Embed Size (px)
Citation preview

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002) 1
VETERINARIASociedad de Medicina Veterinaria del Uruguay
Año LXII Vol. 37 N° 147-148 Abril - Setiembre de 2002
Cerro Largo 1895 - Montevideo - Uruguay Tel-Fax (598-2) 408 6174 - 409 9458 - Email: [email protected]
Esta edición consta de 1600 ejemplares y se distribuye sin costo a todos los socios de la SMVU.Los contenidos y opiniones incluidos en los artículos son responsabilidad exclusiva de los autores.Se autoriza la reproducción parcial o total de lo editado mencionando la fuente.Por convenio de la SMVU y Facultad de Veterinaria (16-12-1988), el Dpto. de Documentación y Biblioteca de la Fac. deVeterinaria. Se realiza el canje internacional por otras publicaciones científicas.
Contenido
Editorial
Artículo Original
Estudio de la estructura genética de la raza Holando uruguayo mediante marcadores genéticos.Comparación intraracial.Kelly, E.L.; Mortari, N.; De Andrés; D., Postiglioni, A.. ................................................... 7
Artículo Original
Caracterización de un herpesvirus de un terneroAlonzo, P., et al. ........................................................................................................................15
Comunicación Corta
Divisiones del ciego y colon ascendente del conejo (Oryctolagus cuniculus)Möller, R.; Pérez, W.; Martin, E.. ..........................................................................................23
Diagnóstico
Presencia de nuevos géneros y especies de estrongilidos en los equinos en UruguayFalcón, J. D... ...........................................................................................................................27
Conferencia
La RabiaRossi, D... ...................................................................................................................................31

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002)2 2002

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002) 3
VETERINARIAISSN 0376 - 4362 - Indizada en: Vet-CD/BEASTCD
REDACTOR RESPONSABLE: Analía Cobo, DMVCONSEJO EDITOR “Profesor Walter García Vidal”: Pedro Bañales, DMV
Gonzalo Leaniz, DMVLuis Barros, DV, MSV, PhDDaniel Elhordoy, DV, FRCVSJacqueline Maisonnave, DMV, PhDMaría A. Solari, DV
Asesor Bibliotecológico: Elba Domínguez
ARBITROS de los TRABAJOS CIENTÍFICOS (1997 - 2001)
Berthelot, X. (DMV) FRANCIA Martin, E. (DMV) URUGUAYCamarotte, D. (DMV) URUGUAY Pérez Clariget, R. (DMV) URUGUAYCardelino, R. (Ing. Agr.) URUGUAY Pimentel, C. (DMV) BRASILCardozo, E. (DMV) URUGUAY Riet Correa, F. (DMV) BRASILCardozo, H. (DMV) URUGUAY Rodríguez, H. (DMV) SUECIACastells, D. (DMV) URUGUAY Theis, J.H. (DVM) USACattaneo, G. (DMV) CHILE Traldi, A. (DMV) BRASILEddi, C. (DMV) ARGENTINA Trejo González, A. (DC) MÉXICOFeinstein, R. (DMV) SUECIA Trica, G. (DMV) URUGUAYFlores, E. (DMV) CHILE Tortora, J. (DMV) MÉXICOLazaneo, E. (DMV) URUGUAY Uriarte, G. (DMV) URUGUAYLeites, O. (DMV) URUGUAY Weiblen, R. (DMV) BRASIL
CONSEJO DIRECTIVO (2000 - 2004)
Presidente: Dra. Analía Cobo LeturiaPresidente Suplente: Dr. Humberto Tommasino
Titulares: Dr. José Gallero Quadros Comisión Fiscal: Dr. Oscar FerreiraDr. Roberto Acuña Dr. Daniel AlzaDr. Carlos Morón Dr. Omar LandeiraDr. Eduardo Galagorri
: 2002

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002)4
INTEGRACIÓN de COMISIONESSEDE SOCIAL FINANZAS CURSOS Y ASUNTOSRafael Varela Oscar Ferreira CAPACITACION UNIVERSITARIOSJorge Butthyany Rafael Varela Oscar Ferreira Julio García LagosJuan José Mari Ariel Saez Eduardo Galagorry Angela RistaAlicia Baldovino BOLETÍN Y R.R.P.P. Juan José Mari Luis AlberteMERCOSUR Luis Delucchi Inés Sienra Gastón CossiaHugo Fontaiña Daniel Alza Ana de León Mario AlvarezJulio García Lagos M. Guadalupe CULTURA Y Carlos PereiraIgnacio Pereira Daniel Rossi DEPORTES Gabriel MaruriEugenio Perdomo Fernando Echezarreta Walter Faliveni DECRETO 160/97Angela Rista Alvaro Fernández Raúl Piaggio G. De GregorioLuis Barros Viviana Cuñarro Raquel Pérez Luis DelucchiJorge Baraibar REVISTA J. de Miquelerena Alvaro TrinidadOrgelio Cabrera María Solari ESTATUTOS REPRODUCCIONFESTEJOS Jacqueline Maisonnave Eduardo Galagorry Pedro BañalesElbio Sosa Daniel Elhordoy Joaquín Rossi Guillermo de NavasRafael Varela Luis Barros Gastón Casaux A. Durán del CampoAnalía Cobo Pedro Bañales Oscar Ferreira Luis CuencaMagela Damiani Margarita de Miquelerena Gabriel DuránMaría Raimondi
CENTRO DE VETERINARIOS DE LA SMVU
ARTIGAS DURAZNO MALDONADO RIO NEGRO SAN JOSERamón Rodríguez Ana Acuña Juan C. Dibarboure Carlos De Mateo Joaquín RossiLavalleja 234 Artigas 375 Veterinaria Maldonado Young, 19 de Abril 1920 Colón 523
Velázquez esq. MitreCANELONES FLORES PASO DE LOS TOROS RIVERA SORIANORamiro Díaz Héctor García Pintos Carlos Casadei Rafael Piazze Edgardo BelliniBatlle 304 Trinidad,Granja Roland Leandro Gómez 514 Luis A. de Herrera 536 Mercedes, Sánchez 811
CERRO LARGO FLORIDA PAYSANDU ROCHA TACUAREMBOAlberto Sanner Luis Albornoz Carlos Pepe Omar Pereyra Pedro DutraMelo, Esteban Vieira 658 Luis A. de Herrera 481 Uruguay 1189 Zorrilla de San Martín 157 Lab. Vet. “El Campo”
COLONIA LAVALLEJA PANDO SALTO TREINTA Y TRESHugo Betancour Amalia Villalba Alberto Varela Francisco Hermann Mónica BurgosJosé Artigas s/n Minas, Rodó 424 Wilson Ferreira 1017 Washinton Beltrán 69 Basilio Araújo 1038 AColonia Miguelete
RIO BRANCOPedro FleitasVirrey Aredondo
DELEGATURAS DE LA SMVU
CONAHSA FUNDACION “MARCOAníbal Ibarburu PODESTA”Oscar Ferreira Alvaro OliveraAgustín Landeira
COMISION ASESORRAAUDU C.J.P.P.U.Ana Terzhagui Walter FaliveniEduardo Galagorry Julia Saizar
Alicia BaldovinoC.H.L.C.H.Mariano CarballoJesús Falcón
ASOCIACIONES ESPECIALIZADAS QUEINTEGRAN LA SMVU
Comisión de Reproducción e Inseminación Artificial (CRIA)
Sociedad de Buiatría del Uruguay
Soc. Uruguaya de Vet. Especialistas enPequeños Animales (SUVEPA)
Soc. Uruguaya de Vet. Especialistas en
Animales Silvestres (SUVEAS)
Soc Veterinaria Especialistas en Cerdos (SVEC)
Asoc. Uruguaya de Veterinarios Laboratoristas (AUVELA)
Asoc. Vet. Esp. Protección Alimentos (ANEPA)
2002

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002) 5
Editorial
:5-5 (2002)
Estimados colegas, hemos sido llamadosa escribir el editorial de este número 147/148. Para ello hemos estado re-leyendoa muchos de los que nos precedieron yhemos podido comprobar muchas y muyinteresantes cosas.Algunos editoriales nos han recordadolos 91 años de la propia asociación gre-mial a la cual pertenecemos, los 93 añosde los estudios veterinarios en elUruguay, la creación allá por la décadadel 50 de la Asociación de Colaborado-ras de la Medicina Veterinaria, tan estre-chamente ligada a nuestra profesión.
Hemos repasado como se creó la Acade-mia Nacional de Veterinaria, logro biencercano en el tiempo y producto del es-fuerzo de muchos colegas.Hemos visto como los veterinariostrabajamos incansablemente en la luchacontra la Fiebre Aftosa, por mediode aquellos colegas que participaron encada etapa, desde «Di.L.F.A.» y desdeantes hasta la fecha, trabajando por másde 40 años, siendo creativos, dándoleimpulso y llevando sobre los hombrosesta campaña, muchas veces «luchan-do» en condiciones adversas.Hemos comprobado también que en todolo que a campañas sanitarias se refiere, seha desempeñado un rol protagónico,dándole a nuestro país un status sanita-rio reconocido en el mundo, ya se tratede bovinos, de aves, de sussubproductos, en fin, en todas aquellasáreas donde hay un veterinario trabajan-do, abriendo así lasposibilidades de nue-vos mercados, certificando exportacio-nes, en definitiva, eliminando barreras noarancelarias.
Respecto a la salud humana, hemos en-contrado en nuestra Revista muchísimotrabajo al respecto y si bien parece algotan obvio para el mundo, creemos quemerece la pena que los uruguayos lorecordemos una vez más, que tengamospresente el trabajo de los veterinarios enHidatidosis, en Toxoplasmosis, en Bru-celosis, en Leptospirosis, por citar sola-mente algunas enfermedades.Y cómo no encontrar en nuestra revisiónde Revistas anteriores, su número100, en ese momento bajo la responsa-bilidad del Dr. García Vidal. Y el conve-nio que nuestra SMVU, representada porel entonces Presidente Dr. García Lagos,y el Decano de nuestra Facultad deVeterinaria, Dr. MarcoPodestá, firmaron, haciendo posible unmuy fructífero intercambio con todo elmundo académico.También hemos recordado como se hanrealizado ya seis Congresos Nacionales.Todos ellos con el esfuerzo de muchos delos integrantes de nuestro gremio, quecada uno en su área de trabajo, ya sea enproducción, en reproducción, en tecno-logía de los alimentos, en animales decompañía, de zoológicos, y en un muylargo etcétera, dieron muchas horas de sutiempo para lograr el éxito que se alcan-zó en todos y en cada uno de ellos.Y por supuesto el trabajo que duranteaños aquellos veterinarios del áreade buiatría han realizado, esfuerzo quese ha concretado en las Jornadas de Buia-tría, ya van treinta, organizadas por elCentro Veterinario de Paysandú en for-ma ininterrumpida, correspondiendo ensiete oportunidades a Congresos o Jor-nadas Latinoamericanas. Lo que sin duda
avala un logro que nadie nos regaló, sinoque la profesión se ganó, y nos referi-mos a la nominación como país sede del XXI Congreso Mundial de Buiatria delAño 2000, desafío que no solo convocóa los colegas buiatras, sino a la profesióntoda y también al país.Sin duda no nos sería suficiente un edi-torial para ayudarnos a refrescar nuestramemoria colectiva, repasando todosaquellos logros y espacios de trabajo yde participación por los que hemos tran-sitado a lo largo de nuestra historia pro-fesional y gremial. Y reconociendo queno siempre han sido recordados ni valo-rados con justicia, aún por nosotros mis-mos, pero que han hecho a la propia ra-zón de ser de nuestra profesión e íntima-mente unida a ella, a la existencia denuestra «S.M.V.U.».Con la certeza de que en la vida sepuede actuar de dos maneras, «PORACCION O POR OMISION»; y luegode haber repasado tantas y tan fructífe-ras acciones desarrolladas por nuestraprofesión a lo largo de tantos años, invi-tamos a toda ella a emprender uncamino de ACCION, participativo yde trabajo, ya sea creando espa-cios o impulsando criterios en los quecreamos (de creer), mediante los quepodamos hacer nuestro aporte para me-jorar la calidad de vida de nuestra gente,de nuestras familias, de nosotros mis-mos...

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002) 7
Artículo Original
Recibido: 20/06/02 Aprobado: 22/07/02
7-14(2002)
Estudio de la estructura genética de la raza Holando Uruguayo mediantemarcadores genéticos. Comparación intraracial
Kelly, E.L.1 ; Mortari, N. 2 ;De Andrés, D. 3; Postiglioni, A. 1.
1Area Genética. Facultad de Veterinaria. Universidad de la República Oriental del Uruguay.Montevideo. A. Laspaces 1550. CP 11600., e-mail: [email protected].
2Universidad Federal de San Carlos. S.P. Brasil.3CSIC. España.
RESUMENEl estudio de la estructura genética de la raza Holando Urugua-yo (HU) se realiza en una muestra de 357 animales, correspon-diendo a 138 madres, 42 padres y sus 177 crías. Se testan 8sistemas de grupos sanguíneos (A, B, C, F, J, L, S, Z), obte-niéndose un menor número de alelos del sistema B en las crías.Para evaluar la variabilidad de nuestra población se calcula elíndice de heterocigosidad medio (IH). Se demuestra que ésta esmayor en las madres que en las crías. Se comparan las frecuen-cias genotípicas observadas en la generación filial con las fre-cuencias esperadas de acuerdo al cruzamiento al azar entre lospadres y las madres para los sistemas A, F, J, L y Z. Lossistemas J y Z presentaron un X2 significativo, siendo susposibles causas el efecto producido por el uso de pocos padrescon gran número de crías. Se calcula la distancia genética (DG)e identidad (genética) de Nei entre la muestra de HU y lapoblación de Holstein-Friesian (HF) de USA, por ser su prin-cipal ancestro. Esta se estima a partir de la frecuencia génica delos 7 sistemas, siendo la identidad promedio entre HU y HF deUSA muy alta (0,9807). En relación a los marcadores analiza-dos, se concluye que existiría una disminución de la variabili-dad genética en la siguiente generación y una gran similitudentre las poblaciones de HF de USA y Uruguay.
Palabras clave: Grupos sanguíneos, diversidad bovina, Holando Uruguayo.
INTRODUCCIÓNEl estudio de la estructura y variabilidad poblacional a travésde los marcadores genéticos, nos permite evaluar los efectosproducidos en las poblaciones por diferentes métodos de cría yla relación genética entre ellas. Los grupos sanguíneos, porposeer sistemas complejos con elevado grado de polimorfis-mo, se consideran marcadores genéticos apropiados para reali-zar este tipo de estudios (11, 20).Por otro lado es de gran importancia conservar la variabilidadgenética en las poblaciones de animales de producción ya quees un pre-requisito para el progreso de la selección y para susobrevivencia, pues si todos los individuos tuvieran los mis-mos genes, no existiría la capacidad de adaptarse a los cambiosambientales y se produciría la extinción de esa raza o población(15, 18). Entre las causas que disminuyen la variabilidad pro-
SUMMARYA total of 357 blood samples were collected from HolsteinFriesian cattle in Uruguay, distributed as follows: 138 cows,42 bulls and 177 calves. Blood groups were determined byusing 39 reagents from 8 systems (A, B, C, F, J, L, S, Z). Inorder to evaluate the degree of genetics variability in our sam-ple, the average Heterozygosity was calculated. On the otherhand, the average Heterozygosity was higher in cows than incalves, that together with a diminished number of alleles in Blocus for progeny, points out a fall in genetic variability in thenext generation. The significance of differences between theobtained and expected values of genotypic frequencies foundin the progeny for systems J and Z seem to be caused by alimited number of sires rather than by other factors effectingthe changes in gene frequencies. Nei´s measures of normalizedgenetic identity and standard genetic distance between twopopulations were calculated for populations from Uruguay andUSA over 7 loci. Those measures show that the HF populatio-ns from the USA and Uruguay are the closest ones. Summingup all the results, we conclude that there is a decrease in thegenetic variability of the Uruguayan HF progeny generationand a high similarity between HF populations from the USAand Uruguay.Keywords: Blood groups, diversity cattle, Holstein-Friesian-
Uruguay.
duciendo una erosión genética tenemos: número reducido deindividuos fundadores, consanguinidad y uso extensivo de lainseminación artificial. Por ejemplo la raza HF de USA cuyapoblación excede los 10 millones, tiene un tamaño efectivomuy bajo (< 1000), como resultado del uso sistemático de lainseminación artificial permitiendo que unos pocos machos dejendescendencia (7). Esta erosión genética limita la adaptación dela población a nuevas circunstancias y afecta el futuro de la críabovina al reducirse la base genética sobre la que se seleccionanlos fenotipos deseables en el futuro (www.ri.bbsrc.ac.uk/cdiv_www/inform.htm). Recientes estudios sobre la evoluciónde la variabilidad genética en diferentes razas en base a losmarcadores genéticos sanguíneos han demostrado los efectosproducidos por la intensa selección sobre la reducción de losniveles de la misma (14; 3; 10).

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002)8
en la bibliografía (9; 23). Para ello se es-tima la DG estándar y la identidad gené-tica (IG) normalizada (16) mediante elprograma informático descrito por Do-wling y Moore (6). La DG e IG se esti-ma a partir de las frecuencias génicas ycalcula la probabilidad de que dos alelostomados de cada población sean idénti-cos, si tienen alelos iguales su valor es 1y ninguno es 0 (2). Para un locus se cal-cula como:
IG=∑ai.bi/√∑ai2. ∑bi2
Siendo: ai la frecuencia de los alelos en la pobla-ción A y bi los de la población B.La DG estima el número de sustitucio-nes alélicas por locus y se calcula como:
DG= -log.n .I También se calculó el índice de hetero-cigosidad medio esperado que mide lavariabilidad de una población como:
IH= 1-∑p2
Siendo p la frecuencia de los diferentesalelos de un locus. Como el sistema B de grupos sanguí-neos fue analizado con diferentes núme-ros de reactivos en las poblaciones deUruguay y de USA, para realizar la com-paración de las frecuencias génicas fuenecesario igualar los fenogrupos, agru-pándose aquellos fenogrupos semejan-tes y sus frecuencias génicas en la po-blación de USA.
RESULTADOSEn el Cuadro 1 se detallan los sistemasde grupos sanguíneos del HU con susalelos y sus respectivas frecuencias gé-nicas. Se determinaron 39 fenogrupos di-ferentes en el sistema B encontrándose 4fenogrupos propios de nuestra pobla-ción. El test de homogeneidad entre lasmuestras de los 16 establecimientos, fueno significativo, reuniéndose por lo tan-to como una sola población.El test de equilibrio génico para el siste-ma F demostró ser levemente significati-vo (X2 =3.94; P<0.05) para las madres,con un coeficiente de Consanguinidadde f= 0.1797 (Cuadro 2).En la trasmisión de los alelos de la gene-ración parental a la filial se observó una
La raza Holando del Uruguay representael 90 % del ganado lechero de nuestropaís (DICOSE, 1988). Su origen data delaño 1889 con las primeras importacio-nes de animales de Holanda, pero reciéna principios de siglo comienza la crea-ción de la raza con el mayor ingreso deanimales provenientes de USA y Argen-tina. En el año 1971 con la importaciónde semen congelado de USA y Canadácomienza la expansión de la raza (5).El objetivo del presente trabajo es anali-zar la variabilidad genética del HolandoUruguayo a través de marcadores san-guíneos y su relación con otras pobla-ciones HF de diferentes países, especial-mente con su principal ancestro el HFde USA.
MATERIALES Y MÉTODOSSe estudia una muestra al azar de 357bovinos Holando de pedigree, constitui-da por 138 madres 42 padres (90% eransemen importado de USA y Canadá) ysus 177 crías. La misma pertenece a 16establecimientos de la cuenca lechera deMontevideo. Por lo cual se realiza untest de homogeneidad entre las muestrasantes de reunirlas como una población.A cada animal se le extrajo una muestrade sangre con anticoagulante y fue anali-zada según la técnica serológica de he-mólisis descrita por Stormont y col. paradeterminar 8 sistemas de grupos sanguí-neos (21,22). La tipificación se realizóincubando a 25°C una mezcla de 50 μl dereactivo de grupos sanguíneos, 25 μl deuna suspensión de glóbulos rojos lava-dos al 3% en solución fisiológica de cadaindividuo a tipificar y 25 μl de comple-mento de conejo fresco. La lectura de lareacción se registra a los 90 minutos y alas 3 hs. a partir del inicio de la incuba-ción. Los 37 reactivos de grupos sanguí-neos fueron aislados mediante inmuni-zaciones (12), siendo la batería la siguien-te: sistema A (A2); B (B1,G2,I1,I2,O1,Q,T1,Y2,P1,A,B,D,E2,G,I1,J,O2,Q,Y,G”);C (C1,C2, W, R1,X1,L´); F ( F1, V2); J(J); L (L); S (S, H¨,U1,U´,U”); Z (Z1).La estandarización de los reactivos fuerealizada a través del test de comparacióninternacional organizado por la Internatio-nal Society for Animal Genetics (ISAG),lo cual permitió realizar los estudios com-parativos con otras poblaciones de HF.
A partir de la lectura del test se determi-naron los grupos sanguíneos para cadaanimal o sea su hemotipo o genotipo san-guíneo.La caracterización racial se realizó me-diante la estimación de las frecuenciasgénicas de la población a partir de losgenotipos sanguíneos determinados. Di-chas frecuencias se calcularon por sepa-rado para madres, padres y crías con elfin de evitar la superposición de genera-ciones, ilustrar las diferencias entre ma-chos y hembras y conocer la evoluciónde la variabilidad de la raza Holando uru-guayo. La determinación de los fenogru-pos (alelos) de los sistemas complejos By S en los productos se realizaron poranálisis de segregación de factores en fa-milias de toros. La frecuencia génica deestos sistemas se calcularon por el mé-todo de alocación (17). Para el sistema Cno se le calcularon las frecuencias géni-cas debido a que en muchos casos no sepudo determinar los fenogrupos. El cál-culo de las frecuencias génicas del siste-ma codominante F se hizo por recuentode genes y la de los sistemas con domi-nancia completa (A, J, L, Z) con 2 alelosmediante la raíz cuadrada de la frecuen-cia relativa del genotipo homocigota re-cesivo. El sistema A se lo incluyó dentrode este grupo de sistemas por ser testa-do para un solo factor sanguíneo. Para elsistema F se realizó el test de equilibriogénico Hardy-Weinberg con el método deX2. Además, se evaluó por éste métodosi las frecuencias genotípicas observa-das en la generación filial se desviabansignificativamente de las esperadas porel cruzamiento de las generaciones pa-rentales.La variabilidad genética se estima me-diante el coeficiente de consanguinidady el índice de heterocigosidad medio es-perado (1). El coeficiente de consangui-nidad (f) (13) se calcula como:
f = H.esp-H.obs/ H.esp.
Donde:H representa la frecuencia relativa de losheterocigotas esperados y observadosrespectivamente.La comparación del HU con las pobla-ciones de HF de USA y la de España serealizó en base a los resultados descritos
7-14

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002) 9
SISTEMAS ALELOS FRECUENCIAS Padres Madres Crías
A A2 .225 .335 .226a .775 .665 .774
B b .0169 .0105 .0186BG2I1O1A’ .0122 .0 .0BG2KO’G» .0 .0036 .0BG2K(Y2)E’2O’ .0122 .0109 .0056BG2KA’O’ .0 .0036 .0BG2KA’I’O’G» .0 .0036 .0056BG2KY2A’I’O’G» .0122 .0 .0028BI1 .0 .0078 .0BO1 .0122 .0380 .0199BO1Y2 .0 .0266 .0166BO1B’ .0122 .0151 .0113BO1D’(Y2) .0122 .0109 .0028BO1G» .0 .0036 .0030BY2A’G’Q’G» .0 .0257 .0085G2I1 .0123 .0493 .0226G2O1Y2 .0 .0 .0028G2Y2D’ .0 .0036 .0028G2Y2E’2Q’ .2446 .2073 .2386I1 .0 .0049 .0028I2 .0366 .0726 .0706I2D’G’ .0 .0 .0028O1Y2G’G» .0244 .0073 .0169O1A’ .0244 .0620 .0452O1A’I’Q’ .0 .0036 .0028P .0 .0036 .0056Q .0 .0036 .0Y2A’ .0854 .0923 .0814Y2A’D’G’I’Q’ .0 .0036 .0028Y2A’Y’ .0 .0036 .0113A’ .0122 .0115 .0098D’G’O’ .1601 .0292 .0791D’G’O’G» .0 .0073 .0028G’G» .0366 .0074 .0313G’O’G» .0 .0073 .0
J’O’ .0610 .1273 .0932I’ .0 .0036 .0Q’ .0953 .0992 .1008G’ .0443 .0 .0057G” .0732 .0259 .0731
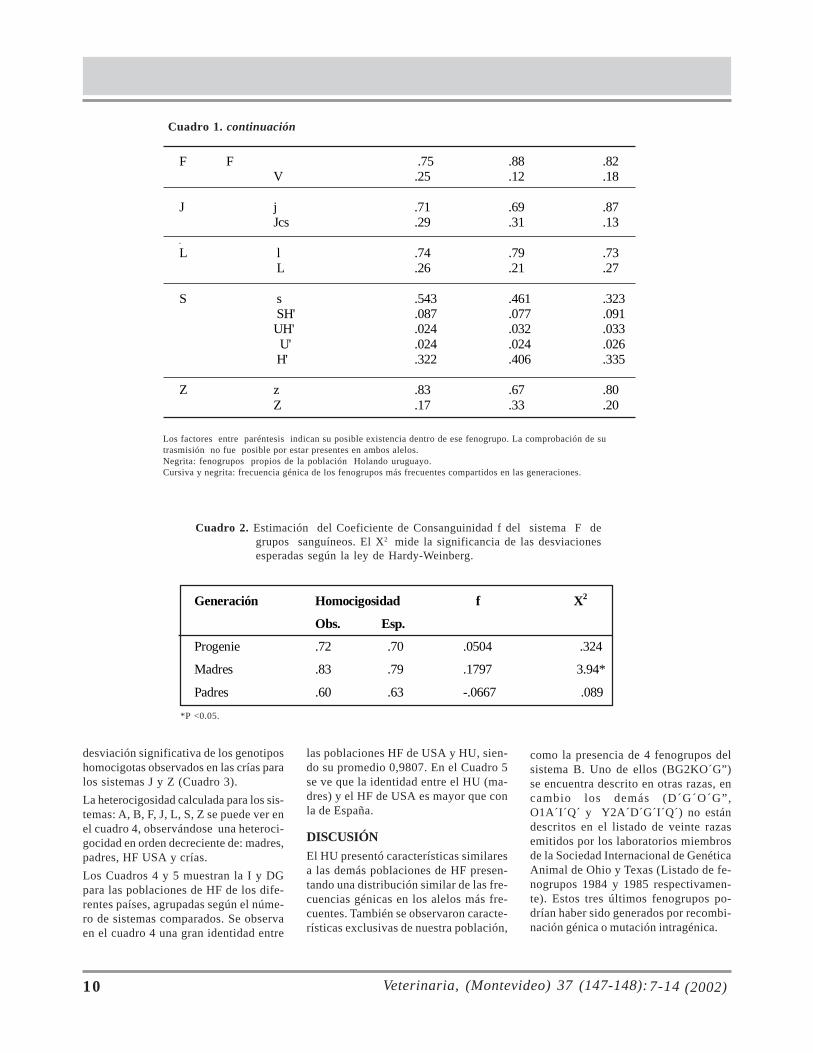
Cuadro 1. Frecuencia alélicas de los grupos sanguíneos de la raza Holando del Uruguay.
7-14
Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002)10 7-12 (2002)
F F .75 .88 .82 V .25 .12 .18 ____________________________________________________________________ J j .71 .69 .87 Jcs .29 .31 .13 _____________________________________________________________________ L l .74 .79 .73
L .26 .21 .27 _____________________________________________________________________ S s .543 .461 .323
SH' .087 .077 .091 UH' .024 .032 .033 U' .024 .024 .026 H' .322 .406 .335
______________________________________________________________________ Z z .83 .67 .80 Z .17 .33 .20
Cuadro 1. continuación
Los factores entre paréntesis indican su posible existencia dentro de ese fenogrupo. La comprobación de sutrasmisión no fue posible por estar presentes en ambos alelos.Negrita: fenogrupos propios de la población Holando uruguayo.Cursiva y negrita: frecuencia génica de los fenogrupos más frecuentes compartidos en las generaciones.
Cuadro 2. Estimación del Coeficiente de Consanguinidad f del sistema F degrupos sanguíneos. El X2 mide la significancia de las desviacionesesperadas según la ley de Hardy-Weinberg.
Generación Homocigosidad
Obs. Esp.
f X2
Progenie
Madres
Padres
.72 .70
.83 .79
.60 .63
.0504 .324
.1797 3.94*
-.0667 .089
*P <0.05.
desviación significativa de los genotiposhomocigotas observados en las crías paralos sistemas J y Z (Cuadro 3).La heterocigosidad calculada para los sis-temas: A, B, F, J, L, S, Z se puede ver enel cuadro 4, observándose una heteroci-gocidad en orden decreciente de: madres,padres, HF USA y crías.Los Cuadros 4 y 5 muestran la I y DGpara las poblaciones de HF de los dife-rentes países, agrupadas según el núme-ro de sistemas comparados. Se observaen el cuadro 4 una gran identidad entre
las poblaciones HF de USA y HU, sien-do su promedio 0,9807. En el Cuadro 5se ve que la identidad entre el HU (ma-dres) y el HF de USA es mayor que conla de España.
DISCUSIÓNEl HU presentó características similaresa las demás poblaciones de HF presen-tando una distribución similar de las fre-cuencias génicas en los alelos más fre-cuentes. También se observaron caracte-rísticas exclusivas de nuestra población,
como la presencia de 4 fenogrupos delsistema B. Uno de ellos (BG2KO´G”)se encuentra descrito en otras razas, encambio los demás (D´G´O´G”,O1A´I´Q´ y Y2A´D´G´I´Q´) no estándescritos en el listado de veinte razasemitidos por los laboratorios miembrosde la Sociedad Internacional de GenéticaAnimal de Ohio y Texas (Listado de fe-nogrupos 1984 y 1985 respectivamen-te). Estos tres últimos fenogrupos po-drían haber sido generados por recombi-nación génica o mutación intragénica.
7-14

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002) 11
Los resultados obtenidos indican unatendencia a conservar cierta variabilidadgenética, teniendo en cuenta el tamañode la muestra y la cantidad de fenogru-pos detectados en el sistema B (39 en138 individuos). Considérese a modo deejemplo que Bouw (4) detectó 42 alelosen una muestra de 1200 individuos deHF de Holanda y que Hines (9) observó82 en una muestra de 8630 HF de USAde los cuales 22 estaban presentes en unsolo animal y que por lo tanto es pocoprobable que estén presentes en nuestramuestra.Consideraremos ahora los 10 fenogruposmás comunes en el sistema B pues es undato que nos permite caracterizar la po-blación y conocer los cambios que pre-senta una raza (23). En el caso del HUtotalizan en promedio el 83%, existien-do un alelo con una alta frecuencia(G2Y2E´2Q´: 23%), siendo también elmás frecuente en las otras poblacionesde HF. El sistema B por lo tanto presen-ta un gran número de alelos, de los cua-
les sólo unos pocos presentan frecuen-cias altas (cuadro 1). Por otro lado siobservamos las diferentes generacionestenemos que los 10 alelos B más comu-nes en los productos suman el 84 % adiferencia de las madres que totalizan el80 %, por lo tanto existe una mayor va-riabilidad genética en las madres que enlas crías, ya que en éstas últimas aumen-tan las frecuencias de los alelos más co-munes, pero disminuyen el número dealelos de dicho sistema (34 en las madresy 32 en las crías: Cuadro 1). Al compararestos fenogrupos vemos que las críascomparten 9 de los 10 alelos más fre-cuentes con los padres y 8 con las ma-dres. Por lo tanto las crías se parecenmás a los padres que a las madres, locual se confirma al presentar las críasuna mayor identidad genética (0.9968) yla menor distancia de Nei (0.0032) conlos padres (Cuadro 4).Como conclusión podemos decir que lavariación de las frecuencias génicas y ladisminución de los alelos de una genera-
ción a otra puede ser debido al uso depocos padres que dejan un gran númerode crías, lo cual es típico de la situacióngeneral como vimos anteriormente conel HF de USA (7). Como consecuenciase producen cambios en las frecuenciasgénicas de las poblaciones con la perdidade polimorfismo. De acuerdo a estos re-sultados existiría una tendencia en el HUhacia la disminución de la variabilidadgenética en la siguiente generación, yaque las crías no solamente presentan unnúmero menor de alelos en el sistema B,sino que también hay una disminucióndel coeficiente de heterocigosidad (Cua-dro 4). Estos resultados también se hanobservado en estudios realizados en ra-zas Nórdicas (10). En ellas se evaluó loscambios de la variación genética en 13razas mediante la comparación de datospublicados en marcadores genéticos san-guíneos desde 1956 a 1975, donde hu-bieron pérdida de alelos que tenían bajafrecuencia. También según dichos estu-dios se esperaría una pérdida del 1 al 11%
Cuadro 3. Comparación de las frecuencias genotípicas observadas en lageneración filial con las frecuencias esperadas de acuerdo alcruzamiento al azar entre los padres y las madres.
Locus Genotipo Fr. obs. Fr. Esp. X2
A
F
J
L
Z
a/a
V/V
j/j
l/l
z/z
.59
.04
.75
.54
.64
.51
.03
.49
.58
.56
3.51
1.18
48.95*
1.30
4.74*
Cuadro 4. Identidad y distancia genética (I/D)* para 7 sistemas de grupos sanguíneos (A, B, F, J, L, S, Z) entrelas generaciones de HU y USA (9). En diagonal y negrita se encuentran los valores del Indice deHeterocigosidad promedio esperado y su error estándar.
Población Padres Madres Productos USA Padres .4739±.0788 .9850 .9968 .9752 Madres .0151 .4837±.0849 .9793 .9896 Productos .0032 .0209 .4454±.0872 .9773 USA .0251 .0105 .0229 .4699±.090
* Se expresa la identidad en el triángulo superior del cuadro y en el inferior la distancia.
7-14 (2002)

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002)12
de su heterocigosidad durante un perío-do de 20 a 40 años, de acuerdo al númeroefectivo actual de las razas nórdicas queva de 30 a 257. En el HF se vería agrava-da la situación por la disminución del N°efectivo de 1000 a 100 individuos de1996 (7) a 1999 (19) para una poblaciónde 10 millones de individuos para Nor-teamérica y Oeste Europeo.El equilibrio génico examinado en el sis-tema F mostró en las madres un aumentodel coeficiente de consanguinidad(f= 3.94; P< 0.05), al compararlo con lascrías y los padres. El valor de f positivoindica un déficit de heterocigotas, lo cualpuede ser causa de apareamientos endo-gámicos. Sin embargo considerando quese estimó a través de un locus simple yel alto coeficiente de heterocigosidad delas madres la consanguinidad no sería tanalto como lo indica el valor de f (Cua-dros 1 y 4).Las frecuencias genotípicas observadasen las crías al ser comparadas con lasesperadas (cuadro 3), nos muestra dife-rencias significativas en los sistemas J yZ, adjudicando estas diferencias a unaumento de los homocigotas recesivosobservados (j/j, z/z). Para estos siste-
mas, se analizó el porcentaje de hijos quedejan los padres homocigotos recesivos(sistema J, 20 de los 40 padres totales,es decir el 50%; el sistema Z 27 de los 40totales, esto es , el 67,%), que resultóser, respectivamente el 54,4% y el 65%de los hijos de la población. La pequeñadesviación en la contribución a la gene-ración siguiente, y su corrección no ex-plica el excesivo aumento de crías j/j.Considerando los padres cuyo genotipose ha podido determinar para éste siste-ma, vemos que la mayoría son heteroci-gotas por lo cual la frecuencia de recesi-vos sería superior a lo estimado. Unaposible causa de este resultado podríaser que esté actuando la selección a fa-vor de los recesivos en este sistema yaque según Hafez (8) el antígeno J delsuero estaría asociado con un aumentode la mortalidad y fallas en la fertiliza-ción por causas de incompatibilidad in-munológica.La IG entre la población de USA y HUtiene un alto valor promedio de 0,9807(0.9896 a 0.9751: Cuadro 4) lo que indi-ca que entre ambas poblaciones existeun alto grado de semejanza genética quecorrespondería a poblaciones sin aisla-
miento reproductivo (2). La poblaciónde España ocupa un lugar más alejado(Cuadro 5), reflejando probablemente lamayor influencia en ella de las FrisonasEuropeas. Por lo tanto se observa unagran similitud entre las poblaciones HFcomparadas, lo que nos indica una granuniformidad genética dentro de la raza.Con respecto a este tema debemos con-siderar los efectos producidos por la in-trogresión de genes HF en otras pobla-ciones como en el ganado negro y blancoeuropeo, en el cual si bien han mejoradola producción lechera también han teni-do un efecto desfavorable sobre la ferti-lidad (14).Si consideramos que nuestra raza Ho-lando se ha formado a partir de cruza-mientos absorbentes con la poblaciónH.F de USA y que en momento actualexiste una gran identidad entre ambas(cercana a 1) se debería hacer una estra-tegia para conservar los genes nativos delHU producto de su adaptación al medioy evitar la erosión genética observada ennuestra raza en el presente trabajo comoconsecuencia de una pérdida de la varia-bilidad genética de una generación a otra.
Población Uruguay USA España Uruguay .3719±.0457 .9972 .9783 USA .0028 .3421±.0377 .9878 España .0219 .0122 .3471±.0535
Cuadro 5. Identidad y distancia genética para 5 sistemas (A, F, J, L, Z), entre Uruguay, USA (9) yEspaña (23). En diagonal IH y su error estándar.
CONCLUSIONESLos resultados obtenidos del análisis deuna muestra de HU mediante grupos san-guíneos, nos permite concluir que:1) Presentan características de la raza HF
(frecuencias génicas similares) y ex-clusivas de nuestra población, debidoa la presencia de 4 fenogrupos del sis-tema B.
2) Existiría una tendencia en el HU haciala disminución de la variabilidad ge-nética en la siguiente generación, yaque las crías presentan un número me-
nor de alelos en el sistema B y unadisminución del coeficiente de hete-rocigosidad.
3) Existiría cierto grado de consaguini-dad debido a un coeficiente f positi-vo para el sistema F en las madres yun desvio significativo hacia los ge-notipos homocigotas en la progeniede los sistemas J y Z.
4) Se observa una gran similitud entrelas poblaciones HF comparadas, es-pecialmente entre HU y HF de USA,lo que nos indica una gran uniformi-dad genética dentro de la raza.
AgradecimientosEn el presente trabajo colaboraron va-rias personas e instituciones gracias a lascuales fue posible la realización del mis-mo. Al PEDECIBA y el ICI que finan-ciaron las pasantías en los Laboratoriosde Inmunogenética de la UniversidadFederal de San Carlos (Brasil) y en laDepartamento de Genética y Zootecniade la Facultad de Veterinaria de la Uni-versidad de Córdoba (España). A la Fa-cultad de Veterinaria, especialmente a losBr. José Payque y Luis Roses por suasistencia en el área de informática.
7-14

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002) 13
Referencias Bibliográficas1. Ayala, F.J. (1982). Population and
Evolutionary genetics. California. Ed.Cumnings Publishing Company.
2. Ayala F.J.; Kiger J.K. (1984).Modern genetics. Ed.2ª. California.Ed. Cumnings Publishing Company.
3. Blott, S.C.; Williams J.L.; HaleyC.S. (1998). Genetic variation withinthe Hereford breed of cattle. Animal.Genetics 29: 202-211.
4. Bouw, J. (1960). The geticalcomposition of the Dutch cattlebreeds as determined by thefrequencies of blood groups. Z.Tierzucht. Zucht. Biol. 74: 248-266.
5. Caorsi, C. (1973). HolandoUruguayo. Historia y presente deuna raza. Soc. Criadores Holando delUruguay.p 39.
6. Dowling ,T.E.; Moore W.S. (1984).A program for estimating geneticvariabil i ty within and betweenpopulations. Journal of Heredity.75: 416.
7. Georges, M.; Andersson, L. (1996).Livestock Genomics Comes of Age.Genome Research.6:907-921.
8. Hafez E.S.E. (1996). Reproducción einseminación artificial en animales,6a Ed. México, Nueva EditorialInteramericana, S.A.542p.
9. Hines, H.C.; Haenlein, G.F.W.;Zikakis, J.P.; Dickey, H.C. (1977).Blood Antigen, Serum Protein, andMilk Protein Gene sequencies andGenetic Interrelationships inHolstein Cattle. Jour. Dairy Sci.60(7):1143.1151.
10.Kantanen, J.; Olsaker, I .;Adalsteinsson, S; Sandberg, K.;Eythorsdottir, E.; Pirhonen, K.;Holm L-E. (1999). Temporal changesin genetic variation of NorthEuropean cattle breeds. AnimalGenetics.30: 16-27.
11. Kelly, L. (1988). Grupos sanguíneosBovinos. Veterinaria. N° 99. 17-20.
12. Kelly, L. (1988). Obtención de losprimeros reactivos de grupossanguíneos de Bovinos en el Uruguay.Jornadas Científico Técnicas deProducción Animal.1988.
13. Kidd, K.K.; Stone, W.H.;Crimella, C.; Carenzi, C.; Casati,M.; Rognoni, G. (1980).Inmunogenetic and populationgenetic analyses of Iberian cattle.Anim. Blood Grps. biochem. 11:21-38.
14. Lidauer, M.; Mantysaari, E.(1996). Genetic constitution of theFinnish black and white catt lepopulation and the influence ofHosteinization on protein yield, days openand somatic cell count. Acta AgriculturaeScandinavica. 46: 193-200.
15. Mac Gregor, H. (1995). DNA andall that. Equinet. The equine geneticsand evolution research informationNetwork.
16. Nei, M.(1972). Genetic distancebetween populations. TheAmer.Naturalist.106:283-292.
17. Neimann-Sorensen, A.(1956).Blood groups and breed structure asexemplified by three Danish breeds.Acta Agr. Scand. Acta Agr. Scand.6:115.
18. Rendel,J. (1967). Studies of BloodGroups and Protein Variants as aMeans of reveling similarities anddifferences between animalpopulations. ABA Vol.35, N°3:371-383.
19.Riquet, J.; Coppieters, W.;Cambisano, N.; Arranz, J.J.;Berzi, Davis, S.K.; Grisart, B.;Farnir, F.; Karim, L.; Mni, M.;Simon, P.; Taylor J.F.;Vanmanshoven, P.; Wagenaar, D.;Womack, J.E.; Georges M. (1999).Fine-mapping of quantitative traitloci by identity by descent in outbredpopulations: Application to milkproduction in dairy cattle. Proc. Natl.Acad. Sci. USA.Vol.96:9252-9257.
20. Rodero, A.; Garzón, R.; Llanes, D.;Zarazaga, I.; Vallejo, M.; Monje,E. (1982). Genetic distances betweenspanish sheep breeds. Archivos deZootecnia, 31: 97-108.
21. Stormont C. (1981). The B and Csystems of cattle revisited. Frontiersin Inmunogenetics. Ed W.H.Hildemann. New York p 31-43.
22. Stormont,C., Owen, R.D., Irwin,M.R. (1951). The B and C systemsof bovine blood groups. Genetics.36:134.
23. Vallejo, V.M. (1978). Razas vacunasautóctonas en vías de extinción.Aportes al estudio genético. SerieUniversitaria 69. Fundación JuanMarch.
7-14

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002) 15
INTRODUCCIÓNEl Herpesvirus bovino tipo-1 (HVB-1),pertenece a la familia Herpesviridae (43,48), subfamilia alphaherpesvirinae, gé-nero varicellovirus (6). Puede causar Ri-notraqueitis Infecciosa Bovina (IBR),Vulvovaginitis Pustular Infecciosa (IPV),Balanopostit is Pustular Infecciosa(IPB), conjuntivitis, abortos, infeccióngeneralizada en el neonato y meningoen-cefalitis (20, 43, 48). Es una enfermedadcontagiosa de distribución mundial (43).El análisis de HVB-1, basado en el pa-trón de digestión por enzimas de restric-ción, permite identificar 3 subtipos(HVB1.1; HVB1.2a; HVB1.2b). La en-zima HindIII, corta el ADN y da patro-
nes de bandas que diferencian los subti-pos de HVB-1 (12, 25, 28). Los anti-cuerpos monoclonales pueden ser utili-zados también, para reconocer subtiposde HVB-1 (26).Los subtipos HVB-1.1 y HVB-1.2a seasocian a la forma respiratoria de la en-fermedad (IBR) y clínicamente puedencursar con síntomas respiratorios, con-juntivitis, signos clínicos nerviosos, in-fertilidad y abortos (46). El subtipoHVB-1.1 puede causar abortos 30 a 60días post signos respiratorios (10, 11).Si bien es una enfermedad en la cual, nose ha observado una alta mortalidad, 3%en ganado lechero, su importancia radicaen las pérdidas económicas debidas a
abortos, pérdidas de neonatos, pérdidadel estado general, y caída de la produc-ción lechera, dando lugar en algunos ca-sos a complicaciones por infeccionesbacterianas secundarias (27). El HVB-1.2b, esta asociado a enfermedad genital(IPV/IBP) y no causa abortos (11, 45).Otro HVB que se disemina rápidamenteal SNC y que fue clasificada como tipo5, ocasiona meningoencefalitis severa, fueaislada en Australia, Argentina, Brazil yUSA (14).El genoma de HVB-1 esta compuesto por136 kilobases (Kb) y posee al menos 10genes con potencial para codificar glico-proteínas. Por su localización en la su-perficie del virión, estas glicoproteínas
Artículo Original
Caracterización de un herpesvirus, aislado de un ternero, con signosnerviosos y sin respuesta inmune humoral específica
Alonzo, P.1; Benavides, U. 1; Isnardi, F.1; Puentes, R.1; Carol, H.1; Clavijo, A.1; del Campo, R.2;Bonnevaux, J.3; Weiblen, R.4; Fondevila, N.5; Romera, S.A.5; Sadir, A.M.5; Maisonnave, J.1
RESUMENSe caracteriza como Herpesvirus bovino (HVB)1.1, un aisla-miento viral, realizado de un ternero de nueve meses de edadcon signos neurológicos, corrimiento ocular y nasal unilateral.Muestras de hisopados nasal, ocular y prepucial, se inocularonen cultivos de la línea celular de riñón bovino, Madin DarbyBovine Kidney. Los inóculos de los tres primeros días postinicio de signos clínicos, produjeron efecto citopático caracte-rístico de HVB. El aislamiento (Uy-1999) fue identificado comoherpesvirus por inmunofluorescencia directa y caracterizadomediante enzimas de restricción, reacción en cadena de polime-rasa e inmunohistoquímica. Aunque la cepa Uy-1999 es inmu-nogénica para otros bovinos, el animal del cual se aisló el HVB,nunca produjo anticuerpos específicos anti-HVB, pero sí con-tra otros patógenos, como el virus de diarrea viral bovina. Losresultados obtenidos hasta el momento, sugieren que la falta derespuesta inmune humoral no es debida a una variación antigé-nica del aislamiento. Los métodos diagnósticos en que se basanlas campañas de control, deberían ser re-evaluados, si la exis-tencia de bovinos latentemente infectados con HVB-1, y sinanticuerpos específicos detectables, no es un hecho aislado.
Palabras clave: herpesvirus bovino, aislamiento,caracterización
SUMMARYAn herpesvirus, isolated from a nine month old calf, with neu-rological clinical signs, unilateral ocular and nasal discharge,was characterized as Bovine Herpes Virus-1.1 (BHV-1.1). Na-sal, ocular and prepucial swabs, were collected and superna-tants were inoculated in Madin Darby Bovine Kidney cells.Only the nasal and ocular inoculates from the first three daysafter clinical signs appeared, produced herpesvirus characte-ristic citopathic effect. The isolate (Uy-1999) was identifiedas herpesvirus by direct immunofluorescence, characterizedby restriction enzyme genomic profile, polymerase chain reac-tion and immunohistochemistry. Even though the isolate Uy-1999 is immunogenic for other bovines, the animal from whichthe isolation was made, never produced BHV specific antibo-dies, while it did to other pathogens as bovine viral diarrheavirus. The results obtained up to now, suggest that the lack ofhumoral immune response, is not due to an antigenic variationof the isolate. If the existence of BHV-1 latently infected bovi-nes, is not an isolated issue, the tests used in the control cam-paigns should be reviewed.
Keywords: bovine herpesvirus, field isolate, characterization
1Facultad de Veterinaria, Dpto.de Ciencias Microbiológicas, Area Inmunología, Lasplaces 1550, Monteviddeo, C.P. 11600, Uruguay. Tel: (598) (2) 6281303,Fax: (598) (2) 6280130, E-mail: [email protected]
2Instituto de Investigaciones Biólogicas "Clemente Estable", 3Transgenes, 4Universidad Federal de Santa María, R.S. Brasil, 5INTA Castelar, Argentina.
Recibido: 19/08/02 Aprobado: 09/12/02
15-22

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002)16
son importantes blancos del sistema in-mune del hospedador y tienen un rolfundamental en la patogenicidad, pormediar la entrada, fusión y dispersióndel virus célula-célula. Las más estudia-das por la importancia en estos eventosson gB, gC y gD, pero actualmente otrasglicoproteínas, enzimas y proteínas re-guladoras, son incluidas en el repertoriodel HVB (3, 41). Luego de la infección primaria el HVBse hace latente en ganglios nerviosos dela puerta de entrada, (19, 30, 33) y pue-de ser re-activado y re-excretado cuandoel portador se encuentra bajo condicio-nes de estrés o mediante inmunodepre-sión con corticoides (34, 47). La latenciase define como una infección persistenteen el organismo, donde el genoma viralestá presente en ausencia de virus infec-cioso (20). El HVB-1 reactivado esusualmente excretado por la vía de in-fección primaria, dado que ésta determi-na el lugar de latencia. Pero hay veces enque la infección primaria intra nasal vaseguida de una infección generalizada, encuyo caso se puede instalar latencia enotros ganglios nerviosos y el virus pue-de ser re-excretado por semen (45).Cuando hay re-activación viral, la inmu-nidad humoral específica, ejerce controlde la viremia, evitando la re-excresión ydiseminación del virus (20). Los anti-cuerpos (Acs) normalmente son detec-tables 7 a 10 días post-infección. Losisotipos de inmunoglobulinas M (IgM)y G (IgG), llegan a su título máximo alos 14 y 35 días post-infección respecti-vamente (17). Sin embargo la infeccióncon HVB-1 vía genital, puede inducirrespuesta inmune humoral débil, o pue-de no inducir respuesta inmune detecta-ble (45). La diseminación del HVB-1 a lamayoría de los animales de un rodeo esel resultado de una infección respirato-ria, la diseminación es limitada en casosde infección genital. Durante la infecciónprimaria aguda el virus se excreta en se-creciones nasales durante un período de4 a 17 días, los títulos más altos son 4 a6 días post infección. Animales que ex-cretan el virus por la mucosa nasal lotransmiten por contacto directo y porgotitas en aerosol a una distancia corta(8 metros). Animales que excretan HVB-1.2b (virus de vulvo vaginitis pustularinfecciosa y balanopostitis) por vagina
o prepucio, lo transmiten menos eficien-temente, y un menor número de anima-les se infecta. La dosis infectante nece-saria para infectar un animal ha sido pocodocumentada y dependerá de la virulen-cia de la cepa (46).El HVB ha sido detectado en Uruguay,clínica y epidemiológicamente desde1970. Fue aislado por primera vez en1981, de un animal seropositivo, inmu-nodeprimido con corticoides (16). En1987, se describió un caso de granulomanasal bovino en ganado Jersey, con altamorbilidad, oportunidad en que se logróel aislamiento de HVB-1 de hisopadosnasales (31). Se ha descrito que en el 70%de los tumores de ojo se aisla HVB-1(42). En Uruguay existe un 75-100% deestablecimientos con presencia de anti-cuerpos a HVB y la prevalencia a nivelnacional es de 45% en ganado de carne y48% en ganado de leche (38).Con respecto al control de la enferme-dad, se han descrito varios métodos, se-gún la prevalencia de infección. En luga-res de muy baja prevalencia se puedeemprender la erradicación, eliminando losanimales seropositivos. Donde la preva-lencia es alta, esta medida es poco viabley la vacunación es la indicada para esti-mular la inmunidad específica e impedirla re-excresión viral. La vacunación noimpide la infección pero si la sintomato-logía (20). En un rodeo con un alto por-centaje de animales seropositivos, lashembras adultas multíparas muy proba-blemente ya hayan sido infectadas y susistema inmune evitará que aborte unasegunda vez a causa de HVB-1. Por lotanto, teniendo en cuenta el costo-bene-ficio de vacunar para evitar síntomas,abortos por ejemplo, sería convenientevacunar únicamente las vaquillonas an-tes del primer entore (Dr. E. Dubovi,comunicación personal).Las vacunas utilizadas en Uruguay sonmayormente fabricadas con cepas de HVBde referencia. Si los aislamientos realiza-dos en el país son caracterizados, habríaposibilidad de incluir en las vacunas cepasnacionales. Así como tener un stock decepas actuantes en el país y la región.La inmunotolerancia no se ha descritopara el HVB, pero sí para otros viruscomo el de Diarrea Viral Bovina (BVDV).Embriones bovinos infectados con virus
no citopático de BVDV in útero, antesque su sistema inmune este desarrolla-do, reconocen como propio al virus, yno producen anticuerpos específicos, yquedan persistentemente infectados (PI).Estos terneros son la fuente de infecciónen un rodeo y por lo general tienen me-nor desarrollo corporal y pelo hirsuto(2, 9).El propósito del presente trabajo es iden-tificar y caracterizar una cepa de HVBaislada a partir de un ternero de 9 mesesde edad, con sintomatología nerviosa ymenor desarrollo corporal que los bovi-nos de la misma categoría. Este ternerose recuperó rápidamente a pesar de quenunca produjo anticuerpos anti-HVB ypasó a ser un toro adulto saludable y conaptitudes reproductivas normales.
MATERIAL Y MÉTODOS
1. Aislamiento viralSe obtienen hisopados oculares, nasalesy prepuciales durante 7 días post iniciode signos clínicos (21 muestras), de unternero Limousin con nueve meses deedad, menor desarrollo corporal que losbovinos de su misma categoría, pelo hir-suto, corrimiento ocular y nasal serosounilateral, y ataxia. Los hisopados, seinocularon en cultivos de células de ri-ñón bovino, línea celular Madin DarbyBovine Kidney (MDBK), mantenidos enmedio Eagle modificado (MEM), 50 g/mL de gentamicina y 10% de suero fetalbovino (GIBCO) irradiado. A los 4 díaspost-inoculación, el cultivo que no mues-tra efecto citopático (CPE), se congela,descongela y centrifuga a 400g por 15minutos, para luego inocular el sobrena-dante en una monocapa de células vir-gen. Este procedimiento se repite 3 ve-ces (3 pases ciegos), para dar como ne-gativa una muestra (7).
2. Identificación y caracterizaciónviral
a. Identificación viral con anticuer-pos específicos. Las células con CPE ca-racterístico de herpesvirus, fueron fijadascon acetona e incubadas 1 hora a 37ºC conanticuerpo policlonal anti-HVB conjuga-do con isotiocianato de fluoresceina FITC(VMRD, USA), y observadas al micros-copio de luz ultravioleta (UV) (7, 24).
15-22

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002) 17
Para cuantificación mediante citofluoro-metría (40), las células infectadas quemuestran CPE y los controles negativosse resuspendieron por tratamiento contripsina y se incubaron con el mismoconjugado policlonal FICT por 2 hs. a4ºC en PBS 0.1% azida, lavadas por cen-trifugación dos veces y analizadas me-diante un citómetro de flujo (FAC Scan,Becton Dickinson). Se definieron regio-nes en cuadrantes, los límites fueron fi-jados en 197 para el valor de luz disper-sada a 180 grados (Forward SCatteredHeigth=FSC-H) y 189 para el valor deintensidad de FICT (FL-1). Los even-tos en la región superior derecha fueronconsiderados positivos y se registraronlos porcentajes de células positivas en10.000 eventos (40). Para ambas técni-cas, se utilizan, como control positivocélulas infectadas con HVB-1 de refe-rencia cepa Los Angeles (LA), y comocontrol negativo células sin inocular.b. Inmunohistoquímica. Células infec-tadas con el aislamiento (Uy-1999), seincuban 1 hora a 37ºC con anticuerposmonoclonales (Mab) anti-HVB-1.1(Mab60), y anti-HVB-5 (Mab2915).Luego de tres lavados se agrega conjuga-do a peroxidasa anti-inmunoglobulina deratón, se incuba 1 hora a 37ºC, se lava yse revela con sustrato Di-amino bencidi-na (DAB) (26).c. Estudios moleculares. La técnica dereacción en cadena de la polimerasa(PCR) Multiplex (1), se utilizó para di-ferenciar ADN de HVB-1 y HVB-5 enuna sola reacción. La extracción de ADNse realizó mezclando, 250 μL de sobre-nadante de células infectadas con: Uy-1999, HVB-1.1 (LA), y HVB-5 (663),con igual volumen de solución tampónde extracción (Tris10mM, disodiumEDTA 1mM, 0,5% SDS, Proteinasa K1,6U/mL, pH 8). La mezcla se incuba atemperatura ambiente durante 5 minu-tos y se adicionan 500 μL de (25 fenol:24 Cloroformo: 1 alcohol isoamilico).Luego de centrifugar a 16.000g por 2minutos, el sobrenadante se somete a otraextracción con cloroformo-alcoholisoamilico (24:1) y centrifuga nuevamen-te. La precipitación del ADN se realizacon 1 mL de etanol absoluto, y 100 μLde NaCl 3M pH 5,2, centrifugando a16.000g por 30 minutos. El pellet se lavacon 1 mL de etanol 70%, se seca y resus-
pende en 10μL de agua bi-destilada, al-macenadose a -20ºC hasta su uso. En cadaextracción, se utilizan como controlespositivos cepas de referencia de HVB-1.1 y HVB-5 y sobrenadante de célulassin inocular como control negativo.El ADN y los primers (TK1, TK2, GD1,GD2) se incubaron a 99ºC (comienzocaliente) por 10 minutos y luego a 88ºCcuando los demás reactivos se adiciona-ron: (buffer Taq polimerasa, glicerol,Cl2Mg, dNTP, Taq polimerasa). Se rea-lizaron 35 ciclos de 1 minuto a 95ºC (des-naturalización), 1 minuto a 61ºC (hibri-dización) y 1 minuto a 72ºC (síntesis),finalmente 5 minutos a 72ºC. El produc-to amplificado se analizó por electrofo-resis en gel de agarosa horizontal 1,8 %(100V, 30 minutos) en solución tampóncon bromuro de etidio y visualizado enun transiluminador de UV.Para la caracterización genómica del vi-rus, se realizó análisis del ADN viral conenzimas de restricción (HindIII) (25, 28).Cuando células infectadas con Uy-1999y cepas de referencia HVB1.1 y HVB-5mostraban 50% de CPE, se lisaron con0,2 mL de Sodium Dodecyl Sulfate (SDS)al 10%. El ADN celular fue precipitadocon 0,5 mL de NaCl 5M, y el pellet sedescartó luego de la centrifugación. ElADN viral presente en el sobrenadante,fue incubado con proteinasa K 20μg/mL,durante 2 horas a 37ºC. Las proteínasfueron removidas mediante extraccióncon fenol:cloroformo y eter etílico satu-rado en agua, y el ADN precipitado conisopropanol durante 1 hora a -70ºC. Elpellet de ADN viral fue resuspendido en50μl de Tris-HCl 10mM, pH 8, EDTA1mM (TE), y el corte con HindIII serealiza, de acuerdo a las instrucciones delfabricante (Biolabs). Los fragmentosobtenidos, se separan por electroforesisen geles de agarosa 0,7%, en solucióntampón Tris-Acetato-EDTA (TAE), avoltaje (30V) constante durante 5 horas.Se tiñen con bromuro de etidio y las ban-das se visualizan con un transiluminadorde UV (25, 28).d. Identificación de la glicoproteínaC (gC) en la cepa Uy-1999.Las proteínas virales se separan en gelesde poliacrilamida al 12% (SDS-PAGE),y se transfieren a una membrana immu-nolon P (Millipore). El bloqueó se reali-
za con una solución 5% leche e soja enPBS-tween 20, y se lava 3 veces conPBS-Tween 0,05% agitando. Se incubadurante 1 hora a 37ºC con anticuerpomonoclonal anti-gC (Dr. L. Babiuk,VIDO, Saskatoon, Canada), se lava 3veces, y se incuba en agitación durante 1hora a 37ºC con conjugado a peroxidasaanti-ratón. Luego de 3 lavados se revelacon el kit Renaissance, según indicacio-nes del fabricante (NEN Life Science).(3, 18).
3. Respuesta Inmune Humorala. Serología. Se obtienen muestras desuero del ternero Limousin, durante losprimeros 7 días post inicio de signos clíni-cos, cada 15 días durante 5 meses y cada 6meses hasta la actualidad. Las muestras seestudian para detectar anticuerpos (Acs)específicos anti-HVB, mediante ELISA(HIPRA-España). La presencia de anti-cuerpos neutralizantes se determina, me-diante la técnica de seroneutralización invitro (SN), en placas de 96 hoyos. Lasmuestras de suero se incuban 1 hora a 37ºCcon 100 unidades infectantes de la cepaLA título 106,7 dosis infectante de cultivode tejido 50% (DICT50) /50 μL. Luego seagregan 20.000 células/hoyo y se incuban72 horas a 37ºC y 5% CO2. El título seexpresa como el logaritmo de la inversa dela última dilución que protege el desafío de100 DICT50.(29).La presencia de anticuerpos anti-virus deDiarrea Viral Bovina (BVDV) se determi-na mediante SN, utilizando para desafío100 UI de la cepa de referencia Singer (tí-tulo 105,5 DICT50/50 μL) (32) y ELISA(HIPRA-España). Anticuerpos anti-virusde Leucosis Bovina (BLV) fueron evalua-dos mediante inmunodifusión en gel de agar(AGID).La técnica de microaislamiento viral (39)se utiliza para detectar virus de BVDV enlas muestras de suero antedichas.Se realiza un muestreo serológico estratifi-cado por categorías, del 10% del rodeo alcual pertenecía el ternero Limousin(n=130). Muestreandose 28 ternero/as 9 a12 meses de edad, 15 bovinos entre 1 y 1año y medio, 32 de 1 año y medio a 3 añosy 55 bovinos adultos. Los sueros se pro-cesan para la detección de anticuerpos neu-tralizantes de HVB, por la técnica de SNin vitro (29).
15-22

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002)18
b. Estudios de inmunogenicidad dela cepa Uy-1999. Se formula una vacu-na inactivada utilizando Uy-1999 (35,36) con adyuvante formulado por INTA(Arlacel C, Markol 52 y Tween 80) (37).Se inmunizaron por vía intramusculardos bovinos Holstein de 10 meses deedad (seronegativos para HVB-1 por SNy ELISA). Muestras de suero para de-tectar anticuerpos anti-HVB fueron ob-tenidas 17 y 25 días post vacunación(dpv). Los anticuerpos totales se cuan-tifican mediante el test de ELISA, y seexpresan como el logaritmo de la inversade la máxima dilución del suero, cuya den-sidad óptica (DO) es igual o mayor al40% de la DO del control positivo (35).Los anticuerpos neutralizantes se titula-ron por SN (29).
RESULTADOSSe logró el aislamiento de un HVB a par-tir de hisopados nasales y oculares obte-nidos en los tres primeros días post ini-cio de signos clínicos (Cuadro 1). Loscultivos de células MDBK inoculadoscon estas muestras, presentaron CPE ca-racterístico de HVB (células redondea-das en racimos).El virus fue identificado como HVB, ob-servandose fluorescencia específica in-tracitoplasmática de las células inocu-ladas con el aislamiento Uy-1999 (Fig.1). Los estudios de citofluorometría con-firman este resultado, dado que célulasinoculadas con Uy-1999 y con cepa dereferencia HVB-1 (LA), presentaron 26%y 29% de eventos positivos respectiva-
mente, mientras que células no inocula-das presentaron menos de 2,5%.La técnica de PCR Multiplex amplificaun segmento de 183 pares de bases (pb)del ADN Uy-1999, al igual que en la cepade referencia HVB-1.1 (LA) (Fig. 2).Luego del corte con la enzima HindIII, elADN Uy-1999 presenta el siguientepatrón de bandas: 6 bandas entre 23.1 y9.4 kb, 3 bandas entre 9.4 y 6.5 kb y 2bandas alrededor de 4.4 kb, que corres-ponde al patrón de HVB-1.1.Células MDBK infectadas con Uy-1999,e incubadas con el monoclonal Mab60anti-HVB-1.1, dan coloración positiva,comparado con la no coloración al utilizarMab2915, específico de HVB-5 (fig. 3).El Western blot muestra la presencia dela glicoproteína C de envoltura, en el ais-lamiento Uy-1999 (Fig. 4).Nunca se detectaron anticuerpos anti-HVB en el ternero del cuál se aisló lacepa Uy-1999, ni por la técnica de sero-neutralización ni por ELISA.La prevalencia de anticuerpos anti-HVB,en el rodeo al que pertenece el animal delcual se aisló HVB, es de 8.5%. El cuadro2 muestra la prevalencia de anticuerposanti-HVB por categoría.Las muestras resultaron negativas a BLV,sin embargo se pudo verificar la sero-conversión específica a BVDV, 7 mesespost inicio de signos clínicos. No se ais-ló BVDV en las muestras de suero postcomienzo de signos clínicos.La cepa Uy-1999 mostró ser inmunogé-nica, al detectarse anticuerpos totales yneutralizantes específicos anti-HVB, enlos bovinos inmunizados con la vacunainactivada. Los títulos de anticuerpos
Hisopos
Días
(post inicio de síntomas)
Nasales
Oculares
Prepuciales
1 + + -
2 + + -
3 + + -
4 - - -
5 - - -
6 - - -
7 - - -
Cuadro 1. Resultados de aislamiento en cultivos celulares de la líneacelular MDBK
15-22

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002) 19
Figura 2. PCR Multiplex: Carril 1: controlpositivo = HVB-1 de referencia (LA), Carril2: control negativo = HVB-5 de referencia(663), Carril 3 y 4: aislamiento Uy-1999.
Referencias: Calle 1: Control positivo HVB-1. Calle 2: Control positivo HVB-5
Calle 3-4: Uy 1999.
Figura 3. Inmunohistoquímica en células MDBK infectadas con aislamiento Uy-1999: 3ª = Anticuerpo monoclonal (Mab) anti-BHV-1.1 (Mab 60), 3b =Mab anti-BHV-5 (Mab2915), 400X.
Figura 4. Detección de glicoproteína C por Westernblot . Carri l 1: Uy-1999,. Carri l 2:Marcador de peso molecular.
totales obtenidos a los 17 dpv fueron de3,40, y a los 25 dpv en uno de los bovi-nos aumentó a 4,01, permaneciendo cons-tante en el otro bovino vacunado (Gráfi-ca 1). Los títulos de anticuerpos neutra-lizantes obtenidos a los 17 dpv fueronde 1 y 1,3, y a los 25 dpv aumentaron a1,9 y 2,2 respectivamente (Gráfica 2).
DISCUSIÓNPrevio a la caracterización se pensó queel aislamiento era un HVB-5, dado que lameningoencefalitis por infección con her-pesvirus, usualmente esta asociada a estetipo viral (8, 12, 15). Sin embargo se con-firmó que el aislamiento corresponde altipo HVB-1.1, que también puede pro-vocar alteraciones nerviosas (4, 13, 23).Las características de menor peso cor-poral que los bovinos de su edad y pelohirsuto, del ternero Limousin, nos llevóa pensar que podría tratarse de un bovi-no persistentemente infectado (PI) conBVDV. La ausencia de virus en las mues-tras de suero post inicio de signos clíni-cos, y la aparición de anticuerpos espe-cíficos anti-BVDV siete meses más tar-de, descartan esta hipótesis.
1 2
0
0,5
1
1,5
2
2,5
0 17 25
Tiempo en días
Títu
lo S
.N
bovino 1 bovino 2
Gráfica 1. Evolución del título de anticuerpos totales(medido por ELISA) en bovinosvacunados con la cepa Uy-1999.
00,5
11,5
22,5
33,5
44,5
0 17 25
Tiem po en días
Títu
lo E
LISA
bovino 1 bovino 2
Gráfica 2. Evolución del tí-tulo de anticuerpos seroneu-tralizantes en bovinos vacu-nados con la cepa Uy-1999.
15-22

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002)20
La presencia del HVB en el estableci-miento se confirma al encontrar anima-les seropositivos. La prevalencia seroló-gica encontrada en el rodeo al que perte-necía el ternero del cuál se aisló HVB, esmenor a la reportada en el país (38), y elhecho de que solo un animal mostró sig-nos clínicos en el rodeo, podrían sugerirque la cepa viral Uy-1999 es de baja vi-rulencia. Se podría plantear, en este caso,que existen diferencias antigénicas entrela cepa Uy-1999 y la de referencia. Perolos resultados obtenidos de la inmuno-genicidad y la identificación de la glico-proteína C en la cepa Uy.1999, nos llevaa rechazar esta hipótesis. Sin embargo,la confirmación debe ser realizada conestudios moleculares más precisos.La inmunogenicidad de la cepa aislada serealizó en bovinos de distinta raza, debi-do a que no existe literatura que especi-fique diferencias entre razas en sensibi-lidad a la infección por HVB.Nunca se detectó respuesta inmune hu-moral anti-HVB en el ternero del cual seaisló Uy-1999, lo que llevaría a pensaren un animal con dificultad en la respues-ta a determinados patógenos. Sin embar-go, la presencia de respuesta inmune hu-moral especifica anti-BVDV y la recu-peración post infección con HVB, des-cartan la hipótesis de inmunodeficien-cia.La posibilidad de una infección leve opor vía venérea, que no desencadene unarespuesta inmune humoral (45) en el ter-nero Limousin, se descarta, por la apari-ción de signos clínicos, y la convivenciacon bovinos del mismo sexo y edad, almomento del aislamiento. Además, comola cepa Uy-1999 se aisló de hisopadosnasal y ocular, es más factible, que lainfección haya sido vía aerógena y la la-tencia este establecida en ganglios satéli-tes de la puerta de entrada (19). Si la
viremia fue suficiente para establecer la-tencia en ganglios alejados de la puertade entrada, el virus se podría llegar a ex-cretar por semen también (45).Se detectó, por PCR, ADN de HVB enmuestras de liquido seminal del terneroLimousin post inmunodepresión concorticoides (34). Si bien no se logró re-aislar Uy-1999, se confirma latencia vi-ral en ganglios alejados de la puerta deentrada. Estos resultados no se presen-tan en la sección correspondiente por noser un objetivo del presente trabajo.Otra hipótesis posible, es que el animalse infectó cuando aún tenía anticuerposcalostrales, neutralizando el virus y evi-tando signos clínicos, pero no la infec-ción y latencia (5, 21, 22). Los signosclínicos observados en el ternero Li-mousin, en esta hipótesis, corresponde-rian a una reactivación del virus, cuandolos anticuerpos calostrales ya no exis-ten. Por lo tanto, el animal debería haberproducido anticuerpos propios anti-HVB, post signos clínicos. El no haberdetectado inmunidad humoral anti-HVB,y la edad del ternero al momento del ais-lamiento (nueve meses de edad), descar-tan esta hipótesis.En estudios futuros, será evaluada la res-puesta inmune celular (RIC), mediantela técnica de linfoproliferación in vitro.Si no existe RIC específica anti-HVB enel bovino Limousin, se podría plantearotra hipotesis, la respuesta inmune in-nata, fue, en este caso, suficiente paracontrolar la infección viral y lograr la re-cuperación clínica.La posibilidad de que el animal se infec-tara in útero, antes de que su sistemainmune estuviese maduro, y establecieratolerancia a HVB, no ha sido descrito.Sin embargo, la tolerancia inducida porinfección viral temprana se da en otrosvirus como el de BVDV. Para confirmar
o descartar esta posibilidad, son necesa-rios estudios de infección experimentalen distintas etapas de la gestación.Las campañas de control o erradicaciónde la enfermedad, se basan en pruebasque detectan anticuerpos específicosanti-HVB. Por lo tanto, la existencia debovinos latentemente infectados conHVB, y sin anticuerpos detectables, im-plicaría sin duda, un riesgo epidemioló-gico, para la diseminación del virus y laenfermedad. Por lo tanto si se confirmala existencia de estos bovinos, habría quere-evaluar las pruebas en que se basanlas campañas de control y se deberíanutilizar pruebas mas sensibles como laPCR (44).
CONCLUSIONESSe caracterizó el aislamiento Uy-1999como un HVB1.1, y se comprobó su ca-pacidad inmunogénica, al responder bo-vinos inoculados con la vacuna elabora-da con la cepa aislada.Con los resultados obtenidos hasta elmomento, concluimos que la falta de res-puesta inmune humoral en el bovino delcuál se aisló Uy-1999, no es debida avariaciones en la cepa aislada.La cepa aislada y caracterizada se ex-pandió y almacenó a -80ºC y nitrógenolíquido, para poder ser utilizada en laelaboración de vacunas con cepas nacio-nales.
AgradecimientosOsvaldo Zabal y Teresa Moral, Labora-torio de virología, INTA Castelar, Ar-gentina, por consejos en técnicas de cul-tivo celular.Juan Cristina y Heber Espino, Centrode Investigaciones Nucleares, por traba-jos de irradiación de SFB.
15-22

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002) 21
Referencias Bibliográficas1. Alegre, M.; Nanni, M.; Fondevila,
N. (2001). Development of aMultiplex Polymerase ChainReaction for the Differentiation ofBovine Herpesvirus-1 and 5. J. Vet.Med. B 48, 613-621
2. Baker, J.C. (1987). Bovine viraldiarrhea virus: a review.J.Am.Vet.Med.Assoc. 190:1449-1458.
3.Baranowski, E.; Dubuisson, J.;Pastoret, P.P.; Thiry, E. (1993).Identification of 108k, 93K and 42kglycoproteins of bovine herpesvirus-1 by monoclonal antibodies.Arch.Virol. 133 (1-2): 97-111.
4. Barenfus, M.; Delli Quadri, C.A.;McIntyre, R.W.; Schroeder, R.J.(1963). Isolation of infectious bovinerhinotracheitis virus from calves withmeningoencephalitis. J. Am.Vet.Med. Assoc. 143: 725-728.
5. Bradshaw, B.J.F.; Edwards,S . (1996). Antibody isotyperesponses to experimental infectionwith bovine herpesvirus 1 in calveswith calostrally derived antibody.Vet. Microbiol. 53 (1-2): 143-151.
6. Brown, F. (1989). The classificationand nomenclature of viruses.International Committee onTaxonomy of viruses in Edmonton,Canada, 1987. Intervirology 30: 181-186.
7. Carbey, E.A. (1971). RecomendedStandard laboratory techniques fordiagnosing infectious bovinerhinotracheit is , bovine virusdiarrhea, and shipping fever(Parainfluenza3) Proc. 75th Annu.Meet. U.S. Animal health Assoc.,pp629-648.
8. Carril lo, B.J.; Pospischil , A.;Dahme, E. (1983). Pathology of abovine viral necrotizing encephalitisin Argentina. Zentralb Vetinarmed(B) 30: 161-168.
9. Cutlip, R.C.; McClurkin, A.W.;Coria, M.F. (1980). Lesions inclinically healthy cattle persistentllyinfected with the virus of bovine viraldiarrhea- glomerulonephritis andencephalitis.J.Am Vet.Res.,41:1938-1941.
10. d´Offay, J.M.; Mock, R.E.; Fulton,R.W. (1993). Isolation andcharacterization of encephaliticbovine herpesvirus type 1 isolatesfrom cattle in North America. Am. J.Vet. Res. 54 (4): 534-539.
11. Edwards, S.; White, H.; Nixon, P.(1990). A study of the predominantgenotypes of bovine herpesvirus 1,found in the U. K. Vet. Microbiol.22 (2-3): 213-223.
12. Engels, M.; Steck, F.; Wiler, R.(1981). Comparison of the genomesof infectious bovine rhinotracheitisand infectious pustularvulvovaginit is virus strains byrestriction endonuclease analysis.Arch. Virol. 67 (2):169-174.
13. Engels, M.; Guiliani, C.; Wild, P.;Beck, T.M.; Loeple, E.; Wiler, R.(1986). The genome of bovineherpesvirus 1 (BHV-1) strains,exhibit ing a neuropathogenicpotential compared to known BHV-1 strains by restriction site mappingand cross-hybridization. Virus Res.6: 57-73.
14. Flores, E.F.; Silva, A.M.; Weiblen,R. (1998). Neuropatogenicidade doHerpesvirus Bovino Tipo 5 (HVB-5). Simposio Internacional sobreHerpesvirus Bovino (tipo 1 e 5) eVirus da Diarreia Viral Bovina(BVDV). Santa Maria, RS, Brasil.127-137.
15. French, E.L. (1962). A specific virusencephalitis in calves: isolation andcharacterization of the causal agent.Aust. Vet. J. 38: 216-221.
16. Guarino, H.; Maisonnave, J.;Capano, F.; Pereira, J. (1982).Primer aislamiento e identificacióndel virus de la Rinotraqueít isInfecciosa Bovina en Uruguay.Veterinaria (Montevideo) 78: 131-134.
17. Guy, J.S.; Potgieter, L.N.D. (1985).Bovine herpesvirus-1 infection ofcatt le: Kinetics of antibodyformation after intranasal exposureand abortion induced by the virus.Am.J.Vet.Res. 46 (4): 893-898.
18. Herring, A.J.; Sharp, J.M. (1984).Protein blotting: the basic method andits role in viral diagnosis. In:
McNulty, M.S. & McFerran, J.B.(eds.). Recent advances in virusdiagnosis. Martinus Nijhoff, TheHague, pp 115-124.
19. Karhs, R.F. (1987). InfectiousBovine Rhinotracheitis: A Reviewand Update. J.Am.Vet.Med.Assoc.171: 1055-1064.
20.Lemaire, M.; Pastoret, P.; Thiry,E.P. (1994). Le controle de l´infectionpar le virus de la rhinotrachéiteinfectieuse bovine. Ann. Méd. Vét.138: 167-180.
21.Lemaire M. V.; Weynants, J.;Godfrold, F.; Chynts, G.; Meyer,J.J.; Letesson and E. Thiry. (2000).Effects of bovine herpes virus tipo 1infection in calves with maternalantibodies on immune response andvirus latency. J. Clin. Microbiol.38:1885-1894.
22.Lemaire M.; G. Meyer; E.Baranowski; F. Schynts.;Wellemans G.; Kerkohfs P.; andThiry. E. (2000). Production ofbovine herpesvirus type 1-seronegative latente carriers byadministrations of a live-attenuatedvaccine in passively immunizedcalves. J Clin. Microbiol. 38 (11):4233-4238.
23.Ludwig,H. (1983). Bovineherpesviruses. In: Roizman B, (ed.)The Herpesviruses. Vol .2. New YorkPlenum Press, pp. 135-214.
24.Lupton, H.W.; Barns, H.J.; Reed,D.E. (1980). Evaluation of the rabbitas laboratory model for InfectiousBovine Rhinotracheit is Virusinfection. Cornell Vet. 70:77-95.0
25.Mayfield, J.E.; Good, P.J.; van Oort,H.J.; Campbell , A.R.; Reed,D.E.(1983). Cloning and cleavage sitemapping of DNA from bovineherpesvirus 1 (Cooper strain). J.Virol. 47 (1): 259-264.
26.Metzler, A.E.; Matilde, H.;Gassmann, U.; Engels, M.; Wyler,R. (1985). European isolates ofbovine herpesvirus-1: a comparisonof restriction endonuclease sites,polypeptides, and reactivity withmonoclonal antibodies. Arch.Virol.85: 57-69.
15-22

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002)22
27. Miller, J. 1991. Special Symposium.The multiple manifestation of IBRinfection. Vet.Med. 84-98.
28.Misra, V.; Babiuk, L.A.; Darcel,C.L. (1983). Analysis of bovineherpes virus type 1 isolates byrestrict ion endonucleasefingerprinting. Arch. Virol. 76 (4):341-354.
29.Msolla, P.M.; Wiseman, A.;Selman, I.E. (1981)- The prevalenceof serum neutralizing antibodies toinfectiours bovine rhinotracheitis virusin Scotland. J. Hyg. 86: 209-215.
30. Pastoret, P.P.; Thiry, E.; Brocher,B.; Derboven, G. (1982). BovidHerpesvirus 1 Infection of Cattle:Pathogenesis, Latency,Consequences of Latency. Ann.Rech. Vet. 13: 221-235.
31. Rivero, R.; Haedo, F.; Feola, R.;Capano, F.; Guarino, H.; Saizar,J.; Bermúdez, J. (1987). GranulomaNasal Bovino: descripción de un casocolectivo y discusión sobre suprobable etiología. Veterinaria(Montevideo) 98:5-11.
32. Robison, D.S.; Gillespie, J.H.;Baker, J.A. (1960). Theneutralization test as an indicator ofimmunity to Bovine viral diarrheavirus . Cornell Vet., 50:503-509.
33. Rock, D.L. (1994). Latent infectionwith bovine herpesvirus type 1.Seminars in Virology. 5: 233-240.
34. Rock, D.L.; Lokensgard, J.; Lewis,T.; Kutish, G. (1992).Characterization of dexamethasone-induced reactivation of latent bovineherpesvirus 1. J. Virol., 66 (4):2484-2490.
35. Romera, S.; Zamorano, P.I.; Alcón,V.L.; Puntel, M.; Ferrari, P.N.;Borca, M.V.; Sadir, A.M. (1999).Estrategia de inmunización enbovinos: empleo de RN-205 paraincrementar la respuesta humoral ycelular contra herpesvirus bovino.1.Therios 28 (149) : 216-225.
36. Romera, S.; Hilgers, L.; Puntel,M.; Zamorano, P.; Alcón, V.; DusSantos, M.; Blancovierra, J.;Borca, M.V.; Sadir A, M. (2000).Adjuvant effects of sulfolipo-cyclodextrin in a squalane-in wateremulsión and water-in-mineral oilemulsions for BHV-1 vaccines incattle. Vaccine.19:132-141.
37.Sadir, M.A.; Zamorano, P.I.;Romera, A.; Wigdorovitz, A.;Smitsaart, E.; Marangunich, L.;Schiappacassi, C.; Borca, M.V.(1999). Improvement of the immuneresponse to foot and mouth diseasevirus vaccine in calves by usingAvridine as adjuvant. Vet. Immunol.and Immunopathol. 69: 11-22.
38. Saizar, J. (1997). Determinación dela prevalencia de la RinotraqueítisBovina Infecciosa en rodeos de lechey carne en Uruguay. Veterinaria(Montevideo). 33.133-136.
39.Saliki, J.T.; Fulton, R.W.; Hull,S.R.; Dubovi, E.J. (1997).Microti ter Virus Isolation andEnzyme Immunoassays forDetection of Bovine Viral DiarrheaVirus in Cattle Serum. J. ClinicalMicrobiology, 35 (4) : 803-807.
40. Sharrow, S.; Segal, D. (1995).Immunofluorescence and cell sorting,in Current Protocols in Immunology.Coligan, J.; Kruisbeck, A.; Margulies,D.; Shevach, E. and Strober, W. Eds.John Wiley and Sons, New York, Unit5.1-5.6.
41. Schwyzer, M.; and Ackerman, M.(1996). Molecular virology ofrumiant herpesviruses. VeterinaryMicrobiology 53:17-29.
42.Spadrow, B.; Hoffmann, D. (1980).Bovine Ocular Squamous CellsCarcinoma. Veterinary Bulletin50(6):449-459.
43. Tikoo, S.K.; Campos, M.; Babiuk,L.A. (1995). Bovine herpesvirus 1(BHV-1): biology, pathogenesis andcontrol. Ad. Virus Res. 45: 191-223.
44. van Engelenburg, F.A.C.; vanSchie, F.W.; Fijsewijk, F.A.M.; vanOirschot, J.T. (1993). Excretion ofBovine Herpesvirus 1 in semen isdetected much longer by PCR thanby virus isolation. J. of ClinicalMicrobiology. 33(2):308-312.
45. van Oirschot, J.T. (1995). BovineHerpesvirus 1 in semen of bulls andrisk of transmision: a brief review. TheVeterinary Quaterly.17(1): 29-33.
46. Wentink, GH.; Van Oirschot JT.;Verhoeff J. (1993). Risk of infectionwith Bovine Herpes virus 1 (BHV1):A review. The Veterinary Quaterly15(1):30-33
47. Whetstone, C.A.; Miller, J.M.;Seal, B.S.; Bello, L.J.; Lawrence,W.C. (1992). Latency andreactivation of a thymidine kinase-negative bovine herpesvirus 1deletion mutant. Arch. Virol., 107(1-2): 27-34.
48. Wyler, R.; Engels, M.; Schwyzer,M. (1989). Infectious bovinerhinotracheitis/vulvovaginitis (BHV-1). In: Wittmann, G. (Ed.).Herpesvirus Disease of Cattle, Horseand Pigs. Developments inVeterinary Virology. Kluwer AcademicPublishers, Boston, pp. 1-72.
15-22

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002) 2323-26
INTRODUCCIÓNDebido al aumento en la utilización delconejo como mascota y como animal deconsumo, hay unmayor interés en el conocimiento de todolo relacionado con esta especie.El Intestino grueso tienen una confor-mación muy diferente a la de los demásherbívoros domésticos. Además, el co-nejo no ha sido incluido en la NominaAnatomica Veterinaria (NAV) (7) y porlo tanto no existe un acuerdo internacio-nal sobre la nomenclatura anatómica paraesta especie. Por este motivo, diferentesautores han usado nombres diferentespara estructuras similares. En el caso delintestino y en particular para el ciego y
colon ascendente, los autores citados eneste trabajo utilizan nomenclatura dife-rente y confusa. El objetivo de este tra-bajo es proponer una división de las par-tes del ciego y colon ascendente del co-nejo con una nomenclatura clara, instruc-tiva y con valor descriptivo, de acuerdoa los principios establecidos por la Aso-ciación Mundial de Anatomistas Veteri-narios (7).
MATERIALES Y MÉTODOSSe estudiaron 18 conejos adultos, 10hembras y 8 machos. Las razas usadasfueron: californiano: 6 hembras y 2 ma-chos; neocelandés: 4 hembras y 2 ma-chos; chinchilla: 2 machos y cruza deneocelandés x californiano: 2 machos. La
edad de los animales osciló entre 1 y 2años y el peso entre 6 y 7 kilos. Fueronsacrificados por sobredosis de anestési-co, tiopental sódico i/v, tras la venoclisisde una vena auricular. Diez animales seestudiaron mediante disección en frescoy los otros fueron previamente fijadoscon solución de formol al 10 % por víaintra-arterial. La técnica de fijación con-sistió en canular en sentido caudal la ar-teria aorta a nivel de tórax por un abor-daje intercostal izquierdo e inyectar lasolución de formol al 10 %.El abordaje del abdomen se hizo por unaincisión paramediana; desde el apéndicexifoides hasta el pubis, luego se proce-dió a realizar una incisión sobre cada arco
Divisiones del ciego y colon ascendente del conejo (Oryctolagus cuniculus)
Comunicación Corta
Möller, R.1; Pérez, W.1; Martin, E.1
RESUMENActualmente en el Uruguay, el conejo se está popularizandocomo mascota y también como animal de consumo, cosa que seevidencia en los comercios de venta de mascotas y en las carni-cerías. Esto implica un mayor interés en el conocimiento detodo lo relacionado con esta especie.La anatomía del conejo no es bien conocida y no existe unacuerdo sobre la nomenclatura para esta especie. El objetivo deeste trabajo es proponer una división del ciego y colon de acuerdocon los principios establecidos en la Nomina Anatomica Vete-rinaria, designar cada parte con un nombre corto y simple faci-litando su recuerdo; que tenga valor instructivo y descriptivo.Se estudiaron 18 conejos adultos. Algunos se disecaron en fres-co y otros fijados en formol al 10 %. En el ciego, enrollado enuna vuelta y media, se describe una base, un cuerpo con trespartes y el apéndice vermiforme. En el colon ascendente sedescriben también tres partes: un asa proximal, un asa interme-dia con dos partes (mayor y menor) y un asa distal unida almesoduodeno y mesenterio. Se concluye con una lista de nom-bres al estilo de la Nómina Anatómica Veterinaria.
Palabras clave: anatomía, conejos, ciego, colon ascendente,intestino.
SUMMARYAt present the rabbit is becoming more popular as a pet and asspecial dish. As can be see at pet shops and butcheries. There-fore there is more interest in everything related to the species.The anatomy of the rabbit is not well known and there is noagreement in the nomenclature.The objective of the present work is to propose division of thecaecum and colon according to the “Nomina Anatómica Veteri-naria”, to name each part short and simple, easy to rememberso it is instructive and descriptive.Eighteen adult rabbits were studied. Some were dissected freshand others fixed in 10 % formaline. The caecum is haze in oneand a half loop; it has a base, a body with 3 parts and thevermiform appendix.The ascendent colon also has 3 parts: a proximal loop, an inter-mediate loop with two parts (major and minor) and a distalloop attached to the mesoduodenum and mesentery.A list as names in the Nomina Anatomica Veterinaria style isproposed.
Keywords: anatomy, rabbits, caecum, ascendent colon,intestine.
Recibido: 27/05/02 Aprobado: 28/10/021Facultad de Veterinaria. Universidad de la República Oriental del Uruguay. Montevideo.A. Laspaces 1550. CP 11600., e-mail: [email protected].

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002)24
costal hacia dorsal, para tener una am-plia vista de los órganos abdominales.Una vez expuestos los órganos, se pro-cedió a la observación de cada una de laspartes del intestino grueso comenzandodesde el ciego a nivel de la desembocadu-ra del íleon.Las referencias usadas para los esque-mas y las fotos son:a, apéndice vermiforme del ciego; b, basedel ciego; c, parte descendente del cuer-po del ciego; c‘, parte ascendente delcuerpo del ciego; c“, parte transversa delcuerpo del ciego; d, duodeno; e, estóma-go; h, hígado; i, ileon; j, asa proximal delcolon ascendente; k, asa intermedia -parte mayor del colon ascendente; l, asaintermedia - parte menor del colon as-cendente; m, asa distal del colon ascen-dente; n, colon transverso; o, colon des-cendente; y, yeyuno-ileon.
RESULTADOSEn todos los animales se observó la mis-ma disposición del ciego y colon ascen-dente.El ciego, relativamente muy largo al com-pararlo con otros mamíferos domésticos,se encontró siempre enrollado en unavuelta y media. Esta conformación per-mitió dividirlo en tres partes principa-les. Una parte inicial, la base (Basis ceci)(b, Esquema 1), seguida de un cuerpo(Corpus ceci) (Figura 1 y 2 y Esquema1) y una parte terminal más delgada, elapéndice vermiforme (Appendix vermi-formis ceci) (a; Esquema 1 y Figura 1 y2). En el cuerpo del ciego también se ob-servaron tres partes evidentes: una par-te descendente ubicada a la derecha delplano medio (pars descendens corporisceci) y extendida hacia caudal (c, Esque-ma 1 y Figura 1), una parte ascendente
(pars ascendens corporis ceci) extendi-da de caudal a craneal ubicada a la iz-quierda del plano medio (c‘, Esquema 1y Figura1) y una parte transversal (parstransversa corporis ceci), que describíauna curva de izquierda a derecha (c“, es-quema 1), de transición entre el cuerpo yel apéndice vermiforme.El colon ascendente se encontró tambiéncon tres partes o asas fácilmente recono-cibles (Esquema 2). La primera, a partirdel ciego y unida a él en toda su exten-sión presentaba saculaciones en su parteexterna. Ésta podría llamarse asa proxi-mal (Ansa proximalis coli) (j; esquema),siendo su limite a la altura del borde libredel pliegue cecocólico que era lo que lasostenía. La segunda, estaba situada casitoda a la izquierda del plano mediano ydorsalmente. Esta segunda parte o asaintermedia (Ansa intermedia coli) des-cribió, en todos los casos, 2 flexuras con
Esquema 1. Ciego. Vista central.
Figura 2. Vista derecha de los órganos digestivos.
Figura 1. Vista izquierda de los órganos digestivos.
14-17 23-26

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002) 25
vértice hacia caudal. La primera flexura(k, Esquema 2 y Figura 1) fue siempre lamás grande, sujeta por un amplio meso-colon, con su superficie externa con sa-culaciones (Haustrae) al igual que el asaproximal. La segunda flexura del asa in-termedia, tenía una superficie lisa, esta-ba adherida a la raíz del mesenterio en unanimal y en los restantes estaba sosteni-da por un muy corto mesocolon (l, Es-quema 2 y Figura 1). En base a las dife-rencias de extensión y conformación,cada uno de estos tramos del asa inter-media, es decir, las flexuras, podrían lla-marse parte mayor (Pars magna, ansaeintermedialis coli) y parte menor (Parsparva, ansae intermedialis coli) respec-tivamente (k y l en Esquema 2 y Figura1). El último tramo del colon ascenden-te, el asa distal (Ansa distalis coli) (m;Figura 2), estaba a la derecha del planomediano y dorsalmente a las otras dosasas cólicas. Esta asa, estaba siempre fi-jada a la raíz del mesenterio y almesoduodeno y se extendía desde el asa
Esquema 2. Colon ascendente. Vistaventral.
Nota: para dar mejor entendimiento dela continuidad de las partes del colonascendente, no se han respetado lasproporciones y a cada parte se le ha dadoun relleno diferente.
La tesis de Heldsdörfer (6) es la que ofre-ce la descripción más detallada y es laque mejor concuerda con nuestra des-cripción. Este autor propone dividir elcolon ascendente en porción inicial y asasprimera, segunda y tercera. La primera ysegunda asa con sus vértices hacia cau-dal. La concordancia de la parte inicial ytercera asa con las asas proximal y distalde nuestra propuesta es total. Las asasprimera y segunda corresponderían a laspartes mayor y menor del asa interme-dia de éste trabajo. La ventaja de nuestrapropuesta, creemos que radica en que seajusta a los principios generales de no-menclatura anatómica acordados por laAsociación Mundial de Anatomistas Ve-terinarios y que resulta, por lo tanto,descriptiva, fácil de recordar y adecuadapara el estudio funcional de sus partes.
CONCLUSIONESEl siguiente listado al estilo de la Nómi-na Anatómica Veterinaria concluye estapropuesta de división y nomenclaturadel ciego y colon del conejo:
CecumBasis ceciCorpus ceci
Pars descendensPars ascendensPars transversa
Appendix vermiformis ceciColon ascendens
Ansa proximalis coliAnsa intermedia coli
Pars magnaPars parva
Ansa distalis
AgradecimientosAgradecemos al Área de Animales deGranja y Pilíferos de la Facultad de Ve-terinaria por cedernos las instalacionesy especialmente a la Dra. Beatriz Cava-llero.
intermedia hasta la flexura cólica dere-cha del colon transverso, describiendo enun animal una doble curvatura.
DISCUSIÓNLa descripción del ciego está basada enel trabajo de Helsdörfer (6). Sin embar-go, hemos adaptado la misma al criteriomás moderno y acorde a la NAV (7) em-pleado por Barone (1,2,3). Según esteúltimo, la base del ciego es parte del co-lon ascendente, al igual que en el caballo(2), luego describe un cuerpo pero sinasignarle partes. Nosotros proponemospor lo tanto incorporar a la nomenclatu-ra del ciego, la base y el cuerpo con trespartes: descendente, ascendente y trans-versa. Si bien se incorpora el término“base” (a semejanza del caballo) no con-sideramos aplicar el término de “ápex”.Esto es debido a que no se ajusta a lascaracterísticas generales del ciego de lasdemás especies domésticas, por la pre-sencia del apéndice vermiforme en el ex-tremo del cuerpo.Las descripciones encontradas en la bi-bliografía consultada sobre el colon as-cendente del conejo, son poco explíci-tas. Bensley (4) se limita a describirlecinco ramas o tramos separados porflexuras, sin especificar la topografía yconformación de cada parte. Barone(1,2,3), en cambio, simplifica la descrip-ción a sólo dos tramos. El primero, alque llama asa espiral, está asociado alciego e ileon, lo que corresponde a lo quenosotros proponemos llamar asa proxi-mal. Al segundo tramo le describe dosamplias inflexiones en S. Este segundotramo sería comparable a las asas inter-media y distal que describimos, aunqueno es posible establecer una correspon-dencia con las inflexiones descritas poreste autor.El trabajo publicado por Bonfert (5) ofre-ce una descripción del colon ascendentealgo más detallada; al igual que Barone(1,2,3) lo divide en dos partes, pero a lasegunda parte la subdivide en tres asas.Las dos primeras asas de Bonfert (5)corresponden a las dos partes, mayor ymenor, del asa intermedia descrita pornosotros, mientras que la tercera asa con-cuerda con el asa distal de nuestra pro-puesta.
14-1723-26

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002) 27
INTRODUCCIÓNEn Uruguay, existe muy poca informa-ción científica nacional referente a loshelmintos que parasitan a los equinos(1). Desde que la economía del país de-pende en un altísimo porcentaje de laexportación de productos y subproduc-tos derivados de la explotación de ovi-nos y bovinos, es racionalmente a estasespecies que se les asigna prioridad altiempo de invertir esfuerzos de investi-gación. Sin embargo, el equino continúasiendo imprescindible en las tareas rura-les. Además, varias actividades deporti-vas de naturaleza hípica como las carre-ras de caballos, el polo, los raids, la equi-tación, etc., tienen en el país una tradi-ción de larga data y la obtención de equi-nos que puedan desarrollar su máximopotencial competitivo requiere que es-tos hayan estado sometidos desde sunacimiento a un programa sanitario óp-timo el cual tiene como uno de sus pila-res fundamentales la sanidad parasitaria.Existe además una actividad de exporta-ción, fundamentalmente de carne equinaprocesada. En este rubro Uruguay ex-porta anualmente aproximadamente 40millones de dólares de acuerdo a datosde INAC.De acuerdo a las investigaciones realiza-das en otros países el equino es la espe-
cie doméstica que puede albergar la ma-yor cantidad de géneros y especies denematodos parásitos en su tubo digesti-vo(10). La gran mayoría de ellos perte-necen a la familia Strongylidae y dentrode esta se agrupan en dos subfamiliasStrongylinae y Cyathostominae cuyasformas adultas se encuentran distribuí-das a lo largo del intestino grueso. Den-tro de la subfamilia Strongylinae se handescrito cuatro géneros que son Stron-gylus, Triodontophorus, Oesophago-dontus y Craterostomum. Las especiesdel género Strongylus han sido llamadasprincipalmente por autores sajones,como los “grandes estrongilidos” mien-tras que las especie pertenecientes a losotros tres géneros se les conoce comolos “medianos estrongilidos” (17). Den-tro de la subfamilia Cyathostominae queagrupa a los “pequeños estrongilidos” sehan descrito en el mundo 13 géneros con51 especies (11).Desde siempre se ha reconocido funda-mentalmente la importancia patógena delgénero Strongylus debido a que sus esta-dos larvarios realizan migraciones pordiversos órganos internos, causandolesiones que pueden originar cuadros clí-nicos desde leves a muy graves o inclusola muerte del hospedero mientras que susestados adultos, provistos de una gran
cápsula bucal, injurian ampliamente lamucosa intestinal y sustraen a su hospe-dero considerable volumen sanguíneo(17).Menor importancia, desde el punto devista patógeno, se le atribuído a los de-más géneros de la subfamilia Strongili-nae y a los géneros que integran la subfa-milia Cyathostominae principalmenteporque sus estados adultos tienen unmenor tamaño relativo, con cápsulas bu-cales menos desarrolladas en el caso delos estrongylinos o de forma cilíndricapoco profunda como en el caso de loscyathostominos con lo que su acciónsobre la mucosa intestinal es poco con-siderable. Además la fase parasitaria delciclo biológico de estos nematodos trans-curre enteramente en el tubo intestinaldel equino sin lesionar por lo tanto órga-nos internos (17).Sin embargo, en los últimos años se harealizado una profusa actividad de in-vestigación en los pequeños estrongyli-dos debido fundamentalmente a tres he-chos: 1) el desarrollo y uso, en el últimocuarto del siglo pasado, de drogas anti-helmínticas de gran eficacia contra losgrandes estrongylidos ha determinadoque éstos hayan perdido significación(12). Por otra parte, estas drogas, hanmostrado menor eficacia frente a los pe-
Presencia de nuevos géneros y especies de estrongilidos en los equinosen Uruguay
Falcón, J.D. 1
RESUMENSe comunica la presencia, por primera vez, en equinos de Uru-guay, de las siguientes especies de nematodos: Triodontopho-rus brevicauda (Loos, 1902), (Strongylidae – Strongylinae) yCylicodontophorus bicoronatus (Loos, 1900), Cylicocyclusbrevicapsulatus (Ihle, 1920), Cylicocyclus elongatus (Loos,1900) (Strongylidae – Cyathostominae).
Palabras clave: Triodontophorus, Cylicodontophorus,Cylicocyclus.
SUMMARYThe presence, for the first time, in equines of Uruguay of thefollowing species of nematodes Triodontophorus brevicauda(Loos, 1902), (Strongylidae – Strongylinae) and Cylicodonto-phorus bicoronatus (Loos, 1900), Cylicocyclus brevicapsula-tus (Ihle, 1920), Cylicocyclus elongatus (Loos, 1900) (Stron-gylidae–Cyathostominae) is reported.
Keywords: Triodontophorus, Cylicodontophorus, Cylicocyclus.
Recibido: 30/09/02 Aprobado: 28/10/021Departamente de Parasitología y Enfermedades Parasitarias. Facultad de Veterinaria. Universidad de la RepúblicaOriental del Uruguay. Montevideo. A. Laspaces 1550. CP 11600.
27-30
Comunicación Corta

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002)28
queños estrongylidos y desde finales dela década de los setenta se vienen publi-cando trabajos que dan cuenta de la exis-tencia, en diferentes partes del mundo(5, 6, 7, 8), y también en Uruguay (3)del fenómeno de resistencia de este gru-po de nematodos hacia los benzimidazo-les , 2) se ha reconocido en algunos tra-bajos la importancia de los estados lar-varios de los pequeños estrongylidos,cuando emergen de la mucosa de intesti-no grueso, ciego y colon, en la etiologíade un importante síndrome que puedecausar un severo cuadro de colitis o lamuerte en equinos (13, 14, 19), 3) se hanobtenido promisorios resultados para elcontrol biológico de este grupo de pará-sitos a través del uso de determinadasespecies de hongos nematófagos (2, 9).Lo anteriormente mencionado ha hechoque la infección con cyathostominos seahoy un problema parasitario emergentede los equinos.En su revisión sobre la fauna parasitoló-gica diagnosticada en Uruguay, Castro yTrenchi (4) mencionan solamente den-tro de la familia Strongylidae a los géne-ros Strongylus con sus especies S. vul-garis, S. equinus y S. edentatus. Otracomunicación mencionada en dicha pu-blicación se refiere a “Cylicostoma”, noaclarando a que se refiere el autor conesta denominación.Es necesario conocer cual es la situaciónactual a nivel nacional con respecto a esteproblema sobre el que se deberá seguirinvestigando en el futuro y creemos queel primer paso es determinar claramentecuales son los géneros y eventualmentelas especies de cyathostominos presen-tes en el país.
MATERIALES Y MÉTODOSSe utilizó el tubo gastrointestinal com-pleto de dos equinos, no pudiendo de-terminarse datos de los animales debidoa que las plantas de faena adquieren losanimales a vendedores que los obtienende diferentes lugares del país.En el laboratorio se procesó el materialincidiendo con enterótomo la pared delciego y colon en toda su longitud e ins-peccionando cuidadosamente tanto lasuperficie de la mucosa de dichos órga-nos como su contenido, el que fue sepa-rado en pequeñas porciones para facili-
tar la visualización de los vermes. Loshelmintos adultos se separaron de lamateria fecal mediante aguja parasitoló-gica depositándolos primeramente ensolución salina fisiológica para ser lava-dos y posteriormente se transfirieron arecipientes adecuados con solución deformol comercial al 10% para su conser-vación.Previo aclarado con lactofenol se proce-dió a montarlos en gelatina para su pos-terior estudio al microscopio óptico a400 aumentos. Se determinó la longituden milímetros, y se estudió fundamen-talmente la morfología de la cápsula bu-cal y de la extremidad posterior tanto delos ejemplares machos como de las hem-bras. Para la identificación de géneros yespecies se usó la clave de Lichtenfels(10) y el trabajo monográfico ilustradode Tolliver (18). Los nombres de las es-pecies de acuerdo a lo recomendado porLichtenfels et al. (11).
RESULTADOSSe determinó la presencia de las siguien-tes especies de nematodos: Triodonto-phorus brevicauda (Loos, 1902) ( Stron-gylidae – Strongylinae) y Cylicodonto-phorus bicoronatus (Loos, 1900), Cyli-cocyclus brevicapsulatus (Ihle, 1920),Cylicocyclus elongatus (Loos, 1900)(Strongylidae – Cyathostominae)..
DISCUSIÓNLa principal característica morfológicadistintiva de los nematodos pertenecien-tes al género Triodontophorus es su cáp-sula bucal de forma subglobular en la quese destacan 3 grandes dientes que pare-cen emerger desde la entrada del esófago(10). En los ejemplares observados almicroscopio óptico pudimos reconoceren su cápsula bucal las característicasmorfológicas (Figura 1) que nos llevarona al diagnóstico de la especie Triodonto-phorus brevicauda, fundamentalmente laforma de sus dientes, cada uno, con unasuave ranura en forma de U en el medioy la extremidad anterior del nematodosimilar a un pequeño plato boca arriba(Tolliver). Las medidas de longitud ob-tenidas en dos machos fueron de 14,6 y15 mm. las que coinciden con las men-cionadas por otros autores (15).Dentro de la subfamilia Cyathostominaeel género Cylicodontophorus se distin-gue morfológicamente por poseer unacápsula bucal corta y de paredes gruesascon coronas de dentículas interna y ex-terna bien notorias. Lichtenfeld (10) des-cribe, dentro de este género, tres espe-cies que son: C. bicoronatus, C. euproc-tus, y C. mettami, estableciendo comocaracterística morfológica diferencial masimportante entre ellas la relación entrelargo y ancho de los elementos que for-
Figura 1. Triodontophorus brevicauda.Extremidad anterior 400 x. Seobserva lacápsula bucal con los dientescaracteristicos del género en el fondo. Enla parte más apical de la extremidad seobserva una estructura con apariencia de«platillo boca arriba» que distingue a estaespecie.
27-30

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002) 29
man las coronas radiadas interna y exter-na, siendo las dentículas de ambas coro-nas de aproximadamente el mismo tama-ño en C. bicoronatus y notoriamente di-ferentes entre si en las otras dos espe-cies. Otros investigadores (18) comple-mentan la descripción morfológica de C.bicoronatus mencionando que la coronaradiada interna está formada de elemen-tos fuertes que semejan los dientes de unpeine (Figura 2). Las medidas de longi-
tud obtenidas en dos ejemplares hem-bras fueron de 13,7 y 12,5 mm. las quecoinciden con las mencionadas por otrosautores (15).Los nematodos pertenecientes al géneroCylicociclus se caracterizan fundamen-talmente por ser nematodos de media apequeña talla y provistos de una cápsu-la bucal que en general es rectangular enla mayoría de las especies que han sidodescritas. Destacan también las papilas
laterales que son anchas y prominentes(10). Se comprobó la presencia de dosespecies: C. brevicapsulatus (Figura 3)que posee una cápsula bucal muy pocoprofunda e inaparente con elementos dela corona radiada externa anchos y biennotorios. También comprobamos la pre-sencia de C. elongatus (Figura 4) especiecaracterizada por su tamaño similar al delos medianos estrongilidos y morfológi-camente distinguibles por su cápsula
Figura 2. Cylicodontophorus bicoronatus. Extremidad anterior 400 x. Seobserva la cápsula bucal muy corta y con paredes que se abrenhacia fuera y los fuertes elementos de la corona interna quesemejan a los «dientes de un peine»
Figura 3. Cylicociclus brevicapsulatus. Extremidad anterior 400 x. Seobserva la cápsula bucal extremadamente corta e inaparente.
Figura 4. Cylicociclus elongatua. Extremidad anterior 400 x. Seobserva la cápsula bucal con pocos y muy anchoselementos de la corona externa. Escala: Línea en bordesuperior izquierdo corresponde a 100 micras.
27-30

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002)30
bucal bastante ancha con pocos y an-chos elementos de la corona radiada ex-terna y un esófago muy largo y recto(10). En el caso de C. brevicapsulatusobtuvimos medidas de longitud total de8 mm. en los machos y entre 9,7 y 11,5mm en la hembras mientras que para C.elongatus obtuvimos medidas de 18 mm.en dos hembras. En ambos casos estasmedidas coinciden con las mencionadaspor otros autores (15).
CONCLUSIONES YRECOMENDACIONESDe acuerdo a los datos obtenidos en estetrabajo se concluye que las especies men-cionadas se encuentran integrando la fau-na parasitaria en los equinos de Uruguay.Investigaciones en países limitrofes (16)comunican la presencia en ellos de unaamplia gama de especies de Cyathosto-minos, situación ésta que seguramente
es similar en Uruguay. Es necesario con-tinuar investigando hasta dilucidar total-mente el espectro de especies presentesen el país para en etapas posteriores en-carar estudios epidemiológicos que nosconduzcan a establecer planes de con-trol mas eficaces.
Referencias Bibliográficas1. Amaro J., Ormaechea D., Capurro
F., Diana V., Pessano G., Sallúa S.(1992). Presencia y prevalencia deFasciola hepática y helmintos gas-trointestinales en una muestra deequinos deportivos en el Uruguay.Veterinaria, Vol. 28 No. 116.
2. Bird J., Herd, R.P. (1995). In vitroassessment of two species ofnematophagous fungi (Arthrobotrysoligospora and Arthrobotrysflagrans) to control the developmentof infective cyathostome larvae fromnaturally infected horses. Vet.Parasitol. 56: 181-187.
3. Castells, D.; Trezza, C.; Sacco, G.;Ponce de León, L.I. (1995).Resistencia antihelmíntica deCiatostomas (pequeños estróngilusdel equino) al fenbendazol.Veterinaria. Vol. 31, No. 127, julio-setiembre 1995.
4. Castro, E.R. y Trenchi, H. (1954).Fauna parasitológica comprobada enel Uruguay. Revista de MedicinaVeterinaria. Tomo VII, Año XIX.
5. Gawor, J. (1995). Resístanse ofCyathostominae to bencimidazolepreparation in horses. MagazynWeterynarijny 4: 101-102.
6. Genchi, C., Sacco, B. di., Traldi. G.,Nogara. B., Quintavalla. F., Di.Sacco. B. (1992).First observationsin Italy on the resistance of smallstrongyles (Cyathostominae) to
benzimidazoles and the efficacy ofpyrantel pamoate. Ippologia. 3: 2, 77-80.
7. Herd, R.P.; Coles, G.C. (1995).Slowing the spread of anthelminthicresistant nematodes of horses in theUnited Kigdom. Vet. Rec. 136:481-485.
8. Ihler, C.F. (1995). A field study onanthelmintic resistance in equinesmall strongyles in Norway. ActaVeterinaria Scandinavica 36: 135-143.
9. Larsen, M.; Nansen, P.; Grondahl,C.; Thamsborg, S.M.; Gronvold,J.; Wolstrup, J.; Henriksen, S.A.;Monrad, J. (1996) The capacity ofthe fungus Duddingtonia flagrans toprevent strongyle infection in foalson pasture. Parasitology 113: 1-6.
10. Lichtenfels J.R. (1975). Helminthsof Domestic Equids. Proceedings ofThe Helminthological Society ofWashington. Vol. 42.
11. Lichtenfels J.R.; Kharchenko,V.A.; Krecek, R.C.; Gibbons, L.M.(1998). An annotated checklist bygenus and species of 93 species levelnames for 51 recognized species ofsmall strongyles (Nematoda:Strongyloidea: Cyathostominea) ofhorses, asses and zebras of the world.Vet. Parasitol. 79: 65-79
12. Love, S., Murphy, D., Mellor D.(1999). Pathogenicity ofcyathostome infection. Vet.Parasitol. 85: 113-122.
13.Mair, T.S. (1994). Outbreak of larvalcyathostomiasis among a group ofyearling and two-years-old horses.Vet. Rec. 135: 598-600.
14. Mansmann, R. (1997). Clinicalaspects of equine larvalcyathostomiasis. Large AnimalPractice 18: 30-32.
15. Quiroz Romero, H. (1990)Parasitología y EnfermedadesParasitarias de los AnimalesDomésticos. 4ª. Ed. México, Limusa.876 p.
16. Silva, A.V.M.; Costa, H.M.A.;Santos, H.A.; Carvalho, R.O.(1999). Cyathostominae (Nematoda)parasites of Equus caballus in someBrazilian states. Vet. Parasitol. 86:15-21.
17. Soulsby, E.J.L. (1987) Parasitologíay Enfermedades Parasitarias en losAnimales Domésticos. 7ª . Ed.México, Interamericana. 823 p.
18. Tolliver S.C. (Monografía) (2000).A Practical Method of Identificationof the North AmericanCyathostomes (Small Strongyles) inEquids in Kentucky. University ofKentucky. College of Agriculture.Department of Veterinary Science.Lexington, Kentucky 40546.
19. van Loon, G., Deprez, P., Muylle,E., Sustronk, B. (1995). Larvalcyathostomiasis as a cause of deathin two regularly dewormed horses.J. Vet. Med. Series A842). 301-306.
27-30

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002) 319-12 (2001)
La rabia es una enfermedad muy vieja, talvez tan vieja como la propia humanidad.Tres mil años antes de Jesucristo ya seencuentra el origen de la palabra "rabia"en la lengua sánscrita, donde "Rabhas" sig-nifica "agredir".La palabra griega "lyssa" viene de la raíz"lud": "violento". La primera descripciónde la enfermedad se remonta a muchosaños antes de Jesucristo, en el Código Es-huma en Babilonia. Desde la antigüedad yase había establecido la relación entre larabia humana y las mordeduras de los ani-males rabiosos (especialmente perros).Girolamo Fracastoro, sabio italiano naci-do en Verona, describió la enfermedad (quehabía podido observar en numerosos pa-cientes) y sus modos de contaminación, yesto en 1530, es decir ¡350 años antes deque Luis Pasteur trabajara en la vacunaDurante el siglo XIX la rabia canina o ra-bia de la calle es por dondequiera un verda-dero flagelo, particularmente en Europa.El miedo a la rabia, debido a su modo decontaminación y a la ausencia de trata-miento eficaz, se había vuelto irracional.Las personas mordidas por un perro sos-pechoso de rabia se suicidaban o eran sa-crificadas.En este mundo de miedo irracional, el pri-mer tratamiento post-exposición realiza-do en 1885 por Luis Pasteur dio a estegran sabio una aura internacional que nohabían sido suscitado hasta entonces susotros importantes trabajos científicos.
LOUIS PASTEUR (1822-1895)Fue este tal vez el investigador que másvidas haya salvado. Sus estudios orienta-dos demostrar su creencia de la falsedad dela Teoría de la Generación espontánea, levalieron honores y enemigos. Pero al mis-mo tiempo y gracias a su tesón y enfervo-rizado trabajo, hicieron que la ciencia mé-dica diera su más trascendental paso: elconocimiento y dominio de los gérmenespatógenos.
Sus estudios se abocaron a las más diversaspatologías animales y humanas: desde unaenfermedad de los gusanos de seda, el car-bunco, y por supuesto la Rabia.El dos de junio de 1878, Pasteur en una pruebade campo que involucró a 60 carneros, de-mostró ante numeroso público la eficacia dela primera vacuna, cuando de la poblacióninfectada con Ántrax solamente permane-cieron vivos y sanos los veinticinco anima-les que previamente se habían inoculado conaquella primitiva vacuna y diez individuostestigos que no habían tenido contacto conla enfermedad.En 1880 Pasteur se aboca de lleno al estu-dio de la rabia, apoyándose en estudios deotros investigadores no tan conocidos hoydía, tales como Pierre Galtier, quien entreotras cosas se le considera el descubridorde la presencia del virus de la rabia en lasaliva de los infectados.Comienza Pasteur a trabajar en la búsque-da de la vacuna de la rabia, motivado porla profusión de casos de la enfermedad enFrancia. Comienza pues, a experimentarcon perros rabiosos. Había descubierto queel virus (aunque ni por asomo sospecharde que tipo de microorganismo se trataba)además de en la saliva, se encontraba enlos tejidos nerviosos, y es así que comien-za a experimentar con perros rabiosos ysanos, así como con conejos.Luego de varios experimentos logra ate-nuar el virus y con ese material logra lasprimitivas vacuna, las que fueron usadashasta entrado el sigo veinte. Luego de Pas-teur, el avance de la técnica permitió puri-ficar la vacuna de la rabia hasta llegar a laque hoy tenemos, de cultivos celulares.El virus de la Rabia es, junto con el de lapoliomielitis, el más neurotropo de losvirus.
GÉNERO LYSSAVIRUS :En este género, diferentes serotipos ha-bían sido caracterizados gracias al empleode una bacteria de anticuerpos monoclo-
nales. Es así que 4 principales serotiposhan sido identificados.El serotipo 1 comprende todas las cepasde virus rábico (rabia salvaje, rabia de lascalles, las cepas de rabia fijas y las cepasvaccinales).Los otros serotipos son consideradoscomo virus emparentados con la rabia. Observado con microscopio electrónico,el virión rábico presenta un diámetro de75 nm y un largo que varía entre 130 nm; La envoltura viral está erizada de espícu-las de 9 nm de longitud.Una sóla molécula de A.R.N. está presentepor virión, constituída de 11932 nucleó-tidos, de un peso molecular de 4,6.106daltones. El A.R.N. del genoma es de pola-ridad negativa, indicando que no es infec-cioso en sí mismo. Luego de la penetra-ción en la célula, el A.R.N. genómico, debeser transcripto en moléculas complemen-tarias positivas, capaces de producir lasproteínas virales.Contrariamente a los virus transportadospor vía sanguínea, el virus rábico es trans-portado por vía nerviosa, es decir que tomaprestado el sistema de transporte de sucélula -blanco (la neurona) para progresarde la periferia (sitio de inoculación) hacialos centros nerviosos. Este sistema detransporte hace intervenir los microtúbu-los de la célula.El neurotropismo del virus de la rabia, esevidente en la patogenia de la enferme-dad. Desde la lesión local por donde se in-troduce en los tejidos, pasa a lo largo delos nervios hasta el SNC.La viremia es rara, pero una diseminaciónhematógena puede ocurrir en condicionesexperimentales, como al infectar perrospor vía sanguínea. La infección da por re-sultado una extensa destrucción de la cor-teza cerebral y cerebelosa , cerebro medio,ganglios basales, protuberancia y bulbo, condegeneración y desmielinización neuronal.En la médula espinal se observan altera-ciones similares, más intensas en las astasposteriores. Hay una hiperemia e infiltra-
Diagnóstico
La rabia
Rossi, D.1
Recibido: 08/07/02 Aprobado: 02/09/02Conferencia dictada en el Hospital de Clínicas, UDELAR.1Ejercicio liberal E-mail: [email protected]
31-34

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002)32
ción general, infiltración mononuclear ya veces pequeñas hemorragias peri vascu-lares . La extensión de estas lesiones va adepender de la duración de la enfermedad.Cuando la muerte ocurre tempranamente,las lesiones son mínimas. En forma cen-trífuga, a partir del SNC, el virus tambiénllega a las glándulas salivales, haciendo dela saliva la principal fuente de infección.
DIAGNÓSTICOEs muy importante el diagnóstico de labo-ratorio en el animal que ha mordido. Lasaliva es infecciosa antes de que aparezcanlos síntomas de la rabia. Básicamente seintenta conservar al animal sospechosovivo por lo menos por diez días, y de esamanera permitir el desarrollo de la enfer-medad, y así facilitar el diagnóstico. Estese basa - tanto en animales inferiores,como en el hombre en encontrar los cor-púsculos de Negri, fundamentalmente enel Asta de Ammon por frotis por impre-sión.Los corpúsculos de Negri, son formacio-nes acidófilos de inclusión, que son patog-nomónicos de la rabia. Los encontramosen el citoplasma de las grandes células gan-glionares , más abundantemente en el Astade Ammon, en la capa de células piramida-les de la corteza cerebral y la capa decélulas de Purkinje del cerebelo y los gan-glios basales. Miden de 2 a 10 micras, peroeste tamaño puede ser mayor o menor. Sepueden teñir con Giemsa , aunque muchosinvestigadores opinan que es superior latinción con el método de Sellers (fucsinabásica y azul de metileno en metanol) ti-ñendo los cuerpos de Negri de magenta ylos gránulos internos de azul. Si bien comodije son patognomónicos de la rabia, en un10 al 25 por ciento de los casos de infec-ciones en las que se ha aislado el virus, nohan podido ser encontrados. Son básica-mente agrupaciones de virus.
Durante años se usaron las tinciones deGiemsa o de Sellers, pero últimamente seprefiere la técnica de Inmunofluorecen-cia. Esta permite visualizar cuerpos me-nores que no pueden verse con tinciones yademás para ubicar el virus en las glándu-las salivales. El virus también se puede ais-lar por inoculación de cerebro de ratón, yla visualización de los corpúsculos de Ne-gri en las células cerebrales del mismo.
LOS VECTORES DE LA RABIAEl principal vector y reservorio en el mun-do es el perro. Los animales salvajes pue-
den agredir directamente al hombre o demanera indirecta contaminando a los ani-males domésticos. Es inmensa la cantidadde vectores que pueden transmitir la rabia,y la diversidad va a depender de la zona delglobo que se trate.Está presente en todos los continentes, ysegún la OMS, solo en 1992 murieron enel mundo miles de muertes por rabia hu-mana.Por ejemplo en América del Sur, constitu-yen un grave peligro los murciélagos he-matófagos (Desmodus rotundus rotundus), así como los insectívoros.Entre el Río Grande y el estuario del Pla-ta, los primeros se cuentan por millones,y son los responsables de cómo mínimounas 150 muertes por rabia humana, y de500.000 a 1:000.000 de muertes de resespor rabia bovina, con el consiguiente per-juicio económico.En los lugares donde este problema es gra-ve, se ha recurrido a la unión del conoci-miento con el ingenio para poder comba-tir el flagelo.
Estudios realizados en los vampiros, de-mostraron que estos animales carecen deelementos en sus organismos para regularla coagulación de su sangre, es por eso quenecesitan de sangre de otros mamíferos.De no conseguirla, la muerte del quirópte-ro es inminente. Basados en esto, es quelos investigadores comenzaron a inyectara las reses cantidades inofensivas de anti-coagulantes. Los vampiros que muerdenesas reses, mueren por falta de factores dela coagulación en un plazo máximo de tresdías.En Nicaragua, estas técnicas de luchas con-tra los murciélagos hematófagos hizo quela producción de leche aumentara un 8%.En un solo rebaño de vacas que registrabaunas 1500 mordeduras diarias de murciéla-go, con una pérdida diaria de 90 litros desangre por día, mediante la inoculación deanticoagulantes, se logró llegar a tan solosiete al cabo de una semana.
En nuestro país hay grandes colonias deDesmodus rotundus, pero afortunadamen-te se trata de individuos libres del mal. Espor eso, y ante la indicación de los biólo-gos y expertos en el tema, que no es con-veniente su exterminio, ya que en caso dequedar sus cuevas y refugios vacíos, el ries-go que se corre es que los mismos seanocupados por colonias de animales infec-tados con la rabia.
En América del Norte, los reservorios de larabia, son las mofetas, los zorros, los mapa-ches y los murciélagos insectívoros.Solo en 1995, fueron declarados en Américadel Norte 7877 casos no - humanos de rabia.En Europa, la rabia ha evolucionado mu-cho en el curso del último siglo. Luego dela erradicación de la rabia canina gracias ala vacunación de los perros domésticos yde la eliminación de perros callejeros, variospaíses de Europa del Oeste han permanecidoindemnes de rabia durante períodos más omenos largos.Al final de la última guerra mundial, la adap-tación del virus rábico a los zorros ha per-mitido a la rabia invadir numerosos países.La caza de zorros y la utilización de vene-no no disminuyeron la rabia vulpina enEuropa. No ha sido hasta la utilización devacunas antirábicas orales contenidas enlos cebos que el número de casos de rabiaha sensiblemente disminuído en toda Eu-ropa. En Francia, ésta disminución ha sidoparticularmente remarcable.Los casos de rabia humana originarios sonescasos en Europa. Ocurren sobre todo enlos países del este europeo donde la rabiacanina no es importante (menos de 10casos por año).Excepcionalmente, las personas mordidasen regiones de rabia endémica (África,Asia) desarrollan la enfermedad en un paíseuropeo.El principal vector de la rabia en Asia es elperro, allí en la mayoría de los casos derabia humana son identificados. Es gene-ralmente admitido que el número estima-do de muertos (cifra superior al númerooficialmente declarado) es del orden de40.000, de los cuales la mayoría de loscasos se encuentran en India. Ciertos paí-ses han establecido programas nacionalesde lucha contra la rabia que han hecho dis-minuir sensiblemente el número de muer-tos (China, Indonesia, Malasia, Tailandia).El perro continúa siendo el principal vec-tor de rabia en África (alrededor del 90%).Más de 4000 casos de rabia animal hansido diagnosticados (informe de la OMS)que no reflejan sino una parte de la si-tuación de la rabia en África.Alrededor de 100-200 personas muerende rabia cada año. Sin embargo, en lamayoría de esos casos, el diagnóstico esúnicamente clínico. Como en Asia, hayverdaderamente una subestimación delnúmero de casos de rabia.
31-34

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002) 33
En nuestro país, podemos decir que larabia está con nosotros desde antes queseamos una nación independiente. JoséP. Barrán habla en la su obra historia dela sensibilidad uruguaya, que ya en losalbores del siglo XIX, dentro de las mu-rallas de Montevideo, uno de los peli-gros que podían acechar a los poblado-res al pasar por uno de los tantos terre-nos baldíos que había en la plaza fuerteera, el ataque de algún perro rabioso.Agrega para graficar, que los cadáveresde perros con rabia, que las autoridadeso los ciudadanos mataban, quedaban ti-rados en las calles, por falta de un servi-cio que se encargara de la limpieza de lasmismas, aumentando y haciendo inso-portable el olor que había en la ciudad.La Rabia, tiene el raro privilegio de ha-ber sido la primera enfermedad animaldocumentada en una publicación en Uru-guay, y es gracias al trabajo del Dr. Ani-bal Durán del Campo, Uruguay: Pro-ducción de carne, perspectiva históricay contribución de la profesión veterina-ria a la misma (Prácticas veterinarias Nº3,Diciembre 1999) que tomamos conoci-miento de ello., Quien lo hizo no fue otroque Dámaso Antonio Larrañaga, en sulibro "Viaje de Montevideo a Paysandú".El relato de este viaje de 1815, en el queLarrañaga concurre a reunirse con el Ge-neral Artigas y algunos Diputados de laProvincia Oriental en Paysandú, setransforma en una maravillosa crónica dela realidad de la campaña y sus habitan-tes, así como una minuciosa descripciónde la flora y la fauna de la región, de lacual el sacerdote era un profundo cono-cedor.En una parte de la misma, Larrañaga, re-firiéndose a una de las paradas del viaje,dice:"Tratamos de cenar prontamente y me-ternos en nuestras camas tendidas en elsuelo sobre cueros a fin de abrigarnoscon nuestras cubijas. Nuestro mayor cui-dado en medio de tantas incomodidades,era atar bien los cueros que servían deparapeto, ya no tanto para el frío cuantopor temor a los perros rabiosos que, paranuestra desgracia hay muchos en estacampaña, y acababan de matar uno eneste mismo día, que vino a los ranchos.Esta plaga la experimentamos desde laguerra última de los ingleses."Mas adelante en el relato, Larrañaga re-lata unos consejos que les dio a los luga-
reños de la forma de proceder en caso demordedura por un perro rabioso:"...procuré recomendar a estos vecinosque inmediatamente que se sintiesenmordidos tratasen de dilacerar la herida,cuidando de no dejarla cerrar auxiliándo-se de algún cáustico, aunque fuese conun hierro caldeado, pues esta es la únicae infalible medicina contra la hidrofobiao rabia. También les hablé de algunasyerbas que recomienda últimamente elSr. Cavanilles en sus Anales, como el"Echium Vulgare" o borraja cimarrona yotras plantas , que cubren nuestros ca-minos." "De todas estas plantas secas ypulverizadas se toman como dos nariga-das por dos veces en diez o doce días, ysin más régimen, asegura dicho autor ha-ber hecho curas prodigiosas. Yo he he-cho la experiencia y surtió buen efectoen un pobre paisano, bien que no puedoasegurar se debiese a esto solo la cura,pues ya se había aplicado otros innume-rables tratamientos."También en su obra, "La Banda Orientalhacia 1880", el autor Felix de Azara, abor-da el tema. Allí sugiere un reglamentogeneral de la campaña, en el que toca in-numerables tópicos atinentes a la desor-denada extramuros, y en el último puntodice:"duodécimo: exterminar los perros ci-marrones, lo que no se conseguirá porlos medios que se practican , sino tra-yendo de Cataluña la fruta silvestre lla-mada Mataca, para echar sus polvossobre las reses muertas, porque así pe-recerían todos sin remedio, y lo mismolos tigres y leones." Como vemos por las crónicas de la épo-ca, de las cuales estos son solamente dosde la gran cantidad de testimonios que serefieren al tema, era una gran preocupa-ción la que se sentía con respecto al temade la Rabia, y la forma en que se luchabafundamentalmente era con las matanzasdel principal vector: el perro. Eran varios los tratamientos que se prac-ticaban para curar el mal, aunque comosabemos una vez que han aparecido lossíntomas nerviosos de la enfermedad, lamuerte es cosa inexorable.Todavía no se conocían los trabajos dePasteur, en procura de la vacuna. Y sepuede decir que el método radical de lamatanza de perros, en forma permanen-te y sistemática dio en cierta forma susresultados.
Los exterminios eran realmente asombro-sos, se habla de que "se formaban ejérci-tos de paisanos que acorralaban a la pe-rrada en las rinconadas de ríos y arro-yos, para matarlos a lanza y bala"Hay documentos de la época que men-cionan que utilizando ese procedimien-to, se habrían llegado a matar en una solajornada 10.000 perros en Rincón de SanJosé, y en 1852, en Rincón de Tacuarí,más de 13.000.
LA ESPADA DE DAMOCLES Hoy en día, en nuestro país aparentemen-te no hay rabia. Y digo aparentemente,porque no se efectúan de rutina los análi-sis de los animales muertos con sintoma-tología nerviosa, como se hacía antaño.Cualquier informe de los organismosmundiales o continentales de la salud,cuando hacen un relevamiento de la si-tuación de la rabia, país por país, se re-fieren a Uruguay como: "sin apariciónde casos de la enfermedad".Esto que debería ser una tranquilidad yun orgullo, se ve opacado es en realidaduna pesada espada de Damocles balan-ceándose sobre nuestras cabezas. Lo queantes nos enorgullecía, hoy nos aver-güenza.Siempre fuimos pioneros en la lucha con-tra el flagelo. Contábamos un Institutoantirrábico que era orgullo en el mundoentero, donde no solo se analizaban ani-males y personas sospechosas de tenerla enfermedad, se preparaba el conjuga-do necesario para los estudios de Inmu-nofluorecencia y se garantizaba la segu-ridad de las vacunas que había en plaza,sino que además de ello se planificabanlas campañas tendientes a lograr la erra-dicación de la enfermedad, que eran ejem-plo para otros países.En el año 1967,por ejemplo, Uruguaytrajo a un eminente experto en el tema,como lo era el médico peruano Dr. Ave-lio Málaga Alba, quien organizó y dirigióla campaña en dicho año. Al terminar lamisma, el Dr. Málaga comentó que nun-ca en todos sus años de dirigir campañasen los más diversos países, había podidollevar a cabo una tan excepcional como laque hizo en Uruguay.Toda la población montevideana se mo-vilizó en aras de ese objetivo. Se hizouna vacunación masiva de canes con pro-pietario, se capturó a conciencia a pe-
31-34

Veterinaria, (Montevideo) 37 (147-148): 1-20 (2002)34
rros vagabundos. En años anterioresMontevideo había sufrido una gran can-tidad de casos de perros rabiosos, losque habían ocasionado tres muertes enseres humanos, los que murieron respec-tivamente en los hospitales Vilardebó,Pedro Visca y Pereira Rossell.Los últimos casos de rabia canina fuerondetectados escapando de Montevideohacia el este, con el consiguiente peligrode que el mal se transformara de urbana asub urbana, con el desastre que ello sig-nificaría.Ya en el año 1965, el Uruguay tambiénfue pionero con la vacunación pre expo-sición de profesionales, con una vacunade cerebro de ratón lactante, la que sehacía en el Instituto antirrábico, dirigidopor los doctores Dilandro y Lorenzo.Veterinario uno y médico el otro.Hoy la realidad es otra: el instituto dezoonosis está totalmente desmantelado.En el año 1995, atendiendo los clamoresde determinados grupos de protección alos animales, se suspendió la captura decanes en la vía pública, y al mismo tiem-po se le sacó todo -o casi todo- al Insti-tuto de Zoonosis (antiguamente antirrá-bico). No hay personal, ni camionetas nimedios para realizar un diagnóstico.La realidad regional no es para nada tran-quilizadora,Tanto en la Argentina, como en Brasil yespecialmente en Paraguay los casos derabia van en aumento. Hasta en Chile,que cuentan con la excepcional barreraque significa la Cordillera de los Andes,recientemente han tenido la muerte deuna niña mordida por un gato, infectadoa su vez, por un murciélago insectívoro.La comunidad científica debe reaccionarde una manera drástica y contundente.Se deben ponderar las opiniones y po-ner en su exacto punto quién y cómopuede opinar de este tema. Es así que novale lo mismo la opinión de un técnicoque haya dedicado su vida a estudiarlo,que el de una ama de casa que siente com-pasión por los perritos.La profesión veterinaria sufrió en carnepropia este tipo de desatinos, cuando enel año 1994 se impuso al Uruguay "librede aftosa sin vacunación".En ese momento, en un Foro sobre laenfermedad que se realizó en el seno dela Sociedad de Medicina Veterinaria del
Uruguay, se concluyó que de ningunamanera nuestro país estaba preparadopara ser considerado así.No obstante, por parte del ministerio deganadería, agricultura y pesca (que comoanécdota diré que estaba dirigido por unabogado) se dijo que eso era erróneo, yse procedió contrario a la opinión de losexpertos.Las consecuencias de ello las estamosviendo hoy, y las pagamos todos.No debemos dejar que con la rabia ocu-rra lo mismo. La campaña debe estar enmanos de científicos, médicos, veterina-rios, expertos en epidemiología y en edu-cación, así como de todos aquellos quese hayan especializado en el tema.Se debe proceder de la misma manera quese hizo cuando el último brote de rabiaen el país: lograr que la población se com-prometa con la campaña. Es la únicamanera.Los que debemos hablar, y para ello esmenester contar con el total apoyo ofi-cial, solamente pueden ser los que sabe-mos del asunto.Hace poco hubo una reunión en el des-pacho de un diputado. Allí fui invitado,y compartí la mesa con una serie de se-ñoras protectoras de animales, y en de-terminado momento, mientras yo inten-taba decir lo que pensaba acerca del temarabia, fui interrumpido por una de estasmujeres, la que con una autoridad queasustaba me espetó en la cara:Eso es todo mentira. La rabia solo des-apareció en el siglo XIX, y tan solo fue-ron unos cientos de casos.A eso es a lo que me refiero: La bibliajunto al calefón.Debemos contar con una buena ley detenencia responsable de los animales decompañía, no un mamarracho. La profe-sión veterinaria (Sociedad de MedicinaVeterinaria, la Facultad y la AcademiaNacional de Veterinaria) elaboró un an-teproyecto de ley, el que actualmente estásiendo analizado en el parlamento y enello depositamos nuestras mayores ex-pectativas de llegar a un buen puerto.Se debe jerarquizar y apoyar el trabajode los técnicos del instituto zoonosis,del laboratorio Rubino y de todas lasdependencias de Salud Pública que tra-bajan en el problema.
También hay que abordar el tema delperro vagabundo, así como del callejero.Y este tema es de por sí espinoso, por-que no hay duda que es el talón de Aqui-les de toda campaña que se encare, yaque tiene un componente de sensibilidadmuy fuerte, que generalmente hace quela gente ponga el grito en el cielo, y porlo tanto que muchos políticos -por mie-do a perder popularidad- miren para elcostado cuando se les exige aplicación denormativas claras.La mayoría de las protectoras de anima-les se arrogan el título de ser represen-tantes de organizaciones mundiales deprotección a los animales, y en realidades mentira. La prueba de esto está en quela Sociedad Mundial de Protección a losAnimales, recomienda la captura y euta-nasia del perro vagabundo.¿Qué quiere decir esto?¿Qué es gente queodia a los animales y quiere matarlos poralgún tipo de placer sádico? Para nada,es gente que básicamente está asesoradapor veterinarios, por médicos o por bió-logos, que son quienes dirigen las cam-pañas sanitarias, no una manga de sabi-hondos, muchas veces con objetivos nomuy claros.Pero, como ya dije, para que esto ocurradebe haber un verdadero compromisopolítico de las autoridades. Sin el mis-mo, nada, ni las reuniones ni las campa-ñas, sirve para nada.Se debe realizar una verdadera campañade información a la población, y de estamanera contar con un pueblo conscientey comprometido con la campaña.Hoy vemos con agrado, que existe unaintención por parte de las autoridades deSalud Pública en hincarle el diente al asun-to, y lo aplaudimos. No solo lo aplaudi-mos, sino que además participamos delemprendimiento con muchísimas espe-ranzas.Es nuestra obligación como universita-rios aportar a la sociedad en lo que sabe-mos. Somos los referentes del pueblo eneste tema, y es nuestra obligación retri-buir -más que con impuestos-, con nues-tro trabajo y opinión la educación queaquel nos ha pagado.
31-34





























