Embed Size (px)
Citation preview

Sex Differences in a C. elegans Sensory Behavior
by
Kyung Hwa Lee
Submitted in Partial Fulfillment
of the
Requirements for the Degree
Doctor of Philosophy
Supervised by
Professor Douglas S. Portman
Interdepartmental Graduate Program in Neuroscience
School of Medicine and Dentistry
University of Rochester
Rochester, New York
2009

ii
To my family whose love and prayer has carried me through this endeavor

iii
Curriculum Vitae
The author was born in Seoul, Korea on August 5, 1980. She attended
Handong Global University from March 1999 to December 2002, and graduated with
a Bachelor of Science degree in February 2003. She came to the University of
Rochester in the Summer of 2003 and began graduate studies in the Interdepartmental
Graduate Program in Neuroscience. She pursued her research in “Sex differences in
C. elegans olfactory behavior” under the direction of Professor Douglas S. Portman
and received the Master of Science degree from the University of Rochester in March
2006. A part of the thesis is published in the journal: Current Biology 17, 1858–
1863, November 6, 2007.

iv
Acknowledgements
I am grateful to all members of the Portman lab for their challenging hard
work, encouragements, and accountability to discuss science and to share life. In
particular, I thank Dr. Renee Miller, William Mowrey and a former member, Dr.
Adam Mason for insightful suggestions and discussions throughout years. Their
valuable technical contributions to my thesis work are noted within the text.
Foremost I offer my gratitude to my advisor, Dr. Douglas S. Portman, who
has supported me throughout my graduate studies with his patience, knowledge, and
encouragement. He was always available for discussions in person and online. One
simply could not wish for a better advisor.
I would like to thank my thesis committee members: Dr. Robert S. Freeman
and Dr. Kathy W. Nordeen for their time and advice. I also appreciate Dr. Oliver
Hobert for his time and kindness to serve on my committee and deliver valuable
suggestions and insights.
I thank Dr. White and Dr. Jorgensen for sharing oxEx862 and oxEx863 and for
communicating unpublished data, Dr. Schwarz and Dr. Horvitz for generously
providing ceh-30(n4289) mutants, and Dr. Bargmann for helpful suggestions.
I cannot end without thanking my family, on whose constant love and prayer I
have relied throughout my journey at the Academy.

v
Abstract
Sex differences in the structure and function of the nervous system exist
throughout the animal kingdom. Together with sex-biases in neurological diseases,
this highlights the importance of studying how sexual differentiation modifies neural
circuits and function. Taking advantage of the unique strengths of the nematode C.
elegans, we explore how “neural sex”, the sexual state of a given neuron established
by cell-intrinsic sex determination, regulates the function of the “core” neural
circuitry composed of neurons common to both sexes. To ask how neural sex
influences behavior, we have examined olfaction, well-described in the C. elegans
hermaphrodite but previously unstudied in the male. Using a novel assay involving
the simultaneous presentation of two attractants, we have observed characteristic and
distinct sex differences in olfactory preference behaviors. These sex differences were
prominent before sexual maturation and did not require the gonad or germline,
suggesting that core neural circuitry itself may be the cellular focus of sexually
different shared behavior. To address this directly, we switched the sexual state of
subsets of core neurons by cell-type specific expression of sexual regulators. We
found that the neural sex of even a single sensory neuron, AWA, can determine the
sexual phenotype of olfactory preference, indicating that AWA itself possesses
sexually different functional properties. Moreover, at least some of these functional
properties arise through sex differences in the expression of the odorant receptor
ODR-10, providing a molecular mechanism for the generation of sexually different
shared sensory function. This work has revealed a novel pathway for bringing about

vi
sex differences in the function of shared neural circuitry, and may shed light on the
nature of sexual dimorphisms in the vertebrate nervous system.

vii
Table of Contents
Chapter 1 Introduction 1
1 The general problem of sex differentiation in the nervous system 1
Sex, brain, and behavior 1
Sex differences are prominent in the neuroanatomy for sex-specific 2 behaviors
Sex differences also occur in the areas of the brain not relevant to 2 reproductive behavior
Sex differences are observed even in common behaviors non-relevant to 3 reproduction
Sex-biases are prevalent in the nature and/or incidences of neurological 3 diseases
Sex differences in behavioral symptoms of some neurological disorders 4
2 Sex hormones and the sex of the brain 5
Activity of sex hormones has been thought to regulate sex differences in 5 the brain
Some sexually different behaviors are not explained by sex hormones 5
3 Chromosomal sex also control properties of neural structures and 6 behaviors
Sex differences in neurological diseases are not all explained by the 6 activity of sex hormones
Evidences of sex hormone-independent sexual differentiation in the 6 vertebrate system
Cell-intrinsic sex regulators generate sex-specific behaviors in 7 invertebrate organisms
The pathway of chromosomal sex regulation on the properties of the 8 neural circuit is largely unknown

viii
4 Neural circuits and behaviors 8
Some gene expression differences change behaviors 8
Sex-specific behaviors are generated by sexually different 9 interpretations of the same sensory stimuli as a result of differences in gene expression
Common behaviors between sexes or between species are modified 9 by gene expression differences to confer sex difference or species difference
A complete diagram of the neural circuit for any complex behavior 10 is generally not described
5 C. elegans as a system to study sex differences in shared behaviors 10
C. elegans is an ideal model for neuroscience and for studying sexual 10 dimorphism in the nervous system
The cell-intrinsic sex determination pathway regulates all known somatic 11 sex differences in C. elegans
The C. elegans core neural circuitry has molecular sex difference 12
Sex differences in the C. elegans core neural function 12
6 C. elegans olfactory behaviors 17
The hermaphrodite olfactory system is well characterized in its structure 17 and function
Olfactory neural circuit possess molecular properties for behavioral 21 plasticity
Chapter 2 Neural sex modifies the function of a C. elegans sensory circuit 22
1 Introduction 22 2 Materials and Methods 23 3 Results 29
C. elegans exhibit significant sex difference in olfactory behaviors 29
Each sex displays distinct and characteristic olfactory preferences 30

ix
Sexually different olfaction is not the secondary effect of male-specific 34 behaviors
The male-specific CEM neurons do not have a primary role in the 37 sexually different shared sensory function
Gonad signaling is not necessary for sex difference in olfaction 37
Sexual differences in olfaction are prominent before sexual maturation 38
Neural sex determines the sex phenotype of a common sensory function, 41 olfactory preference
Pan-neural sex-transformation 45
Sex-transformation on sensory, interneuron and motor neurons 45
4 Discussion 49
Why are there sex differences in C. elegans olfaction? 49
Developmental regulation of sexually different olfactory behaviors 51
How does sex modify olfaction? 52
Neural Sex regulation on Behaviors 53
Chapter 3 Neural sex modifies the properties of a single sensory neuron to 55 generate sexually different olfactory behaviors
1 Introduction 55 2 Materials and Methods 56 3 Results 58
Sexually different olfaction arises through neural sex modification on 58 sensory neurons
A single sensory neuron, AWA, generates sexually different olfactory 62 preference
Neural sex regulation on sexually different olfaction is a property of the 65 C. elegans olfactory circuit
Neural sex modifies a target gene in AWA neurons to bring about sex 69

x
difference in olfaction
4 Discussion 80
Core sensory circuitry controls a sex difference in olfactory preference 80
A single sensory neural-switch between hermaphrodite and male olfaction 80
Neural sex regulates an effector gene critical for the function of a neural 82 circuit
Appendix 1 Potential sex difference in AWA connectivity 85
Chapter 4 Discussion 87
The control of sex differences in a C. elegans sensory behavior 87
Insights on sexual dimorphisms in neurological diseases 89
References 91
Appendix 2 Strains 102

xi
List of Tables
Table A2.1 Nematode strains 102

xii
List of Figures
Figure 1.1 C. elegans Sexes 13
Figure 1.2 Somatic sex determination in C. elegans 14
Figure 1.3 The C. elegans core nervous system 15
Figure 1.4 Sexually dimorphic gene expressions in the C. elegans core 16 nervous syste
Figure 1.5 Does the C. elegans core nervous system mediate sexually 19 dimorphic behaviors?
Figure 1.6 The C. elegans olfactory neural circuit is a part of the core 20 nervous system
Figure 2.1 Single odorant assay 25
Figure 2.2 Olfactory preference assay 27
Figure 2.3 Male olfaction is significantly different to hermaphrodite 31 olfaction
Figure 2.4 Each sex has distinct and characteristic olfactory preferences 33
Figure 2.5 Sexually different olfactory preferences are generated neither 35 by sex-specific behaviors nor by structures
Figure 2.6 Sex differences in olfaction precede sex-specific differentiation 39
Figure 2.7 The terminal sex regulator, tra-1, controls sex difference 43 in olfactory preference
Figure 2.8 Sex-transformation of the nervous system 44
Figure 2.9 Neural sex-transformation 46
Figure 2.10 Neural sex determines the sex phenotype of olfactory preference 48
Figure 2.11 Sexually different properties of the sensory neurons control 50 the sex phenotype of olfactory preference
Figure 3.1 Neural sex governs sex difference in olfactory preference 60

xiii
Figure 3.2 A neural sex mechanism modifies properties of the sensory 61 neurons to bring about sexually dimorphic olfactory preference
Figure 3.3 The sexual state of a single sensory neuron, AWA, is 63 sufficient to impart sex differences in olfactory preference
Figure 3.4 How AWA neurons are modified by neural sex? 66
Figure 3.5 Neural sex regulates sex differences in both AWA and AWC 67 olfactory preference behaviors
Figure 3.6 Sexually different mechanisms underlying responses to 71 hermaphrodite AWA odorants
Figure 3.7 How do male AWA neurons respond to hermaphrodite AWA 73 odorants?
Figure 3.8 Male odr-10 expression is significantly reduced compared to that 75 of hermaphrodites
Figure 3.9 ODR-10 expression is sex-specifically regulated: strong ODR-10 78 in hermaphrodites but weak ODR-10 in males
Figure 3.10 Neural sex in AWA neurons regulates sexually dimorphic ODR-10 79 expression
Figure 3.11 Neural sex generates sex difference in a target gene for sexually 84 different common sensory function
Figure A1.1 AWA connectivity is similar between sexes 86

xiv
List of Symbols
ANOVA Analysis of variance
ATRX X-linked α thalassaemia/mental retardation syndrome
AWA Amphid Wing type neuron A sense volatile attractants
AWB Amphid Wing type neuron B sense repellents
AWC Amphid Wing type neuron C sense volatile attractants
bu 2-butanone (hermaphrodite AWC odorant)
bz benzaldehyde (hermaphrodite AWC odorant)
C. elegans Caenorhabditis elegans
C.I. Chemotaxis Index
CA Male-specific ventral cord motor neurons
cAMP cyclic Adenosine MonoPhosphate
CEM Male-specific cephalic sensory neurons
CP Male-specific tail interneurons
da diacetyl (hermaphrodite AWA odorant)
DIC Differential Interference Contrast
DM doublesex and mab-3
Ex Extrachromosomal array
fem-3 FEMinization of XX and X0 animals

xv
FMR1 fragile X mental retardation 1
GFP Green Fluorescence Protein
glp-1 abnormal germlilne proliferation defective
glr-1 glutamate receptor family (AMPA)
H Hermaphrodite
him-5 mutant strain for the high incidence of male progenies
HSN Hermaphrodite-specific serotonergic motor neurons
iaa isoamylalcohol (hermaphrodite AWC odorant)
Is Integrated strain
JARID1C Jumonji, AT rich interactive domain 1C
M Male
MECP2 methyl CpG binding protein 2 (Rett syndrome)
odr odorant response abnormal
OPI Olfactory Preference Index
osm-5 OSMotic avoidance abnormal
Pglr-1 promoter specific for interneurons, head motor neurons
Podr-7 AWA neural specific promoter
Posm-5 promoter specific for sensory neurons
pkd-2 marks CEM neurons in the male head regions
Prab-3 pan-neural promoter

xvi
PUFA Poly Unsaturated Fatty Acid
py pyrazine (hermaphrodite AWA odorant)
RFP Red Fluorescence Protein
RnA Ray neuron type A
RnB Ray neuron type B
SEM Standard Errors of the Mean
SNB-1 C. elegans Synaptobrevin
srd-1 male specifically expressed 7-transmembrane receptor
srj-54 male specifically expressed 7-transmembrane receptor
tph-1 marks Herm.-specific HSNs, male-specific CP neurons
tra transformer
VC Hermaphrodite-specific cholinergic motor neurons

1
Chapter 1 Introduction
1 The general problem of sex differentiation in the nervous system
Sex, brain, and behavior
The sex of the brain has been historically discussed in terms of the relationships
between sexual dimorphism in the hypothalamus, sex hormones, and sex behaviors
[Levine, 1966]. Investigations in the past decade, however, have revealed abundant
evidence that sex of the brain is not limited to the neural machinery of sexual activity.
Rather, sex of the brain affects many different areas of brain and behaviors. Many
areas of brain responsible for sensory processing, cognition, pain and stress response,
and reward are sexually differentiated. Sex differences in a variety of behaviors
common to both sexes, not related to reproductive behaviors, are well observed
throughout the animal kingdom and in humans. Regarding these sex differences, the
action of sex hormones during development and in adulthood has been thought to
exclusively set up the neural substrates of sex differences in behavior. Any sex
differences in a common behavior were suggested to result from sexual dimorphism
in the structure of brain areas: significant sex differences in the size of nucleus of sub-
cortical structures and/or in the connectivity of neurons (i.e., differences in cell
numbers, thickness of cortical layers, numbers of spines, and electrophysiological
properties). However, it has been revealed that anatomically common structures
between sexes also give rise to sex differences in some behaviors and even in
neurological diseases. Furthermore, some sexually different behaviors are not
explained by sex hormones and recent studies reveal that cell-intrinsic sex
differentiation is a significant regulator of the sex of the brain. Therefore, some
behavioral sex differences may arise through sex differences in the molecular
properties of common neural circuitry. A better understanding of these issues will
shed light on how biological sex interacts with developmental control to impart
plasticity to neural circuitry to bring about sex differences in behaviors.

2
Sex differences are prominent in the neuroanatomy for sex-specific behaviors
In mammals, reproductive behaviors are significantly different by sex. In some cases,
neural substrates that may underlie these behaviors have been identified. For
example, the rat hypothalamus was first described to have sexual dimorphism [Gorski
et al., 1978]. The SDN-POA (sexually dimorphic nucleus of preoptic area) in the
hypothalamus regulates male copulatory behaviors. It was revealed that lesions in the
entire anterior preoptic area eliminate male courtship behavior and lesions restricted
to SDN-POA significantly slow acquisition of male copulatory behavior. The volume
of SDN-POA is about seven times larger in male rats than in female rats [Morris et
al., 2004]. Another well-characterized sexually dimorphic area of the rat brain is the
posterodorsal medial amygdala (MePd). This area receives pheromone stimuli and
main olfactory inputs, which triggers male responses to female pheromones, male
dominance social behavior, and female relations with litters. Lesions in MePd result
in severe deficits in those behaviors. Consistent with obvious sex difference in the
function of MePd, its volume is about 1.5 times larger in male rats than in female rats
and its neurochemical characteristics are also sexually different [Cooke et al., 2005].
Together, SDN-POA and MePd are representative sexually dimorphic areas in the
brain, offering good model systems to study the relationship between the brain sex
and the sex-specific behaviors.
Sex differences also occur in the areas of the brain not relevant to reproductive
behavior
Sex differences in the brain are not limited to areas dedicated to reproductive
behavior. Sex differences also exist in many ‘cognitive’ regions such as
hippocampus, amygdala, and neocortex [Juraska, 1991]. The hippocampus, the most
well-known structure regulating learning and memory, is also sexually dimorphic in
its structure and function [Juraska et al., 1985; McEwan et al, 2000]. Its normalized
size compared to the whole brain is larger in women than in men. Specifically, the
volume of the CA1 region, the number of pyramidal cells in CA1, and the neuronal

3
density of the dentate gyrus are larger in males [Madeira et al., 1995]. The
neurochemical systems within the hippocampus are also sexually different.
Furthermore, the reactivity of hippocampus to stressful situations in both rats and
humans are sexually different. In the amygdala of rat pups, the basomedial nucleus
displays sexually different changes in serotonin receptor expressions upon separation
from mother rodents [Ziabreva et al., 2003]. The amygdala of human brain also
exhibits sex difference in the hemispheric lateralization of amygdala function for
memory with emotional events [Cahill et al, 2004]. Together, these suggest that
sexual dimorphisms in these ‘cognitive’ regions bring about sex differences even in
behaviors common to both sexes. The mechanistic relationships between these
anatomical and functional sex differences are largely unclear.
Sex differences are observed even in common behaviors non-relevant to
reproduction
Recent findings reveal that sex differences in common behaviors are apparent
between sexes. Behaviors fundamental to both sexes were different in mammalian
model systems. Those behaviors encompass sensorimotor behaviors, hippocampal,
striatal learning strategies, and drug-addiction [Dewing et al., 2006; Korol et al.,
2004; Becker et al., 1999]. Furthermore, emotion, memory, vision, hearing, feeding,
face processing, pain perception, navigation, and the effects of stress in the brain were
also sexually differentiated in animals and humans. However, due to the complexity
of the neural circuitry and the behavior itself, the mechanisms that give rise to sex
differences even in common behaviors are not well understood.
Sex-biases are prevalent in the nature and/or incidences of neurological diseases
Clinical observations have reported that there are significant sex-biases in variety of
aspects of diseases affecting the nervous system. These sex-biases affect the nature
of disease, its incidence, and recovery ability [Cahill et al., 2006]. First,
schizophrenia displays sex differences in both nature and incidences. For example,

4
the morphology of the diseased brain areas is sexually different in that the normally
sex-biased ratio of the size of amygdala to that of the orbitofrontal cortex is increased
in men with psychosis but decreased in women with psychosis [Gur et al., 2004].
This disease occurs about 2.7 times more often in men than in women. Second,
autism displays the extreme sex-bias in its incidences in that males get autism four
times more often than females [Swaab et al., 2003]. Third, Parkinson’s disease (PD)
reveals sex differences in its symptoms and drug responses. For example, male PD
patients go through more serious rigidity than female PD patients and drug-related
dyskinesias are more frequently observed in female PD patients [Brann et al., 2007].
These sex differences in the neurological diseases are suggested to result from
sexual dimorphisms in the brain itself [Cahill, 2006; Wizemann et al., 2001; Swaab et
al., 2003]. Substrates for sex-bias in brain diseases may be apparent sex differences
in the structure and function of the affected brain regions. In addition, extensive sex
differences in many neurotransmitter systems arise as important possible molecular
substrates of sex-bias in neurological disorders. In particular, unipolar depression,
which occurs more often in females than in males, may be understood better by
studying the functional role of the higher mean rate of serotonin synthesis in males
than in females [Nishizawa et al., 1997].
Sex differences in behavioral symptoms of some neurological disorders
Certain neurological diseases such as mental retardation, neurodegenerative diseases,
and neuropsychiatric disorders seriously impair a plethora of behavioral symptoms in
humans. The affected behaviors in these diseases are influenced by gender, brining
about sexually different behavioral symptoms in patients. For example, the
APOE*E4 allele associated with increased risk of Alzheimer’s Disease (AD) is linked
with significantly more serious memory disruption in female than in male AD
patients [Fleisher et al., 2005]. In addition, the deficits in social behaviors of the
valproic acid-induced rat autism model were similar to behavioral symptoms of
autism patients and were present only in male rats [Schneider et al., 2008]. This

5
suggests that autism, with a strong (4:1) male-biased incidence, can at least in part be
attributed to higher susceptibility of male behavioral substrates to disease risk factors.
Therefore, investigations on how common behaviors between sexes are also sexually
differentiated will improve our understanding of the pathology underlying behavioral
symptoms of brain diseases and our chances of developing treatments.
2 Sex hormones and the sex of the brain
Sex hormones regulate sex differences in the brain
Scientists first demonstrated that testosterone masculinizes the brain by exposing
female guinea pigs to testosterone in utero, which permanently hindered normal
female reproductive behaviors in adulthood [Phoenix et al., 1959]. Organizing sexual
differentiation of the brain by sex hormones during development was thought to be
the major mechanism by which sex-specific behaviors are generated [Cooke et al.,
1998]. In addition to the role of sex hormones during development, the activity of
sex hormones in adulthood gives rise to sex-specific behaviors. For example, the
treatment of the testosterone in the adult female canary brain at least temporarily
generates male-specific courtship songs [Goldman et al., 1983].
Some sexually different behaviors are not explained by sex hormones
The activity of circulating sex hormones, particularly testosterone and estrogen, has
been thought to govern all known sexual differentiation of the brain and behaviors.
However, findings over the past several decades have revealed that that is not the
case. One major finding is that the rat dopaminergic neurons display sex differences
even before sexual hormones are active [Reisert and Pilgrim, 1991]. In addition,
many studies on sex differences in behaviors show that sex hormones cannot account
for all those differences [Arnold, 2004]. For example, male aggression and female
parental behaviors are also at least in part attributed to the role of sex chromosome

6
complement other than the testis-determining gene Sry on the Y chromosome
[Gatewood et al., 2006]. Furthermore, social interactions are sexually differentiated
by the sex chromosomal complement [McPhie-Lalmansingh, 2008].
3 Chromosomal sex also control properties of neural structures and behaviors
Sex differences in neurological diseases are not all explained by the activity of
sex hormones
Sex-biases in many aspects of brain diseases are thought to result from the activity of
sex hormones during development and in adult. The simple notion was that sex
hormones establish sexually different neuroanatomy and molecular substrates more
susceptible to certain neurological diseases in one sex than in the other. Furthermore,
sex hormones, especially estrogen, give rise to sex differences in post-injury brain
damage and recovery. However, recent clinical reports reveal that sex differences
exist in the developing brain, at times when the circulating sex hormonal activity is
absent. For example, both post-ischemic brain damage and pattern of apoptosis were
significantly different in two sexes of the postnatal day 7 rats at which sexual
maturation is not established [Renolleau et al., 2008; Hurn et al., 2005; Edwards,
2004]. Thus, these findings highlight the importance of studying how the cell-
intrinsic sex determination mechanisms give rise to sexual differentiation of the brain.
Evidences of sex hormone-independent sexual differentiation in the vertebrate
system
Consistent with the insufficiency of sex hormonal activity to bring about sexual
dimorphisms in the brain, behavior, and diseases, several lines of evidence reveal that
the sex chromosomes themselves harbor information regulating neural sex
differentiation and the generation of sexual dimorphisms. The ground-breaking
finding of a rare gynandromorphic zebra finch that has two sex phenotypes
throughout its body shows that sex chromosomal gene expression correlates with

7
sexually lateralized brain phenotype irrespective of the gonadal sex [Agate et al.,
2003]. Furthermore, a series of experiments support the idea that the cell-intrinsic
sex determination mechanisms control sexual differentiation in the brain in a variety
of species [Arnold, 2004; Carruth et al., 2002; Dewing et al., 2006; Gahr et al.,
2003]. In particular, Dewing et al., revealed that the Y chromosome-linked male-
determining gene Sry may directly control the higher expression of tyrosine
hydroxylase in dopaminergic neurons of the adult male substantia nigra system,
imparting sex differences to the sensorimotor behaviors. Together, these studies
reveal that cell-intrinsic sex regulators may directly regulate molecular sex
differences to bring about behavioral sex differences. However, very little is known
about the regulatory mechanisms that might link chromosomal sex to differences in
neural development or function.
Cell-intrinsic sex regulators generate sex-specific behaviors in invertebrate
organisms
Invertebrate systems have been an advantageous tool to study sex differences in
neurobiology and behavior since their nervous systems and behaviors are relatively
simple and tractable. Studies on sexual differentiation in the nervous system utilizing
invertebrate systems, in particular, Drosophila, revealed how the cell-intrinsic
mechanisms differentiate sex-specific neurons and generate sex-specific behaviors -
i.e., behaviors present in one sex and unnecessary for the survival of the other sex -
for reproduction. In Drosophila, a sex determination gene, fruitless (fru), has been
shown to control male-specific behaviors by specifying masculine properties in
multiple types of fly neurons, suggesting that fru might be a master regulator of male
behavioral circuitry [Billeter et al., 2006; Manoli et al., 2006; Vrontou et al., 2006;
Datta et al., 2008]. In addition, possibly in a smaller portion of male neurons, another
sex determination gene, doublesex (dsx), also contributes to the development of
masculine properties [Kimura et al., 2008; Rideout et al., 2007].

8
The pathway of chromosomal sex regulation on the properties of the neural
circuit is largely unknown
Altogether, in addition or in parallel to the sex hormonal control over all known sex
differences in animals, the cell-intrinsic sex regulators are suggested to establish
changes in the properties of the neural circuitry itself to generate sex differences in
behaviors. However, due to the complexity of the neural circuitry for a given
behavior, the lack of knowledge on downstream effectors of the sex chromosomes
and target genes of those effectors, the specific mechanism through which the sex
chromosomal signaling modifies the molecular/cellular/physiological properties of
neural circuitry is not described.
4 Neural circuits and behaviors
Some gene expression differences change behaviors
Understanding how behaviors are generated by the neural circuitry is a major problem
in neuroscience. In some cases, it has been suggested that behavior can be hard-wired
into the corresponding neural circuitry. However, the surprising plasticity in behavior
suggests that the neural circuitry can modify the behavior it generates to meet the
needs of animals in survival and reproduction throughout generations. Consistent
with this idea, studies in different systems have shown that some gene expression
changes in neural circuitry give rise to differences in behaviors between sexes,
species, and even in an individual animal.
Sex-specific behaviors are generated by sexually different interpretations of the
same sensory stimuli as a result of differences in gene expression
Mouse pheromone responses require the vomeronasal organ (VNO) and the main
olfactory epithelium (MOE). A recent study reveals that mouse TRPC2, an ion
channel specific to the VNO, is responsible for sex-specific processing of the same

9
sensory stimuli (mouse pheromones) to generate sex-specific responses according to
the sex of the individual [Kimchi et al., 2007]. This study reveals that the same
sensory stimuli trigger sexually different downstream signaling mediated by TRPC2
in the VNO sensory neurons to bring about activation of sex-specific behavior.
In Drosophila, male flies execute male-specific courtship song mediated by
the pattern generator. By light-activated channel expression in the motor circuits,
song-like wing movement and sound were generated by both sexes. However,
authentic male courtship song was only revealed in normal males and in females with
expression of the male form of fruitless (fru, a master sex regulator of fly neurons).
This indicates that fru sets up male characteristics to process the same stimuli in a
male way and brings about the normal male singing [Clyne et al., 2008]. Together,
these studies suggest that changes in gene expression in an otherwise common neural
circuit between sexes can generate sexually different behaviors.
Common behaviors between sexes or between species are modified by gene
expression differences to confer sex difference or species difference
Fiddler crabs display sexually different responses to food-related cues: the
chemosensory neurons of male fiddler crabs are less sensitive to low concentrations
of food cues than those of female fiddler crabs. Changes in the expression of cAMP
signaling between male and female crabs underlie generation of sex differences in
food-sensation [Weissburg et al., 2001]. In rodents, different expression levels of the
oxytocin receptor (OTR) and vasopressin 1a receptor (V1aR) in the ventral forebrain
reward circuitry results in species differences in social behaviors. High levels of
these molecules contribute to monogamous social behavior of the prairie vole
whereas low levels of them generate polygamous behavior of the montane and
meadow voles [Hammock et al., 2006]. Therefore, these studies reveal that gene
expression changes bring about modification even in common behaviors between
sexes or between species. Since common behaviors are presumably generated by the
same neural circuitry, these findings surprisingly suggest that the activity or

10
physiology of the appropriate neural circuit for these behaviors may be made different
by changes in gene expression. Furthermore, these findings indicate that the extent of
plasticity in neural circuitry to bring about differences in behaviors must be quite
broad.
A complete diagram of the neural circuit for any complex behavior is generally
not described
Growing evidence, as described above, reveal that discrete changes in gene
expression modify the properties of neural circuitry to generate sex differences or
species differences in behaviors. However, due to the limited knowledge on the
neural circuit sufficient or necessary for any given behavior, elucidation of the tight
control of changes in molecular components on behaviors is challenging in vertebrate
systems. By utilizing relatively simple model systems, this problem could be
alleviated and understanding the relationship of gene, neural circuit, and behavior can
be expedited.
5 C. elegans as a system to study sex differences in shared behaviors
C. elegans is an ideal model for neuroscience and for studying sexual
dimorphism in the nervous system
C. elegans has a relatively simple nervous system, harboring about 300 neurons. The
nervous system (Figure 1.3) mediates a plethora of behaviors that can be categorized
into two classes: one, shared behaviors fundamental to both sexes mediated by the
“core” nervous system (neural circuitry comprising common neurons between sexes)
and the other, sex-specific behaviors for reproduction generated by sex-specific
neural circuitry. With sophisticated genetic tools and the complete neuronal wiring
diagram for adults of one sex (hermaphrodites), C. elegans provides a unique
opportunity to study how neural circuits generate behavior at the resolution of single
genes and single neurons.

11
The cell-intrinsic sex determination pathway regulates all known somatic sex
differences in C. elegans
C. elegans has two sexes: XX hermaphrodite and X0 male (Figure 1.1).
Hermaphrodites are essentially somatic females except it can self-fertilize in the
absence of males. As in most animals, C. elegans sex determination depends on sex
chromosomes: XX or X0. However, in contrast to the vertebrate sex determination in
which the early gonad primes tissues to adopt sex-specific characteristics through the
influence of gonadal steroids [Morris et al., 2004], most sex differences in C. elegans
extragonadal tissues do not depend on gonads. This has been experimentally
demonstrated by laser ablation of early gonadal primordium cells, which had no
effect on sex-specific somatic development [Kimble, 1981; Klass et al., 1976].
Furthermore, C. elegans does not have such sex hormones (e.g., estrogen) produced
by its gonad, although the presence of potential gonad signaling is reported [Lipton et
al., 2004; Kleemann et al., 2008].
C. elegans somatic sex determination mostly relies on the sexual state of each
cell (Figure 1.2). By measuring the X chromosome dosage, an inhibitory genetic
cascade converges onto the terminal sex regulator gene, tra-1 (transformer 1). tra-1
then directs sexual differentiation throughout the soma. Elegant genetic studies on
the tra-1 null mutant revealed that tra-1 acts cell-autonomously in the specification of
nearly all sexually dimorphic cell fates in the C. elegans soma [Hunter and Wood,
1990]. This indicates that TRA-1 activity imparts a sexual identity to each cell, either
hermaphrodite (TRA-1 ON) or male (TRA-1 OFF) sexual fate. However, the
downstream effectors of TRA-1 are not completely known.
The C. elegans core neural circuitry has molecular sex difference
In contrast to the conspicuous sex-specific neurons in each sex, it has been thought
that the core nervous system possesses only minor, ultrastructural sex differences in
connectivity. However, two recent findings revealed that the core nervous system is

12
also sexually differentiated at the molecular level (Figure 1.4). First, srd-1, a seven
transmembrane receptor, is specifically expressed in the male ADF sensory neuron
pair [Troemel et al., 1995]. Second, srj-54, a seven transmembrane receptor, is
specifically expressed in the male AIM interneurons [D.S.P., unpublished data].
These ADF and AIM neurons are present in both sexes - i.e., both neurons are part of
the core nervous system - and these genes exist in both sexes, however, gene
expression was observed specifically in males. Despite their unknown functional
significance, sex-specific gene expression in core neurons may give rise to sexual
modifications in behavior since gene expression differences can result in functional
differences. Together, these findings indicate that the core nervous system is also
sexually differentiated at the molecular level. These molecular substrates of sexual
dimorphism in the C. elegans core nervous system prompted us to initiate this
investigation on potential sex difference in a shared sensory behavior, olfaction since
chemosensation/olfaction may be affected by gene expression differences in neurons.
Sex differences in the C. elegans core neural function
The core nervous system mediates a variety of behaviors displayed by both
hermaphrodites and males. In spite of sexually differentiated non-reproductive
behaviors in the vertebrate systems, such as learning and memory, addiction, and
stress responses, sex differences in the C. elegans shared behaviors of two sexes have
not been understood well. Recently, some learning and memory functions of C.
elegans, mediated by core neurons, have been reported to be sexually different [Vellai
et al., 2006]. Furthermore, it has been found that the core nervous system mediates
sexually different locomotion [W. Mowrey, unpublished data]. These results, along
with molecular evidence of sex differences, suggest that other C. elegans core neural
functions may be also sexually differentiated.
13
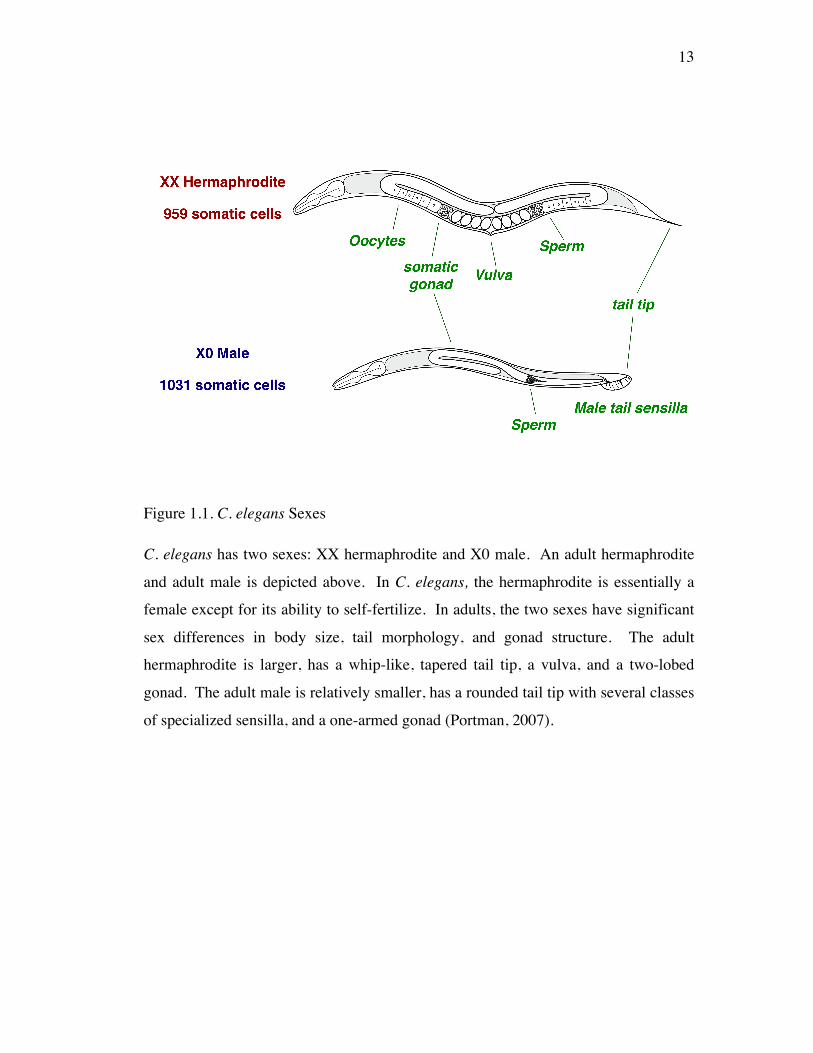
Figure 1.1. C. elegans Sexes
C. elegans has two sexes: XX hermaphrodite and X0 male. An adult hermaphrodite
and adult male is depicted above. In C. elegans, the hermaphrodite is essentially a
female except for its ability to self-fertilize. In adults, the two sexes have significant
sex differences in body size, tail morphology, and gonad structure. The adult
hermaphrodite is larger, has a whip-like, tapered tail tip, a vulva, and a two-lobed
gonad. The adult male is relatively smaller, has a rounded tail tip with several classes
of specialized sensilla, and a one-armed gonad (Portman, 2007).

14
Figure 1.2. Somatic sex determination in C. elegans
All known C. elegans sex differences are derived from the action of the cell-intrinsic
sex determination pathway. The pathway originates from the ratio of sex
chromosomes to autosomes signaled by the signal-element genes on X chromosomes
and A autosomes (Rhind et al., 1995). Downstream of the sdc genes, dosage
compensation (not shown) reduces gene expression from the X chromosomes in half
and is controlled independently from the differentiation of somatic characteristics
(Meyer, 2005). A repressive genetic cascade that ultimately regulates the global sex-
determining gene tra-1 controls all known somatic sex characteristics. In XX
animals, tra-1 is active, repressing male differentiation and promoting hermaphrodite
differentiation. In X0 animals, tra-1 is inactive, allowing male differentiation.
15
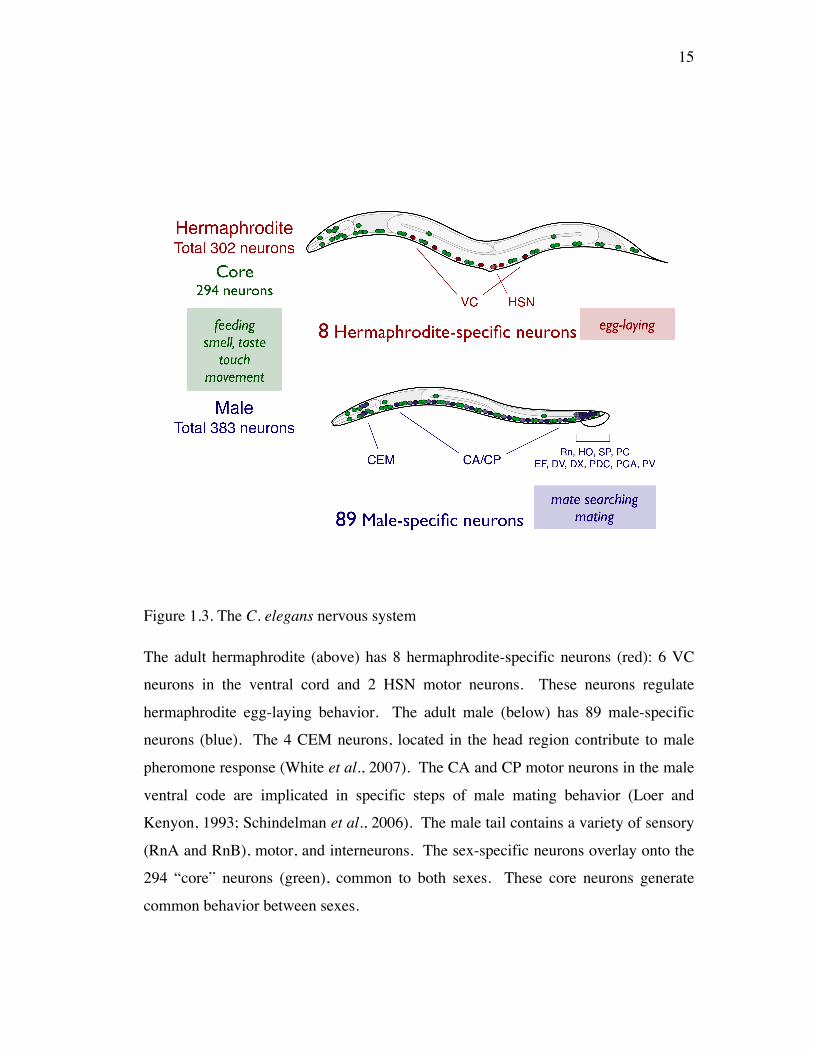
Figure 1.3. The C. elegans nervous system
The adult hermaphrodite (above) has 8 hermaphrodite-specific neurons (red): 6 VC
neurons in the ventral cord and 2 HSN motor neurons. These neurons regulate
hermaphrodite egg-laying behavior. The adult male (below) has 89 male-specific
neurons (blue). The 4 CEM neurons, located in the head region contribute to male
pheromone response (White et al., 2007). The CA and CP motor neurons in the male
ventral code are implicated in specific steps of male mating behavior (Loer and
Kenyon, 1993; Schindelman et al., 2006). The male tail contains a variety of sensory
(RnA and RnB), motor, and interneurons. The sex-specific neurons overlay onto the
294 “core” neurons (green), common to both sexes. These core neurons generate
common behavior between sexes.
16
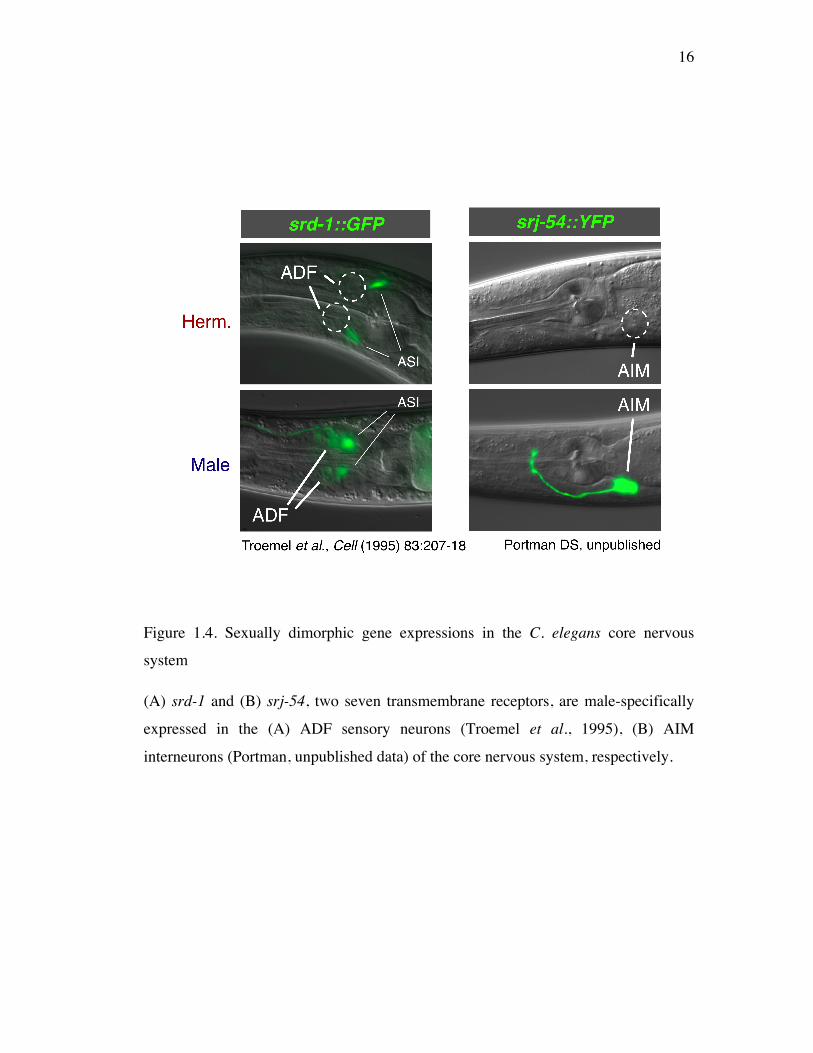
Figure 1.4. Sexually dimorphic gene expressions in the C. elegans core nervous
system
(A) srd-1 and (B) srj-54, two seven transmembrane receptors, are male-specifically
expressed in the (A) ADF sensory neurons (Troemel et al., 1995), (B) AIM
interneurons (Portman, unpublished data) of the core nervous system, respectively.

17
6 C. elegans olfactory behaviors
The hermaphrodite olfactory system is well characterized in its structure and
function
To address potential sex differences in the core nervous system, we systematically
approached this question by studying behaviors (Figure 1.5). Among the many
behaviors mediated by the core nervous system, we chose to study C. elegans
olfactory behaviors, which are relatively simple, easily tractable, and well
characterized in hermaphrodites. C. elegans must encounter thousands of chemicals
in the wild environment (soil) and must be able to sort out massive information to
find food sources, to locate mating partners, and to avoid harmful toxicants. As a
result, worms execute attraction and repulsion behaviors in response to different
sensory cues. In C. elegans, chemotaxis to volatile attractant sources is referred to as
olfactory behavior and is mediated by the core nervous system. All data on the
structure and function of C. elegans olfactory system are based on hermaphrodites
[Bargmann et al., 1993]. In contrast, male olfaction has never been examined. The
olfactory neuronal circuit (Figure 1.6) is composed of a set of sensory (olfactory)
neurons, layers of interneurons, and motor neurons. These neurons are common
between male and hermaphrodite. The connectivity between these neurons is
completely identified in hermaphrodites. In addition, the characterization of the male
neuronal wiring diagram is under way [S. Emmons, Albert Einstein College of
Medicine; http://worms.aecom.yu.edu/pages/all%20male%20neurons.htm]. The
olfactory sensory neurons (Figure 1.6) are AWA, AWB, and AWC, located in the
amphid of head region. Each of these neurons is left and right paired, finger-shaped,
contains branched ciliary endings, is located under the sheath cells, and is indirectly
exposed to the environment [Bargmann et al., 1993; Bargmann, 2006]. AWA and
AWC neurons sense attractive cues and AWB neurons detect aversive molecules.
Olfactory neurons synapse onto intervening layers of interneurons, which are in turn
connected to motor neurons. Through complex activity in the olfactory neural circuit,

18
C. elegans navigates a chemical gradient using temporal comparisons of encountered
concentrations of chemical cues [Pierce-Shimomura et al., 1999; Dusenbery et al.,
1980; Miller et al., 2005]. As with vertebrate and fly olfaction, C. elegans olfaction
also conveys information through G-protein coupled receptor signaling [Colosimo et
al., 2004; Komatsu et al., 1999; Troemel et al., 1995]. However, in contrast to
vertebrate neurons that sense thousands of chemicals through thousands of olfactory
neurons, each of them responding to single odorant, C. elegans can detect thousands
of odorants with a few olfactory neurons. Only one chemical has been linked to its
cognate receptor and the ligands for other receptors are as yet unknown. The only
one de-orphanized odorant receptor in C. elegans is the diacetyl receptor, ODR-10
[Sengupta et al., 1996]. As in other chemosensory systems, interaction of an odorant
with its cognate receptor is predicted to either activate or inhibit synaptic output of a
chemosensory neuron [Wakabayashi et al., 2004; Gray, 2005; Chalasani et al., 2007].

19
Figure 1.5. Does the C. elegans core nervous system mediate sexually dimorphic
behaviors?
The C. elegans core nervous system may also be sexually differentiated and generate
sexually different core neural functions.
20
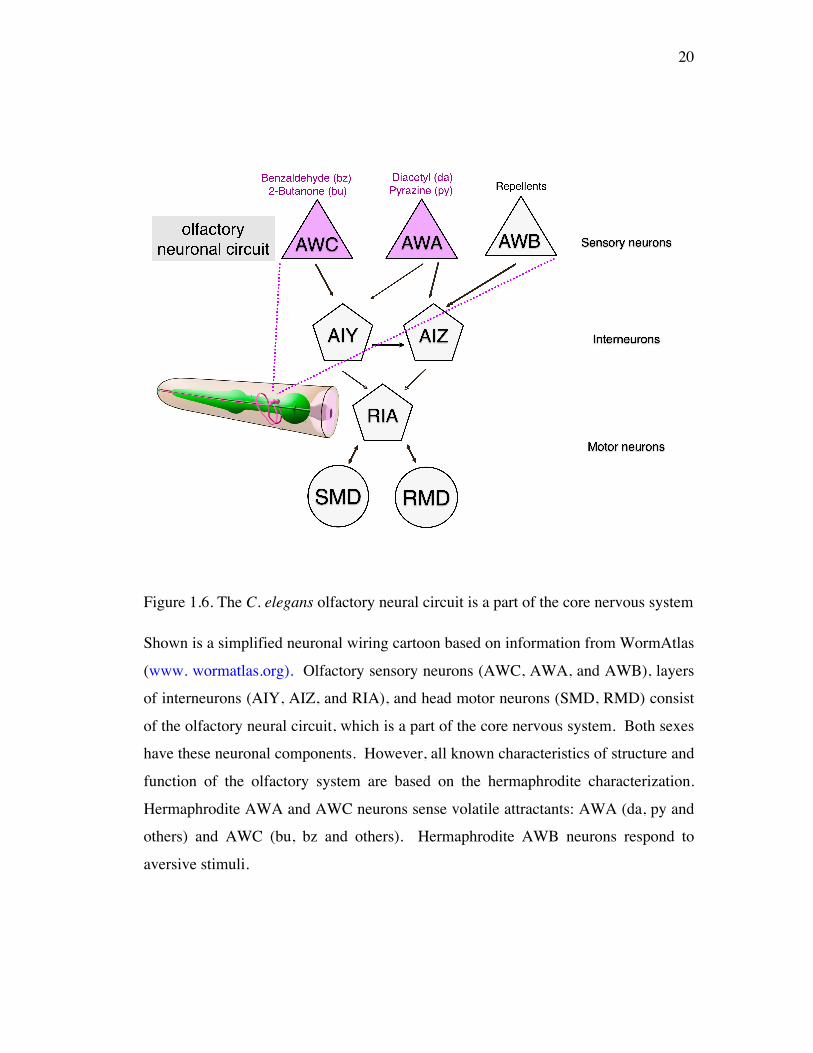
Figure 1.6. The C. elegans olfactory neural circuit is a part of the core nervous system
Shown is a simplified neuronal wiring cartoon based on information from WormAtlas
(www. wormatlas.org). Olfactory sensory neurons (AWC, AWA, and AWB), layers
of interneurons (AIY, AIZ, and RIA), and head motor neurons (SMD, RMD) consist
of the olfactory neural circuit, which is a part of the core nervous system. Both sexes
have these neuronal components. However, all known characteristics of structure and
function of the olfactory system are based on the hermaphrodite characterization.
Hermaphrodite AWA and AWC neurons sense volatile attractants: AWA (da, py and
others) and AWC (bu, bz and others). Hermaphrodite AWB neurons respond to
aversive stimuli.

21
Olfactory neural circuits possess molecular properties for behavioral plasticity
The C. elegans chemosensory system is highly developed to mediate recognition,
discrimination, and adaptation to chemical cues. Moreover, like sensory behaviors in
other higher organisms, C. elegans can also modulate sensory behaviors according to
its internal and external state such as memory of past experience, feeding state, and
contextual cues. Furthermore, some genes involved in food/odor learning behaviors
have been identified [Ishihara et al., 2002; Remy and Hobert, 2005]. In particular,
cGMP and PKC signaling acts as a behavioral switch in a single neuron (AWC) to
switch between odor preference and avoidance [Tsunozaki et al., 2008]. This
behavioral switch is suggested as a mechanism underlying AWC olfactory
sensitization/adaptation and provides an example of how changes in the molecular
properties of a neuron can give rise to modification of behavior. Together with the
fact that some chemosensory receptors are involved in the sensation of internal state
modulating behaviors, modification of neural circuit function by gene expression
changes may be an important property of behavioral plasticity in C. elegans.

22
Chapter 2 Neural sex modifies the function of a C. elegans sensory circuit
1 Introduction
In spite of the importance of studying sex difference in the brain, it is not understood
how chromosomal sex generates complex sex differences in the neural circuit and
behavior. The relatively simple and well-characterized C. elegans nervous system
and tractable innate behaviors make it feasible and interesting to study this complex
issue. C. elegans has two sexes: XX hermaphrodites and X0 males. The cell-
intrinsic sex determination pathway determines the sexual state of each somatic cell
[Hodgkin, 1987; Hodgkin and Brenner, 1977; Hunter and Wood, 1990]. In the
pathway, the TRA-1 (a terminal sex regulator) activity determines either
hermaphrodite or male fate of a given cell. The C. elegans nervous system harbors
extensive sexual dimorphism, including the structurally and functionally well-
characterized sex-specific nervous system. The sex-specific nervous system is
composed of distinct sets of sex-specific neurons, which mediate sex-specific
behaviors for reproduction. This sex-specific nervous system connects to the “core”
nervous system comprised of 294 common neurons between sexes. The core nervous
system has been thought to exhibit only a few ultrastructural level sex differences.
However, recent findings on two male-specific gene expressions in core neurons
reveal that the core nervous system is also sexually differentiated in its molecular
properties. Although the functional significance of these molecular sex differences in
the core neurons is not understood, these could contribute to sex differences in neural
function. Therefore, the core nervous system may also be sexually differentiated in
its functions imparting sex differences to shared behaviors between the two sexes in
C. elegans.
To ask how core neural functions may differ between sexes, we choose to
examine C. elegans olfactory behavior, a function of the core neural circuitry.
Because male olfaction has never been carefully examined, we began characterizing
male responses to simple canonical odorants for comparison to the well-characterized

23
hermaphrodite olfaction. My results reveal that the “neural sex”, the sexual state of a
given neuron established by cell-intrinsic sex determination, determines the sexual
phenotype of olfaction. Each sex revealed distinct and characteristic olfaction giving
rise to significant sex differences. This suggests that neural sex establishes sexually
different properties in core neural circuitry to bring about sex differences in a shared
sensory behavior.
2 Materials and Methods
Nematode Genetics, Strains, and Transgenes
C. elegans cultures were grown on nematode growth medium (NGM) plates seeded
with E. coli OP50 as described [Brenner et al., 1974]. him-5 (high incidence of
males) was used as our lab stock for stable and abundant generation of male
progenies in self-fertilizing population by him-5 hermaphrodites. him-5(e1490)
mutation increases the chance of meiotic disjunction of chromosomes leading to the
higher ratio of spontaneous male progenies in a given population. In all my
experiments, him-5 genetic background was used except in strains containing tra-1.
him-5 is noted as the wild-type group on figures and in the text.
The following mutant alleles were used: tra-1(e1099) III, pha-1(e2123ts) III,
him-5(e1490) V, lin-15(n765) X, and ceh-30(n4289) X. tra-1 XX pseudomales were
obtained from the self progeny of tra-1(e1099)/pha-1(e2123) hermaphrodites.
Sex-transformation constructs were generated using the GatewayTM cloning kit
(Invitrogen). EG4391 and EG4392 strains containing the Prab-3::fem-
3(+)_mCherry::unc-54 3’UTR transgenes oxEx862 and oxEx863 were generously
provided by J. White and E. Jorgensen [White et al., 2007]. To make Posm-5::fem-
3(+)_mCherry::unc-54 3’UTR and Pglr-1::fem-3(+)_mCherry::unc-54 3’UTR, we
polymerase chain reaction (PCR) amplified the osm-5 [Qin et al., 2001; Haycraft et
al., 2001] and glr-1 [Zheng et al., 1999] promoters and made 4-1 Entry clones for use
in the Multisite Gateway System as described [White et al., 2007].

24
Extrachromosomal arrays were generated by the coinjection of the fem-3(+)
construct at 50–75ng/ml with a coelomocyte::GFP marker (75ng/ml). UR226,
UR227 strains containing the transgenes (fsEx160, fsEx161) and UR224, UR225
strains containing the transgenes (fsEx158, fsEx159) were cordially generated by D.A.
Mason. The expression pattern of each transgene was verified by the observation of
fluorescence from mCherry, encoded by the distal open reading frame (ORF) in these
operon-based constructs. In behavioral experiments on worms with these transgenes,
only those showing clear mCherry expression were assayed.
The following transgenes were used for the marker-gene expression studies
(fsIs6[srj-54::YFP + cc::GFP], bxIs14[pkd-2::GFP + pBX1], oxEx862[Prab-3::fem-
3(+)::mCherry + pkd-2::GFP + lin-15(+)], and zdIs13[tph-1::GFP]) (Figure 2.9).
Behavioral assays
Single odorant assay
The single odorant assay (Figure 2.1) is essentially the same as the classical
chemotaxis assay [Bargmann et al., 1993]. It measures worms’ responses to a single
odorant diluted in 100% EtOH (1ul) placed on the left spot, 0.5cm apart from the
edge of the assay plate. Opposite to the odorant spot, 100% EtOH as a control was
placed on the right spot, 0.5cm apart from the edge of the plate. 100% EtOH is
neutral to worms. It was used for a control spot to distinguish the specific response of
worms to a single odorant presented simultaneously. To paralyze worms that get to
the odorant source, 1 ul of 0.3 M NaN3 was placed on both EtOH and odorant spot.
Populations of single sex worms were placed on the 2% agar assay plate without food
and after 45 minutes the number of worms at both spots was counted for the
quantification of olfactory behaviors. For quantification, the Chemotaxis Index (C.I.)
which equals to (B-A)/Total was used. A and B represent the number of worms at
odorant spots. The C.I. varies from +1 (complete attraction) to -1 (complete
repulsion) [Bargmann et al., 1993]. An average of 50 animals were subjected to each

25
Figure 2.1. Single odorant assay
“Single odorant assay” is essentially the same as the classical chemotaxis assay
(Bargmann et al., 1993)

26
assay. Four to twelve assays of each sex at each dilution of an odorant were
performed. For statistical significance, we have used a two-sample Student’s t-test
assuming equal variances between sexes of each odorant at each dilution.
To remove any possible effect of age variance in olfactory behavior, we used
age-synchronized cultures for all behavior assays otherwise it is noted. This was
carried out by synchronized egg-laying rather than hypochlorite treatment to avoid
any potential side effects of larval starvation. Briefly, 20 gravid hermaphrodites were
allowed to lay eggs on a seeded plate for 2 h and were then removed. The resulting
progeny matured in a relatively synchronous manner. To avoid any potential
influence of interaction between sexes onto olfaction, in all assays, animals were sex-
segregated as L4 larvae, before male mating structures have not yet developed, and
transferred to single-sex “holding plates” overnight before behavioral assays.
Olfactory preference assays (Figure 2.2)
The olfactory preference assay is done with basically the same set up and procedure
as the single odorant assay [Chapter 2.1 Materials and Methods] except that another
volatile attractant is placed on the left spot instead of 100% EtOH. Both odorants
used for this assay are diluted into 100% EtOH and a 1/100 dilution of each odorant
was used in all my olfactory preference assays.
I quantified behaviors of single sex populations of worms in this assay using
an Olfactory Preference Index (OPI), defined as (b-a)/(a+b), where a and b represent
the numbers of animals migrating to odorants A and B, respectively. The OPI can
vary from -1 (indicating a complete preference for odorant A) to +1 (a complete
preference for odorant B). An OPI of 0 indicates that equal numbers of animals
migrate to each of the two spots. Behaviors of wild-type animals were analyzed by
one way analysis of variance (ANOVA) with equal variance with Bonferroni post-
hoc test.

27
Figure 2.2. Olfactory preference assay
(A) The olfactory preference assay (Lee and Portman, 2007) is a modification of the
single-odorant assay in which the control spot (“A”) is replaced with a second
attractive odorant. All attractants were diluted to 1/100. A sex-segregated population
of worms is placed 1 cm below the center of the plate. After 45 min, the number of
animals within 2 cm of each spot is counted and used to calculate the olfactory
preference index (OPI). This assay eliminates any potential confounding effects of
other sexually different behaviors (e.g., movement rate or mating drive) or overall
sensitivity on olfactory response, as it measures the relative difference in attraction to
two different odorants. (B) OPI can range from -1 (strong preference for odorant A)
and to +1 (strong preference for odorant B). OPI of +1 means more worms get to the
odorant B spot than to the A spot. OPI of 0 indicates similar preference of both
odorants as the assay plate reveals approximately equal number of worms gets to both
spots.

28
Larvae cultures and assays
I have used synchronized cultures set up from different time points to get
simultaneous populations of L3, L4, and adult animals. L3 (36-40 hr), L4 (44-48 hr),
and adult (~72 hr after laid egg) were used. L4 animals were separate-sexed and
grown as adult for adult experimental group. To test statistical significance between
sexes of each developmental stage, I used the Student t-test.
Laser ablation and assays
I followed a standard protocol to ablate gonad precursor cells [Bargmann et al.,
1995]. Laser ablations were performed on gonad primordium cells (Z1, Z4) and
germline precursor cells (Z2, Z3) in early larval stage 1 (L1) animals. Operated
animals were rescued, grown up to larval stage 4 (L4) and separated by sex. Animals
that did not undergo laser ablation (mock) were also separated by sex at L4 and
assayed as adults. Animals were scored as responding to a particular odorant if, after
30 min on the assay plate, their distance from that odorant source was less than 40%
of the distance to the other odorant source. (This constraint traces an arc around each
odorant source, the radius of which varies from ~2.5 cm at the plate’s equator to ~2.8
cm at its edge.) Data from assays of laser-ablated animals were nonparametric and
analyzed by logistic regression.
Sexual mosaics
Because hermaphrodites carrying Prab-3::fem-3(+)::mCherry::unc-54::3’UTR
(oxEx862 and oxEx863) and Posm-5::fem-3(+)::mCherry::unc-54::3’UTR transgenes
(fsEx160 and fsEx161) laid very late-stage eggs, these animals were manually staged
as mid-L4s. In all assays, animals were sex-segregated as L4 larvae and transferred
to single-sex “holding plates” overnight before behavioral assays. In behavioral
experiments on worms with these transgenes, only those showing clear mCherry
expression (or in the case of oxEx862 and oxEx863, the rescue of the lin-15 Muv
(Multi-vulva) phenotype) were assayed. Comparisons of the behavior of wild-type

29
and transgenic animals were carried out with two way analysis of variance (ANOVA)
with Bonferroni post-hoc tests.
Statistical Analyses
For all behavior assays, weighted means and standard errors of the mean (SEMs)
were calculated with Stata 9 (StataCorp LP [College Station, TX]). I used the total
number of worms in each single odorant assay or the number of responders in each
olfactory preference assay to weigh the mean and SEM. Comparisons of the behavior
of wild-type single sex population and mutant or other groups, depending on the
experimental design, were carried out with one way or two way analysis of variance
(ANOVA) with Bonferroni post-hoc tests.
3 Results
C. elegans exhibit significant sex difference in olfactory behaviors
Though hermaphrodite olfaction to a variety of odorants has been well characterized
[Bargmann et al., 1993; Ward et al., 1975; Ware et al., 1975], male olfactory
responses have never been systematically examined. To explore C. elegans male
olfaction, we compared the responses of adults of each sex to four volatile attractants
of hermaphrodites: diacetyl (da), benzaldehyde (bz), pyrazine (py), and 2-butanone
(bu). I used a single odorant assay, previously described as the chemotaxis assay, for
measuring olfactory behaviors to a single volatile attractant [Bargmann et al., 1993].
I have tested three serial dilutions of each odorant noted on the X axis. The response
measured as the Chemotaxis Index (C.I.) is displayed as columns of each chart for
each odorant.
I examined male responses to three serial dilutions of all four volatile
attractants of hermaphrodites. Male responses were all attractive (C.I. > 0) revealing
that male olfactory responses are similar overall to the previously described

30
hermaphrodite olfactory responses [Bargmann et al., 1993]. However, male
responses, in many cases, were significantly lower than those of hermaphrodites
(Figure 2.3). Specifically for da (an AWA odorant in hermaphrodites) and bz
(AWC), male responses were significantly reduced compared to hermaphrodites’ at
all dilutions of both odorants except at bz 1/1000. In responses to py (AWA) and bu
(AWC), male responses were similar to hermaphrodites at all dilutions of both
odorants except at py 1/100 and bu 1/100. Together, these reveal that male responses
to some, but not all, olfactory attractants are lower than hermaphrodite responses.
Moreover, this suggests that C. elegans has odorant- and concentration-specific sex
differences in olfaction.
Each sex displays distinct and characteristic olfactory preferences
To define sex differences in C. elegans olfaction more specifically, we have
developed an olfactory preference assay in which two different odorants were
simultaneously presented to single-sex population. Two attractants were placed on
the opposing sides of an assay plate without food (Figure 2.2). Migration to one
odorant source or the other should depend on an animal’s relative preference for each
odorant. If male olfactory function is simply less efficient than that of
hermaphrodites, males would be expected to distribute themselves among the two
odorant spots in the same relative number as hermaphrodites do. If, however, there
are more specific sex differences in olfactory behavior, males and hermaphrodites
might exhibit differences in their relative attraction to the two odorants.
Using the olfactory preference assay, we examined the behavior of wild-type
adult hermaphrodites and males to four pairwise combinations of hermaphrodite
attractants. I used 1/100 dilutions of each odorant, as this concentration usually
results in peak responses under single-odorant conditions (Figure 2.3) [Bargmann et
al., 1993]. Additionally, the response to each of these odorants at this concentration
is known to be mediated predominantly by a hermaphrodite single sensory neuron
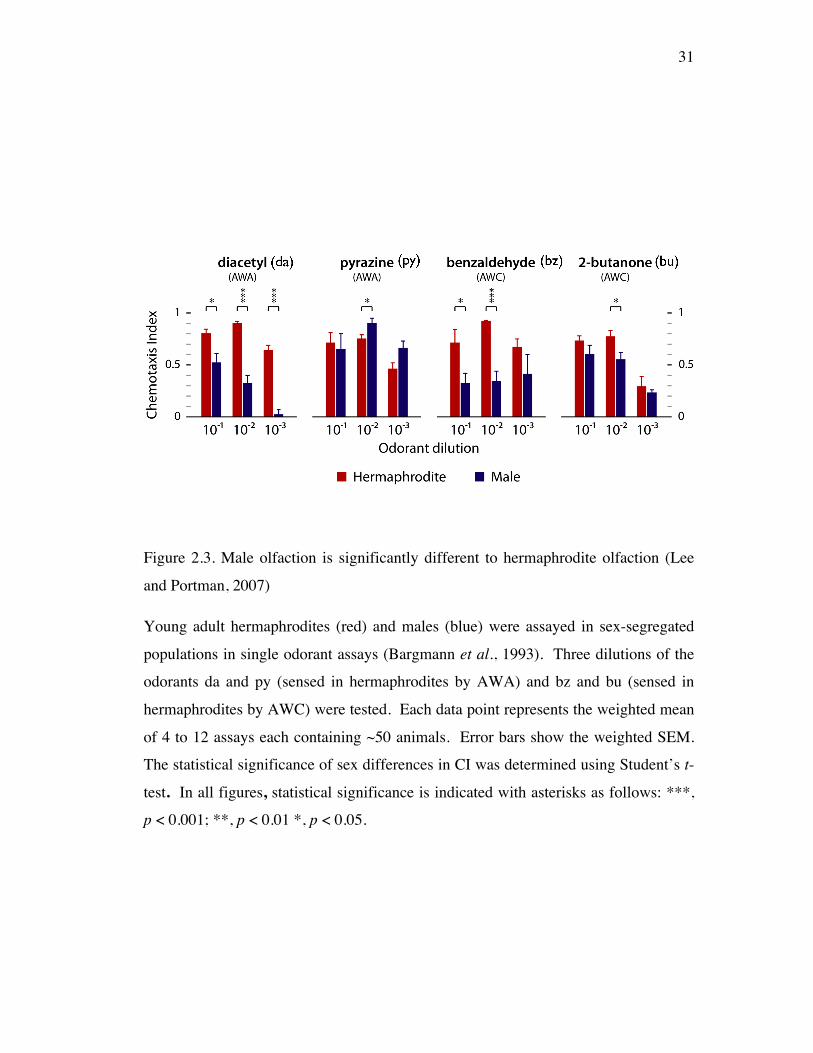
31
Figure 2.3. Male olfaction is significantly different to hermaphrodite olfaction (Lee
and Portman, 2007)
Young adult hermaphrodites (red) and males (blue) were assayed in sex-segregated
populations in single odorant assays (Bargmann et al., 1993). Three dilutions of the
odorants da and py (sensed in hermaphrodites by AWA) and bz and bu (sensed in
hermaphrodites by AWC) were tested. Each data point represents the weighted mean
of 4 to 12 assays each containing ~50 animals. Error bars show the weighted SEM.
The statistical significance of sex differences in CI was determined using Student’s t-
test. In all figures, statistical significance is indicated with asterisks as follows: ***,
p < 0.001; **, p < 0.01 *, p < 0.05.

32
pair: hermaphrodite AWA neurons for da and py, hermaphrodite AWC neurons for bz
and bu [Bargmann et al., 1993; Chou et al., 2001; Sengupta et al., 1994].
I first examined the responses of animals to opposing pairs of attractants
sensed by two different sensory neurons of hermaphrodites (Figure 2.4). For both
pairs tested, da-bz and py-bz, we found significant and robust sex differences in
olfactory preference behavior. In the da-bz assay, males strongly preferred da
although hermaphrodites displayed approximately equal preference for both odorants.
This sex difference in OPIda-bz was statistically significant. Sex differences in the py-
bz assay were also statistically significant; in this case hermaphrodites preferred bz
while males preferred py. A model in which male olfactory responses are simply less
efficient than those of hermaphrodites cannot easily account for these results.
Instead, these pronounced disparities in OPI between males and hermaphrodites
indicate that there are qualitative sex differences in C. elegans olfactory function.
I also found significant sex differences in olfactory responses to two odorants
sensed by the same sensory neuronal pair (Figure 2.4). In the da-py assay, in which
both odorants are sensed primarily by hermaphrodite AWA neurons, hermaphrodites
demonstrated a strong preference for da, while males showed a similarly strong
preference for py. Again, this difference was statistically significant. I also observed
sex differences in the bu-bz assay, in which both odorants are sensed by the
hermaphrodite AWC neuron, and observed less pronounced sex difference.
In another combination of hermaphrodite AWC odorants, bu and iaa
(isoamylalcohol), male and hermaphrodite olfaction was significantly different
(Figure 2.4). These results suggest that sex differences in olfactory behaviors can
arise from olfactory signaling via a single sensory neuron pair and raise the
possibility that sexual differentiation may influence the properties of sensory neurons
themselves.

33
Figure 2.4. Each sex has distinct and characteristic olfactory preferences
(A) The OPI of adult hermaphrodites (open red circles) and males (closed blue
circles) is shown for each of the four odorant pairs (Lee and Portman, 2007) and the
fifth odorant pair indicated at the left and right side of each cart. Four volatile
attractants of hermaphrodites: diacetyl (da), benzaldehyde (bz), pyrazine (py), and 2-
butanone (bu) were paired in various combinations for these olfactory preference
assays. Each point represents the weighted mean of at least 7 olfactory preference
assays each containing ~50 animals. Error bars indicate weighted SEM. The
significance of sex differences in OPI to each odorant pair was determined using
Student’s t-test.

34
Sexually different olfaction is not the secondary effect of male-specific behaviors
To investigate how these highly characteristic and distinct sex differences in a shared
sensory function of C. elegans come about, we tested the role of sex-specific
behaviors and sex-specific structures.
Sexually different olfactory preference might have resulted from the
secondary effect of previously described sex-specific behaviors, particularly male-
specific mating behaviors. My typical observations on male behaviors in a male only
population were that males attempt to mate with neighboring worms when they ran
into each other regardless of the sex of the neighbor. Together, male mating
behaviors somehow might impart sex differences to olfaction.
To examine the potential contribution of male-specific behaviors to male
olfactory preference, we compared male behaviors in single-sex population,
individuals, and mixed-sex population (Figure 2.5A). First, we tested the role of male
mating behaviors. The single sex population behaviors were the control to which we
compared the individual behaviors. In isolation, the mating behaviors should be less
frequent than in population with neighbors since individual worm’s
mechanosensation-mediated mating attempt is suppressed. Male mating attempt
could be suppressed at least in part in isolation. I found that males in isolation exhibit
the same olfactory preferences as males in male population, indicating that male
olfactory preference is not a secondary effect of the male mating behavior. Second,
the role of male mate-searching behaviors was examined by comparing male
behaviors in male only population to that in mixed-sex population. Male mate-
searching drive is a male-specific behavior in that males leave the food area, the
bacterial lawn on the culture dish, in the absence of hermaphrodites. If male mate-
searching drive is the primary contribution to sexual difference in olfactory
preference, male behaviors in a mixed-sex population should be different from male
behaviors in a male-only population.
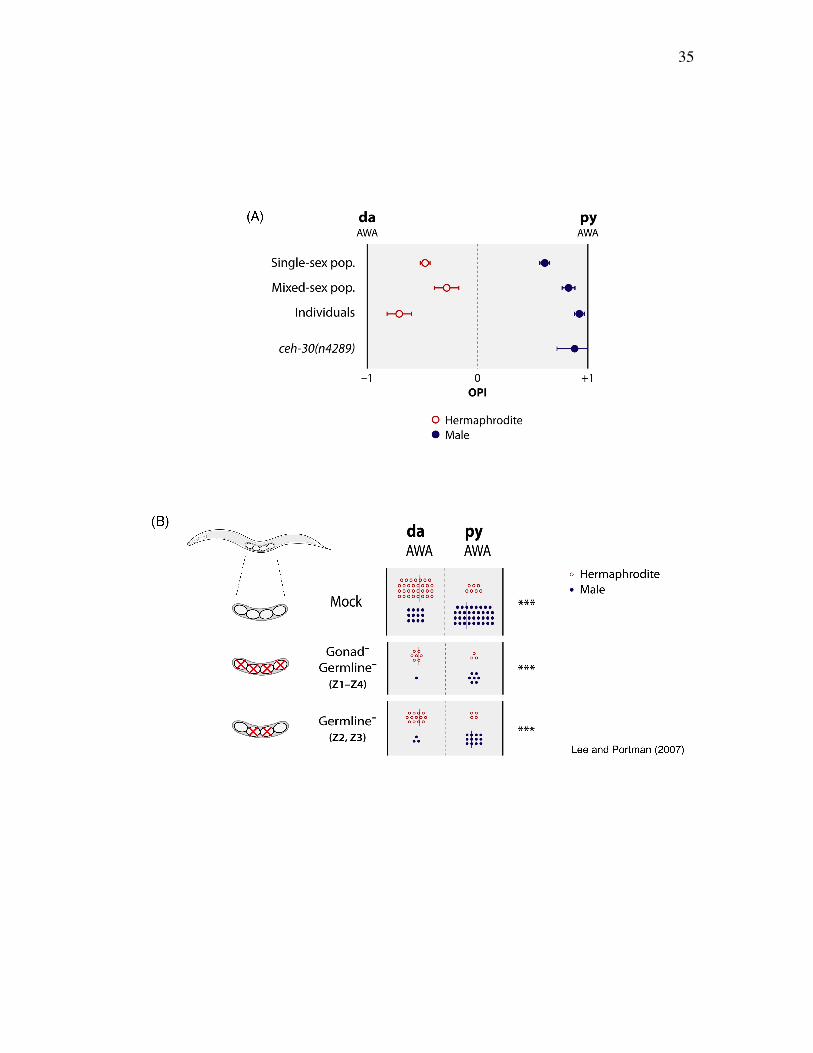
35

36
Figure 2.5. Sexually different olfactory preferences are generated neither by sex-
specific behaviors nor by structures (Lee and Portman, 2007)
(A) Data are shown for animals assayed in standard single-sex population assays,
mixed-sex population assays, and animals assayed individually. ceh-30 mutant males
were assayed in standard, single sex population assays. Each data point represents
the weighted mean and weighted SEM of at least ten assays, each containing ~50
animals, except for ceh-30, which represents six assays of ~40 animals each.
Statistical significance was determined by ANOVA with Bonferroni post-hoc tests.
Each experimental group revealed sex differences (***). ceh-30 males and wild-type
males are significantly different (***).
(B) Mock-ablated, germline and gonad (Z1-Z4)-ablated, and germline (Z2, Z3)-
ablated animals were tested individually for da-py olfactory preference behavior as
young adults. Behavioral responses were determined using a modified olfactory
preference assay as described in Experimental Procedures. Each point represents
odorant-preference behavior of single animal (open red circle, hermaphrodite; closed
blue circle, male). Vertical bars indicate the median response of each group of
animals. Logistic regression was used to determine the statistical significance of the
sex difference in behavior in each of the three groups.

37
However, in the mixed-sex population, male behaviors were still significantly
different from hermaphrodite behaviors. This indicates that male olfactory preference
is not a secondary effect of the male mate-searching drive. Therefore, sexually
different olfactory preferences might be attributed to specific modification in the
machinery for olfactory behaviors rather than to an influence from the machinery for
male-specific behaviors.
The male-specific CEM neurons do not have a primary role in the sexually
different shared sensory function
Sex-specific neurons may contribute to sex differences in olfactory preferences
through direct and/or indirect input to the core neurons. The male-specific CEM
sensory neurons are located in the head region where the olfactory neural circuit
resides. CEM neurons regulate male-specific responses to hermaphrodite conditioned
medium, which contains hermaphrodite pheromone [White et al., 2007]. Therefore, I
postulated that male-specific CEM neurons might be responsible for generating male
olfactory preference. To address this, ceh-30 mutant males, in which all four CEM
neurons are absent, were subjected to the da-py olfactory preference assay.
Intriguingly, ceh-30 males behaved as the wild-type males indicating that male-
specific CEM neurons are not required for the sexually different olfactory preference
behavior (Figure 2.5A). This stands in contrast to the contribution of male-specific
CEM neurons to the male-specific hermaphrodite pheromone response and suggests
that the core nervous system itself may generate sex difference in a shared behavior,
olfactory preference.
Gonad signaling is not necessary for sex difference in olfaction
It has been suggested that gonad signaling exists and regulates sex-specific behaviors
in C. elegans, such as male mate-searching behaviors [Lipton et al., 2004; Kleemann
et al., 2008]. I, therefore, asked whether gonad signaling influences olfactory
preference. To address this idea, I first tested behaviors of glp-1 mutants, which lack

38
a germline. I found that adult glp-1 hermaphrodites have similar olfactory preference
as wild-type hermaphrodites (data not shown). To directly test the role of gonad and
germline, we generated worms without both gonad and germline or without germline
only by laser ablation and tested their behaviors in the da-py olfactory preference
assay. Adult worms without both gonad and germline or without germline
maintained intact sex differences in their olfactory preference (Figure 2.5B).
Therefore, gonad signaling is not necessary for sex difference in olfactory preference
at least in the da-py assay and it is consistent with the possibility that the core nervous
system itself imparts sexual dimorphism to olfactory preference.
Sexual differences in olfaction are prominent before sexual maturation
C. elegans life span (Figure 2.6B) begins with embryonic development and
encompasses four larval stages and adulthood. Late in larval development,
significant changes to the nervous system take place in both sexes. In L4
hermaphrodites, the egg-laying system matures [Schafer, 2006; Bany et al., 2003]; in
L4 males, sensory circuits necessary for mating develop in the tail, neurons are added
to the ventral cord, and the CEM head neurons undergo maturation [Sulston and
Horvitz, 1977]. In addition, extensive gonad maturation occurs in both sexes at L4
stage [Kimble, 2005]. I therefore asked whether the appearance of sex differences in
olfactory preference behavior coincided with any of these sex-specific developmental
events.
To address this, we compared the olfactory preferences of larvae to those of
adults (Figure 2.6A). Since extensive sexual differentiation occurs at the late larval
developmental stages, we asked whether the olfactory preference of larval stage 3
(L3) and larval stage 4 (L4) worms might be similar or different from that of adults.
In the py-bz assay, both L3 and L4 larvae showed the same general trend in
olfactory preference behavior exhibited by adults: males preferred py to bz more
strongly than hermaphrodites did. This difference was marginally significant in L4
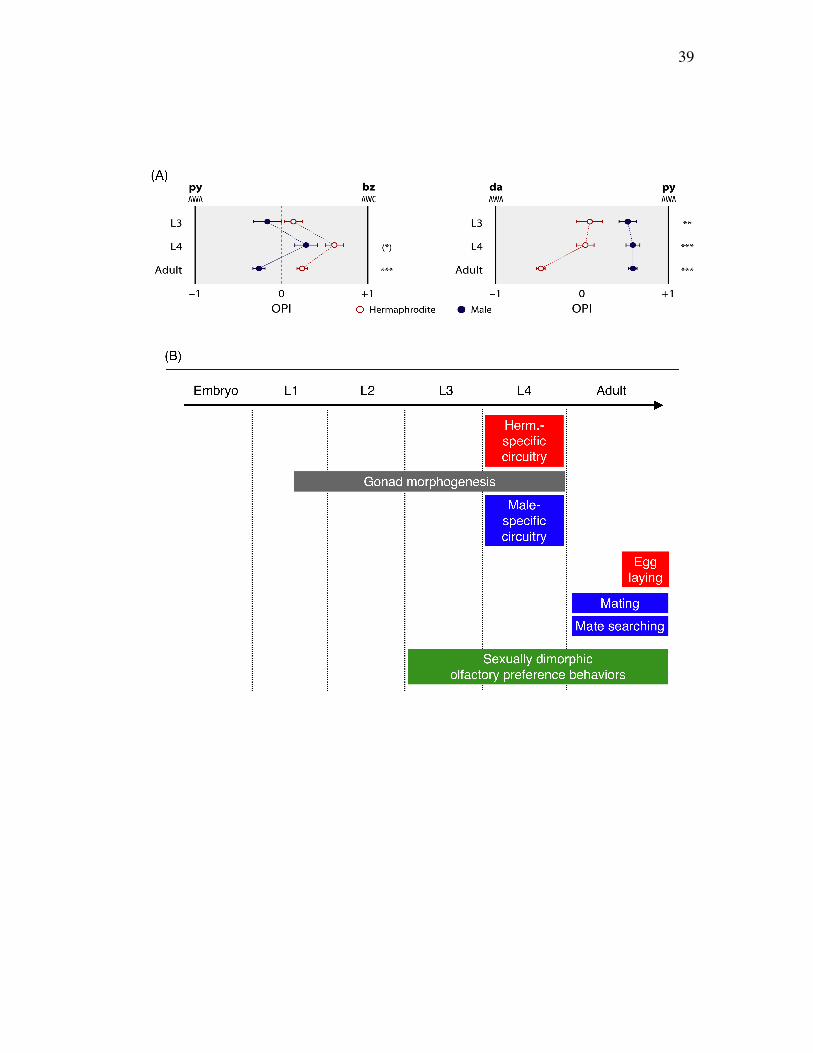
39

40
Figure 2.6. Sex differences in olfaction precede sex-specific differentiation
L3, L4 and young adult animals of both sexes were tested in the py-bz (A, Left) and
da-py (A, Right) assays. Hermaphrodite behavior is shown with open red circles;
male behavior with closed blue circles. Each data point represents the weighted mean
OPI and standard error of at least 10 assays each containing roughly 50 animals. The
significance of sex differences in OPI at each developmental stage was determined
using Student’s t-test and is indicated with asterisks: ***, p < 0.001; (*), p = 0.059.
(B) Major sexual differentiation events in both sexes are noted including sexually
different olfaction.

41
animals (p = 0.059); however, it was not significant in L3 animals. Additionally, L4
animals of both sexes showed a temporary but marked positive shift in OPIpy-bz, the
significance of which is unclear. In the da-py assay, developmental changes were
also apparent. Both L3 and L4 animals showed significant sex differences in OPIda-py,
indicating that clear sex differences are present well before the majority of the sex-
specific nervous system develops. Interestingly, hermaphrodites, but not males,
undergo a significant change in da-py preference behavior as they mature from L4s to
adults, such that the magnitude of the sex difference in OPIda-py is much greater in
adults than in larvae. This suggests that an alteration of hermaphrodite behavior
coinciding with maturation of the reproductive system accounts for the full extent of
sex differences in adult da-py preference. Together, these data demonstrate that
developmental changes in larvae influence sex differences in olfaction, but that some
sex differences in behavior clearly precede the maturation of sex-specific neurons
indicating that the core nervous system possesses properties for sexually different
olfaction even before construction of the sex-specific nervous system.
Neural sex determines the sex phenotype of a common sensory function,
olfactory preference
All known somatic sex differences in C. elegans are regulated by the terminal sex
regulator gene, tra-1. tra-1 loss in XX hermaphrodites results in development of tra-
1 pseudomales, which are virtually identical to wild-type X0 males. Since tra-1
regulates sex-specific development in the C. elegans soma, we postulated that tra-1
also controls sex differences in olfactory preference. In both da-py and py-bz assays,
the behavior of tra-1 XX pseudomales was statistically indistinguishable from that of
the wild-type X0 males (Figure 2.7A). tra-1 XX pseudomales also exhibited male-
like responses in da, py single odorant assays (Figure 2.7B). Therefore, tra-1 also
determines the sex phenotype of a shared behavior, olfactory preference.
Furthermore, it confirms that the cell-intrinsic sex determination pathway controls sex
differences in both structure and function of the nervous system.

42
Next, we explored directly whether the sexual state of the core nervous system
governs sex differences in olfactory preferences. If it does, switching the sexual state
of the nervous system should reverse the sex phenotype of an olfactory behavior.
Studies on both naturally occurring gynandromorph animals and artificial
gynandromorph animals have shown that behavioral sex is determined by the sexual
state of neural substrates for the behavior [Agate et al., 2003; Hall et al., 1977;
Weissburg et al., 2001]. To investigate directly whether the sexual state of neurons
regulates sexually different behaviors, we reprogrammed the sexual state of neurons
by taking advantage of the cell-intrinsic sex determination in C. elegans. By
generation of sexual mosaics – i.e., animals possessing neurons of the opposite sex in
the body of one sex – we could test if changing in sexual state of certain neurons also
changes the sex phenotype of its neural functions (Figure 2.8B).
To reprogram neural sex, we modified the tra-1 activity by manipulation of
the somatic sex determination pathway. In this pathway, FEM-3, a ubiquitin-ligase
cofactor, degrades TRA-1 and promotes male development. Overexpression of FEM-
3 is sufficient to masculinize XX animals [Mehra et al., 1999]. By overexpression of
FEM-3 specifically in the nervous system using a pan-neural promoter (Figure 2.8A),
Prab-3, we switched the sexual state of the nervous system from hermaphrodite to
male. To confirm that sex-transformation on the core neurons was successful, we
asked if male-specific gene expression in core neurons came on in the hermaphrodites
with masculinized neurons. The male-specific expression of a putative seven
transmembrane receptor, srj-54 in the AIM core interneuron of wild-type animals was
used as a reporter for the sex-transformation of core neurons (Figure 1.4). I first
crossed the srj-54::yfp reporter strain with the strain carrying masculinizing transgene
(Prab-3::fem-3) and found that the male-specific srj-54::yfp expression came on in
the AIM interneuron of hermaphrodites by the Prab-3::fem-3 transgene activity
(Figure 2.9). This indicates that the core nervous system is sexually transformed by
the masculinizing transgene even at the gene expression level.
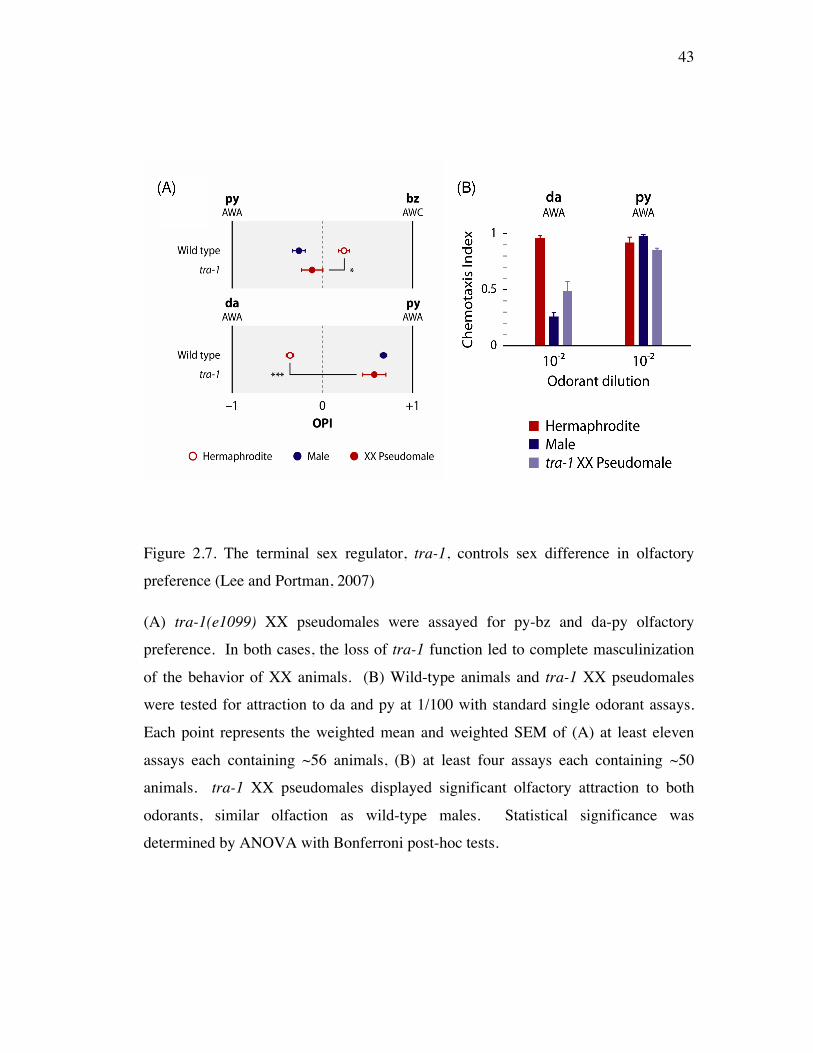
43
Figure 2.7. The terminal sex regulator, tra-1, controls sex difference in olfactory
preference (Lee and Portman, 2007)
(A) tra-1(e1099) XX pseudomales were assayed for py-bz and da-py olfactory
preference. In both cases, the loss of tra-1 function led to complete masculinization
of the behavior of XX animals. (B) Wild-type animals and tra-1 XX pseudomales
were tested for attraction to da and py at 1/100 with standard single odorant assays.
Each point represents the weighted mean and weighted SEM of (A) at least eleven
assays each containing ~56 animals, (B) at least four assays each containing ~50
animals. tra-1 XX pseudomales displayed significant olfactory attraction to both
odorants, similar olfaction as wild-type males. Statistical significance was
determined by ANOVA with Bonferroni post-hoc tests.
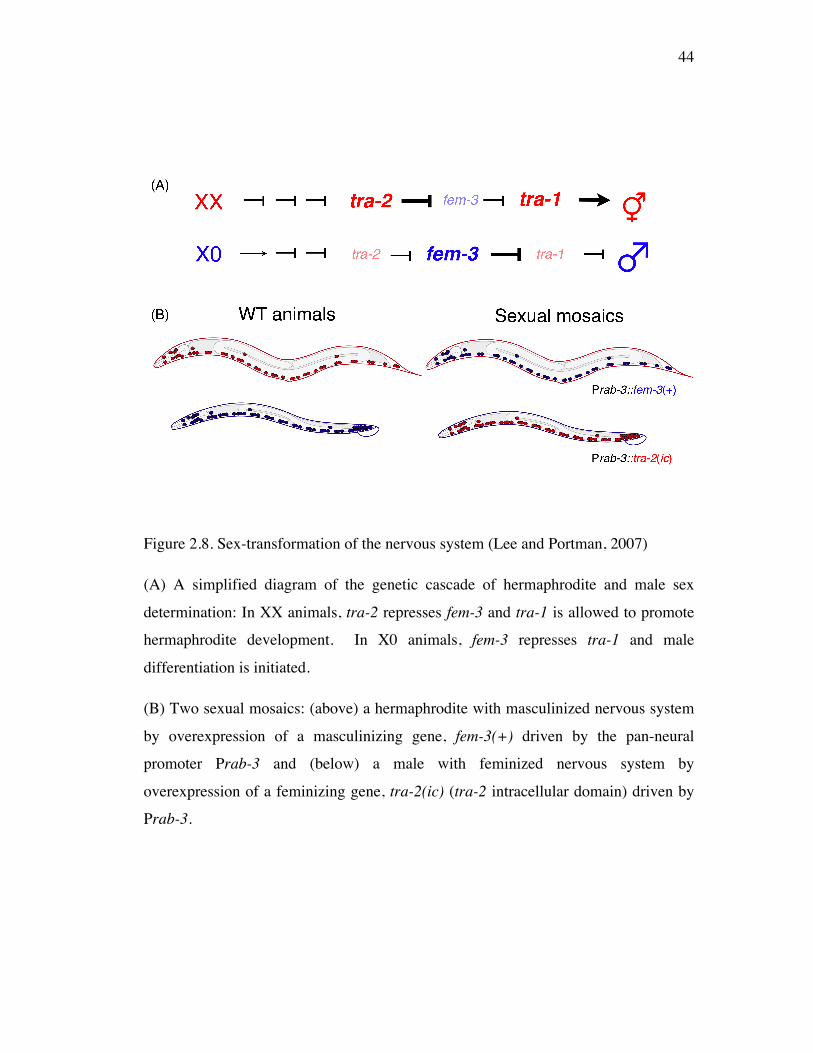
44
Figure 2.8. Sex-transformation of the nervous system (Lee and Portman, 2007)
(A) A simplified diagram of the genetic cascade of hermaphrodite and male sex
determination: In XX animals, tra-2 represses fem-3 and tra-1 is allowed to promote
hermaphrodite development. In X0 animals, fem-3 represses tra-1 and male
differentiation is initiated.
(B) Two sexual mosaics: (above) a hermaphrodite with masculinized nervous system
by overexpression of a masculinizing gene, fem-3(+) driven by the pan-neural
promoter Prab-3 and (below) a male with feminized nervous system by
overexpression of a feminizing gene, tra-2(ic) (tra-2 intracellular domain) driven by
Prab-3.

45
I additionally checked if the pan-neural masculinizing transgene could
generate male-specific neurons in hermaphrodites. As pkd-2::gfp reporter of male-
specific neurons was not revealed in hermaphrodites carrying the pan-neural
masculinizing transgene (Figure 2.9), we, therefore, concluded that masculinizing the
nervous system is sufficient to switch the sexual state of the core neurons only. Male-
specific neurons were not generated by the masculinizing transgene (Prab-3::fem-3)
due to the later expression of the transgene than the sex determination of sexually
dimorphic cell lineages [Portman, 2007]. This technique ensured me to examine the
sole role of the core neurons in sexually different-shared behaviors and to exclude
potential role of the sex-specific neurons.
Pan-neural sex-transformation
Using these sexual mosaics, hermaphrodites with masculinized neurons and males
with feminized neurons, we looked for sex reversal in olfactory preference due to the
sex-transformation on core neurons. In the da-py assay, the response of
hermaphrodites with masculinized neurons was statistically different from the
response of the wild-type hermaphrodites (Figure 2.10). Unlike the wild-type
hermaphrodites, hermaphrodites from two different lines strongly preferred py,
similar to the behavior of wild-type males. Thus, switching the neural sex is
sufficient to transform the sexual phenotype of behavior. This indicates that the
sexual state of the core neural circuit represents the sex phenotype of olfactory
preference.
Sex-transformation on sensory, interneuron and motor neuron
I have identified the core neurons as the generator of sex differences in olfactory
preference. Among various types of neurons in the core neural circuit, we have
selectively sex-transformed most of the sensory neurons or subsets of interneurons
and motorneurons. To target neuronal subtypes for sex-transformation, we have used
cell-type specific promoters: for sensory neurons, we have used the Posm-5 [Qin et

46
Figure 2.9. Neural sex-transformation (Lee and Portman, 2007)
(A) Whole-animal views of young adults showing a wild-type hermaphrodite, an
oxEx862 hermaphrodite, and a wild-type male. The transgenic hermaphrodite has a
normal soma but retains extra eggs (bracketed area), indicating that the
masculinization of the nervous system disrupts egg-laying behavior. A similar
phenotype was seen in the Posm-5::fem-3(+) hermaphrodites (data not shown),
indicating that this phenotype might stem from defects in hermaphrodite-specific
sensory control of egg laying.
(B) Roughly half of oxEx862 adult hermaphrodites express srj-54::GFP in the head
AIM neuron (arrowhead). This expression is never seen in wild-type hermaphrodites
but is observed in nearly all wild-type adult males.
(C) The pkd-2::GFP transgene marks the CEM neurons in the head of adult males

47
(arrowheads). Expression is only very rarely observed in wild-type or oxEx862
hermaphrodites, indicating that oxEx862 does not result in the generation of CEM
neurons in hermaphrodites.
(D) In the male tail, pkd-2::GFP is expressed in the male-specific RnB and HOB
neurons (bracket). No expression was observed in the tails of wild-type or oxEx862
hermaphrodites, indicating that these male-specific neurons do not form in oxEx862
hermaphrodites.
(E) tph-1::GFP is expressed in the hermaphrodite-specific neuron HSN in wild-type
and oxEx862 hermaphrodites (arrowheads). Interestingly, tph-1::GFP expression is
usually reduced in the HSNs of oxEx862 hermaphrodites, and the HSN neurons in
these animals are sometimes mispositioned (data not shown), indicating that fem-3(+)
expression might disrupt HSN differentiation. I also sometimes observed tph-1::GFP
expression in two cells flanking the vulva in oxEx862 hermaphrodites; this might
indicate that the hermaphrodite-specific cells VC4 and VC5 can adopt a CP-like fate
in these animals. In adult males, tph-1::GFP marks these male-specific CP ventral
cord neurons. Four of the CP neurons (open arrowheads) are visible in this view of a
wild-type adult male.

48
Figure 2.10. Neural sex determines the sex phenotype of olfactory preference
Animals carrying fem-3(+) overexpression transgene were assayed in the da-py
olfactory preference assay. Two different transgenic lines are shown. Prab-3::fem-
3(+) (oxEx862 and oxEx863) expresses fem-3(+) throughout the nervous system.
Each point represents the weighted mean and standard error of at least 6 assays with
an average of 56 animals per assay (wild-type), 25 animals per assay (Prab-3::fem-
3(+)). Statistical significance was determined by ANOVA with Bonferroni post-hoc
tests.

49
al., 2001; Haycraft et al., 2001] and for many interneurons (and a small set of head
motor neurons), we have used Pglr-1 [Maricq et al., 1995].
In the da-py assay, the response of hermaphrodites with masculinized sensory
neurons was statistically different from the response of the wild-type hermaphrodites
(Figure 2.11). Unlike the wild-type hermaphrodites, hermaphrodites from two
different lines strongly preferred py, similar to the behavior of the wild-type males in
the da-py assay. This indicates that the sensory neural circuit itself is sufficient to
bring about sexually different olfactory preference. Furthermore, it suggests that the
sensory neural circuit must have sexually different properties giving rise to two
different forms of the shared sensory function.
In contrast, the response of hermaphrodites with masculinized glr-1-
expressing cells was not different from the response of the wild-type hermaphrodites
(Figure 2.11). As the wild-type hermaphrodites, hermaphrodites from two different
lines strongly preferred da in the da-py assay. This indicates that the properties of
glr-1-expressing interneurons and motor neurons are not modified by neural sex.
4 Discussion
Why are there sex differences in C. elegans olfaction?
I have found for the first time that there is a great deal of sex difference in C. elegans
olfactory behaviors. Unlike sex-specific chemosensory behaviors utilized for the
reproduction, the olfactory behaviors we characterized here are exhibited by both
sexes. In the sense that behaviors are naturally selected and evolve according to
beneficiary effects, the sex difference in olfaction we observed may have adaptive
value in worm survival. One possible reason for this sex difference in olfaction is
males’ necessity for finding mates. In spite of incomplete identification of the
hermaphrodite pheromone components, canonical attractants we have examined are

50
Figure 2.11. Sexually different properties of the sensory neurons control the sex
phenotype of olfactory preference
Animals carrying three different fem-3(+) overexpression transgenes were assayed in
the da-py olfactory preference assay. Two different transgenic lines are shown for
each construct. Prab-3::fem-3(+) (oxEx862 and oxEx863) expresses fem-3(+)
throughout the nervous system, Posm-5::fem-3(+) (fsEx160 and fsEx161) expresses
fem-3(+) in ~60 sensory neurons (Qin et al., 2001; Haycraft et al., 2001), and Pglr-
1::fem-3(+) (fsEx158 and fsEx159) expresses fem-3(+) in a large set of interneurons
and motor neurons (Maricq et al., 1995).
Each point represents the weighted mean and standard error of at least 6 assays with
an average of 56 animals per assay (wild-type), 25 animals per assay (Prab-3::fem-
3(+)), 43 animals per assay (Posm-5::fem-3(+)), and 38 animals per assay (Pglr-
1::fem-3(+)). Statistical significance was determined by ANOVA with Bonferroni
post-hoc tests.

51
not thought to be pheromones in C. elegans. Rather, it is more probable that sex
difference in the necessity of nutrition results in sex difference in olfactory behaviors.
Together with the fact that many olfactory attractants are bacterial metabolites, we
prefer this speculation that C. elegans olfaction is sex-specifically modified for biased
nutrition necessity. Males may need more carbohydrates for their high motility and
hermaphrodites may need more proteins for their large volume of oocyte cytoplasms.
For most insects, adult food preferences were dependent on their nutrition
necessity for the reproduction [Cornelius et al., 2000]. Sex differences in non
mating-related chemosensory behaviors were also revealed in Fiddler crabs (Uca
spp.). Their feeding behaviors were sexually different in that male fiddler crabs
confine their feeding in a more food rich regions and display less sensitivity to blends
of food-related chemicals than females having higher sensitivity to low
concentrations of foods [Weissburg and Derby, 1995]. It is suggested that these sex
differences may also arise from sex difference in the necessity of sufficient food.
Therefore, it is a property of evolution of animal behaviors to adopt discrete
modification even in a common behavior between sexes for the advantage of each
sex.
Developmental regulation of sexually different olfactory behaviors
Sexually different olfactory preference was found in developing worms in which no
sex-specific neural circuitry and gonads were established (Figure 2.6). This shows
that the core neural circuitry itself rather than the sex-specific systems generates
sexually different olfaction. However, the diverging hermaphrodite olfactory
preference during the development suggests some influence of the sex-specific
systems on the olfactory preference. The single odorant responses of L4
hermaphrodites to da 1/100 are significantly smaller than that of adult hermaphrodites
(data not shown). L4 males however do not have any difference from adult males in
their single odorant response to da 1/100 (data not shown). Consistent with the
diverging responses of hermaphrodite responses in the da-py assay throughout the L3,

52
L4, and adult stages (Figure 2.6A, Right), the single odorant response of L4 animals
suggest that some non cell-autonomous influences from the development of sex-
specific structures on the core neural circuitry may also contribute to the complete
extent of sexually different olfaction in the da-py assay. Furthermore, in the bz-py
assay, the marked positive shift in the larvae OPI (Figure 2.6A, Left) may be either
revealing unstable olfaction at the sexually differentiating developmental stages or
displaying variability in the larvae olfaction more prominently in the py-bz assay than
in other assays. Altogether, sexually different olfactory preferences seem to be
certainly controlled throughout development. Analysis of the behavior of animals
with temporal manipulation of the sex determinants will give a better understanding
of how sex and development factors coordinate together to establish the complete set
of sex differences in a sensory behavior.
How does sex modify olfaction?
Opposing the dogma that sexual hormones regulate all known sex differences in the
structure and function of the nervous system, my findings reveal that the cell-intrinsic
sex regulators establish sexually different properties in the common neurons between
sexes. In particular, sensory neurons are the generator of sexually different olfactory
preference. Sensory neurons themselves should produce sexually different neural
output giving rise to actual display of sex difference in behavior. Assuming the
connectivity of olfactory neurons to others is the same between sexes, we suggest that
sensory neurons may send sexually different neurotransmission to the postsynaptic
neurons. The neurotransmission may be either qualitatively and/or quantitatively
different between sexes. As the chemosensory neurons of fiddler crabs display
sexually different neural response by modulation of the same cAMP-signaling
pathway [Weissburg, 2001], sensory neurons in C. elegans may also convey two
different outputs by differential regulation on the same signaling components. At one
extreme, however, qualitative and/or quantitative sex difference in critical signaling
molecules may bring about behavioral sex difference. Since the sufficiency of

53
sensory neurons for sexually different responses were revealed in the da-py assay in
which AWA odorants are tested, it is likely that AWA neurons itself possess sex
differences in substrates such as a critical AWA signaling component: da receptor
ODR-10. Although sensory neurons were sufficient for determining the sex
phenotype of olfactory preference, other neurons may also be required to display the
complete extent of sex difference in olfactory preference. Two lines of
hermaphrodites with masculinized sensory neurons display intermediate OPIs
between the OPI of wild-type hermaphrodites and males (Figure 2.11). It is possible
that other subsets of core neurons together with sensory neurons may bring about the
complete sex difference in olfactory preference. Major postsynaptic neurons of
olfactory sensory neurons (AWA and AWC) are AIY and AIZ interneurons. Despite
no contribution of glr-1-expressing neurons, interneurons and head neurons in
connectivity to olfactory sensory neurons may contribute to sexual difference in
olfaction. I suggest that sex-transforming both olfactory neurons and the AIY and
AIZ interneurons might fully reverse the sex phenotype of olfactory preference.
Alternatively, male-specific sensory neurons in addition to core sensory neurons may
be required for the full extent of sex difference in olfactory preference. Since the
sensory neural masculinizing transgene in the hermaphrodites does not establish
male-specific sensory neurons, these sensory neural masculinized hermaphrodites
could have displayed the incomplete sex-switch in behaviors (Figure 2.11).
Neural Sex regulation on Behaviors
Together with recent work in Drosophila [Billeter et al., 2006; Manoli et al., 2006;
Vrontou et al., 2006; Certel et al., 2007; Datta et al, 2008], my work emphasizes
advantages of utilizing the invertebrate systems to elucidate the mechanisms by
which neural sex establishes sexual dimorphisms in the nervous system. I investigate
how neural sex generates sexually different-shared behaviors mediated by the
common neural circuit and this study is distinct from studies on sex-specific neurons
and behaviors. My findings reveal that the cell-intrinsic sex determinants establish

54
sex difference in the properties of the core nervous system and generate sexually
different-shared behaviors. Together with recent evidences that the sex chromosomal
signaling determines sex of the brain, independently from sex hormones [Arnold et
al., 2004; Dewing et al., 2006], my findings may shed light on understanding how
brain is modified by neural sex to accommodate substrates for sex-biases in the nature
and incidences of neurological diseases. Last, studying sex differences in the
common neurons will help unravel the mechanisms by which neural circuitry is
plastic and brings about a variety of behaviors that enhance animals’ survival and
adaptation to the environment.

55
Chapter 3 Neural sex modifies the properties of a single sensory neuron to generate
sex differences in olfactory behaviors
5 Introduction
Understanding the neurobiological substrates of behavior is one of the main problems
in neuroscience. Due to the complexity of the nervous system, it has been
challenging to elucidate mechanisms that innate behaviors, prevalent and conserved
across species, are generated. Sex differences in behaviors have provided a unique
opportunity to study how behaviors are encoded in neural circuitry. Most studies of
sex differences in behavior have focused on sex-specific behaviors, mostly for the
reproduction, present in one sex but not in the other. Extensive studies in C. elegnas,
Drosophila, and mouse have shown that sex-specific neural circuits generate sex-
specific behaviors [Portman, 2007; Villella and Hall, 2008].
The relatively simple C. elegans nervous system allows me to directly address
how common neural circuit is modified to impart sex differences to a shared sensory
behavior. The absence of sex hormones and sex differences even in common
behaviors [Lee and Portman, 2007] are the advantages for utilizing this system to
explore the cell-intrinsic sex regulator, properties of common neural circuit, and
shared behavior. I previously revealed that neural sex, the sexual state of a given
neuron established by cell-intrinsic sex determination, determines sex phenotype of a
shared sensory behavior. Furthermore, sensory neurons are the sufficient core
neurons for sex differences in olfaction [Lee and Portman, 2007]. However, the
mechanisms that neural sex modifies the properties of sensory neurons to generate
sexually different olfaction were not understood.
A pair of left and right AWA olfactory neurons has its cell bodies positioned
in the amphid and its long dendrites project to the tip of nose at which branched
ciliated endings reside [Bargmann et al., 1993]. In hermaphrodites, the AWA
neurons sense volatile odorants: diacetyl (da), pyrazine (py), and 2,4,5-

56
trimethylthiazole (tmt). Responses to these canonical attractants generally peak at the
1/100 dilution of each odorant. The identity of AWA neurons is specified by AWA-
exclusive expression of ODR-7, the only member of the large divergent nuclear
receptor family, which activates the expression of AWA-specific signaling genes to
establish AWA function [Colosimo et al., 2003]. Hermaphrodites carrying the null
mutation odr-7(ky4) fail to express AWA-specific signaling genes [Sengupta et al.,
1996] and fail to respond to all odorants sensed by the AWA neurons [Sengupta et
al., 1994] despite intact AWA neural morphology. odr-7 controls expression of the
single ligand-identified odorant receptor, odr-10 (the diacetyl receptor), in AWA
neurons.
Here, I examine how neural sex generates sexually different olfactory
behaviors and reveal that neural sex modifies target gene expression in the AWA
neuron to bring about sexually different olfactory behaviors.
2 Materials and Methods
Nematode Genetics, Strains, and Transgenes
C. elegans strains were cultured as described in Chapter 2.
Sex-transformation constructs were generated by the GatewayTM cloning kit
from Invitrogen. To make Prab-3::tra-2(ic)_mCherry::unc-54 3’UTR and Posm-
5::tra-2(ic)_mCherry::unc-54_3’UTR, I polymerase chain reaction (PCR) amplified
the rab-3 [Nonet et al., 1997] and the osm-5 [Qin et al., 2001; Haycraft et al., 2001]
promoters and made 4-1 Entry clones for use in the Multisite Gateway System as
described [Hartley et al., 2000]. Extrachromosomal arrays were generated by the
coinjection of the tra-2(ic) (75ng/ul) construct with a coelomocyte::GFP marker
(75ng/ul) into him-5.
UR249, UR250 strains containing the Prab-3::tra-2(ic)_mCherry::unc-54

57
3’UTR transgenes (fsEx187, fsEx188) and UR247, UR248 strains containing the
Posm-5::tra-2(ic)_mCherry::unc-54_3’UTR transgenes (fsEx185, fsEx186) were
generated by W. Mowrey.
To make Podr-7::fem-3(+)_mCherry::unc-54 3’UTR and Podr-7::tra-
2(ic)_mCherry::unc-54 3’UTR, I polymerase chain reaction (PCR) amplified the odr-
7 [Sengupta et al., 1994, Colosimo et al., 2003] promoter and made 4-1 Entry clones
for use in the Multisite Gateway System as described [Hartley et al., 2000].
Extrachromosomal arrays were generated by the coinjection of the fem-3(+) construct
(75ng/ml) with a coelomocyte::GFP marker (75ng/ul).
UR245, UR246 strains containing the Podr-7::fem-3(+)_mCherry::unc-54
3’UTR transgenes (fsEx202, fsEx203) were generated by myself and D.A. Mason,
respectively. UR462, UR463 containing the Podr-7::tra-2(ic)_mCherry::unc-54
3’UTR transgenes (fsEx204, fsEx205) were generated by W. Mowrey. The
expression pattern of each transgene was verified by the observation of fluorescence
from mCherry, encoded by the distal open reading frame (ORF) in these operon-
based constructs. In behavioral experiments on worms with these transgenes, only
those showing clear mCherry expression were assayed.
UR223 (odr-7 (ky4); him-5), UR463 (odr-10 (ky32); him-5), UR458 (kyIs37;
him-5), UR460 (kyIs53; him-5) and UR488 (kyIs53; him-5; fsEx204[Podr-7::tra-
2(ic)_mCherry::unc-54 3’UTR]) were generated by crosses, utilizing the following
mutant strains obtained from the Caenorhabditis Genetics Center: CX4 (ky4), CX32
(ky32), CX3260 (kyIs37), and CX3344 (kyIs53).
The following reporters were used for the odr-10 expression studies:
transcriptional reporter (kyIs37[Podr-10::GFP]) and translational reporter
(kyIs53[Podr-10::ODR-10::GFP]).
The pENTRY[SNB-1_eGFP] construct containing the sequence of SNB-1 and
eGFP translational fusion was generated by performing the B-P recombination

58
reaction utilizing the GatewayTM cloning kit from Invitrogen (W. Mowrey). Podr-
7::SNB-1_eGFP::unc-54 3’UTR, an expression clone encoding SNB-1 only in AWA
neurons, was generated by performing the multi-site gateway L-R recombination
reaction. To locate AWA neurons relevant to adjacent neurons, I selected the AIY
neural-specific reporter strain OH1098 carrying otIs133[Pttx-3::rfp]. The
fsEx201[Podr-7:: SNB-1_eGFP::unc-54 3’UTR] was generated in the OH1098 strain
by W. Mowrey. To examine male expression of SNB-1 compared to hermaphrodites,
the transgenic animals were crossed with fsIs14; him-5 and obtained the UR459
(otIs133; him-5; fsEx201).
Behavioral assays
Single odorant assays and olfactory preference assays were carried out as described in
Chapter 2.
3 Results
Sexually different olfaction arise through neural sex modification on sensory
neurons
In chapter 2, I demonstrated that the masculinization of hermaphrodite sensory
neurons is sufficient to generate masculinized sensory function, namely male
olfactory preference in hermaphrodites [Lee and Portman, 2007]. To more
thoroughly examine the role of neural sex in sexually different sensory function, I
asked if the feminization of male neurons could feminize male olfactory preference.
To feminize all neurons, I overexpressed TRA-2(ic), the constituitively active
intracellular domain of TRA-2, in the nervous system using the pan-neural promoter
Prab-3 [Nonet et al., 1997]. In the sex determination pathway, TRA-2 inhibits FEM-
3 allowing the activation of TRA-1 to result in hermaphrodite development (Figure
2.8). Expression of TRA-2(ic), the predicted intracellular carboxy-terminal domain

59
of TRA-2A, can feminize X0 males [Kuwabara and Kimble 1995]. Utilizing this
technique, I could generate sexual mosaic worms in which the male nervous system is
feminized. This provides mosaics that are complementary to hermaphrodites with the
masculinized nervous system [Lee and Portman, 2007]. I then examined whether
change in the sexual state of neurons reverses the sex phenotype of neural function.
Two lines of males with feminized nervous system revealed hermaphrodite-like
olfactory preference in da-py assay (Figure 3.1). This indicates that feminization of
all neurons in males is sufficient to generate hermaphrodite olfactory preference.
Furthermore, sexual properties in the neural circuitry are plastic enough to be
modified in both masculinization and feminization directions to generate sexually
different neural function. Together, this confirms that neural sex governs sex
difference in olfactory preference.
To feminize only sensory neurons, I overexpressed TRA-2(ic) in the ~60
sensory neurons driven by a sensory neural promoter, Posm-5. osm-5 encodes cilia
protein, which regulates cilia development and osm-5 is expressed in all ciliary
neurons including both core neurons and male-specific neurons in the male tail [Qin
et al., 2001; Haycraft et al., 2001]. Utilizing this technique, I could generate males
with the feminized sensory neurons, in addition to hermaphrodites with masculinized
sensory neurons [Lee and Portman, 2007]. Then I examined whether change in the
sexual state of sensory neurons reverses the sex phenotype of sensory function. Two
lines of males with feminized sensory neurons behaved significantly different from
wild-type males and rather revealed similar response as wild-type hermaphrodite
olfactory preference in da-py assay (Figure 3.2). This indicates that the sexual state
of sensory neurons determines sex phenotype of olfactory preference in da-py assay.
Since the osm-5 promoter affects even male-specific sensory neurons in addition to
common sensory neurons, this result supports the idea that the male-specific sensory
neurons are not required for the olfactory behavioral sex. Together with the
sufficiency of masculinized sensory neurons for male olfactory preference [Lee and
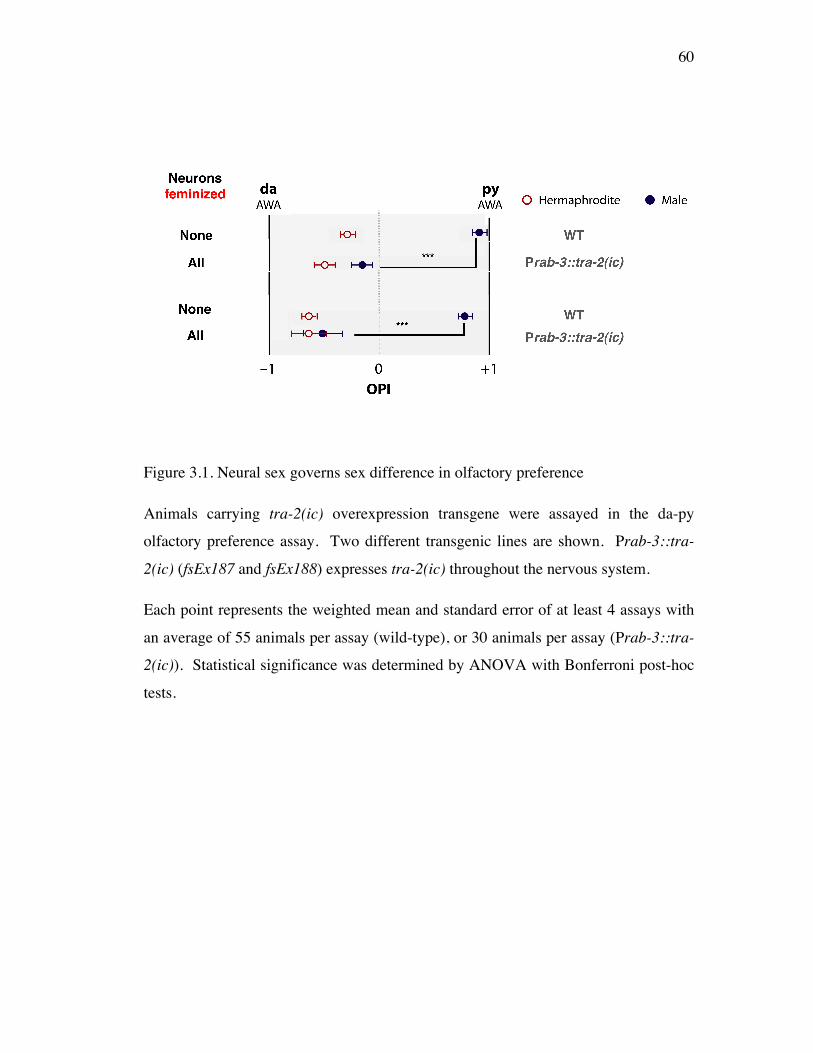
60
Figure 3.1. Neural sex governs sex difference in olfactory preference
Animals carrying tra-2(ic) overexpression transgene were assayed in the da-py
olfactory preference assay. Two different transgenic lines are shown. Prab-3::tra-
2(ic) (fsEx187 and fsEx188) expresses tra-2(ic) throughout the nervous system.
Each point represents the weighted mean and standard error of at least 4 assays with
an average of 55 animals per assay (wild-type), or 30 animals per assay (Prab-3::tra-
2(ic)). Statistical significance was determined by ANOVA with Bonferroni post-hoc
tests.
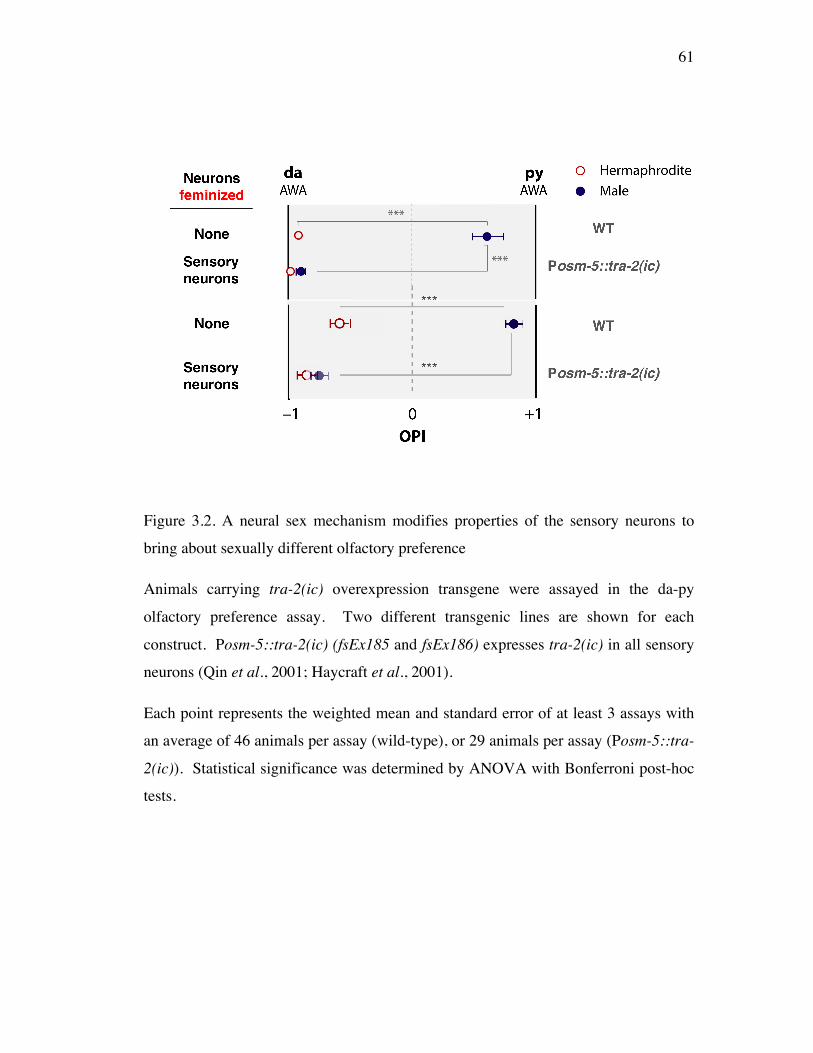
61
Figure 3.2. A neural sex mechanism modifies properties of the sensory neurons to
bring about sexually different olfactory preference
Animals carrying tra-2(ic) overexpression transgene were assayed in the da-py
olfactory preference assay. Two different transgenic lines are shown for each
construct. Posm-5::tra-2(ic) (fsEx185 and fsEx186) expresses tra-2(ic) in all sensory
neurons (Qin et al., 2001; Haycraft et al., 2001).
Each point represents the weighted mean and standard error of at least 3 assays with
an average of 46 animals per assay (wild-type), or 29 animals per assay (Posm-5::tra-
2(ic)). Statistical significance was determined by ANOVA with Bonferroni post-hoc
tests.

62
Portman, 2007], this indicates that the neural sex mechanism modify properties of the
sensory neural circuit to bring about sexually different olfactory preference.
A single sensory neuron, AWA, generates sexually different olfactory preference
Since both da and py odorants are sensed by hermaphrodite AWA olfactory neurons
and sensory neurons are sufficient to mediate sexually different AWA-mediated
olfactory preference, I speculated that AWA neurons themselves may contribute to
sexually different olfactory preference. To explore if AWA olfactory neurons possess
sexual dimorphism, I tested olfaction in animals in which only AWA was sex-
reversed. To switch the sexual state of AWA neurons, I overexpressed sex factors
utilizing the AWA-specific promoter Podr-7 [Sengupta et al., 1994]. odr-7 is a
member of the nuclear receptor family that regulates the AWA neural fate. The
timing of the odr-7 promoter activity, therefore, must be early enough for
modification of the sexual state of AWA neurons. I found that hermaphrodites with
masculinized AWA neurons behave significantly different from the wild-type
hermaphrodites (Figure 3.3A). Two lines of hermaphrodites with masculinized AWA
neurons strongly preferred py as the wild-type males in da-py assay. This indicates
that masculinization of AWA neurons is sufficient to bring about male olfactory
preference. It is fascinating that a single sensory neuron can determine behavioral sex
of the AWA-mediated olfactory preference. Conversely, I examined olfactory
preference of males with feminized AWA neurons. I found that two lines of males
with feminized AWA neurons exhibit wild-type hermaphrodite-like olfactory
preference (Figure 3.3B). This strengthens the idea that neural sex of AWA neurons
is sufficient to generate sexually different olfactory preference at least in da-py
(AWA/AWA) olfactory preference assay. Altogether, this indicates that neural sex
regulates the molecular properties of AWA neurons to bring about sexually different
olfactory preference.
I speculate that masculinization or feminization of the molecular properties of
the AWA neurons must be the basis of the sexually different olfactory preference in
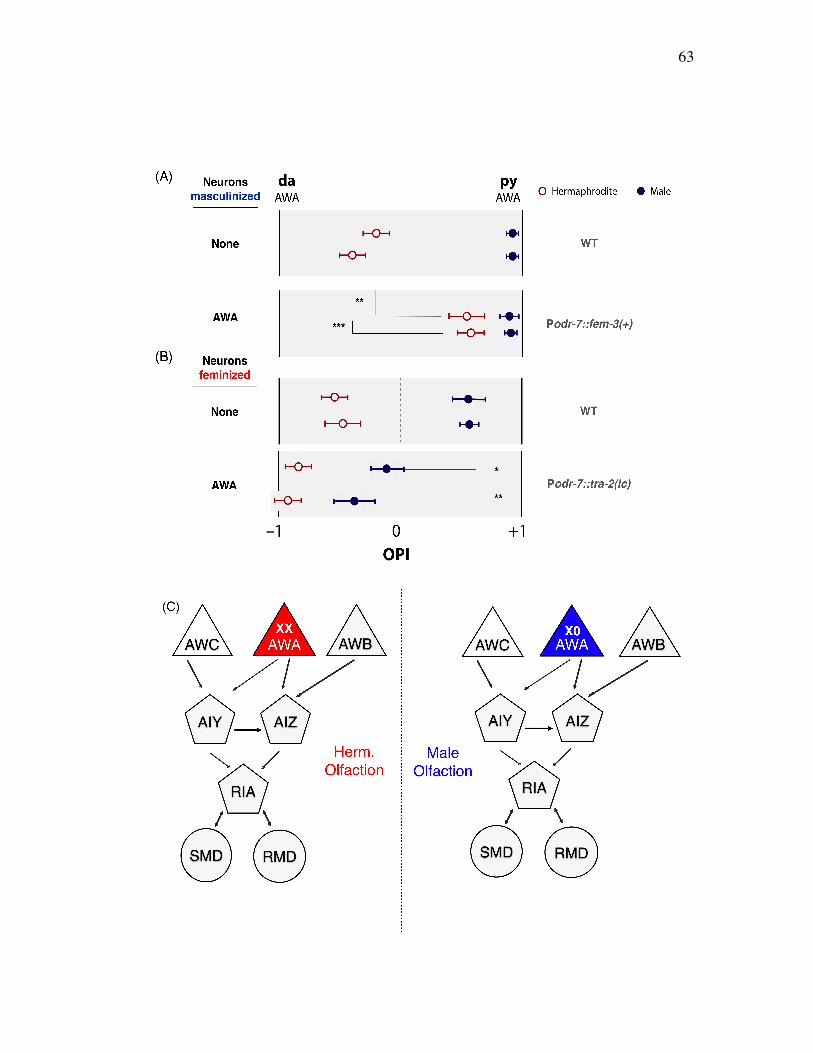
63

64
Figure 3.3. The sexual state of a single sensory neuron, AWA, is sufficient to impart
sex differences in olfactory preference
(A) Animals carrying fem-3(+) overexpression transgene and (B) tra-2(ic)
overexpression transgene were assayed in the da-py olfactory preference assay. Two
different transgenic lines are shown for each construct. (A) Podr-7::fem-3(+)
(fsEx202 and fsEx203) expresses fem-3(+), (B) Podr-7::tra-2(ic) (fsEx204 and
fsEx205) expresses tra-2(ic) only in AWA single neurons.
Each point represents the weighted mean and standard error of (A) at least 6 assays
with an average of 36 animals per assay (wild-type), or 9 animals per assay (Prab-
3::fem-3(+)), (B) 4 assays with an average of 25 animals per assay (wild-type), or 17
animals per assay (Prab-3::tra-2(ic)). Statistical significance was determined by
ANOVA with Bonferroni post-hoc tests.
(C) In my model, neural sex, defined as cell-autonomous signals originated from sex
chromosomes in the neuron itself, feminizes or masculinizes the AWA neuron,
modifies the function of the olfactory circuit, and generates sex differences in
olfactory preference.

65
response to AWA odorants (Figure 3.3C). The potential mechanisms underlying
sexual differentiation of AWA neurons by sex determinants may be through
modification on AWA olfactory signaling molecules, synaptic changes, and/or6
differential activity of the circuit (Figure 3.4).
Neural sex regulation on sexually different olfaction is a property of the C.
elegans olfactory circuit
Neural sex generates sexually different olfactory preference in response to
simultaneous presentation of AWA odorants. The extent to which neural sex
establishes sex differences in neural functions is not fully understood. In other words,
I asked if neural sex controls sexually different olfactory preference in response to
other combinations of odorants. I therefore tested sex-transformed animals in
response to two AWC odorants, bu and iaa. By spontaneous chromosomal
integration of an extrachromosomal array, oxEx862[Prab-3::fem-3_mCherry::unc-54
3’UTR], I obtained fsIs15[Prab-3::fem-3_mCherry::unc-54 3’UTR]; him-5 animals.
I first tested fsIs15; him-5 in the da-py assay in which both odorants are detected by
AWA neurons and confirmed that the integrated masculinizing gene works as the
transgene (Figure 3.5A). Then I explored whether sexually different olfactory
preference is revealed in another olfactory preference assay. Among the different
combinations of odorants sensed by AWC neurons, I selected bu-iaa assay in which
the most sex difference between wild-type male and hermaphrodite was observed
(Figure 2.4). Hermaphrodites with pan-neural masculinized neurons (fsIs15; him-5
Herm.) behaved as wild-type males with marginal statistical difference (p-value =
0.059) (Figure 3.5B). This indicates that neural sex is sufficient to determine sexually
different olfactory preference in the bu-iaa assay in which both odorants are detected
by AWC. Therefore, neural sex regulation of behavior is not a specific property of
AWA neurons. Furthermore, this suggests that neural sex control of core neural
functions is a property of C. elegans sexual differentiation.
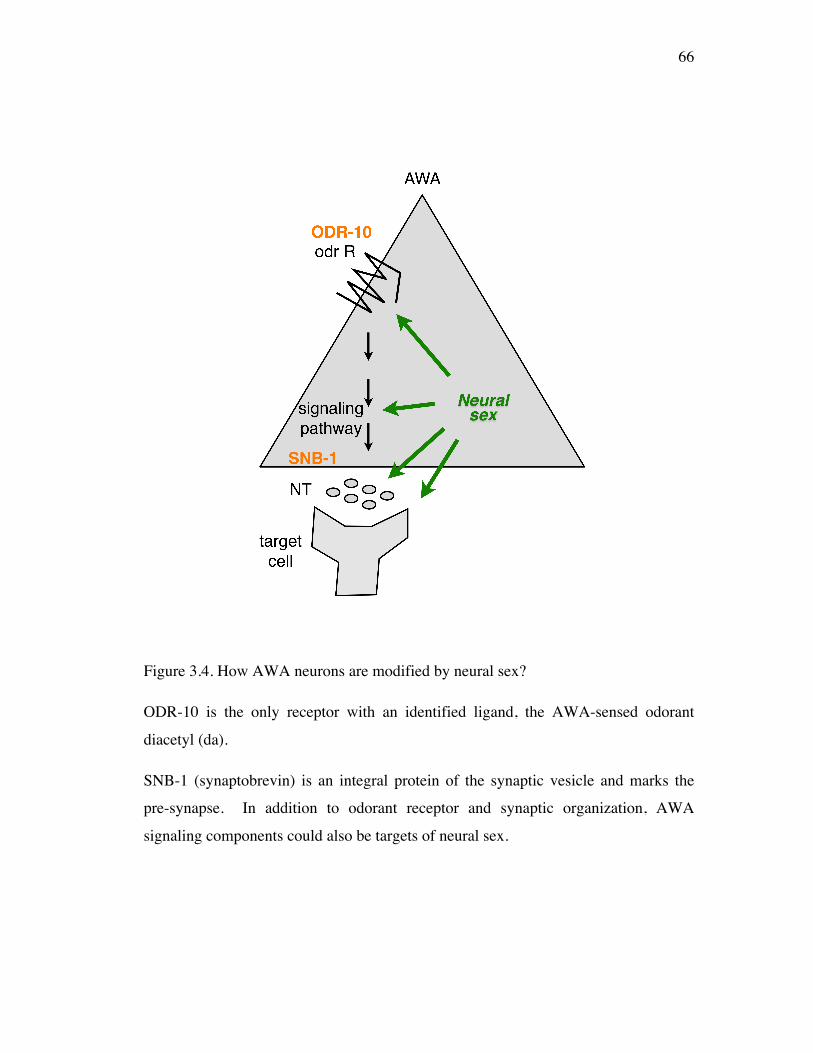
66
Figure 3.4. How AWA neurons are modified by neural sex?
ODR-10 is the only receptor with an identified ligand, the AWA-sensed odorant
diacetyl (da).
SNB-1 (synaptobrevin) is an integral protein of the synaptic vesicle and marks the
pre-synapse. In addition to odorant receptor and synaptic organization, AWA
signaling components could also be targets of neural sex.
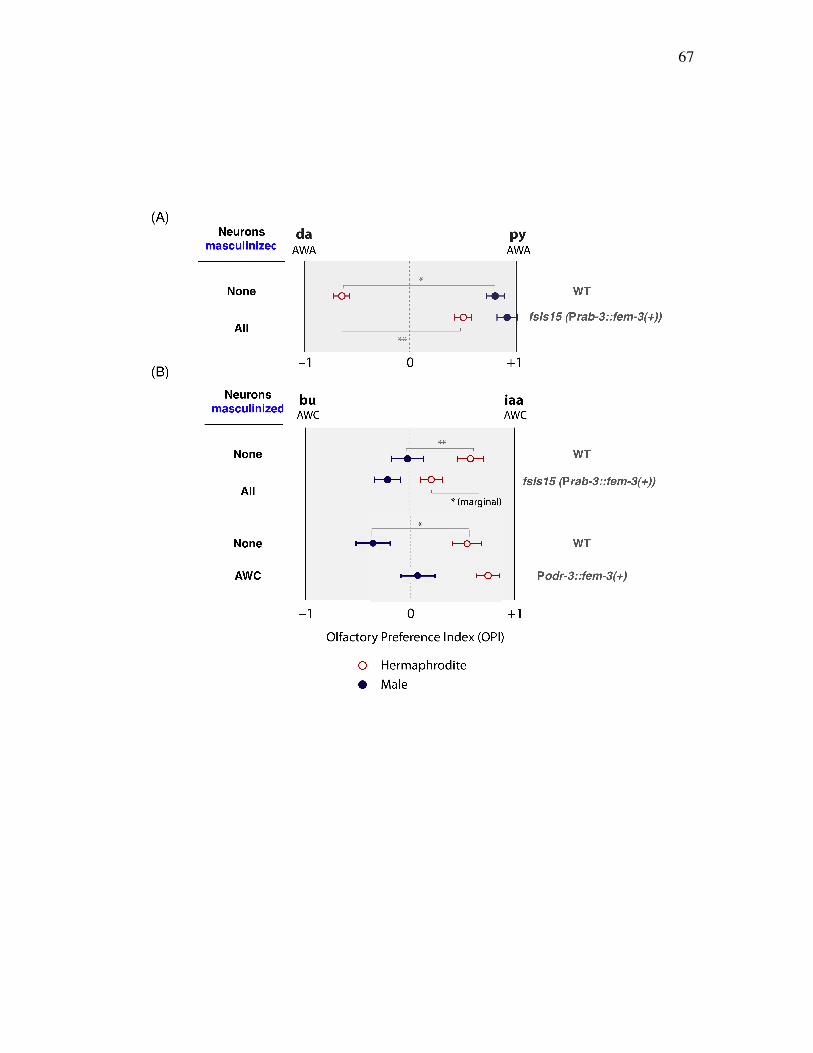
67

68
Figure 3.5. Neural sex regulates sex differences in both AWA and AWC olfactory
preference behaviors
Animals with the integrated transgene fsIs15 (Prab-3::fem-3(+)) were tested in the
da-py (AWA/AWA) assay (A) and in the bu-iaa (AWC/AWC) assay (B) (Top).
Animals carrying the fem-3(+) overexpression transgene: Podr-3::fem-3(+) (fsEx206)
in a small number of neurons including AWC neurons were assayed in the bu-iaa
(AWC/AWC) assay (B) (Bottom).
Each point represents the weighted mean and standard error of (A) 5 assays with an
average of 40 animals per assay (wild-type), or 37 animals per assay (fsIs15), (B)
(Top) 13 assays with an average of 42 animals per assay (wild-type), or 40 animals
per assay (fsIs15) (Bottom) 4 assays with an average of 22 animals per assay (wild-
type), or 16 animals per assay (Podr-3::fem-3(+)). Statistical significance was
determined by ANOVA with Bonferroni post-hoc tests.

69
Since both bu and iaa odorants are sensed by hermaphrodite AWC olfactory neurons,
I speculated that AWC neurons themselves may contribute to sexually different
olfactory preference. To explore if AWA olfactory neurons possess sex differences
that generate sexually different preferences in the bu-iaa (AWC/AWC) assay, I tested
the olfaction of animals in which AWC was sex-reversed. To switch the sexual state
of the AWC neurons, I overexpressed the masculinizing gene fem-3(+) utilizing the
Podr-3 expressed majorly in AWC neurons [Roayaie et al., 1998]. odr-3 encodes a
Gi/Go-like Ga protein that regulates at least in part of the AWC neural fate. I found
that hermaphrodites with masculinized AWC neurons display similar behavior as the
wild-type hermaphrodites (Figure 3.5B). This indicates that masculinization of AWC
neurons is not sufficient to bring about male olfactory preference. Together with the
sufficiency of pan-neural masculinization to masculinize behaviors in the bu-iaa
(AWC/AWC) assay, this suggests that core neurons other than AWC and/or non-cell
autonomous influence from the sex-specific structures are responsible for sexually
different olfactory preference in response to AWC-odorants.
Neural sex modifies a target gene in AWA neurons to bring about sex difference
in olfaction
Although I know that AWA neurons are sufficient for sex-specific AWA-mediated
olfactory preference and AWA neurons must have sexual dimorphism, I do not know
how AWA neurons are sexually different. Therefore, I investigated how AWA
neurons are sexually different by examining how male AWA neurons respond to
hermaphrodite AWA neurons. I already know that males also display attraction
behaviors to the hermaphrodite AWA odorants (Figure 2.3). However, how males
sense those odorants have never been carefully examined. To ask whether male
AWA neurons also detect da and py, as they do in hermaphrodites, I used odr-7 (ky4)
mutants in which AWA function is absent despite intact AWA morphology. odr-7
hermaphrodites fail to respond to da and py [Sengupta et al., 1994]. If male AWA

70
neurons also sense da and py, odr-7 males should also fail to respond to da and py as
odr-7 hermaphrodites do.
I tested male and hermaphrodite responses to each of py 1/100, da 1/100 in
single odorant assays. According to the previous work [Sengupta et al., 1994], odr-7
hermaphrodite response to py 1/100 and da 1/100 is greatly reduced down to C.I. = 0.
I found that both odr-7 hermaphrodites and males exhibited significantly reduced
response to py 1/100 (p < 0.001, p < 0.001) (Figure 3.6A). This indicates that male
AWA neurons are required for py 1/100 response, just as hermaphrodite AWA
neurons are (Figure 3.7).
odr-7 hermaphrodites respond to da 1/100 significantly lower than the wild-type
hermaphrodites with statistical significance (p < 0.05) (Figure 3.6B). However, da
1/100 response of odr-7 males was similar to that of the wild-type males (Figure
3.6B). It reveals that male AWA neurons are not required for da 1/100 response and
suggests that male response to da 1/100 is mediated by other neurons (Figure 3.7).
Together, it suggests that the ability of AWA neurons to mediate da response is a
property of sexual dimorphism in AWA neurons.
The odr-7 hermaphrodite response (C.I. = ~0.5) is AWC neuron-mediated
response to da at this concentration (Figure 3.7) (Chou et al., 2001). Together with
the previous finding that hermaphrodite AWC neurons sense da at high
concentrations ( >= da 1/100), the odr-7 male response (C.I. = ~0.5) may result from
male AWC neurons detecting da at 1/100 (Figure 3.7).
Since male responses to da 1/100 do not require AWA neurons, I asked how
male AWA neurons do not mediate da response. One simple potential mechanism is
that the da receptor ODR-10 is either absent or weakly expressed in male AWA
neurons giving rise to its inability to detect da. Therefore, I began to examine ODR-
10 expression by comparing both odr-10 transcriptional reporter and ODR-10
translational reporter between the sexes.
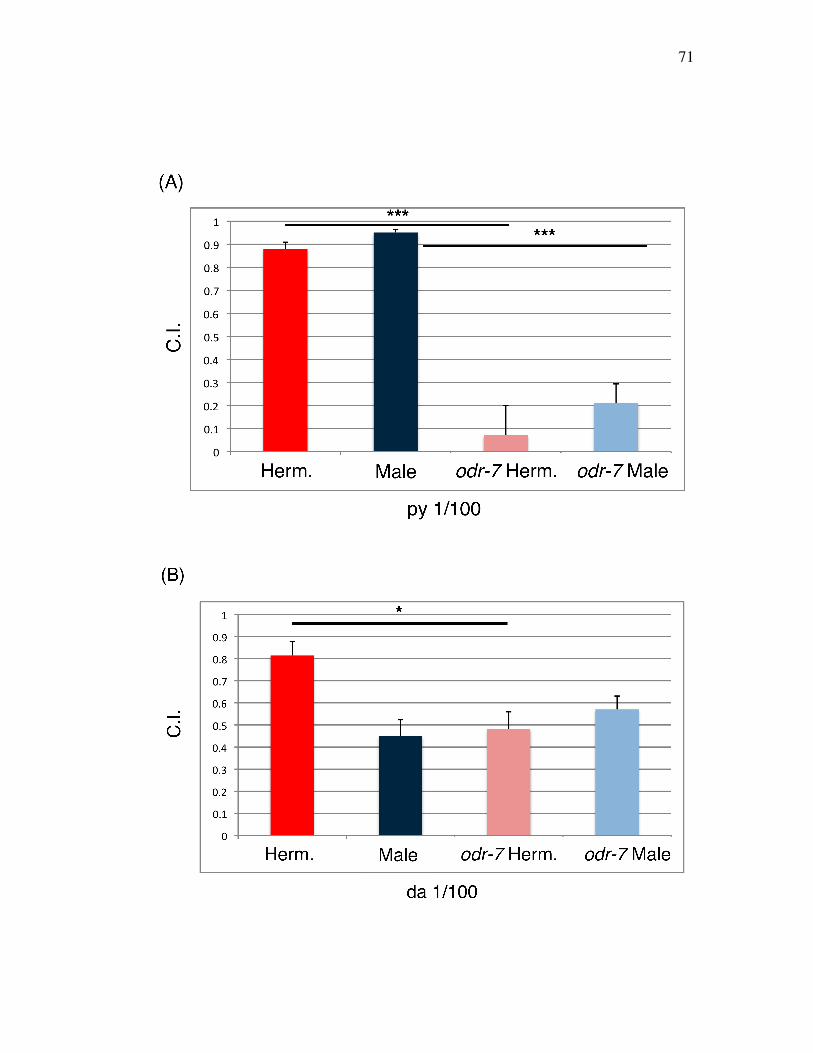
71

72
Figure 3.6. Sexually different mechanisms underlying responses to hermaphrodite
AWA odorants
Young adult wild-type hermaphrodites (red), wild-type males (blue), odr-7 (ky4);
him-5 hermaphrodites (light pink), and odr-7(ky4); him-5 males (light blue) were
assayed in sex-segregated populations in the (A) py 1/100 single odorant assay and
the (B) da 1/100 single odorant assay. Each data point represents the weighted mean
of 4 to 7 assays each containing ~43 animals. Error bars show the weighted SEM.
Statistical significance was determined by ANOVA with Bonferroni post-hoc tests.

73
Figure 3.7. How do male AWA neurons respond to hermaphrodite AWA odorants?
(Top) Male AWA neurons detect py as they do in hermaphrodites. (Bottom) In
contrast to hermaphrodite AWA neurons, male AWA neurons are not required for
response to da. This sex difference in AWA neurons to respond to da could be based
on a sex difference in the expression of the da receptor ODR-10.

74
odr-10 transcriptional reporter gene expression was examined in kyIs37[Podr-
10::GFP]; him-5 animals. I first compared Podr-10::GFP (transcriptional reporter)
expression of ~20 adult worms of each sex. I visually scaled the intensity of GFP
signal of AWA neurons in each worm from 1 (dimmest) to 5 (brightest) and
determined the number of worms at each intensity level. This preliminary study was
performed under the upright compound microscope. Despite bright Podr-10::GFP
expression in hermaphrodites (15/16 had an expression level of 5), male Podr-
10::GFP expressions was mostly dim (9/13 had an expression level of 2) (Figure
3.8B). A representative picture of the Podr-10::GFP expression of both sexes is
shown in Figure 3.8A. This significant sex difference suggests that the promoter
activity of the odr-10 may be sexually different, such that males have lower
transcription of odr-10. In spite of this significant sex difference, I cannot yet
eliminate the possibility that males in general exhibit lower intensities of fluorescence
reporter genes. The ODR-10 protein expression was examined in the kyIs53[Podr-
10::ODR-10::GFP]; him-5 animals under the upright compound microscope. I first
compared Podr-10::ODR-10::GFP (translational reporter) expression between sexes.
In hermaphrodites, bright ODR-10 receptor expression was observed in the ciliary
endings of the AWA dendrites at the tip of the nose with relatively consistent
intensity and was rarely observed in the cell body of AWA neurons as previous
studies have shown [Dwyer et al., 1998]. In males, however, ODR-10 protein
expression was either absent or very weak at the ciliary endings of the AWA neurons
and anywhere else in the amphid (Figure 3.9A).
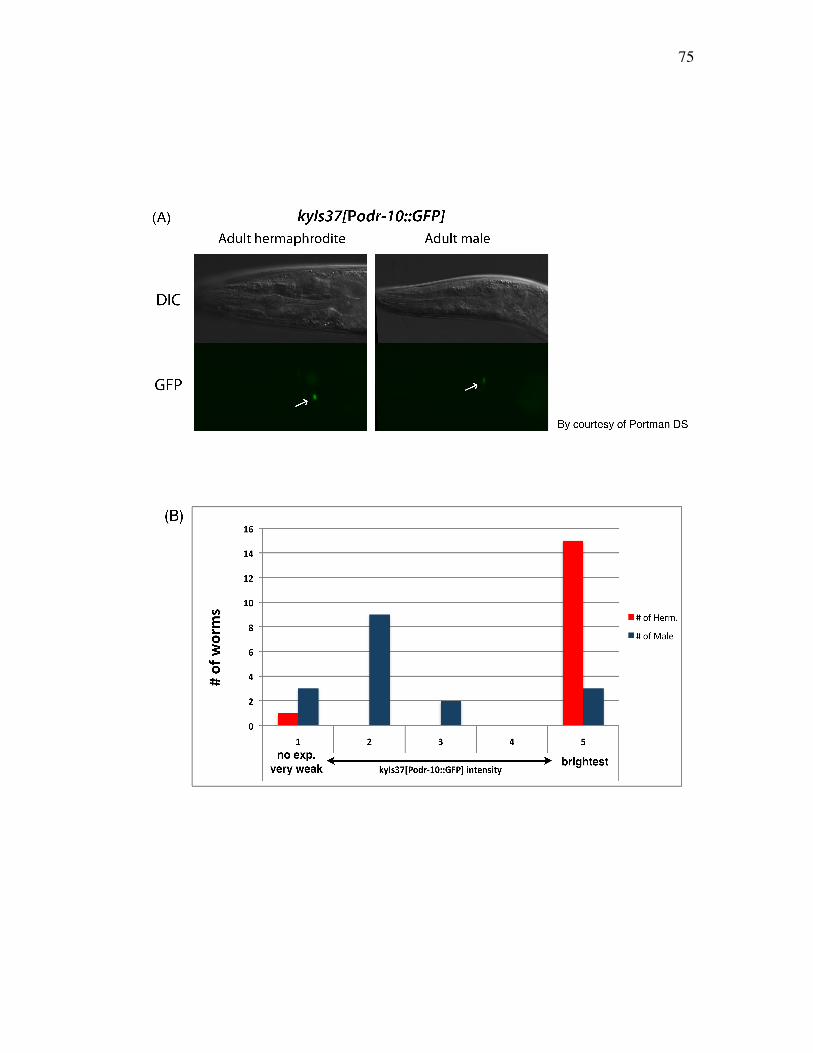
75

76
Figure 3.8. Male odr-10 expression is significantly reduced compared to that of
hermaphrodites
(A) Expression of an integrated Podr-10::GFP transcriptional reporter in wild-type
animals (lateral views). (Top) DIC of an adult hermaphrodite (Left) and an adult
male (Right). (Bottom) GFP of an adult hermaphrodite and an adult male. The solid
arrow indicates the position of the AWA cell body showing Podr-10::GFP
expression.
(B) Visually measured qualitative sex difference in kyIs37[Podr-10::GFP] intensity
is displayed on the X axis as a scale from 1 (dimmest) to 5 (brightest). The number
of worms in each category of brightness is noted on the Y axis. ~20 age-matched
adult worms of each sex were examined.

77
To verify the absence of ODR-10 expression in males with higher magnification, I
imaged males and hermaphrodites using LEICA confocal microscopy. Preliminary
evidence confirmed that males do not express ODR-10 [4/4 males] although
hermaphrodites have strong ODR-10 expression [4/4 hermaphrodites]. To visually
measure the qualitative sex difference in ODR-10 expression, I again scaled the
intensity of the GFP signal of each worm from 1 (dimmest) to 5 (brightest) and
charted the number of worms at each intensity level. Despite bright ODR-10
expression in hermaphrodites [26/33 had signal intensities of 4 or 6], male ODR-10
expression was mostly dim [33/34 showed no signal] (Figure 3.9B). This indicates
that there is a significant sex difference in the expression of ODR-10, the da receptor.
Next, I asked how this dramatic sex difference in ODR-10 expression came
about. I postulated that the neural sex controls ODR-10 expression giving rise to
sexually different level of ODR-10. To test this, I generated the kyIs53; him-5;
fsEx204[Podr-7::tra-2(ic)_mCherry::unc-54 3’UTR] which have both the ODR-10
translational reporter (kyIs53) and the AWA-feminization construct (fsEx204). If
neural sex regulates the ODR-10 expression, males with feminized AWA neurons
should reveal increased ODR-10 level similar to the ODR-10 expression in the wild-
type hermaphrodites. Intriguingly, AWA-feminized males revealed ODR-10
expression not as bright as but similar to the wild-type hermaphrodite ODR-10
expression (Figure 3.10). This indicates that neural sex controls the sexually different
ODR-10 expression. Moreover, this identifies ODR-10 expression as the first
identified molecular property that may regulate sex differences in olfaction.
Together, this work reveals that neural sex controls sexually different expression of a
target gene that may directly give rise to sexually different sensory function.
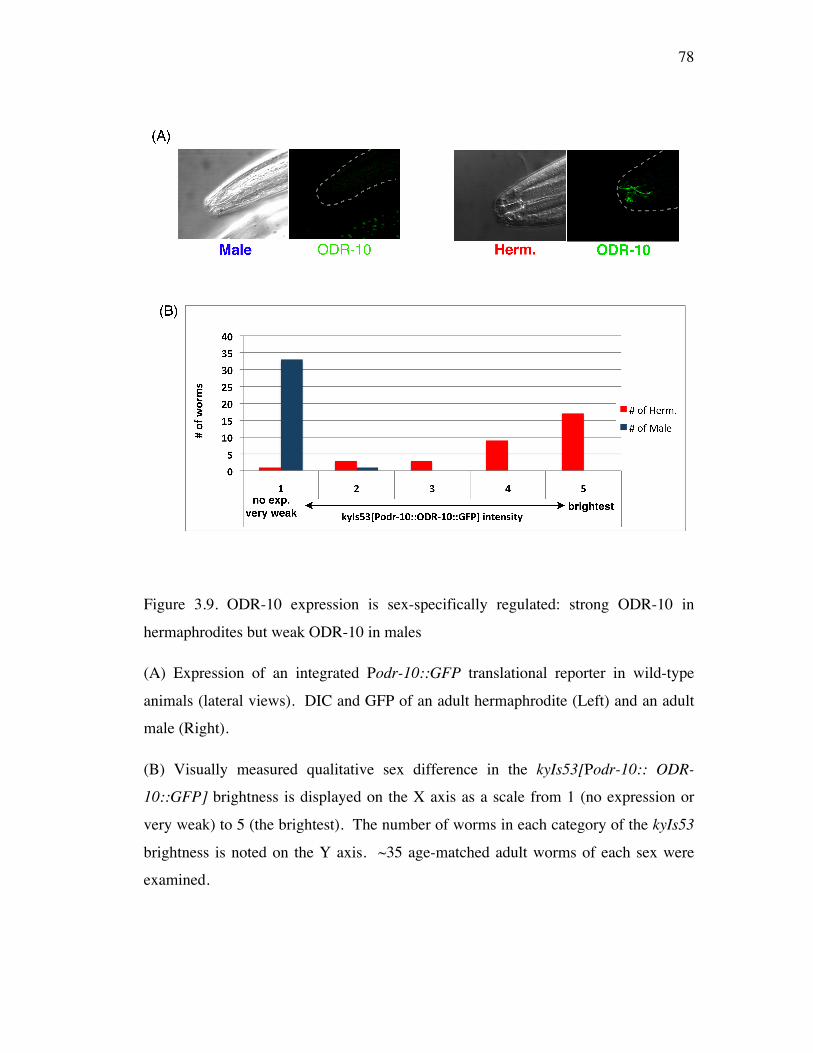
78
Figure 3.9. ODR-10 expression is sex-specifically regulated: strong ODR-10 in
hermaphrodites but weak ODR-10 in males
(A) Expression of an integrated Podr-10::GFP translational reporter in wild-type
animals (lateral views). DIC and GFP of an adult hermaphrodite (Left) and an adult
male (Right).
(B) Visually measured qualitative sex difference in the kyIs53[Podr-10:: ODR-
10::GFP] brightness is displayed on the X axis as a scale from 1 (no expression or
very weak) to 5 (the brightest). The number of worms in each category of the kyIs53
brightness is noted on the Y axis. ~35 age-matched adult worms of each sex were
examined.
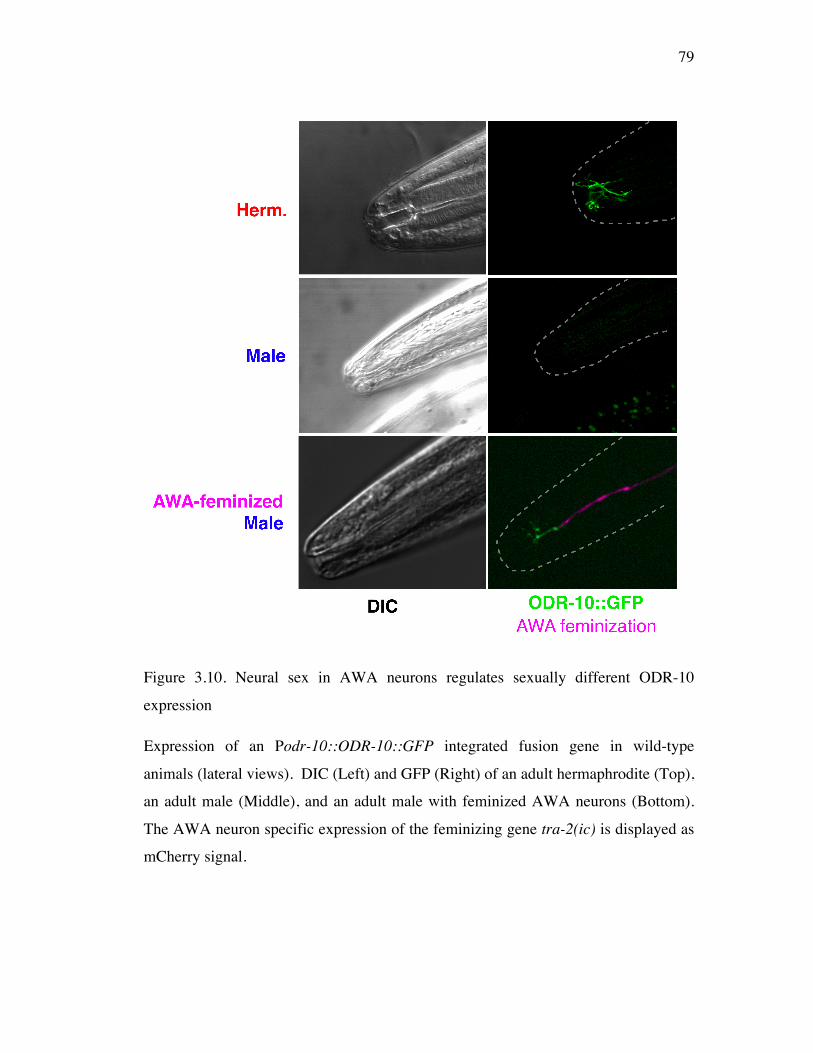
79
Figure 3.10. Neural sex in AWA neurons regulates sexually different ODR-10
expression
Expression of an Podr-10::ODR-10::GFP integrated fusion gene in wild-type
animals (lateral views). DIC (Left) and GFP (Right) of an adult hermaphrodite (Top),
an adult male (Middle), and an adult male with feminized AWA neurons (Bottom).
The AWA neuron specific expression of the feminizing gene tra-2(ic) is displayed as
mCherry signal.

80
4 Discussion
Core sensory circuitry controls a sex difference in olfactory preference
The feasibility of behavioral sex-switching experiments in both directions by sex-
transformation of subsets of neurons reveals two important points. First, both
masculinization and feminization transgenes identified the same sufficient sets of
neurons for sexually different olfactory preference. If any neurons absolutely
required for sex difference in a sensory function were sexually reversed by
masculinization but not by feminization, the sensory function would be sex-
transformed by masculinization but not by feminization. Second, the core neural
circuitry is sufficiently plastic to be modified by neural sex in both directions for
behavioral specialization. Together, sexually different olfactory preference may be
attributed to sex differences in molecular and/or physiological properties of the
sensory neural circuit rather than to sexually different neural connectivity.
Furthermore, it reveals that the cell-intrinsic sex regulators must modify the
properties of core neurons to impart sex differences in the shared sensory function.
I have examined the cellular focus of sex difference in the da-py assay in
which the most significant sex difference in olfactory preference is observed among
five preference assays with different odorant combinations [Lee and Portman, 2007].
Sex differences in olfactory preferences upon exposure to other combinations of
odorants may also be regulated by neural sex. I found that hermaphrodites with
masculinized neurons could also reverse hermaphrodite behavior to males’ in the bu-
iaa (AWC/AWC) assay (Figure 3.5). This indicates that neural sex regulation of sex
differences in olfaction is not limited to the single AWA pair neurons and is a
property of the C. elegans olfactory circuit.
A single sensory neural-switch between hermaphrodite and male olfaction
Sex-transformation of the single sensory neuron pair AWA was sufficient to reverse
the sex phenotype of AWA-mediated olfactory preference in both directions.

81
Masculinization of AWA neurons generated male-like olfactory preference of
otherwise wild-type hermaphrodites and feminization of AWA neurons brought about
hermaphrodite-like olfactory preference of otherwise wild-type males. These results
clearly reveal that neural sex of a single sensory neuron type modifies a shared
sensory behavior.
A decision on the sex phenotype of olfactory preference at the single sensory
neural level may contribute to regulation of innate behaviors across generations.
Since a few sensory neurons are exposed to thousands of other kinds of sensory
stimuli and also informed with internal states, it could be advantageous for a system
to segregate behavioral substrates for two different behaviors at the level of sensory
neurons. This discovery of the sensory neural segregation of two different behaviors
is also observed in behaviors of other systems. This sensory neural role for turning
on a behavior of either sex resembles the mouse pheromone activation of sex-specific
behavior by VNO (vomeronasal organ). In mouse, VNO of each sex determines sex
difference in response to pheromones. Male pheromone increases the females’
receptivity to male sexual behavior although it induces male-male aggression in
males. These different responses are controlled by VNO sensory neural input to the
downstream neural circuits and independent of disruption in estrous cycle or sex
hormones [Kimchi et al., 2007]. Drosophila male courtship song generation also
resembles this mechanism in which sexually different sensory neural input (this case,
either presence or absence of the male-specific input) modifies common postsynaptic
effector machinery to bring about male singing only in male flies [Clyne et al, 2008].
Taken together, my findings that sex differences in sensory neurons presynaptic to
common neural circuitry generates sex differences in a shared behavior may be a
general mechanism for behavioral plasticity at the circuit level and for regulation of
behaviors across generations.
The analogy between systems above suggests that the properties of sensory
receptor neurons responding to sensory stimuli should be sexually different to bring

82
about sex difference in a sensory behavior. The dramatic sex difference in ODR-10
expression may be the direct molecular substrate for sex difference in AWA neural
function including AWA-mediated olfactory preference. The degree of ODR-10
function corresponding to the ODR-10 expression level could affect ODR-10
downstream signaling upon exposure to da. I suggest that the differences in ODR-10
expression lead to changes in PUFA (Poly Unsaturated Fatty Acids)-mediated AWA
olfactory signaling which in turn results in different AWA neural activity. Sex
difference in the AWA neural output to postsynaptic neurons modulating locomotion
would be ultimately displayed as sex difference in the AWA-mediated olfaction.
Together with preliminary results from the measurements that synaptic properties
(connectivity) of AWA neurons are not sexually different (Figure A1.1), I speculate
that sex differences in AWA-mediated olfactory behaviors may rely on the sex
difference in the AWA neurons itself which may be expressed as differential
modulation of AWA synaptic transmission.
A recent study reveals an ability of another olfactory neuron, AWC, to switch
between attraction and repulsion by the activity of olfactory signaling components
[Tsunozaki et al., 2008]. Together with my speculation that a difference in ODR-10
expression level may generate sexually different behaviors, the modulation of critical
sensory neural genes may be a common principle for imparting plasticity to a sensory
behavior.
Neural sex regulates an effector gene critical for the function of a neural circuit
Neural sex controls sex difference in ODR-10 expression: strong ODR-10 expression
in hermaphrodites and weak expression/absence of ODR-10 in males. Feminization
of AWA male neurons could switch very faint male ODR-10 expression to
significantly brighter ODR-10 expression in males similar to the level of the ODR-10
expression in wild-type hermaphrodites. I suggest that the increased ODR-10
expression in feminized male AWA neurons brings about hermaphrodite AWA
functions. Cell-intrinsic sex regulators, tra-1 downstream effectors, must therefore

83
regulate ODR-10 to confer sex differences in AWA neurons. Although the link
between the terminal sex regulator tra-1 and target gene odr-10 is not understood yet
(Figure 3.11), my findings give insights on how chromosomal sex may establish sex
difference in molecular/cellular substrates.
Together, I have found that neural sex modifies the expression of a target gene
in a single sensory neuron to confer sex differences in a shared sensory behavior. Our
study differs from other studies describing mechanisms underlying sex-specific
neuron-regulated sex-specific behaviors. I suggest that cell-intrinsic sex regulators in
a sensory neuron, acting independently from external factors (e.g., sex-specific
structures/functions in the system), modify the activity of sensory neural circuit to
generate either feminized or masculinized shared sensory function. Further work in
this area will shed light on understanding how brain is sexually differentiated to
impart sex differences to common behaviors of the vertebrate system.
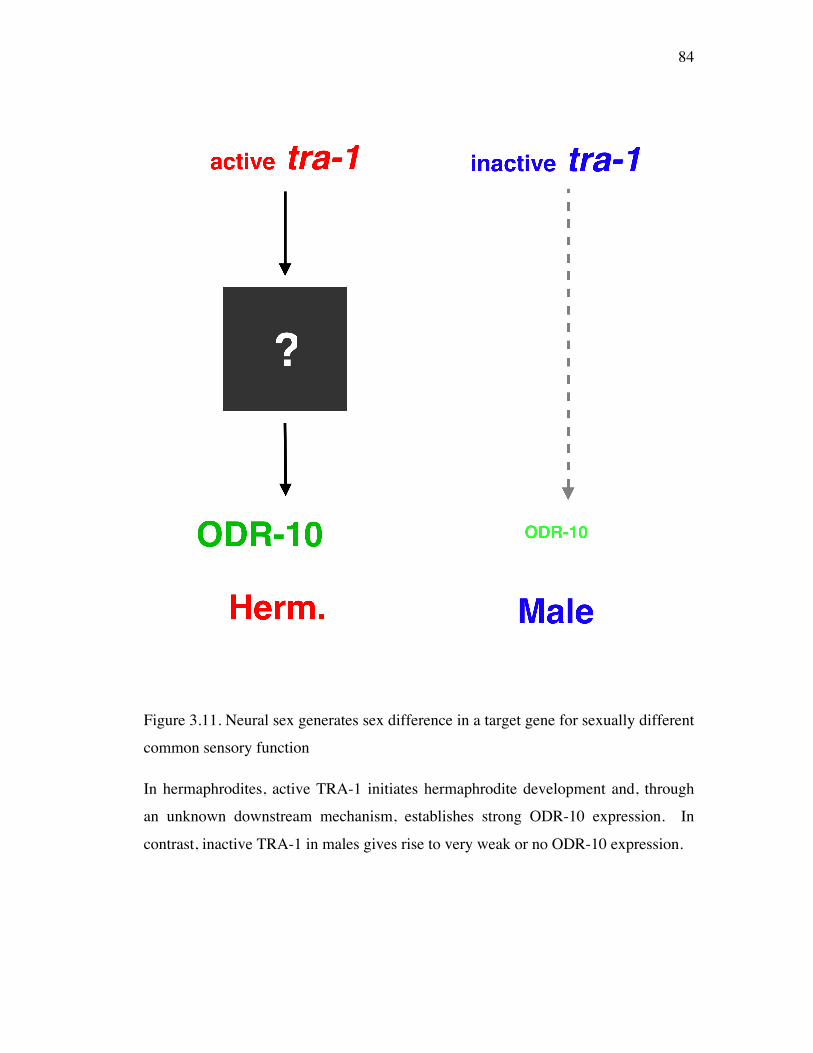
84
Figure 3.11. Neural sex generates sex difference in a target gene for sexually different
common sensory function
In hermaphrodites, active TRA-1 initiates hermaphrodite development and, through
an unknown downstream mechanism, establishes strong ODR-10 expression. In
contrast, inactive TRA-1 in males gives rise to very weak or no ODR-10 expression.

85
Appendix 1 Potential sex difference in AWA connectivity
A simple mechanism underlying AWA-mediated sexually different olfactory
preference may be the sex difference in AWA synaptic properties. I chose to use the
synaptobrevin-1 localization as a read-out of synapse properties. I generated worms,
which specifically express snb-1, synaptobrevin in C. elegans, driven by the AWA-
specific promoter Podr-7. SNB-1 is a synaptic vesicle integral membrane protein and
marks the presynaptic region. In vivo, it shows up as multiple punctate structures. As
a guide to the localization of AWA synapses, the AIY interneuron is displayed with
the otIs133(Pttx-3::rfp). Overall, I found that the qualitative properties of synapses,
namely their number, location, and intensity of puncta (Podr-7::SNB-1_eGFP), were
similar between sexes (Figure A1.1). This indicates that sex difference in AWA-
mediated olfactory preference is not attributed to sex difference in the AWA
connectivity.
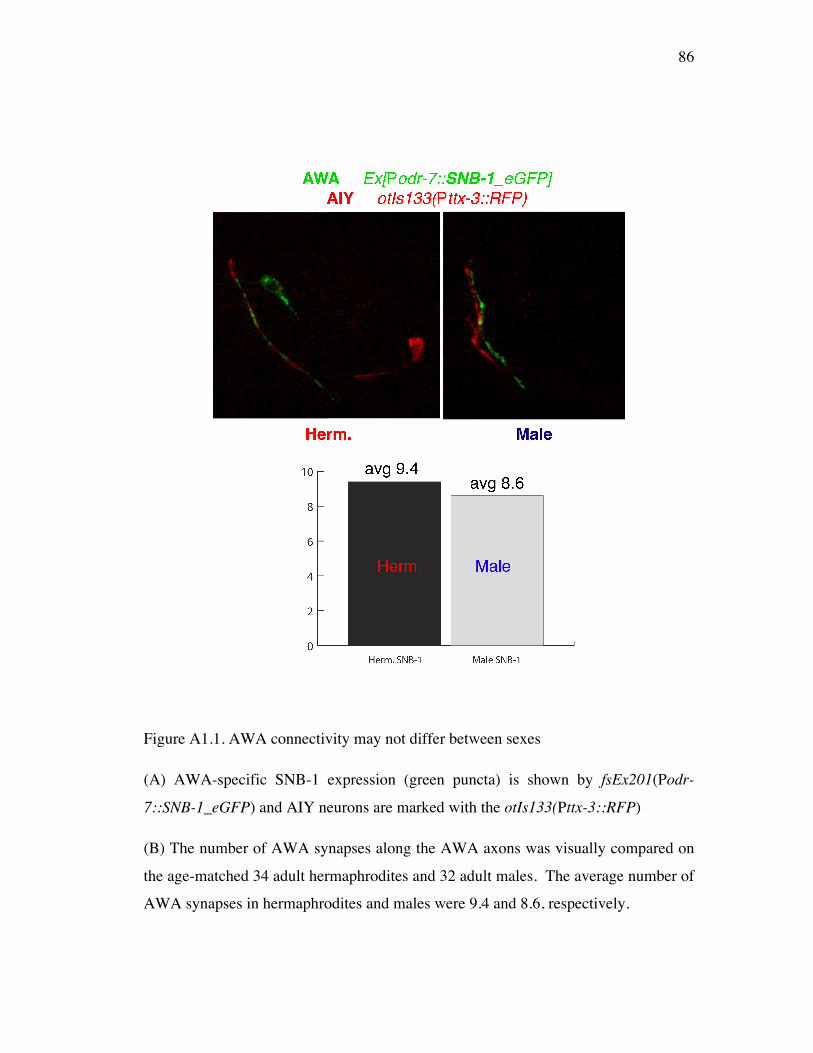
86
Figure A1.1. AWA connectivity may not differ between sexes
(A) AWA-specific SNB-1 expression (green puncta) is shown by fsEx201(Podr-
7::SNB-1_eGFP) and AIY neurons are marked with the otIs133(Pttx-3::RFP)
(B) The number of AWA synapses along the AWA axons was visually compared on
the age-matched 34 adult hermaphrodites and 32 adult males. The average number of
AWA synapses in hermaphrodites and males were 9.4 and 8.6, respectively.

87
Chapter 4 Discussion
The control of sex differences in a C. elegans sensory behavior
I have, for the first time, characterized the significant sex differences in a C. elegans
common sensory behavior, olfaction. These sex differences are controlled by neural
sex, the sexual state of a neuron determined by the cell-intrinsic sex regulators. I
identified that the single sensory neuron pair AWA is sufficient cellular foci to bring
about sexually different AWA-mediated olfactory preference. Furthermore, I
revealed the significant sex difference in the ODR-10 (the da receptor, critical for
AWA function). Finally, I elucidated that the neural sex controls the sex difference in
ODR-10 expression in AWA.
I have found that neural sex controls the expression of a specific target gene,
ODR-10, in the sensory neuron pair AWA (Figure 3.10). However, the functional
role of the dramatic sex difference in ODR-10 expression has yet to be examined.
Increasing the level of ODR-10 expression in male AWAs to the level seen in wild-
type hermaphrodites may bring hermaphrodite-like behavior in the da-py assay
(AWA/AWA). This would indicate that neural sex brings about sexually different
olfactory behaviors through differential regulation on the common sensory genes.
Furthermore, it would reveal that changes in gene expression is directly associated
with modification in a sensory behavior.
Second, the missing link between the cell-intrinsic sex regulators and the
sexually different gene expressions must be addressed (Figure 3.11). Since odr-10
upstream region does not seem to contain a consensus binding site for the terminal
sex regulator TRA-1, it is unlikely that sexually different ODR-10 expression is
directly controlled by TRA-1 activity. TRA-1 downstream effectors, the conserved
DM domain genes, are the likely upstream regulators of ODR-10. These DM
(doublesex and mab-3)-domain genes specify partially overlapping subsets of male
characteristics in C. elegans [Lints and Emmons, 2002; Shen and Hodgkin, 1988; Yi

88
et al., 2000; Mason et al., 2008]. By epistatic analysis (genetic interaction tests) on
DM domain genes and ODR-10 expression, the role of DM domain genes in the
sexually different target gene expressions may be revealed. If these conserved DM
domain genes, known to regulate sex differentiation in other metazoans [Zarkower,
2006], modulate ODR-10 expression, it would suggest that neural sex regulation on a
critical signaling gene is the common principle to generate sexually different sensory
behaviors in different animal systems.
Last, the full extent of neural sex regulation on behavior has not yet been
thoroughly explored. I defined the AWA neurons as a sufficient cellular focus of the
sex difference in AWA-mediated olfactory preference behavior. Our results in four
other olfactory preference settings (Figure 2.4) show significant sex differences,
suggesting that other neurons may also have sexually different properties established
by neural sex. The neural sex control on the behavioral sex in AWC/AWC olfactory
preference (Figure 3.5) supports this idea. Testing sex-transformed animals in
different kinds of olfactory preference assays will widen the spatial map of neural sex
control on sensory behaviors. Sex-transformation on combinations of different single
neurons or subsets of sensory neurons and interneurons will reveal the complete
neural components for the full extent of sex difference in olfactory preferences.
The temporal activity of neural sex is another intriguing question (Figure
2.6B). Understanding when neural sex is established will tell me if the determination
on the sex phenotype of a behavior is based on adult behavioral plasticity or
developmental control. If switching neural sex in adult worms is sufficient to reverse
behavioral sex, it indicates that neural circuit plasticity in the adult (i.e., including
changes in the molecular properties of already established neural circuits) may give
rise to sexually different behaviors. Otherwise it suggests that some of
developmental sexual differentiation is critical for sexually different behaviors.
Furthermore, it is possible that many other target genes critical for neural function are
regulated by neural sex. The AWA neurons may have sex differences in the

89
expression of other olfactory signaling genes in addition to ODR-10. To elucidate the
spectrum of molecular sex differences in any neural focus of behavior, systemic
comparisons of mRNA profiles of single neurons or subsets of sensory neurons could
be useful. This approach may identify discrete sex differences in gene expressions.
Insights on sexual dimorphisms in neurological diseases
Increasing attention on the significant sex differences in many aspects of neurological
diseases has captured the interest of both physicians and neurobiologists. However,
with the complexity of brain diseases and the nervous system itself, the etiology of
many neurological diseases is poorly understood. Studying how the nervous system
is sexually differentiated utilizing simple invertebrate animal models will yield
insights on the sex-biases in multiple brain diseases of the vertebrates and human and
on the development of better treatments for neurological diseases.
Cell-intrinsic regulation on sexually different gene expressions may be substrates of
the sex-biases in diseases
Historically, epidemiological findings reported that sex hormones control sex
differences in the pathologies of diseases and responses to therapeutics [Grady et al.,
1992]. However, recent clinical reports highlight the role of cell-intrinsic sex
regulators of sex differences in brain diseases, independently from the role of sex
hormones. Sex differences in post-ischemic brain damages and the recovery were
observed in the developing brains of child patients before sexual maturation.
Interestingly, the cell death pathways acting upon the ischemic injury in the
developing rat brain (postnatal day 7) exhibited significant sex differences. These
indicate that prominent sex differences exist in the developing brain that contribute to
sex differences in the pathology of brain diseases [Renolleau et al., 2008; Hurn et al.,
2005; Edwards, 2004]. However, the exact mechanism of sex-specific regulation on
cell death pathways has not been understood. Our finding on neural sex-regulated
gene expression (ODR-10, critical for sensory function) suggests that chromosomal

90
sex may somehow modify conventional cell death pathways to give rise to sexually
different pathogenesis upon brain injury.
Sex differences in gene expressions may give rise to sex differences in behavioral
symptoms of diseases
Many neurological diseases that occurr with sex-biased incidence are
associated with a variety of disrupted behaviors. As I discussed in Chapter 1, for
example, autism patients and autism mouse models both display a plethora of
disrupted social behaviors only in males [Schneider et al., 2008]. In addition,
epigenetic control on sexually different gene expression (i.e., MECP2, FMR1, ATRX
and JARID1C) in both human and mouse models is suggested as a mechanism for
sex-biases in various aspects of neurobehavioral disease [Tsai et al., 2009; Shibayama
et al., 2004; Iwase et al, 2007; Hagerman et al., 2006; Berube et al, 2002; Weaving et
al., 2004]. Our studies on how sex differences in gene expressions modify behaviors
give insights on the mechanisms underlying sex-biased behavioral symptoms of
multiple neuropsychiatric disorders.

91
References
Agate, R.J., Grisham, W., Wade, J., Mann, S., Wingfield, J., Schanen, C., Palotie, A., and Arnold, A.P. (2003). Neural, not gonadal, origin of brain sex differences in a gynandromorphic finch. Proc Natl Acad Sci U S A 100, 4873-4878.
Arnold, A.P. (2004). Sex chromosomes and brain gender. Nat Rev Neurosci 5, 701-708.
Bany, I., Dong, M., and Koelle, M. (2003). Genetic and cellular basis for acetylcholine inhibition of Caenorhabditis elegans egg-laying behavior. J Neurosci 23, 8060-8069.
Bargmann, C.I. (2006). Chemosensation in C. elegans. WormBook.
Bargmann, C.I., Hartwieg, E., and Horvitz, H.R. (1993). Odorant-selective genes and neurons mediate olfaction in C. elegans. Cell 74, 515-527.
Barr, M.M., and Sternberg, P.W. (1999). A polycystic kidney-disease gene homologue required for male mating behaviour in C. elegans. Nature 401, 386-389.
Becker, J.B., Arnold, A.P., Berkley, K.J., Blaustein, J.D., Eckel, L.A., Hampson, E., Herman, J.P., Marts, S., Sadee, W., Steiner, M., et al. (2005). Strategies and methods for research on sex differences in brain and behavior. Endocrinology 146, 1650-1673.
Berube, N.G., Jagla, M., Smeenk, C., De Repentigny, Y., Kothary, R., and Picketts, D.J. (2002). Neurodevelopmental defects resulting from ATRX overexpression in transgenic mice. Hum Mol Genet 11, 253-261.
Billeter, J., Rideout, E., Dornan, A., and Goodwin, S. (2006). Control of male sexual behavior in Drosophila by the sex determination pathway. Curr Biol 16, R766-776.
Brann, D.W., Dhandapani, K., Wakade, C., Mahesh, V.B., and Khan, M.M. (2007). Neurotrophic and neuroprotective actions of estrogen: basic mechanisms and clinical implications. Steroids 72, 381-405.
Bratus, A., and Slota, E. (2006). DMRT1/Dmrt1, the sex determining or sex differentiating gene in Vertebrata. Folia Biol (Krakow) S 54, 81-86.
Brenner, S. (1974). The genetics of Caenorhabditis elegans. Genetics 77, 71-94.
Cahill, L. (2006). Why sex matters for neuroscience. Nat Rev Neurosci 7, 477-484.
Cahill, L., Uncapher, M., Kilpatrick, L., Alkire, M.T., and Turner, J. (2004). Sex-

92
related hemispheric lateralization of amygdala function in emotionally influenced memory: an FMRI investigation. Learn Mem 11, 261-266.
Carruth, L.L., Reisert, I., and Arnold, A.P. (2002). Sex chromosome genes directly affect brain sexual differentiation. Nat Neurosci 5, 933-934.
Certel, S., Savella, M., Schlegel, D., and Kravitz, E. (2007). Modulation of Drosophila male behavioral choice. Proc Natl Acad Sci U S A 104, 4706-4711.
Chalasani, S., Chronis, N., Tsunozaki, M., Gray, J., Ramot, D., Goodman, M., and Bargmann, C. (2007). Dissecting a circuit for olfactory behaviour in Caenorhabditis elegans. Nature 450, 63-70.
Chalfie, M., Horvitz, H.R., and Sulston, J.E. (1981). Mutations that lead to reiterations in the cell lineages of C. elegans. Cell 24, 59-69.
Chou, J., Bargmann, C., and Sengupta, P. (2001). The Caenorhabditis elegans odr-2 gene encodes a novel Ly-6-related protein required for olfaction. Genetics 157, 211-224.
Colosimo, M.E., Brown, A., Mukhopadhyay, S., Gabel, C., Lanjuin, A.E., Samuel, A.D., and Sengupta, P. (2004). Identification of Thermosensory and Olfactory Neuron-Specific Genes via Expression Profiling of Single Neuron Types. Curr Biol 14, 2245-2251.
Colosimo, M.E., Tran, S., and Sengupta, P. (2003). The divergent orphan nuclear receptor ODR-7 regulates olfactory neuron gene expression via multiple mechanisms in Caenorhabditis elegans. Genetics 165, 1779-1791.
Conradt, B., and Horvitz, H.R. (1998). The C. elegans protein EGL-1 is required for programmed cell death and interacts with the Bcl-2-like protein CED-9. Cell 93, 519-529.
Cooke, B., Hegstrom, C.D., Villeneuve, L.S., and Breedlove, S.M. (1998). Sexual differentiation of the vertebrate brain: principles and mechanisms. Front Neuroendocrinol 19, 323-362.
Cooke, B.M., and Woolley, C.S. (2005). Sexually dimorphic synaptic organization of the medial amygdala. J Neurosci 25, 10759-10767.
Cornelius, M.L., Duan, J.J., and Messing, R.H. (2000). Volatile host fruit odors as attractants for the oriental fruit fly (Diptera: Tephritidae). J Econ Entomol 93, 93-100.
Datta, S.R., Vasconcelos, M.L., Ruta, V., Luo, S., Wong, A., Demir, E., Flores, J., Balonze, K., Dickson, B.J., and Axel, R. (2008). The Drosophila pheromone cVA

93
activates a sexually dimorphic neural circuit. Nature 452, 473-477.
de Bono, M., Zarkower, D., and Hodgkin, J. (1995). Dominant feminizing mutations implicate protein-protein interactions as the main mode of regulation of the nematode sex-determining gene tra-1. Genes Dev 9, 155-167.
Dewing, P., Chiang, C.W., Sinchak, K., Sim, H., Fernagut, P.O., Kelly, S., Chesselet, M.F., Micevych, P.E., Albrecht, K.H., Harley, V.R., et al. (2006). Direct Regulation of Adult Brain Function by the Male-Specific Factor SRY. Curr Biol 16, 415-420.
Dewing, P., Shi, T., Horvath, S., and Vilain, E. (2003). Sexually dimorphic gene expression in mouse brain precedes gonadal differentiation. Brain Res Mol Brain Res 118, 82-90.
Doniach, T., and Hodgkin, J. (1984). A sex-determining gene, fem-1, required for both male and hermaphrodite development in Caenorhabditis elegans. Dev Biol 106, 223-235.
Dwyer, N.D., Troemel, E.R., Sengupta, P., and Bargmann, C.I. (1998). Odorant receptor localization to olfactory cilia is mediated by ODR-4, a novel membrane-associated protein. Cell 93, 455-466.
Edwards, D. (2004). Brain protection for girls and boys. J Pediatr 145, 723-724.
Ember, C.R., and Ember, M. (2003). Encyclopedia of sex and gender : men and women in the world's cultures, (New York: Kluwer Academic/Plenum Publishers).
Erdman, S., and Burtis, K. (1993). The Drosophila doublesex proteins share a novel zinc finger related DNA binding domain. EMBO J 12, 527-535.
Fleisher, A., Grundman, M., Jack, C.R., Jr., Petersen, R.C., Taylor, C., Kim, H.T., Schiller, D.H., Bagwell, V., Sencakova, D., Weiner, M.F., et al. (2005). Sex, apolipoprotein E epsilon 4 status, and hippocampal volume in mild cognitive impairment. Arch Neurol 62, 953-957.
Gahr, M. (2003). Male Japanese quails with female brains do not show male sexual behaviors. Proc Natl Acad Sci U S A 100, 7959-7964.
Gatewood, J.D., Wills, A., Shetty, S., Xu, J., Arnold, A.P., Burgoyne, P.S., and Rissman, E.F. (2006). Sex chromosome complement and gonadal sex influence aggressive and parental behaviors in mice. J Neurosci 26, 2335-2342.
Goldman, S.A., and Nottebohm, F. (1983). Neuronal production, migration, and differentiation in a vocal control nucleus of the adult female canary brain. Proc Natl Acad Sci U S A 80, 2390-2394.

94
Goodman, M.B. (2006). Mechanosensation. WormBook.
Gorski, R.A., Gordon, J.H., Shryne, J.E., and Southam, A.M. (1978). Evidence for a morphological sex difference within the medial preoptic area of the rat brain. Brain Res 148, 333-346.
Grady, D.a.o. (1992). Hormone therpay to prevent disease and prolong life in postmenopausal women. Ann Intern Med 117, 1016-1037.
Gray, J.M., Hill, J.J., and Bargmann, C.I. (2005). A circuit for navigation in Caenorhabditis elegans. Proc Natl Acad Sci U S A 102, 3184-3191.
Gruninger, T., Gualberto, D., LeBoeuf, B., and Garcia, L. (2006). Integration of male mating and feeding behaviors in Caenorhabditis elegans. J Neurosci S 26, 169-179.
Hagerman, R.J. (2006). Lessons from fragile X regarding neurobiology, autism, and neurodegeneration. J Dev Behav Pediatr 27, 63-74.
Hall, D., Lints, R., and Altun, Z. (2006). Nematode neurons: anatomy and anatomical methods in Caenorhabditis elegans. Int Rev Neurobiol S 69, 1-35.
Hall, J.C. (1977). Portions of the central nervous system controlling reproductive behavior in Drosophila melanogaster. Behav Genet 7, 291-312.
Hammock, E.A., and Young, L.J. (2006). Oxytocin, vasopressin and pair bonding: implications for autism. Philos Trans R Soc Lond B Biol Sci 361, 2187-2198.
Hartley, J.L., Temple, G.F., and Brasch, M.A. (2000). DNA cloning using in vitro site-specific recombination. Genome Res 10, 1788-1795.
Haycraft, C.J., Swoboda, P., Taulman, P.D., Thomas, J.H., and Yoder, B.K. (2001). The C. elegans homolog of the murine cystic kidney disease gene Tg737 functions in a ciliogenic pathway and is disrupted in osm-5 mutant worms. Development 128, 1493-1505.
Hodgkin, J. (1987). A genetic analysis of the sex-determining gene, tra-1, in the nematode Caenorhabditis elegans. Genes Dev 1, 731-745.
Hodgkin, J., and Brenner, S. (1977). Mutations causing transformation of sexual phenotype in the nematode Caenorhabditis elegans. Genetics 86, 275-287.
Hong, C.S., Park, B.Y., and Saint-Jeannet, J.P. (2007). The function of Dmrt genes in vertebrate development: it is not just about sex. Dev Biol 310, 1-9.
Horvitz, H.R., and Sulston, J.E. (1980). Isolation and genetic characterization of cell-lineage mutants of the nematode Caenorhabditis elegans. Genetics 96, 435-454.

95
Hunter, C.P., and Wood, W.B. (1990). The tra-1 gene determines sexual phenotype cell-autonomously in C. elegans. Cell 63, 1193-1204.
Hurn, P.D., Vannucci, S.J., and Hagberg, H. (2005). Adult or perinatal brain injury: does sex matter? Stroke 36, 193-195.
Ishihara, T., Iino, Y., Mohri, A., Mori, I., Gengyo-Ando, K., Mitani, S., and Katsura, I. (2002). HEN-1, a secretory protein with an LDL receptor motif, regulates sensory integration and learning in Caenorhabditis elegans. Cell 109, 639-649.
Iwase, S., Lan, F., Bayliss, P., de la Torre-Ubieta, L., Huarte, M., Qi, H.H., Whetstine, J.R., Bonni, A., Roberts, T.M., and Shi, Y. (2007). The X-linked mental retardation gene SMCX/JARID1C defines a family of histone H3 lysine 4 demethylases. Cell 128, 1077-1088.
Juraska, J.M. (1991). Sex differences in "cognitive" regions of the rat brain. Psychoneuroendochrinology 16, 105-109.
Juraska, J.M. (1991). Sex differences in "cognitive" regions of the rat brain. Psychoneuroendocrinology 16, 105-109.
Juraska, J.M., Fitch, J.M., Henderson, C., and Rivers, N. (1985). Sex differences in the dendritic branching of dentate granule cells following differential experience. Brain Res 333, 73-80.
Kahn-Kirby, A., Dantzker, J., Apicella, A., Schafer, W., Browse, J., Bargmann, C., and Watts, J. (2004). Specific polyunsaturated fatty acids drive TRPV-dependent sensory signaling in vivo. Cell 119, 889-900.
Kimble, J., and Crittenden, S.L. (2005). Germline proliferation and its control. WormBook, 1-14.
Kimble, J., Edgar, L., and Hirsh, D. (1984). Specification of male development in Caenorhabditis elegans: the fem genes. Dev Biol 105, 234-239.
Kimble, J.E., and White, J.G. (1981). On the control of germ cell development in Caenorhabditis elegans. Dev Biol 81, 208-219.
Kimchi, T., Xu, J., and Dulac, C. (2007). A functional circuit underlying male sexual behaviour in the female mouse brain. Nature 448, 1009-1014.
Kimura, K., Hachiya, T., Koganezawa, M., Tazawa, T., and Yamamoto, D. (2008). Fruitless and doublesex coordinate to generate male-specific neurons that can initiate courtship. Neuron 59, 759-769.

96
Klass, M., Wolf, N., and Hirsh, D. (1976). Development of the male reproductive system and sexual transformation in the nematode Caenorhabditis elegans. Dev Biol 52, 1-18.
Kleemann, G., Jia, L., and Emmons, S. (2008). Regulation of Caenorhabditis elegans Male Mate Searching Behavior by the Nuclear Receptor DAF-12. Genetics.
Komatsu, H., Jin, Y.H., L'Etoile, N., Mori, I., Bargmann, C.I., Akaike, N., and Ohshima, Y. (1999). Functional reconstitution of a heteromeric cyclic nucleotide-gated channel of Caenorhabditis elegans in cultured cells. Brain Res 821, 160-168.
Komatsu, H., Mori, I., Rhee, J., Akaike, N., and Ohshima, Y. (1996). Mutations in a cyclic nucleotide-gated channel lead to abnormal thermosensation and chemosensation in C. elegans. Neuron 17, 707-718.
Korol, D.L. (2004). Role of estrogen in balancing contributions from multiple memory systems. Neurobiol Learn Mem 82, 309-323.
Kuwabara, P.E., and Kimble, J. (1995). A predicted membrane protein, TRA-2A, directs hermaphrodite development in Caenorhabditis elegans. Development 121, 2995-3004.
Lee, K., and Portman, D. (2007). Neural sex modifies the function of a C. elegans sensory circuit. Curr Biol 17, 1858-1863.
Levine, S. (1966). Sex-role identification and parental perceptions of social competence. Am J Ment Defic 70, 822-824.
Lints, R., and Emmons, S.W. (2002). Regulation of sex-specific differentiation and mating behavior in C. elegans by a new member of the DM domain transcription factor family. Genes Dev 16, 2390-2402.
Madeira, M.D., and Lieberman, A.R. (1995). Sexual dimorphism in the mammalian limbic system. Prog Neurobiol 45, 275-333.
Madl, J., and Herman, R. (1979). Polyploids and sex determination in Caenorhabditis elegans. Genetics 93, 393-402.
Manoli, D., Meissner, G., and Baker, B. (2006). Blueprints for behavior: genetic specification of neural circuitry for innate behaviors. Trends Neurosci 29, 444-451.
Maricq, A.V., Peckol, E., Driscoll, M., and Bargmann, C.I. (1995). Mechanosensory signalling in C. elegans mediated by the GLR-1 glutamate receptor. Nature 378, 78-81.

97
Mason, D., Rabinowitz, J., and Portman, D. (2008). dmd-3, a doublesex-related gene regulated by tra-1, governs sex-specific morphogenesis in C. elegans. Development 135, 2373-2382.
McEwen, B.S. (2000). The neurobiology of stress: from serendipity to clinical relevance. Brain Res 886, 172-189.
McPhie-Lalmansingh, A.A., Tejada, L.D., Weaver, J.L., and Rissman, E.F. (2008). Sex chromosome complement affects social interactions in mice. Horm Behav 54, 565-570.
Mehra, A., Gaudet, J., Heck, L., Kuwabara, P.E., and Spence, A.M. (1999). Negative regulation of male development in Caenorhabditis elegans by a protein-protein interaction between TRA-2A and FEM-3. Genes Dev 13, 1453-1463.
Miller, A., Thiele, T., Faumont, S., Moravec, M., and Lockery, S. (2005). Step-response analysis of chemotaxis in Caenorhabditis elegans. J Neurosci 25, 3369-3378.
Morris, J.A., Jordan, C.L., and Breedlove, S.M. (2004). Sexual differentiation of the vertebrate nervous system. Nat Neurosci 7, 1034-1039.
Nelson, G., Lew, K., and Ward, S. (1978). Intersex, a temperature-sensitive mutant of the nematode Caenorhabditis elegans. Dev Biol 66, 386-409.
Nishizawa, S., Benkelfat, C., Young, S.N., Leyton, M., Mzengeza, S., de Montigny, C., Blier, P., and Diksic, M. (1997). Differences between males and females in rates of serotonin synthesis in human brain. Proc Natl Acad Sci U S A 94, 5308-5313.
Nonet, M., Staunton, J., Kilgard, M., Fergestad, T., Hartwieg, E., Horvitz, H., Jorgensen, E., and Meyer, B. (1997). Caenorhabditis elegans rab-3 mutant synapses exhibit impaired function and are partially depleted of vesicles. J Neurosci 17, 8061-8073.
Nopoulos, P., Flaum, M., and Andreasen, N.C. (1997). Sex differences in brain morphology in schizophrenia. Am J Psychiatry 154, 1648-1654.
O'Hagan, R., and Chalfie, M. (2006). Mechanosensation in Caenorhabditis elegans. Int Rev Neurobiol 69, 169-203.
Phoenix, C.H., Goy, R.W., Gerall, A.A., and Young, W.C. (1959). Organizing action of prenatally administered testosterone propionate on the tissues mediating mating behavior in the female guinea pig. Endocrinology 65, 369-382.
Pierce-Shimomura, J.T., Dores, M., and Lockery, S.R. (2005). Analysis of the effects

98
of turning bias on chemotaxis in C. elegans. J Exp Biol 208, 4727-4733.
Pierce-Shimomura, J.T., Morse, T.M., and Lockery, S.R. (1999). The fundamental role of pirouettes in Caenorhabditis elegans chemotaxis. J Neurosci 19, 9557-9569.
Portman, D. (2007). Genetic Control of Sex Differences in C. elegans Neurobiology and Behavior. Adv Genet 59, 1-37.
Qin, H., Rosenbaum, J.L., and Barr, M.M. (2001). An autosomal recessive polycystic kidney disease gene homolog is involved in intraflagellar transport in C. elegans ciliated sensory neurons. Curr Biol 11, 457-461.
Raymond, C.S., Shamu, C.E., Shen, M.M., Seifert, K.J., Hirsch, B., Hodgkin, J., and Zarkower, D. (1998). Evidence for evolutionary conservation of sex-determining genes. Nature 391, 691-695.
Reisert, I., and Pilgrim, C. (1991). Sexual differentiation of monoaminergic neurons--genetic or epigenetic? Trends Neurosci 14, 468-473.
Remy, J.J., and Hobert, O. (2005). An interneuronal chemoreceptor required for olfactory imprinting in C. elegans. Science 309, 787-790.
Renolleau, S., Fau, S., and Charriaut-Marlangue, C. (2008). Gender-related differences in apoptotic pathways after neonatal cerebral ischemia. Neuroscientist 14, 46-52.
Rideout, E.J., Billeter, J.C., and Goodwin, S.F. (2007). The sex-determination genes fruitless and doublesex specify a neural substrate required for courtship song. Curr Biol 17, 1473-1478.
Roayaie, K., Crump, J.G., Sagasti, A., and Bargmann, C.I. (1998). The G alpha protein ODR-3 mediates olfactory and nociceptive function and controls cilium morphogenesis in C. elegans olfactory neurons. Neuron 20, 55-67.
Schafer, W. (2006). Genetics of egg-laying in worms. Annu Rev Genet 40, 487-509.
Schneider, T., Roman, A., Basta-Kaim, A., Kubera, M., Budziszewska, B., Schneider, K., and Przewlocki, R. (2008). Gender-specific behavioral and immunological alterations in an animal model of autism induced by prenatal exposure to valproic acid. Psychoneuroendocrinology 33, 728-740.
Segal, S.P., Graves, L.E., Verheyden, J., and Goodwin, E.B. (2001). RNA-Regulated TRA-1 nuclear export controls sexual fate. Dev Cell 1, 539-551.
Sengupta, P. (2007). Generation and modulation of chemosensory behaviors in C.

99
elegans. Pflugers Arch 454, 721-734.
Sengupta, P., Chou, J.H., and Bargmann, C.I. (1996). odr-10 encodes a seven transmembrane domain olfactory receptor required for responses to the odorant diacetyl. Cell 84, 899-909.
Sengupta, P., Colbert, H.A., and Bargmann, C.I. (1994). The C. elegans gene odr-7 encodes an olfactory-specific member of the nuclear receptor superfamily. Cell 79, 971-980.
Shen, M.M., and Hodgkin, J. (1988). mab-3, a gene required for sex-specific yolk protein expression and a male-specific lineage in C. elegans. Cell 54, 1019-1031.
Shibayama, A., Cook, E.H., Jr., Feng, J., Glanzmann, C., Yan, J., Craddock, N., Jones, I.R., Goldman, D., Heston, L.L., and Sommer, S.S. (2004). MECP2 structural and 3'-UTR variants in schizophrenia, autism and other psychiatric diseases: a possible association with autism. Am J Med Genet B Neuropsychiatr Genet 128B, 50-53.
Sulston, J.E., and Horvitz, H.R. (1977). Postembryonic cell lineages of the nematode Caenorhabditis elegans. Dev Biol 56, 110-156.
Sulston, J.E., Schierenberg, E., White, J.G., and Thomson, J.N. (1983). The embryonic cell lineage of the nematode Caenorhabditis elegans. Dev Biol 100, 64-119.
Swaab, D.F., Chung, W.C., Kruijver, F.P., Hofman, M.A., and Hestiantoro, A. (2003). Sex differences in the hypothalamus in the different stages of human life. Neurobiol Aging 24 Suppl 1, S1-16; discussion S17-19.
Swaab, D.F., and Hofman, M.A. (1988). Sexual differentiation of the human hypothalamus: ontogeny of the sexually dimorphic nucleus of the preoptic area. Brain Res Dev Brain Res 44, 314-318.
Thorne, N., Chromey, C., Bray, S., and Amrein, H. (2004). Taste perception and coding in Drosophila. Curr Biol 14, 1065-1079.
Trent, C., Tsuing, N., and Horvitz, H. (1983). Egg-laying defective mutants of the nematode Caenorhabditis elegans. Genetics 104, 619-647.
Troemel, E.R., Chou, J.H., Dwyer, N.D., Colbert, H.A., and Bargmann, C.I. (1995). Divergent seven transmembrane receptors are candidate chemosensory receptors in C. elegans. Cell 83, 207-218.
Tsai, H.W., Grant, P.A., and Rissman, E.F. (2009). Sex differences in histone

100
modifications in the neonatal mouse brain. Epigenetics 4.
Tsunozaki, M., Chalasani, S.H., and Bargmann, C.I. (2008). A behavioral switch: cGMP and PKC signaling in olfactory neurons reverses odor preference in C. elegans. Neuron 59, 959-971.
Vellai, T., McCulloch, D., Gems, D., and Kovacs, A. (2006). Effects of sex and insulin/insulin-like growth factor-1 signaling on performance in an associative learning paradigm in Caenorhabditis elegans. Genetics S 174, 309-316.
Villella, A., and Hall, J. (1996). Courtship anomalies caused by doublesex mutations in Drosophila melanogaster. Genetics 143, 331-344.
Villella, A., and Hall, J.C. (2008). Neurogenetics of courtship and mating in Drosophila. Adv Genet 62, 67-184.
Von Stetina, S., Treinin, M., and Miller, D.r. (2006). The motor circuit. Int Rev Neurobiol S 69, 125-167.
Vrontou, E., Nilsen, S., Demir, E., Kravitz, E., and Dickson, B. (2006). fruitless regulates aggression and dominance in Drosophila. Nat Neurosci 9, 1469-1471.
Wakabayashi, T., Kitagawa, I., and Shingai, R. (2004). Neurons regulating the duration of forward locomotion in Caenorhabditis elegans. Neurosci Res 50, 103-111.
Wang, Z., Singhvi, A., Kong, P., and Scott, K. (2004). Taste representations in the Drosophila brain. Cell 117, 981-991.
Weaving, L.S., Christodoulou, J., Williamson, S.L., Friend, K.L., McKenzie, O.L., Archer, H., Evans, J., Clarke, A., Pelka, G.J., Tam, P.P., et al. (2004). Mutations of CDKL5 cause a severe neurodevelopmental disorder with infantile spasms and mental retardation. Am J Hum Genet 75, 1079-1093.
Weissburg, M.J. (2001). Sex, sensitivity, and second messengers: differential effect of cyclic nucleotide mediated inhibition in the chemosensory system of fiddler crabs. J Comp Physiol [A] 187, 489-498.
Weissburg, M.J., and Derby, C.D. (1995). Regulation of sex-specific feeding behavior in fiddler crabs: physiological properties of chemoreceptor neurons in claws and legs of males and females. J Comp Physiol [A] 176, 513-526.
Weissburg, M.J., Derby, C.D., Johnson, O., McAlvin, B., and Moffett, J.M., Jr. (2001). Transsexual limb transplants in fiddler crabs and expression of novel sensory capabilities. J Comp Neurol 440, 311-320.

101
White, J., Nicholas, T., Gritton, J., Truong, L., Davidson, E., and Jorgensen, E. (2007). The sensory circuitry for sexual attraction in C. elegans males. Curr Biol 17, 1847-1857.
White, J.G. (1988). The anatomy. In The nematode Caenorhabditis elegans, W.B. Wood, ed. (Cold Spring Harbor Laboratory Press).
White, J.G., Southgate, E., Thomson, J.N., and Brenner, S. (1986). The structure of the nervous system of the nematode Caenorhabditis elegans. Phil Trans R Soc Lond B 314, 1-340.
Wizemann, T.M., and Pardue, M.L. (2001). Exploring the biological contributions to human health: Does sex matter?, (Washington, D.C: National Academy Press).
Yi, W., Ross, J.M., and Zarkower, D. (2000). mab-3 is a direct tra-1 target gene regulating diverse aspects of C. elegans male sexual development and behavior. Development 127, 4469-4480.
Zarkower, D. (2001). Establishing sexual dimorphism: conservation amidst diversity? Nat Rev Genet 2, 175-185.
Zarkower, D. (2006). Somatic sex determination. WormBook, 1-12.
Zarkower, D., and Hodgkin, J. (1992). Molecular analysis of the C. elegans sex-determining gene tra-1: a gene encoding two zinc finger proteins. Cell 70, 237-249.
Zarkower, D., and Hodgkin, J. (1993). Zinc fingers in sex determination: only one of the two C. elegans Tra-1 proteins binds DNA in vitro. Nucleic Acids Res 21, 3691-3698.
Zheng, Y., Brockie, P., Mellem, J., Madsen, D., and Maricq, A. (1999). Neuronal control of locomotion in C. elegans is modified by a dominant mutation in the GLR-1 ionotropic glutamate receptor. Neuron 24, 347-361.
Ziabreva, I., Poeggel, G., Schnabel, R., and Braun, K. (2003). Separation-induced receptor changes in the hippocampus and amygdala of Octodon degus: influence of maternal vocalizations. J Neurosci 23, 5329-5336.
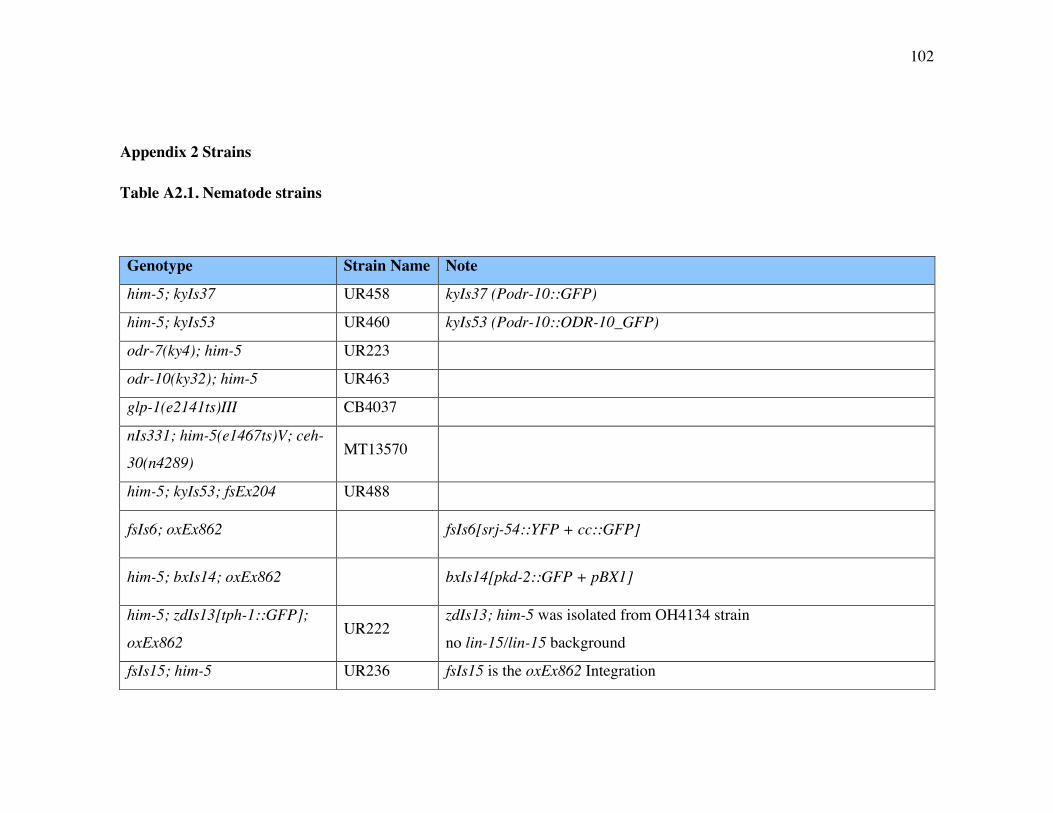
102
Appendix 2 Strains
Table A2.1. Nematode strains
Genotype Strain Name Note
him-5; kyIs37 UR458 kyIs37 (Podr-10::GFP)
him-5; kyIs53 UR460 kyIs53 (Podr-10::ODR-10_GFP)
odr-7(ky4); him-5 UR223
odr-10(ky32); him-5 UR463
glp-1(e2141ts)III CB4037
nIs331; him-5(e1467ts)V; ceh-
30(n4289) MT13570
him-5; kyIs53; fsEx204 UR488
fsIs6; oxEx862 fsIs6[srj-54::YFP + cc::GFP]
him-5; bxIs14; oxEx862 bxIs14[pkd-2::GFP + pBX1]
him-5; zdIs13[tph-1::GFP];
oxEx862 UR222
zdIs13; him-5 was isolated from OH4134 strain
no lin-15/lin-15 background
fsIs15; him-5 UR236 fsIs15 is the oxEx862 Integration
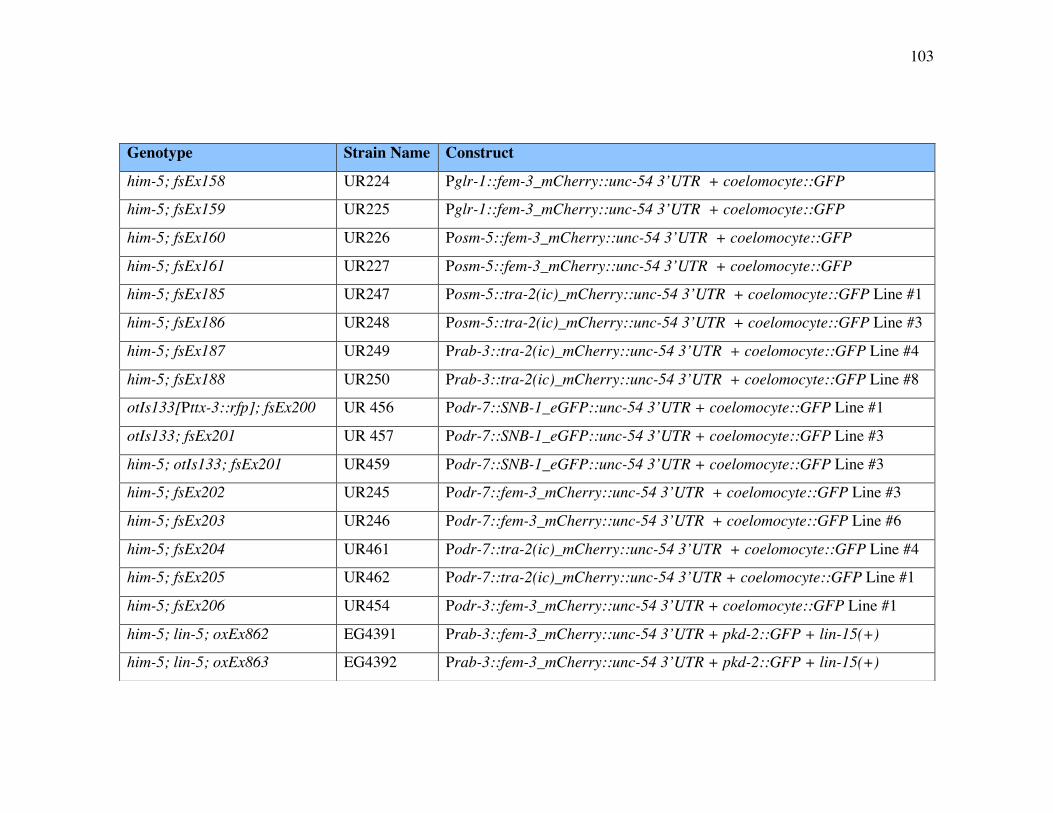
103
Genotype Strain Name Construct
him-5; fsEx158 UR224 Pglr-1::fem-3_mCherry::unc-54 3’UTR + coelomocyte::GFP
him-5; fsEx159 UR225 Pglr-1::fem-3_mCherry::unc-54 3’UTR + coelomocyte::GFP
him-5; fsEx160 UR226 Posm-5::fem-3_mCherry::unc-54 3’UTR + coelomocyte::GFP
him-5; fsEx161 UR227 Posm-5::fem-3_mCherry::unc-54 3’UTR + coelomocyte::GFP
him-5; fsEx185 UR247 Posm-5::tra-2(ic)_mCherry::unc-54 3’UTR + coelomocyte::GFP Line #1
him-5; fsEx186 UR248 Posm-5::tra-2(ic)_mCherry::unc-54 3’UTR + coelomocyte::GFP Line #3
him-5; fsEx187 UR249 Prab-3::tra-2(ic)_mCherry::unc-54 3’UTR + coelomocyte::GFP Line #4
him-5; fsEx188 UR250 Prab-3::tra-2(ic)_mCherry::unc-54 3’UTR + coelomocyte::GFP Line #8
otIs133[Pttx-3::rfp]; fsEx200 UR 456 Podr-7::SNB-1_eGFP::unc-54 3’UTR + coelomocyte::GFP Line #1
otIs133; fsEx201 UR 457 Podr-7::SNB-1_eGFP::unc-54 3’UTR + coelomocyte::GFP Line #3
him-5; otIs133; fsEx201 UR459 Podr-7::SNB-1_eGFP::unc-54 3’UTR + coelomocyte::GFP Line #3
him-5; fsEx202 UR245 Podr-7::fem-3_mCherry::unc-54 3’UTR + coelomocyte::GFP Line #3
him-5; fsEx203 UR246 Podr-7::fem-3_mCherry::unc-54 3’UTR + coelomocyte::GFP Line #6
him-5; fsEx204 UR461 Podr-7::tra-2(ic)_mCherry::unc-54 3’UTR + coelomocyte::GFP Line #4
him-5; fsEx205 UR462 Podr-7::tra-2(ic)_mCherry::unc-54 3’UTR + coelomocyte::GFP Line #1
him-5; fsEx206 UR454 Podr-3::fem-3_mCherry::unc-54 3’UTR + coelomocyte::GFP Line #1
him-5; lin-5; oxEx862 EG4391 Prab-3::fem-3_mCherry::unc-54 3’UTR + pkd-2::GFP + lin-15(+)
him-5; lin-5; oxEx863 EG4392 Prab-3::fem-3_mCherry::unc-54 3’UTR + pkd-2::GFP + lin-15(+)

104





























