
Plant Cell, Tissue and Organ Culture 28: 261-266, 1992. © 1992 Kluwer Academic Publishers. Printed in the Netherlands.
Cryopreservation of in vitro-grown shoot tips of apple and pear by vitrification
T. Niino 1, A. Sakai 2, H. Yakuwa ~, & K. Nojiri ~ 1Laboratory of Plant and Tissue Preservation, Department of Genetic Resources II, National Institute of Agrobiological Resources. 6000-1, Tohkamachi, Shinjo, Yamagata, 996 Japan; 2Asabucho 1-5-23, Kitaku, Sapporo, 001 Japan
Received 24 April 1991; accepted in revised form 17 October 1991
Key words: apple, cryopreservation, fruit trees, Malus, pear, Pyrus, shoot tips, vitrification
Abstract
In vitro-grown shoot tips of apples (Malus domestica Borkh. cv. Fuji) were successfully cryopreserved by vitrification. Three-week-old in vitro apple plantlets were cold-hardened at 5°C for 3 weeks. Excised shoot tips from hardened plantlets were precultured on a solidified Murashige & Skoog agar medium (MS) supplemented with 0.7M sucrose for 1 day at 5°C. Following preculture shoot tips were transferred to a 2 ml plastic cryotube and a highly concentrated cryoprotective solution (designated PVS2) was then added at 25°C. The PVS2 contains (W/V) 30% glycerol, 15% ethylene glycol and 15% dimethylsulfoxide in medium containing 0.4 M sucrose. After dehydration at 25°C for 80 min, the shoot tips were directly plunged into liquid nitrogen. After rapid warming, the shoot tips were expelled into 2 ml of MS medium containing 1.2 M sucrose and then plated on agar MS medium. Direct shoot elongation was observed in approximately 3 weeks. The average rate of shoot formation was about 80%. This vitrification method was successfully applied to five apple species or cultivars and eight pear cultivars. This method appears to be a promising technique for cryopreserving shoot tips from in vitro-grown plantlets of fruit trees.
Abbreviations: D M S O - dimethylsulfoxide, E G - ethylene glycol, PVS2- vitrification solution, L N - liquid nitrogen, B A - 6-benzylaminopurine, N A A - a-naphthaleneacetic acid, S E - standard error, ABA - abscisic acid
Introduction
Cryopreservation may be a useful method for long-term storage of germplasm using a mini- mum of space and maintenance. The develop- ment of techniques to successfully store apical meristems in liquid nitrogen (LN) is needed for conservation of genetic resources of fruit trees.
Winter vegetative buds from field grown fruit trees were stored in LN and successfully grafted onto rootstocks (Sakai & Nishiyama 1978; Tyler & Stushnoff 1988). However, in vitro cultured
plantlets are a preferable source of material for germplasm preservation. Plantlets are cultured under disease-free conditions, they are mass propagated and growth can be easily controlled. Until now, few papers reported cryopreservation of apical meristems from proliferating cultures of apple. Apical meristems from stock cultures of apple (cv. Jonathan) were first tested for survival in LN (Kuo & Lineberger 1985). In their study, apical meristems were dissected from cold-hard- ened plantlets and subsequently precultured for 48 h on medium supplemented with 5 or 10%

262
DMSO. These meristems were frozen slowly to -40°C in the presence of cryoprotectants prior to a plunge in LN. Regrowth following freezing to -196°C was in the form of callus, indicating meristem injury such that organized growth of the shoot meristems was precluded (Kuo & Lineberger 1985).
A simple vitrification procedure for cryo- preservation of cultured plant cells was pre- sented by Sakai & Kobayashi (1990) and Sakai et al. (1990, 1991). Nucellar cells of navel orange were treated at 25°C for 3 to 5 min with a highly concentrated cryoprotective solution (designated PVS2), followed by a direct plunge in LN. This method was successfully applied to apical meris- tems from in vitro plantlets of white clover. Excised shoot tips were precultured on medium supplemented with 1.2M sorbitol at 4°C for 2 days. However, this method has not produced a high level of survival in apple shoot tips. Recent- ly, we found that cold-hardening significantly improved the recovery rates of shoot tips cooled to -196°C by the simple vitrification procedure. We report a method for successful cryopreserva- tion of shoot tips from tissue-cultured plantlets of apple and pear.
adjusted to pH 5.6 prior to autoclaving at 121°C for 15 min. The stock cultures were sub- cultured every 40 days. The stock cultures of pear plants were grown on Murashige & Skoog (MS) medium containing 2.5 mg1-1 BA, 2.5% sucrose and 0.8% agar (Wako Chemical Co, Japan) at pH 5.6. These tissue-cultured plantlets of apple and pear were all grown on 25ml medium in 100 ml glass flasks (6 cm long, 9 cm base diameter) under white fluorescent light (52 p, mol s -I m-2), 16h photoperiod at 25°C.
Cold-hardening and precultures
Three week old plantlets were cold-hardened at 5°C for 3 weeks under 8 h photoperiods. Shoot tips were dissected cold-hardened and non-hard- ened ptantlets. Shoot tips (1.5-2mm long, 1.5 mm base diameter) consisted of the apical meristem and 4 to 5 leaf primordia (Fig. 1). Cold-hardened and non-hardened shoot tips were precultured at 5°C for 1-2 days on MS medium supplemented with 0.7 M sucrose under 8 h photoperiod.
Materials and methods
Materials
Tissue-cultured apple plants Malus domestica Borkh. cv. Fuji were mainly used in the present study. Stock cultures of five other apple cultivars Golden Delicious, and hybrids 423-1 (Fu- ji x Mohe-7); the apple rootstocks M. 9 and M. 26 (Malus paradisiaca Shneid.) and Malus prunifolia Borkh. were also used. Eight tissue- cultured pear cultivars were also tested for sur- vival following immersion in LN by vitrification: Pyrus pyrifolia (Burro.) Nakai Hokkaiwase, Yoshino and Senryo; Pyrus communis L. Beurr6 d'Amanlis, Beurr6 Jean Van Geert, Doyenn6 du Cornice, Early Seckel and Fondante Thirriot.
Stock cultures of apple plants were maintained on Murashige & Skoog (MS) medium containing 1.0mg1-1 BA, 2.5% sucrose and 0.7% Bacto- agar (Difco, Detroit, USA). The medium was
L
~--1.0 -1.5 " ~ m m
Fig. 1. Longitudinal section of an excised shoot tip. L, young leaf; LP, leaf primordia; M, apical meristem. (A)
upper and (B) lower ends of an excised shoot tip.
Vitrification procedure
Following preculturing, ten shoot tips were transferred to 1.0 ml of PVS2 in a 2 ml plastic cryotube (Wheaton, Scientific, Millville, USA) at 25°C. The vitrification solution PVS2 (Sakai & Kobayashi 1990; Sakai et al. 1990, 1991), con- tains 30% (W/V) glycerol, 15% (W/V) EG and 15% (W/V) DMSO in MS basal medium con- taining 0.4 M sucrose (pH 5.8). After a 20 min- treatment, the PVS2 was removed using a Pas- teur pipette, and replaced twice with fresh PVS2 and held at 25°C for different lengths of time. The cryotubes in which shoot tips were finally suspended in 0.6 ml of fresh PVS2 immediately before being directly plunged and held in LN for one day. The cooling rate was about 280°C min-~. Shoot tips from LN were rapidly warmed in a water bath at 25°C (warming rate: about 280°C min- ~ ).
Viability and regrowth
After rapid warming, PVS2 was drained from the cryotubes and replaced twice with liquid MS medium containing 1.2 M sucrose and held for 30min. After blotting the surface solution, the shoot tips were transferred onto 0.8% agar MS medium excluding NH4NO 3 (Kuriyama et al. 1990) and cultured under standard conditions described previously.
Recovering shoot tips were observed for 20 and 40 days. Shoot formation was recorded as percent of total number of shoot tips forming shoots 40 days after plating. Twenty shoot tips were tested for each of two replicates for each experiment.
R e s u l t s
Cold-hardening significantly improved the re- covery rates of shoot tips cooled to -196°C by vitrification. At one day following hardening, the recovery rate of precultured shoot tips was 80% (Table 1). However, in non-hardened shoot tips, the recovery rate was only 25% after one day of preculturing.
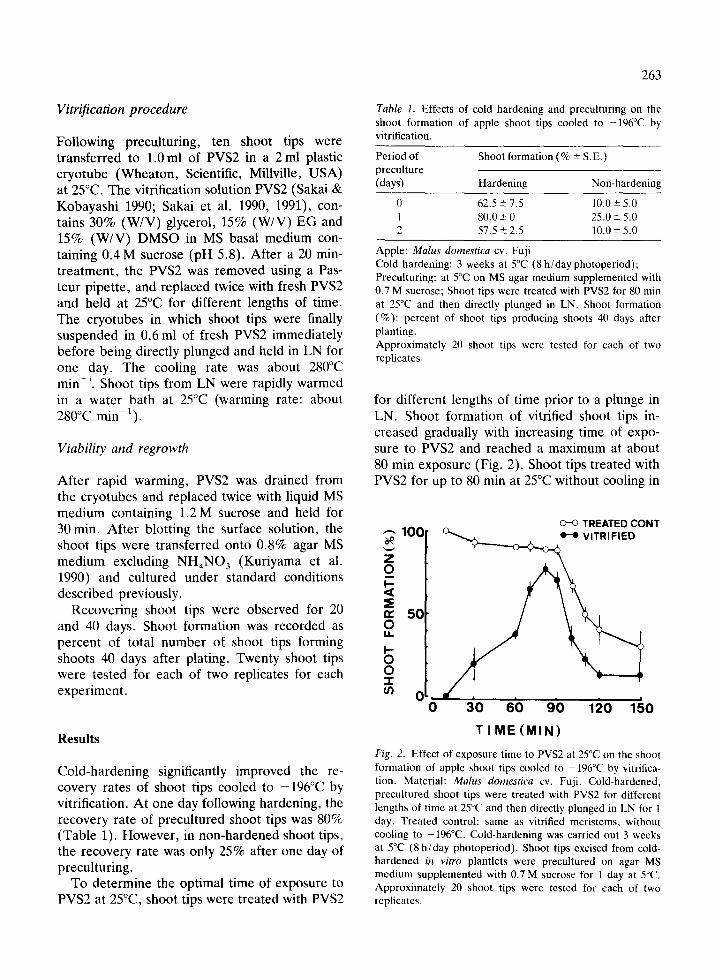
To determine the optimal time of exposure to PVS2 at 25°C, shoot tips were treated with PVS2
263
Table 1. Effects of cold hardening and preculturing on the shoot formation of apple shoot tips cooled to -196°C by vitrification.
Period of preculture (days)
Shoot formation (% +- S.E.)
Hardening Non-hardening
0 62.5±7.5 10.0±5.0 1 80 .0±0 25.0±5.0 2 57.5±2.5 10.0±5.0
Apple: Malus domestica cv. Fuji Cold hardening: 3 weeks at 5°C (8 h/dayphotoperiod); Preculturing: at 5°C on MS agar medium supplemented with 0.7 M sucrose; Shoot tips were treated with PVS2 for 80 min at 25°C and then directly plunged in LN. Shoot formation (%): percent of shoot tips producing shoots 40 days after planting. Approximately 20 shoot tips were tested for each of two replicates.
for different lengths of time prior to a plunge in LN. Shoot formation of vitrified shoot tips in- creased gradually with increasing time of expo- sure to PVS2 and reached a maximum at about 80 min exposure (Fig. 2). Shoot tips treated with PVS2 for up to 80 min at 25°C without cooling in
TREATED CONT 100
z o_ I-.- ,¢[ =E n, 50
I-.
8 ~ 0 ,
0 30 60 9 0 120 150
T I M E ( M I N )
Fig. 2. Effect of exposure time to PVS2 at 25°C on the shoot formation of apple shoot tips cooled to -196°C by vitrifica- tion. Material: Malus domestica cv. Fuji. Cold-hardened, precultured shoot tips were treated with PVS2 for different lengths of time at 25°C and then directly plunged in LN for 1 day. Treated control: same as vitrified meristems, without cooling to -196°C. Cold-hardening was carried out 3 weeks at 5°C (8 h/day photoperiod). Shoot tips excised from cold- hardened in vitro plantlets were precultured on agar MS medium supplemented with 0.7 M sucrose for 1 day at 5°C. Approximately 20 shoot tips were tested for each of two replicates.

264
LN ( t reated control) retained high levels of shoot format ion (about 90%). However , longer exposures decreased shoot formation to nearly the same level as vitrified shoot tips (Fig. 2).
Shoot formation by the same vitrification pro- cedure was compared to five other apple cul- tivars or species. High levels of shoot formation were obtained in the apples tested (Table 2).
High levels of shoot formation were also ob- tained for eight pear cultivars cooled to -196°C by the same vitrification procedure (Table 3).
Successfully vitrified apple shoot tips resumed
Table 2. Shoot formation of apple shoot tips cooled to -196°C by vitrification.
Cultivar or species Shoot formation (% ± S.E.)
cv. Golden Delicious (Malus domestica) cv. a hybrid 423-1 (Malus domestica) M. 9 ( Malus paradisiaca) M. 26 ( Malus paradisiaca) Malus prunifolia
55.0±5.0 77.5±7.5 45.0±5.0 67.5±2.5 70.0±5.0
Cold hardening: 3 weeks at 5°C (8 h/day photoperiod); Preculturing: at 5°C for 1 day on MS agar medium supple- mented with 0.7 M sucrose. Hardened, precultured shoot tips were treated with PVS2 for 80 min at 25°C prior to a plunge in LN. Shoot formation (%): percent of shoot tips producing shoots 40 days after planting. Approximately 20 shoot tips were tested for each of two replicates.
Table 3. Shoot formation of pear shoot tips cooled to -196°C by vitrification.
Pyrus cultivars Shoot formation (% - S.E.)
Pyrus pyrifolia c.v. Hokkaiwase 70.0 ± 0 c.v. Yoshino 40.0--- 0 c.v. Senryo 60.0- 0
Pyrus communis cv. Beurr6 d'Amanlis 72.5 ± 2.5 cv. Beurr6 Jean Van Geert 67.0 ± 3.0 cv. Doyenn6 du Comice 50.0 -+ 15.0 cv. Early Seckel 61.1 -+ 11.1 cv. Fondante Thirriot 57.5 -+ 7.5
Cold hardening: 3 weeks at 5°C (8 h/day photoperiod); Preculturing: at 5°C for 1 day on MS agar medium supple- mented with 0.7 M sucrose. Hardened, precultured shoot tips were treated with PVS2 for 90 min at 25°C prior to a plunge in LN. Shoot formation (%): percent of shoot tips producing shoots 40 days after planting. Approximately 20 shoot tips were tested for each of the two replicates.
Fig. 3. Plantlets developed from apple shoot tips cooled to -196°C by vitrification, at 30 days after reculture. Material: Malus domestica cv. Fuji.
growth in about 3 days after reculture, and star- ted to develop shoots within two or three weeks without intermediary callus formation. Figure 3 shows shoots formed from the vitrified apical shoots tips after one month of culture. Almost all shoot formed roots on modified MS medium (half-strength inorganic salt macronutrients) con- taining 1 mg NAA1-1, and were successfully t ransferred to pots. No morphological abnor- malities were observed in the plants developed f rom cryopreserved shoot tips.
Discussion
Cold-hardening, preculturing and post-thaw handling of apical meristems from in vitro plant- lets of fruit trees are essential to successful cryopreservation. Reed (1990) clearly demon- strated that cold-hardening significantly im- proved the recovery rates of apical meristems of all four pear cultivars tested. A high level of sugar (Dereuddre et al. 1988) or sorbitol (Yamada et al. unpublished) during preculture was repor ted to he important in improving sur- vival of cryopreserved meristems. In the present study, preculturing at 5°C for 1 day following cold-hardening improved the recovery rates of hardened shoot tips cooled to -196°C by vitrifi- cation. However , alterations associated with enhanced shoot tip survival during precultur- ing is not well characterized. Anandarajah &

265
McKersie (1990) reported high sucrose concen- trations in the maturation medium of somatic embryo of alfalfa to substitute for exogenous ABA induced desiccation tolerance of somatic embryos.
Successful vitrification requires careful control of the highly concentrated vitrification solution to prevent injury by chemical toxicity or excess osmotic stresses during dehydration. To establish this objective, a stepwise equilibration procedure at about 20°C and 5 or 0°C has been used (Uragami et al. 1989; Towill 1990). The one step method for dehydration presented by Sakai & Kobayashi (1990) and Sakai et al. (1990, 1991) simplified the procedure and enhanced survival. This simple vitrification method was successfully applied to the cryopreservation of meristems of white clover (Yamada et al. unpublished). Pre- cultured meristems (0.5 to 1.0 mm in length) of white clover which were treated with PVS2 at 25°C for 5 min or at 0°C for 15 min prior to a plunge in LN produced high levels of plant re- generation (about 80%). However, incubation time in PVS2 appears to be species-specific. The protocol for white clover (5 min at 25°C or 15 min at 0°C) was not applicable to apple and pear. In the present study, optimal exposure time to PVS2 for apple and pear was 80-90 m i n - considerably longer than that required for white clover (Yamada et al. unpublished) and carnation (Langis et al. 1990) meristems. Apple and pear meristems used in this study were two to three times the length of white clover. Con- ceivably, incubation time in PVS2 and thus meristem dehydration time may necessarily in- crease with size of the excised shoot tips. Further investigations of the relationship between ex- cised shoot tip size and optimal PVS2 exposure time are required.
It is particularly important that cryopreserved cells and meristems be capable of producing plants identical to the non-treated phenotype. Haskins & Kartha (1980) clearly demonstrated renewed growth in frozen-thawed pea meristems to be frozen tissue other than the original meris- tematic 'dome' and that renewed growth led to differentiation and whole plant regeneration. Formation of a callus phase prior to shoot initia- tion is undesirable since callus potentially in- creases the frequency of variants. The pattern of
growth of cryopreserved meristems varies con- siderably with cryogenic protocols (Reed 1990) and with regrowth medium (Withers et al. 1988). Frozen meristems of cold hardened Pyrus pro- duced a high portion of normal (non-callused) shoots after thawing (Reed 1990). Cryopreser- ved clover meristems following preculture by vitrification resumed growth three days after plating and developed shoots within 10 days without intermediary callus formation (Yamada et al. unpublished). In the present study, we also observed the same recovery process in the cold hardened shoot tips of apple and pear cooled to -196°C by vitrification.
Shoot formation of apple shoot tips treated with PVS2 for 90 min. without cooling in LN (treated control) was similar to vitrified apple meristems (Fig. 2). Thus, vitrification under adequately controlled conditions did not cause additional loss beyond that produced by the vitrification solution. Complete vitrification eliminates concern for potentially damaging ef- fects of intra- and extracellular crystallization producing high levels of cell and meristem sur- vival. Vitrified clover meristems in LN were rapidly transferred into a bath held at -70°C before being warmed rapidly in water at 25°C. A 1 min exposure to -70°C caused serious injury, but a slight decrease in survival was obtained in the meristems held for 15 s at -70°C (Yamada et al. unpublished). These results indicate that in- tracellular crystallization occurs in the vitrified cells during slow warming and causes cell death (Sakai et al. 1991).
Acknowledgements
The authors wish to thank Dr. A. Ishihara, Iwate University for supplying apple stock cul- tures, Dr. Karen Tanino, Department of Hor- ticulture, University of Saskatchewan for review- ing the manuscript, and Mrs. Y. Takeda for her technical assistance.
References
Anandarajah K & McKersie BD (1990) Manipulating the desiccation tolerance and vigor of dry somatic embryos of

266
Medicago sativa L. with sucrose, heat shock and abscisic acid. Plant Cell Rep. 9:451-455
Dereuddre J, Fabre J & Bassaglia C (1988) Resistence to freezing in liquid nitrogen of carnation (Dianthus caryophyllus L. var Kolo) apical and axially shoot tips excised from different aged in vitro plantlets. Plant Cell Rep. 7:170-173
Haskins RH & Kartha KK (1980) Freeze preservation of pea meristems: Cell survival. Can. J. Bot. 58:833-840
Kuo CC & Lineberger BD (1985) Survival of in vitro cul- tured tissue of Jonathan apples exposed to -196°C. HortScience 20:764-767
Kuriyama A, Watanabe K, Ueno S & Mituda M (1990) Inhibitory effect of ammonium ion on recovery of cryop- reserved rice cells. Plant Sci 64:231-235
Langis R, Sohnabel B, Earle ED & Steponkus PL (1989) Cryopreservation of Brassica compestris L, cell suspensions by vitrification. Cryo-Letters 10:421-428
Langis R, Sohnabel-preikstas B J, Earle ED & Steponkus PL (1990) Cryopreservation of carnation shoot tips by vitrifi- cation. Cryobiology 27:657-658
Reed B (1990) Survival of in vitro-grown apical meristems of Pyrus following cryopreservation. HortScience 25:111-113
Sakai A & Nishiyama Y (1978) Cryopreservation of winter vegetative buds of hardy fruit trees in liquid nitrogen. HortScience. 13:225-227
Sakai A & Kobayashi S (1990) A simple and efficient procedure for cryopreservation of nucellar cells of navel orange by vitrification. Cryobiology 27:657
Sakai A, Kobayashi S & Oiyama I (1990) Cryopreservation of nucellar cells of navel orange (Citrus sinensis var. brasiliensis Tanaka) by vitrification. Plant Cell Rep. 9: 30-33
Sakai A, Kobayashi S & Oiyama I (1991) Survival by vitrification of nucellar cells of navel orange (Citrus sinen- s/~ var. brasiliensis Tanaka) cooled to -196°C. J. Plant Physiol. 137:465-470
Tyler NJ & Stushnoff C (1988) The effects of prefreezing and controlled dehydration on cryopreservation of dormant vegetative apple buds. Can. J. Plant Soil. 68:1163-1167
Towill LE (1990) Cryopreservation of isolated mint shoot tips by vitrification. Plant Cell Rep. 9:178-180
Uragami A, Sakai A, Nagai M & Takahashi TA (1989) Survival of cultured cells and somatic embryos of As- paragus officinalis L. cryopreserved by vitrification. Plant Cell Rep. 8:418-421
Withers LA, Benson EE & Martin M (1988) Cooling rate/ culture medium interactions in the survival and structure stability of cryopreserved shoot-tips of Brassica napus. Cryo-Letters 9:114-119
Recommended

















