
Vol. 30, No. 10JOURNAL OF CLINICAL MICROBIOLOGY, OCt. 1992, p. 2748-27520095-1137/92/102748-05$02.00/0
Colony Morphotype on Sabouraud-Triphenyltetrazolium Agar:a Simple and Inexpensive Method for Candida
Subspecies DiscriminationGUILLERMO QUINDOS,* MANUEL FERNANDEZ-RODRIGUEZ, ANGEL BURGOS,
MATILDE TELLAETXE, RAMON CISTERNA, AND JOSE PONTON
Departamento de Microbiologia e Inmunologia, Facultad de Medicina y Odontologia,Universidad del Pais Vasco, Bilbao, SpainReceived 27 January 1992/Accepted 27 June 1992
A new method of Candida subspecies discrimination on Sabouraud-triphenyltetrazolium agar is reported.Eive hundred sixty-two strains of Candida and Torulopsis glabrata, previously identified by conventionalmycological methods, were studied. Each strain received a three-letter code and a number based on its colonialmorphology. Sixteen morphotypes were found for Candida albicans, 6 were found for Candida parapsilosis, 4were found for both Candida guilliermondii and Candida krusei, and 12 were found for Candida tropicalis. Noneof the 56 T. glabrata strains studied grew on this agar. A reproducibility of 95% was found for C. albicans. Thesimplicity and low cost could make this method useful for typing Candida spp.
Candida albicans and other Candida spp. are opportunis-tic pathogens involved in an increasing number of infectionsof patients with immunodeficiency. Different typing meth-ods, including biotyping (11), enzyme typing (15), morpho-typing (13), serotyping (1), resistotyping (8), killer typing(14), protein typing (2, 7), and karyotyping (10), have beendescribed previously, and attempts have been made todetect a possible correlation of some C. albicans types withthe virulence of the isolates (5, 9). However, it is still notpossible to type C. albicans with a degree of certainty, andthe lack of a suitable typing scheme has undoubtedly hin-dered studies of the epidemiology and virulence of thispathogenic yeast (3). These typing methods have usually notbeen applied to other Candida spp. (9), and most of theminvolve a relatively complicated methodology. In this article,we report the development of a new and inexpensive typingmethod and its application to evaluation of the morphotypespresent in 562 strains of Candida spp. from different clinicalspecimens.Data were obtained from 562 unrelated strains of C.
albicans, Torpulopsis glabrata, and other medically impor-tant Candida spp., previously identified by conventionalmycological methods, from a variety of clinical samples.Most of them were from skin and mucosae, with 384 isolatesfrom oral, vaginal, and respiratory samples and 178 fromblood, peritoneal fluid, or organ biopsies. C. albicans 10261and 3153 and their mutants IR2h (proteinase deficient) andCA2 (germ tube deficient), respectively, sent by A. Cassone(Istituto Superiore di Sanita, Rome, Italy), were also evalu-ated. Fungal identification was based on morphology onSabouraud dextrose agar (Difco, Detroit, Mich.) and cornmeal extract-Tween 80 agar (Oxoid, Basingstoke, UnitedKingdom), germ tube formation, as well as biochemicalcharacterization with the use of the API 20C Aux (APISystem S.A., Montalieu Vercieux, France) and the 16-disccarbon auxanogram (Diagnostics Pasteur, Paris, France)systems.The Sabouraud-triphenyltetrazolium (STTZ) agar, con-
* Corresponding author.
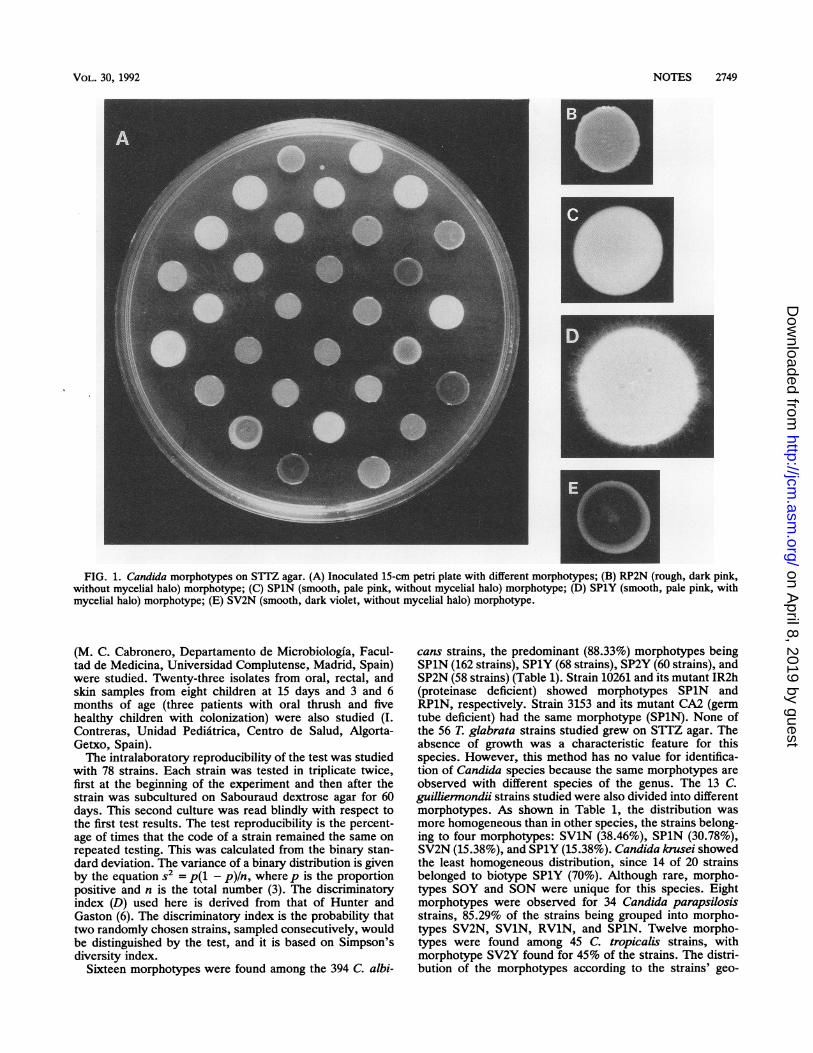
taining 2,3,5-triphenyltetrazolium chloride (Sigma ChemicalCo., St. Louis, Mo.) (0.1 g), peptone (10 g), glucose (20 g),agar (20 g), and distilled water (1 liter), was distributed in 40-to 50-ml quantities into 15-cm petri dishes. Plates were eitherinoculated immediately or stored at 4°C for no longer than 8weeks. Each isolate was maintained by monthly transfers inSabouraud dextrose agar slants and storage at 4°C. Beforepreparation of inocula, each isolate was grown on a freshSabouraud dextrose agar plate at 37°C for 24 h. Three or fourcolonies were resuspended in 1 ml of distilled water andadjusted to a MacFarland standard of 5 to prepare eachinoculum. Before inoculation, all plates were incubatedinverted at 37°C for 1 h to dry agar surfaces. Plates wereinoculated in triplicate with 10 ,ul from the cell suspension byusing a micropipette and allowing the inoculum drop to dryon the agar surface. Cultures were incubated at 37°C for 6days. After incubation, a three-letter code and a numberwere given to each strain, according to its colonial morphol-ogy (Fig. 1): texture, S = smooth and R = rough; color, P =
pink (P1 = pale pink = Pantone 176C, P2 = dark pink =Pantone 1785C), V = violet (Vi = pale violet = Pantone251C, V2 = dark violet = Pantone 262C) (Pantone Inc.), 0 =
orange, W = White, etc.; and presence of mycelial halo(hyphae or pseudohyphae), Y = yes and N = no. The codeNG was used if no growth was observed. Pantone is aregistered trademark for color specification that is orientedto facilitation and color standardization of publishing andgraphic design work.We were not able to make a direct comparison between
the morphotype method with STTZ agar and referencemethods, which are not well established for Candida typing;therefore, we attempted to subdivide the strains within C.albicans species on the basis of their likelihood of diversity.We compared strains from various geographical and epide-miological situations with those from infected and uninfectedpersons. With this purpose, 277 additional C. albicansstrains from Barcelona (J. M. Torres Rodriguez, InstitutoMunicipal de Investigaciones Medicas-Hospital del Mar,Barcelona, Spain), London (G. C. White, Candida Unit,Division of Hospital Infection, Central Public Health Labo-ratory, Colindale, London, United Kingdom), and Madrid
2748
on April 8, 2019 by guest
http://jcm.asm
.org/D
ownloaded from
NOTES 2749
FIG. 1. Candida morphotypes on S1TZ agar. (A) Inoculated 15-cm petri plate with different morphotypes; (B) RP2N (rough, dark pink,without mycelial halo) morphotype; (C) SPlN (smooth, pale pink, without mycelial halo) morphotype; (D) SPlY (smooth, pale pink, withmycelial halo) morphotype; (E) SV2N (smooth, dark violet, without mycelial halo) morphotype.
(M. C. Cabronero, Departamento de Microbiologia, Facul-tad de Medicina, Universidad Complutense, Madrid, Spain)were studied. Twenty-three isolates from oral, rectal, andskin samples from eight children at 15 days and 3 and 6months of age (three patients with oral thrush and fivehealthy children with colonization) were also studied (I.Contreras, Unidad Pediatrica, Centro de Salud, Algorta-Getxo, Spain).The intralaboratory reproducibility of the test was studied
with 78 strains. Each strain was tested in triplicate twice,first at the beginning of the experiment and then after thestrain was subcultured on Sabouraud dextrose agar for 60days. This second culture was read blindly with respect tothe first test results. The test reproducibility is the percent-age of times that the code of a strain remained the same onrepeated testing. This was calculated from the binary stan-dard deviation. The variance of a binary distribution is givenby the equation s2 = p(l - p)In, where p is the proportionpositive and n is the total number (3). The discriminatoryindex (D) used here is derived from that of Hunter andGaston (6). The discriminatory index is the probability thattwo randomly chosen strains, sampled consecutively, wouldbe distinguished by the test, and it is based on Simpson'sdiversity index.
Sixteen morphotypes were found among the 394 C. albi-
cans strains, the predominant (88.33%) morphotypes beingSPiN (162 strains), SPlY (68 strains), SP2Y (60 strains), andSP2N (58 strains) (Table 1). Strain 10261 and its mutant IR2h(proteinase deficient) showed morphotypes SPiN andRP1N, respectively. Strain 3153 and its mutant CA2 (germtube deficient) had the same morphotype (SP1N). None ofthe 56 T. glabrata strains studied grew on STTZ agar. Theabsence of growth was a characteristic feature for thisspecies. However, this method has no value for identifica-tion of Candida species because the same morphotypes areobserved with different species of the genus. The 13 C.guilliermondii strains studied were also divided into differentmorphotypes. As shown in Table 1, the distribution wasmore homogeneous than in other species, the strains belong-ing to four morphotypes: SV1N (38.46%), SPiN (30.78%),SV2N (15.38%), and SPlY (15.38%). Candida krusei showedthe least homogeneous distribution, since 14 of 20 strainsbelonged to biotype SPlY (70%). Although rare, morpho-types SOY and SON were unique for this species. Eightmorphotypes were observed for 34 Candida parapsilosisstrains, 85.29% of the strains being grouped into morpho-types SV2N, SV1N, RV1N, and SP1N. Twelve morpho-types were found among 45 C. tropicalis strains, withmorphotype SV2Y found for 45% of the strains. The distri-bution of the morphotypes according to the strains' geo-
VOL. 30, 1992
on April 8, 2019 by guest
http://jcm.asm
.org/D
ownloaded from
J. CLIN. MICROBIOL.
TABLE 1. Morphotypes of 562 strains of Candida on STTZ agar
Species Morphotype' No. of(no. of isolates) Mrhteastrains (%)
C. albicans (394) SPlN 162 (41.12)SPlY 68 (17.26)SP2Y 60 (15.23)SP2N 58 (14.72)NG 11 (2.79)RP1N 6 (1.52)RP2Y 6 (1.52)RP1Y 6 (1.52)SV1Y 4 (1.03)RP2N 3 (0.76)SV2Y 3 (0.76)RV2Y 2 (0.51)RV2N 2 (0.51)RV1Y 1 (0.25)SV1N 1 (0.25)SV2N 1 (0.25)
T. glabrata (56) NG 56 (100)
C. guilliernondii (13) SV1N 5 (38.46)SPlN 4 (30.78)SV2N 2 (15.38)SPlY 2 (15.38)
C. krusei (20) SPlY 14 (70)SPlN 4 (20)SOY 1 (5)SON 1 (5)
C. parapsilosis (34) SV2N 11 (32.35)SV1N 8 (23.53)RV1N 5 (14.71)SPlN 5 (14.71)RP1N 2 (5.88)RV2N 1 (2.94)SPlY 1 (2.94)NG 1 (2.94)
C. tropicalis (45) SV2Y 18 (45)SV1Y 5 (12.5)RV2Y 5 (12.5)SV2N 4 (10)NG 4 (10)SP2Y 3 (7.5)SPlN 1 (2.5)SPlY 1 (2.5)RP1N 1 (2.5)RP2Y 1 (2.5)RV1Y 1 (2.5)RV1N 1 (2.5)
a The code used is explained in Materials and Methods.
graphical and clinical origins is shown in Tables 2 and 3.Although differences in morphotypes were observed, SPiNwas predominant among strains from the four geographicallocations and from most of the clinical sites. MorphotypeSV2N, which was very infrequent among our C. albicansstrains, was the second most frequent among strains fromMadrid.Morphotype SPiN was predominant in the pediatric iso-
lates (data not shown). However, in all patients, the samemorphotypes were present among isolates from differentlocations and at different times of sampling if the cultureswere positive. Patients with oral thrush had strains with theSPiN morphotype isolated from the mouth, anus, and skin
TABLE 2. Morphotypes of C. albicans isolates from differentgeographical origins
No. of strains (%) from source or geographical originMorphotypea
Our study Barcelona London Madrid
SPlN 162 (41.12) 29 (52.73) 38 (33.63) 61 (55.96)SPlY 68 (17.26) 7 (12.73) 12 (10.62) 1 (0.92)SP2Y 60 (15.23) 2 (3.64) 30 (26.55) 0 (0)SP2N 58 (14.72) 4 (7.27) 24 (21.24) 0 (0)NG 11 (2.79) 5 (9.09) 2 (1.77) 0 (0)RP1N 6 (1.52) 1 (1.82) 2 (1.77) 12 (11.01)RP2Y 6 (1.52) 1 (1.82) 2 (1.77) 0 (0)RP1Y 6 (1.52) 1 (1.82) 0 (0) 0 (0)SV1Y 4 (1.03) 1 (1.82) 0 (0) 3 (2.75)RP2N 3 (0.76) 0 (0) 1 (0.88) 0 (0)SV2Y 3 (0.76) 0 (0) 0 (0) 0 (0)RV2Y 2 (0.51) 3 (5.44) 2 (1.77) 0 (0)RV2N 2 (0.51) 0 (0) 0 (0) 0 (0)RV1Y 1 (0.25) 0 (0) 0 (0) 0 (0)SV1N 1 (0.25) 1 (1.82) 0 (0) 28 (25.69)SV2N 1 (0.25) 0 (0) 0 (0) 0 (0)RV1N 0 (0) 0 (0) 0 (0) 4 (3.67)
a The code used is explained in Materials and Methods.
of the napkin area. In some oral and rectal samples, twodifferent morphotypes were present, but this was not ob-served with skin samples.Ten SPlN-morphotype C. albicans isolates from eight
patients with denture stomatitis (two strains from one pa-tient, another two strains from another patient, and six fromsix different persons) were evaluated by Susan A. Howell
TABLE 3. Relationship of morphotypes of C. albicans isolates totheir clinical and geographical origins
Origin of clinical Morphotypea (no.) of isolates from:sample Madrid Barcelona London
Blood SPlN (4) SPlN (2) SPlN (12)SV1N (2) SP2N (1) SP2Y (10)RP1N (1) SP2N (9)SV1Y (1) RV2Y (2)
SPlY (2)Other (4)
Mouth and SPlN (7) SPlN (12) SPlN (18)digestive tract SV1N (6) SPlY (4) SP2Y (14)
RP1N (4) NG (4) SPlY (9)Other (4) SP2N (7)
Other (3)
Respiratory tract SPlN (7) SV2Y (3) SPlN (4)RV1N (3) SPlY (2) SP2N (3)SV1N (2) Other (3) SP2Y (3)SV1Y (2) SPlY (1)RP1N (1)
Nail and skin SPlN (13) SPlN (4) SP2N (3)SV1N (7) RP1N (1) SPlN (2)RP1N (2)Other (2) SP2Y (1)
Urogenital tract SPiN (30) SPlN (11) SPlN (2)SV1N (10) Other (4) SP2N (2)RP1N (4) SP2Y (2)SPlY (1)
a The code used is explained in Materials and Methods.
2750 NOTES
on April 8, 2019 by guest
http://jcm.asm
.org/D
ownloaded from

NOTES 2751
from the Department of Microbial Diseases, Lambeth Hos-pital (London, United Kingdom) by restriction fragmentlength polymorphism analysis. DNA was digested withrestriction endonucleases EcoRI, Hinfl and BglII (12). Eightdifferent karyotypes were found, and only isolates from thesame patient had the same karyotype (data not shown).Morphotype SPlN appeared to be that for a subspeciescomposed by genetically different strains.The typeabilities for the different species tested, if absence
of growth was not considered a morphotype, were 91.11%for C. tropicalis, 97.21% for C. albicans and C. parapsilosis,and 100% for C. guilliennondii and C. krusei. The reproduc-ibilities of the test after the strains were subcultured on
STTZ agar for 2 months were 85% for 6 C. guilliennondiistrains, 92% for 12 C. tropicalis strains, 95% for 49 C.albicans strains, and 100% for 4 C. krusei strains and 7 C.parapsilosis strains. According to Simpson's index of diver-sity, biotyping on STTZ agar had a D = 0.757 for C.albicans. The method showed discriminatory indices forCandida spp. different from that of C. albicans; 0.769 for C.guilliermnondii, 0.416 for C. krusei, 0.835 for C. parapsilosis,and 0.811 for C. tropicalis.
Sixty strains of six species of Candida were cultured indifferent conditions on S1TZ agar for the selection of theappropriate conditions of test performance. A 10-,ul inocu-lum from a cellular suspension adjusted to a MacFarlandstandard of 5 was chosen since lesser concentration suspen-
sions gave unsatisfactory results. Incubation of cultures for6 days at 37°C was selected versus incubation at 37°C for 1,2, 4, 5, 8, and 10 days or the same periods of incubations at
37°C plus 1 additional day at 25°C. The reason for this choicewas the consolidation of the colonial features used for thecode (texture, color, or presence of mycelial halo) on day 6of incubation in 98% of the strains. No improvement of theseresults was observed with other incubation conditions.When assessing the value of any typing method, the three
main characteristics that need to be considered are typeabil-ity, reproducibility, and discriminatory power. The cost,
ease of use, and rapidity should also be considered. Thetypeability of a method is the proportion of a population ofisolates that can be typed by that method. All existing typingmethods for C. albicans have 100% typeability (2). Ourmethod gave 97.21% typeability, since 11 of 394 C. albicansstrains typed did not grow in the medium. These strains maybe considered to be a different morphotype (NG). Themethod described here was not able to discriminate differentmorphotypes of T. glabrata, since none of the strains testedgrew in STTZ agar.
Reproducibility of the method for expression and stabilityof strain codes ranged from 85 to 100%, depending on thespecies tested, and was 95% for C. albicans. Overheating ofthe medium during its preparation and marked changes in theincubation temperature resulted in differences among mor-
photypes. As the evaluation of a morphotype is subjectiveand different appreciations of color intensity are unavoid-able, we recommend the use of Pantone color standards. Thesame results are thus obtained with different readers. At thesame time, variation in colony morphology of some strainsduring subculturing could be explained by high-frequencyswitching (16). Estimates of reproducibility of different phys-iochemical typing methods for C. albicans have shownvariations better and worse than those described here for thetyping on STTZ agar. McCreight et al. (8) found a 96% invitro reproducibility for resistotyping tests. However,Hunter and Fraser (4), using a modification of that system,
found reproducibilities ranging from 77 to 100%, depending
on the number of test differences required to distinguishamong strains. In contrast, a reproducibility lower than 95%has been found for morphotyping on malt extract agar anddifferent biotyping systems (3, 13). Other typing methods,based on assimilation of carbon sources, detection of extra-cellular enzymes, or biochemical reactions, have also shownwide variations in their reproducibilities (11, 15, 17, 18).The discrimination of a method is an estimate of its ability
to differentiate between two unrelated strains. According toSimpson's index of diversity, the typing on SITZ had aD =0.757 for C. albicans. This means that if two strains of C.albicans are taken randomly from the population, then75.7% of the time they would fall into different types. Themethod showed higher discriminatory indices for C. guillier-mondii, C. parapsilosis, and C. tropicalis. The lowest dis-criminatory index was found for C. krusei, since 70% of theisolates fell into the same biotype. The discriminatory abilityof C. albicans typing on STTZ agar seems to be in anintermediate position among previously published methods.It was lower than resistotyping (D = 0.899) and DNA typing(D = 0.868) but higher than the killer system (D = 0.724),immunoblotting (D = 0.679), enzyme biovars (D = 0.549),monoclonal antibodies (D = 0.492), and serotyping (D =0.438) (3, 6).Compared with other systems for strain differentiation,
the S1TZ agar morphotyping system has the advantages ofsimplicity, low cost (morphotype determination for ninestrains costs approximately the same as a Sabouraud dex-trose agar plate), and the potential for typing C. albicans andother medically important Candida species. The main disad-vantage is the impossibility of typing T. glabrata strains.
We thank D. W. R. Mackenzie and Tyrone Pitt for criticalrevisions of the manuscript; A. Cassone for strains 10261, 3153,CA2, and IR2h; Susan Howell for work on karyotyping; J. M.Torres Rodriquez, G. C. White, and M. C. Cabronero for strains;and E. Gonzalez-Miranda for estimable help with the graphic work.
This work was supported by grant UPV 093.327-0119/89 from theUniversidad del Pais Vasco-Euskal Herriko Unibertsitatea andgrant SGV 093.327-SC 04/90 from the Departamento de Educaci6n,Universidades e Investigaci6n del Gobierno Vasco.
REFERENCES1. Hasenclever, H. F., and W. 0. Mitchell. 1963. Antigenic studies
of Candida. IV. The relationship of the antigenic groups ofCandida albicans to their isolation from various clinical speci-mens. Sabouraudia 2:201-204.
2. Howell, S. A., and W. C. Noble. 1990. Typing tools for theinvestigation of epidemic fungal infection. Epidemiol. Infect.105:1-9.
3. Hunter, P. R. 1991. A critical review of typing methods forCandida albicans and their applications. Crit. Rev. Microbiol.17:417-434.
4. Hunter, P. R., and C. A. M. Fraser. 1988. Use of modifiedresistogram to type Candida albicans isolated from cases ofvaginitis and from faeces in the same geographical area. J. Clin.Pathol. 40:1159-1161.
5. Hunter, P. R., C. A. M. Fraser, and D. W. R. Mackenzie. 1989.Morphotype markers of virulence in human candidal infections.J. Med. Microbiol. 28:85-91.
6. Hunter, P. R., and M. A. Gaston. 1988. Numerical index of thediscriminatory ability of typing systems: an application ofSimpson's index of diversity. J. Clin. Microbiol. 26:2465-2466.
7. Lehmann, P. F., B. J. Kemker, C. B. Hsiao, and S. Dev. 1989.Isoenzyme biotypes of Candida species. J. Clin. Microbiol.27:2514-2521.
8. McCreight, M. C., D. W. Warnock, and M. V. Martin. 1985.Resistogram typing of Candida albicans isolates from oral andcutaneous sites in irradiated patients. Sabouraudia 23:403-406.
9. Merz, W. G. 1990. Candida albicans strain delineation. Clin.
VOL. 30, 1992
on April 8, 2019 by guest
http://jcm.asm
.org/D
ownloaded from

J. CLIN. MICROBIOL.
Microbiol. Rev. 3:321-334.10. Merz, W. G., C. Connelly, and P. Hieter. 1988. Variation of
electrophoretic karyotypes among clinical isolates of Candidaalbicans. J. Clin. Microbiol. 26:842-845.
11. Odds, F. C., and A. B. Abbott. 1980. A simple system for thepresumptive identification of Candida albicans and differentia-tion of the strains within the species. Sabouraudia 18:301-317.
12. Pearce, M., and S. A. Howell. 1991. Restriction fragment lengthpolymorphism analysis of azole-resistant and azole-susceptibleCandida albicans strains. J. Clin. Microbiol. 29:1364-1367.
13. Phongpaichit, S., D. W. R. Mackenzie, and C. Fraser. 1987.Strain differentiation of Candida albicans by morphotyping.Epidemiol. Infect. 99:421-428.
14. Polonelli, L., C. Archibusacci, M. Sestito, and G. Morace. 1983.
Killer system: a simple method for differentiating Candidaalbicans strains. J. Clin. Microbiol. 17:774-780.
15. Roman, M. C., and M. J. L. Sicilia. 1983. Preliminary investi-gation of Candida albicans biovars. J. Clin. Microbiol. 18:430-431.
16. Slutsky, B., J. Buffo, and D. R. Soll. 1985. High-frequencyswitching of colony morphology in Candida albicans. Science230:666-669.
17. Williamson, M. I., L. P. Samaranayake, and T. W. MacFarlane.1986. Biotypes of oral Candida albicans and Candida tropicalisisolates. J. Med. Vet. Mycol. 24:81-84.
18. Williamson, M. I., L. P. Samaranayake, and T. W. MacFarlane.1987. A new simple method for biotyping Candida albicans.Microbios 51:159-167.
2752 NOTES
on April 8, 2019 by guest
http://jcm.asm
.org/D
ownloaded from
Recommended

















