carried out by virally coded proteases. Some of these proteases can work even while part of the polyprotein.
Products of cleavage include: An RNA polymerase (replicase), Structural components of the virion, and Proteases.5
RNA replication
We now have newly made viral proteins to support replication.
1. Viral RNA polymerase copies plus-sense genomic RNA into complementary minus-sense RNA using VPg (or precursor containing VPg), Viral RNA polymerase (replicase), and Certain Host proteins. VPg may act as a primer for RNA synthesis, this would explain why it is at the 5' end of all newly synthesized RNA molecules.
2. New minus sense strands serve as template for new plus sense strands. Again, RNA polymerase and VPg are needed. VPg is linked to the 5' ends of the new plus sense strands (again, it probably functions as a primer).
The new plus strand has three alternative fates:

It may serve as a template for more minus strands. It may be packaged into progeny virions. It may be translated into polyprotein (In this case VPg is usually removed prior to
translation).
Assembly
When sufficient plus-sense progeny RNA and virion proteins have accumulated, assembly begins. Particles assemble with VPg-RNA inside and 3 proteins in the capsid [VP0, 1 and 3]. VP0 is then cleaved to VP2 and VP4 as the virions mature and these mature virions are infectious. Virions are released following cell lysis. Excess capsids are formed and inclusion bodies may be seen in the cytoplasm.
Single stranded RNA Segmented Viruses:
The term for RNA viruses that explicitly excludes retroviruses is ribovirus. Their RNAs are called segmented because each segment is coding for a particular gene & trait.
Segmented Negative Strand Viruses:
Examples: Orthomyxoviruses , Bunyaviruses (include Hantavirus genus), Arenaviruses
ORTHOMYXOVIRUSES (ORTHOMYXOVIRIDAE)
It is also called Influenza virus. There are three groups of influenza virus: A, B and C. Influenza A virus is most intensively studied and influenza A and B are the most important in human disease. Influenza viruses are pleomorphic virions (that is, they vary in shape). They have negative-sense, single-stranded RNA and an RNA genome that is SEGMENTED. There are eight RNA segments in influenza A. The nucleocapsid is helical. Virions contain RNA polymerase packaged within the virus particle. These viruses are enveloped and have two membrane glycoproteins:
HA - hemaglutinin - This is the attachment and fusion protein NA - neuraminidase - This is important in release. It removes sialic acid from
proteins of the virus and the host cell.
Adsorption and penetration:
The virus adsorbs to receptors on the cell surface and is internalized by endocytosis. At acidic pH of an endosome, HA undergoes a conformational change and fusion occurs. Nucleocapsids are released to cytoplasm and are transported into the nucleus. The mRNA synthesis and replication of viral RNA occurs in the nucleus. This is very unusual for an RNA virus.

Double Stranded RNA Virus:
The double-stranded (ds)RNA viruses represent a diverse group of viruses that vary widely in host range (humans, animals, plants, fungi, and bacteria), genome segment number (one to twelve), and virion organization (T-number, capsid layers, or turrets). Members of this fascinating group include the rotaviruses, renowned globally as the commonest cause of gastroenteritis in young children, and bluetongue virus, an economically important pathogen of cattle and sheep.Viruses with dsRNA genomes are currently grouped into six families: Reoviridae, Birnaviridae, Totiviridae, Partitiviridae, Hypoviridae, and Cystoviridae. Of these six families, the Reoviridae is the largest and most diverse in terms of host range.The double stranded RNA viruses replicate in the cytoplasm, and their replication is monocistronic(mRNA that can encode only one polypeptide per RNA molecule). Example is the Reovirus.
REOVIRUS FAMILY (REOVIRIDAE)
The Reovirus family includes:
the members of the reovirus genus the members of the rotavirus genus the members of the orbivirus genus Colorado tick fever virus
Reoviruses have icosahedral symmetry and a multiple layered capsid (inner and outer capsid). The RNA is double stranded. There are 10-12 segments (depending on the genus of the Reovirus family). There are some significant differences in the life cycle of members of the reovirus family and of the rotavirus family. Due to their clinical importance in humans, focus is on rotaviruses.
ROTAVIRUSES(rota = wheel (from appearance of virions in the electron-microscope))

Adsorption, penetration and uncoating:
It is still not clear what exactly what happens in-vivo. There appears to be a need for a protease to remove some of the outer layer of the capsid and to generate an "intermediate sub-viral particle" (ISVP) before the virus can enter the cytoplasm. In vivo, the ISVPs are probably generated by protease digestion in the GI tract. A viral attachment protein is then exposed on the ISVP, probably at the vertices, and binds to host cell receptors. The activated ISVP enters the cytoplasm directly or via endocytosis. In the cytoplasm, the virion RNA is copied by the viral RNA polymerase while still in a nucleocapsid that has fewer proteins associated with it than are associated with the ISVP or the virion.
Transcription and translation:
Double stranded RNA does not function as an mRNA and so the initial step is to make mRNA (transcription). The mRNAs are made by virally-coded RNA polymerase packaged in the virion. The RNA is capped and methylated by virion packaged enzymes. It is then extruded from the vertices of the capsid. The mRNAs are translated and the resulting viral proteins assemble to form an immature capsid. The mRNAs are packaged into the immature capsid and are then copied within the capsid to form double stranded RNAs (It is not known how the virus ensures that each particle gets one copy of the 11 different mRNAs) .More mRNAs are now made by the newly formed immature capsids.
Assembly
More proteins are made and eventually the immature capsids bud into the lumen of the endoplasmic reticulum. In doing so, they acquire a transient envelope which is lost as they mature. This is a very odd feature of the rotaviruses. Then the release probably occurs via cell lysis.
Double-stranded RNA Segmented viruses:
Double-stranded RNA viruses (Group III) contain from one to a dozen different RNA molecules, each of which codes for one or more viral proteins. They include: orthoreoviruses, rotavirus, phytoreoviruses, and bluetongue virus
Structure of Orthoreoviruses ( reoviruses ):
Reoviruses are non-enveloped and characterized by concentric capsid shells that encapsidate a segmented dsRNA genome. In particular, reovirus has eight structural proteins and ten segments of dsRNA. A series of uncoating steps and conformational changes accompany cell entry and replication. The icosahedral structures of intact virions, infectious subviral particles that bind to cell surface receptors and core particles, which mediate RNA transcription, ths is how reovirus completes its lifecycle.

Structure and Functions of the Orthoreovirus σ3 Protein:
Protein s3 serves a number of distinct roles in the orthoreovirus life cycle. It forms the outermost layer of the reovirus particle, it imparts significant environmental stability to virions. Virion σ3 also plays a critical role as a determinant of cell entry, as it must be degraded from particles during their activation for membrane penetration. Early in infection σ3 is thought to serve a regulatory function through its capacity to bind dsRNA and interfere with dsRNA-activated innate immune pathways.
Retroviruses:
Retroviruses comprise a large and diverse family of enveloped RNA viruses defined by common taxonomic denominators that include structure, composition, and replicative properties. The virions are 80–100 nm in diameter, and their outer lipid envelope incorporates and displays the viral glycoproteins
The shape and location of the internal protein core are characteristic for various genera of the family. The virion RNA is 7–12 kb in size, and it is linear, single-stranded, non-segmented, and of positive polarity. The hallmark of the family is its replicative strategy which includes as essential steps reverse transcription of the virion RNA into linear double-stranded DNA and the subsequent integration of this DNA into the genome of the cell. Retroviruses contain RNA as the hereditary material in place of the more common DNA. In addition to RNA, retrovirus particles also contain an enzyme called reverse transcriptase (or RTase) that can both copy minus strand DNA from genomic RNA, catalyze the synthesis of a complementary plus DNA strand. The resulting double stranded DNA is integrated in the host chromosome and is transcribed by the host's own machinery. The resulting transcripts are either used to synthesize proteins or produce new viral particles. These new viruses are released by budding, usually without killing the host cell. Both HIV and HTLV viruses belong to this class of viruses.

Retroviruses are broadly divided into two categories—simple and complex—distinguishable by the organization of their genomes. The retrovirus family is split up into 7 genera: the Alpha retroviruses, the Beta retroviruses, Gamma retroviruses, Delta retroviruses, Epsilon viruses (all of which used to be classified as one genus, the oncoviruses), the Lentiviruses (which includes HIV) and the Spumaviruses.
All retroviruses contain three major coding domains with information for virion proteins: gag, which directs the synthesis of internal virion proteins that form the matrix,
the capsid, and the nucleoprotein structures; pol, which contains the information for the reverse transcriptase and integrase
enzymes; and env, from which are derived the surface and transmembrane components of the
viral envelope protein.
An additional, smaller, coding domain present in all retroviruses is pro, which encodes the virion protease. Simple retroviruses usually carry only this elementary information, whereas complex retroviruses code for additional regulatory non-virion proteins derived from multiply spliced messages.
Virus Replication
A detailed summary of the steps of virus replications is as follows:
1) Receptor binding and membrane fusion.2) Internalization and un-coating of virion.
3) Reverse transcription of RNA to form double-stranded DNA
4) Entry of viral DNA, into host nucleus.
5) Integration of viral DNA into cellular DNA to form the provirus.
6) Transcription of provirus to form viral RNA.
7) Splicing of viral RNAs and export from host nucleus.
8) Translation of viral RNAs to form viral proteins.
9) Assembly of virion and packaging of viral RNA genome.
10) Budding and release of new virions.
11) Proteolytic processing of proteins and virion maturation.
Human Retroviruses:

Human T-Cell lymphotropic Virus Types I and II:
HTLV- I and II is included in the retroviridae family because of its nucleotide sequence and genome structure. Morphologically they are named the primate T-cell leukemia/lymphoma viruses. They are biologically distinct from the human immunodeficiency viruses in the lentivirus genus. Clinical manifestations of HTLV-I are linked with the development of adult T-cell leukemia/lymphoma and a progressive neurological disease called HTLV-I associated myelopathy. HTLV-II is also linked with leukemia and neurologic disease cases as well.
Human Immunodeficiency Viruses(HIV)
Human immunodeficiency viruses are part of the lentivirus genus. It includes the disease subtypes HIV-1 and HIV-2, the third and fourth human retroviruses discovered. HIV enters the host cell through the CD4 molecule and chemokine receptor as a dual receptor system. The biology of HIV-1 has been highly researched due to the pressing concerns of an HIV global pandemic and push for vaccine and treatment development.
DNA Viruses:Dna virus is a virus that has DNA (deoxyribonucleic acid) as its genetic material. They are usually Large, Icosahedral, enveloped in Lipoproteins, Do not have polymerase enzymes, and cause Latent infection. Examples include Poxviruses, Herpesviruses, hepadnaviruses, Hepatitis b.

HERPESVIRUSES :
Larger virions than adenoviruses(180 - 200nm). Larger genome (three to five times) than adenoviruses. Linear, double-stranded DNA, Enveloped, icosahedral virus (this means that lipid solvents readily inactivate these viruses)
Adsorption and penetration
Many herpesviruses, including herpes simplex virus, can fuse directly with the plasma membrane (results in partial uncoating). Such fusion with the plasma membrane has implications for both the virus and the host cell. Among these are:i) Since the fusion protein is active at physiological pH, if it is inserted into the host cell membrane during the virus growth cycle, the infected cell can potentially fuse with other cells and form syncytia. ii) The viral membrane leaves a "footprint" in the cell plasma membrane and this is a possible clue that the cell is infected. Capsids are transported towards the nucleus and the DNA passes into the nucleus (probably via nuclear pores).
Early phase
Early transcription (the mRNAs made during this phase are the alpha and beta mRNAs). Herpes viruses use host RNA polymerase. However, a virion tegument protein (VP16) enters the nucleus upon infection and is important as part of the transcription factor complex recognized by the host RNA polymerase. The virus uses host mRNA modification enzymes.
Initially, alpha-mRNAs are transcribed. These are the immediate early mRNAs and are exported to the cytoplasm and translated into alpha-proteins. The α-proteins translated in the cytoplasm are transported into nucleus where they enable the beta-promoters to be used by the host RNA polymerase.
Beta-mRNAs are transcribed by the host RNA polymerase again. (Beta-genes are still "early" since they are transcribed prior to DNA synthesis. Sometimes alpha-genes are called "immediate early" and beta-genes are called "early"). Beta proteins are involved in gene expression regulation. They decrease alpha-gene expression and are needed for gamma gene expression. They are also involved in various aspects of DNA synthesis; for example, herpes beta -genes code for a variety of proteins including DNA polymerase, DNA binding proteins, thymidine kinase, ribonucleotide reductase etc.
Since these beta proteins are virally-coded and not host-coded enzymes, they are potentially weak links in the virus life cycle and thus promising targets for viral chemotherapy

Late phase
DNA replication
Herpesviruses code for several proteins, in addition to the DNA polymerase, that are needed for DNA replication. The precise mechanism of DNA replication is not known. DNA replication is accompanied by a lot of recombination. The replicated DNA is present as long concatameric molecules (tandem repeats of the genome linked head-to-tail). These are cleaved to genome-size lengths when DNA is packaged into the virion.
Late transcription:
By definition, late transcription occurs after DNA replication. Gamma mRNAs are made and are translated in the cytoplasm. Gamma proteins are predominantly structural. There is decreased expression of beta genes in the late stage. This is probably due to down-regulation of transcription of beta genes, by gamma proteins. In herpes viruses there is no apparent organization of the genome into blocks for either early or late transcription.
Assembly
Assembly occurs in the nucleus. A capsid is formed and the DNA enters the capsid. The capsids acquire an envelope by budding through areas of the inner nuclear membrane which have viral membrane proteins inserted into them. These areas have tegument proteins associated with the inner face of the inner nuclear membrane. The virus envelope then fuses with the outer nuclear membrane and the de-enveloped nucleocapisid is delivered into the cytoplasm, where it acquires a more mature tegument. It then becomes re-enveloped by budding into Golgi-derived vesicles and is then released.
The late protein required for transcription of immediate early mRNAs in the next round of infection is packaged in the virion.
Evolution of Viruses:Viral genomes undergo genetic change by mutation, either spontaneous or induced mutation, and by recombination. Recombination may be either intramolecular or, among viruses with divided genomes, by reassortment.
Mutations:
These arise naturally during viral replication: e.g. due to errors by the genome-replicating polymerase or as a result of the incorporation of tautomeric forms of the bases. Mutation in RNA viruses may be extremely rapid because RNA is a less thermodynamically stable molecule than is DNA, many of the mutations which give rise to the diversity seen with the HIV virus is not a result of the fact that the virus uses an RNA genome, but of the

nature of the polymerase enzyme that the virus uses. There is no proof-reading mechanism for RNA polymerases, as there is for DNA polymerases. This situation is compounded in the retroviruses, for there is no proof-reading mechanism for the reverse transcriptase either. Most of these mutations result in non-viable phenotypes. Whether the genetic changes lead to emergence of an altered phenotype depends on natural selection, which may occur within the infected cell, during spread of virus in the body, or the transmission of the virus from one host to the next. Mutations can manually b introduced by Agents acting directly on bases, e.g. nitrous acid, Agents acting indirectly, e.g. base analogs which mis-pair more frequently than normal bases or even by Agents such as UV light or X-rays
Exchange of Genetic material
This involves breaking of covalent bonds within the nucleic acid, exchange of genetic information, and reforming of covalent bonds. This kind of break/join recombination is common in DNA viruses or those RNA viruses which have a DNA phase (retroviruses). The host cell has recombination systems for DNA. If a virus has a segmented genome and if two variants of that virus infect a single cell, progeny virions can result with some segments from one parent, some from the other. This is an efficient process - but is limited to viruses with segmented genomes - so far the only human viruses characterized with segmented genomes are RNA viruses
For the practicing physician, virus evolution may appear to be an academic matter, because evolutionary changes usually occur over a time scale that is long compared with human life. However, sometimes genetic changes in viruses may occur rapidly as a result of evolutionary pressure. For instance, the highly virulent myxoma virus introduced into Australia to control the wild rabbit population evolved in a few years to a much more attenuated strain, enabling infected rabbits to survive for weeks instead of days, thereby increasing chances for transmission. Among influenza viruses, antigenic variation evolves toward decreased affinity for preexisting neutralizing antibodies during the course of an outbreak. Because survival of a virus depends largely on its ability to circulate among its natural hosts, natural selection tends to favor those viruses that are better transmitted (usually less virulent), have a lower susceptibility to antibody, and have a greater ability to persist. Also, the ability of the virus to produce reactions that promote excretion, such as coughing and sneezing in respiratory infections and diarrhea in many enteric infections, is likely to be retained.
The mutations and evolution in viruses is supported by the human population explosion. Evolution allows influenza to remain potentially the most important of all human viral diseases. Genetic reassortment and exchange of influenza viruses between humans and animals, producing antigenic shift, periodically introduce new viruses to the human population; mutation and selection, producing antigenic drift, accounts for year-to-year variations in influenza A subtypes.

References: www.Stanford.edu www.ncbi.nlm.nih.gov http://pathmicro.med.sc.edu/mhunt/dna1.htm http://science.jrank.org/pages/7190/Viral-Genetics.html