Embed Size (px)
Citation preview

Universidade São Judas Tadeu
Programa de Pós-Graduação Stricto Sensu em Educação Física
Paula Andréa Malveira Cavalcante
Contratilidade miocárdica de ratos submetidos a protocolo de treinamento
linear de natação: papel da intensidade do exercício
São Paulo
2015

1
Paula Andréa Malveira Cavalcante
Contratilidade miocárdica de ratos submetidos a protocolo de
treinamento linear de natação: papel da intensidade do exercício
Dissertação apresentada ao Programa de
Mestrado em Educação Física da Universidade
São Judas Tadeu como requisito parcial à
obtenção do título de Mestre em Educação Física.
Linha de pesquisa: Fenômeno Esportivo
Orientador: Prof. Dr. Danilo Sales Bocalini
São Paulo
2015

2
Ficha catalográfica elaborada pela Biblioteca da
Universidade São Judas Tadeu
Bibliotecária: Daiane Silva de Oliveira - CRB 8/8702
Cavalcante, Paula Andréa Malveira
C376c Contratilidade miocárdica de ratos submetidos a protocolo de
treinamento linear de natação: papel da intensidade do exercício / Paula
Andréa Malveira Cavalcante. - São Paulo, 2015.
81 f. : il. ; 30 cm.
Orientador: Danilo Sales Bocalini.
Dissertação (mestrado) – Universidade São Judas Tadeu, São Paulo, 2015.
1. Coração - Contração. 2. Miocárdio. 3. Natação - Treinamento. I. Bocalini,
Danilo Sales. II. Universidade São Judas Tadeu, Programa de Pós-Graduação
Stricto Sensu em Educação Física. III. Título CDD 22 – 613.7

3
AGRADECIMENTOS
Ao meu orientador, Professor Dr. Danilo Sales Bocalini pela confiança no meu
trabalho e suporte no decorrer do curso.
Ao querido Professor Dr. Aylton Figueira Jr. que desde a graduação me incentivou
e acreditou em mim.
Aos professores do curso de Mestrado da Universidade São Judas Tadeu pela
enriquecedora convivência neste período.
Aos meus amigos e familiares que sempre torceram por mim.

4
SUMÁRIO
LISTA DE ABREVIAÇÕES.......................................................................................... 06
LISTA DE FIGURAS.................................................................................................... 08
LISTA DE QUADROS.................................................................................................. 09
LISTA DE TABELAS.................................................................................................... 10
RESUMO...................................................................................................................... 11
ABSTRACT................................................................................................. 13
1. INTRODUÇÃO.................................................................................................... 15
1.1. Minha trajetória.................................................................................................... 15
1.2. Justificativa.......................................................................................................... 17
1.3. Objeto do estudo................................................................................................. 19
1.4. Objetivos.............................................................................................................. 20
1.4.1. Objetivo geral............................................................................................. 20
1.4.2. Objetivos específicos................................................................................. 20
1.5. Hipóteses............................................................................................................. 21
2. REVISÃO DE LITERATURA.............................................................................. 22
2.1. Adaptações cardíacas no exercício aeróbio: papel da modalidade.................... 22
2.2. Adaptações cardíacas no exercício aeróbio: papel da intensidade.................... 23
2.3. Avaliação da aptidão aeróbia em roedores.................................................... 29
2.4. Remodelamento cardíaco................................................................................... 30
2.5. Alterações na contratilidade miocárdica.............................................................. 39
3. MATERIAIS E MÉTODOS.................................................................................. 47
3.1. Animais................................................................................................................ 47
3.2. Protocolo de exercício físico................................................................................ 47
3.3. Experimento 1 - Avaliações “in vivo”................................................................... 49
3.3.1. Avaliação da capacidade física.................................................................. 49
3.3.2. Ecodoplercardiograma............................................................................... 50
3.4. Experimento 2 - Avaliação “pós morten”............................................................. 51
3.4.1. Biometria das massas cardíacas............................................................... 51
3.4.2. Avaliação da mecânica miocárdica............................................................ 51
3.5. Análise estatística................................................................................................ 53
4. DESENHO EXPERIMENTAL.............................................................................. 54
5. RESULTADOS.................................................................................................... 55
6. DISCUSSÃO....................................................................................................... 64

5
7. CONCLUSÃO...................................................................................................... 74
8. REFERÊNCIAS................................................................................................... 75
9. COMITÊ DE ÉTICA............................................................................................. 93

6
LISTA DE ABREVIAÇÕES
AS Área de secção transversa do músculo papilar
AT Átrios
C Coração
CS citrato sintase
CT Carga de treino
DC Débito cardíaco
DD Diâmetro diastólico
DDfVE Diâmetro diastólico final do ventrículo esquerdo
dP/dt Primeira derivada temporal da pressão intraventricular
dT/dt Primeira derivada temporal da tensão
DS Diâmetro sistólico
DSfVE Diâmetro sistólico final do ventrículo esquerdo
DSVE Diâmetro sistólico do ventrículo esquerdo
ERP Espessura sistólica e diastólica da parede ventricular.
EPAD Espessura da parede anterior na diástole
EPPD Espessura da parede posterior na diástole
FC Frequência cardíaca
FEDT Fração de encurtamento do diâmetro
Lmáx Comprimento máximo
MCP Miosina de cadeia pesada
MOD Grupo de treinamento moderado
MVE Massa do ventrículo esquerdo
PA Pressão arterial
PC Peso corpóreo
PGC-1α Proteína peroxissoma ativado pelo proliferador do receptor gama
coativador 1-alfa
PLB Fosfolamban
PLB-Tre17 Fosfolamban fosforilado na treonina 17
RC Remodelamento cardíaco
RS Retículo sarcoplasmático
SED Grupo controle
Serca2a Ca2+-ATPase
INT Grupo de treinamento intenso

7
TD Tensão isométrica desenvolvida
TPT Tempo para TD atingir o seu valor máximo
TR Tensão isométrica de repouso ou diastólica
TS Tempo total da sessão
TT Trabalho total do programa de treino
VD Ventrículo direito
VE Ventrículo esquerdo
VO2máx Consumo máximo de oxigênio
VO2pico Consumo de oxigênio de pico

8
LISTA DE FIGURAS
Figura 1. Resultado da busca de artigos científicos na PubMed
Figura 2. Resultado da busca de artigos científicos na PubMed
Figura 3. Concentrações de lactato sanguíneo de ratos sedentários e treinados
com natação
Figura 4. Esquema das vias de sinalização envolvidas na indução da hipertrofia
cardíaca fisiológica e patológica
Figura 5. Mecanismo da hipertrofia
Figura 6. Fluxo do cálcio celular
Figura 7. Fases de treinamento
Figura 8. Desenho experimental do estudo
Figura 9. Carga de treino
Figura 10. Tempo de treino
Figura 11. Trabalho total
Figura 12. Comparação da intensidade, tempo e trabalho total dos grupos de
treinamento
Figura 13. Teste de capacidade física
Figura 14A. Aumento da TD no tempo de pausa
Figura 14B. Área abaixo da curva
Figura 15A. % do Lmax da TD
Figura 15B. Coeficiente angular da TD
Figura 16A. % do Lmax da TR
Figura 16B. Constante de rigidez da TR

9
LISTA DE QUADROS
Quadro 1. Adaptações cardiovasculares relacionadas ao treinamento
aeróbio...........17

10
LISTA DE TABELAS
Tabela 1. Massa corporal inicial e final dos grupos
Tabela 2. Parâmetros ecocardiográficos
Tabela 3. Parâmetros biométricos
Tabela 4. Parâmetros biométricos dos músculos papilares
Tabela 5. Parâmetros da contratilidade miocárdica

11
RESUMO
Introdução: O treinamento físico aeróbio exerce influências fisiológicas positivas
relacionadas ao remodelamento cardíaco, porém, pouco ainda se sabe sobre as
alterações provocadas pelas diferentes intensidades. Objetivo: investigar as alterações
estruturais e contráteis do miocárdio de ratas, após 12 semanas de treinamento de
natação com diferentes intensidades de esforço. Métodos: Trinta ratas livres de agentes
patogênicos foram randomizadas em três grupos: sedentário (SED), 10 animais
permaneceram alojados em repouso durante 12 semanas, treinamento moderado
(MOD), 10 animais foram submetidos a exercício físico de intensidade moderada
durante 12 semanas, treinamento intenso (INT), 10 animais foram submetidos a
exercício físico de alta intensidade durante 12 semanas. O protocolo de exercício físico
consistiu de natação. Após uma semana de adaptação, os animais do protocolo de
natação moderada treinaram com sobrecarga de 2% aderida a cauda do animal, já para
o protocolo de alta intensidade, o percentual de carga foi controlado de maneira linear
respeitando a seguinte ordem: 1º semana: sem carga; 2º semana: 5% do peso corporal,
da 3º a 6º semana: 6% do peso corporal, 7º a 8º semana: 7% do peso corporal e da 9º
a 12º semana: 8% do peso corporal fixado na cauda do animal. O treinamento foi
continuado por período total de 12 semanas, 5 dias por semana, 60 minutos/dia. Foram
analisadas as alterações dos parâmetros envolvidos no remodelamento cardíaco pela
avaliação da biometria, ecodoplercardiografia e contratilidade miocárdica. Resultados:
Após doze semanas de treinamento a capacidade física do grupo SED (141 ± 12 seg)
foi inferior (p< 0,001) aos demais grupos, que também diferiram (p < 0,01) entre si (MOD:
293 ± 10 vs. INT: 371 ± 20; seg). Todos os grupos aumentaram o peso corpóreo após
doze semanas, contudo, não foram encontradas diferenças entre os grupos. O peso dos
átrios e do VD dos grupos SED, MOD e INT não diferiram entre si, contudo, o peso tanto
do VE quanto do coração do grupo SED, foi menor que os grupos MOD e INT que
também diferiram entre si. Não foram encontradas diferenças entre os parâmetros
biométricos do músculo papilar. Em relação aos parâmetros ecocardiográficos, não
foram encontradas diferenças entre os grupos antes da randomização, mas após doze
semanas de treinamento, os grupos MOD e INT apresentaram bradicardia, aumento na
EPPD, DDfVE e DSfVE em relação ao grupo SED, sendo mais evidente no grupo INT.
Não foram encontradas diferenças EPAD e na FE. Os valores da TD, +dT/dt e da –dT/dt
do grupo SED (TD: 0,85 ± 0,4 g/mm2/mg; +dT/dt: 7 ± 0,97 g/mm2/mg/s; -dT/dt: 5 ± 0,34
g/mm2/mg) foram inferiores (p< 0,001) aos grupos MOD (TD: 1,07± 0,1 g/mm2/mg;
+dT/dt: 11 ± 1,07 g/mm2/mg/s; -dT/dt: 7 ± 0,52 g/mm2/mg) e INT (TD: 1,27± 0,7
g/mm2/mg; +dT/dt: 15 ± 0,59 g/mm2/mg/s; -dT/dt: 10 ± 1,06 g/mm2/mg) que também
diferiram (p< 0,01) entre si. Não foram encontradas diferenças entre os grupos na TR
(SED: 0,20 ± 0,05, MOD: 0,21 ± 0,03, Int: 0,20± 0,02; g/mm2/mg), no TPT (SED: 183±
6, MOD: 172 ± 9, INT: 163 ± 5; ms) e na TR50% (SED: 142± 9, MOD: 148 ± 10, INT: 147
± 4; ms). Foi possível visualizar o comportamento da TD após interrupções no tempo de
estímulos. A força desenvolvida aumentou conforme aumentou o intervalo de estimulo,
contudo, diferenças foram encontradas quando analisado o percentual de aumento
quando comparados a 120 segundos de intervalo. Um aumento de 52 3% foi
encontrado no grupo INT, estatisticamente diferente (p<0,03) do aumento externado
pelo grupo MOD. Adicionalmente, ambos os grupos treinados foram superiores ao grupo

12
SED que tiveram aumento de 32 4%. O desenvolvimento de força em todos os pontos
percentuais diferiu (p<0,01) entre os grupos MOD (92: 0,67 0,09, 94: 0,83 0,11, 96:
0,94 0,11, 98: 1,10 0,13, 100: 1,30 0,12; %) e Int (92: 0,89 0,12, 94: 1,06 0,13,
96: 1,21 0,14, 98: 1,38 0,15, 100: 1,57 0,11; %), contudo, ambos foram superiores
(p< 0,001) ao grupo SED (92: 0,34 0,05, 94: 0,48 0,07, 96: 0,67 0,11, 98: 0,75
0,12; 100: 0,93 0,13; %). Não foram encontradas diferenças entre os grupos em
relação aos pontos percentuais de Lmax da TR. Os valores dos coeficientes de correlação
linear (r) variaram entre 0,9897 para o grupo SED, 0,9910 para o grupo MOD e 0,9986
para o grupo INT não diferiram entre si. Quando se consideram tanto os coeficientes
angulares das retas de Frank-Starling quanto os coeficientes de rigidez da TR, não
foram encontradas diferenças entre os grupos SED (0,072 0,012 e 0,545 0,061;
g/mm2/mg/%Lmax), MOD (0,072 0,011 e 0,567 0,074; g/mm2/mg/%Lmax) e INT (0,083
0,003 e 0,0616 0,077 g/mm2/mg/% Lmax). Conclusão: O exercício aeróbio realizado
de forma crônica é um eficiente aliado nas adaptações cardiovasculares, em alta e
moderada intensidade, contudo, o treinamento físico aeróbio de alta intensidade foi mais
favorável para adaptações na aptidão cardiorrespiratória e na função cardíaca do que o
exercício de intensidade baixa a moderada.
Palavras chaves: contratilidade miocárdica; remodelamento cardíaco; treinamento com
natação.

13
ABSTRACT
The aerobic exercise has physiological positive influences related to cardiac remodeling,
however, little is known about the changes caused by different intensities. Objective:
The aim of this study was investigate the myocardial structural and contractile changes
in rats after 12 weeks of swimming training with different intensities of effort. Methods:
Thirty rats were randomized into three groups: sedentary (SED), moderate training
(MOD) and intense training (INT) swimming for 12 weeks. In moderate swimming
protocol, the animals were trained with overhead of 2%. In the high intensity protocol,
the load was controlled linearly in the following order: 1st week: without load; 2nd week:
5% of body weight, 3 to 6 week: 6% of body weight, 7 to 8 week: 7% of body weight and
9 to 12 week: 8% of body weight. The changes of the parameters involved were analyzed
in the evaluation of biometrics, ecodoplercardiografia and myocardial contractility.
Results: After twelve weeks of training the physical capacity of the SED group (141 ±
12 sec) was lower (p <0.001) to the other groups, which also differ (p <0.01) between
them (MOD: 293 ± 10 vs. INT: 371 ± 20; sec). All groups increased body weight after
twelve weeks, however, no differences were found between groups. The weight of the
atria and VD of the SED, MOD and INT did not differed, however, the weight of both,VE
and the heart in the group SED was lower than the MOD and INT groups and which also
differed from each other. No differences were found between the biometric parameters
of the papillary muscle. Regarding echocardiographic parameters, there were no
differences between the groups before randomization, but after twelve weeks of training,
the MOD group and INT presented bradycardia, increased EPPD, DDfVE and DSfVE,
being more evident in the group INT. It was possible to view of the TD behavior after
interruption in stimulus time. The force increased with increased stimulation interval,
however, differences were found when analyzed the percentage increase compared to
interval of 120 seconds. An increase of 52 ± 3% was found in the INT group, statistically
different (p <0.03) of MOD. Additionally, both the trained groups were higher than the
SED group with 32 ± 4%. The development of strength in all percentage points were
different (p <0.01) between the groups: MOD (92: 0,67 0,09, 94: 0,83 0,11, 96: 0,94
0,11, 98: 1,10 0,13, 100: 1,30 0,12; %) and INT (92: 0,89 0,12, 94: 1,06 0,13,
96: 1,21 0,14, 98: 1,38 0,15, 100: 1,57 0,11; %), however, both were higher (p
<0.001), to the SED group (92: 0,34 0,05, 94: 0,48 0,07, 96: 0,67 0,11, 98: 0,75
0,12; 100: 0,93 0,13; %). Conclusion: Aerobic exercise performed chronically is an
efficient ally in cardiovascular adaptations in high and moderate intensity, however, the

14
high-intensity aerobic training was more favorable for adaptations in cardiorespiratory
fitness and cardiac function than intensity exercise low to moderate.
Key words: myocardial contractility; cardiac remodeling; training swimming.

15
1. INTRODUÇÃO
1.1. Minha trajetória
Na minha concepção, o curso de Mestrado representa um enorme privilégio e
trata-se de uma oportunidade rica de amadurecimento profissional e pessoal.
Entretanto, seguindo a minha natureza aquariana e levando em consideração que tenho
sido um ser senciente, percebo que esse aprimoramento é uma etapa importante, mas
compõe apenas parte de um processo infindável na busca de conhecimento.
Hoje consigo perceber que a chave para encontrar o saber traduz-se de forma
simples na curiosidade, na humildade e na dedicação. Acredito que os instrumentos
necessários para buscar uma melhor compreensão do mundo e da natureza humana
estão ao alcance das minhas mãos e assemelham-se ao trabalho das formigas,
construído diariamente com muito esforço e determinação.
Esse presente trabalho foi uma grata oportunidade para refletir sobre a minha
construção profissional e um pretexto para recordar fatos da infância e adolescência
que de certa forma já apontavam nessa direção. Assim, de forma muito resumida,
compartilho aqui alguns parágrafos da minha trajetória.
No ensino fundamental eu costumava participar de todas as feiras de ciências e
os assuntos sobre o corpo humano já me despertavam interesse. Mais tarde, no ensino
médio no Mackenzie, escolhi a área de biológicas e acredito que dois fatos contribuíram
para que eu vislumbrasse a minha verdadeira vocação.
O primeiro deles foi no primeiro ano, quando me delegaram a tarefa de apresentar
um seminário sobre os sistemas do corpo humano. Confeccionar todos os desenhos em
cartolinas foi uma experiência prazerosa, mas muito trabalhosa. Naquela época, não
contávamos com o PowerPoint ou qualquer outro recurso tecnológico, então, os
desenhos eram feitos à mão para serem apresentados aos colegas de classe, de
preferência da forma mais didática possível. Então, além de desenhar os cartazes e
colá-los na lousa, tive a ideia de apresentar o “sangue venoso”, “sangue arterial”,
“glóbulo branco”, “glóbulo vermelho”, entre outros, como se fossem personagens
personificados, no intuito de tentar explicar de forma simples, um assunto tão complexo.
Os alunos conseguiram entender a matéria e lembro do contentamento da professora
com o resultado.
No último ano do ensino médio, fui agraciada com uma nota dez em biologia. Esse
fato marcou muito a minha adolescência, pois a professora Marilu contou para a turma

16
que após alguns anos lecionando biologia no Mackenzie, aquela teria sido a primeira
nota dez registrada na sua carreira.
Depois desse episódio, eu comecei a fazer as provas de biologia em frente à
parede e longe dos meus colegas. Era constrangedor, um exagero, mas esse foi o preço
que tive que pagar por me empenhar na disciplina que tanto me fascinava. Um ano
depois, encontrei a professora Marilu no campus do Mackenzie que mostrou-se
desapontada quando contei que havia iniciado o curso de Arquitetura.
Na época, a minha primeira escolha teria sido Medicina, mas na impossibilidade
de obter apoio financeiro, segui adiante com a segunda opção. Gostava de arte e
gostava de desenhar, mas isso não foi suficiente para permanecer na profissão de
arquiteta e anos mais tarde decidi pela Educação Física.
Partindo desse ponto, o gosto e interesse pela biologia aliou-se à curiosidade
sobre os efeitos da atividade física sobre a saúde. Percebi, então, que de certa forma
eu havia me tornado por analogia, uma suposta “médica” que hoje prescreve
tratamentos não medicamentosos.
Depois de graduada, iniciei uma capacitação profissional no Incor, ministrando
aulas de condicionamento físico para grupos especiais no setor de reabilitação cardíaca.
Concomitantemente, ingressei em uma especialização de “Musculação e
Condicionamento Físico” e o tema do meu trabalho de conclusão foi direcionado à área
cardiovascular. Entretanto, apesar do meu interesse na área cardiovascular, o estudo
do ser humano e a integração dos seus vários sistemas biológicos, sempre me
despertou curiosidade e fascínio.
Até o final de 2015, finalizarei mais uma etapa de aprendizado, pesada e robusta,
mas vestindo sentimentos antagônicos, de alegria e de angústia, pois me vejo diante da
suposta constatação de Sócrates: "só sei que nada sei...” e completou o filósofo: “mas
o fato de saber isso, me coloca em vantagem sobre aqueles que acham que sabem
alguma coisa”. Daqui em diante, a minha única certeza é que essa estrada longa na
busca do conhecimento será fascinante e interminável.

17
1.2. Justificativa
Os efeitos benéficos do exercício crônico sobre o sistema cardiovascular
encontram-se bem descritos na literatura, no entanto, quando o tema discorre sobre a
intensidade do exercício sobre os aspectos biológicos e clínicos, as diretrizes são
controversas tanto na prevenção quanto na reabilitação cardiovascular (BARINAGA,
1997; MANSON et al, 2002; KEMI et al, 2005). Embora a intensidade elevada de
treinamento induza a aumentos maiores em aptidão física e consumo máximo de
oxigênio (ROGNMO et al, 2012), a intensidade moderada é frequentemente
recomendada como igualmente eficaz (KEMI et al, 2005; ALVES et al, 2010).
Nessa perspectiva, muitos estudos ainda são necessários, pois pouco se sabe
sobre a influência do treinamento físico em parâmetros morfo-funcionais cardíacos,
tanto em humanos quanto em animais de experimentação independente da presença
de agentes patogênicos. Existe disponível na literatura 318 estudos publicados na base
de dados da PubMed relacionando as palavras remodelamento cardíaco e exercício
(Figura 1).
Figura 1. Resultado da busca de artigos científicos na PubMed (05/09/2015), de 1991 a 2015, utilizando as palavras chaves: cardiac remodeling exercise training.
Após o refinamento na busca, tendo em vista a influência do treinamento por
natação no remodelamento cardíaco (Figura 2), os números são ínfimos, perfazendo
um total de 25 estudos disponíveis na base de dados.
0
10
20
30
40
50
199
1
199
3
199
4
199
5
199
7
199
9
200
0
200
1
2002
2003
2004
2005
2006
2007
2008
2009
201
0
201
1
201
2
201
3
201
4
201
5
AR
TIG
OS
PU
BLI
CA
DO
S (N
°)
ANO DE PUBLICAÇÃO
Palavras-chave: cardiac remodeling, exercise training.
Total de artigos = 318
18
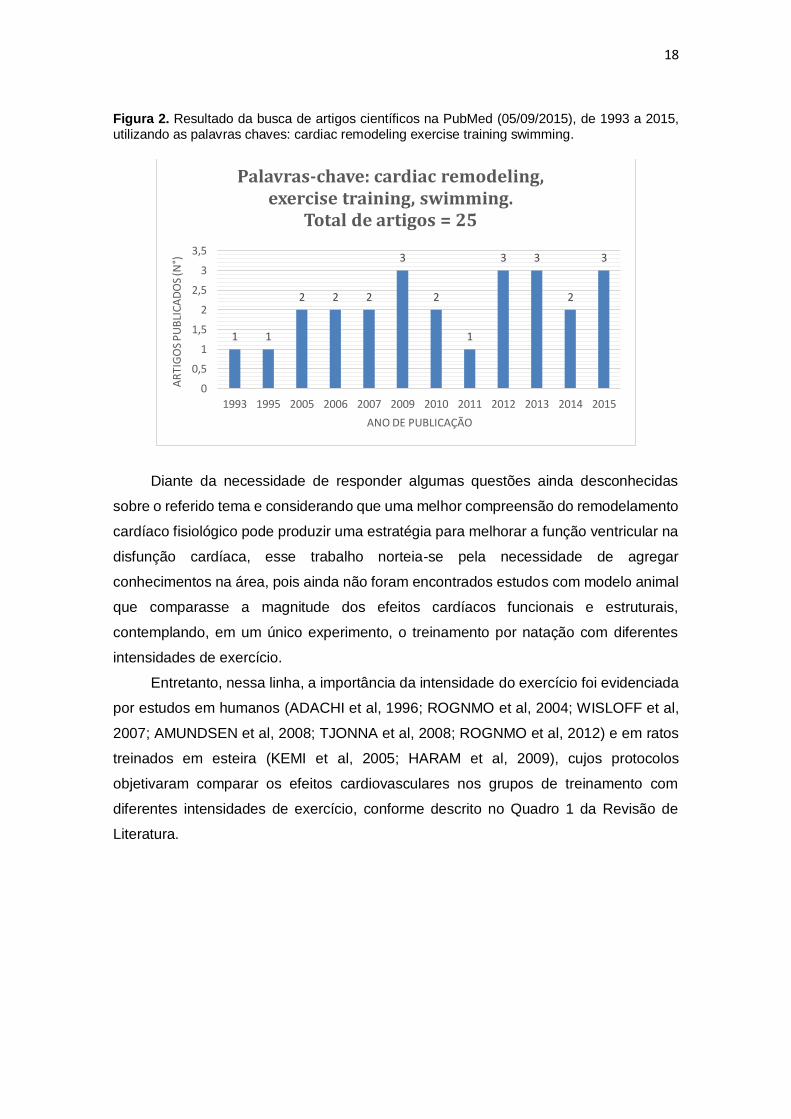
Figura 2. Resultado da busca de artigos científicos na PubMed (05/09/2015), de 1993 a 2015, utilizando as palavras chaves: cardiac remodeling exercise training swimming.
Diante da necessidade de responder algumas questões ainda desconhecidas
sobre o referido tema e considerando que uma melhor compreensão do remodelamento
cardíaco fisiológico pode produzir uma estratégia para melhorar a função ventricular na
disfunção cardíaca, esse trabalho norteia-se pela necessidade de agregar
conhecimentos na área, pois ainda não foram encontrados estudos com modelo animal
que comparasse a magnitude dos efeitos cardíacos funcionais e estruturais,
contemplando, em um único experimento, o treinamento por natação com diferentes
intensidades de exercício.
Entretanto, nessa linha, a importância da intensidade do exercício foi evidenciada
por estudos em humanos (ADACHI et al, 1996; ROGNMO et al, 2004; WISLOFF et al,
2007; AMUNDSEN et al, 2008; TJONNA et al, 2008; ROGNMO et al, 2012) e em ratos
treinados em esteira (KEMI et al, 2005; HARAM et al, 2009), cujos protocolos
objetivaram comparar os efeitos cardiovasculares nos grupos de treinamento com
diferentes intensidades de exercício, conforme descrito no Quadro 1 da Revisão de
Literatura.
1 1
2 2 2
3
2
1
3 3
2
3
0
0,5
1
1,5
2
2,5
3
3,5
1993 1995 2005 2006 2007 2009 2010 2011 2012 2013 2014 2015
AR
TIG
OS
PU
BLI
CA
DO
S (N
°)
ANO DE PUBLICAÇÃO
Palavras-chave: cardiac remodeling, exercise training, swimming.
Total de artigos = 25

19
1.3. Objeto do estudo
O objeto do presente estudo investigou os efeitos do exercício aeróbio de
diferentes intensidades em ratos fêmeas, relacionando-os às mudanças cardíacas
estruturais e contráteis, influenciadas pelo ganho da capacidade funcional adquirida
pelo treinamento por natação.

20
1.4. Objetivos
1.4.1. Objetivo geral
O objetivo do estudo foi investigar as alterações biométricas e contráteis do
miocárdio de ratas, submetidas a 12 semanas de treinamento de natação com diferentes
intensidades de esforço.
1.4.2. Objetivos específicos
Avaliar os efeitos de 12 semanas de treinamento de natação com diferentes
intensidades de esforço nas seguintes variáveis:
a. Capacidade física;
b. Massa corporal;
c. Massa cardíaca total, atriais e ventriculares;
d. Função ventricular;
e. Contratilidade miocárdica;
f. Potenciação pós pausa;
g. Curva de estiramento tensão (Curva de Frank-Starling).

21
1.5. Hipóteses
Considerando que as evidências científicas apresentam muitos aspectos
relacionados à influência do treinamento aeróbio por natação nas alterações estruturais
cardíacas e contráteis foram consideradas as seguintes hipóteses para o
desenvolvimento do estudo:
a) Espera-se que não sejam encontradas diferenças na massa corporal entre os
grupos de intervenção.
b) Possivelmente, a capacidade física dos animais treinados será melhor do que
o grupo controle. Entretanto, o grupo de treinamento intenso poderá apresentar
melhores adaptações em relação ao grupo moderado.
c) Nos parâmetros correspondentes a hipertrofia cardíaca, pela avaliação
biométrica espera-se que em ambos os grupos treinados seja encontrado aumento da
massa cardíaca quando comparado ao grupo controle. Entretanto, acreditamos que o
grupo de treinamento intenso apresentará maiores adaptações cardíacas em relação ao
grupo de treinamento moderado.
d) Em relação aos parâmetros funcionais, a função miocárdica poderá ser
aprimorada nos grupos treinados em relação ao grupo controle, contudo o grupo de
treinamento intenso possivelmente apresentará maiores adaptações cardíacas em
relação ao grupo de treinamento moderado.
e) A potenciação pós-pausa poderá ser aprimorada nos grupos treinados em
relação ao grupo controle, contudo esperamos que o grupo de treinamento intenso
apresente maiores adaptações cardíacas em relação ao grupo de treinamento
moderado.
f) A função inotrópica dos grupos, representada pela curva de estiramento
tensão, possivelmente será preservada ou aumentada, contudo o grupo submetido ao
treinamento intenso poderá apresentar melhor desempenho contrátil em relação aos
grupos de treinamento moderado e controle. Adicionalmente, o grupo controle
certamente apresentará menor desempenho contrátil em relação ao grupo de
treinamento de intensidade moderada.

22
2. REVISÃO DE LITERATURA
Esse capítulo pretendeu coletar informações da literatura científica pertinentes ao
tema e às principais inquietações que nortearam o presente trabalho.
Inicialmente, essa revisão discorreu sobre a ação do exercício aeróbio crônico
sobre as adaptações cardíacas, enfatizando a discussão sobre as diferentes
intensidades de exercício e também foi abordado o entendimento sobre o processo de
remodelamento cardíaco e as suas peculiaridades, bem como os achados sobre a
função contrátil e as demais etapas desse processo de adequação cardíaca ao
treinamento físico.
2.1. Adaptações cardíacas no exercício aeróbio: papel da modalidade
O efeito crônico do exercício tem sido estudado ao longo dos anos na busca do
entendimento efetivo sobre as adaptações provocadas no organismo humano. Nessa
perspectiva, a ação do exercício físico regular e a melhora da capacidade funcional
apresentam-se como importantes fatores de implemento de saúde e os seus efeitos
benéficos sobre o sistema cardiocirculatório encontram-se bem descritos na literatura
(ELLIOTT et al, 2013; ROGNMO et al, 2012; FREIMANN et al, 2005; STROM et al, 2005;
WAGNER et al, 2002; SHEPARD e BALADY, 1999; POWERS et al, 1998; BERLIN e
COLDITZ, 1990).
Dentre os protocolos de exercícios utilizados, efeitos positivos do treinamento
físico aeróbio tanto em humanos (ADACHI et al, 1996; ROGNMO et al, 2004; WISLOFF
et al, 2007; AMUNDSEN et al, 2008; TJONNA et al, 2008; ROGNMO et al, 2012) quanto
em animais (SCHAIBLE e SCHEUER, 1981; KEMI et al, 2005; HARAM et al, 2009)
mostraram melhora na função ventricular, evidenciando aumento do débito cardíaco e
do volume sistólico. Outros estudos também avaliaram a performance em corações
isolados (SCHAIBLE e SCHEUER, 1979; FULLER e NUTTER, 1981), músculos
papilares (BOCALINI et al, 2010; CHEN et al, 2001) e miócitos isolados (BOZI et al,
2013; KEMI et al, 2005; WISLOFF et al, 2002) e todos eles ressaltaram aumento da
função cardíaca.
Outro aspecto que deve-se levar em consideração é a diferença na magnitude da
hipertrofia cardíaca que está diretamente relacionada ao protocolo de treinamento
utilizado (WANG et al, 2010; HASHIMOTO et al, 2011; FERNANDES et al, 2011;
BARROS, 1994). Nesse aspecto, a natação é frequentemente utilizada em estudos de

23
fisiologia do exercício envolvendo ratos, especialmente por tratar-se de uma habilidade
inata (KREGEL, 2006; de ARAUJO et al, 2007) e por apresentar custos mais baixos em
comparação com a corrida em esteira rolante (de ARAUJO et al, 2007). Além disso,
estudos utilizando este modelo revelou semelhanças nas adaptações ao exercício em
relação aos observados em seres humanos (de ARAUJO et al, 2007; WANG et al, 2010).
Sendo assim, o treinamento de natação em ratos é reconhecido por sua eficácia
em induzir maior hipertrofia cardíaca quando comparado ao exercício em esteira
(WANG et al, 2010; SCHAIBLE e SCHEUER, 1981), promovendo aumento significativo
no volume diastólico final do ventrículo esquerdo (GEENEN et al, 1988; SCHAIBLE e
SCHEUER, 1981). Seguindo essa linha, SCHAIBLE e SCHEUER (1981) mostraram que
o aumento no fluxo coronariano é proporcional ao grau de hipertrofia induzido pelo
treinamento físico resultante de um aumento do leito vascular coronário e concluíram
que o treinamento crônico de natação em ratas promoveu aprimoramento do
desempenho contrátil do miocárdio, apesar de um acentuado grau de hipertrofia,
contrastando com treinamentos em esteira onde não foram observados nem hipertrofia
nem melhora na função.
De fato, a maioria dos estudos de corrida em esteira não conseguiu mostrar
hipertrofia cardíaca em ratos (FITZSIMONS et al, 1990; SCHAIBLE e SCHEUER, 1981;
HAN, 2013), no entanto, alguns pesquisadores observaram hipertrofia cardíaca em
ratos treinados em protocolos de corrida (KEMI et al, 2002; ALLEN et al, 2001). Em
ambos estudos de KEMI (2002 e 2007) com treinamento de alta intensidade em esteira,
verificou-se um aumento significativo no tamanho dos cardiomiócitos, bem como uma
melhora da contratilidade e da função cardíaca, sugerindo que a alta intensidade nos
programas de esteira ou roda de corrida voluntária também parecem eficazes para
indução de hipertrofia fisiológica (WANG et al, 2010).
2.2. Adaptações cardíacas no exercício aeróbio: papel da intensidade
A adaptação ao treinamento é dependente de fatores como carga, duração e
frequência, porém, do ponto de vista das adaptações miocárdicas relativas a diferentes
intensidades pouco se sabe. Ainda que a intensidade elevada de treinamento induza a
aumentos maiores em aptidão física e consumo máximo de oxigênio (VO2máx)
(ELLIOTT et al, 2013; TJONNA et al, 2008; WISLOFF et al, 2007; KEMI et al, 2005), a
intensidade moderada é frequentemente recomendada como igualmente eficaz (LAVIE
et al, 2015; ALVES et al; 2010; KEMI et al, 2005).

24
Profissionais e instituições mais conservadoras costumam prescrever a
intensidade do exercício físico para jovens e idosos, portadores ou não de fatores de
risco para a doença cardiovascular, pelos limiares ventilatórios fornecidos pela
ergoespirometria (ALVES et al, 2010). Contudo, para indivíduos com insuficiência
cardíaca, a prescrição do limite superior é estabelecida 10% menor que o valor
registrado no ponto de compensação respiratória, evitando que o exercício seja
realizado em acidose metabólica descompensada (ALVES et al, 2010).
Embora o exercício de alta intensidade seja comumente evitado devido aos riscos
para cardiopatas, em 2012, ROGNMO e colegas revelaram que a taxa de evento
adverso associado a este modo de treinamento é baixa e, portanto, adequado para
pacientes com doença arterial coronariana.
Vários estudos (WISLOFF et al, 2007; TJONNA et al, 2008; ROGNMO et al, 2012;
ELLIOTT et al, 2013) mostraram maior efetividade no ganho de capacidade física,
qualidade de vida e no controle de fatores de risco, quando essa população foi
submetida a um treinamento de maior intensidade, evidenciando a importância e a
segurança da intensidade mais alta e demonstrando que esses achados podem mudar
alguns paradigmas no futuro (ALVES et al, 2010).
Defende-se também que o exercício vigoroso pode agudamente e
transitoriamente aumentar o risco de morte súbita cardíaca e infarto do miocárdio em
pessoas susceptíveis (THOMPSON et al, 2007).
Nessa análise, a dose ideal do treinamento de alta intensidade ainda é objeto de
estudos. Já foi demonstrado que uma única sessão semanal de exercício de alta
intensidade reduziu o risco de morte cardiovascular, em homens e mulheres livres de
doenças cardiovasculares (WISLOFF et al, 2006) e em pacientes com doença coronária
estabelecida (MOHOLDT et al, 2008).
A recomendação mínima do ACSM e AHA (HASKELL et al, 2007) para evitar
doenças crônicas em adultos e idosos é de 30 minutos por dia, durante 5 dias na
semana de atividade aeróbia moderada ou 20 minutos por dia, durante 3 dias na
semana de atividade intensa, ou ainda a combinação das intensidades. Essas
orientações também sugerem que dose mais elevada de atividade aeróbia pode ser
necessária em alguns grupos que tentam impedir a transição para o excesso de peso
ou obesidade (O'KEEFE et al, 2012).
Em contrapartida, não há uma prescrição para evitar a overdose do exercício e
suas doenças relacionadas, mas parece aconselhável, pelo menos do ponto de vista da
saúde pública, limitar o treinamento físico vigoroso a 60 min/dia, perfazendo um

25
acumulado semanal de não mais do que 5 horas, considerando ainda 1 ou 2 dias off por
semana de exercícios de alta intensidade (LAVIE et al, 2015; O'KEEFE et al, 2012).
De fato, o excesso de exercício pode ser cardiotóxico em indivíduos predispostos
(MONT et al, 2009; SARQUELLA-BRUGADA et al, 2012; MAILLET et al, 2013) e, nessa
linha, existe uma relação entre as horas acumuladas de prática e o risco de fibrilação
atrial (MONT et al, 2009). Estudos em ratos, sem a presença de agentes patogênicos
(BENITO et al, 2011) e em atletas maratonistas (SHARMA e ZAIDI, 2012) que treinaram
excessivamente ao longo dos anos, mostraram associação com inflamação do
miocárdio e fibrose, resultando em um substrato para arritmogênese e disfunção do
miocárdio, no entanto, os mecanismos subjacentes ainda devem ser elucidados,
embora as alterações estruturais atriais como dilatação e fibrose estejam presentes
(MONT et al, 2009).
Adicionalmente, estudos recentes (McCULLOUGH e LAVIE, 2014; SCHWARTZ
et al, 2014) sugerem que corredores de longa distância podem ter, ao longo dos anos
de treinamento ininterrupto, níveis aumentados de aterosclerose e doenças
coronarianas. Doses muito elevadas de exercício de resistência em alguns indivíduos
com predisposição genética ou com doenças pré-existentes (SARQUELLA-BRUGADA
et al, 2012; MAILLET et al, 2013), pode levar à dilatação e disfunção cardíaca (LAVIE
et al, 2015), especialmente no lado direito do coração e septo ventricular que por sua
vez, conduz à libertação de marcadores cardíacos, tais como a troponina e peptídeo
natriurético cerebral, resultando em arritmias ventriculares malignas e aumento do risco
de morte súbita cardíaca (LAVIE et al, 2015), contudo, ainda há lacunas na literatura
abordando temas que permitam estabelecer o limite de exercício para a toxicidade
cardíaca potencial, o rastreio de indivíduos em risco, e os programas de treinamento
físico ideal para otimizar a saúde cardiovascular (O'KEEFE et al, 2012).
Por essa razão, parece relativamente fraca a evidência de que vários anos de alta
intensidade possa provocar a fibrose cardíaca ou acelerar a aterosclerose, tendo em
vista que não há nenhum argumento contundente de que o treinamento excessivo de
endurance possa ser potencialmente prejudicial para o coração (LAVIE et al, 2015). Em
última análise, evidências norteiam estudos (BHELLA et al, 2014; LEW, 2014),
sinalizando que níveis muito elevados de treinamento de endurance podem impedir a
redução da complacência e distensibilidade dos vasos sanguíneos, observado no
processo de envelhecimento saudável e sedentário.
A importância da intensidade do exercício foi evidenciada por estudos em
humanos (ROGNMO et al, 2012; WISLOFF et al, 2007; SWAIN e FRANKLIN, 2006,

26
ROGNMO et al, 2004) e em ratos treinados em esteira (KEMI et al, 2005; WISLOFF et
al, 2002; KEMI et al, 2002) que mostraram que o treinamento físico aeróbio de alta
intensidade pode ser mais favorável para adaptações na aptidão cardiorrespiratória e
na função cardíaca do que o exercício de intensidade baixa a moderada, sugerindo
ainda que as mudanças vasculares e periféricas podem contribuir para os efeitos
dependentes da intensidade, como a função endotelial arterial e seus mecanismos
reguladores (HARAM et al, 2009).
NUTTER (1981) demonstrou que o treinamento de resistência em intensidade
moderada não resultou em hipertrofia cardíaca significativa, rigidez miocárdica alterada,
ou mudanças consistentes na contratilidade miocárdica. Nessa linha, o estudo de KEMI
(2005) com ratos treinados em esteira, sem a presença de agentes patogênicos,
demonstrou que as adaptações cardiovasculares ao treinamento dependem da
intensidade. A estreita correlação entre VO2máx, dimensões dos cardiomiócitos e
capacidade contrátil sugeriu significativamente maior benefício com alta intensidade, em
contraste, os efeitos sobre a função endotelial foram induzidos pelo exercício regular de
intensidade moderada (KEMI et al, 2005).
Ainda nesse aspecto, abordagens epidemiológicas e clínicas tem mostrado que
as adaptações do exercício físico podem ser dependentes tanto da intensidade quanto
no volume total do treinamento (ADACHI et al, 1996; BELARDINELLI et al, 1999;
TANASESCU et al, 2002; GREGG et al, 2003; LEE et al, 2003; ROGNMO et al, 2004).
Tal fato pode estar associado ao equivalente metabólico ou ao consumo máximo de
oxigênio, cuja função é transportar o oxigênio da atmosfera e entregá-lo para os
músculos, onde é usado durante a respiração mitocondrial para produzir ATP
(BASSETT e HOWLEY, 2000).
Nessa análise, pode-se concluir que o sistema de transporte de oxigênio sofre
uma adaptação favorável que se exterioriza através de maiores valores de VO2máx
como consequência das adaptações funcionais decorrente do processo de
remodelamento cardíaco induzido pelo treinamento físico aeróbio (SBC, 2005). Dessa
forma, o consumo de oxigênio de pico (VO2pico) tem emergido como uma importante
referência clínica que o identificaram como um dos maiores preditores independente de
mortalidade tanto em indivíduos saudáveis quanto em cardiopatas (PAFFENBARGER
et al, 1993; BELARDINELLI et al, 1999; MYERS et al, 2002; KAVANAGHT et al, 2002;
GULATI et al, 2003).
Enquanto um adulto jovem saudável, não atleta, apresenta VO2máx da ordem de
35 ml/kg/min, em atletas de resistência aeróbia, essa variável aumenta

27
significantemente, 50%, em média (BARROS 1994), podendo ultrapassar 70ml/kg/min
(GHORAYEB e BATLOUNI, 1998). Esse índice fisiológico é também considerado o
principal indicador das adaptações cardiovasculares e, provavelmente, o indicador
isolado mais objetivo do nível de treinamento físico (FLETCHER et al, 2001;
GHORAYEB et al, 2005), observando ainda que a melhora da potência aeróbica máxima
costuma ser inversamente proporcional à capacidade física antes do treinamento, sendo
que os pacientes mais comprometidos são aqueles que, proporcionalmente, obtêm as
melhorias mais significativas (SBC, 2005).
Entretanto, apesar da importância desse marcador, quando analisamos as
recomendações gerais tanto para prevenção quanto para reabilitação, encontramos na
literatura variações de intensidade correspondente de 40% a 85% do VO2máx (ACSM,
1994; FLETCHER et al, 2001), causando controvérsias sobre os efeitos biológicos e
clínicos entre os exercícios de moderada e alta intensidade (BARINAGA, 1997;
MANSON et al, 2002; KEMI et al, 2005).
Dessa forma, do ponto de vista da aptidão física, sabe-se que a variação da
intensidade difere entre os ganhos do VO2máx tanto em homens (AMUNDSEN et al,
2008; GIBALA e McGEE, 2008) quanto em animais (KEMI et al, 2008a; KEMI et al,
2008b; KEMI et al, 2007; KEMI et al, 2005; KEMI et al, 2004; WISLOFF et al, 2002), sem
doenças (GIBALA e McGEE, 2008; KEMI et al, 2008a; KEMI et al, 2008b; KEMI et al,
2007; KEMI et al, 2005; KEMI et al, 2004; WARBURTON et al, 2005; WISLOFF et al,
2002; WENGER e BELL, 1986; WISLOFF et al, 2007) ou cardiopatas (AMUNDSEN et
al, 2008; ROGNMO et al, 2004; ADACHI et al, 1996), reforçando a ideia de que as
adaptações são condicionadas à intensidade do exercício.
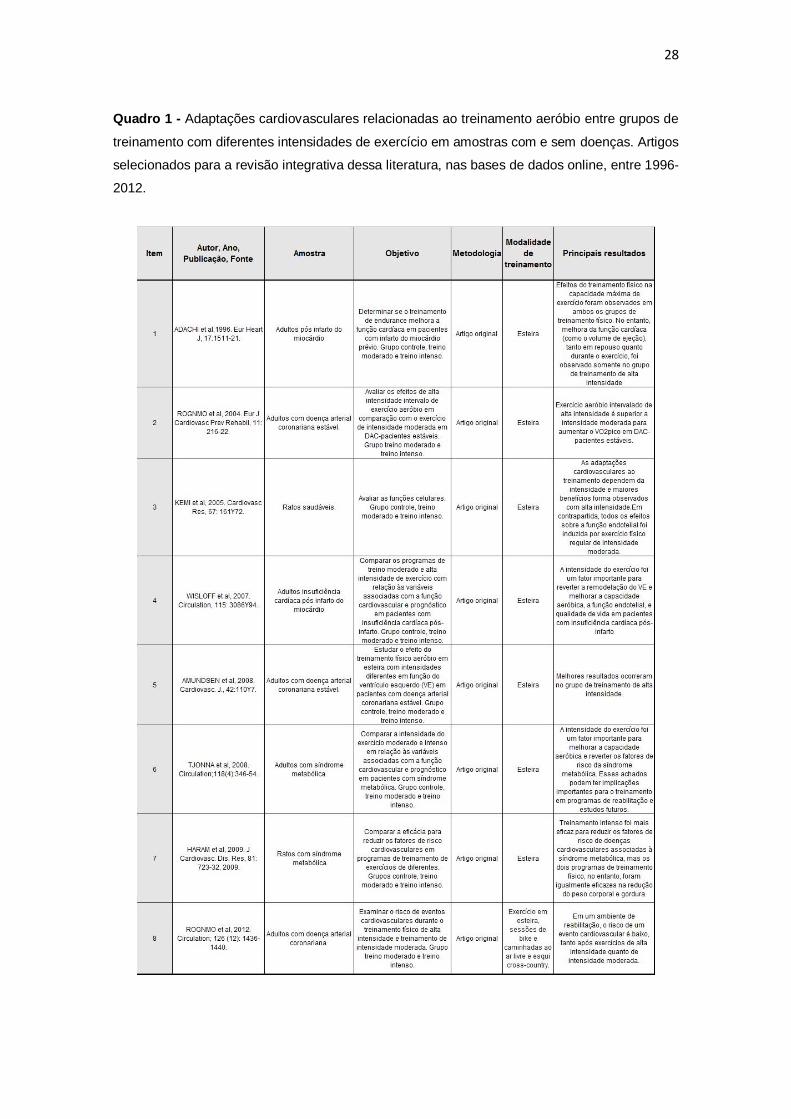
O Quadro 1 descreve os artigos produzidos na literatura que compararam os
efeitos cardiovasculares entre grupos de treinamento aeróbio com diferentes
intensidades de exercício em amostras com e sem doenças.
28
Quadro 1 - Adaptações cardiovasculares relacionadas ao treinamento aeróbio entre grupos de
treinamento com diferentes intensidades de exercício em amostras com e sem doenças. Artigos
selecionados para a revisão integrativa dessa literatura, nas bases de dados online, entre 1996-
2012.

29
2.3. Avaliação da aptidão aeróbia em roedores
A maioria dos estudos sobre a resposta do lactato ao exercício foram realizados
em seres humanos, no entanto, para investigar aspectos das adaptações cardíacas que
não podem ser realizadas em humanos, mais atenção tem sido dada para o
desenvolvimento de métodos capazes de avaliar a aptidão aeróbia em roedores
considerando as respostas do lactato ao exercício (VOLTARELLI et al, 2002,
CARVALHO et al, 2005).
GOBATTO (2001) determinaram a máxima fase estável de lactato em ratos,
submetendo os animais a esforços diários de natação contínua com a duração de 20
minutos, suportando cargas equivalentes a 5, 6, 7, 8, 9 e 10% do peso do corpo.
Amostras de sangue foram coletadas a cada 5 min. Os autores observaram
estabilização do lactato sanguíneo em 5,5 mmol/l, correspondente a cargas de 5 e 6%
do peso corporal.
Em um estudo anterior em ratos, PILLIS (1993) observaram que durante um teste
em esteira, o limiar anaeróbio ocorreu quando a concentração de lactato sanguíneo foi
de aproximadamente 4,0 mmol/l, sugerindo que o tipo de exercício usado, corrida ou
natação, podem afetar o valor de lactato sanguíneo no limiar anaeróbio.
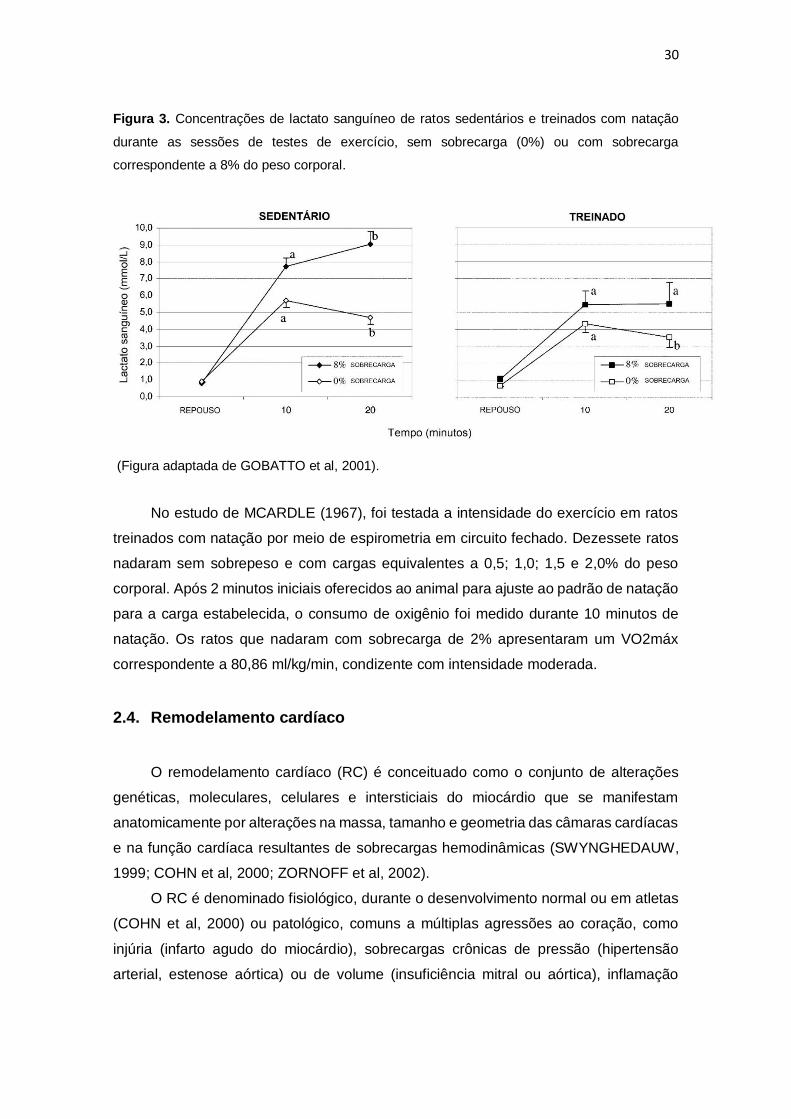
Conforme demonstrado na Figura 3, GOBATTO (2001) verificaram que os ratos
sedentários treinados com natação não apresentaram estabilização do lactato
sanguíneo, mas o grupo treinado conseguiu estabilizar na concentração apontada como
estado estável de lactato máximo sanguíneo de 5,5 mmol/l. Esses dados sugerem que
a capacidade de endurance dos animais foi melhorada com sobrecarga de 8% e a
concentração de lactato máximo permaneceu estável em 5,5 mmol/l nos animais
treinados.
30
Figura 3. Concentrações de lactato sanguíneo de ratos sedentários e treinados com natação
durante as sessões de testes de exercício, sem sobrecarga (0%) ou com sobrecarga
correspondente a 8% do peso corporal.
(Figura adaptada de GOBATTO et al, 2001).
No estudo de MCARDLE (1967), foi testada a intensidade do exercício em ratos
treinados com natação por meio de espirometria em circuito fechado. Dezessete ratos
nadaram sem sobrepeso e com cargas equivalentes a 0,5; 1,0; 1,5 e 2,0% do peso
corporal. Após 2 minutos iniciais oferecidos ao animal para ajuste ao padrão de natação
para a carga estabelecida, o consumo de oxigênio foi medido durante 10 minutos de
natação. Os ratos que nadaram com sobrecarga de 2% apresentaram um VO2máx
correspondente a 80,86 ml/kg/min, condizente com intensidade moderada.
2.4. Remodelamento cardíaco
O remodelamento cardíaco (RC) é conceituado como o conjunto de alterações
genéticas, moleculares, celulares e intersticiais do miocárdio que se manifestam
anatomicamente por alterações na massa, tamanho e geometria das câmaras cardíacas
e na função cardíaca resultantes de sobrecargas hemodinâmicas (SWYNGHEDAUW,
1999; COHN et al, 2000; ZORNOFF et al, 2002).
O RC é denominado fisiológico, durante o desenvolvimento normal ou em atletas
(COHN et al, 2000) ou patológico, comuns a múltiplas agressões ao coração, como
injúria (infarto agudo do miocárdio), sobrecargas crônicas de pressão (hipertensão
arterial, estenose aórtica) ou de volume (insuficiência mitral ou aórtica), inflamação

31
(miocardites) ou expressão de um programa genético que leva à miocardiopatia
(PONTES e LEÃES, 2004). Esses eventos permitem que o coração se adapte às novas
condições de funcionamento e ativem importantes mediadores que incluem estresse
parietal, neuro hormônios (sistema simpático, renina-angiotensina, aldosterona e
endotelina), citocinas, produção de óxido nítrico e estresse oxidativo (COHN et al, 2000).
A principal adaptação do coração ao treinamento físico é o RC (CRAIG et al, 1991;
WISLOFF et al, 2001a; WISLOFF et al, 2001b; WISLOFF et al, 2002; KEMI et al, 2004),
e os efeitos cardioprotetores estão relacionados às mudanças cardíacas estruturais e
funcionais, influenciadas pelo ganho da capacidade funcional adquirida pelo
treinamento. Os principais mecanismos fisiológicos atribuídos ao RC são o aumento da
contratilidade (WISLOFF et al, 2001b; WISLOFF et al, 2002; ZHANG et al, 2000a;
ZHANG et al, 2000b), melhora no transiente de Ca2+ no meio intracelular, afetando a
função ventricular (WISLOFF et al, 2001b; DIFFEE e NAGLE, 2003; DIFFEE et al, 2001),
aumento da dilatação com expressivas melhoras na oxigenação miocárdica
(HAMBRECHT et al, 1998) e adicionais funções endotélio-dependente que impedem
eventos isquêmicos (KEMI et al, 2005).
Entretanto, as respostas do RC fisiológico não são uniformes e variam de acordo
com o tipo e a intensidade do exercício físico realizado, assim como ao programa de
treinamento físico (MORGANROTH et al, 1975), e está diretamente relacionado à
capacidade aeróbia máxima ou VO2máx (BOOTH et al, 1998; MONTGOMERY,1997).
Dessa forma, o RC, considerado um dos mais significativos resultados da adaptação
crônica ao exercício, deriva na hipertrofia do miócito para adequação do desempenho
cardíaco às demandas do organismo (NATALI, 2004; GHORAYEB et al, 2005).
As adaptações ao treinamento físico ocorrem ao longo de um processo integrado,
do nível sistêmico ao nível molecular (de OLIVEIRA et al, 2010). O avanço do
conhecimento nesta área se renova à medida que novas tecnologias surgem como
ferramentas para estudos experimentais (de OLIVEIRA et al, 2010) e o entendimento
desses conceitos torna-se fundamental, desde a reabilitação até o esporte de alto
rendimento, pois dá suporte aos profissionais que trabalham com a atividade física na
avaliação de certos procedimentos, no tratamento de doenças, na recuperação de
lesões e na seleção de talentos esportivos (de OLIVEIRA et al, 2010).
O início do processo de RC é motivado por um estímulo hemodinâmico
proveniente de uma sobrecarga transitória, como a que ocorre no crescimento e no
treinamento físico ou de uma sobrecarga persistente, advinda de eventos como o infarto
do miocárdio ou de algumas doenças como a hipertensão.

32
Os fatores desencadeantes do RC sensibilizam receptores de membrana ligados
ao sistema de sinalização intracelular pelo qual o estímulo atua sobre o núcleo e
condiciona às modificações da síntese proteica (dos SANTOS e TUCCI, 2007). Com o
aumento no conteúdo das proteínas contráteis, o resultado da adaptação deriva na
hipertrofia do cardiomiócito (NATALI, 2004; GHORAYEB et al, 2005) com consequentes
adaptações na funcionalidade do órgão. Nessa perspectiva, o resultado final do RC
pode ter diferentes contornos e consequências de acordo com o tipo e o tempo do
estímulo gerador (DORN, 2007; McMULLEN e JENNINGS, 2007).
O crescimento do coração, do nascimento à idade adulta, é dirigido pela hipertrofia
cardíaca fisiológica também referida como hipertrofia pós-natal, responsável pelo
crescimento do coração materno durante a gravidez e pelo crescimento do coração de
atletas como resultado de exercícios intensos e/ou repetitivos (MAILLET et al, 2013).
Assim, a fase de crescimento e o treinamento físico induzem o coração à
hipertrofia fisiológica caracterizada pela inexistência de fibrose e alteração na expressão
de genes hipertróficos pós-natal, não ocorrendo, nesse caso, alterações na expressão
dos genes fetais que são classicamente associados com o desenvolvimento de
hipertrofia patológica (KEMI et al, 2007; KEMI et al, 2008; WISLOFF et al, 2009; KEHAT
e MOLKENTIN, 2010; MAILLET et al, 2013). Isso sugere que a hipertrofia fisiológica
induzida por exercício depende mais da tradução do mRNA, incluindo a sua eficiência,
ou seja, a síntese de proteína por mRNA transcrito, e da manutenção das proteínas
sintetizadas (WISLOFF et al, 2009).
Em contraste, as adaptações hipertróficas patológicas são acompanhadas por
alterações nos genes fetais que permaneciam latentes e que voltam a reger a síntese
proteica (dos SANTOS e TUCCI, 2007), conferindo características fenotípicas de
imaturidade à célula hipertrofiada (de OLIVEIRA et al, 2012). Nesse tipo de hipertrofia,
os padrões de expressão dos genes mimetizam aqueles observados durante o
desenvolvimento embrionário (FREY et al, 2004), passando a compor o miocárdio com
características funcionais diferentes daquelas sintetizadas sob a regência de genes da
vida extrauterina (dos SANTOS e TUCCI, 2007).
Dessa forma, verificamos que os mecanismos moleculares ativados no
treinamento aeróbio são diferentes dos encontrados nas hipertrofias patológicas
(McMULLEN et al, 2003; McMULLEN e JENNINGS, 2007; WISLOFF et al, 2009; KEHAT
e MOLKENTIN, 2010; HASHIMOTO et al, 2011; FERNANDES et al, 2011; MAILLET et
al, 2013).

33
Esforços consideráveis de investigação têm sido centralizados sobre os
mecanismos moleculares responsáveis pela transdução de carga hemodinâmica em
crescimento cardíaco induzido pelo exercício físico (FERNANDES et al, 2011). Nesse
aspecto, um número elevado de vias de sinalização intracelular foi identificado como
transdutores importantes na resposta hipertrófica fisiológica em cardiomiócitos (DORN,
2007).
Embora tanto a hipertrofia cardíaca fisiológica quanto a patológica estejam
associadas à um aumento da massa do coração, a hipertrofia fisiológica está associada
a uma estrutura e função cardíaca normal ou aumentada (McMULLEN e JENNINGS,
2007; McMULLEN et al, 2003; WISLOFF et al, 2009; KEHAT e MOLKENTIN, 2010;
HASHIMOTO et al, 2011; FERNANDES et al, 2011; MAILLET et al, 2013), enquanto a
hipertrofia patológica associa-se a uma série complexa de eventos, incluindo a
regulação positiva de genes fetais, histopatologia e disfunção cardíaca (McMULLEN e
JENNINGS, 2007).
Levando-se em consideração que as alterações fisiológicas causadas pelo
exercício têm efeitos favoráveis sobre a hipertrofia patológica, uma compreensão do
remodelamento cardíaco fisiológico pode fornecer uma estratégia para melhorar a
função ventricular na disfunção cardíaca (FERNANDES et al, 2011; McMULLEN e
JENNINGS, 2007; WISLOFF et al, 2007). Nesse aspecto, intervenções terapêuticas
com base em estímulos de genes que conduzem ao crescimento fisiológico pode
proporcionar uma adicional estratégia para tratar ou reverter a insuficiência cardíaca
(FERNANDES et al, 2011; McMULLEN e JENNINGS, 2007).
As diferenças bioquímicas e moleculares entre as hipertrofias fisiológica e
patológica foram demonstradas em vários estudos (McMULLEN et al, 2003; McMULLEN
e JENNINGS et al, 2007; WISLOFF et al, 2009; KEHAT e MOLKENTIN, 2010;
HASHIMOTO et al, 2011; MAILLET et al, 2013), conforme modelo adaptado na Figura
3.
De maneira geral, a hipertrofia fisiológica é iniciada por sinais finitos, que incluem
hormonas de crescimento (tais como a hormona da tiróide, insulina, fator de crescimento
tipo insulina 1 (IGF-1) e fator de crescimento do endotélio vascular) e as forças
mecânicas que convergem para um pequeno número de vias de sinalização
intracelulares (tais como PI3K, AKT, ativada por AMP proteína quinase e mTOR) para
afetar a transcrição do gene, tradução proteica e o metabolismo (MAILLET et al, 2013;
McMULLEN e JENNINGS, 2007). Por outro lado, estímulos neuro-humorais como a
angiotensina II (AngII) e endotelina 1 (ET-1) resultam na ativação da proteína G, por

34
meio dos receptores Gαq que estão associados à hipertrofia patológica e consequente
progressão para a insuficiência cardíaca (McMULLEN e JENNINGS, 2007).
Figura 4. Esquema das vias de sinalização envolvidas na indução da hipertrofia cardíaca
fisiológica e patológica.
RTK, receptor tirosina-quinase; mTOR, alvo da rapamicina em mamíferos; NA, noradrenalina; PLC, fosfolipase C; DAG, diacilglicerol; IP3, inositol 1,4,5-trifosfato; MAPK, mitogenativado proteína-quinase; JNK, c-jun amino-terminal quinase; PKC, proteína quinase C; PI3-K, fosfatidilinositol-3 quinase; ERK, sinal extracelular quinase regulamentado; Ang II, angiotensina II; ET-1, endotelina-1; GPCR, L- receptor acoplado a proteína; NFAT, fator nuclear de células T ativadas; IGF1, fator semelhante à insulina de crescimento 1; S6Ks, ribossomal S6 quinases (Figura modificada de McMULLEN e JENNINGS, 2007).
Após a reativação de genes como resposta ao estímulo hemodinâmico, as
transformações miocárdicas próprias do remodelamento são fundamentadas pelas
modificações qualitativas e quantitativas do metabolismo proteico (dos SANTOS e
TUCCI, 2007).
Nesse contexto, o crescimento do músculo cardíaco causado por sobrecargas
hemodinâmicas pode conduzir à hipertrofia adaptada ou fisiológica, ou mal adaptada,
também denominada patológica (FREY et al, 2004; KEMPF e WOLLERT, 2004) e uma
35
das diferenças entre a hipertrofia cardíaca fisiológica e patológica pode consistir no
tempo de estímulo (DORN, 2007; McMULLEN e JENNINGS, 2007).
Dessa forma, a hipertrofia fisiológica é aquela desenvolvida em decorrência da
sobrecarga hemodinâmica transitória (Figura 4), como as observadas no crescimento
cardíaco durante a adolescência, na gestação e em resposta a exercícios regulares,
enquanto a hipertrofia patológica é aquela onde o coração trabalha continuamente
contra uma sobrecarga pressórica ou volumétrica aumentada (MAILLET et al, 2013;
KEMPF e WOLLERT, 2004; DORN, 2007) e caracteriza-se não só pelo crescimento das
fibras do miocárdio, mas também pelas mudanças na arquitetura cardíaca e no
metabolismo celular ocorrendo disfunção miocárdica com aumento da morbidade e
mortalidade (FLORESCU e VINEREANU, 2006).
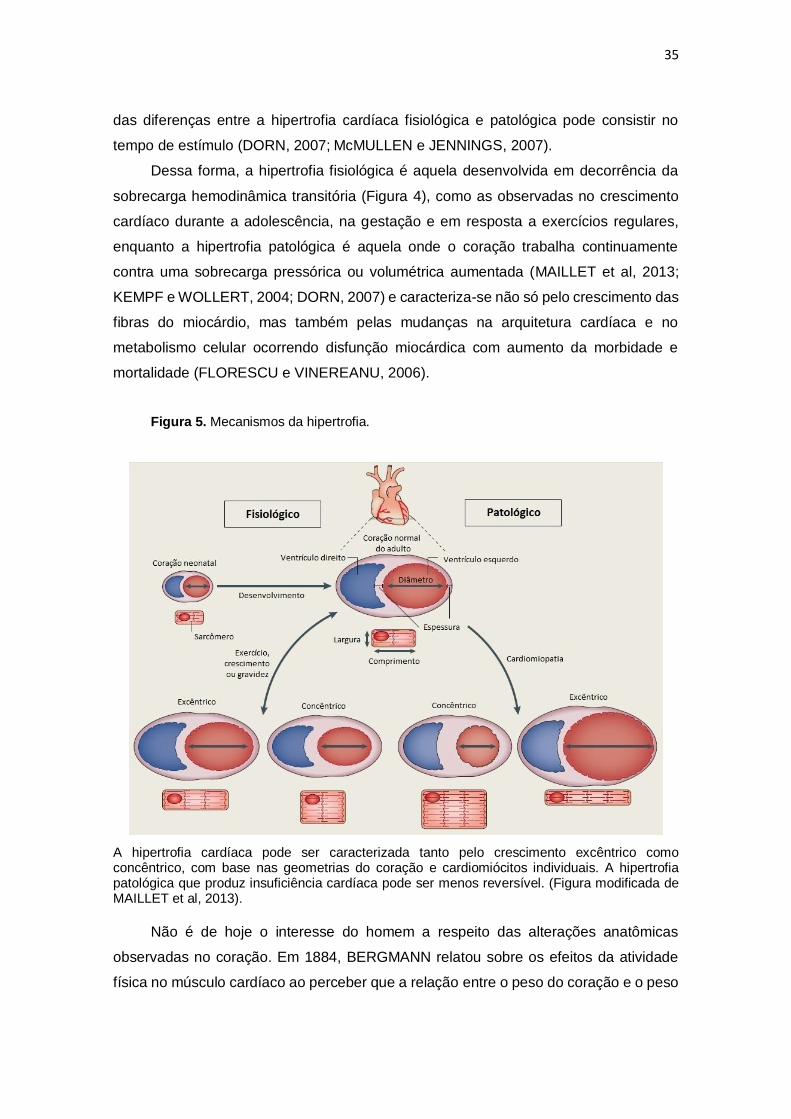
Figura 5. Mecanismos da hipertrofia.
A hipertrofia cardíaca pode ser caracterizada tanto pelo crescimento excêntrico como concêntrico, com base nas geometrias do coração e cardiomiócitos individuais. A hipertrofia patológica que produz insuficiência cardíaca pode ser menos reversível. (Figura modificada de MAILLET et al, 2013).
Não é de hoje o interesse do homem a respeito das alterações anatômicas
observadas no coração. Em 1884, BERGMANN relatou sobre os efeitos da atividade
física no músculo cardíaco ao perceber que a relação entre o peso do coração e o peso

36
corporal de animais selvagens era muito maior quando comparados com os animais
domésticos (ROST, 1997). Anos à frente, em 1899, HENSCHEN descreveu o coração
de esquiadores de campo por meio de auscultação torácica antes e após uma corrida.
Tempos depois, esses resultados se confirmaram por radiografia e por evidências em
autópsia (ROST, 1997).
Em condições fisiológicas como na prática de exercícios físicos, dois tipos de
sobrecarga intermitente podem levar à hipertrofia cardíaca de maneiras diferentes,
porém desenvolvidas de forma simétrica no coração (de OLIVEIRA et al, 2010;
FERNANDES et al, 2011). Entretanto, as alterações estruturais resultantes do
treinamento físico, dependem da natureza, duração e intensidade do exercício
(GHORAYEB et al, 2005). Com relação à natureza do treinamento, a hipertrofia cardíaca
é dependente do tipo de exercício realizado (HASHIMOTO et al, 2011, FERNANDES et
al, 2011, BARROS, 1994).
No exercício estático ou isométrico, como por exemplo, levantadores,
arremessadores de peso e martelo, praticantes de luta romana e fisiculturistas, a força
é desenvolvida com pouco ou nenhum movimento (de OLIVEIRA et al, 2010;
FERNANDES et al, 2011). Esse tipo de exercício apresenta como consequência
hemodinâmica uma ligeira elevação do débito cardíaco, resultante do aumento da
frequência cardíaca e grande elevação da pressão arterial, levando à sobrecarga de
pressão no coração, que resulta em espessamento da parede ventricular esquerda (de
OLIVEIRA et al, 2010) induzindo à hipertrofia concêntrica, com adição de sarcômeros
em paralelo, sem alteração do tamanho da câmara cardíaca ou prejuízo da função
(MORGANROTH et al, 1975; BARROS, 1994; FERNANDES et al, 2011).
No exercício dinâmico, em que os atletas realizam exercícios isotônicos como
nadar, pedalar, correr e andar, os principais padrões hemodinâmicos são o aumento na
frequência cardíaca e no volume sistólico, os dois componentes do débito cardíaco (de
OLIVEIRA et al, 2010). Nesse caso, a sobrecarga sobre o coração é
predominantemente volumétrica, induzindo à hipertrofia ventricular esquerda excêntrica,
com adição de sarcômeros em série que resulta em aumento da câmara cardíaca, sem
prejuízo de função (MORGANROTH et al, 1975; BARROS, 1994; FERNANDES et al,
2011). Esta forma de remodelamento é homogênea e tem como consequência o
aumento da força contrátil (WEBER, 2000).
Em geral, na maioria dos tipos de exercícios físicos ou programas de
condicionamento físico, há uma associação de componentes estático e dinâmico (de
OLIVEIRA et al, 2010). Portanto, a hipertrofia fisiológica que ocorre normalmente é uma

37
combinação de diferentes graus de ambas, hipertrofia concêntrica e excêntrica, levando
à hipertrofia cardíaca mista, como a observada em triatletas (CLAESSENS et al, 1999).
A hipertrofia fisiológica é uma forma suave de crescimento, tipicamente
caracterizada por um aumento de 10-20% no peso do coração, normalizados pelo peso
corporal e totalmente reversível. (MAILLET et al, 2013). Entretanto, em alguns casos, a
hipertrofia fisiológica desenvolvida por atletas de resistência de alto nível poderia ser
incorretamente interpretada como patológica por apresentar uma estrutura
macroscópica semelhante a hipertrofia patológica (de OLIVEIRA et al, 2010;
FERNANDES et al, 2011), no entanto, semelhantes adaptações são geralmente
encontradas em atletas que usam esteroides anabolizantes associados ao treinamento
de resistência (RIEBE et al, 1992). Sendo assim, é importante ressaltar que a hipertrofia
cardíaca concêntrica induzida pelo treinamento de resistência também é fisiológica
(FERNANDES et al, 2011; MAILLET et al, 2013).
De um modo geral, isso sugere que a hipertrofia fisiológica induzida pelo exercício
ou pela gravidez é provavelmente inofensiva e talvez até benéfica em indivíduos
saudáveis, contudo, na presença de doenças pré-existentes, como miocardiopatias
hereditárias, canalopatias arritmogênicas ou desequilíbrio angiogênico pode-se
desenvolver a doença e até mesmo a morte prematura em atletas e mães
(SARQUELLA-BRUGADA et al, 2012; MAILLET et al, 2013).
O exercício físico dinâmico realizado de forma crônica mostra-se um eficiente
aliado nas adaptações cardiovasculares (ELLIOTT et al, 2013; ROGNMO et al, 2012;
FREIMANN et al, 2005; STROM et al, 2005; WAGNER et al, 2002; SHEPARD e
BALADY, 1999; POWERS et al, 1998; BERLIN e COLDITZ, 1990). Entretanto, estudos
(WISLOFF et al, 2002; WISLOFF et al, 2007; SWAIN e FRANKLIN, 2006; ROGNMO et
al, 2004; ROGNMO et al, 2012; KEMI et al, 2002; KEMI et al, 2005) demonstraram que
o treinamento físico aeróbio de alta intensidade foi mais favorável para adaptações na
aptidão cardiorrespiratória e na função cardíaca do que o exercício de intensidade baixa
a moderada.
Entre os principais parâmetros cardiovasculares que sofrem adaptações com o
treinamento está a frequência cardíaca (FC) (de OLIVEIRA et al, 2010). O treinamento
aeróbico reduz tanto a FC em repouso (SBC, 2005; SCHAIBLE e SCHEUER, 1979;
GEENEN et al, 1988) como durante o exercício realizado em cargas submáximas de
trabalho e esses efeitos parecem decorrer da redução da hiperatividade simpática, do
aumento da atividade parassimpática, da mudança no marca-passo cardíaco ou mesmo
da melhora da função sistólica (SBC, 2005). Entretanto, apesar de o treinamento físico

38
induzir melhora da potência aeróbica máxima, ele não modifica de modo apreciável, a
FC máxima (SBC, 2005).
Outro parâmetro passível de adaptação com o treinamento é a pressão arterial
(PA), cuja manutenção é fundamental para a perfusão adequada dos tecidos e é
determinada por variáveis como o débito cardíaco (DC) e a resistência vascular
periférica (ANGHEBEN e OGIHARA, 2013). O treinamento físico reduz a pressão
arterial (PA) de repouso e durante o exercício submáximo, no entanto, da mesma forma
que ocorre com a FC, o treinamento parece provocar pouca alteração na PA máxima
aferida no pico do esforço (SBC, 2005).
A bradicardia de repouso tem sido utilizada como marcador dos efeitos do
treinamento físico aeróbio sobre o sistema cardiovascular e ocorre devido a adaptação
da função ventricular, verificada por um volume sistólico aumentado nos indivíduos
treinados, de modo a compensar essa redução na FC para gerar um DC de mesma
magnitude (de OLIVEIRA et al, 2010). Estudos tanto em humanos (WISLOFF et al,
2007) quanto em animais treinados (SCHAIBLE e SCHEUER, 1979; FULLER e
NUTTER, 1981; WISLOFF et al, 2002; KEMI et al, 2005; BOCALINI et al, 2010)
mostraram melhora na função ventricular.
Do ponto de vista da economia do organismo, a função crucial do coração é a
ejeção ventricular, pois o volume sistólico precisa ser assegurado em nível mínimo
compatível com a perfusão tissular (dos SANTOS e TUCCI, 2007). Nessa perspectiva,
o RC induzido pelo exercício físico é o responsável pela melhora da capacidade do
sistema cardiovascular e a principal resposta adaptativa é o aumento do volume de
ejeção que contribui para um maior DC observado em indivíduos treinados (BAGGISH
e WOOD, 2011; WEINER e BAGGISH, 2012).
O volume de ejeção ou débito sistólico corresponde à quantidade de sangue
ejetada pelos ventrículos a cada batimento, sendo proporcional à força determinada pela
contração do músculo cardíaco. Portanto, quanto maior a força de contração, maior será
o volume de ejeção (ANGHEBEN e OGIHARA, 2013). Entretanto, a força de contração
do músculo cardíaco depende do comprimento da fibra muscular no início da contração
e da característica de contratilidade do coração (ANGHEBEN e OGIHARA, 2013).
O estiramento do miocárdio devido a maiores enchimentos ventriculares eleva a
afinidade da troponina C pelo Ca2+ e também pode aumentar a liberação e absorção do
Ca2+ do retículo sarcoplasmático e esses dois efeitos aumentam a força desenvolvida
quando o comprimento da fibra do músculo cardíaco aumenta (LEVY et al, 2006). Assim,
quando o comprimento do sarcômero aumenta, ocorre aumento da tensão criada

39
durante a contração e, consequentemente, aumenta o volume de ejeção. Portanto, se
houver aumento da quantidade de sangue que chega aos ventrículos, as fibras se
alongarão mais e aumentará a força de contração, promovendo a ejeção de mais
sangue (ANGHEBEN e OGIHARA, 2013).
A Lei de Frank Starling determinou essa relação entre o estiramento e a força de
contração no coração sadio, ou seja, quanto maior a quantidade de sangue que chega
ao coração maior será a força de contração. (ANGHEBEN e OGIHARA 2013; LEVY et
al, 2006). Dessa forma, a contratilidade pode ser determinada experimentalmente como
a modificação no pico de força isométrica (pressão isovolumétrica) em um determinado
comprimento inicial da fibra (volume sistólico final) (LEVY et al, 2006).
2.5. Alterações na contratilidade miocárdica
Para discussão sobre as adaptações mecânicas ao exercício é importante
considerar que neste processo outras adaptações ocorrem simultaneamente. Dentre
essas, destaca-se a hipertrofia cardíaca, pois está relacionada diretamente com o
desenvolvimento da força celular (NATALI, 2004). Portanto, no treinamento de
resistência aeróbia, o alargamento do miocárdio ocorre acompanhado de função
contrátil reforçada (WISLOFF et al, 2001b).
A contratilidade dos cardiomiócitos depende da expressão de α e β-MCP, assim,
a miosina de cadeia pesada (MCP) é a principal proteína contrátil do coração e ela é
crucial para a eficiência do desempenho cardíaco. Além disso, a relação α/β-MCP varia
em resposta à sinalização fisiológica e patológica (FERNANDES et al, 2011).
Nesse aspecto, de OLIVEIRA (2010) esclareceram as características das
principais proteínas regulatórias que constituem o sarcômero: miosina de cadeia pesada
(MCP); miosina de cadeia leve (MCL1 e MCL2); tropomiosina, o complexo troponina
(TnT, TnI e TnC); e actina. No ventrículo da maioria das espécies de mamíferos,
inclusive o homem, foi identificada pelo menos uma das três isoformas de miosina (V1,
V2 e V3). Essas três isoformas são constituídas de somente dois tipos de MCP, α e β.
V1 e V3 são hemodímeros α/α e β/ β, respectivamente, enquanto V2 é heterodímeros
α/β (Figura 10) (FRANCHINI, 2001; de OLIVEIRA et al, 2010).
A α-MCP apresenta maior atividade ATPásica e maior velocidade de
encurtamento dos sarcômeros, enquanto a β-MCP apresenta menor velocidade de
encurtamento dos mesmos (SWYNGHEDAUW, 1986). Entretanto, a intensidade com
que esta alteração é observada na hipertrofia cardíaca depende do tipo de sobrecarga

40
e da espécie estudada (FRANCHINI, 2001). Em ratos a isoforma αα predomina em
corações normais enquanto em seres humanos a isoforma que predomina é a ββ
(SWYNGHEDAUW, 1999).
As alterações hemodinâmicas que ocorrem após o nascimento representam um
estímulo para a regulação dessas isoformas (de OLIVEIRA et al, 2010). Durante a vida
fetal, a grande maioria dos mamíferos expressa a β-MCP no ventrículo. Nos mamíferos
pequenos (rato e coelho), a α-MCP aumenta de forma rápida imediatamente antes do
parto e corresponde à isoforma dominante durante toda a vida adulta. Portanto, do ponto
de vista molecular, ocorre diminuição da expressão do gene da β-MCP ventricular e
aumento da expressão gênica da α-MCP (de OLIVEIRA et al, 2010). Nos mamíferos
maiores (cão, porco e homem), ocorre o contrário, a α-MCP é dominante apenas
transitoriamente após o nascimento, sendo o gene da β-MCP expresso de forma
dominante durante toda a vida. Nos átrios a situação é diferente, uma vez que a isoforma
α é expressa de forma dominante durante toda a vida, em todos os mamíferos (de
OLIVEIRA et al, 2010). Entretanto, a distribuição das isoformas da MCP pode ser
modificada em resposta a sobrecarga de trabalho, tanto em condições fisiológicas
quanto em condições patológicas.
No homem e em animais maiores em que a frequência cardíaca de repouso é
relativamente menor do que no rato, e a isoforma predominante no ventrículo da MCP
é exclusivamente a V3, o treinamento físico melhora a função ventricular sem modificar
a atividade ATPase miosínica ou a composição das isoformas da miosina (de OLIVEIRA
et al, 2010). Entretanto, em condições de sobrecarga patológica, o padrão de miosina
nos ventrículos não é modificado, uma vez que já predomina a isoforma V3 da miosina,
compreendendo cerca de 95% do total da miosina expressa. O que modifica na
hipertrofia cardíaca patológica é a quantidade de proteína expressa; a isoforma V3
aumenta para quase 100% (de OLIVEIRA et al, 2010).
No estudo de SCHAIBLE e SCHEUER (1981), ratos treinados com natação, sem
a presença de agentes patogênicos, demonstraram aumento na atividade da ATPase
miosínica no ventrículo esquerdo, o qual foi secundário ao aumento da expressão da
isoforma V1 (α-MCP), melhorando a função sistólica do animal treinado.
Por outro lado, nessa espécie com predomínio da isoforma V1 e alta atividade da
ATPase miosínica, a imposição de sobrecarga patológica como na hipertensão, resulta
em rápida mudança (dois a três dias) no padrão das isoenzimas para a isoforma V3 (β-
MCP), a qual está associada com a diminuição da atividade da ATPase miosínica (de
OLIVEIRA et al, 2010). Essas alterações impostas por condições patológicas levam, na

41
realidade, a uma mudança fisiológica importante que implica na expressão de um
fenótipo fetal, ou seja, na reprogramação gênica (de OLIVEIRA et al, 2010). A isoforma
de actina α, encontrada no músculo esquelético de animais adultos e no miocárdio na
fase embrionária também é expressa em ventrículo esquerdo submetido a sobrecarga
crônica (SWYNGHEDAUW, 1999).
No nível celular, a hipertrofia dos cardiomiócitos é caracterizada por aumento no
tamanho celular, melhora na síntese da proteínas e aumento na organização do
sarcômero (FREY et al, 2004). O rearranjo gênico induz a síntese proteica com o
aumento na produção de proteínas constituintes das membranas, das organelas e das
unidades morfofuncionais do coração: os sarcômeros (dos SANTOS e TUCCI, 2007).
Em paralelo, ocorre aumento do retículo sarcoplasmático e no número e tamanho das
mitocôndrias, para manter um estado funcional adequado ao número dos componentes
contráteis (dos SANTOS e TUCCI, 2007; GARCIA e INCERPI, 2008).
No RC fisiológico, decorrente do treinamento físico, crescimento ou gravidez, o
aumento de volume dos miócitos se faz pela síntese de novos componentes, como o
aumento no conteúdo das proteínas contráteis que levam ao aumento predominante
das miofibrilas, não ocorrendo grandes alterações nas características do estroma,
portanto sem prejuízo funcional do órgão (STROM et al, 2005; de OLIVEIRA et al, 2010;
FERNANDES et al, 2011). Em contraste, no RC patológico, como o que ocorre na
hipertensão arterial ou na cardiomiopatia hipertrófica, o aumento no volume dos
miócitos, provocado pelo aumento do número de miofibrilas é acompanhado de
aumento dos componentes do estroma, que em geral se apresentam de forma
desproporcional à resposta dos miócitos (WILLIAMS et al, 2000), sobretudo com
aumento do conteúdo do colágeno, podendo levar à deficiência no processo de
relaxamento do miocárdio (de OLIVEIRA et al, 2010).
O processo conhecido como acoplamento excitação-contração, responsável pela
contração muscular cardíaca, é o resultado do acoplamento entre o sinal elétrico que
chega à célula cardíaca e a geração da força mecânica (de OLIVEIRA et al, 2012). No
plano celular, o miócito ventricular cardíaco é a principal célula responsável pelo
acoplamento excitação-contração e consequente geração de força contrátil (de
OLIVEIRA et al, 2012).
Entretanto, para que o miócito cardíaco contraia, a concentração de Ca2+ deve
aumentar da faixa de nanomolar, encontrada na célula em repouso, na diástole, para a
micromolar, na sístole (de OLIVEIRA et al, 2012). Esse aumento na concentração de
Ca2+ intracelular, também conhecido como transiente de Ca2+ devido a sua duração

42
limitada, ocorre em cada ciclo cardíaco em resposta ao potencial de ação (de OLIVEIRA
et al, 2012). Adicionalmente, vários sensores intracelulares de cálcio estão presentes
na célula cardíaca, tais como a calsequestrina, calmodulina cinase II (CaMKII),
calmodulina, calcineurina e troponina C. Juntos, estes sensores de Ca2+ modulam o
acoplamento excitação-contração, assim como podem participar no RC patológico (de
OLIVEIRA et al, 2012).
A concentração do Ca2+intracelular livre é, portanto, o fator regulador crítico da
função dos miócitos cardíacos e o principal responsável pelo estado contrátil do
miocárdio. (FEARNLEY et al, 2011; LEVY et al, 2006; MARKS, 2003; BERS, 2001). Além
disso, o Ca2+ também controla numerosas outras atividades dos miócitos, incluindo a
transcrição do gene (FEARNLEY et al, 2011).
Mudanças sutis nos componentes da engrenagem da sinalização do Ca2+, ainda
que através de mutação, doença ou alteração crônica da demanda hemodinâmica, pode
ter consequências profundas para a função e fenótipo dos miócitos. (FEARNLEY et al,
2011; MARKS, 2003). Anormalidades na cinética do Ca2+ no cardiomiócito pode levar a
depressões do inotropismo miocárdico e do relaxamento. Além disso, evidências
apoiam que a intensidade das modificações guarda relação com a intensidade e o tempo
de atuação do agente despertador do remodelamento (dos SANTOS e TUCCI, 2007).
Para um melhor entendimento do fluxo do cálcio no ambiente celular, cabe
destacar o papel do retículo sarcoplasmático (RS) que é um sistema de membrana
intracelular em células cardíacas que desempenha um papel predominante no
acoplamento excitação-contração e contratilidade cardíaca (FRANK et al, 2003). Além
disso, o RS é a organela com maior foco de interesse científico nas repercussões do
RC, pois é provedora da maior parcela de Ca2+ ativador da contração permanecendo
armazenado no RS em ligação com a proteína calsequestrina (dos SANTOS e TUCCI,
2007).
Durante o potencial de ação, a permeabilidade do Ca2+ do sarcolema aumenta e
penetra na célula por meio dos canais para Ca2+ ativados pela voltagem no sarcolema
e nas invaginações, os túbulos T (Figura 5). Entretanto, a quantidade de cálcio que
penetra na célula do espaço extracelular não é suficiente para induzir a contração das
miofibrilas, mas serve como gatilho para liberar o Ca2+ do Ca2+ intracelular armazenado
no RS e esse processo é conhecido como libertação de cálcio induzido por cálcio (LEVY
et al, 2006; SEJERSTED, 2011; FEARNLEY et al, 2011). Dessa forma, os íons que se
interiorizam reagem com os receptores de rianodina (RyR) e promovem a abertura dos
canais liberadores de cálcio. Por esse mecanismo, grandes quantidades de cálcio são

43
liberadas no citoplasma a partir do RS (LEVY et al, 2006; SEJERSTED, 2011; de
OLIVEIRA et al, 2012). Nessas condições, a concentração de Ca2+ citosólica, que se
encontra na faixa de nanomolar, aumenta para a faixa de micromolar e isso é reflexo do
recrutamento de diferentes grupos de receptores de rianodina pela corrente de Ca2+
simultaneamente em toda a célula (de OLIVEIRA et al, 2012).
O Ca2+ liberado no citoplasma se liga à proteína troponina C que interage com a
tropomiosina para desbloquear os sítios ativos entre os filamentos de actina e miosina,
permitindo a contração das miofibrilas (sístole) (LEVY et al, 2006; SEJERSTED, 2011).
A miosina é dotada de capacidade ATPásica e hidrolisa o ATP, liberando o fósforo
dotado de energia que suporta a contração, ocorrendo, então, o desenvolvimento da
força e o encurtamento do miocárdio (dos SANTOS e TUCCI, 2007).
Ao final da sístole, o influxo de Ca2+ é interrompido e o RS não é mais estimulado
a liberar Ca2+. Nesse momento, o RS recaptura Ca2+ citosólico por meio de uma bomba
de cálcio, denominada Serca2a que é a principal estrutura responsável pela
manutenção da homeostasia do Ca2+ intracelular, envolvida no processo de
relaxamento miocárdico (de DEUS et al, 2012). Em humanos, a Serca2a remove 70 a
80% dos íons de cálcio (FRANK et al, 2003), já em ratos, esta remoção é de 92% (OPIE,
1998). A maior ou menor ação da Serca2a é regulada por outra proteína, a fosfolambam
(PLB) (dos SANTOS e TUCCI, 2007). Na sua forma não fosforilada, a proteína PLB inibe
a Serca2a. Quando fosforilada, o efeito inibidor é suprimido e a bomba de cálcio do RS
promove a recaptação de Ca2+ ocorrendo o relaxamento (diástole).
O aumento da expressão proteica da Serca2a parece melhorar o inotropismo
cardíaco, devido ao prolongamento do tempo de enchimento ventricular e ao maior
acúmulo de Ca2+ no RS (KEMI et al, 2008a). Outros estudos também mostraram que o
exercício físico aeróbio de alta intensidade aumentou a expressão proteica da Serca2a
(WISLOFF et al, 2001b; KEMI et al, 2007; KEMI et al, 2008a), da fosfolamban fosforilada
na treonina 17 (PLB-Tre17) (KEMI et al, 2007), sem alteração dos níveis proteicos da
FLB (KEMI et al, 2007, KEMI et al, 2008a), no ventrículo esquerdo de ratas
normotensas.
O trocador de Na/Ca também participa da homeostase do Ca2+, porém, sem gasto
energético. O objetivo do trocador Na/Ca é trocar, por meio da membrana, um átomo de
cálcio por três de sódio, no entanto, o sentido da troca depende das condições do pH,
da concentração intracelular de cada íon e do potencial da membrana. Em função
desses fatores a proteína trocadora pode interiorizar ou eliminar Ca2+ para manter a
homeostase do Ca2+ intracelular (LEVY et al, 2006; dos SANTOS e TUCCI, 2007).

44
Dessa forma, conclui-se que no cardiomiócito, o grande sítio do acoplamento
excitação-contração é o citosol, local onde o aumento do Ca2+ citosólico dispara a
contração celular (de OLIVEIRA et al, 2012). A concentração de cálcio no miócito
cardíaco é controlada de forma dinâmica, no entanto, na hipertrofia patológica ou
durante o desenvolvimento de arritmias cardíacas a regulação do Ca2+ estará
comprometida devido às diversas alterações nas proteínas envolvidas no controle do
acoplamento excitação-contração (de OLIVEIRA et al, 2012).
A hipertrofia dos cardiomiócitos é a base estrutural do sinal clínico do
remodelamento mais facilmente detectado na prática médica (dos SANTOS e TUCCI,
2007). Para avaliar a adaptação estrutural do miocárdio em humanos e animais, o
padrão geométrico do ventrículo esquerdo é obtido por meio do
Dopplerecocardiograma, com base no valor do índice de massa do ventrículo esquerdo
(MVE) e da espessura relativa da parede (ERP), que é calculada como a soma da
espessura do septo interventricular e da parede inferolateral dividida pelo diâmetro
diastólico do ventrículo esquerdo (TSUTSUI et al, 2010).
Outra técnica experimental comumente utilizada em ratos para estimar a
hipertrofia miocárdica é o estudo biométrico (ANTONIO et al, 2009; HELBER et al, 2009;
BOCALINI et al, 2010). Logo após o sacrifício dos animais e retirada do músculo papilar,
os ventrículos esquerdos (VE) e direitos (VD) são rapidamente pesados para avaliação
das massas brutas que são corrigidas pelas massas corpóreas de cada animal.
Figura 6. Fluxo do cálcio celular.

45
As proteínas marcadas na figura são as principais responsáveis pela homeostasia do Ca2+ intracelular: RyR, receptores de rianodina; PLB, fosfolambam; trocador Na+/Ca2 e Serca2a, Ca2+-ATPase. Organela: retículo sarcoplasmático, RS; TnC, troponina C. (Figura modificada de BERS, 2001).
Vários estudos (CHEN et al, 2001; SONG et al, 2013; VEIGA et al, 2013;
BOCALINI e TUCCI, 2009; BOCALINI et al, 2010; BOCALINI et al, 2014) testaram a
contratilidade em ratos por meio da análise de músculos papilares e, nesse aspecto, os
músculos papilares anteriores e posteriores do ventrículo esquerdo são maiores e mais
fortes quando comparados com aqueles encontrados no ventrículo direito
(GARBELOTTI e PELOZO, 2013). Dessa forma, para análise dos parâmetros
mecânicos convencionais no comprimento máximo (Lmáx), calculados em contração
isométrica (BOCALINI et al, 2010; SERRA et al, 2010), são utilizados
experimentalmente os músculos papilares e não as fibras musculares do próprio
miocárdio, pois estas são organizadas de forma bastante complexa (GARBELOTTI e
PELOZO, 2013), inviabilizando a análise desses parâmetros.
A mecânica da contração dos miócitos cardíacos pode ser estudada no
laboratório, examinando o comportamento de uma tira de músculo isolado e o músculo
papilar é conveniente para isso, pois suas fibras correm mais ou menos na mesma
direção (PINNELL et al, 2007). Uma tira de músculo é ancorada em ambas as
extremidades e é submetido a contração isométrica. Com o aumento do comprimento
inicial, a tensão gerada durante a contração isométrica cresce e a alteração no
comprimento da fibra inicial é análogo à pré-carga (PINNELL et al, 2007), que resulta
em um volume diastólico final ventricular esquerdo aumentado, expandindo assim o
comprimento da fibra. Isto produz um aumento da força de contração e um aumento do
volume da pulsação resultante da curva de Frank Starling (PINNELL et al, 2007).
A explicação para isso é que no comprimento normal de repouso, a sobreposição
de actina e miosina não é ótima. O aumento do comprimento inicial melhora o grau de
sobreposição e, por conseguinte, aumenta a tensão desenvolvida (PINNELL et al,
2007). O alargamento do músculo aumenta a sensibilidade da troponina ao cálcio
(sensibilidade de cálcio dependente de comprimento) e pode também conduzir a um
reforço de cálcio livre intracelular (PINNELL et al, 2007).
Estudos em miócitos cardíacos isolados (BOZI et al, 2013; KEMI et al, 2005;
NATALI, 2004; WISLOFF et al, 2002) também têm sido realizados com modelos animais
buscando revelar os mecanismos responsáveis pela melhora na função contrátil
(NATALI, 2004). Entender as alterações das propriedades mecânicas dos miócitos
cardíacos, a menor unidade contrátil do músculo cardíaco, em resposta ao exercício é

46
fundamental para explicar os mecanismos responsáveis pelas adaptações do músculo
cardíaco ao exercício crônico (NATALI, 2004). Isso porque os miócitos cardíacos
isolados demonstram propriedades contráteis similares às do músculo intacto (BRADY,
1991).
Os estudos com células isoladas indicaram que o exercício crônico não é capaz
de alterar a concentração intracelular de cálcio e a tensão passiva dos miócitos
cardíacos, entretanto, pode aumentar a sensibilidade dos miofilamentos ao cálcio,
especialmente quando os miócitos são estendidos, gerando uma maior tensão ativa
(NATALI, 2004).

47
3. MATERIAIS E MÉTODOS
3.1. Animais
Após aprovação do Comitê de Ética da Universidade São Judas Tadeu
(011/2014), 30 ratos fêmeas (Rattus norvegicus albinus, rodentia Mammalia), da
linhagem Wistar, com idade aproximada de 8 semanas, pesando 180–200g no início do
estudo, sem a presença de agentes patogênicos, com certificado de saúde animal
provenientes do Centro Multidisciplinar para Investigação Biológica (CEMIB –
UNICAMP, Campinas, SP), foram introduzidas no protocolo experimental. Optamos por
fêmeas, pois apresentam peso corpóreo menor do que os machos. A quantidade de
animais informada ao Comitê de Ética (180) considerou outros experimentos,
relacionados a este, que serão realizados posteriormente.
Os animais foram mantidos em caixas, fazendo uso “ad libitum” de água e ração
para ratos Nuvital CR-1, em ambiente com controle de temperatura em torno de 22ºC,
umidade do ar em 54% e ciclo claro/escuro invertido de 12 h claro e 12 h escuro (18:00-
06:00 horas: luzes acesas). A intensidade luminosa para o período de luz no biotério e
sala de laboratório foi mantida entre 50 e 60 lux e 300 e 400 lux, respectivamente. A
cada 48 horas foram trocadas as serragens das caixas e verificadas as condições dos
animais, da água e da ração.
Para a realização do estudo, os animais foram distribuídos aleatoriamente em três
grupos experimentais:
Sedentário (SED; n: 10): animais que não foram submetidos a nenhum tipo de
treinamento e permaneceram alojados durante 12 semanas.
Treinamento moderado (MOD; n: 10): animais que permaneceram alojados, mas
foram submetidos a exercício físico de intensidade moderada durante 12 semanas.
Treinamento intenso (INT; n: 10): animais que permaneceram alojados, mas foram
submetidos a exercício físico de alta intensidade durante 12 semanas.
3.2. Protocolo de exercício físico
O protocolo de exercício físico consistiu de natação, conforme protocolo
padronizado por MCARDLE (1967) e comumente utilizado pela literatura (BOCALINI et
al, 2010; VEIGA et al, 2011), em piscina construída utilizando caixa d’água de policloreto
de polivinila com diâmetro de 20 cm e altura de 60 cm.

48
O programa de treinamento foi individualizado e dividido em duas fases: Fase 1
(adaptação ao meio líquido), que possuiu duração de seis dias. O período de adaptação
ao meio líquido tem como finalidade reduzir o stress dos animais sem promover
adaptações fisiológicas ao treinamento físico. No primeiro dia, os animais foram
deixados nadando em piscina durante dez minutos. Nos dias seguintes, cada sessão de
exercício foi acrescida em dez minutos (BOCALINI et al, 2010). Ao final desse período,
os animais nadaram 60 minutos consecutivos por sessão. Após cada sessão de
natação, os ratos foram secados com toalha seca antes de serem devolvidos às suas
gaiolas.
Fase 2 (treinamento principal): os animais participaram de dois modelos diferentes
de treinamento, em sessões de natação de 60 minutos por dia, cinco vezes por semana,
até totalizar 12 semanas de treinamento.
Figura 7. Fases de treinamento.
Fase 1: Adaptação ao meio líquido. Fase 2: Treinamento principal.
O protocolo de natação linear de intensidade moderada foi realizado conforme
prévias publicações (MCARDLE, 1967; BOCALINI et al, 2010; VEIGA et al, 2011) sendo
constituído em natação com sobrecarga aderida a cauda do animal com peso relativo
de 2% do peso corpóreo. Já para o protocolo linear de alta intensidade, um peso relativo
variando entre 5% a 8% do peso corpóreo foi fixado na cauda do animal, conforme
proposto por VLADIMIR (2006). Neste protocolo o ajuste da carga foi alterado
semanalmente pelo peso corporal, contudo, o percentual de carga foi controlado de
maneira linear respeitando a seguinte ordem: 1º semana: sem carga; 2º semana: 5% do
peso corporal, da 3º a 6º semana: 6% do peso corporal, 7º a 8º semana: 7% do peso
corporal e da 9º a 12º semana: 8% do peso corporal. Para efeito de comparações o
protocolo de treinamento foi organizado em blocos sendo apresentado como: 1º bloco:

49
1º semana de treinamento; 2° bloco: 2º semana de treinamento; 3° bloco:
correspondente da 3º a 6º semana de treinamento, 4° bloco: correspondente da 7º a 8º
semana de treinamento e 5° bloco: correspondente da 9º a 12º semana de treinamento.
A intensidade utilizada no grupo de treinamento intenso, corresponde a carga de
treino predominantemente anaeróbia, conforme estudo de GOBATTO et al (2001) que
observaram estabilização do lactato sanguíneo em 5,5 mmol/l, correspondente a cargas
de 5 a 6% do peso corporal.
Foi quantificado a carga de treino (CT), o tempo total da sessão (TS) e o trabalho
total (TT) do programa de treino em cada bloco. Para a quantificação do trabalho
semanal total do programa em cada bloco de treino foi considerando a seguinte
equação:
TT semanal= (CT diário x TS diário) x 5, sendo 5 dias da semana.
A carga de trabalho semanal (TT), em unidades arbitrais (ua) foi quantificada pela
relação do volume (TS) e da intensidade (CT) de cada sessão de treinamento ao longo
dos 5 dias da semana (MUJIKA et al, 1995; de ARAUJO et al, 2011). O volume semanal
ou o tempo total da sessão (TS), foi a somatória dos minutos diários de treino
multiplicado por 5 dias da semana. Os ratos do grupo INT que não completaram a
sessão de treino, tiveram o tempo de treino efetivo registrado (minutos) (MUJIKA et al,
1995; de ARAUJO et al, 2011).
O trabalho total (TT) semanal do programa de treino em cada bloco foi o produto
entre o tempo (minutos) e a carga de treinamento (gramas), ajustada pelo peso corporal.
Cada grupo de animais teve o seu peso corporal total aferido semanalmente em
uma balança Filizolla® modelo EF1100 com acuidade de 1,0g (um grama).
A água da piscina foi constantemente agitada com auxílio de uma bomba para
evitar que os animais mergulhassem ou flutuassem ao longo das sessões de
treinamento. O aquecimento da água foi feito por meio de termostato associado ao
sistema de aquecimento elétrico promovendo manutenção da temperatura entre 32-
34°C, de acordo com recomendações para prescrição de exercícios para roedores
(KREGEL et al, 2006).
3.3. Experimento 1
Avaliações “in vivo”: no início e no final do treinamento.
3.3.1. Avaliação da capacidade física

50
A capacidade física foi avaliada em todos os grupos do experimento. No início e
no final da 1º, 2º, 6º, 8º e 12º semana de protocolo de treinamento os animais foram
pesados e pesos equivalentes a 10% do peso corporal foram fixados em seus tórax para
a avaliação da capacidade física. O tempo máximo de natação até a exaustão completa
do animal foi cronometrado, tendo como critério de exaustão a permanência submersa
por, pelo menos, 10 segundos conforme modelo utilizado no laboratório (BOCALINI et
al, 2010).
3.3.2. Ecodoplercardiograma
O ecodoplercardiograma foi realizado em todos os ratos dois dias após o termino
do protocolo de treinamento físico conforme técnica padronizada no laboratório (PERON
et al. 2005; ANTONIO et al. 2009; HELBER et al. 2009; VEIGA et al. 2011; VEIGA et al,
2013). Para tanto, as ratas foram anestesiadas (400 mg/kg de peso corporal) e
submetidas à tricotomia do tórax. Os animais foram posicionados em decúbito lateral
esquerdo para obtenção das imagens ecocardiográficas. Três eletrodos foram aderidos
às patas para registro simultâneo do traçado eletrocardiográfico, para identificação dos
acidentes do ciclo cardíaco e determinação da freqüência cardíaca. O aparelho utilizado
foi da marca Sonos 5500 (Philips, Andover, MA, USA), com capacidade de produzir
imagens nos Modos unidimensional e bidimensional e analisar a velocidade do fluxo
sanguíneo pela técnica de Doppler espectral e mapeamento de fluxo a cores. Foi
utilizado transdutor de fusão (5 a 12 MHz) com profundidade de 2 a 3 cm. Os registros
dos traçados do Modo M e do Doppler tiveram velocidade de 100 mm/s. As imagens
foram gravadas em fitas de vídeo VHS para análise posterior, sendo o resultado final
proveniente da média das obtidas de três ciclos cardíacos diferentes. Em todo o exame
foram seguidas as normas preconizadas pela American Society of Echocardiography.
A função ventricular foi analisada pela fração de encurtamento da área
transversa (FEAT), representada pela média das medidas obtidas nos planos basal,
médio e apical do ventrículo esquerdo, no corte transversal e paraesternal segundo a
fórmula:
FEAT= [(AD-AS/AD)] x 100%
sendo AD a área diastólica e AS a área sistólica.

51
3.4. Experimento 2
Avaliações “pós morten”
3.4.1. Biometria das massas cardíacas
O peso corporal de cada animal foi determinado no início, semanalmente durante
o período de treinamento e ao final do protocolo experimental. As massas cardíacas
foram determinadas conforme modelo já padronizado em nosso laboratório (ANTONIO
et al, 2009; HELBER et al, 2009; BOCALINI et al, 2010) e foram normalizadas pelo peso
corpóreo de cada animal: peso dos átrios (AT/PC), massa do ventrículo direito (VD/PC)
e massa do ventrículo esquerdo (VE/PC).
3.4.2. Avaliação da mecânica miocárdica
Após eutanásia, foi realizada toracotomia mediana, os corações foram retirados
rapidamente e colocados em solução de Krebs-Henseleit previamente aquecida a 29oC
e borbulhada com oxigênio conforme técnica padronizada utilizada em nosso laboratório
(PERON, 2005; BOCALINI e TUCCI, 2009; ANTONIO et al, 2009; HELBER et al, 2009;
BOCALINI et al, 2010; SERRA et al, 2010). A parede livre do ventrículo direito foi
extirpada com o objetivo de expor o septo interventricular, que foi dividido ao meio para
exposição dos dois músculos papilares do ventrículo esquerdo. Os corações foram
rapidamente transferidos para placa de Petri contendo solução de Krebs-Henseleit
adequadamente oxigenada e aquecida a 29oC e o músculo papilar posterior foram
cuidadosamente dissecado. A duração máxima de todo o procedimento, desde a
retirada do coração até a colocação do músculo papilar na cuba experimental foi de 5
minutos. Cada extremidade dos músculos foi fixada por anéis de aço inoxidável. Em
seguida, os músculos foram transferidos para câmara de vidro e permaneceram
submersos em solução de Krebs-Henseleit constantemente oxigenada por
borbulhamento e mantida em temperatura constante de 29oC. Um dos anéis foi fixado a
gancho existente no fundo da câmara de vidro e o outro ligado à extremidade de
transdutor de força (Modelo FT03E, Grass Instruments, Quincy, MA, USA) por meio de
fio de aço inoxidável. O transdutor de força foi conectado a micro manipulador (Modelo
2046F, Mitutoyo Sul Americana, São Paulo, SP, Brasil) que permitiu deslocamentos
verticais de 10 micrômetros a cada volta do micrométrico. A composição da solução de
Krebs-Henseleit, em milimoles por litro, foi de 132 NaCl; 4,69 KCl; 1,5 CaCl; 1,5 MgSO4;

52
1,18 KH2PO4; 5,50 glicose e 20 de HEPES como tampão. A pressão parcial de oxigênio
(pO2) da solução foi mantida entre 550 e 600 mmHg. Paralelamente ao músculo papilar
existiu dois eletrodos de platina que liberaram ativação de campo gerada por
estimulador (Modelo AVS-100, AVS Projetos Especiais, São Paulo, SP, Brasil). Os
estímulos elétricos foram liberados em ondas quadradas com duração de 5 ms e
voltagem aproximadamente 10 a 15% maior que o mínimo necessário para provocar
resposta mecânica do músculo, sob frequência de 0,2 Hz.
Durante período de 60 minutos os músculos foram deixados para contrair
isotonicamente contra pré-carga baixa. A contração isotônica foi possibilitada pela
existência de mola (tensão nominal de 0,3 g/cm) entre o anel superior e o transdutor, de
modo a permitir a ocorrência de encurtamento miocárdico. Em seguida, a mola foi
retirada do sistema e as contrações passaram a ser isométricas. Os músculos
permaneceram em contração isométrica durante 20 minutos para que a estabilização
da preparação pudesse ser alcançada. Com auxílio do micro manipulador o músculo foi
cuidadosamente estirado até atingir o ápice da curva comprimento/tensão (comprimento
diastólico do músculo no qual tensão isométrica desenvolvida atinge valor máximo:
Lmáx). Definido o estiramento do músculo que propiciou o maior valor da tensão
desenvolvida, foi medido o seu comprimento com auxílio de paquímetro Mitutoyo.
Após a determinação de Lmáx, os músculos permaneceram em contração
isométrica por 20 minutos e ao final deste período foram considerados como valores
basais correspondentes a 100% Lmáx os seguintes parâmetros: tensão isométrica
desenvolvida (TD); tensão isométrica de repouso (TR); taxa da variação da tensão
desenvolvida, sendo consideradas a taxa positiva (+dT/dt), negativa (-dT/dt) máximas,
tempo para TD atingir seu valor máximo (TPT) e tempo para TD decair 50% (TR50%).
Após obtenção dos dados em condições basais foram realizadas análises com os
seguintes protocolos experimentais:
Potenciação pós-pausa: a partir de pausas correspondentes a 5, 10, 15, 30, 45, 60 e
120 segundos a contração que se seguiu imediatamente à pausa foi usada para
fornecer informação sobre a função do retículo sarcoplasmático, sendo os dados
normalizados pela contração em condições basais (BOCALINI et al 2010).
Curva estiramento-tensão: Após os músculos permanecerem em nova estabilização
em Lmáx, a TD foi determinada nos comprimentos correspondentes a 92%, 94%, 96%,
98% Lmáx possibilitando a determinação da curva estiramento-tensão desenvolvida
(relação de Frank-Starling). A determinação da TR nos mesmos comprimentos

53
permitiu estimar a constante de rigidez miocárdica (BOCALINI et al, 2010; SERRA et
al, 2010).
Terminadas as manobras de avaliação da mecânica miocárdica, os músculos
foram retirados do sistema, o segmento contido entre os anéis de aço foi isolado e
pesado em balança modelo OCCU-124 (Fisher Scientific, Pittsburg, PA, USA).
Considerando-se a densidade muscular igual a 1, temos:
1 = m/V, onde m é a massa do músculo papilar e V o seu volume. Portanto:
m = V
Por outro lado, considerando o músculo papilar como um cilindro, o seu volume foi
determinado por:
V = L × AS, sendo
L o comprimento do músculo e AS a área de secção. Substituindo o valor de V temos:
m = L × AS e, portanto, AS = m /L
Os dados de tensão obtidos em grama foram corrigidos pela área de secção
(g/mm2) e posteriormente normalizados pelo peso do músculo (g/mm2/mg) conforme
estudos realizados em nosso laboratório (BOCALINI e TUCCI, 2009).
3.5. Análise estatística
O teste de D'Agostino-Pearson foi aplicado para análise da distribuição e
normalidade. Os dados foram expressos como médias ± erro padrão da média e
comparados por ANOVA um fator, dois fatores ou por medidas repetidas,
complementada com o teste de Tukey ou Bonferroni, conforme necessário. As análises
estatísticas foram realizadas com o auxílio do programa SPSS para Windows (versão
21.0, SPSS Inc., Chicago, Illinois, USA) com nível de significância de 5%.
A curva de estiramento-tensão desenvolvida (curva de Frank Starling) foi avaliada
por análise de regressão linear e as inclinações das retas (slopes) foram utilizadas como
parâmetros de comparação. Já a complacência miocárdica foi avaliada pela relação
estiramento-tensão diastólica ajustadas pela equação monoexponencial, como segue:
y = β0eβ1x, onde β0, β1 são constantes da curva. Essas relações não lineares foram
comparadas entre os grupos por valores de rigidez constante.

54
4. DESENHO EXPERIMENTAL
Como ilustrado na Figura 8, no início do Experimento 1, que se estende até o final
do período de treinamento, os animais foram pesados e submetidos ao teste de
capacidade física e ecodoplercardiograma. Após randomização, os grupos TM e TI
foram submetidos a 12 semanas de treinamento e foram pesados semanalmente. Ao
final da 12º semana, os animais foram submetidos ao Experimento 2 que avaliou a
biometria e a mecânica miocárdica.
Figura 8. Desenho experimental do estudo.
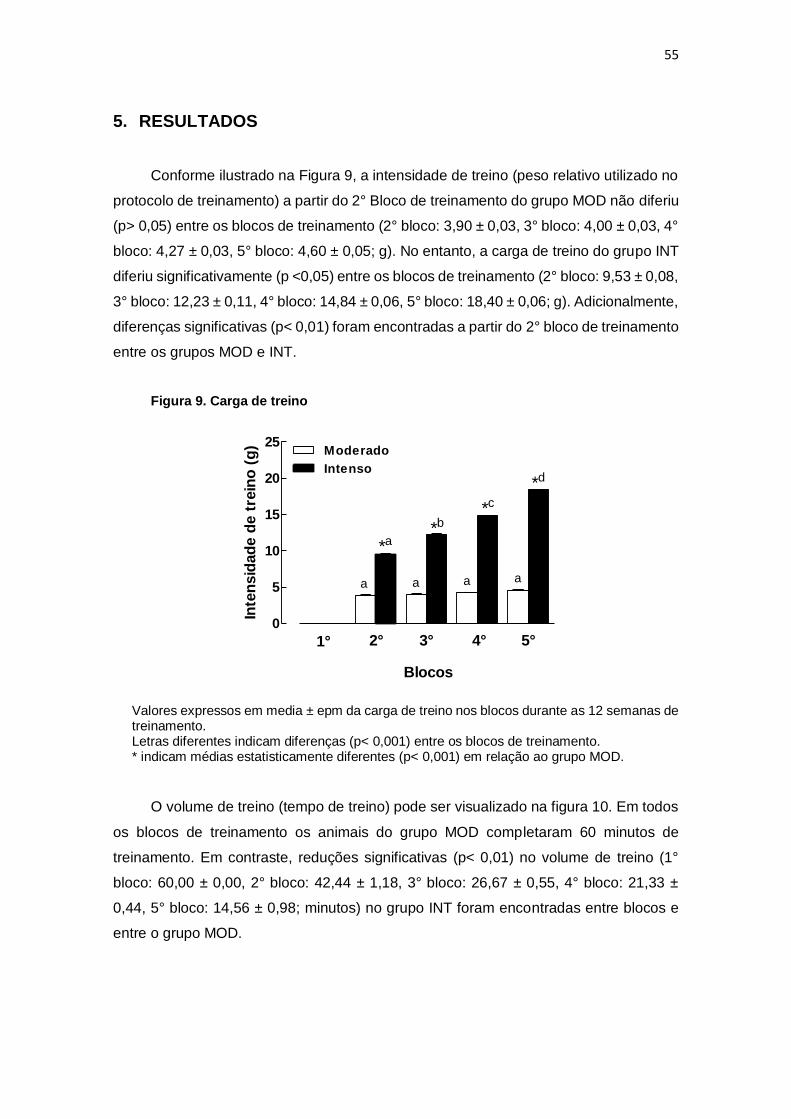
55
5. RESULTADOS
Conforme ilustrado na Figura 9, a intensidade de treino (peso relativo utilizado no
protocolo de treinamento) a partir do 2° Bloco de treinamento do grupo MOD não diferiu
(p> 0,05) entre os blocos de treinamento (2° bloco: 3,90 ± 0,03, 3° bloco: 4,00 ± 0,03, 4°
bloco: 4,27 ± 0,03, 5° bloco: 4,60 ± 0,05; g). No entanto, a carga de treino do grupo INT
diferiu significativamente (p <0,05) entre os blocos de treinamento (2° bloco: 9,53 ± 0,08,
3° bloco: 12,23 ± 0,11, 4° bloco: 14,84 ± 0,06, 5° bloco: 18,40 ± 0,06; g). Adicionalmente,
diferenças significativas (p< 0,01) foram encontradas a partir do 2° bloco de treinamento
entre os grupos MOD e INT.
Figura 9. Carga de treino
0
5
10
15
20
25Moderado
*a
1°
Blocos
Intenso
2° 3° 4° 5°
*b*c
*d
Inte
nsid
ad
e d
e t
rein
o (
g)
a a a a
Valores expressos em media ± epm da carga de treino nos blocos durante as 12 semanas de treinamento. Letras diferentes indicam diferenças (p< 0,001) entre os blocos de treinamento. * indicam médias estatisticamente diferentes (p< 0,001) em relação ao grupo MOD.
O volume de treino (tempo de treino) pode ser visualizado na figura 10. Em todos
os blocos de treinamento os animais do grupo MOD completaram 60 minutos de
treinamento. Em contraste, reduções significativas (p< 0,01) no volume de treino (1°
bloco: 60,00 ± 0,00, 2° bloco: 42,44 ± 1,18, 3° bloco: 26,67 ± 0,55, 4° bloco: 21,33 ±
0,44, 5° bloco: 14,56 ± 0,98; minutos) no grupo INT foram encontradas entre blocos e
entre o grupo MOD.
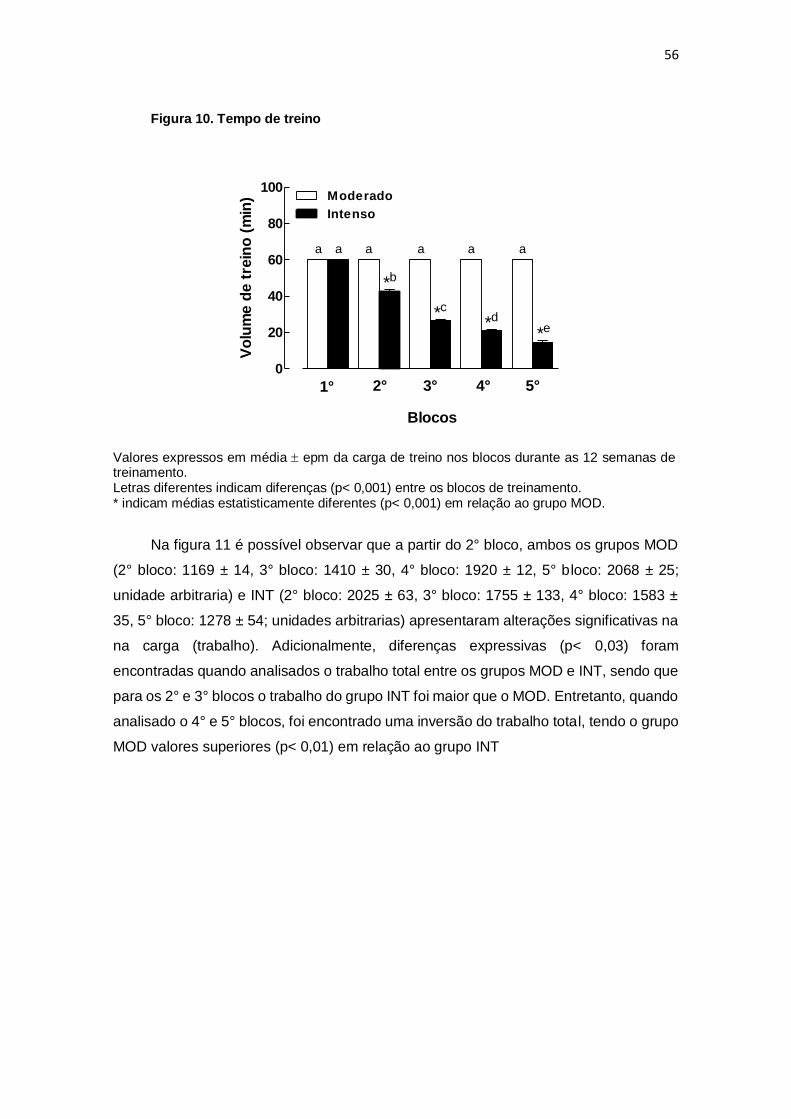
56
Figura 10. Tempo de treino
0
20
40
60
80
100Moderado
1°
Blocos
Intenso
2° 3° 4° 5°
*b
*c
*d
*e
aa a a a aV
olu
me d
e t
rein
o (
min
)
Valores expressos em média epm da carga de treino nos blocos durante as 12 semanas de treinamento. Letras diferentes indicam diferenças (p< 0,001) entre os blocos de treinamento. * indicam médias estatisticamente diferentes (p< 0,001) em relação ao grupo MOD.
Na figura 11 é possível observar que a partir do 2° bloco, ambos os grupos MOD
(2° bloco: 1169 ± 14, 3° bloco: 1410 ± 30, 4° bloco: 1920 ± 12, 5° bloco: 2068 ± 25;
unidade arbitraria) e INT (2° bloco: 2025 ± 63, 3° bloco: 1755 ± 133, 4° bloco: 1583 ±
35, 5° bloco: 1278 ± 54; unidades arbitrarias) apresentaram alterações significativas na
na carga (trabalho). Adicionalmente, diferenças expressivas (p< 0,03) foram
encontradas quando analisados o trabalho total entre os grupos MOD e INT, sendo que
para os 2° e 3° blocos o trabalho do grupo INT foi maior que o MOD. Entretanto, quando
analisado o 4° e 5° blocos, foi encontrado uma inversão do trabalho total, tendo o grupo
MOD valores superiores (p< 0,01) em relação ao grupo INT
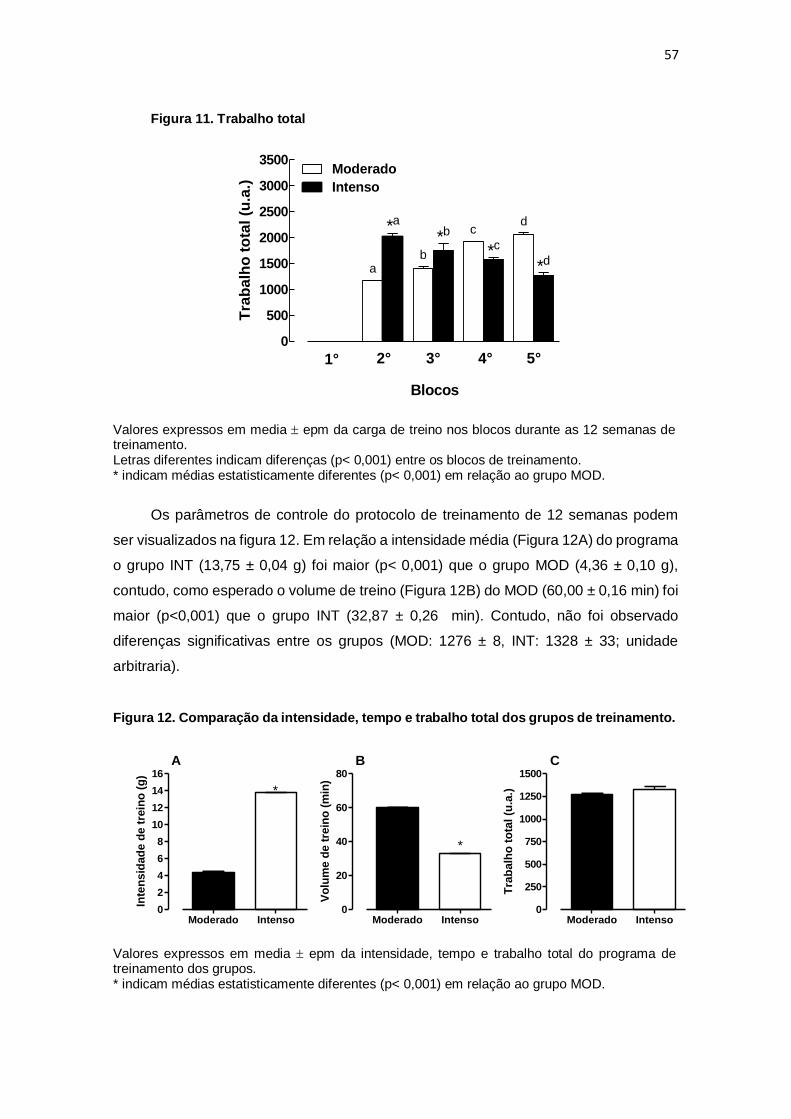
57
Figura 11. Trabalho total
0
500
1000
1500
2000
2500
3000
3500Moderado
*b
1°
Blocos
Intenso
*c
*d
2° 3° 4° 5°
*a
ab
cd
Tra
ba
lho
to
tal
(u.a
.)
Valores expressos em media epm da carga de treino nos blocos durante as 12 semanas de treinamento. Letras diferentes indicam diferenças (p< 0,001) entre os blocos de treinamento. * indicam médias estatisticamente diferentes (p< 0,001) em relação ao grupo MOD.
Os parâmetros de controle do protocolo de treinamento de 12 semanas podem
ser visualizados na figura 12. Em relação a intensidade média (Figura 12A) do programa
o grupo INT (13,75 ± 0,04 g) foi maior (p< 0,001) que o grupo MOD (4,36 ± 0,10 g),
contudo, como esperado o volume de treino (Figura 12B) do MOD (60,00 ± 0,16 min) foi
maior (p<0,001) que o grupo INT (32,87 ± 0,26 min). Contudo, não foi observado
diferenças significativas entre os grupos (MOD: 1276 ± 8, INT: 1328 ± 33; unidade
arbitraria).
Figura 12. Comparação da intensidade, tempo e trabalho total dos grupos de treinamento.
Moderado Intenso0
2
4
6
8
10
12
14
16A
*
Inte
ns
idad
e d
e t
rein
o (
g)
Moderado Intenso0
20
40
60
80B
*
Vo
lum
e d
e t
rein
o (
min
)
Moderado Intenso0
250
500
750
1000
1250
1500C
Tra
ba
lho
to
tal
(u.a
.)
Valores expressos em media epm da intensidade, tempo e trabalho total do programa de treinamento dos grupos. * indicam médias estatisticamente diferentes (p< 0,001) em relação ao grupo MOD.
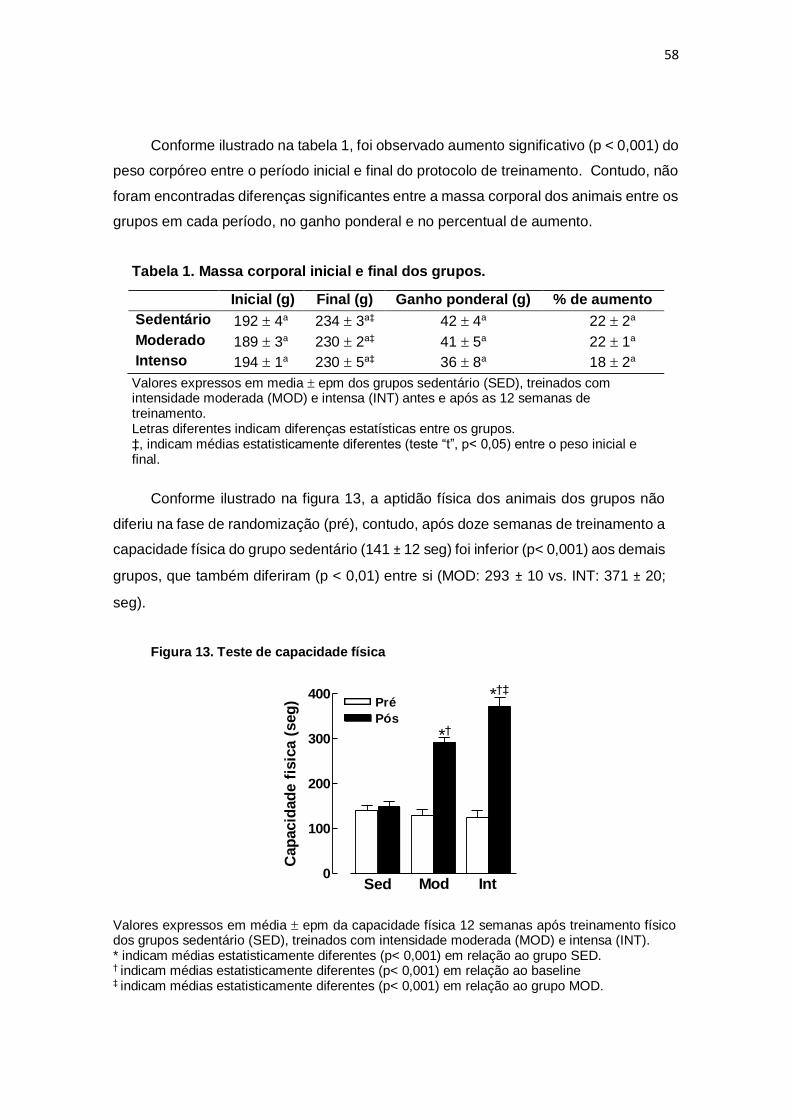
58
Conforme ilustrado na tabela 1, foi observado aumento significativo (p < 0,001) do
peso corpóreo entre o período inicial e final do protocolo de treinamento. Contudo, não
foram encontradas diferenças significantes entre a massa corporal dos animais entre os
grupos em cada período, no ganho ponderal e no percentual de aumento.
Tabela 1. Massa corporal inicial e final dos grupos.
Inicial (g) Final (g) Ganho ponderal (g) % de aumento
Sedentário 192 4a 234 3a‡ 42 4a 22 2a
Moderado 189 3a 230 2a‡ 41 5a 22 1a
Intenso 194 1a 230 5a‡ 36 8a 18 2a
Valores expressos em media epm dos grupos sedentário (SED), treinados com intensidade moderada (MOD) e intensa (INT) antes e após as 12 semanas de treinamento. Letras diferentes indicam diferenças estatísticas entre os grupos. ‡, indicam médias estatisticamente diferentes (teste “t”, p< 0,05) entre o peso inicial e final.
Conforme ilustrado na figura 13, a aptidão física dos animais dos grupos não
diferiu na fase de randomização (pré), contudo, após doze semanas de treinamento a
capacidade física do grupo sedentário (141 ± 12 seg) foi inferior (p< 0,001) aos demais
grupos, que também diferiram (p < 0,01) entre si (MOD: 293 ± 10 vs. INT: 371 ± 20;
seg).
Figura 13. Teste de capacidade física
0
100
200
300
400Pré
Pós
*†
*†‡
Sed Mod Int
Cap
acid
ad
e f
isic
a (
seg
)
Valores expressos em média epm da capacidade física 12 semanas após treinamento físico dos grupos sedentário (SED), treinados com intensidade moderada (MOD) e intensa (INT). * indicam médias estatisticamente diferentes (p< 0,001) em relação ao grupo SED. † indicam médias estatisticamente diferentes (p< 0,001) em relação ao baseline ‡ indicam médias estatisticamente diferentes (p< 0,001) em relação ao grupo MOD.
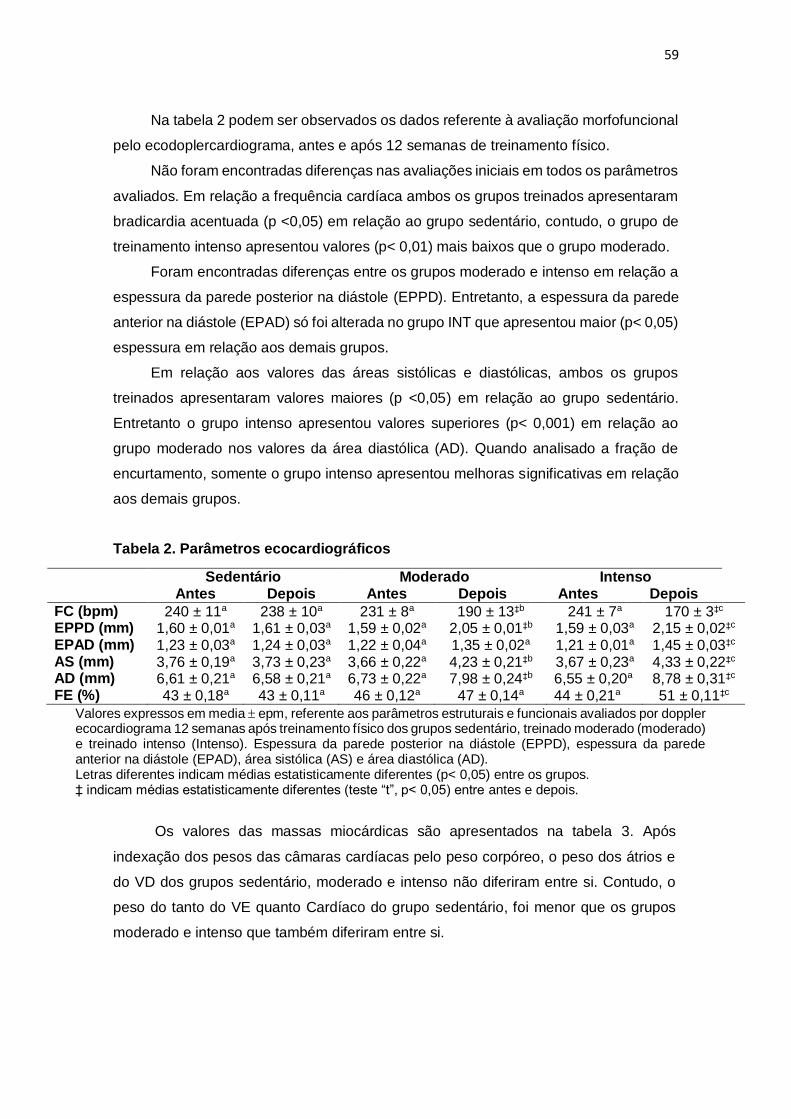
59
Na tabela 2 podem ser observados os dados referente à avaliação morfofuncional
pelo ecodoplercardiograma, antes e após 12 semanas de treinamento físico.
Não foram encontradas diferenças nas avaliações iniciais em todos os parâmetros
avaliados. Em relação a frequência cardíaca ambos os grupos treinados apresentaram
bradicardia acentuada (p <0,05) em relação ao grupo sedentário, contudo, o grupo de
treinamento intenso apresentou valores (p< 0,01) mais baixos que o grupo moderado.
Foram encontradas diferenças entre os grupos moderado e intenso em relação a
espessura da parede posterior na diástole (EPPD). Entretanto, a espessura da parede
anterior na diástole (EPAD) só foi alterada no grupo INT que apresentou maior (p< 0,05)
espessura em relação aos demais grupos.
Em relação aos valores das áreas sistólicas e diastólicas, ambos os grupos
treinados apresentaram valores maiores (p <0,05) em relação ao grupo sedentário.
Entretanto o grupo intenso apresentou valores superiores (p< 0,001) em relação ao
grupo moderado nos valores da área diastólica (AD). Quando analisado a fração de
encurtamento, somente o grupo intenso apresentou melhoras significativas em relação
aos demais grupos.
Tabela 2. Parâmetros ecocardiográficos
Sedentário Moderado Intenso
Antes Depois Antes Depois Antes Depois
FC (bpm) 240 ± 11a 238 ± 10a 231 ± 8a 190 ± 13‡b 241 ± 7a 170 ± 3‡c
EPPD (mm) 1,60 ± 0,01a 1,61 ± 0,03a 1,59 ± 0,02a 2,05 ± 0,01‡b 1,59 ± 0,03a 2,15 ± 0,02‡c
EPAD (mm) 1,23 ± 0,03a 1,24 ± 0,03a 1,22 ± 0,04a 1,35 ± 0,02a 1,21 ± 0,01a 1,45 ± 0,03‡c AS (mm) 3,76 ± 0,19a 3,73 ± 0,23a 3,66 ± 0,22a 4,23 ± 0,21‡b 3,67 ± 0,23a 4,33 ± 0,22‡c
AD (mm) 6,61 ± 0,21a 6,58 ± 0,21a 6,73 ± 0,22a 7,98 ± 0,24‡b 6,55 ± 0,20a 8,78 ± 0,31‡c
FE (%) 43 ± 0,18a 43 ± 0,11a 46 ± 0,12a 47 ± 0,14a 44 ± 0,21a 51 ± 0,11‡c
Valores expressos em media epm, referente aos parâmetros estruturais e funcionais avaliados por doppler ecocardiograma 12 semanas após treinamento físico dos grupos sedentário, treinado moderado (moderado) e treinado intenso (Intenso). Espessura da parede posterior na diástole (EPPD), espessura da parede anterior na diástole (EPAD), área sistólica (AS) e área diastólica (AD). Letras diferentes indicam médias estatisticamente diferentes (p< 0,05) entre os grupos. ‡ indicam médias estatisticamente diferentes (teste “t”, p< 0,05) entre antes e depois.
Os valores das massas miocárdicas são apresentados na tabela 3. Após
indexação dos pesos das câmaras cardíacas pelo peso corpóreo, o peso dos átrios e
do VD dos grupos sedentário, moderado e intenso não diferiram entre si. Contudo, o
peso do tanto do VE quanto Cardíaco do grupo sedentário, foi menor que os grupos
moderado e intenso que também diferiram entre si.

60
Tabela 3. Parâmetros biométricos.
Biometria cardíaca Sedentário Moderado Intenso
A (mg/g) 0,12 0,01 0,12 0,01 0,13 0,01 VD (mg/g) 0,60 0,01 0,66 0,02 0,61 0,03 VE (mg/g) 2,38 0,06 2,92 0,06* 3,33 0,15*‡ Car (mg/g) 2,98 0,05 3,44 0,07* 3,78 0,07*‡
Valores expressos em média epm dos parâmetros biométricos cardíacos dos grupos sedentário (SED), treinados com intensidade moderada (MOD) e intensa (INT) referentes a massa dos átriais (A), do ventrículo direito (VD), do ventrículo esquerdo (VE) e cardíaco (Car) normalizados pela massa corporal. * indicam médias estatisticamente diferentes (p< 0,001) em relação ao grupo SED. ‡indicam médias estatisticamente diferentes (p< 0,001) em relação ao grupo MOD.
Não foram encontradas diferenças entre os parâmetros biométricos do músculo
papilar (Tabela 4), no entanto, os parâmetros da função miocárdica podem ser
visualizados na tabela 5.
Tabela 4. Parâmetros biométricos dos músculos papilares.
Biometria papilar Sedentário Moderado Intenso
Massa do papilar (mg) 5,27 0,49 4,91 0,60 5,44 0,42 Lmax (mm) 6,65 0,43 6,50 0,35 6,98 0,35 AS (mm2) 0,79 0,06 0,78 0,10 0,73 0,06
Valores expressos em média epm dos grupos sedentário, treinados com intensidade moderada e intensa referente ao comprimento ótimo (Lmax) e área de secção transversa do músculo papilar (AS).
Os parâmetros da função sistólica (TD e +dT/dt) e diastólica (-dT/dt) dos animais
treinados foram superiores em relação aos animais sedentários (Tabela 5). Contudo,
ambos os parâmetros (sistólicos e diastólicos) do grupo INT foram superiores (p< 0,001)
em relação ao grupo MOD. Não foram encontradas diferenças entre os grupos na TR
bem como nas variáveis temporais (TPT e TR50%).
Tabela 5. Parâmetros da contratilidade miocárdica.
Sedentário Moderado Intenso
TD (g/mm2/mg) 0,85 0,4 1,07 0,1* 1,27 0,7*‡ +dT/dt (g/mm2/mg/s) 7 0,97 11 1,07* 15 0,59*‡ –dT/dt (g/mm2/mg) 5 0,34 7 0,52* 10 1,06*‡ TR (g/mm2/mg) 0,20 0,05 0,21 0,03 0,20 0,02 TPT (ms) 183 6 172 9 163 5 TR50% (ms) 142 9 148 10 147 4
Valores expressos em média epm dos grupos sedentário (SED), treinados com intensidade moderada (MOD) e intensa (INT) referentes a tensao desenvolvida (TD), as primeiras derivadas temporais de contração (+dT/dt) e relaxamento (-dT/dt), a tensão de repouso (TR), tempo para TD atingir o pico máximo (TPT) e o tempo para TD decair 50%.
* indicam médias estatisticamente diferentes (p< 0,001) em relação ao grupo SED.
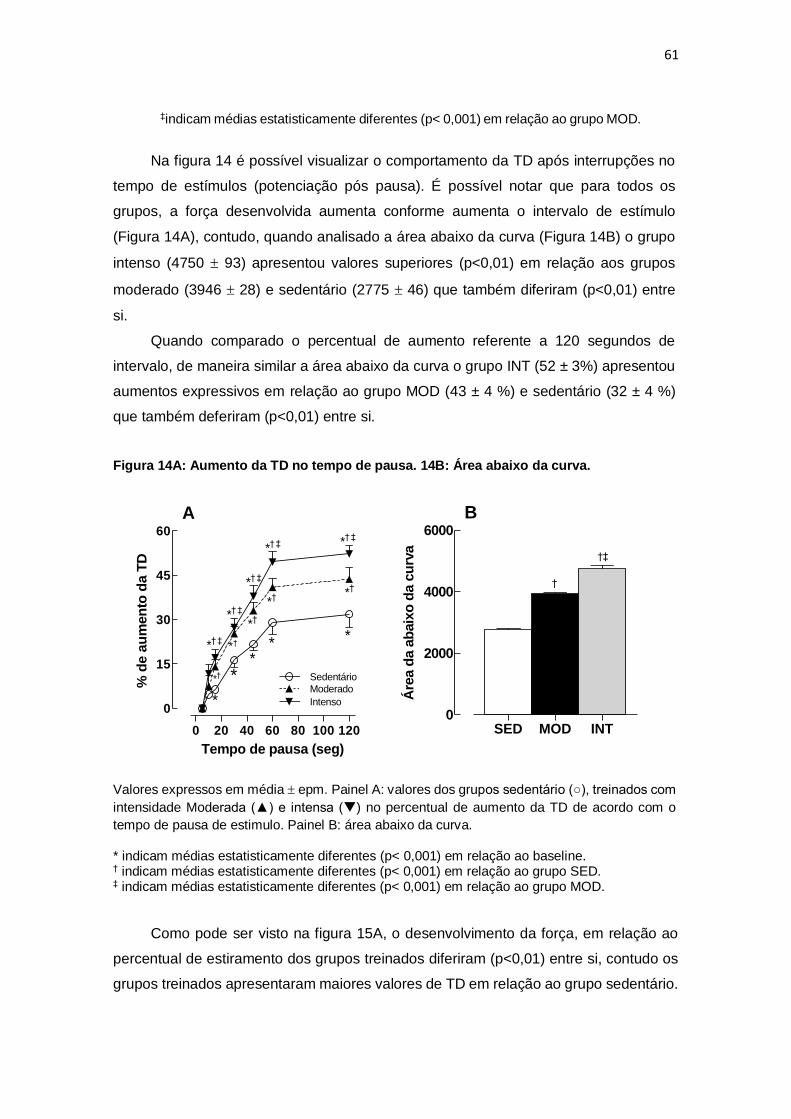
61
‡indicam médias estatisticamente diferentes (p< 0,001) em relação ao grupo MOD.
Na figura 14 é possível visualizar o comportamento da TD após interrupções no
tempo de estímulos (potenciação pós pausa). É possível notar que para todos os
grupos, a força desenvolvida aumenta conforme aumenta o intervalo de estímulo
(Figura 14A), contudo, quando analisado a área abaixo da curva (Figura 14B) o grupo
intenso (4750 93) apresentou valores superiores (p<0,01) em relação aos grupos
moderado (3946 28) e sedentário (2775 46) que também diferiram (p<0,01) entre
si.
Quando comparado o percentual de aumento referente a 120 segundos de
intervalo, de maneira similar a área abaixo da curva o grupo INT (52 ± 3%) apresentou
aumentos expressivos em relação ao grupo MOD (43 ± 4 %) e sedentário (32 ± 4 %)
que também deferiram (p<0,01) entre si.
Figura 14A: Aumento da TD no tempo de pausa. 14B: Área abaixo da curva.
0 20 40 60 80 100 120
0
15
30
45
60
Sedentário
Moderado
Intenso
*†
*
**
**
*†
*†
*† *†
*†‡
*†‡
*†‡
*†‡ *†‡
A
Tempo de pausa (seg)
% d
e a
um
en
to d
a T
D
SED MOD INT0
2000
4000
6000
B
Áre
a d
a a
baix
o d
a c
urv
a
†
†‡
Valores expressos em média epm. Painel A: valores dos grupos sedentário (○), treinados com
intensidade Moderada (▲) e intensa () no percentual de aumento da TD de acordo com o
tempo de pausa de estimulo. Painel B: área abaixo da curva.
* indicam médias estatisticamente diferentes (p< 0,001) em relação ao baseline. † indicam médias estatisticamente diferentes (p< 0,001) em relação ao grupo SED. ‡ indicam médias estatisticamente diferentes (p< 0,001) em relação ao grupo MOD.
Como pode ser visto na figura 15A, o desenvolvimento da força, em relação ao
percentual de estiramento dos grupos treinados diferiram (p<0,01) entre si, contudo os
grupos treinados apresentaram maiores valores de TD em relação ao grupo sedentário.
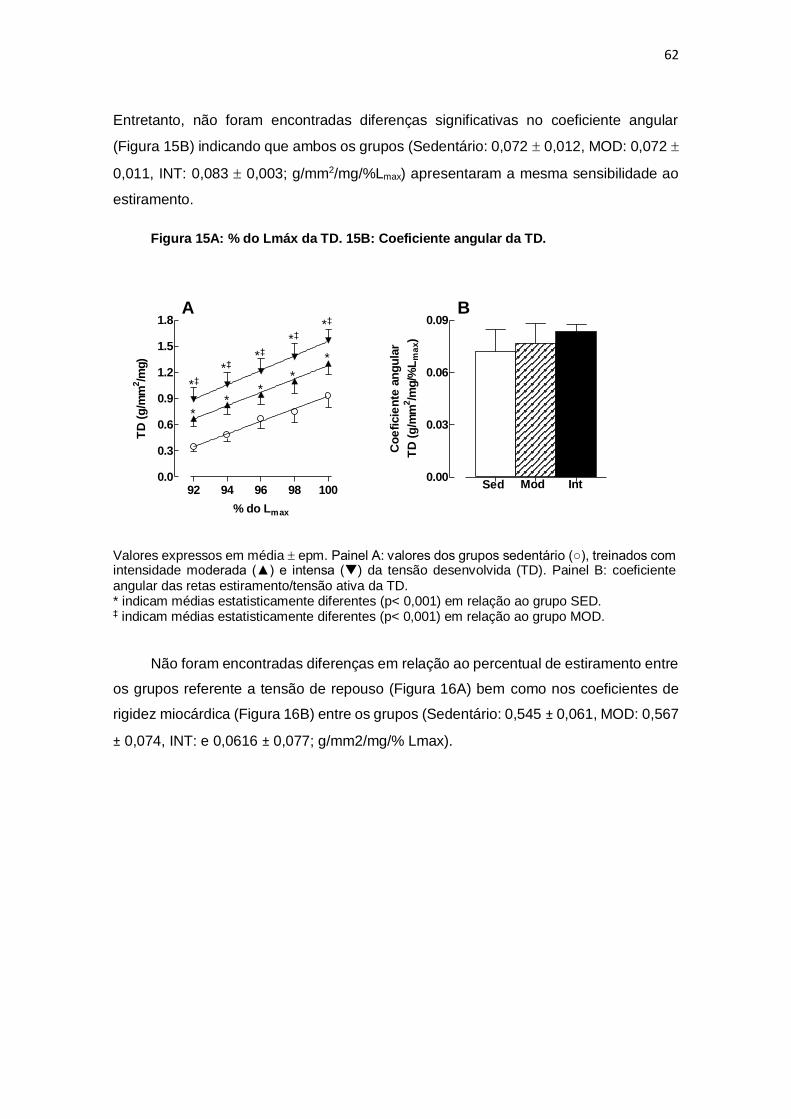
62
Entretanto, não foram encontradas diferenças significativas no coeficiente angular
(Figura 15B) indicando que ambos os grupos (Sedentário: 0,072 0,012, MOD: 0,072
0,011, INT: 0,083 0,003; g/mm2/mg/%Lmax) apresentaram a mesma sensibilidade ao
estiramento.
Figura 15A: % do Lmáx da TD. 15B: Coeficiente angular da TD.
92 94 96 98 1000.0
0.3
0.6
0.9
1.2
1.5
1.8A
*‡
*‡
*‡
*‡
*‡
**
**
*
% do Lmax
TD
(g
/mm
2/m
g)
0.00
0.03
0.06
0.09B
Sed Mod Int
Co
efi
cie
nte
an
gu
lar
TD
(g
/mm
2/m
g/%
Lm
ax)
Valores expressos em média epm. Painel A: valores dos grupos sedentário (○), treinados com intensidade moderada (▲) e intensa () da tensão desenvolvida (TD). Painel B: coeficiente angular das retas estiramento/tensão ativa da TD. * indicam médias estatisticamente diferentes (p< 0,001) em relação ao grupo SED. ‡ indicam médias estatisticamente diferentes (p< 0,001) em relação ao grupo MOD.
Não foram encontradas diferenças em relação ao percentual de estiramento entre
os grupos referente a tensão de repouso (Figura 16A) bem como nos coeficientes de
rigidez miocárdica (Figura 16B) entre os grupos (Sedentário: 0,545 ± 0,061, MOD: 0,567
± 0,074, INT: e 0,0616 ± 0,077; g/mm2/mg/% Lmax).
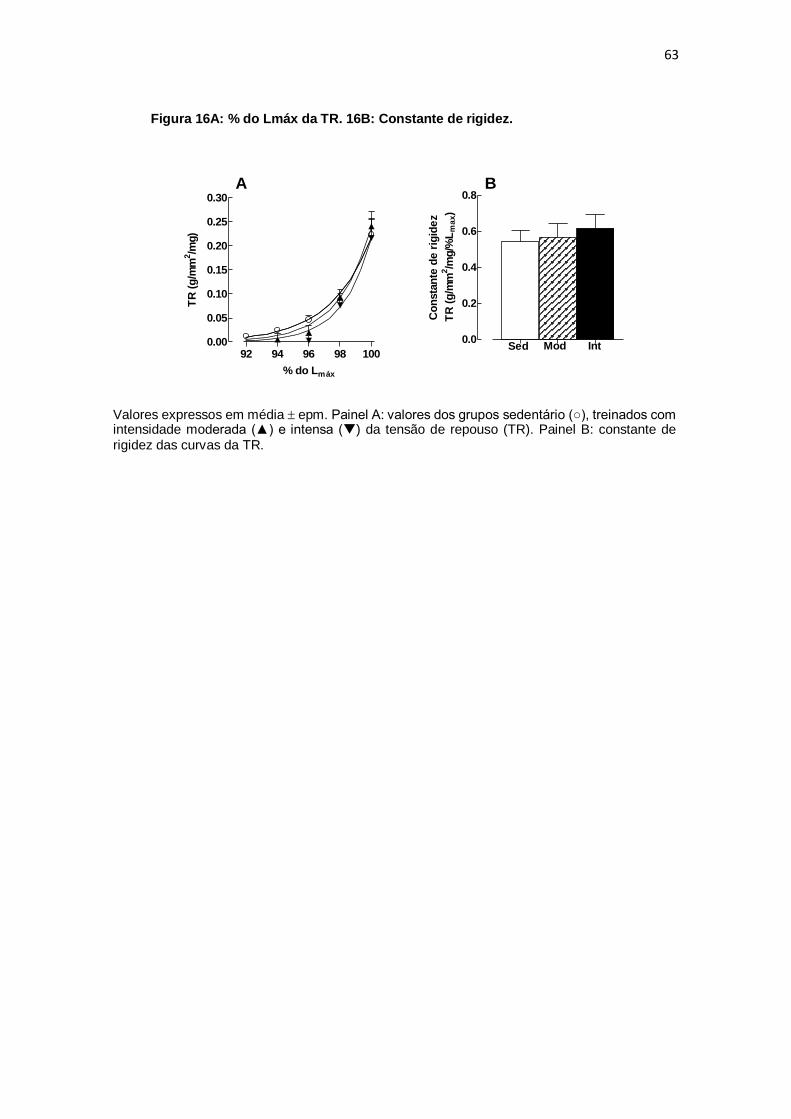
63
Figura 16A: % do Lmáx da TR. 16B: Constante de rigidez.
92 94 96 98 1000.00
0.05
0.10
0.15
0.20
0.25
0.30A
% do Lmáx
TR
(g
/mm
2/m
g)
0.0
0.2
0.4
0.6
0.8B
Sed Mod Int
Co
nsta
nte
de
rig
ide
z
TR
(g
/mm
2/m
g/%
Lm
ax)
Valores expressos em média epm. Painel A: valores dos grupos sedentário (○), treinados com intensidade moderada (▲) e intensa () da tensão de repouso (TR). Painel B: constante de rigidez das curvas da TR.

64
6. DISCUSSÃO
A ação do exercício físico regular e a melhora da capacidade funcional
apresentam-se como importantes fatores de implemento de saúde e os seus efeitos
benéficos sobre o sistema cardiocirculatório encontram-se bem descritos na literatura
(ELLIOTT et al., 2013; ROGNMO et al., 2012; FREIMANN et al., 2005; STROM et al.,
2005; WAGNER et al., 2002; SHEPARD e BALADY, 1999; POWERS et al., 1998;
BERLIN e COLDITZ, 1990). Entretanto, ainda há controvérsias sobre os efeitos
biológicos e clínicos entre os exercícios de moderada e alta intensidade (BARINAGA,
1997; MANSON et al., 2002; KEMI et al., 2005).
Há evidências consideráveis que apoiam o papel do baixo volume e alta
intensidade como sendo um método de treinamento potente e eficiente para induzir
adaptações tanto centrais (cardiovasculares) quanto periféricas (músculo esqueléticas),
ligadas a melhores resultados para a saúde e performance (GIBALA et al, 2009 e 2012).
Sabe-se que a variação dos parâmetros básicos de treinamento (frequência,
intensidade e duração) exercem importante influência no processo adaptativo (ADACHI
et al, 1996; TANASESCU et al, 2000; EVANGELISTA et al, 2003; KEMI et al., 2005)
sendo de suma importância a equalização destes parâmetros para discussões mais
homogêneas do real processo adaptativo oriundo de diferentes estratégias de
treinamento (TJONNA et al 2008; HARAM et al, 2009; ROGNMO et al 2004).
No presente estudo, houve a preocupação de equalizar o programa de
treinamento, tendo sido efetivo quando não observado diferença significativa no trabalho
total dos grupos treinados (Figura 12C), comprovando que as expressivas adaptações
cardíacas demonstradas pelo grupo intenso, foram efetivamente atribuídas ao programa
de alta intensidade. Estes resultados corroboram com outros estudos (KEMI et al 2005;
WISLOFF et al. 2002; WISLOFF et al. 2007; GIBALA et al. 2009, GIBALA et al. 2012;
ROGNMO et al. 2012; ELLIOTT et al. 2013) que avaliaram as adaptações do
treinamento de alta intensidade nas adaptações cardíacas, contudo, sem equalização
do programa de treinamento.
Existem alguns estudos, cuja proposta de periodização têm sido bem sucedidas
quando o intuito foi analisar adaptações bioquímicas e músculo esqueléticas (MUJIKA
et al, 1995; de ARAÚJO et al, 2013, de ARAÚJO et al 2012, de ARAÚJO et al 2007), no
entanto, para nosso conhecimento, não existe disponível na literatura estudos com
programa de treinamento periodizado em ratos avaliando as adaptações cardíacas. No
protocolo utilizado nesse estudo, o ajuste da carga foi alterado semanalmente pelo

65
percentual da massa corporal, técnica bastante utilizada na literatura (MEDEIROS et al,
2000; VOLTARELLI et al, 2002; MEDEIROS et al, 2004; EVANGELISTA et al, 2003;
VLADIMIR et al, 2006) quando o objetivo foi aumentar a intensidade em programas de
treinamento utilizando a modalidade natação.
Quando analisamos a massa corporal, observamos aumento significante em todos
os grupos experimentais após as doze semanas de observação no grupo SED e nos
grupos de treinamento MOD e INT. Embora não significativo, foi observado maior
tendência de redução (18%) no grupo INT quando comparado aos outros grupos.
Recentemente vem sendo discutido (RAMOS-FILHO et al, 2015; BOUTCHER,
2011; HARAM et al, 2009; TRAPP et al, 2008; WANG et al, 2008) que exercícios de alta
intensidade podem ser mais eficazes na redução de gordura corporal subcutânea e
abdominal do que exercício com menor intensidade. Nessa análise, estudos tanto em
humanos quanto em animais (RAMOS-FILHO et al, 2015; BOUTCHER, 2011; HARAM
et al, 2009; TRAPP et al, 2008; WANG et al, 2008; TJONNA et al, 2008) evidenciaram
que o treinamento de alta intensidade foi superior ao treinamento com intensidade
moderada na redução da massa e da gordura corporal. Em contraste, estudos utilizando
modelo animal (WISLOFF et al 2001a; WISLOFF et al, 2001b; DIFFEE e NAGLE, 2003;
KEMI et al,2005) apresentaram tendência de aumento na massa corporal de animais
dos grupos treinados.
Uma característica importante no controle, bem como na redução da massa
corporal é o controle da ingestão calórica (WASHBURN et al, 2014). Tendo em vista
que o treinamento físico induz a um aumento do consumo energético (McARDLE et al,
2008), associado ao não controle da ingestão calórica desses animais no presente
estudo, e a variação (ganho) da massa corporal natural de ratas fêmeas na margem
250 a 350 gramas na fase adulta (ILAR, 2003), é possível especular que a estabilização
da massa corporal dos grupos treinados em nosso estudo tenha ocorrido tanto pelo
crescimento natural do animal como, concomitante, pela perda de gordura corporal e
equilíbrio da demanda energética.
No que diz respeito ao aumento na capacidade física dos animais dos grupos
treinados, especial atenção pode ser direcionada ao resultado do grupo INT, que obteve
melhores resultados em relação ao grupo MOD. Esses resultados corroboram com
outros estudos (WISLOFF et al., 2007; TJONNA et al., 2008; ROGNMO et al., 2012;
ELLIOTT et al., 2013) que mostraram maior efetividade no ganho de capacidade física
e no controle de fatores de risco em treinamento de alta intensidade. Nesse aspecto, o
exercício é o estímulo fisiológico mais importante para o aumento da demanda de

66
oxigênio do miocárdio e, para suprir essa demanda, ocorrem adaptações centrais e
periféricas (EGAN et al, 2010; DUNCKER e BACHE, 2008).
Para um funcionamento satisfatório, a produção de energia do coração provém da
produção de energia oxidativa, resultante do metabolismo glicolítico (OPIE e
LOPASCHUK, 2004). Assim, os aumentos da atividade cardíaca dependem,
paralelamente, da disponibilidade de oxigênio (DUNCKER e BACHE, 2008). Em
repouso, o consumo de oxigénio normalizado por grama de miocárdio é 20 vezes mais
elevada do que a do músculo esquelético (LAUGHLIN et al, 1996). Dessa forma, para
se adaptar às elevadas exigências de oxigênio, o coração mantém um nível muito
elevado de extração de oxigênio do sangue arterial, em torno de 70 a 80%, superior à
do músculo esquelético, com 30 a 40% (LAUGHLIN et al, 1996).
Como consequência, o nível de extração de oxigênio no coração é facilitado por
uma elevada densidade capilar de 3,000 a 4,000 / mm2 (LAUGHLIN MH e TOMANEK,
1987), substancialmente mais elevada do que os capilares encontrados no músculo
esquelético, de 500 a 2000 / mm2 (GUTE et al, 1996). Devido ao alto nível de extração
de oxigênio pelo miocárdio durante condições de repouso, o aumento da demanda de
oxigênio produzido pelo exercício, em torno de até 6 vezes durante o exercício máximo,
é mediado principalmente por um aumento no fluxo sanguíneo coronariano (DUNCKER
e BACHE, 2008).
É importante salientar que o entendimento dos efeitos do exercício físico sobre a
rede de vasos sanguíneos é essencial para a prescrição de exercícios tanto na
prevenção quanto no tratamento de doenças, como nas cardiopatias ou angiopatias
(DENIPOTI et al, 2006). Nessa análise, o aumento do fluxo de sangue é responsável
pela dilatação no calibre dos vasos arteriais decorrente de um aumento real de tecido
endotelial e da musculatura lisa do vaso, fenômeno chamado de arteriogênese (EGAN,
2013; DUNCKER e BACHE, 2008).
Concomitantemente, ocorre a angiogênese, formação de novos capilares a partir
de capilares pré-existentes e, nesse aspecto, a hipóxia é reconhecidamente um potente
fator angiogênico (PRIOR et al., 2004). De acordo com DENIPOTI et al, (2006), a queda
nos níveis circulantes de oxigênio age como um estímulo para as células endoteliais
liberarem o fator de transcrição induzível por hipóxia (HIF) que, por sua vez, induz a
expressão do fator de crescimento endotelial vascular (VEGF). Dessa forma, a
expressão de VEGF é requerida no treinamento de endurance para induzir aumentos
das fibras capilares muscular e densidade capilar (BOOTH et al, 2015). Nessa linha,
acredita-se também que a produção de óxido nítrico (NO) seja o principal mecanismo

67
regulador da angiogênese, em resposta à pressão exercida pelo fluxo sanguíneo na
parede do vaso (PRIOR et al., 2004).
Além de melhorar a vasculatura e a estrutura contrátil, o exercício regular também
promove a remodelação da rede mitocondrial, apresentando expressiva melhora na
qualidade e na quantidade de mitocôndrias (BOOTH et al, 2015). Há evidências
(BOOTH et al, 2015; EGAN, 2013 e 2010; BURGOMASTER et al. 2008) que o conteúdo
mitocondrial da musculatura esquelética após treinamento aeróbio, promove
modificações positivas na capacidade respiratória e na extração de oxigênio,
melhorando a performance e resistência dos sujeitos treinados. O mecanismo para o
aumento do condicionamento físico aeróbio está associado a um teor elevado da
proteína peroxissoma ativado pelo proliferador do receptor gama coativador 1-alfa
(PGC-1α) (BOOTH et al, 2015; GIBALA et al, 2012; EGAN et al, 2013 e 2010; HARAM
et al, 2009; GIBALA et al, 2009; BURGOMASTER et al. 2008). Sabe-se que essa
proteína coordena a biogênese mitocondrial, interagindo com vários genes que
codificam para proteínas nucleares mitocondriais (EGAN, 2013; ROHAS et al 2007;
WENDE et al 2007).
Em 2000, GOTO et al. foram os primeiros a relatar que o PGC-1α aumentou
significativamente no músculo esquelético em ratos treinados por natação em
comparação ao grupo controle não exercitado. Mais tarde, outros grupos também
observaram maior teor de PGC-1α em modelo animal (TERADA et al, 2002) e seres
humanos (NORRBOM et al 2004). Evidências mais recentes (GIBALA et al, 2012; ENGA
et al., 2013 e 2010; GIBALA et al, 2009) sugerem que a intensidade do exercício é o
fator responsável pela ativação da PGC-1α no músculo esquelético humano.
O treinamento físico com natação é um eficiente aliado no aumento da atividade
oxidativa muscular e essa marcante adaptação do músculo esquelético ao treinamento
aeróbio também pode ser observada por meio do aumento da atividade da enzima
citrato sintase (CS) (EGAN, 2013; GIBALA et al, 2012; DIFFEE GM e NAGLE, 2003;
MEDEIROS et al, 2000), enzima que controla o primeiro passo do ciclo de Krebs.
RAMOS-FILHO et al (2015) demonstraram que depois de treinamento de baixo volume
e alta intensidade (HIIT), a atividade da CS aumentou 1,3 vezes no músculo tibial
anterior em ratos treinados com natação. MEDEIROS et al. (2000) observaram que os
ratos treinados com natação aumentaram a atividade da enzima citrato sintase no
músculo sóleo em 52%.
Outro ponto de extrema relevância diz respeito a adaptação crônica induzida pelo
ao exercício, sobretudo na resposta hipertrófica do cardiomiócito para adequação do

68
desempenho cardíaco às demandas do organismo (NATALI et al., 2004; GHORAYEB
et al., 2005). A hipertrofia cardíaca fisiológica é aquela desenvolvida em decorrência da
sobrecarga hemodinâmica transitória, como as observadas no crescimento cardíaco
durante a adolescência, na gestação e em resposta a exercícios regulares (MAILLET et
al., 2013; KEMPF E WOLLERT, 2004; DORN, 2007).
Não diferente de outros estudos (NUNES et al, 2013; BOCALINI et al, 2010; KEMI
et al, 2005; WANG et al, 2008; ANTONIO et al, 2009; HELBER et al, 2009; BOCALINI
et al, 2010; ZHANG et al, 2000a; ZHANG et al, 2000b; WISLOFF et al., 2001b; WISLOFF
et al, 2002) os dados referente ao aumento da massa cárdica, bem como as alterações
dos parâmetros estruturais analisados por meio do ecodoplercardiograma, são
considerados como indicadores de hipertrofia cardíaca nos animais treinados nessa
pesquisa.
O treinamento físico induz o coração à hipertrofia fisiológica, caracterizada pela
inexistência de fibrose e alteração na expressão de genes hipertróficos pós-natal, não
ocorrendo nesse caso alterações na expressão dos genes fetais que são classicamente
associados com o desenvolvimento de hipertrofia patológica (KEMI et al., 2007, 2008a,
2008b; WISLOFF et al., 2009; KEHAT E MOLKENTIN, 2010; MAILLET et al., 2013).
Isso sugere que a hipertrofia fisiológica depende mais da síntese de proteína por mRNA
transcrito, e da manutenção das proteínas sintetizadas (WISLOFF et al., 2009).
Nessa análise, a hipertrofia fisiológica é iniciada por sinais finitos, que incluem
hormonas de crescimento (tais como a hormona da tiróide, insulina, fator de crescimento
tipo insulina 1 (IGF-1) e fator de crescimento do endotélio vascular) e as forças
mecânicas que convergem para um pequeno número de vias de sinalização
intracelulares (tais como PI3K, AKT, ativada por AMP proteína quinase e mTOR) para
afetar a transcrição do gene, tradução proteica e o metabolismo (MAILLET et al., 2013;
MCMULLEN E JENNINGS et al., 2007).
Assim, neste tipo de hipertrofia, o aumento de volume dos miócitos se faz pela
síntese de novos componentes, como o aumento no conteúdo das proteínas contráteis
que levam ao aumento predominante das miofibrilas, não ocorrendo grandes alterações
nas características do estroma, portanto sem prejuízo funcional do órgão (STROM et
al., 2005, de OLIVEIRA, 2010, FERNANDES et al., 2011). Em paralelo, ocorre aumento
do retículo sarcoplasmático (RS) e no número e tamanho das mitocôndrias, para manter

69
um estado funcional adequado ao número dos componentes contráteis (dos SANTOS
e TUCCI, 2007; GARCIA e INCERPI, 2008).
Entretanto, a magnitude da hipertrofia cardíaca está diretamente relacionada ao
protocolo de treinamento utilizado (HASHIMOTO et al., 2011; FERNANDES et al., 2011;
WANG et al., 2010; BARROS, 1994). Nesse aspecto, a natação é frequentemente
utilizada em estudos de fisiologia do exercício envolvendo ratos, especialmente por
tratar-se de uma habilidade inata (KREGEL, 2006; de ARAUJO et al., 2007). Além disso,
estudos utilizando este modelo revelou semelhanças nas adaptações ao exercício em
relação àqueles observados em seres humanos (de ARAUJO et al., 2007; WANG et al.,
2010).
Nesse aspecto, o treinamento de natação em ratos é reconhecido por sua eficácia
em induzir hipertrofia cardíaca robusta quando comparado ao exercício em esteira
(WANG et al., 2010; SCHAIBLE e SCHEUER, 1981), no entanto, em ambos estudos de
KEMI (2002, 2007) com treinamento de alta intensidade em esteira, verificou-se um
aumento significativo no tamanho dos cardiomiócitos, bem como um reforço da
contratilidade e da função cardíaca, sugerindo que a alta intensidade nos programas de
esteira ou roda de corrida voluntária também parecem eficazes para indução de
hipertrofia fisiológica (WANG et al., 2010).
Considerando que a hipertrofia cardíaca fisiológica é uma forma suave de
crescimento, tipicamente caracterizada por um aumento de 10-20% no peso do coração,
normalizados pelo peso corporal e totalmente reversível (MAILLET et al., 2013), no
presente estudo foi verificado que o peso do VE no grupo intenso aumentou 39,91% e
o peso do coração aumentou 26,85%, comparados ao grupo sedentário. Nessa mesma
análise, o grupo moderado também apresentou hipertrofia significativa, com aumento
de 22,69% no peso do VE e 15,43% no peso do coração.
No estudo de MEDEIROS et al (2004) com ratos treinados com natação, utilizando
carga de 5% do PC, por 8 semanas, a aumento na massa ventricular esquerda e
cardíaca foi de 13%, respectivamente, quando comparado ao grupo sedentário. No
experimento de DIFFEE et al (2001), o treinamento em esteira incluiu um aumento de
5,9% na massa cardíaca, diferente de KEMI et al (2002) que demonstrou um aumento
no tamanho dos cardiomiócitos de 20 a 32% em ratos treinados em esteira com alta
intensidade.
Assim, no presente estudo, os grupos treinados foram induzidos ao
desenvolvimento de remodelamento cardíaco (aumento nas espessuras das paredes e
das áreas sistólicas e diastólicas), com maiores implementos no grupo que treinou em

70
alta intensidade, confirmando, assim, que a hipertrofia cardíaca proveniente do exercício
físico está associada a uma estrutura e função cardíaca normal ou aumentada
(McMULLEN et al., 2003; McMULLEN e JENNINGS, 2007; WISLOFF et al., 2009;
KEHAT e MOLKENTIN, 2010; BERNARDO et al., 2010; HASHIMOTO et al., 2011;
FERNANDES et al., 2011; MAILLET et al., 2013).
Quando analisamos a fração de encurtamento pela ecodoplercardiografia,
principal indicador de desempenho ventricular, nossos resultados mostraram que a
função cardíaca foi reforçada no grupo de treino intenso, que apresentou melhoras
significativas em relação aos demais grupos. De fato, o treinamento aeróbio melhora a
força de contração e aumenta as taxas de encurtamento de cardiomiócitos (KEMI et al,
2008c). Estudos tanto em humanos (WISLOFF et al., 2007) quanto em animais
treinados (SCHAIBLE e SCHEUER, 1979; FULLER e NUTTER, 1981; WISLOFF et al.,
2002; KEMI et al., 2005; BOCALINI et al., 2010) mostraram melhora na função cardíaca,
evidenciando aumento do débito cardíaco e do volume sistólico.
Ambos os grupos treinados apresentaram bradicardia acentuada em relação ao
grupo sedentário, contudo, o grupo intenso apresentou menores valores comparado aos
demais grupos. Nessa análise, sabe-se que a frequência cardíaca é mediada pela
atividade do sistema nervoso autônomo, por meio dos ramos simpático e
parassimpático, predominando a atividade vagal ou parassimpática em repouso e a
simpática durante o exercício (ALMEIDA e ARAÚJO, 2003). A bradicardia induzida pelo
exercício pode ainda ser consequência da adaptação intrínseca no nódulo sinusal
(CATAI et al, 2002; ALMEIDA e ARAÚJO, 2003).
Estudos sugerem que indivíduos bem condicionados com treinamento aeróbio,
possuem menor frequência cardíaca de repouso (MEDEIROS et al, 2000; ALMEIDA e
ARAÚJO, 2003; WANG et al 2008), indicando uma maior atividade parassimpática
(AUBERT et al, 2001; DIXON et al, 1992) ou menor atividade simpática (CHACON-
MIKAHIL et al, 1998). Valores menores de frequência cardíaca podem ocorrer ainda em
função de outros fatores decorrentes de um programa de treinamento, como o aumento
do retorno venoso e do volume sistólico (BONADUCE et al, 1998), bem como na
manutenção do débito cardíaco em repouso constante frequentemente encontradas em
indivíduos com melhor condicionamento aeróbico, independentemente da função
autonômica (CLAUSEN, 1977).
Vários estudos (CHEN et al., 2001; SONG et al., 2013; VEIGA et al., 2013;
BOCALINI e TUCCI, 2009; BOCALINI et al., 2010 e 2014) avaliaram a contratilidade
miocárdica em ratos por meio da análise de músculos papilares e, nesse aspecto, os

71
músculos papilares do ventrículo esquerdo são maiores e mais fortes quando
comparados com aqueles encontrados no ventrículo direito (GARBELOTTI e PELOZO,
2013). No presente estudo, foi utilizado a sugestão de estudos prévios (BOCALINI e
TUCCI 2009, BOCALINI et al. 2010, SERRA et al. 2010) realizados, utilizando o músculo
papilar posterior.
A influência do comprimento do músculo sobre a contratilidade (mecanismo de
Frank-Starling) é um fator crucial na contração do miocárdio (NATALI, 2004). Os
resultados da mecânica miocárdica observados nessa investigação, demonstraram que
os grupos de treinamento moderado e intenso tiveram melhoras na função sistólica (TD
e +dT/dt) e diastólica (–dT/dt) do miocárdio com destaque para o grupo intenso.
Contudo, similar a outros estudos (BOCALINI et al, 2014; WISLOFF et al, 2001b), nesse
experimento, não foi observado melhora do desempenho do mecanismo da curva
estiramento-tensão (FIGURA 15B) indicando que todos grupos experimentais
apresentaram a mesma sensibilidade ao estiramento.
Entretanto, quando nos referimos a força contrátil (TABELA 5), é possível
visualizar um aumento expressivo nos grupos treinados. Uma possível explanação para
tal melhora relaciona-se à variação na concentração da cinética do Ca2+ intracelular
como regulador crítico do inotropismo miocárdico (FEARNLEY et al., 2011; LEVY et al.,
2006; MARKS, 2003; BERS, 2002).
No presente estudo não verificamos a expressão das proteínas responsáveis pela
manutenção da homeostase do Ca2+ intracelular, no entanto, foi possível notar que a
força contrátil aumentou com consequente aumento do intervalo de estímulo na análise
da contração pós-pausa, indicando, mesmo que indiretamente, melhora da cinética do
Ca2+ com concomitante aumento do inotropismo miocárdico (BOCALINI et al, 2012;
PIESKE et al, 1996).
Já é conhecido na literatura (RAVENS et al.; 1995; BERS et al., 1993; MILL et al.,
1992; WENDT-GALLITELLI & ISENBERG, 1992; SCHOUTEN, 1990; VASSALO &
MILL, 1988; MANTELLI et al., 1984; RAVENS, 1992; ALLEN, et al., 1977) que a
potenciação pós pausa é um fenômeno que acontece em alguns tecidos cardíacos, no
qual o primeiro batimento é potencializado e regularizado com os subsequentes, após
o prolongamento de uma pausa. Esse fenômeno conta com alterações na liberação do
Ca2+ do estoque intracelular durante a contração, pela atividade dos trocadores
Na+/Ca2+, e alterações na SERCA 2a, com mudanças na captura de Ca2+ pelo retículo
sarcoplasmático (SOARES et al. 2005; DAMIANI et al., 2004; RAVENS et al., 1995;

72
MILL et al. 1992; VASSALO & MILL, 1988; BOSE et al., 1988; ABREU et al., 1987,
SUTSKO et al., 1986; ALLEN et al., 1977).
Alterações na taxa e ritmo são variáveis comumente utilizadas como manobras
experimentais para avaliar o comportamento ou anormalidades na cinética do Ca2+, no
acoplamento excitação-contração (ENDOH, 2004; BOCALINI et al, 2012). Vários
estudos relataram que o aumento da contratilidade está associado à frequência de
estímulos no músculo cardíaco (ENDOH, 2004; BERS, 2001; SCHOUTEN et al., 1987;
LEWARTOWSKI & PYKOWSKY, 1987; RAVENS, 1992). Em contraste, utilizando
amostra de ratos infartados, BOCALINI et al (2012) confirmaram que a redução na força
de contração cardíaca foi atribuída à captação reduzida do Ca2+ pela SERCA2a e pela
excessiva expressão do trocador Na+/ Ca2+.
Ao final da sístole, o influxo de Ca2+ é interrompido e o RS não é mais estimulado
a liberar Ca2+. Nesse momento, o RS recaptura Ca2+ citosólico por meio de uma bomba
de cálcio, denominada Serca2a que é a principal estrutura responsável pela
manutenção da homeostasia do Ca2+ intracelular, envolvida no processo de
relaxamento miocárdico (de Deus et al., 2012). Em humanos, a Serca2a remove 70 a
80% dos íons de cálcio (Frank et al., 2003), já em ratos, esta remoção é de 92% (Opie,
1998). A maior ou menor ação da Serca2a é regulada por outra proteína, a fosfolambam
(PLB) (Levy et al., 2006; dos Santos e Tucci, 2007). Na sua forma não fosforilada, a PLB
inibe a Serca2a. Quando fosforilada, o efeito inibidor é suprimido e a bomba de cálcio
do RS promove a recaptação de Ca2+ ocorrendo o relaxamento (diástole) (Levy et al.,
2006).
Nossos resultados mostraram que após interrupções no tempo de estímulos,
ocorreu um aumento na força de contração de 52% no grupo que treinou intenso e de
43% no grupo de treino moderado, sendo superiores a resposta em relação ao grupo
SED (32%). Embora sendo uma medida indireta, os resultados da análise da
potenciação pós pausa, associado a um aumento da função contrátil apresentada pelos
grupos treinados, com resposta superior para o grupo INT, torna-se um dos maiores
achados desse estudo como um dos mecanismos responsáveis por justificar tal
aumento.
Nessa linha, KEMI et al. (2005) e KEMI et al. (2008a) mostraram que o aumento
da expressão proteica da Serca2a parece melhorar o inotropismo cardíaco, devido ao
prolongamento do tempo de enchimento ventricular e ao maior acúmulo de Ca2+ no
reticulo sarcoplasmático, demonstrando que a sensibilidade do Ca2+ e, por conseguinte,
a atividade máxima da SERCA-2a, aumentou 40% e 30% em alta e moderada

73
intensidade, respectivamente. Adicionalmente, outros estudos também mostraram que
o exercício físico aeróbio de alta intensidade é eficiente para aumentar a expressão
proteica da Serca2a (WISLOFF et al., 2002; KEMI et al., 2007), da fosfolamban
fosforilada na treonina 17 (PLB-Tre17) (KEMI et al., 2007), sem alteração dos níveis
proteicos da FLB (KEMI et al., 2007; KEMI et al., 2008a) no ventrículo esquerdo.

74
7. CONCLUSÃO
Os resultados desse estudo confirmam que o exercício aeróbio realizado de forma
crônica é um eficiente aliado nas adaptações cardiovasculares, em alta e moderada
intensidade. Contudo, o treinamento aeróbio de alta intensidade foi mais favorável em
promover adaptações na aptidão cardiorrespiratória, no aumento da massa cardíaca e
na função contrátil do que o exercício de intensidade moderada.
A maior limitação desse estudo foi o fato de não ter sido verificado a expressão
das proteínas envolvidas na cinética do cálcio, como a Serca 2a e a fosfolamban,
contudo, com as avaliações realizadas nesse trabalho, permite-nos especular a
existência de uma forte relação dessas proteínas na função contrátil e no processo de
indução da hipertrofia miocárdica nos grupos de treinamento. Nessa perspectiva, mais
estudos são necessários para uma melhor compreensão dos mecanismos associados
às adaptações cardíacas do treinamento aeróbio em diferentes intensidades.

75
8. REFERÊNCIAS
ABREU, G. R., VASSALO, D. V.; MILL, J. G. The Na+-Ca2+ exchange mechanism
as a regular of post rest contractions in cardiac muscle. Brazilian J. Med. Biol.
Res., 20: 817-820, 1987.
ADACHI H, KOIKE A, OBAYASHI T, UMEZAWA S, AONUMA K, INADA M. Does
appropriate endurance exercise training improve cardiac function in patients with
prior myocardial infarction? Eur Heart J, 17:1511 – 21, 1996.
ALLEN, D. G. On the relationship between action potential duration and tension in
cat papillary muscle. Cardiovasc. Res., v.11, p.210-218, 1977.
ALLEN D.L, HARRISON B.C, MAASS A, BELL M.L, BYRNES W.C, LEINWAND
L.A. Cardiac and skeletal muscle adaptations to voluntary wheel running in the
mouse. Journal of Applied Physiology, 90: 1900-1908, 2001.
ALMEIDA M e ARAÚJO CGS. Efeitos do treinamento aeróbico sobre a freqüência
cardíaca. Rev Bras Med Esporte _ Vol. 9, Nº 2, 2003.
ALVES GB, ROVEDA F, CAMARGO EW, NUNES N, NERY SS, da SILVA CEG,
de OLIVEIRA PA. “Reabilitação cardiovascular e condicionamento físico”. In:
Cardiologia do Exercício – Do atleta ao cardiopata, Manole. v.3, 2010.
AMERICAN COLLEGE OF SPORTS MEDICINE POSITION STAND. Exercise for
patients with coronary artery disease. Med Sci Sports Exerc, 26:i– v, 1994
AMUNDSEN B.H, ROGNMO O, HATLEN-REBHAN G, SLORDAHL S.A. High
intensity aerobic exercise improves diastolic function in coronary artery disease.
Scand. Cardiovasc. J., 42:110Y7, 2008.
ANGHEBEN JMM e OGIHARA CA, “Fisiologia cardiovascular”. In: Reabilitação
cardiovascular e metabólica. 1ª ed. São Paulo. Atheneu, 2013.
ANTONIO EL, SANTOS AA, ARAUJO SRR, BOCALINI DS, dos SANTOS L,
FENELON G, FRANCO MF, TUCCI PJF. Left Ventricle radio-frequency Ablation
in the rat: a new model of heart failure due to myocardial infarction homogeneous
in size and low in mortality. J Cardiac Failure, 15(6): 540-548, 2009.
AUBERT AE, BECKERS F, RAMAEKERS D. Short-term heart rate variability in
young athletes. J Cardiol. 37: S85-8, 2001.
BAGGISH AL e WOOD MJ. Athlete's heart and cardiovascular care of the athlete:
scientific and clinical update. Circulation.123 (23):2723-35, 2011.
BARINAGA M. How much pain for cardiac gain? Science, 276:1324– 7, 1997.

76
BARROS NTL. Hipertrofia ventricular secundária ao exercício físico.Rev Soc
Cardiol Est São Paulo; 4: 376-81, 1994.
BASSETT DR Jr e HOWLEY ET. Limiting factors for maximum oxygen uptake and
determinants of endurance performance. Medicine & Science in Sports & Exercise,
32 (1), 70-84, 2000.
BELARDINELLI R, GEORGIOU D, CIANCI G, PURCARO A. Randomized,
controlled trial of long-term Moderate exercise training in chronic heart failure:
effects on functional capacity, quality of life, and clinical outcome. Circulation,
1173– 82, 1999.
BENITO B, GAY-JORDI G, SERRANO-MOLLAR A, et al. Cardiac arrhythmogenic
remodeling in a rat model of long-term intensive exercise training.
Circulation.123:13-22, 2011.
BERLIN J.A e COLDITZ G.A. A meta-analysis of physical activity in the prevention
of coronary heart disease. Am J Epidem, 132: 612-618, 1990.
BERNARDO BC, WEEKS KL, PRETORIUS L, R. McMULLEN JR. Molecular
distinction between physiological and pathological cardiac hypertrophy:
Experimental findings and therapeutic strategies. Pharmacology & Therapeutics.
191–227, 2010.
BERS, D. M.; BASSANI, R. A.; BASSANI, J. W. M.; BAUDET, S.; HRYSHKO, L.
V. Paradoxical twitch potentiation after rest in cardiac muscle: incresead fractional
release of SR calcium. J. Mol. Cell. Cardiol., 25: 1047-1057, 1993.
BERS, D. M. lç ed 2, Kluwer Academic,Dordrecht, Netherlands, 2001.
BHELLA PS, HASTINGS JL, FUJIMOTO N, SHIBATA S; CARRICK-RANSON G,
PALMER DM, BOYD KN, ADAMS-HUET B, LEVINE BD. Impact of Lifelong
Exercise “Dose” on Left Ventricular Compliance and Distensibility. J Am Coll
Cardiol. 64(12):1257-1266, 2014.
BOCALINI DS e TUCCI PJF. Developed force of papillary muscle: what index
correctly indicates contractile capacity? Int Heart J, 50(5): 643-652, 2009.
BOCALINI D.S, VEIGA E.C.A, SOUZA A.F.M, LEVY R.F, TUCCI P.J.F. Exercise
training-induced enhancement in myocardial mechanics is lost after two-weeks of
detraining in rats. Eur J Appl Physiol, 109: 909-914, 2010.
BOCALINI DS, BEUTEL A, BERGAMASCHI CT, TUCCI PJ, CAMPOS RR.
Treadmill Exercise Training Prevents Myocardial Mechanical Dysfunction Induced
by Androgenic-Anabolic Steroid Treatment in Rats PLoS ONE 9(2): e87106, 2014.

77
BOCALINI DS, dos SANTOS L; ANTONIOLL EL; dos SANTOS AA; DAVELL AP;
ROSSONI L; VASSALO DV; TUCCI PJF. Myocardial remodeling after large
infarcts in rat converts post rest-potentiation in force decay. Arq. Bras. Cardiol.
vol.98 no.3, 2012.
BONADUCE D, PETRETTA M, CAVALLARO V, APICELLA C, IANNICIELLO A,
ROMANO M. Intensive training and cardiac autonomic control in high level
athletes. Med Sci Sports Exerc. 30:691-6, 1998.
BOOTH FW, TSENG BS, FLÜCK M, CARSON JA. Molecular and cellular
adaptation of muscle in response to physical training. Acta Physiol Scand,
162:343-50, 1998.
BOOTH FW, RUEGSEGGER GN, TOEDEBUSCH RG, YAN Z. Endurance
Exercise and the Regulation of Skeletal Muscle Metabolism. Prog Mol Biol Transl
Sci.135:129-51, 2015.
BOSE, D.; HRYSHKO, L. V., KING, B. W., CHAU, T. Control of interval-force
relation in canine ventricular myocardium studied with ryanodine. Britsh J.
Pharmacol., 95: 811-820, 1988.
BOUTCHER SH. High-Intensity Intermittent Exercise and Fat Loss. J Obes, 2011.
BOZI LHM, MALDONADO IRSC, BALDO MP, da SILVA MF, MOREIRA JBN,
NOVAES RD, RAMOS RMS, MILL JG, BRUM PC, FELIX LB, GOMES TNP,
NATALI AJ. Exercise training prior to myocardial infarction attenuates cardiac
deterioration and cardiomyocyte dysfunction in rats. Clinics (Sao Paulo). 68(4):
549–556, 2013.
BRADY, A.J. Mechanical properties of isolated cardiac myocytes. Physiol. Rev.
71: 413 – 428, 1991.
BURGOMASTER KA, HOWARTH KR, PHILLIPS AM, Mark RAKOBOWCHUK M,
MACDONALD MJ, McGEE SL and GIBALA MJ. Similar metabolic adaptations
during exercise after low volume sprint interval and traditional endurance training
in humans. The Journal of Physiology. Volume 586, Issue 1, pages 151–160, 2008.
CARVALHO JF, MASUDA MO, POMPEU FA. Method for diagnosis and control of
aerobic training in rats based on lactate threshold. Comp Biochem Physiol A Mol
Integr Physiol;140(4):409-13, 2005.
CATAI AM, CHACON-MIKAHIL MP, MARTINELLI FS, FORTI VAM, SILVA E,
GOLFETTI R. Effects of aerobic exercise training on heart rate variability during
wakefulness and sleep and cardiorespiratory responses of young and middle-age
healthy men. Brazilian J Med Biol Res. 35: 741-52, 2002.

78
CHACON-MIKAHIL MPT, FORTI VAM, CATAI AM, SZRAJER JS, GOLFETTI R,
MARTINS LEB. Cardiorespiratory adaptations induced by aerobic training in
middle-age men: the importance of a decrease in sympathetic stimulation for the
contribution of dynamic exercise tachycardia. BrazilianJ Med Biol Res.31:705-12,
1998.
CHEN J, FELLER GM, BARBATO J C, PERIYASAMY S, XIE Z-J, KOCH LG,
SHAPIRO JI, BRITTON SL. Cardiac performance in inbred rat genetic models of
low and high running capacity. J Physiol; 535(Pt 2): 611–617, 2001.
CLAESSENS C, CLAESSENS P, BLOEMEN H, CLAESSENS M, VERBANCK M,
FAGARD R, ClAESSENS J. Structural heart adaptations in triathletes. Acta
Cardiologica; 54(6):317-325, 1999.
CLAUSEN JP. Effect of physical training on cardiovascular adjustments to exercise
in man. Physiol Rev. 57:779-815, 1977.
COHN J.N, FERRARI R, SHARPE N. Cardiac remodeling--concepts and clinical
implications: a consensus paper from an international forum on cardiac
remodeling. Behalf of an International Forum on Cardiac Remodeling. J Am Coll
Cardiol, 35(3):569-82, 2000.
CRAIG BW, MARTIN G, BETTS J, LUNGREN M, LAMBRET V and KAISERAUER
S. The influence of training-detraining upon the heart, muscle and adipose tissue
of female rats. Mech Ageing Dev, 57:49-61, 1991.
DAMIANI, C. E. N.; MOREIRA, C. M.; ZHANG, H. T.; CREAZZO, T. L.; VASSALO,
D. V. Effects of eugenol, na essential oil, on the mechanical and electrial activities
of cardiac muscle. J. Cardiovasc. Pharmacol, 44(6): 688-695, 2004.
de ARAUJO GG, PAPOTI M, MANCHADO FB, MELLO MA, GOBATTO CA.
Protocols for hyperlactatemia induction in the lactate minimum test adapted to
swimming rats. Com Biochem Physiol A Mol Integr Physiol, 148: 888-892, 2007.
de ARAUJO GG, PAPOTI M, dos REIS IGM, de MELLO MAR, GOBATTO CA.
Physiological responses during linear periodized training in rats. European Journal
of Applied Physiology, Volume 112, Issue 3, pp 839-852, 2012.
de ARAUJO GG, PAPOTI M, DELBIN MA, ZANESCO A, GOBATTO CA.
Physiological adaptations during endurance training below anaerobic threshold in
rats. Eur J Appl Physiol. 113(7):1859-70, 2013.
de DEUS AF, LIMA-LEOPOLDO AP, LEOPOLDO AS, CICOGNA AC, ALVES
CAB, MAZETO CFS, da SILVA DCT, de CAMPOS DHS, de TOMASI LC, FREIRE
PP. “Bomba de Cálcio do Retículo Sarcoplasmático (SERCA) e Bomba de Cálcio

79
da Membrana Plasmática (PMCA)”. In: Sinalização de Cálcio: Bioquímica e
Fisiologia Celulares, Sarvier, 2012.
DENIPOTI CH, MORAES SMF, HERNANDES L. Angiogênese e exercício. Arq
Mudi. 10(2):17-22, 2006.
de OLIVEIRA EM, ALVES GB, BRUM PC, KRIEGER JE. “Aspectos moleculares
da hipertrofia dos músculos cardíaco e esquelético após treinamento físico”. In:
Cardiologia do Exercício – Do atleta ao cardiopata, Manole. v.3, 2010.
de OLIVEIRA MG, LADEIRA MS, ROCHA-RESENDE C, GUATIMOSIM S.
“Sinalização de Ca2+ em Cardiomiócitos”. In: Sinalização de Cálcio: Bioquímica
e Fisiologia Celulares, Sarvier, 2012.
DIFFEE GM, SEVERSEN EA, TITUS MM. Exercise training increases the Ca2+
sensitivity of tension in rat cardiac myocytes. J Appl Physiol, 91:309–15, 2001;
DIFFEE GM e NAGLE DF. Exercise training alters length dependence of
contractile properties in rat myocardium. J Appl Physiol, 94:1137– 44, 2003.
DIXON E, KAMATH MV, MCCARTNEY N, FALLEN E. Neural regulation of the
heart rate in endurance athletes and sedentary controls. Cardiovasc Res. 26:713-
9, 1992.
DORN GW. The fuzzy logic of physiological cardiac hypertrophy. Hypertension.
49(5):962-70, 2007.
dos SANTOS L e TUCCI P. R. Cardiac remodeling: the cardiomyocyte, the cardiac
function. Rev. Soc. Cardiol. Estado de São Paulo;17(3):196-205, 2007.
DUNCKER DJ e BACHE RJ. Regulation of Coronary Blood Flow During Exercise.
Physiol Rev 88: 1009–1086, 2008.
ELLIOTT A, BENTLEY D, AROMATARIS E. Effectiveness of high-intensity interval
training in patients with coronary heart disease: a systematic review protocol. JBI
Database of Systematic Reviews & Implementation Reports;11(8) 13- 22, 2013.
EGAN B, CARSON BP, GARCIA-ROVES PM, CHIBALIN AV, SARSFIELD FM,
BARRON N, McCAFFREY N, MOYNA NM, ZIERATH JR and O’GORMAN DJ.
Exercise intensity-dependent regulation of peroxisomeproliferator-activated
receptor γ coactivator-1α Mrna abundance is associated with differential activation
of upstream signalling kinases in human skeletal muscle. J Physiol 588.10 pp
1779–1790, 2010.
EGAN B, ZIERATH JR. Exercise metabolism and the molecular regulation of
skeletal muscle adaptation. Cell Metab. 17(2):162-84, 2013.

80
ENDOH M. Relação força-freqüência no miocárdio ventricular de mamífero
intacta: relevância fisiológica e fisiopatológica. Eur J Pharmacol. 500 (1-3): 73-86,
2004.
EVANGELISTA FS, BRUM PC and KRIEGER JE. Duration-controlled swimming
exercise training induces cardiac hypertrophy in mice Braz J Med Biol Res vol.36
no.12, 2003
FEARNLEY CJ, RODERICK HL, BOOTMAN MD. Calcium signaling in cardiac
myocytes. Cold Spring Harb Perspect Biol.3(11):a004242, 2011.
FERNANDES T, SOCI UP, OLIVEIRA EM. Eccentric and concentric cardiac
hypertrophy induced by exercise training: microRNAs and molecular determinants.
Brazilian Journal of Medical and Biological Research; 44: 836-847, 2011.
FITZSIMONS D.P, BODELL P.W, HERRICK R.E, BALDWIN K.M. Left ventricular
functional capacity in the endurance-trained rodent. Journal of Applied Physiology,
69: 305-312, 1990.
FLETCHER G.F, BALADY G.J, AMSTERDAM E.A, CHAITMAN B, ECKEL R,
FLEG J, FROELICHER V.F, LEON A.S., PIÑA I.L., RODNEY R., SIMONS-
MORTON D.A., WILLIAMS M.A BAZZARRE T. Exercise standards for testing and
training: a statement for healthcare professionals from the American Heart
Association. Circulation, 104:1694– 740, 2001.
FLORESCU M e VINEREANU D. How to differentiate athlete’s heart from
pathological cardiac hypertrophy? A Journal of Clinical Medicine, Volume1 No.3,
2006.
FRANCHINI KG. Hipertrofia cardíaca: mecanismos moleculares. Rev
BrasHipertens, v. 8, p. 125–142, 2001.
FRANK KF, BÖLCK B, ERDMANN E, SCHWINGER RH. Sarcoplasmic reticulum
Ca2+-ATPase modulates cardiac contraction and relaxation. Cardiovasc Res.
57(1):20-7, 2003.
FREIMANN S, SCHEINOWITZ M, YEKUTIELI D, FEINBERG M.S, ELDAR M,
KESSLER-ICEKSON G. Prior exercise training improves the outcome of acute
myocardial infarction in the rat. Heart structure, function, and gene expression. J
Am Coll Cardiol, 45(6): 931-8, 2005.
FREY N, KATUS HA, OLSON E N, HILL J A. Hypertrophy of the Heart A New
Therapeutic Target? Circulation; 109: 1580-1589, 2004.
FULLER E.O e NUTTER D.O. Endurance training in the rat II. Performance of
isolated and intact heart. J. App. Physiol, 51(4): 941-947, 1981.

81
GARBELOTTI Jr SA e PELOZO Jr O, “Sistema circulatório”. In: Reabilitação
cardiovascular e metabólica. 1ª ed. São Paulo. Atheneu, 2013.
GARCIA JAD e INCERPI EK Factors and Mechanisms Involved in Left Ventricular
Hypertrophy and the Anti-Hypertrophic Role of Nitric Oxide. Arq Bras Cardiol;
90(6): 443-450, 2008.
GEENEN D, BUTTRICK P e SCHEUER J. Cardiovascular and hormonal
responses to swimming and running in the rat. Journal of Applied Physiology, 65:
116-123, 1988.
GIBALA MJ e MCGEE SL. Metabolic adaptations to short-term high intensity
interval training: a little pain for a lot of gain? Exerc. Sport Sci. Rev, 36:58Y63,
2008.
GIBALA MJ, McGEE SL, GAMHAM AP, HOWLETT KF, SNOW RJ &
HARGREAVES M. Brief intense interval exercise activates AMPK and p38 MAPK
signalling and increases the expression of PGC-1α in human skeletal muscle. J
Appl Physiol 106, 929–934, 2009.
GIBALA MJ, LITTLE JP, MAUREEN J MACDONALD MJ and HAWLEY JA.
Physiological adaptations to low-volume, high-intensity interval training in health
and disease J Physiol; 590(Pt 5): 1077–1084, 2012.
GHORAYEB N e BATLOUNI M. Hipertrofia ventricular: mecanismos envolvidos na
indução e possibilidades de regressão. Rev Soc Cardiol Est São Paulo; 8: 298-
301, 1998.
GHORAYEB N, BATLOUNI M, PINTO IMF, GIUSEPPE SD. Hipertrofia ventricular
esquerda do atleta. Resposta adaptativa fisiológica do coração. Arq. Bras. Cardiol.
vol.85 no.3, 2005.
GOBATTO CA, de MELLO MAR, SIBUYA CY, AZEVEDO JRM, dos SANTOS LA,
KOKUBUN E. Maximal lactate steady state in rats submitted to swimming exercise.
Comparative Biochemistry and Physiology Part A 130; 21-27, 2001.
GOTO M, TERADA S, KATO M. cDNA cloning and mRNA analysis of PGC-1 in
epitrochlearis muscle in swimming-exercised rats. Biochem Biophys Res Commun
274(2):350–354, 2000.
GREGG E.W, CAULEY J.A, STONE K, THOMPSON T.J, BAUER D.C,
CUMMINGS S.R, ENSRUD K.E. Relationship of changes in physical activity and
mortality among older women. JAMA, 289: 2379-86, 2003.
GULATI M, PANDEY D.K, ARNSDORF M.F, LAUDERDALE D.S, THISTED R.A,
WICKLUND R.H, AL-HANI A.J.,BLACK H.R. Exercise capacity and the risk of

82
death in women. The St James women take heart project. Circulation, 108:1554–
9, 2003.
GUTE D, FRAGA C, LAUGHLIN MH, AMANN JF. Regional changes in capillary
supply in skeletal muscle of high-intensity endurancetrained rats. J Appl Physiol
81: 619–626, 1996.
HAMBRECHT R. M.J, FIEHN E, WEIGL C, GIELEN S, HAMANN C, KAISER R,
YU J, ADAMS V, NIEBAUER J, SCHULER G. Regular physical exercise corrects
endothelial dysfunction and improves exercise capacity in patients with chronic
heart failure. Circulation, 98: 2709-15, 1998.
HAN GS. Endurance Exercise Effects on Cardiac Hypertrophy in Mice. J Phys Ther
Sci.25(12): 1525–1527, 2013.
HARAM PM, KEMI OJ, LEE SJ, BENDHEIM MO, AL-SHARE QY, WALDUM HL
GILLIGAN LJ, KOCH LG, BRITTON SL, NAJJAR SM, WISLØFF U. Aerobic
interval training vs. continuous moderate exercise in the metabolic syndrome of
rats artificially selected for low aerobic capacity. J Cardiovasc. Dis. Res, 81: 723-
32, 2009.
HASHIMOTO NY, FERNANDES T, SOCI UPR, OLIVEIRA EMO. Molecular
Determinants of Cardiac Hypertrophy Induced by Different Amounts of Aerobic
Exercise Training. Rev Bras Cardiol. 24(3):153-162, 2011.
HASKELL WL, LEE IM, PATE RR, POWELL KE, BLAIR SN, FRANKLIN BA,
MACERA CA, HEATH GW, THOMPSON PD, BAUMAN A. Physical Activity and
Public Health: Updated Recommendation for Adults from the American College of
Sports Medicine and the American Heart Association. MEDICINE & SCIENCE IN
SPORTS & EXERCISE. Med Sci Sports Exerc. 39(8):1423-34, 2007.
HELBER I, ANTONIO E.L, SANTOS A.A, FLUMIGNAN L.G, BOCALINI D.S,
GHEORGHIADE M, TUCCI P.J. Digitoxin prolongs survival of female rats with
heart failure due to large myocardial infarction. J Cardiac Failure, 15(9): 798-804,
2009.
ILAR. Institute of Laboratory Animal Resources. Manual sobre Cuidados e Uso
de Animais de Laboratório. Consulta em 01/11/15:
http://www.usp.br/bioterio/Artigos/Taxonomia_e_biologia.pdf, 2003.
KAVANAGH T, MERTENS D.J, HAMM L.F, BEYENE J, KENNEDY J, COREY P,
SHEPHARD R.J. Prediction of long-term prognosis in men referred for cardiac
rehabilitation. Circulation, 106: 666–671, 2002.

83
KEHAT I e MOLKENTIN JD. Pathways Underlying Cardiac Remodeling During
Pathophysiological Stimulation Circulation. 122:2727-2735, 2010.
KEMI O.J, LOENNECHEN J.P, WISLØFF U, ELLINGSEN Ø. Intensity-controlled
treadmill running in mice: cardiac and skeletal muscle hypertrophy. J Appl Physiol,
93:1301- 9, 2002.
KEMI OJ, HARAM PM, WISLOFF U, ELLINGSEN O. Aerobic fitness is associated
with cardiomyocyte contractile capacity and endothelial function in exercise
training and detraining. Circulation, 109:2897Y904, 2004.
KEMI O.J, HARAM P.M, LOENNECHEN J.P, OSNES J.B, SKOMEDAL T,
WISLOFF U, ELLINGSEN O. Moderate vs. high exercise intensity: differential
effects on aerobic fitness, cardiomyocyte contractility, and endothelial function.
Cardiovasc Res, 67: 161Y72, 2005.
KEMI O.J, ELLINGSEN O, CECI M, GRIMALDI S, SMITH G.L, CONDORELLI G,
WISLOFF U. Aerobic interval training enhances cardiomyocyte contractility and
Ca2+ cycling by phosphorylation of CaMKII and Thr-17 of phospholamban. J. Mol.
Cell. Cardiol, 43: 354Y61, 2007.
KEMI O.J, CECI M, CONDORELLI G, SMITH G.L, WISLOFF U. Myocardial
sarcoplasmic reticulum Ca2+ ATPase function is increased by aerobic interval
training. Eur. J. Cardiovasc. Prev Rehabil, 15: 145Y8, 2008a.
KEMI OJ, CECI M, WISLOFF U, GRIMALDI S, GALLO P, SMITH G.L,
CONDORELLI G, ELLINGSEN O. Activation or inactivation of cardiac Akt/mTOR
signaling diverges physiological from pathological hypertrophy. J. Cell. Physiol,
214: 316Y21, 2008b.
KEMI OJ, ELLINGSEN O, SMITH GL, WISLOFF U. Exercise-induced changes in
calcium handling in left ventricular cardiomyocytes. Front Biosci. 1;13:356-68,
2008c.
KEMPF T & WOLLERT KC. Nitric oxide and the enigma of cardiac hypertrophy.
Bioessays; 6: 608-15, 2004.
KREGEL KC (2006). Resource Book for the design of animal exercise protocols;
Available via http://www.the-
aps.org/mm/SciencePolicy/AnimalResearch/Publications/
Animal-Exercise-Protocols/book14824.pdf Accessed 07 October, 2014.
LAUGHLIN MH e TOMANEK RJ. Myocardial capillarity and maximal capillary
diffusion capacity in exercise-trained dogs. J Appl Physiol 63: 1481–1486, 1987.

84
LAUGHLIN MH, KORTHUIS RJ, DUNCKER DJ, BACHE RJ. Regulation of blood
flow to cardiac and skeletal muscle during exercise.In: Handbook of Physiology.
Exercise: Regulation and Integration of Multiple Systems. Bethesda, MD: Am.
Physiol.Soc, p. 705–769, 1996.
LAVIE C.J, O’KEEFE J.H, SALLIS RE. Exercise and the Heart V the Harm of Too
Little and Too Much. CHEST AND ABDOMINAL CONDITIONS. Volume 14,
Number 2, 2015.
LEE I.M, SESSO H.D, OGUMA Y, PAFFENBARGER R.S.JR. Relative intensity of
physical activity and risk of coronary heart disease. Circulation, 107:1110– 6, 2003.
LEVY MN, KOEPPEN BM, STANTON BA. Fundamentos de Fisiologia. 4ª ed. São
Paulo: Elsevier, 2006.
LEW WY. Exercise: commitment to a young heart. J. Am. Coll. Cardiol. 64: 1267–
9, 2014.
LEWARTOWSKI, B.; PYKOWSKY, B. Cellular mechanism of the relationship
between myocardial force and frequency of contractions. Prog. Biophys. Mol. Biol.,
50: 97-120, 1987.
MAILLET M, van BERLO JH, MOLKENTIN JD. Molecular basis of physiological
heart growth: fundamental concepts and new players. Nature Reviews Molecular
Cell Biology 14, 38-48, 2013.
MANSON JE, GREENLAND P, LaCROIX AZ, STEFANICK ML, MOUTON CP,
OBERMAN A, PERRI MG, SHEPS DS, PETTINGER MB, SISCOVICK DS.
Walking compared with vigorous exercise for the prevention of cardiovascular
events in women. N Engl J Med. 5;347(10):716-25, 2002.
MANTELLI, L.; MANZINI, S.; LEDDA, F.; Differences between the effects of
phenylephrine and other inotropic interventions on post-rest contraction in guinea-
pig ventricular strips. Arch. Intern. Pharmaco., 272: 225-235, 1984.
MARKS A. Calcium and the heart: a question of life and death. J. Clin. Invest.
111:597–600, 2003.
MEDEIROS A, GIANOLLA RM, KALIL LMP, BACURAU RFP, ROSA LFBC,
NEGRÃO CE, BRUM PC. Efeito do treinamento físico com natação sobre o
sistema cardiovascular de ratos normotensos. Rev. Paul. Educ. Fís., São Paulo,
14(1):7-15, 2000.
MEDEIROS A, OLIVEIRA EM, GIANOLLA R, CASARINI DE, NEGRÃO CE and
BRUM PC. Swimming training increases cardiac vagal activity and induces cardiac
hypertrophy in rats. Braz J Med Biol Res, Volume 37(12) 1909-1917, 2004.

85
McARDLE WD. Metabolic stress of endurance swimming in the laboratory rat.
Journal of Applied PhysiologyPublished 1 January Vol. 22no. 1, 50-54, 1967.
McARDLE WD, KATCH FI, KATCH, VL – Fisiologia do exercício: energia, nutrição
e desempenho humano – 6º edição, Guanabara Koogan, Rio de Janeiro, 2008.
McCULLOUGH PA e Lavie CJ. Coronary artery plaque and cardiotoxicity as a
result of extreme endurance exercise. Mo. Med. 111:9Y94, 2014
McMULLEN JR, SHIOI T, ZHANG L, et al. – Phosphoinositide 3-kinase (p110)
plays a critical role for the induction of physiological, but not pathological, cardiac
hypertrophy. PNAS; 21:12355-12360, 2003.
McMULLEN J e JENNINGS GL. Differences between pathological and
physiological cardiac hypertrophy: novel therapeutic strategies to treat heart
failure. Clin Exp Pharmacol Physiol;34(4):255-62, 2007.
MILL, J. G.; VASSALO, D. V.; LEITE, C. M.; Mechanisms undelying the genesis of
post rest contractions in cardiac muscle. Brazilian J. Med. Biol. Res., 25: 399-408,
1992
MOHOLDT T, WISLOFF U, NILSEN TI, SLORDAHL SA. Physical activity and
mortality in men and women with coronary heart disease: a prospective population-
based cohort study in Norway (the HUNT study). Eur J Cardiovasc Prev
Rehabil.15:639–645, 2008.
MONT L, ELOSUA R and BRUGADA J. Endurance sport practice as a risk factor
for atrial fibrillation and atrial flutter. Europace. 11(1): 11–17, 2009.
MONTGOMERY HE. Should the contribution of ACE gene polymorphism to left
ventricular hypertrophy be reconsidered? Heart, 77:89-90, 1997.
MORGANROTH J, MARON BJ, HENRY WL, EPSTEIN SE. Comparative left
ventricular dimensions in trained athletes. Ann Intern Med, 82: 521-4, 1975.
MUJIKA I, CHATARD JC, BUSSO T, GEYSSANT A, BARALE F, LACOSTE L.
Effects of training on performance in competitive swimming. Can J Appl Physiol.
20(4):395-406, 1995.
MYERS J, PRAKASH M, FROELICHER V, DO D, PARTINGTON S, ATWOOD E.
Exercise capacity and mortality among men referred for exercise testing. N Engl J
Med, 346:793– 801, 2002.
NATALI AJ. Effects of chronic exercise on cardiac myocytes: a review about
mechanical adaptations. R. bras. Ci. e Mov. v. 12 n. 1, p. 91-96, 2004.
NORRBOM J, SUNDBERG C, AMELN H, KRAUS W, JANSSON E,
GUSTAFSSON T. PGC-1{alpha}mRNA expression is influenced by metabolic

86
perturbation in exercising human skeletalmuscle. J Appl Physiol. 96(1):189–194,
2004.
NUNES RB, ALVES JP, KESSLER LP, DAL LAGO P. Aerobic exercise improves
the inflammatory profile correlated with cardiac remodeling and function in chronic
heart failure rats. Clinics (Sao Paulo. 68(6):876-82, 2013.
NUTTER DO, PRIEST RE, FULLER EO. Endurance training in the rat. I.
Myocardial T40, 1981.
O'KEEFE JH, PATIL HR, LAVIE CJ, MAGALSKI A, VOGEL RA, McCULLOUGHD
PA. Potential Adverse Cardiovascular Effects From Excessive Endurance
Exercise. Mayo Clin Procv.87(6), 2012
OPIE LH. Myocardial contraction and relaxation. In: Opie LH (ed). The heart
physiology from cell to circulation. 3rd ed. Philadelphia: Lippincott-Raven.p.209-
31, 1998
OPIE LH e LOPASCHUK GD. Fuels: aerobic and anaerobic metabolism. In: Heart
Physiology, From Cell to Circulation, edited by Weinberg RW, Bersin J, Aversa F.
Philadelphia, PA: Lippincot Williams & Wilkins, p. 306–354, 2004.
PAFFENBARGER JR R.S, HYDE R.T, WING A.L, LEE I.M, JUNG D.L, KAMPERT
J.B. The association of changes in physical-activity level and other lifestyle
characteristics with mortality among men. N Engl J Med, 328:538–45, 1993.
PERON A, SARAIVA, RM, ANTONIO EL, ANDRIOLO A, TUCCI PJF. Food
restriction does not impair myocardial mechanics during the healing period of
myocardial infarction in the rat. Nutrition Research. Volume 25, Issue 12, Pages
1075–1084, 2005.
PIESKE B, SÜTTERLIN M, SCHMIDT-SCHWEDA S, MINAMI K, MEYER M,
OLSCHEWSKI M, HOLUBARSCH C, JUST H, AND HASENFUSS G. Diminished
post-rest potentiation of contractile force in human dilated cardiomyopathy.
Functional evidence for alterations in intracellular Ca2+ handling. J Clin Invest.
98(3): 764–776, 1996.
PILIS W, ZARZECZNY R, LANGFORT J, KACIUBA-USCILKO H, NAZAR K,
WOJTYNA J. Anaerobic threshold in ratsComparative Biochemistry and
Physiology Part A: Physiology Volume 106, Pages 285–289, 1993.
PINNELL J, TURNER S, HOWELL S, Cardiac muscle physiology. Critical Care &
Pain. Volume 7, Number 3, 2007.

87
PONTES MRN e LEÃES PE. Remodelamento Ventricular: dos Mecanismos
Moleculares e Celulares ao Tratamento. Revista da Sociedade de Cardiologia do
Rio Grande do Sul - Ano XIII nº 03, 2004.
POWERS SK., DEMIREL HA, VINCENT KH. Exercise training improves
myocardial tolerance to in vivo ischemia-reperfusion in the rat. Am J Physiol, 275:
R1468-1477, 1998.
PRIOR BM, YANG HT, TERJUNG RL. What makes vessels grow with exercise
training? J Appl Physiol. 97:1119-28, 2004.
RAMOS-FILHO D, CHICAYBAM G, DE-SOUZA-FERREIRA E, GUERRA
MARTINEZ C, KURTENBACH E, CASIMIRO-LOPES G, GALINA A. High Intensity
Interval Training (HIIT) Induces Specific Changes in Respiration and Electron
Leakage in the Mitochondria of Different Rat Skeletal Muscles. PLoS One.
10(6):e0131766, 2015.
RAVENS, U. Post-rest potentiation and its decay in NOBLE, M. I. M.; SEED, W. A.
The interval-force relationship of the heart. New York: Cambridge University Press,
p. 245- 258, 1992.
RAVENS, U.; LINK, S.; GATH, J.; NOBLE, M. I. M. Post-rest potentiation and its
decay after inotropic interventions in isolated rat heart muscle. Pharmacol.
Toxicol., 76: 9-16, 1995.
RIEBE D, FERNHALL B, THOMPSON PD. The blood pressure response to
exercise in anabolic steroid users. Med Sci Sports Exerc; 24: 633-637, 1992.
ROGNMO O, HETLAND E, HELGERUD J, HOFF J, SLØRDAHL S.A. High
intensity aerobic interval exercise is superior to moderate intensity exercise for
increasing aerobic capacity in patients with coronary artery disease. Eur J
Cardiovasc Prev Rehabil, 11: 216-22, 2004.
ROGNMO O, MOHOLDT T, BAKKEN H, HOLE T, MOLSTAD P, MYHR NE,
GRIMSMO J, WISLOFF U. Cardiovascular risk of high- versus moderate-intensity
aerobic exercise in coronary heart disease patients. Circulation; 126 (12): 1436-
1440, 2012.
ROHAS LM, St-PIERRE J, ULDRY M, JAGER S, HANDSCHIN C & SPIEGELMAN
BM. A fundamental system of cellular energy homeostasis regulated by PGC-1α.
Proc Natl Acad Sci USA 104, 7933–7938, 2007.
ROST R. “O coração de atleta. Perspectivas históricas – Problemas resolvidos e
não resolvidos”. In: Clínicas Cardiológicas, Interlivros. v.3, 1997.

88
SARQUELLA-BRUGADA G, CAMPUZANO O, IGLESIAS A, SÁNCHEZ-
MALAGÓN J, GUERRA-BALIC M, BRUGADA J, BRUGADA R. Genetics of
sudden cardiac death in children and young athletes. Cardiol. Young 24, 1–15,
2012.
SBC (Sociedade Brasileira de Cardiologia). Diretriz de Reabilitação Cardíaca. Arq.
Bras. Cardiol. vol.84 no.5, 2005.
SCHAIBLE T e SCHEUER J. Effects of physical training by running or swimming
on ventricular performance of rat hearts. J Appl Physiol, 46(4): 854-860, 1979.
SCHAIBLE T e SCHEUER J. Cardiac function in hypertrophied hearts from
chronically exercised female rats. J. Appl Physiol, 50:1140-5, 1981.
SCHOUTEN, V. J. A., VAN DEEN, J. K.; DE TOMBE, P. P.; VERVEEN; A. A.
Force-interval relationship in heart muscle of mammals. A calcium comportament
model. Biophys. J., 51: 13-26, 1987
SCHOUTEN, V. J. A. Interval dependence of force and twitch duration in rat heart
explained by Ca2+ pump inactivation in sarcoplasmic reticulum. J. Physiol., 431:
427-444, 1990.
SCHWARTZ RS, KRAUS SM, SCHWARTZ JG, et al. Increased coronary artery
plaque volume among male marathon runners. Mo. Med. 111:85Y90, 2014
SEJERSTED O. Calcium controls cardiac function – by all means! J Physiol 589.12
pp 2919–2920, 2011.
SERRA A.J, ANTONIO E.L, BOCALINI D.S, SANTOS M.H.H, HIGUCHI M.L,
LEVY R.F, TUCCI PJF. Exercise training inhibits inflammatory cytokines and more
than just prevents myocardial dysfunction in rats with sustained {beta}-adrenergic
hyperactivity. J Physiol, 588(13): 2431-2442, 2010.
SHARMA S e ZAIDI A. Exercise-induced arrhythmogenic right ventricular
cardiomyopathy: fact or fallacy? European Heart Journal. 33, 938–940, 2012.
SHEPARD RJ e BALADY GY. Exercise as cardiovascular therapy. Circulation, 99:
963-72, 1999.
SONG W, VIKHOREV PG, KASHYAP MN, ROWLANDS C, FERENCZI MA,
WOLEDGE RC, MACLEOD K, MARSTON S, CURTIN NA. Mechanical and
energetic properties of papillary muscle from ACTC E99K transgenic mouse
models of hypertrophic cardiomyopathy. Am J Physiol Heart Circ
Physiol;304(11):H1513-24, 2013.

89
STROM CC, APLIN M, PLOUG T et al. Expression profiling reveals differences in
metabolic gene expression between exercise-induced cardiac efects and
maladaptive cardiac hypertrophy. FEBS J.; 272(11):2684– 2695, 2005.
SWAIN DP e FRANKLIN BA. Comparison of cardioprotective benefits of vigorous
versus moderate intensity aerobic exercise. Am J Cardiol, 97:141–147, 2006.
SWYNGHEDAUW B. Molecular mechanisms of myocardial remodeling. Physiol
Ver. 79(1):215-62, 1999.
SWYNGHEDAUW B. Developmental and functional adaptation of contractile
proteins in cardiac and skeletal muscle. Physiol Rev 66: 710-71, 1986
TANASESCU M, LEITZMANN M.F, RIMM E.B, WILLETT W.C, STAMPFER M.J,
HU FB. Exercise type and intensity in relation to coronary heart disease in men.
JAMA, 288:1994-2000, 2002.
TERADA S, GOTO M, KATO M, KAWANAKA K, SHIMOKAWA T, TABATA I.
Effects of low intensity prolonged exercise on PGC-1 mRNA expression in rat
epitrochlearis muscle. Biochem Biophys Res Commun. 296(2):350–354, 2002.
THOMPSON PD, FRANKLIN BA, BALADY GJ, BLAIR SN, CORRADO D, ESTES
NA 3rd, FULTON JE, GORDON NF, HASKELL WL, LINK MS, MARON BJ,
MITTLEMAN MA, PELLICCIA A, WENGER NK, WILLICH SN, COSTA F;
American Heart Association Council on Nutrition, Physical Activity, and
Metabolism; American Heart Association Council on Clinical Cardiology; American
College of Sports Medicine.Exercise and acute cardiovascular events placing the
risks into perspective: a scientific statement from the American Heart Association
Council on Nutrition, Physical Activity, and Metabolism and the Council on Clinical
Cardiology. Circulation. 1;115(17):2358-68, 2007.
TJONNA AE, LEE SJ, ROGNMO Ø, StOLEN TO, BYE A, HARAM PM,
LOENNECHEN JP, AL-SHARE QY, SKOGVOLL E, SLORDAHL SA, KEMI OJ,
NAJJAR SM, WISLOFFf U. Aerobic interval training versus continuous moderate
exercise as a treatment for the metabolic syndrome: a pilot study.
Circulation;118(4):346-54, 2008.
TRAPP EG, CHISHOLM DJ, FREUND J, BOUTCHER SH. The effects of high-
intensity intermittent exercise training on fat loss and fasting insulin levels of young
women. Int J Obes (Lond).32(4):684-91, 2008.
TSUTSUI JM, FALCÃO SNRS, LIMA MM, MATHIAS Jr W. “Ecocardiografia e
exercício físico”. In: Cardiologia do Exercício – Do atleta ao cardiopata, Manole.
v.3, 2010.

90
VASSALO, DV.e MILL, JG. Mechanical behaviour of rest contractions in cardiac
muscle. Acta Physiol. Pharmacolol. Latinoamericana, 38: 87-97, 1988.
VEIGA EC, ANTONIO EL, BOCALINI DS, MURAD N, ABREU LC, TUCCI PJ,
SATO MA. Prior exercise training does not prevent acute cardiac alterations after
myocardial infarction in female rats. Clinics (Sao Paulo); 66(5): 889–893, 2011.
VEIGA ECA, PORTES LA, BOCALINI DS, ANTONIO EL, dos SANTOS AA,
SANTOS MH, SILVA FA, TUCCI PJF. Cardiac Implications after Myocardial
Infarction in Rats previously Undergoing Physical Exercise. Arq Bras Cardiol;
100(1):37-43, 2013.
VLADIMIR I. MOROZOV VI, TSYPLENKOV PV, GOLBERG ND, KALINSKI MI.
The effects of high-intensity exercise on skeletal muscle neutrophil
myeloperoxidase in untrained and trained rats. Eur J App Physiol, 97(6): 716-722,
2006.
VOLTARELLI FA, GOBATTO CA and de MELLO MAR. Determination of
anaerobic threshold in rats using the lactate minimum test. Braz J Med Biol Res;
Volume 35 (11) 1389-1394, 2002.
WAGNER C, SIMON A, EVANS J. Physical activity and coronary event incidence
in Northern Ireland and France: the prospective epidemiological study of myoardial
infarction (PRIME). Circulation, 105: 2247-52, 2002.
WANG S, MA JZ, ZHU SS, XU DJ, ZOU JG CAO KJ. Swimming training can affect
intrinsic calcium current characteristics in rat myocardium. European Journal of
Applied Physiology, Volume 104, Issue 3, pp 549-555, 2008.
WANG Y, WISLOFF U, KEMI O.J. Animal models in the study of exercise-induced
cardiac hypertrophy. Physiol. Res, 59: 633-644, 2010.
WARBURTON D.E, MCKENZIE D.C, HAYKOWSKY M.J. Effectiveness of high-
intensity interval training for the rehabilitation of patients with coronary artery
disease. Am. J. Cardiol, 95:1080Y4, 2005.
WASHBURN RA, SZABO AN, LAMBOURNE K, WILLIS EA, PTOMEY LT, HONAS
JJ, HERRMANN SD, DONNELLY JE. Does the Method of Weight Loss Effect
Long-Term Changes in Weight, Body Composition or Chronic Disease Risk
Factors in Overweight or Obese Adults? A Systematic Review. PLoS One. 9(10):
e109849, 2014.
WEBER K.T. Fibrosis and hypertensive heart disease. Curr Opin Cardiol, 15(4):
264-72, 2000.

91
WEINER RB e BAGGISH AL. Exercise-induced cardiac remodeling. Prog
Cardiovasc Dis. 54(5):380-6, 2012.
WENDE AR, SCHAEFFER PJ, PARKER GJ, ZECHNER C, HAN DH, CHEN MM,
HANCOCK CR, LEHMAN JJ, HUSS JM, McCLAIN DA, HOLLOSZY JO & KELLY
DP (2007). A role for the transcriptional coactivator PGC-1α in muscle refueling. J
Biol Chem 282, 36642–36651, 2007.
WENDT-GALLITELLI, M. F.; ISENBERG, G. Potentiation of contraction as related
to changes in force and total intracelluar calcium. Adv. Exp. Med. Biol., 311: 213-
226, 1992.
WENGER H.A e BELL G.J. The interactions of intensity, frequency and duration of
exercise training in altering cardiorespiratory fitness. Sports Med, 3: 346Y56, 1986.
WILLIAMS AG, RAYSON MP, JUBB M, WORLD M, WOODS DR, HAYWARD M,
MARTIN J, HUMPHRIES S E, MONTGOMERY HE. The ACE gene and muscle
performance. Nature 403, 614, 2000.
WISLOFF U, HELGERUD J, KEMI OJ, ELLINGSEN Ø. Intensity-controlled
treadmill running in rats: Vo2max and cardiac hypertrophy. Am J Physiol Heart Circ
Physiol, 280: H1301-10, 2001a.
WISLOFF U, LOENNECHEN JP, FALCK G, BEISVA°G V, CURRIE S, SMITH GL.
Increased contractility and calcium sensitivity in cardiac myocytes isolated from
endurance trained rats. Cardiovasc Res, 50: 495–508, 2001b.
WISLOFF U, LOENNECHEN J.P, CURRIE S, SMITH G.L, ELLINGSEN O. Aerobic
exercise reduces cardiomyocyte hypertrophy and increases contractility, Ca2+
sensitivity and SERCA-2 in rat after myocardial infarction. Cardiovasc Res, 54:
162-74, 2002.
WISLOFF U, NILSEN TI, DROYVOLD WB, MORKVED S, SLORDAHL SA,
VATTEN LJ. A single weekly bout of exercise may reduce cardiovascular mortality:
How little pain for cardiac gain? The HUNT study, Norway. Eur J Cardiovasc Prev
Rehabil. 13:798–804, 2006.
WISLOFF U, STOYLEN A, LOENNECHEN JP. Superior cardiovascular effect of
aerobic interval training versus moderate continuous training in heart failure
patients: a randomized study. Circulation, 115: 3086Y94, 2007.
WISLOFF U, ELLINGSEN K, KEMI OJ. High-Intensity Interval Training to
Maximize Cardiac Benefits of Exercise Training? Exercise and Sport Sciences
Reviews. V37, Number 3, 2009

92
ZHANG LQ, ZHANG XQ, MUSCH TI, MOORE RL, CHEUNG JY. Sprint training
restores normal contractility in postinfarction rat myocytes. J Appl Physiol, 89:
1099-1105, 2000a.
ZHANG LQ, ZHANG XQ, ROTHBLUM L, MUSCH TI, MOORE RL, CHEUNG JY.
Sprint training normalizes Ca2+ transients and RS function in postinfarction rat
myocytes. J Appl Physiol, 89: 38-46, 2000b.
ZORNOFF LAM, CICOGNA AC, PAIVA SAR, SPADARO J. Remodelamento e seu
impacto na progressão da disfunção ventricular. Rev Soc Cardiol Estado de São
Paulo; 12(3):371-8, 2002.

93
9. COMITÊ DE ÉTICA





























