Embed Size (px)
Citation preview
VERNAL BEHAVIOUR OF THE YELLOW-BELLIED MARMOT (Marmotaflaviventris)
BY KENNETH B. ARMITAGEDepartment of Zoology, The University of Kansas, Lawrence .
The social behaviour of the yellow-belliedmarmot was studied during 4 aestival seasons(Armitage, 1962) . The behaviour was character-ized by a pattern of home ranges, many over-lapping, that changed from year to year . Knownindividuals frequently had different home rangesand occupied different home burrows in success-ive years. Agonistic behaviour among adultfemales and between yearlings and adult femalesevidenced a steady decrease throughout . thesummer and almost completely ceased duringthe onset of hibernation in mid-August . Allthese conditions suggested that the vernalseason was a period of social instability andwas the period during which mating behaviouroccurred and during which home burrows wereselected and home ranges established .
During the spring of 1961 1 had an oppor-tunity to study vernal behaviour . The, object-ives of the study were to provide a quantitativedescriptidu of mating behaviour, to determinehow home ranges were established and toobserve the role of agonistic behaviour in estab-lishing home ranges .
Vernal behaviour is defined as the behaviourcharacteristice of the first 5 weeks post-hiber-nation. During this period mating and gestation(assuming a 30-day gestation) . occur. Becauseprecise reproductive, nutritional and other bio-logical data are unavailable, an ecological group-ing of behavioural patterns is followed ratherthan one based on assumed biological para-meters .
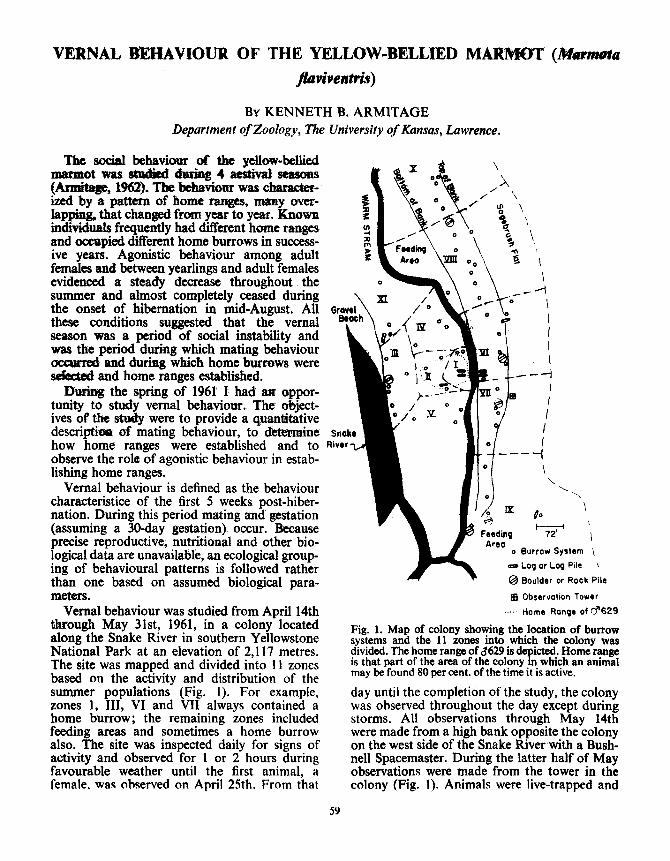
Vernal behaviour was studied from April 14ththrough May 31st, 1961, in a colony locatedalong the Snake River in southern YellowstoneNational Park at an elevation of 2,117 metres .The site was mapped and divided into 11 zonesbased on the activity and distribution of thesummer populations (Fig. 1). For example,zones 1, III, VI and VII always contained ahome burrow ; the remaining zones includedfeeding areas and sometimes a home burrowalso. The site was inspected daily for signs ofactivity and observed for 1 or 2 hours duringfavourable weather until the first animal, afemale, was observed on April 25th. From that
59
o Burrow System ~
® Log or Log Pile
Boulder or Rock Pile
le Observation Tower
Home Range of d'S29
Fig . 1 . Map of colony showing the location of burrowsystems and the 11 zones into which the colony wasdivided . The home range of x629 is depicted . Home rangeis that part of the area of the colony m which an animalmay be found 80 per cent . of the time it is active,day until the completion of the study, the colonywas observed throughout the day except duringstorms. All observations through May 14thwere made from a high bank opposite the colonyon the west side of the Snake River with a Bush-nell Spacemaster . During the latter half of Mayobservations were made from the tower in thecolony (Fig . 1). Animals were live-trapped and
60
ANIMAL BEHAVIOUR, XIII, I
colour marked for identification . A total of 202hours were spent in observation subsequent tothe appearance of the first animal .This study was supported by NSF Grant
G-16354. Mr. Hugh B. House assisted with fieldobservations. The National Park Service pro-vided living quarters and transportation intothe snowbound park . Our deepest appreciationto the many personnel of Grant Teton andYellowstone National Parks who made thisstudy possible.
Activity PatternsThe first animal, X611 was observed at the
log pile in zone I about 9 days after the vegeta-tion began to green-up . She was cautious, atelittle, pumped her tail frequently, but did notcry, even when we approached her. The secondmarmot, x629, was observed late the next after-noon at a burrow in zone VI . He went directlyto the log pile in zone I . The animals copulatedthere during the next morning and were notseen during the afternoon . By 1st May therewere sufficient animals to determine a populationactivity-cycle .
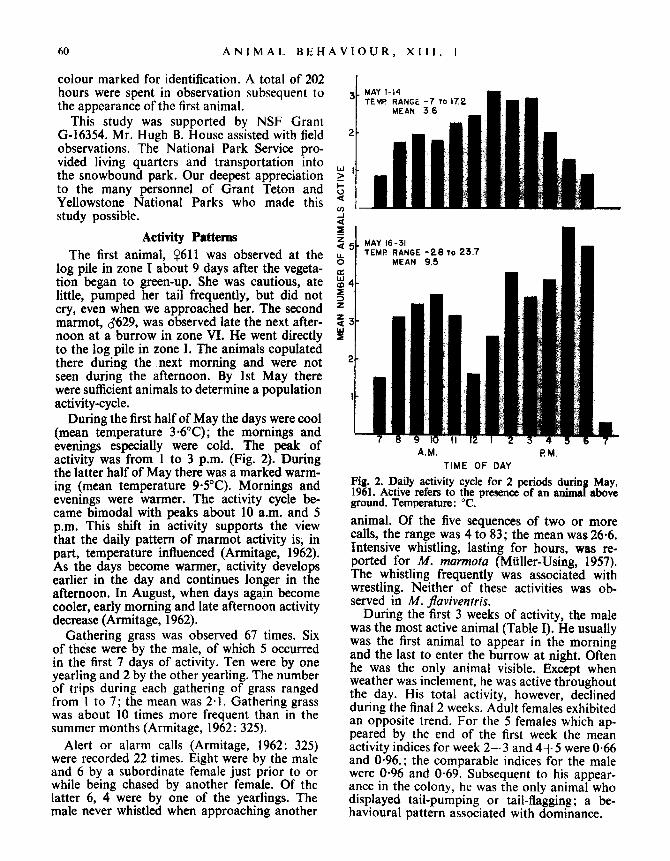
During the first half of May the days were cool(mean temperature 3 .6°C) ; the mornings andevenings especially were cold . The peak ofactivity was from 1 to 3 p.m. (Fig . 2). Duringthe latter half of May there was a marked warm-ing (mean temperature 9 .5°C). Mornings andevenings were warmer. The activity cycle be-came bimodal with peaks about 10 a .m. and 5p.m. This shift in activity supports the viewthat the daily pattern of marmot activity is, inpart, temperature influenced (Armitage, 1962) .As the days become warmer, activity developsearlier in the day and continues longer in theafternoon. In August, when days again becomecooler, early morning and late afternoon activitydecrease (Armitage, 1962) .
Gathering grass was observed 67 times . Sixof these were by the male, of which 5 occurredin the first 7 days of activity . Ten were by oneyearling and 2 by the other yearling . The numberof trips during each gathering of grass rangedfrom I to 7 ; the mean was 2-l . Gathering grasswas about 10 times more frequent than in thesummer months (Armitage, 1962 : 325) .
Alert or alarm calls (Armitage, 1962 : 325)were recorded 22 times . Eight were by the maleand 6 by a subordinate female just prior to orwhile being chased by another female . Of thelatter 6, 4 were by one of the yearlings . Themale never whistled when approaching another
3
2
I
I
MAY 1-14TEMP RANGE -7 To 172
MEAN 3 .6
MAY 16-31TEMP RANGE -2 .8 To 23.7
MEAN 9.5
PMA. M .TIME OF DAY
Fig . 2 . Daily activity cycle for 2 periods during May,1961. Active refers to the presence of an animal aboveground . Temperature : °C .
animal. Of the five sequences of two or morecalls, the range was 4 to 83 ; the mean was 26 .6 .Intensive whistling, lasting for hours, was re-ported for M. marmota (Muller-Using, 1957) .The whistling frequently was associated withwrestling . Neither of these activities was ob-served in M. flaviventris .
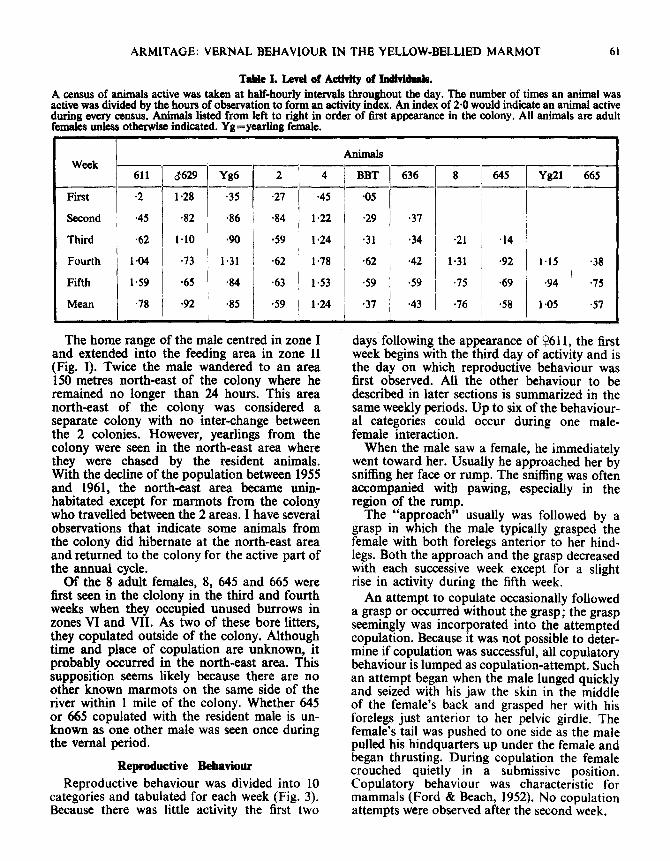
During the first 3 weeks of activity, the malewas the most active animal (Table I) . He usuallywas the first animal to appear in the morningand the last to enter the burrow at night . Oftenhe was the only animal visible. Except whenweather was inclement, he was active throughoutthe day. His total activity, however, declinedduring the final 2 weeks . Adult females exhibitedan opposite trend . For the 5 females which ap-peared by the end of the first week the meanactivity indices for week 24-3 and 4±5 were 0 .66and 0 .96 . ; the comparable indices for the malewere 0 .96 and 0 .69. Subsequent to his appear-ance in the colony, he was the only animal whodisplayed tail-pumping or tail-flagging ; a be-havioural pattern associated with dominance,
ARMITAGE: VERNAL BEHAVIOUR IN THE YELLOW-BELLIED MARMOT
61
Table I. Level of Activity of Individuals .A census of animals active was taken at half-hourly intervals throughout the day . The number of times an animal wasactive was divided by the hours of observation to form an activity index . An index of 2 . 0 would indicate an animal activeduring every census. Animals listed from left to right in order of first appearance in the colony . All animals are adultfemales unless otherwise indicated . Yg=yearling female .
The home range of the male centred in zone Iand extended into the feeding area in zone II(Fig . I). Twice the male wandered to an area150 metres north-east of the colony where heremained no longer than 24 hours . This areanorth-east of the colony was considered aseparate colony with no inter-change betweenthe 2 colonies . However, yearlings from thecolony were seen in the north-east area wherethey were chased by the resident animals .With the decline of the population between 1955and 1961, the north-east area became unin-habitated except for marmots from the colonywho travelled between the 2 areas . I have severalobservations that indicate some animals fromthe colony did hibernate at the north-east areaand returned to the colony for the active part ofthe annual cycle .
Of the 8 adult females, 8, 645 and 665 werefirst seen in the clolony in the third and fourthweeks when they occupied unused burrows inzones VI and VII . As two of these bore litters,they copulated outside of the colony . Althoughtime and place of copulation are unknown, itprobably occurred in the north-east area . Thissupposition seems likely because there are noother known marmots on the same side of theriver within I mile of the colony . Whether 645or 665 copulated with the resident male is un-known as one other male was seen once duringthe vernal period .
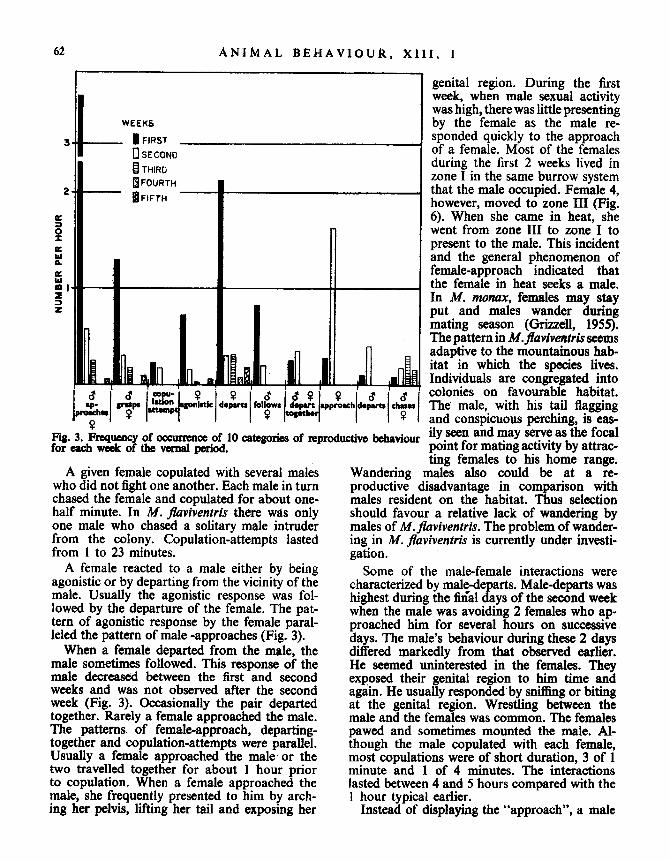
Reproductive BehaviourReproductive behaviour was divided into 10
categories and tabulated for each week (Fig . 3) .Because there was little activity the first two
days following the appearance of Y611, the firstweek begins with the third day of activity and isthe day on which reproductive behaviour wasfirst observed. All the other behaviour to bedescribed in later sections is summarized in thesame weekly periods. Up to six of the behaviour-al categories could occur during one male-female interaction .
When the male saw a female, he immediatelywent toward her. Usually he approached her bysniffing her face or rump . The sniffing was oftenaccompanied with pawing, especially in theregion of the rump .
The "approach" usually was followed by agrasp in which the male typically grasped thefemale with both forelegs anterior to her hind-legs. Both the approach and the grasp decreasedwith each successive week except for a slightrise in activity during the fifth week .
An attempt to copulate occasionally followeda grasp or occurred without the grasp ; the graspseemingly was incorporated into the attemptedcopulation . Because it was not possible to deter-mine if copulation was successful, all copulatorybehaviour is lumped as copulation-attempt . Suchan attempt began when the male lunged quicklyand seized with his jaw the skin in the middleof the female's back and grasped her with hisforelegs just anterior to her pelvic girdle . Thefemale's tail was pushed to one side as the malepulled his hindquarters up under the female andbegan thrusting. During copulation the femalecrouched quietly in a submissive position .Copulatory behaviour was characteristic formammals (Ford & Beach, 1952) . No copulationattempts were observed after the second week .
AnimalsWeek
611 3629 Yg6 2 4
BBT 636 8 645 Yg21 665
First 2 1 .28 3 5 27 4 5 -05
Second 4 5 -82 86 84 1 .22 2 9 37
Third 6 2 1 . 10 90 -59 1 .24 3 1 -34 2 1 14
Fourth 1 .04 -73 1 . 31 62 1 .78 -62 42 1 . 31 92 1 . 15
•3 8
Fifth 1 . 59 65 84 63 1 .53 59 -59 75 69 94 •7 5
Mean 7 8 -92
•85 59I
1 .24 •37 4 3 76 58 1 .05
I
•5 7
11
!prwehal 1w
Fig. 3 . Frequency of occurrence of 10 categories of reproductive behaviourfor each week of the vernal period.
A given female copulated with several maleswho did not fight one another. Each male in turnchased the female and copulated for about one-half minute. In M. flaviventris there was onlyone male who chased a solitary male intruderfrom the colony. Copulation-attempts lastedfrom 1 to 23 minutes .A female reacted to a male either by being
agonistic or by departing from the vicinity of themale. Usually the agonistic response was fol-lowed by the departure of the female . The pat-tern of agonistic response by the female paral-leled the pattern of male -approaches (Fig . 3) .
When a female departed from the male, themale sometimes followed . This response of themale decreased between the first and secondweeks and was not observed after the secondweek (Fig. 3). Occasionally the pair departedtogether. Rarely a female approached the male .The patterns. of female-approach, departing-together and copulation-attempts were parallel .Usually a female approached the male or thetwo travelled together for about 1 hour priorto copulation. When a female approached themale, she frequently presented to him by arch-ing her pelvis, lifting her tail and exposing her
o-I i P~adonhtie I departs
ANIMAL BEHAVIOUR, X111, I
"ARII%1lCT
C?ws p M'"pr~owhldn~ I chases
genital region. During the firstweek, when male sexual activitywas high, there was little presentingby the female as the male re-sponded quickly to the approachof a female . Most of the femalesduring the first 2 weeks lived inzone I in the same burrow systemthat the male occupied . Female 4,however, moved to zone III (Fig .6). When she came in heat, shewent from zone III to zone I topresent to the male. This incidentand the general phenomenon offemale-approach indicated thatthe female in heat seeks a male .In M. monax, females may stayput and males wander duringmating season (Grizzell, 1955) .The pattern in Ms flaviventris seemsadaptive to the mountainous hab-itat in which the species lives .Individuals are congregated into
Icolonies on favourable habitat .The male, with his tail flaggingand conspicuous perching, is eas-ily seen and may serve as the focalpoint for mating activity by attrac-ting females to his home range .
Wandering males also could be at a re-productive disadvantage in comparison withmales resident on the habitat . Thus selectionshould favour a relative lack of wandering bymales of M. flaviventris . The problem of wander-ing in M. flaviventris is currently under investi-gation .
Some of the male-female interactions werecharacterized by male-departs. Male-departs washighest during the final days of the second weekwhen the male was avoiding 2 females who ap-proached him for several hours on successivedays. The male's behaviour during these 2 daysdiffered markedly from that observed earlier .He seemed uninterested in the females. Theyexposed their genital region to him time andagain. He usually responded by sniffing or bitingat the genital region . Wrestling between themale and the females was common . The femalespawed and sometimes mounted the male. Al-though the male copulated with each female,most copulations were of short duration, 3 of 1minute and 1 of 4 minutes. The interactionslasted between 4 and 5 hours compared with theI hour typical earlier.Instead of displaying the "approach", a male
62
WEEKS
I FIRSTD SECOND® THIRDFOURTH
@FIFTH
0X
WW

ARMITAGE: VERNAL $EHAVIOUR IN THE YELLOW-BELLIED MARMOT
63
sometimes chased a female. Chases were un-common during the first week, increased in thesecond and third weeks while "approaches"were decreased. Chases and "c;-approaches"were of equal frequency in the third week . Theinverse relationship between these two activitiesindicates that reproductive behaviour shiftedtoward agonistic behaviour as the season pro-gressed. Chases declined and "S-approaches andgrasps were missing in the fourth week ; thesechanges were associated with a general declinein male activity (Table I) . Chases were missingin the fifth week while "a-approaches" andgrasps occurred infrequently (Fig . 3). Theactivity of the male continued to decline duringearly summer and several observations indicatedthat he became subordinate to the dominantfemale.
The response of the females to being chasedby the male was to depart . Except for the firstweek, the pattern of female-departs is almostidentical to the pattern of male-chases and ofa-approaches of the fifth week .
Thus the agonistic response of a female wasdirected toward a sexual advance of the male,but the agonistic behaviour of the male was metby avoidance .
Agonistic BehaviourAgonistic behaviour was observed in the first
week between a yearling and the adults, the firstday the yearling was observed. The number ofinteractions between the yearling and adultsincreased until the end of the third week ofactivity . During this time the yearling centredher activities in zone I (Figs . 5, 6, 7), but began toextend her home range into other zones (Figs.
6, 7). The yearling occasionally resisted theaggression of the adults, especially that of themale. This resistance reached a peak early in thethird week (Table II). By the end of the thirdweek, the yearling shifted her activity to includezone VI and began to visit zone XI (Fig . 7)Interactions decreased in frequency .
Yearling 21 appeared at the beginning of thefourth week. The frequency of interactionsbetween adult females and yearlings increased .Seventy-two per cent . of the agonistic behaviourbetween adult females and yearlings involvedthe new yearling . During this week yearling Y21wandered widely (Fig . 8) and was chased one ormore times by each of the 5 females living westof the warm stream . Yearling Y6 shifted herhome range to the northern edge of the colony(Fig. 8) and had fewer agonistic interactionswith the adult females . During the fifth week,both yearlings occupied home ranges on thenorthern edge of the colony (Fig. 9). A few con-flicts with adults occurred when the yearlingsoccasionally wandered beyond their home ranges .Most of the conflicts occurred in zones IV andVIII where the home ranges of the yearlingsoverlapped home ranges of adult females.Agonistic behaviour between yearlings andadults decreased and was evenly divided be-tween the 2 yearlings .
During the last 2 weeks there was no resist-ance of females and only minor resistance of themale by the yearlings. All agonistic behaviour be-tween adult females and yearlings during these2 weeks was chases (Table II) .Agonistic behaviour among adult females
was observed first near the middle of the firstweek. During the first week all females used the
Table II. Agonistic Behaviour Between Yearlings and Adults.Total interactions include both the subtle responses, such as avoidance behaviour, and aggressive actions, such as chases .Chases also are recognized separately.
WeekNumber ofyearlings
Interactions with maleNo./hr./yearling
Interactions with femalesNo./hr./yearling/y
Totalinteractions Chases
Yearlingresists
Totalinteractions Chases
Yearlingresists
First 1 235 039 0 -107 102 • 0
Second 1 674 070 070 139 098 008
Third 1 1 . 187 844 1 88 038 025 006
Fourth 2 071 0 -023 -062 -062 0
Fifth 2 016 0 0 027 027
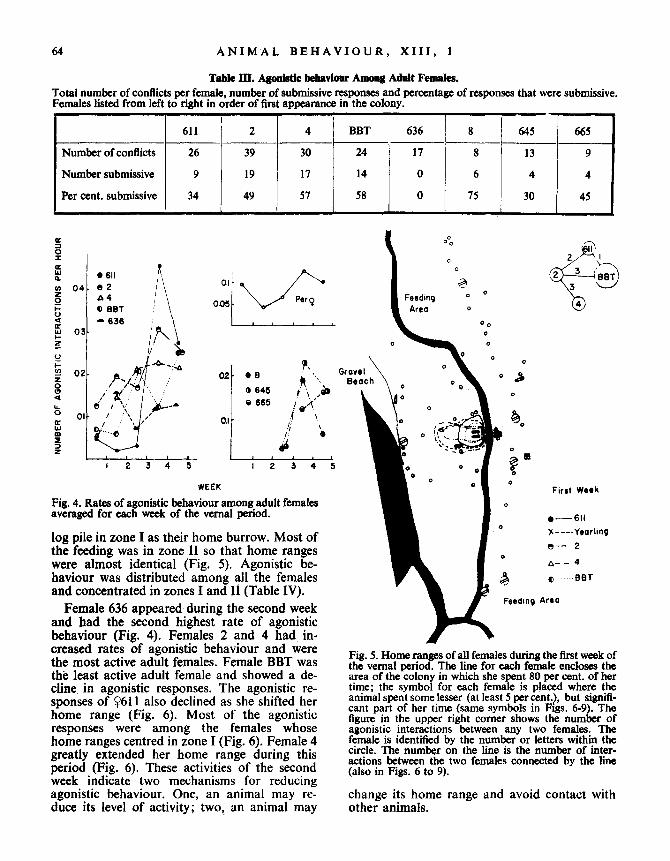
64
Table III. Agonistic behaviour Among Adult Females.Total number of conflicts per female, number of submissive responses and percentage of responses that were submissive .Females listed from left to right in order of first appearance in the colony .
U0
mWm
7z
01
ANIMAL BEHAVIOUR, XIII, 1
0.1
0.05
02
0.1
6
0 645o 665
3 4
WEEKFig. 4 . Rates of agonistic behaviour among adult femalesaveraged for each week of the vernal period .
log pile in zone I as their home burrow . Most ofthe feeding was in zone 11 so that home rangeswere almost identical (Fig . 5). Agonistic be-haviour was distributed among all the femalesand concentrated in zones I and II (Table IV) .
Female 636 appeared during the second weekand had the second highest rate of agonisticbehaviour (Fig. 4). Females 2 and 4 had in-creased rates of agonistic behaviour and werethe most active adult females . Female BBT wasthe least active adult female and showed a de-cline in agonistic responses . The agonistic re-sponses of Y611 also declined as she shifted herhome range (Fig . 6). Most of the agonisticresponses were among the females whosehome ranges centred in zone I (Fig . 6) . Female 4greatly extended her home range during thisperiod (Fig . 6). These activities of the secondweek indicate two mechanisms for reducingagonistic behaviour . One, an animal may re-duce its level of activity ; two, an animal may
a
5
00 0
00
0
0
First Week
3
3
Fig. 5. Home ranges of all females during the first week ofthe vernal period . The line for each female encloses thearea of the colony in which she spent 80 per cent . of hertime ; the symbol for each female is placed where theanimal spent some lesser (at least 5 percent .), but signifi-cant part of her time (same symbols in Figs . 6-9) . Thefigure in the upper right corner shows the number ofagonistic interactions between any two females . Thefemale is identified by the number or letters within thecircle . The number on the line is the number of inter-actions between the two females connected by the line(also in Figs. 6 to 9) .
change its home range and avoid contact withother animals .
611 2 4 BBT
636
8 11
645 665
Number of conflicts 26 39 30 24 17 8 13 9
Number submissive 9 19 17 14 0 6 4 4
Per cent. submissive 34 49 57 58 0 75 30 45
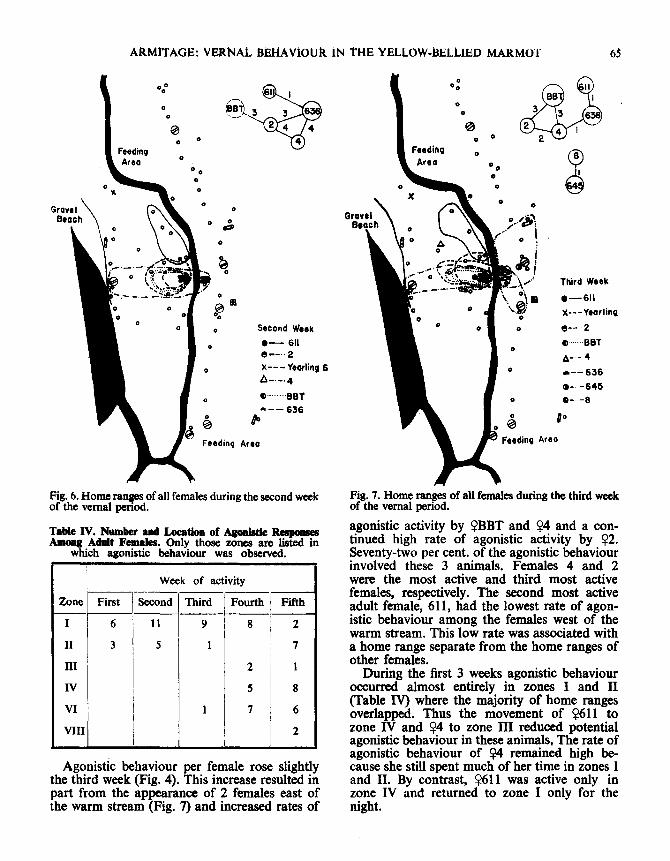
ARMITAGE: VERNAL BEHAVIOUR IN THE YELLOW-BELLIED MARMOT
65
a
0
Fig . 6. Home ranges of all females during the second weekof the vernal period .
Table IV. Number and Location of Agonistic RespomesAmong Adult Females. Only those zones are listed in
which agonistic behaviour was observed .
Agonistic behaviour per female rose slightlythe third week (Fig . 4). This increase resulted inpart from the appearance of 2 females east ofthe warm stream (Fig. 7) and increased rates of
oa 0
Third Week
a
•- 611
\
X---Yearling0o
e-- 2n "BBTO- 4-- 636
0---645Q__8
Fig . 7 . Home ranges of all females during the third weekof the vernal period .
agonistic activity by ?BBT and ?4 and a con-tinued high rate of agonistic activity by ?2 .Seventy-two per cent. of the agonistic behaviourinvolved these 3 animals . Females 4 and 2were the most active and third most activefemales, respectively. The second most activeadult female, 611, had the lowest rate of agon-istic behaviour among the females west of thewarm stream . This low rate was associated witha home range separate from the home ranges ofother females .
During the first 3 weeks agonistic behaviouroccurred almost entirely in zones I and II(Table IV) where the majority of home rangesoverlapped. Thus the movement of ?611 tozone IV and 94 to zone III reduced potentialagonistic behaviour in these animals, The rate ofagonistic behaviour of 94 remained high be-cause she still spent much of her time in zones 1and II. By contrast, X611 was active only inzone IV and returned to zone I only for thenight .
Week of activity
Zone First Second Third Fourth Fifth
I 6 11 9 8 2
II 3 5 1 7
III 2 1
IV 5 8
VI l 1 7 ~
6
VIII 2i
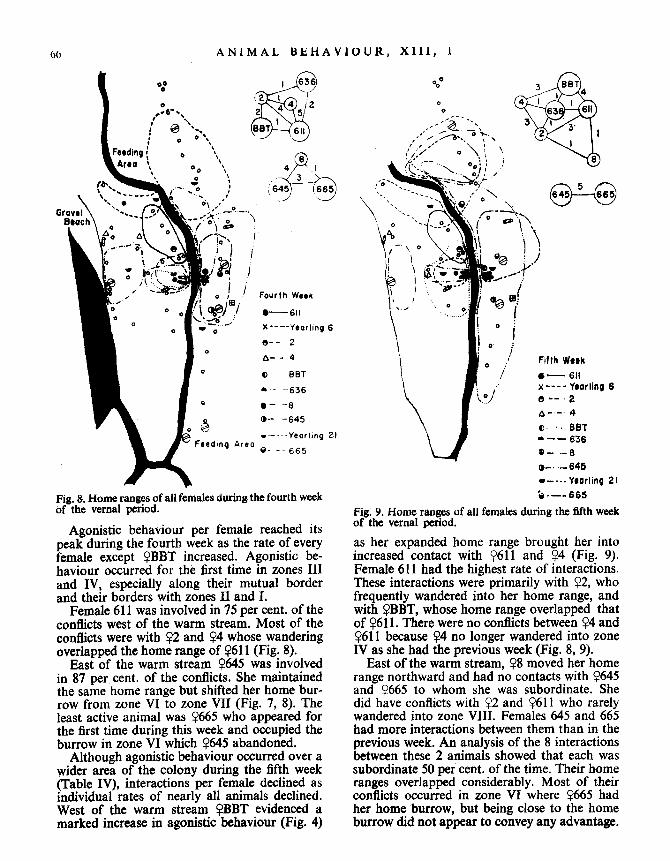
66 ANIMAL BEHAVIOUR, XIII, l
2 5
Fig . &. Home ranges of all females during the fourth weekof the vernal period .
Agonistic behaviour per female reached itspeak during the fourth week as the rate of everyfemale except YBBT increased . Agonistic be-haviour occurred for the first time in zones IIIand IV, especially along their mutual borderand their borders with zones II and I .
Female 611 was involved in 75 per cent . of theconflicts west of the warm stream . Most of theconflicts were with 92 and Y4 whose wanderingoverlapped the home range of Y611 (Fig . 8) .
East of the warm stream Y645 was involvedin 87 per cent. of the conflicts . She maintainedthe same home range but shifted her home bur-row from zone VI to zone VII (Fig. 7, 8). Theleast active animal was ?665 who appeared forthe first time during this week and occupied theburrow in zone VI which Y645 abandoned .
Although agonistic behaviour occurred over awider area of the colony during the fifth week(Table IV), interactions per female declined asindividual rates of nearly all animals declined .West of the warm stream ?BBT evidenced amarked increase in agonistic behaviour (Fig. 4)
00
00
88T
665
Fifth Weeke-611x---- Yearling 68 _2
a-- 4o , • BBT-636636e- -B
._645 - - •- Yearling 21~ •--665
Fig. 9. Home ranges of all females during the fifth weekof the vernal period.
as her expanded home range brought her intoincreased contact with Y611 and Y4 (Fig . 9) .Female 611 had the highest rate of interactions .These interactions were primarily with ?2, whofrequently wandered into her home range, andwith ?BBT, whose home range overlapped thatof ?611. There were no conflicts between ?4 and?611 because ?4 no longer wandered into zoneIV as she had the previous week (Fig. 8, 9) .
East of the warm stream, ?8 moved her homerange northward and had no contacts with ?645and ?665 to whom she was subordinate. Shedid have conflicts with Y2 and Y611 who rarelywandered into zone VIII . Females 645 and 665had more interactions between them than in theprevious week. An analysis of the 8 interactionsbetween these 2 animals showed that each wassubordinate 50 per cent . of the time. Their homeranges overlapped considerably. Most of theirconflicts occurred in zone VI where ?665 hadher home burrow, but being close to the homeburrow did not appear to convey any advantage .

ARMITAGE - VERNAL BEHAVIOUR IN THE YELLOW-BELLIED MARMOT
Field notes indicate that in any particular con-flict the animal that surprised the other seemedto emerge dominant .
Agonistic behaviour continued to declineduring the summer (Armitage, 1962 : 329). Asimilar pattern of a peak of agonistic behaviourfollowing hibernation and a gradual declinethereafter occurs in M. monax (Bronson, 1961) .
Several general conclusions may be drawnfrom the detailed descriptions of agonistic be-haviour .
Dominance by a female did not depend uponearly seasonal appearance in the colony . Thus,9636, dominant west of the warm stream, wasthe fifth animal to appear (Table III) .
Female 636, with no submissive responses,was five years old ; females 645, 611, and 665,with 30 per cent ., 34 per cent. and 45 per cent.submissive responses respectively, were 4 yearsold. The other 4 females were either 2 or 3 yearsold (ages not definitely known) and had from 49per cent . to 75 per cent. submissive responses .The yearlings had 100 per cent . submissiveresponses .
There was a correlation between activity andagonistic behaviour. The correlation coefficient(r) between activity index (Table I) and rate ofagonistic behaviour (Fig . 4) was 0 .571 (p= •001) .Such a correlation means that the more time ananimal spends above ground the more likely itwill come into contact with another animal .Thus an animal can reduce its conflicts either byreducing its level of activity or changing itsplace or time of activity to avoid conflicts .
There was a postive correlation (r=0 .682,.05>p>-02) between mean activity index(Table I) and per cent . of submissive responses(Table 111) for all females . Thus subordinateanimals tended to be more active . However, ifadult females only are considered, there wasonly a slight positive correlation (r=0 . 376, n.s.) .The treatment of the highly active and subordin-ate yearlings in the analysis markedly affectsthe significance of the relationship . Obviouslyfactors other than social rank affect an animal'sactivity and adult females may use means otherthan modifying activity to reduce agonistic be-haviour .
During the vernal period there was a generalmoving out of animals from the centre of con-centration in zone I to other areas of the colony .Agonistic behaviour was reduced by this chang-ing of home ranges . After ?4 centred her activ-ities in zone III (Fig, 7-9), her rate of conflictslevelled off and began to decline (Fig. 4).
67
When ?8 moved from zone VI to zone VIII(Fig. 8, 9), her rate of conflict dropped to thelowest in the colony (Fig. 4). Females BBTand 2 remained in zone I and had continuallyhigh rates of conflict (Fig . 4) . These two femaleswere subordinate to ?636, who harassed themcontinually, and finally left the colony and spentmost of the summer in the area 150 metres to thenorth-east .
Agonistic behaviour occurred between nearestneighbours. As the animals dispersed from zoneI, agonistic behaviour occurred in more areas ofthe colony (Table IV) . There were no interactionsbetween the two groups of marmots, east andwest of the warm stream, until the fifth weekwhen there was some mingling in zone VIII (Fig .9). The patterns of interactions between ?611and other females, described previously, areespecially illustrative (Fig. 5-9) .
Agonistic behaviour may also have affectedreproduction. At least 4 of the females west of thehot stream copulated . Three of these appearedto be pregnant and to give birth, but the youngof only ?636 appeared above ground . The be-haviour of the subordinate animals is typifiedby ?611 who lived in the same burrow systemwith ?636. Female 611 appeared earlier in theday and went north (Fig. 5-7) where she spentmost of the day. She was observed numeroustimes during the day attempting to return to thehome burrow, but if Y636 were visible, sheturned and ran north . Many times there was noindication that ?636 was aware of her activity .She appeared to give birth in late May, but shemay not have been able to care for her youngproperly or the young may have been destroyed .Loss of litters under social stress was reportedfor M. monax (Grizzell, 1955), Mus musculus(Southwick, 1955) and Rattus norvegicus (Cal-houn, 1950). In Rattus, the subordinate femaleshad fewer litters . There is no direct evidence thatbehaviour caused reproductive loss amongmarmots, but the evidence is indicative. Onlythose females subordinate to ?636 failed tolitter ; by contrast Y645 and ?665, on the eastside of the hot creek, littered. The evidence iscircumstantial, but apparently a highly aggress-ive marmot can affect reproduction by sub-ordinate females .
SummaryBehaviour of a colony of yellow-bellied mar-
mots was observed for 5 weeks following emerg-ence from hibernation . The colony consistedof a male, 8 adult females and 2 yearlingfemales .

6 8
Reproductive behaviour was centred in thefirst 2 weeks. The male evidenced a gradualdecline in reproductive behaviour throughout theperiod of observation. Agonistic behaviour to-ward females increased during this decline untilthe last week when it also declined .
All adults were agonistic to yearlings . Therate of agonistic behaviour per female per hourwas highest during the 4th week . There was asignificant positive correlation between activityof adult females and rate of agonistic behaviourand between activity of all females and per cent .of submissive responses. Older animals had alower percentage of submissive responses thanyounger animals . Agonistic behaviour occurredbetween nearest neighbours . The pattern ofnearest neighbour responses changed during thevernal period as females moved from being con-centrated in the centre of the colony to beingmore widely dispersed . The dispersal wasassociated with changes in home ranges andwith the avoidance of dominant animals bysubordinate animals .
Circumstantial evidence indicated that agon-istic behaviour affected reproductive success ofsubordinate animals .
REFERENCESArmitage, K . B. (1962). Social behaviour of a colony
of the yellow-bellied marmot . Anim. Behav., 10,319-331 .
Bronson, F. H . (1961) . An investigation of some aspectsof social pressure in woodchucks . Ph.D. Thesis,The Pennyslvania State University .
Calhoun, J . B . (1950) . The study of wild animals undercontrolled conditions . Ann. N.Y. Acad. Sci., 51,1113-1122 .
Ford, C. S. & Beach, F. A . (1952) . Patterns of sexualbehaviour . New York: Harper and Brothers. 307PP .
ANIMAL BEHAVIOUR, X111, 1
Grizzell, R . A ., Jr. (1955) . A study of the southern wood-chuck, Marmota monax monax . Amer. Midi. Nat.,53, 257-293 .
Muller-Using, D . (1957) . Die Paarungsbiologie desMurmeltieres . Z. Jagdwissensch ., 3, 24-28 .
Southwick, C . H. (1955) . Regulatory mechanisms ofhouse mouse populations : social behavior affect-ing litter survival . Ecology, 36, 627-634 .
(Accepted for publication 12th September, 1964 ;Ms. number : 498) .
AbstractReproductive and agonistic behaviour were
studied in the first 5 weeks post-hibernation.A single male, 5 adult females and one yearlingfemale formed the colony during the first 2weeks when reproductive behaviour occurred .Three adult females and a second yearling femalejoined the colony in the third and fourth weeks .The male evidenced a decline in reproductivebehaviour throughout the 5 weeks. All adultswere agonistic to yearlings. The yearlings did notevidence reproductive behaviour . The rate ofagonistic behaviour per female per hour washighest during the fourth week when parturitionprobably occurred . There was a significant posit-ive correlation between activity of adult femalesand rate of agonistic behaviour and betweenactivity of all females and per cent . of submissiveresponses. Older animals had a power percentageof submissive responses than younger animals .Agonistic behaviour occurred between nearestneighbours . During the vernal period, femalesmoved from being concentrated in the centreof the colony to being more widely dispersed .The dispersal was associated with changes inhome ranges and with the avoidance of domin-ant animals by subordinate animals . Sub-ordinate animals failed to raise litters .





























