
Proc. Natl Acad. Sci. USAVol. 78, No. 2, pp. 810-814, February 1981Biochemistry
Abundant pseudogenes for small nuclear RNAs are dispersed in thehuman genome
(multigene families/Alu family sequence)
RICHARD A. DENISON, SCOTT W. VAN ARSDELL, LAUREL B. BERNSTEIN, AND ALAN M. WEINER
Department of Molecular Biophysics and Biochemistry, Yale University, 333 Cedar Street, P.O. Box 3333, New Haven, Connecticut 06510
Communicated by Donald D. Brown, October 24, 1980
ABSTRACT We have cloned and partially characterized 24loci from the human genome which are complementary to U1, U2,or U3, the three major species of small nuclear RNA (snRNA) inHeLa cells. When compared to the known Ul (human) and U2(rat) snRNA sequences, the DNA sequences we report here forthe complementary regions from two of the clones, Ul.11 andU2.7, reveal the presence of truncated and divergent gene copies.Furthermore, most if not all of the 24 cloned loci contain genecopies that are significantly divergent from the homologous HeLasnRNA species because DNA from every recombinant phage ex-cept U1.7 and U1. 15 proved unable to form snRNA-DNA hybridswhich protect full-length HeLa snRNA from mild digestion withribonuclease T1. Hence, we refer to these loci as snRNA pseu-dogenes. In both clones Ul. 11 and U2.7, an element of the dom-inant middle repetitive DNA sequence family in the human ge-nome, the Alu family, is located upstream from the snRNA pseu-dogene and in the same orientation. Alu elements in the samelocation and orientation relative to bona fide genes have previ-ously been found in the human ,B-globin gene cluster [Duncan, C.H., Biro, P. A., Choudary, P. V., Elder, J. T., Wang, R. C., Forget,G. B., deRiel, J. K. & Weissman, S. M. (1979) Proc. NatL Acad.Sci USA 76, 5095-5099]. We discuss the significance of these find-ings in relation to the nature of snRNA multigene families andother reported examples of pseudogenes.
Among the various classes of molecules found in all eukaryoticcells, the small nuclear RNAs (snRNAs) have received relativelylittle attention until recently. The sequences of eight of thesesmall homogeneous RNA species from mammalian cells (Ul,U2, U3A, U3B, U4, U5, U6, and 4.5S RNA) have been deter-mined (1-6). U3 snRNA is found in the nucleolus, and othersnRNA species are associated with either the nucleoskeleton orthe nucleoplasm (7). At least three of the snRNAs are subjectto considerable evolutionary conservation: Ul and U2 snRNAfrom chicken, rat, mouse, and human cells yield identical ri-bonuclease Ti fingerprints (1, 8, 9), and the RNA sequences ofU1 from HeLa cells and Ula from rat Novikoff hepatoma differin only 2 of 165 positions (1). We have also shown that a snRNAfrom the lower eukaryotic cellular slime mold Dictyosteliumdiscoideum is over 40% homologous to U3 snRNA from the rat(10).Two recent developments have prompted new interest in the
intracellular packaging and function of the snRNAs. First, an-tibodies produced by patients with the autoimmune diseasesystemic lupus erythematosus have been shown to recognizediscrete cellular components that contain snRNAs complexedwith a defined set of proteins (8), thus opening the way for de-tailed structural studies of small nuclear ribonucleoprotein par-ticles. Second, Lerner et al. (9) and Rogers and Wall (11) haveobserved a striking complementarity between the 5' end of UlsnRNA and the sequences that span splice junctions in nuclearRNA, suggesting a possible role, at least for Ul snRNA, in thesplicing of mRNA precursors.
Data concerning the organization and expression of snRNAgenes are scant. Previous estimates of the reiteration frequencyof snRNA genes in mammals had ranged from 100 (12) to 1000(13) copies, based on hybridization of purified snRNA speciesto bulk DNA in solution. Our own recent results suggest thatmany of the sequences complementary to the three majorsnRNA species (U1, U2, and U3) in human cells are not inter-spersed with each other (ref. 14 and unpublished observations).Although no definitive data exist regarding transcription of thesnRNAs, the inhibition of U1 and U2 biosynthesis by both a-amanitin (15) and 5,6-dichloro-1-,8-D-ribofuranosylbenzimida-zole (16) may imply that RNA polymerase II is responsible forthe transcription of at least certain snRNAs. Furthermore, thesensitivity of U1 and U2 biosynthesis to low doses of ultravioletirradiation suggests that these snRNA species are derived fromtranscription units as large as 5 kilobases (kb) (17).We originally decided to clone the genes encoding snRNAs
in the hope that a systematic investigation of their organizationand expression would also open up new avenues for understand-ing the function of the snRNAs themselves. We report here thesurprising result that most, if not all, of the genomic loci com-plementary to snRNAs U1, U2, and U3 that we have examinedcontain divergent, and in some cases truncated, gene copieswhen compared with the corresponding HeLa cell snRNAspecies.
MATERIALS AND METHODSA library of 15-kb partial EcoRI fragments of human placentalDNA in the A vector Charon 4A (18) was kindly supplied by A.Biro, P. V. Choudary, J. T. Elder, and S. M. Weissman. Plaqueswere screened by the method of Benton and Davis (19); we usedas probes U1, U2, and U3 snRNAs isolated from HeLa cells andlabeled in vitro at the 3' end with 5'-32P-labeled pCp and T4RNA ligase as described (14). As little as 106 cpm of snRNA ata specific activity of 106 cpm/,ug was sufficient to screen six 140-mm nitrocellulose filters, each bearing 8000 plaques. With eachprobe, 0. 1% of the plaques were scored as positive and re-purified for further study. Small quantities of recombinantDNA were prepared from 4-ml NZY cultures (20); larger quan-tities were prepared from recombinant phage grown and pu-rified as described (18). EcoRI fragments that hybridized withU1 or U2 were subeloned into pBR322, and the recombinantplasmids were designated pUl. 11 and pU2.7.
All experiments involving recombinant molecules were per-formed under P2-EK2 conditions as specified by the revisedNational Institutes of Health guidelines for recombinant DNAresearch.
RESULTSThe human genomic library in Charon 4A was screened sepa-rately with U1, U2, and U3 snRNA probes to yield 13 recom-
Abbreviations: kb, kilobase; snRNA, small nuclear RNA.
810
The publication costs ofthis article were defrayed inpart by page chargepayment. This article must therefore be hereby marked "advertise-ment" in accordance with 18 U. S. C. §1734 solely to indicate this fact.

Proc. Nati. Acad. Sci. USA 78 (1981) 811
pUI.I
00541282-~~~~~~~~~~C 0f) D0
D__4 LtJ
Alu Family U1 Pseudogene
pU27
lOObp 0 _q-W- ~~~~~~~~~zze "~czz
a GZ nao , .c 00{XO = ,uu xtI m a3AnFi <U2 m
Alu himily U2 Pseudogene
FIG. 1. Restriction maps of the region complementary to U1 andU2 snRNA in the plasmid subclones pU1.11 (Upper) and pU2.7(Lower). The Alu I fragment from pU1.11 was cloned into pBR322 byusing EcoRI linkers. The EcoRi/Hpa I fragment from U2.7 was ob-tained from aHpa I digest ofthe 3-kbEcoRI fragment carried in pU2.7.TheDNA sequencing strategy is shown below each map. bp, Base pairs.
binant phage complementary to U1 (designated Ul.1, U1.2,.), 7 complementary to U2, and 4 complementary to U3.
Each of the 24 phage contains 15 kb of human DNA derived
from a distinct genomic locus because the size of the EcoRIrestriction fragment complementary to the snRNA is differentin each clone (data not shown); moreover, when hybridized tototal cellular RNA labeled in vivo, each recombinant phagetested (Ul. 11, as well as all the U2 and U3 loci) selected onlythe expected snRNA, demonstrating that sequences comple-mentary to one snRNA are not generally interspersed with thosecomplementary to the other two (ref. 14 and unpublished data).We initially chose the recombinant phage Ul.11 and U2.7
for detailed characterization. The unique region complemen-tary to the snRNA in each phage was localized as described (10)and the sequence was determined according to Maxam andGilbert (21). Fig. 1 presents a high-resolution map of the rel-evant restriction fragments from each of the plasmid pBR322subelones pUl.11 and pU2.7; Fig. 2 compares the DNA se-quence of these cloned human genomic loci with the knownsequence of human Ul and rat U2 snRNA (1, 2). We did notexpect such striking divergence between the RNA and DNAsequences, even in the heterologous comparison of rat U2 RNAwith the human genomic clone pU2.7; rat and human U2snRNAs appear indistinguishable as judged both by RNase T1fingerprints and by secondary analysis of T1 oligonucleotidesafter redigestion with pancreatic RNase A (refs. 8 and 9; J. A.Steitz, personal communication). Furthermore, the predictedT1 oligonucleotides for a snRNA transcribed from pUl.l1 or
1 0 2 0 3 0 4 0 5 0
HUMAN Ul RNA m2'2'7Gppp AMACU| AC CUGGCAGGG G AGAUA CC AUGAUCA C GAAGGUGG U UUUCCCAGGG C
pUl.11 HUMAN PSEUDOGENE ATACT CGT CTGGCAGGG A AGATA AA ATGATCA T GAAGGTGG C TTTCCCAGGG T
_ o,00 ,0 190 __ 110 120 130
GAG GC UUAU CC AUUGCACUC C GG A UGUGC U GACCCCUGC G AUUUCCCC A AA U GUGGGAAACU C G A CUGCAUAAUUUGUG
GAG AT TTAT TT ATTGCACTC T GG G TGTGC A GACCCCTGC A ATTTCCCC G AA A GTGGGAAACT T G G CTGCATAATTTGTG
140 150 1960
G U|AG U GGGG G AC U GCGUUCGCGCU U UCCCCUGQOHT T CT T CCTT G GT TAAAAAGAAAAA T AGTAGGC
10 20 30 40 50
RAT U2 RNA m2'2'7Gppp AC GCGG C(CU)(CPP)|CC | UUUUGGCUAAGAUCAAGUGOAGWAlQ GPl UC UAU
pU2.7 HUMAN PSEUDOGENE ATC A C C T C T C AG T G TTTTGGCTAAGATCAAGTGTAS'TATCTGTT CT TATI
60 70 80 90 100 110 120
C AGUU U AAPAUCU UC GAUA C GUCCUCUAUC C GAGGA C A A UAP U APUAAAUGG A UUUUUGGA A C U A GGAG U UGG
A AGTTT AATATCT GATA T GTCCTCTATC T GAGGA T A C TAT ATTAAATGG G TTTTTGGA G C A G GGAG A TGG
130 140 150 160 170 180 190
A AUAGGAGCUU G CUC C GUCCAC C U CACGCAUCGA CC U GGUAUUGC GC AGUACC CU CAGGAA C GGUGCACC AOHG ATAGGAGCTT A CTC T GTCCAC T CC A GGTATTGC AGTACC TC CAGGAA T GGTGCACC C
FIG. 2. Comparison ofRNA sequences for U1 RNA from HeLa cells and U2 RNA from rat Novikoffhepatoma (1, 2) with the corresponding DNAsequences from human clones pUl. 11 and pU2.7. Boxes denote homologous sequences. Numbers refer to positions in theRNA sequences. Ambiguitiesin the reported rat U2 RNA sequence (2) at positions 10-11 and 12-14 are enclosed by parentheses and aligned to give the best match to the pU2.7DNA sequence. Asterisks (*) at positions 6 and 7 in U1 RNA denote modified uridine residues, most likely pseudouridines (1). Modified residues areabbreviated as follows: m, 2'-O-methyl; m2,N6-methyl-2'-O-methyl; 4i, pseudouridine; M2'2'7, N2,2'7-trimethyl.
4
Biochemistry: Denison et aL
p
812 Biochemistry: Denison et aL
pU2.7 are incompatible with the observed Ul and U2 finger-prints. We therefore refer to these loci as pseudogenes.
Both pseudogene sequences exhibit transitional and trans-versional single base changes, and pU2.7 also contains internalinsertions and deletions relative to the rat U2 snRNA sequence.In particular, the pUl. 11 sequence corresponding to the 5'.endof U1 RNA contains three mismatched bases at positions 7-9precisely in the region of U1 that is complementary to splicejunction sequences in nuclear RNA (9, 11). The pUl. 1 pseu-dogene also lacks any sequence corresponding to the last 32nucleotides of U1 RNA, and no sequences resembling the 3'end of U1 RNA could be found in 76 additional base pairs of3' flanking DNA (data not shown). Discounting truncation atthe 3' end, the overall sequence mismatch up to position 133in pUl.11 is 16% (21 differences in 132 nucleotides), and theRNA and DNA sequences maintain perfect registry over thisentire region. The homology between pU2.7 and rat U2 RNAis somewhat better, with only 10% mismatch and no truncation,but the registry is poor (18 differences in the 173 nucleotidesaligned in Fig. 2, with insertion of 4 and deletion of 21 nu-cleotides). The lack of registry might in part reflect sequencedifferences between human U2 RNA and the published rat U2sequence which was determined by classical RNA fingerprint-ing techniques (ref. 2; also see Note Added in Proof).Ti RNase Protection Experiments. To determine whether
the other 22 cloned genomic loci also contain pseudogenes, wescreened DNA from these phage for the ability to formsnRNA-DNA hybrids resistant to mild digestion with T1 RNaseas described (14). Recombinant DNAs were immobilized onnitrocellulose filters and hybridized with purified HeLa U1,U2, or U3 RNA uniformly labeled in vivo with [32P]phosphate.After treatment with 10 units of T1 RNase per ml at high saltconcentration under conditions that leave a perfect RNA-DNAduplex intact (14), the protected snRNA fragments were elutedfrom the filters and their sizes were determined on a 15% poly-acrylamide gel. U2 and U3 snRNA eluted directly from dupli-cate filters without RNase treatment were included as a control(see Fig. 4, lanes U2 and U3).
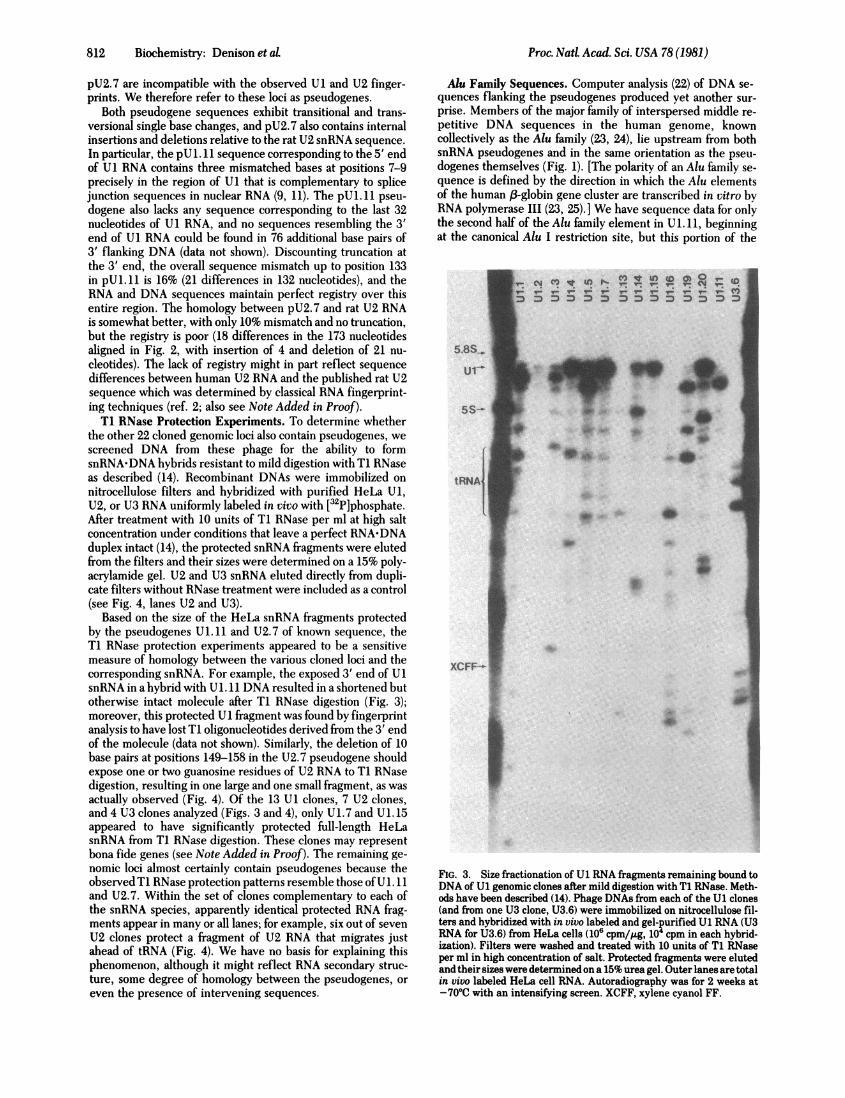
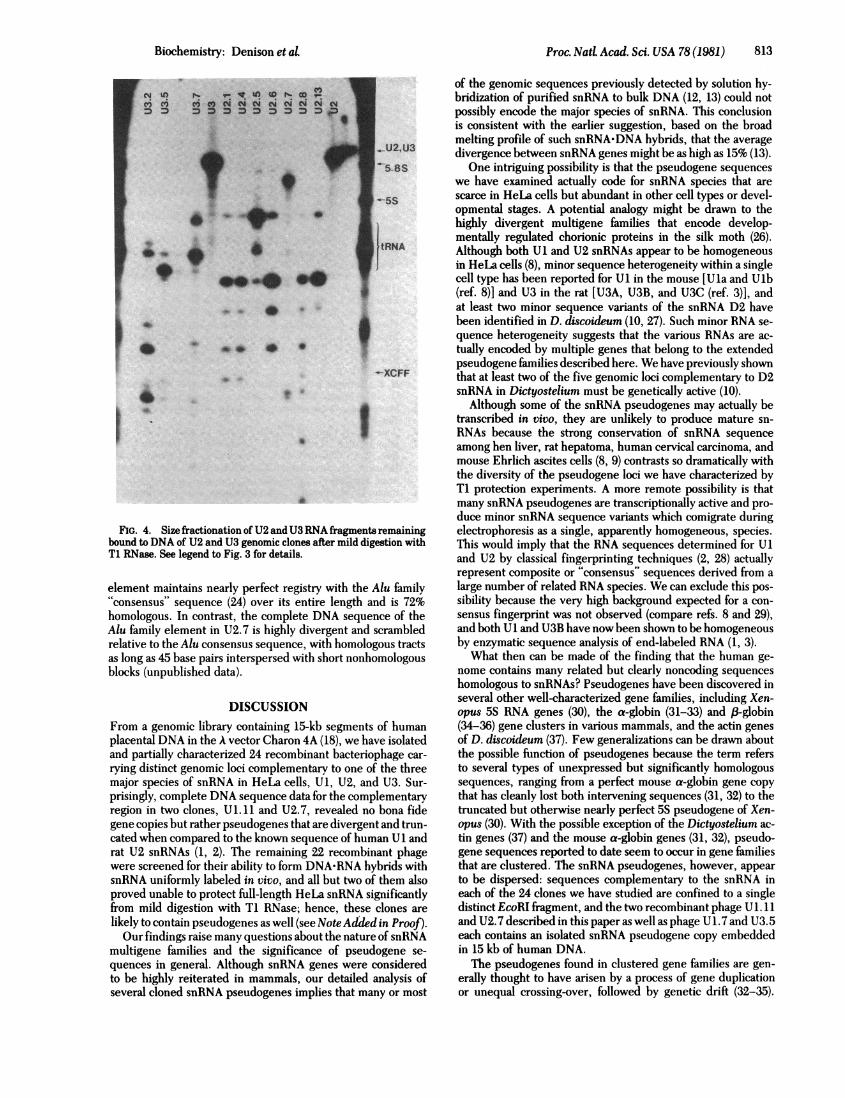
Based on the size of the HeLa snRNA fragments protectedby the pseudogenes Ul. ll and U2.7 of known sequence, theT1 RNase protection experiments appeared to be a sensitivemeasure of homology between the various cloned loci and thecorresponding snRNA. For example, the exposed 3' end of U1snRNA in a hybrid with U1. 11 DNA resulted in a shortened butotherwise intact molecule after T1 RNase digestion (Fig. 3);moreover, this protected Ul fragment was found by fingerprintanalysis to have lost T1 oligonucleotides derived from the 3' endof the molecule (data not shown). Similarly, the deletion of 10base pairs at positions 149-158 in the U2.7 pseudogene shouldexpose one or two guanosine residues of U2 RNA to T1 RNasedigestion, resulting in one large and one small fragment, as wasactually observed (Fig. 4). Of the 13 U1 clones, 7 U2 clones,and 4 U3 clones analyzed (Figs. 3 and 4), only U1. 7 and U1. 15appeared to have significantly protected full-length HeLasnRNA from Ti RNase digestion. These clones may representbona fide genes (see Note Added in Proof). The remaining ge-nomic loci almost certainly contain pseudogenes because theobserved Ti RNase protection patterns resemble those ofU 1. 11and U2.7. Within the set of clones complementary to each ofthe snRNA species, apparently identical protected RNA frag-ments appear in many or all lanes; for example, six out of sevenU2 clones protect a fragment of U2 RNA that migrates justahead of tRNA (Fig. 4). We have no basis for explaining thisphenomenon, although it might reflect RNA secondary struc-ture, some degree of homology between the pseudogenes, oreven the presence of intervening sequences.
Alu Family Sequences. Computer analysis (22) of DNA se-quences flanking the pseudogenes produced yet another sur-prise. Members of the major family of interspersed middle re-petitive DNA sequences in the human genome, knowncollectively as the Alu family (23, 24), lie upstream from bothsnRNA pseudogenes and in the same orientation as the pseu-dogenes themselves (Fig. 1). [The polarity of an Alu family se-quence is defined by the direction in which the Alu elementsof the human /3-globin gene cluster are transcribed in vitro byRNA polymerase III (23, 25). ] We have sequence data for onlythe second half of the Alu family element in U1. 11, beginningat the canonical Alu I restriction site, but this portion of the
5.8S,U'..
M*.'
W:. s#I. **.f::
wAl...:
0-*F * ,Sf~~~~~~~~~~~~~~~~~~i:f
Sff oll
I
xCFF-,-
:
-
---
::
X
a.:
FIG. 3. Size fractionation of U1 RNA fragments remaining bound toDNA of U1 genomic clones after mild digestion with T1 RNase. Meth-ods have been described (14). Phage DNAs from each of the U1 clones(and from one U3 clone, U3.6) were immobilized on nitrocellulose fil-ters and hybridized with in vivo labeled and gel-purified U1 RNA (U3RNA for U3.6) from HeLa cells (106 cpm/,Lg, 104 cpm in each hybrid-ization). Filters were washed and treated with 10 units of T1 RNaseper ml in high concentration of salt. Protected fragments were elutedand their sizes were determined on a 15% urea gel. Outer lanes are totalin vivo labeled HeLa cell RNA. Autoradiography was for 2 weeks at-70TC with an intensifying screen. XCFF, xylene cyanol FF.
Proc. Natl. Acad. Sci. USA 78 (1981)
N m V 0 P- m v 0 0 m 0 V- 0T- V-- -. r: V7 C4 -v-; ip; -: ,-' v-. r: v- r- v- v- r- r; M= :) :3 = M :2 M M = :n M = =
Biochemistry:DenisonetaL~~~~~Proc.NatL Acad. Sci. USA 78(1981) 813
CeGo4)
0
.-U2,U3
5.BS
~-5S
tRNA
W, 40 0
4 *40 * *
~-XCFF
4Y4
0I
FIG. 4. Size fractionation of U2 andU3 RNA fratgments remainingbound to DNA of U2 and U3 genomic clones after mild digestion with
Ti RMase. See legend to Fig. 3 for details.
element maintains nearly perfect registry with the Alu family"tconsensus" sequence (24) over its entire length and is 72%
homologous. In contrast, the complete DNA sequence of the
Alu family element in U2.7 is highly divergent and scrambled
relative to the Alu consensus sequence, with homologous tracts
as long as 45 base pairs interspersed with short nonhomologousblocks (unpublished data).
DISCUSSION
From a genomic library containing 15-kb segments of human
placental DNA in the A vector Charon 4A (18), we have isolated
and partially characterized 24 recombinant bacteriophage car-
rying distinct genomic loci complementary to one of the three
major species of snRNA in HeLa cells, Ul, U2, and U3. Sur-
prisingly, complete DNA sequence data for the complementaryregion in two clones, Ul1. 11 and U2.7, revealed no bona fide
gene copies but rather pseudogenes that are divergent and trun-
cated when compared to the known sequence of human Ul1 andrat U2 snRNAs (1, 2). The remaining 22 recombinant phagewere screened for their ability to form DNA-RNA hybrids with
snRNA uniformly labeled in vivo, and all but two of them also
proved unable to protect full-length HeLa snRNA siguificantlyfrom mild digestion with Ti RNase; hence, these clones are
likely to contain pseudogenes as well (see Note Added in Proof).Our findings raise many questions about the nature of snRNA
multigene families and the siguificance of pseudogene se-
quences in general. Although snRNA genes were considered
to be highly reiterated in mammals, our detailed analysis of
several cloned snRNA pseudogenes implies that many or most
of the genomic sequences previously detected by solution hy-bridization of purified snRNA to bulk DNA (12, 13) could notpossibly encode the major species of snRNA. This conclusionis consistent with the earlier suggestion, based on the broadmelting profile of such snRNA-DNA hybrids, that the averagedivergence between snRNA genes might be as high as 15% (13).One intriguing possibility is that the pseudogene sequences
we have examined actually code for snRNA species that arescarce in HeLa cells but abundant in other cell types or devel-opmental stages. A potential analogy might be drawn to thehighly divergent multigene families that encode develop-mentally regulated chorionic proteins in the silk moth (26).Although both U1 and U2 snRNAs appear to be homogeneousin HeIa cells (8), minor sequence heterogeneity within a singlecell type has been reported for Ul1 in the mouse [Ula and Ulb(ref. 8)] and U3 in the rat [U3A, U3B, and U3C (ref. 3)], andat least two minor sequence variants of the snRNA D2 havebeen identified in D. discoideum (10, 27). Such minor RNA se-quence heterogeneity suggests that the various RNAs are ac-tually encoded by multiple genes that belong to the extendedpseudogene families described here. We have previously shownthat at least two of the five genomic loci complementary to D2snRNA in Dictyostelium must be genetically active (10).
Although some of the snRNA pseudogenes may actually betranscribed in vivo, they are unlikely to produce mature sn-RNAs because the strong conservation of snRNA sequenceamong hen liver, rat hepatoma, human cervical carcinoma, andmouse Ehrlich ascites cells (8, 9) contrasts so dramatically withthe diversity of the pseudogene loci we have characterized byTi protection experiments. A more remote possibility is thatmany snRNA pseudogenes are transcriptionally active and pro-duce minor snRNA sequence variants which comigrate duringelectrophoresis as a single, apparently homogeneous, species.This would imply that the RNA sequences determined for UlIand U2 by classical fingerprinting techniques (2, 28) actuallyrepresent composite or "consensus"' sequences derived from alarge number of related RNA species. We can exclude this pos-sibility because the very high background expected for a con-sensus fingerprint was not observed (compare refs. 8 and 29),and both Ul1 and U3B have now been shown to be homogeneousby enzymatic sequence analysis of end-labeled RNA (1, 3).What then can be made of the finding that the human ge-
nome contains many related but clearly noncoding sequenceshomologous to snRNAs? Pseudogenes have been discovered inseveral other well-characterized gene families, including Xen-opus 5S RNA genes (30), the a-globin (31-33) and &3globin(34-36) gene clusters in various mammals, and the actin genesof D. discoideum (37). Few generalizations can be drawn aboutthe possible function of pseudogenes because the term refersto several types of unexpressed but siguificantly homologoussequences, ranging from a perfect mouse a-globin gene copythat has cleanly lost both intervening sequences (31, 32) to thetruncated but otherwise nearly perfect 5S pseudogene of Xen-opus (30). With the possible exception of the Dictyostelium ac-tin genes (37) and the mouse a-globin genes (31, 32), pseudo-gene sequences reported to date seem to occur in gene familiesthat are clustered. The snRNA pseudogenes, however, appearto be dispersed: sequences complementary to the snRNA ineach of the 24 clones we have studied are confined to a singledistinct EcoRI fragment, and the -two recombinant phage Ul1. 11and U2.7 described in this paper as well as phage U1.-7 and U3.5each contains an isolated snRNA pseudogene copy embeddedin 15 kb of human DNA.The pseudogenes found in clustered gene families are gen-
erally thought to have arisen by a process of gene duplicationor unequal crossing-over, followed by genetic drift (32-35).
0If0
AMM4w
4,~
I
Biochemistry: Denison et aL
I

814 Biochemistry: Denison et aL
Although this mechanism may also account for generation of thepseudogenes discussed here, the dispersion, high reiterationfrequency, and sequence diversity of snRNA pseudogenesmight have functional significance as well: the loci may servein a regulatory capacity (32, 38), for example, by virtue of theirability to form base pairs with the corresponding snRNAs.We were surprised to find upstream Alu family sequences
with the same polarity as the U1. 11 and U2.7 pseudogenes, andwe have recently learned that two Alu family sequences forma hyphenated palindromic sequence surrounding a human U6snRNA pseudogene, with the upstream Alu sequence also inthe same orientation as the pseudogene itself (C. H. Duncan,P. V. Choudary, and S. M. Weissman, personal communica-tion). Middle repetitive DNA sequences belonging to the Alufamily precede both the P- and y-globin gene cluster in humans(23, 25), and when used in vitro as templates for RNA poly-merase III, the human Alu sequences produce discrete RNAtranscripts with the same polarity as the globin genes them-selves (25). A similar arrangement of middle repetitive se-quences may also occur in the -like globin gene cluster of therabbit (39). One reasonable hypothesis is that in mammals, Alufamily sequences or their equivalent (23, 40) often occur up-stream from and in the same orientation as RNA polymerase IItranscription units. Such a sequence arrangement may havesurvived whatever process generated the pseudogenes we havestudied.We have not yet identified bona fide genes encoding U1, U2,
or U3 snRNA. However, we may have selected against theseloci during construction, propagation, or screening of the re-combinant phage library. It should be possible to isolate truesnRNA genes from a more complete library by using proceduresthat distinguish between genes and pseudogenes. We believethat further analysis of snRNA pseudogenes will lead to a betterunderstanding of their origin and possible function.Note Added in Proof. The DNA sequence of clone U1. 15 correspondsto that of human U1 RNA in all but 1 of 165 positions and is thereforelikely to represent a bona fide gene; clone U1.7 contains a slightly di-vergent pseudogene. We have also obtained a revised sequence for ratU2 RNA (R. Reddy, personal communication). The U2.7 pseudogenemaintains nearly perfect registry with this revised sequence except fora single deletion of -12 nucleotides centered at position 150.
We thank Steve Mount and Joan Steitz for many enlightening dis-cussions, and Joe Hendrick for an endless supply of HeLa cells. Thiswork was supported in part by National Science Foundation GrantsPCM 76-81524 and PCM 78-21799 and National Institutes of HealthGrant GM26312.
1. Branlant, C., Krol, A., Ebel, J. P., Lazar, E., Gallinaro, H., Jacob,M., Sri-Widada, J. & Jeanteur, P. (1980) Nucleic Acids Res. 8,4143-4154.
2. Shibata, H., Ro-Choi, T. S., Reddy, R., Choi, Y. C., Henning,D. & Busch, H. (1975)J. Biol Chem. 250, 3903-3920.
3. Reddy, R., Henning, D. & Busch, H. (1979)J. Biol. Chem. 254,11097-11105.
4. Reddy, R. & Busch, H., The Cell Nucleus, in press.5. Epstein, P., Reddy, R., Henning, D. & Busch, H. (1980)J. Biol.
Chem. 255, 8901-8906.
6. Harada, F. & Kato, N. (1980) Nucleic Acids Res. 8, 1273-1285.7. Zieve, G. & Penman, S. (1976) Cell 8, 19-31.8. Lerner, M. R. & Steitz, J. A. (1979) Proc. Natl Acad. Sci. USA 76,
5495-5499.9. Lerner, M. R., Boyle, J. A., Mount, S. M., Wolin, S. L. & Steitz,
J. A. (1980) Nature (London) 283, 220-224.10. Wise, J. A. & Weiner, A. M. (1980) Cell 22, 109-118.11. Rogers, J. & Wall, R. (1980) Proc. Natl Acad. Sci. USA 77,
1877-1879.12. Marzluff, W. F., White, E. L., Benjamin, R. & Huang, H. C. C.
(1975) Biochemistry 14, 3715-3724.13. Engberg, J., Hellung-Larsen, P. & Frederiksen, S. (1974) Eur. J.
Biochem. 41, 321-328.14. Weiner, A. M. (1980) Cell 22, 209-218.15. Jensen, E. G., Hellung-Larsen, P. & Frederiksen, S. (1979) Nu-
cleic Acids Res. 6, 321-330.16. Tamm, I., Kikuchi, T., Darnell, J. E., Jr. & Salditt-Georgiev, M.
(1980) Biochemistry 19, 2743-2748.17. Elicieri, G. L. (1979) Nature (London) 279, 80-81.18. Blattner, F. R., Williams, B. G., Blechl, A. E., Denniston-
Thompson, K., Faber, H. E., Furlong, L. A., Grunwald, D. J.,Kiefer, D. O., Moore, D. O., Schumm, J. W., Sheldon, E. L.& Smithies, 0. (1977) Science 196, 161-169.
19. Benton, W. D. & Davis, R. W. (1977) Science 196, 180-182.20. Blattner, F. R., Blechl, A. E., Denniston-Thompson, K., Faber,
H. E., Richards, J. A., Slightom, J. L., Tucker, P. W. & Smithies,0. (1978) Science 202, 1279-1284.
21. Maxam, A. M. & Gilbert, W. (1980) Methods Enzymol. 65,499-560.
22. Queen, C. & Korn, C. J. (1980) Methods Enzymot 65, 595-609.23. Jelinek, W. R., Toomey, T. P., Leinwand, L., Duncan, C. H.,
Biro, P. A., Choudary, P. V., Weissman, S. M., Rubin, C. M.,Houck, C. M., Deininger, P. L. & Schmid, C. W. (1980) Proc. NatiAcad. Sci. USA 77, 1398-1402.
24. Rubin, C. M., Houck, C. M., Deininger, P. L., Friedmann, T.& Schmid, C. W. (1980) Nature (London) 284, 372-374.
25. Duncan, C., Biro, P. A., Choudary, P. V., Elder, J. T., Wang, R.C., Forget, B. G., deRiel, J. K. & Weissman, S. M. (1979) Proc.Natl Acad. Sci. USA 76, 5095-5099.
26. Jones, C. W., Rosenthal, N., Rodakis, G. C. & Kafatos, F. C.(1979) Cell 18, 1317-1332.
27. Wise, J. A. & Weiner, A. M., J. Biol. Chem., in press.28. Reddy, R., Ro-Choi, T. S., Henning, D. & Busch, H. (1974)J. Biol
Chem. 249, 6486-6494.29. Robertson, H. D., Dickson, E. & Jelinek, W. (1977)J. Mol Biol
115, 571-589.30. Miller, J. R., Cartwright, E. M., Brownlee, G. G., Federoff, N.
& Brown, D. D. (1978) Cell 13, 717-725.31. Nishioka, Y., Leder, A. & Leder, P. (1980) Proc. Natl Acad. Sci.
USA 77, 2806-2809.32. Vanin, E. F., Goldberg, G. I., Tucker, P. W. & Smithies, 0. (1980)
Nature (London) 286, 222-226.33. Proudfoot, N. & Maniatis, T. (1980) Cell 21, 537-544.34. Lacy, E. & Maniatis, T. (1980) Cell 21, 545-553.35. Jahn, C. L., Hutchinson, C. A., III, Phillips, S. A., Weaver, S.,
Haigwood, N. L., Voliva, C. F. & Edgell, M. H. (1980) Cell 21,159-168.
36. Cleary, M. L., Haynes, J. R., Schon, E. A. & Lingrel, J. B. (1980)Nucleic Acids Res. 8, 4791-4802.
37. Firtel, R. A., Timm, R., Kimmel, A. R. & McKeown, M. (1979)Proc. Natl Acad. Sci. USA 76, 6206-6210.
38. Davidson, E. H. & Britten, R. J. (1979) Science 204, 1052-1059.39. Shen, C.-K. J. & Maniatis, T. (1980) Cell 19, 379-391.40. Krayev, A. S., Kramerov, D. A., Skryabin, K. G., Ryskov, A. P.,
Bayev, A. A. & Georgiev, G. P. (1980) Nucleic Acids Res. 8,1201-1215.
Proc. Nad Acad. Sci. USA 78 (1981)
Recommended

















