Embed Size (px)
Citation preview

Transthyretin expression in medulloblastomas and medulloblastoma cell lines S. Albrecht", T.A. Bayer, J.A. Kraus and T. Pietsch Dqmrtriierit of' Neriropnt/iolo,qy, Uiiivrrsity of Bonn Mrdicd Centre. Bonn. Germnny
S. Albrecht. T.A. Bayer. J.A. Kraus and T. Pietsch (1995) Neuropathologg mid Applied NeuroOiologtf 21, 399-409 Transthyretin expression in medulloblastomas and medulloblastoma cell lines
Transthyretin is a protein crucial to the transport of lipophilic molecules such as thyroid hormones and retin- oids. In the central nervous system. large amounts of transthyretin are synthesized by the choroid plexus and are secreted into the cerebrospinal fluid. The choroid plexus is the only site of transthyretin synthesis in the brain. Transthyretin is expressed by most benign and malignant choroid plexus tumours while gliomas and meningiomas do not express transthyretin. Other major sites of transthyretin synthesis are the retinal pigment epithelium and hepatocytes. Medulloblastoma is the prototypical primitive neuroectodermal tumour of the cerebellum and can show multiple lines of differentiation, including the expression of retinal markers. In this study, we examined transthyretin expression both at the RNA and protein level in four medulloblastomas and six medulloblastoma cell lines using Northern and Western blot analysis. reverse transcription polymerase chain reaction (PCR). RNA in sitri hybridization, and immuno- histochemistry. All four medulloblastomas and five of the six medulloblastoma cell lines expressed transthyretin-
Keywords: transthyretin, medulloblastoma, PNET
mRNA as demonstrated by reverse PCR and iu sitii hybridization while three medulloblastomas and one cell line were positive on Northern blot. The medullo- blastoma with the most abundant RNA expression was transthyretin-immunoreactive on cryosections and the medulloblastoma cell line that was positive on Northern blot also expressed transthyretin at levels detectable by Western blot. No transthyretin-immunoreactivity was seen in 16 additional medulloblastomas studied on paraffin sections. These findings indicate that low- level expression of transthyretin-mRNA is common in medulloblastomas and medulloblastoma cell lines. Expression of transthyretin protein occurs rarely but can reach significant levels. Transthyretin expression in medulloblastoma is consistent with retinal pigment epithelium differentiation in medulloblastomas and reflects their pluripotential nature. Furthermore, the potential for transthyretin-immunoreactivity in medullo- blastoma should be kept in mind when performing immunohistochemical studies on poorly differentiated cerebellar tumours.
Introduction
Transthyretin is a serum protein involved in the trans- port of highly lipophilic molecules such as retinoids and thyroid hormones. Serum transthyretin is synthesized and secreted by hepatocytes. However, the choroid
'Present address: Department of Pathology, Sir Mortimer B. Davis- Jewish General Hospital, and Lady Davis Institute for Medical Research, McGill University. Montreal. Canada. Correspondence: Dr T. Pietsch, Department of Neuropathology. University of Bonn Medical Centre. Sigmund-Freud-Strasse 25. D-53105 Bonn. Germany.
Q 199 5 Blackwell Science Ltd
plexus also produces large amounts of transthyretin which are secreted into the cerebrospinal fluid (CSF). Among primary cerebral turnours, choroid plexus papil- lomas and choroid plexus carcinomas often express transthyretin [l. 4, 19, 371. In the eye, transthyretin is synthesized only in the retinal pigment epithelium [ 10, 161. Medulloblastoma is the prototypical primitive neuroectodermal tumour of the cerebellum. Medullo- blastomas can display multiple lines of differentiation, including neuronal, astrocytic, ependymal, and myo-

genic [S. 401. Expression of relinal markers such as rctirial S-antigen and rod-opsin also occur frequently in medulloblastomas [ 6 , 1 3 . 321. In this study, we exam- ined the expression of transthyretin in medulloblastomas and medulloblastoma cell lines. Our results indicate that some medulloblastomas and medulloblastoma cell lines can in fact express transthyretin both at the mRNA and at the protein level. This provides additional evidence for rctinal pigment epithelium differentiation in medullo- blastomas and underscores their pluripotential nature. In addition, the possibility of transthyretin expression in medulloblastoma should be kept in mind when performing imiiiunohistochemical studies on poorly differentiated cerebellar tumours.
Materials and methods
Tumours and cell lines
Tissue was frozen from four medulloblastomas which had been obtained fresh from the surgical resection specimens, snap frozen in liquid nitrogen. and stored at - 80°C until further processing (cases 2462, 2466, 2472 and 305 2). In addition, formalin-fixed. paraffin- embedded tissue from 16 additional medulloblastomas was used. All medulloblastomas had been diagnosed independently by two neuropathologists according to the WHO brain tumour classification. None of these tumours contained areas of papillary architecture or had discern- ible epithelial or glandular features by light microscopy. There was also fresh frozen tissue from a choroid plexus papilloma (case 2410). In addition, we studied six rnedulloblastoma cell lines. These consisted of two previ- ously established lines, D283 Med [17] and DAOY [21], as well as lines MHH-MED-1 to -4 which are four newly described permanent medulloblastoma cell lines [39]. For some experiments, the Hep-T cell line was also included, which is a permanent cell line derived from a hepato- blastoma (T. Pietsch. unpublished data), the MHH- NE-11 cell line derived from a neuroblastoma [38], and the U-87 MG malignant glioma cell line (obtained from ATCC).
Transthyretin probe
The transthyretin-probe was a generous gift of J. Herbert, &ID (Department of Neurology, Columbia University, College of Physicians and Surgeons, New York. USA). It
consists of a 500 bp cDNA fragment that had been originally isolated from an adult human liver cDNA library [42] and later subcloned into the EroRI site of pSP65 [20]. For Northern and Southern blot analysis. probes were generated from the purified insert whereas the probe for non-radioactive RNA in situ hybridization was generated from the linearized plasmid.
RNA extraction
RNA was extracted from tumours 2462. 246h. 2 4 i 2 and 3052 as well as from the six medulloblastoma cell lines. For comparison. RNA was also extracted from the choroid plexus papilloma (case 2410), the Hep-T cell line, and a sample of normal frontal lobe obtained from a surgical specimen. Frozen sections were cut from the medulloblastoma fragments selected for RNA extraction to ensure that they contained only tumour and no contaminating choroid plexus. Solid tissue fragments were pulverized in liquid nitrogen with a mortar and pestle. Total cellular RNA was extracted from the tissue powder using the single step guanidinium thiocyanate/ acid phenol method [Ill. For RNA extraction from cell cultures, cells from one or two 2 5 cm2 culture flasks were spun down, rinsed once with PES and pelleted by centrifugation. The pellets were then treated like the tissue powder obtained from the tissue fragments.
Northern blot analysis
Approximately 1 5 pg of total cellular RNA were loaded onto a 1% agarose-formaldehyde gel, electrophoresed overnight at 1 V/cm. Examination under ultraviolet light confirmed that similar amounts of RNA had been loaded. The RNA was then blotted overnight onto a positively charged nylon membrane (Hybond-N-Plus. Amersham) using 20 x SSC as transfer buffer, and cross-linked to the membrane by irradiation with ultraviolet light (Stratalinker 1800, Stratagene). Approximately 50 ng of the transthyretin-probe were labelled with 50 pCi of (a3’P)-dCTP using a random primer labelling kit (Gibco- BRL) according to the manufacturer’s instructions. The membrane was pre-hybridized overnight at 4 5°C in hybridization buffer (Hybrisol I, Oncor). Hybridization was carried out at 45°C for 16 h. Following hybridiz- ation, the membrane was washed several times under increasingly stringent conditions: the final wash was
0 1995 Blackwell Science Ltd, Neuropntliology and Applied Neurobiology. 21. 399109

performed with 0.25 x SSC with 0.1'3: SDS at 65°C. The membrane was then exposed to X-ray film (Cronex. Dupont) at - 70°C with an intensifying screen for various lengths of time (5 min to 3 days).
Reverse PCR and Southern blot
Approximately 1-2 pg of total cellular RNA were reverse transcribed in a total volume of 20 p1 using a commer- cial cDNA synthesis kit (Superscript preamplification kit, Gibco-BRL) according to the manufacturer's instruc- tions. One microlitre of the cDNA was used for each PCR reaction. PCR was performed in a final volume of 10 p1 in a buffer consisting of 10 r n ~ Tris-HCl (pH 8.3 a t RT), 50 mi KCI, 1 m~ MgCl?. 0.02% gelatin, and 200 FM of each dNTP. Primers were designed based on the pub- lished sequence of the transthyretin gene [45] and had the following sequences: TTR-1. 5'-CGG TGA ATC CAA GTG TCC TC-3'. and TTR-2,5'-AAT GGG GAG ATG CCA AGT GC-3'. TTR-1 corresponds to nucleotides 1024- 1044 in exon 2 and TTR-2 corresponds to nucleotides 3343-3363 in exon 3. The resulting PCR product has a length of 246 base pairs (bp). Since these primers bracket intron 2 of the transthyretin gene, genomic DNA would yield a PCR product of > 2 kb which is not e5ciently amplitled under the chosen PCR conditions. We used 20 pmol of each primer (final concentration 2 p ~ ) and 0.25 units of Taq polymerase (Gibco-BRL) per l o p 1 final reaction volume. The PCR protocol consisted of an initial denaturing step of 95°C x 5 min followed by 30 cycles of 95-C x 30 s, 5 8 T x 60 s, 72°C x 40 s, and a final extension step of 72°C x 10 min.
PCR products were electrophoresed on 2% agarose- ethidium bromide gels. The gel was then washed for 20 min with 0.4 M NaOH. The PCR products were then blotted onto positively charged nylon membranes (Boehringer-Mannheim) using capillary transfer with 0.4 M NaOH for 3 h. The gels were examined under ultraviolet light before and after the blotting procedure to ensure that the PCR products were of the right size and that the transfer was complete. Following the blot, the membrane was baked for 1 5 min at 120°C. Approxi- mately 5 0 n g of the transthyretin probe were labelled with digoxigenin using a random priming kit (Boehringer-Mannheim). The membrane was prehybrid- ized at 45°C in hybridization buffer (Hybrisol I, Oncor) for 4 h and then hybridized overnight (also at 45°C). Post- hybridization stringency washes were as described for
Transthyretiri iri rnedulloblastonln 401
the Northern blot. Bound probe was detected using an anti-digoxigenin antibody linked to alkaline phosphatase with a photoluminescence kit (Boehringer-Mannheim). The membrane was exposed to X-ray film (Cronex. Dupont) for 15 min at RT.
To verify that the PCR product was specific, the product obtained with the choroid plexus papilloma was sequenced using a cycle sequencing protocol with fluo- rescent dideoxynucleotide terminators (Applied Bio- systems) followed by electrophoresis on an automated DNA sequencer (model 373 A, Applied Biosystems). Furthermore, the predicted PCR product contains two HinfI restriction sites at nucleotides 106-1 10 and nucleotides 238-242. Therefore, it ought to yield three fragments of 106, 132. and 8 bp on digestion. To verify this, PCR products from the positive control and all positive medulloblastomas and medulloblastoma cell lines were digested with HinfI. They were then electro- phoresed on an 8% denaturing polyacrylamide gel and detected by silver staining [7]. As a negative control, we substituted water for RNA in the reverse transcription procedure and used the resulting mixture instead of cDNA for PCR.
To ensure that the reverse transcription had indeed yielded cDNA, those cases that were negative for trans- thyretii cDNA were amplified with cDNA-specific primers for the human glyceraldehyde-3 phosphate dehydrogenase (G3PDH) gene (Clonetech).
Non-radioactive RNA in sihi hybridization
Non-radioactive RNA in situ hybridization was per- formed on tumours 2462, 2466. 2472, 3052 and 2410 with the latter serving as positive control. The medullo- blastoma cell lines MHH-MED-2 through -4 and D283 Med were also studied. A digoxigenin-labelled RNA probe was transcribed in vitro from the transthyretin- cDNA probe with SP6 RNA polymerase according to the manufacturer's instructions (Boehringer Mannheim). From the tumours, frozen sections were cut at 7 pm. The sections were mounted on positively charged glass slides (Probeon-Plus slides, Fisher), air dried for 2 h, and stored at - 80°C wrapped in aluminium foil until further use. Cytospins were prepared from the cell lines using a cytocentrifuge (Shandon), air-dried, and stored at - 80°C.
Prior to the hybridization procedure, the sections and cytospins were thawed at room temperature (RT),
0 199 5 Blackwell Science Ltd, Neuropathologg and Applied Neurobiologg. 21, 399-4109

402 S. Albrecht et a].
air-dried again. and then fixed in 4% paraformaldehyde in PBS for 5 min at RT followed by two washes of 5 min each with 2 x SSC. The slides were then treated with 0.25% acetic anhydride in 0.1 M triethanolamine (pH 8.0) for 5 min at RT and then washed 1 min each at RT with 2 x SSC and PBS. This was followed by a 30 min incubation at RT with 0.75% glycine in PBS and a 2 min wash in 2 x SSC. also at RT. The sections were then prehybridized in hybridization solution (50% formamide. 5 x SSC. 0.1% Tween-20) at 50°C for 2 h. Hybridization, was carried out at 50°C overnight in a moist chamber using z 20-100 ng of the digoxigenin-labelled probe per slide. Post-hybridization stringency washes were as fol- lows: 2 x SSC at 5O"C, 2 x SSC with 50% formamide at 50°C, 1 x SSC (at RT), and 0.2 x SSC (at RT), all for 15 min each. This was followed by three washes of 5 min each at RT with 0.1 M maleic acid (pH 7.5) with 0.15 M
NaCI. Bound probe was detected with a digoxigenin detection kit (Boehringer Mannheim) according to the manufacturer's instructions using an anti-digoxigenin antibody linked to alkaline phosphatase with nitroblue tetrazolium as chromogen. Finally, the slides were counterstained with nuclear fast red, dehydrated through graded alcohols, cleared in xylene, and mounted.
Western blot
Western blot analysis of transthyretin protein expression was performed on all six medulloblastoma cell lines and on the Hep-T. MHH-NB-11, and U-87 MG cell lines. Cells were washed three times with ice-cold PBS, and lysed for 15 min on ice in a buffer consisting of 50 rn~ Tris-HC1 (pH 8.0). 120 m NaCl, 1 m phenylmethylsulfonyl fluoride, 10 pg/ml leupeptin (Boehringer-Mannheim), 100 units/ml aprotinin (Calbiochern), and 0.5% Nonidet P-40 (Sigma). We used 1 ml lysis buffer per 10 million cells. Cell debris was removed by centrifugation at 13 000 x g for 10 min at 4°C. Soluble proteins were separated by electrophoresis through 15% SDS- polyacrylamide gels. They were then electroblotted onto a nitrocellulose filter. Prior to immunostaining, the filter was treated with 5% non-fat milk in PBS at 4°C over- night. The filter was then successively incubated with the following reagents (all at RT for 1 h each): rabbit anti-transthyretin antibody (Dako. 1:1000), mouse anti- rabbit immunoglobulin antibody (Jackson Laboratories, 1:lOOO). goat anti-mouse immunoglobulin antibody
(Cappel, 1:200), and APAAP complex (Dianova, 1:SO). Staining was developed with nitroblue tetrazolium/ X-phosphate.
lmmuno histochemistry
Cryosections were cut at 6 pm from the same tissue blocks that had been used for in situ hybridization. The sections were mounted on glass slides, air dried for 2 h, and then fixed in acetone at room temperature for 10 min. In addition, we studied 16 cases of medullo- blastoma on paraffin sections. These were cut at 4 pm, deparaffinized. rehydrated, incubated in 3% H,O, in methanol to block endogenous peroxidase activity. and washed in PBS. Cryosections and para& sections were incubated with the anti-transthyretin antibody (see above) diluted 1:500 and 1:100, respectively, in PBS at RT for 1 h. The biotinylated secondary antibody (swine anti-rabbit, Dako) was applied at a dilution of 1:500 for 1 h at RT. Staining was developed with a commercial avidin-biotin complex kit (Dako). As positive control, we used the same case of choroid plexus papilloma that had been used for RNA extraction and in situ hybridization. For a negative control, non-immune rabbit serum was substituted for the primary antibody.
R esu Its
The results are summarized in Table 1 and are presented in detail below.
Northern blot
The choroid plexus papilloma, three of the four medullo- blastomas, one of the six rnedulloblastorna cell lines, and the Hep-T cell line yielded a band at 0.7 kb which is the expected size of the transthyretin-mRNA [20. 421. The signal produced by the choroid plexus papilloma was very intense even after 5 min of exposure. One of the medulloblastornas (case 2462) produced a band that became discernible after = 10 min of exposure (Figure la). The band produced by rnedulloblastoma 3052 became visible after 3 h of exposure (Figure lb) while the bands produced by medulloblastorna 24 72, the medullo- blastoma cell line MHH-MED-2. and the Hep-T cell.line were seen after 3 days of exposure (Figure lc).
0 1995 Blackwell Science Ltd. Neuroputhology and AppIied Neurobiology, 21, 399409
Trnnsthgretin in rnedulloblnstomn 40 3
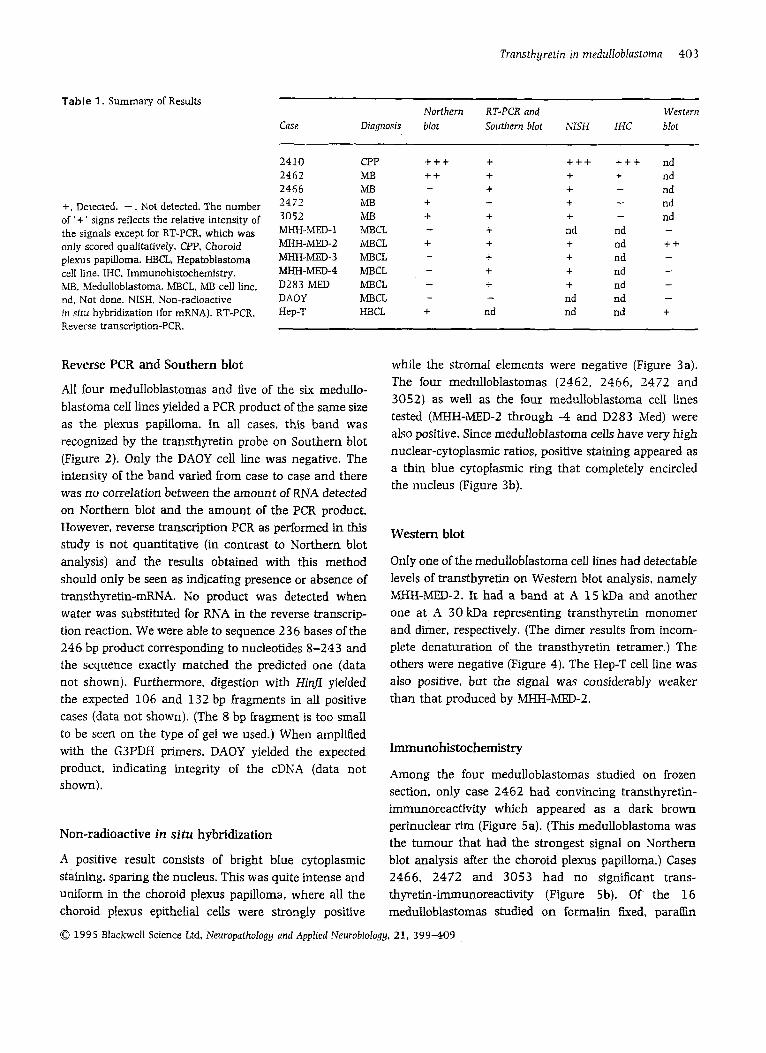
Table 1. Summary of Results Northern RT-PCR and Western
Case Diagnosis blot Southern blot NISH IHC blot
+. Detected. - . Not detected. The number of '+ ' signs reflects the relative intensity of the signals except for RT-PCR, which was only scored qualitatively. CPP, Choroid plexus papilloma. HBCL. Hepatoblastoma cell line. IHC. Irnmunohistochemistry. bB, Medulloblastoma. MBCL. MB cell line. nd. Not done. NISH, Non-radioactive in situ hybridization (for mRNA). RT-PCR. Reverse transcription-PCR.
2410 2462 2466 2471 3052 MHH-bED-1 M H H - m - 2 MHH-bED-3 MHH-MED-4 D283 M W DAOY Hep-T
CPP MB MB MB MI3 MBCL MBCL MBCL MBCL MBCL MBCL HBCL
+++ + ++ +
+ + + + +
+ + +
+ + +
-
-
- - - - - + nd
+++ + + + + nd + + + + nd nd
+++ nd + nd
nd nd nd
- - -
- nd nd ++ nd nd nd nd nd +
- - - -
Reverse PCR and Southern blot while the stromal elements were negative (Figure 3a).
All four medulloblastomas and five of the six medullo- blastoma cell lines yielded a PCR product of the same size as the plexus papilloma. In all cases, this band was recognized by the transthyretin probe on Southern blot (Figure 2). Only the DAOY cell line was negative. The intensity of the band varied from case to case and there was no correlation between the amount of RNA detected on Northern blot and the amount of the PCR product.
The four medulloblastomas (2462. 2466, 2472 and 3052) as well as the four medulloblastoma cell lines tested (MHH-MED-2 through -4 and D283 Med) were also positive. Since medulloblastoma cells have very high nuclear-cytoplasmic ratios, positive staining appeared as a thin blue cytoplasmic ring that completely encircled the nucleus (Figure 3b).
However, reverse transcription PCR as performed in this study is not quantitative (in contrast to Northern blot
blot
analysis) and the results obtained with this method should only be seen as indicating presence or absence of transthyreti-mRNA. No product was detected when water was substituted for RNA in the reverse transcrip- tion reaction. We were able to sequence 2 3 6 bases of the 246 bp product corresponding to nucleotides 8-243 and the sequence exactly matched the predicted one (data not shown). Furthermore, digestion with Hi@ yielded the expected 106 and 132 bp fragments in all positive cases (data not shown). (The 8 bp fragment is too small to be seen on the type of gel we used.) When amplified with the G3PDH primers, DAOY yielded the expected product, indicating integrity of the cDNA (data not shown).
Non-radioactive in situ hybridization
Only one of the meduIIoblastoma cell lines had detectabIe levels of transthyretin on Western blot analysis, namely MHH-MED-2. It had a band at A 15 kDa and another one at A 30 kDa representing transthyretin monomer and dimer, respectively. (The dimer results from incom- plete denaturation of the transthyretin tetramer.) The others were negative (Figure 4). The Hep-T cell line was also positive, but the signal was considerably weaker than that produced by MHH-MED-2.
Irnmunohistochernistry
Among the four medulloblastomas studied on frozen section, only case 2462 had convincing transthyretin- immunoreactivity which appeared as a dark brown perinuclear rim (Figure 5a). (This medulloblastoma was the tumour that had the strongest signal on Northern
A positive result consists of bright blue cytoplasmic blot analysis after the choroid plexus papilloma.) Cases staining, sparing the nucleus. This was quite intense and 2466, 2472 and 3053 had no significant trans- uniform in the choroid plexus papilloma, where all the thyretin-immunoreactivity (Figure 5b). O f the 16 choroid plexus epithelial cells were strongly positive medulloblastomas studied on formalin fixed, p a r a h
0 1995 Blackwell Science Ltd, Neuropathology and Applied NeurobioIogy. 21, 3 9 9 4 0 9

404 S. Alln-eclit el al.
0 t (u
r
1
N a * N I
N LD t N -
N h t N -
9 I:
A 4 5
w z - 2
6 W I: i I z -
M 00 cv n -
Figure 2. Southern blot analysis of PCR products from reverse transcription PCR with transthyretin-cDNA specific primers. Lanes are identified with the tumour number or cell line name. 'Water', negative control for which water was substituted for RNA in the reverse transcription reaction. All four medulloblastomas and five of the six medulloblastoma cell lines are positive. Only the DAOY cell line is negative.
Figure 1 . Northern blot hybridized with transthyretin-cDNA probe and autoradiographed for 10 min (a). 3 h (b) and 3 days (c). Lanes are identified with the tumour number or cell line name. 'Brain'. normal lrontal lobe from a surgical specimen. Cases 2.110 (the choroid plexus papilloma) and medulloblastoma 2462 have a visible band of the expected size (0.7 kb) alter 10 min 01 exposure. The band in case 3052 is visible after 3 h while the bands in case medulloblastoma 2472. the medulloblastoma cell line MHH-MED-2 and the Hep-T cell line were visible only after 3 days. (The band in H e p T appears to be of a larger size due to distortion in the agarose gel.)
embedded sections, only one contained transthyretin-
gested that they were reactive astrocytes of the cerebellar moiecular layer in areas of tumour invasion.
Figure 3. In situ hybridization for transthyretin mRNA. a. AU the epithelial cells of the choroid plexus papilloma are positive while
cell line (MHH-MED-~). Original magnscations x 200 and x 400. respectively.
immunoreactive However, their mo~hology sug- the stroma is negative. b, Positive staining in a medullob]astoma
Discussion
Transthyretin is a serum protein synthesized and secreted by hepatocytes that serves as a transport molecule for highly Iipophilic substances such as thyroid hormones and retiiol. Since it migrates ahead of albumin on serum protein electrophoresis, it is also known -as 'prealbumin.' It is. however, chemically not related to the latter and therefore 'transthyretin' now is the preferred term. Transthyretin is made up of four
identical subunits and has a molecular weight of 5 5 kDa. Transthyretin has been extensively studied in the last decades and both its three-dimensional structure [5] and the complete sequence of the transthyretin gene [45] have been known for several years.
Transthyretin expression is not restricted to the liver, however. In the rat. small amounts of transthyretin- mRNA equivalent to about 1-2% of the levels found in the liver can also be detected on Northern blot analysis in the spleen, stomach, heart and skeletal muscle [42].
0 1995 Blackwell Science Ltd. Neuropathology and Applied Neurobiologg. 21, 399-409

F
rh ? I I P
D
[r) 00 N n
> 0 d
Figure 4. Western blot analysis of cell lines with an anti-transthyretin antibody. The lanes are identified with the cell lines’ names. The lower band represents transthyretin monomer while the upper band (seen in MHH-IVIED-2) consists of transthyretin dirner. Transthyretin-protein is only detectable in the rnedulloblastorna cell line ?VIM-MED-2 and in the Hep-T cell line.
In the rat, high levels of transthyretin-mRNA and immunoreactive transthyretin are also detectable in the pigment epithelium of the retina [lo, 161 and in the fetal visceral yolk sac [43]. In humans. transthyretin is also expressed by the glucagon-producing cells of the pan- creatic islets [9. 12, 23-25, 311 and transthyretin- immunoreactivity is seen in the proximal convoluted tubules of the kidney [3, 311. Scattered transthyretin- immunoreactive cells are also present throughout the gastro-intestinal mucosa from the stomach to the colon [31]. In the human fetus, transthyretin- immunoreactivity of pancreatic islets. gut mucosa and renal tubules can be seen as early as 12 weeks gestation [181.
In recent years, it has become apparent that the choroid plexus is another major site of transthyretin synthesis. Transthyretin-immunoreactivity of both the fetal [36] and adult [2] choroid plexus was described several years ago. Since the staining tends to be more pronounced at the apical pole of the choroid plexus epithelial cells [ 3 61 and because the immunoreactivity could be localized to the Golgi complex and the rough endoplasmic reticulum by immuno-electron microscopy [2], it was suggested that the choroid plexus actually synthesizes transthyretin rather than simply transport- ing serum transthyretin into the CSF. This hypothesis has been confirmed by demonstrating transthyretin- mRNA in the choroid plexus both on Northern blot [20, 421 and by in situ hybridization [ZO]. The choroid plexus secretes its transthyretin into the CSF, where
Figure 5. Immunohistochemistry with anti-transthyreti antibody. The rnedulloblastoma with the highest level of transthyretin-mRNA expression on Northern blot (2462) stains positively (a) while another rnedulloblastoma (2472) is negative (b). Original rnagnscation x 400.
transthyretin constitutes - 5% of the total protein [ZS]. Transthyretin appears to represent a major product of the choroid plexus epithelium, since transthyretin- mRNA levels are up to 30-100 fold higher in the choroid plexus than in liver [14. 27, 351. The precise role of transthyretin in the CSF is not yet clear, although it is thought to facilitate the transport of thyroxin into the CSF [15].
Transthyretin expression by the choroid plexus epi- thelium also appears to be of developmental importance. In the human fetus, transthyretin-immunoreactivity in the choroid plexus can be demonstrated as early as the
0 1995 Blackwell Science Ltd. Neuropnthology nnd Applied Neurobiology. 21. 399-409

406 S. Alhreclzf et al.
sixth week of gestation [36] while in the rat fetus, transthyretin-immunoreactivity and transthyretin- mRNA can already be demonstrated in the tela choroi- dea, which constitutes the primordium of the choroid plexus [27, 441. Several studies both in the rat and in humans have c o h e d that the choroid plexus and its primordia are the only sites of transthyretin synthesis in the CNS [2, 20, 26, 441.
By and large, transthyretin expression in neoplasms parallels its normal distribution pattern. Pancreatic islet cell tumours. especially glucagonomas. non-functioning tumours. and malignant insulinomas are often transthyretin-immunoreactive [9, 22. 241 as are A40-100% of carcinoids depending on their location [9, 341. Renal cell carcinomas can also be transthyretin-immunoreactive [3]. The majority of choroid plexus papillomas and choroid plexus carcinomas are also transthyretin-bunoreactive [ l , 4. 19. 33, 371 and contain transthyretin-mRNA demonstrable by in situ hybridization [ 191. Astrocyto- mas, oligodendrogliomas, glioblastomas, ependymomas. and meningiomas do not have transthyretin- immunoreactivity [4, 19. 411. To our knowledge, medulloblastomas have not been studied previously for transthyretin expression.
In this study, transthyretin-mRNA could be demon- strated in four of four medulloblastomas and in five of six medulloblastoma cell lines. We believe that the transthyretin-mRNA stems from the tumour cells and not from contaminating choroid plexus. First, no choroid plexus was seen on the frozen sections from the tumour fragments that were selected for RNA extraction. Second, transthyretin-mRNA could also be detected by in situ hybridization and was clearly located in tumour cells. Third, transthyretin-mRNA was also found in five of six medulloblastoma cell lies, which do not contain contaminating choroid plexus.
The level of expression of transthyretin mRNA in medulloblastomas is clearly quite variable. Using reverse transcription PCR which is a highly sensitive method, transthyretin cDNA (and therefore mRNA) could be detected in four of four medulloblastomas and five of six medulloblastoma cell lines. However, on Northern blot analysis, a strong signal was detected in only one of the four medulloblastomas (case 2462) and only one of the cell lines was positive (MHH-MED-2). These two cases were also the only ones that expressed transthyretin protein detectable by either i m u n o -
histochemistry or Western blot. The most likely explanation for this discrepancy is that levels of trans- thyretin mRNA and protein expression in most medulloblastomas and medulloblastoma cell lines are too low to be detectable on Northern and Western blot, respectively.
Since we had only four medulloblastomas with fresh frozen tissue at our disposal, we decided to study a number of routinely processed medulloblastomas by transthyretin-immunohistochemistry in order better to define the frequency of transthyretin expression in medulloblastoma. None of these 16 formalin fixed. paraffin embedded medulloblastomas contained transthyretin-immunoreactive cells, except for one, where reactive astrocytes in areas of tumour invasion were faintly positive. (Transthyretin-immunoreactivity of reactive astrocytes has been noted previously [l].) However, formah does reduce transthyretin- immunoreactivity compared with other fixatives [23] and tissues that are transthyretin-immunoreactive on frozen sections can be negative after formalii furation [ 161. Immunohistochemistry on routinely processed histological material may therefore underestimate trans- thyretin expression in medulloblastoma. Nevertheless, the possibility of transthyretin expression in medullo- blastoma should be kept in mind when performing immunohistochemical studies on poorly differentiated ‘small blue cell’ tumours of the cerebellum, especially if the difIerential diagnosis includes a poorly differentiated. mostly solid choroid plexus carcinoma.
hiIedulloblastoma is the prototypical multipotential primitive neuroectodermal tumour of the cerebellum. Medulloblastomas can have multiple lines of differentia- tion, at the light and electron microscopical level and by immunohistochemistry. They can express markers and display features of neuronal, astrocytic. ependymal, retinal, and rnyogenic differentiation [8, 401 and OUT finding of transthyretin expression in medulloblastomas and medulloblastoma cell lines has to be interpreted in this context.
Transthyretin expression in medulloblastoma may be part of the retinal differentiation observed in some medulloblastomas. Up to one half of medulloblastomas contain cells that are immunoreactive for retinal S-antigen and rod-opsin, which are .both markers of photoreceptor differentiation [6, 13, 321. In addition, medulloblastomas can also express another marker of photoreceptor differentiation, namely the interphoto-
0 1995 Blackwell Science Ltd. Neuropathology and Applied Neurobiology. 21. 399-409

Trnnsthyretiri in rnedulioblnstorna 4Oi
receptor retinoid-binding protein [29]. Western blot analysis of a case of medulloblastoma demonstrated expression of retinal S-antigen and rod-opsin [ 301. Inter- estingly. this tumour also contained 1 1-cis-retinal which was converted to all-trans-retinal upon illumination of a tumour homogenate [30]. In the mammalian retina, this conversion takes place only in the retinal pigment epi- thelium. and the authors therefore conclude that some medulloblastomas may also contain retinal pigment epithelium-like cells in addition to cells with photorecep- tor differentiation. Since the retinal pigment epithelium is also the only site of transthyretin synthesis in the eye [lo. 161, our finding of transthyretin expression in medulloblastoma could provide additional evidence for retinal pigment epithelium differentiation in these tumours. Alternatively, transthyretin expression in medulloblastomas may also indicate choroid plexus differentiation. However, we have no additional evidence (either morphological or biochemical) for the latter.
In the end, transthyretin expression in medulloblasto- mas may simply reflect the pluripotential nature of the medulloblastoma tumour cells. An example of this is the hmH-MED-2 cell line. In addition to transthyretin. this line expresses vimentin and markers of early neuronal differentiation such as neuron-specific enolase, synapto- physin. the GD2 and A2B5 gangliosides, and the poly- sialylated 'embryonal' form of NCAM [39]. However, it does not express neurofilaments, which are markers of late neuronal differentiation, and also does not express GFAP or keratin [39]. MHH-MED-2 may therefore have the phenotype of a neuronalh-etinal 'precursor cell and could be useful as a n in vitro model for studying their differentiation.
In summary, our results indicate that low level expression of transthyretin-mRNA occus frequently in medulloblastomas and medulloblastoma cell lines while expression of significant levels of transthyretin-mlWA or expression of transthyretin protein occur more rarely. The expression of transthyretin-mRNA and protein reflects the primitive pluripotential character of medulloblastoma cells. It also limits the useful- ness of transthyretin as a marker of choroid plexus tumours.
Acknowledgements
We wish to thank Dr J. Herbert (Department of Neurology, Columbia University, College of Physicians
and Surgeons. New York) for his generous gift of the transthyretin plasmid and D. Kajetanowicz for excellent technical assistance. S.A. is a recipient of a scholarship from the Alexander-von-Humboldt Foundation. T.B. is supported by the Frite-Thyssen Foundation. This study was supported by grants from the Deutsche Urebshiefe (W42/91 P i l ) and the Deutsche Forschungsqemein- schaft (SFB 400).
References
Albrecht S, Rouah E. Becker LE. Bruner J. Transthyretin immuno-reactivity in choroid plexus neoplasms and brain metastases. Mod Pnthol 1991: -k 610-14 Aleshire SL. Bradley CA. Richardson LD, Par1 FL. Localiz- ation of human prealbumin in choroid plexus epithelium. 1 Histochem Cytochem 1983: 31: 608-12 Ang LC, Debowski T. Michalski R. Immunolocalization of prealbumin (transthyretin) in renal cell carcinoma. Histopathology 1991: 18: 565-8 Ang LC. Taylor AR, Bergin D, Kaufrnann JCE. An immuno- histochemical study of papillary tumours in the central nervous system. Cancer 1990: 65: 2712-19 Blake CCF, Geisow MJ, Oatley SJ. RCrat B. Rerat C. Structure of prealbumin: secondary, tertiary and quaternary inter- actions determined by Fourier rehernent at 1.8 A. ] MoI Biol 1978: 121: 339-56 Bonnin JM, Perentes E. Retinal S-antigen immunoreactivity in medullo-blastomas. Acta h'europathol 1988: 76: 201-7 Budowle B. Chakraborty R. Giusti AM, Eisenberg AJ, Allen RC. Analysis of the VNTR locus DlSSO by the PCR followed by high-resolution PAGE. Am J Hum Genet 1991: 48: 13 7 4 4 Burger PC. Scheithauer BW. Vogel FS. Medulloblastoma. In Surgical Pathology of the Nenlous System and its Coverlngs. New York Churchill Livingstone. 1991: 339-54 Bussolati G. Papotti M. Sapino A. Binding of antibodies against human pre-albumin to intestinal and bronchial carcinoids and to pancreatic endocrine tumours. Virchows Arch [B] 1984; 45: 15-22
10 Cavallaro T. Martone RL, Dwork AJ, Schon EA. Herbert J. The retinal pigment epithelium is the unique site of trans- thyretin synthesis in the rat eye. Invest Opthnlrnol Vis Sci
11 Chomczynski P. Sacchi N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol- chloroform extraction. Anal. Biochem. 1987; 162: 156-9
12 Collins VP, Jacobson B. Pettersson T, Carlstrom A. Mono- clonal antibodies to transthyretin. Scand ] Clin Lab Invest
13 Czerwionka M. Korf HW. Hoffmann 0. Busch H. Schachenmayr W. Differentiation in medulloblastoma: cor- relation between immunocytochemical demonstration of
1990: 31: 497-501
1986: 46: 761-9
0 199 5 Blackwell Science Ltd. Neurapafhalagy arid Applied Neurobiology. 21, 3 9 9 4 0 9

408 S. AIbrecht et al.
photoreceptor markers (S-antigen. rod-opsin) and the sur- vival rate in 66 patients. Actn Neuroptkol 1989: 78:
14 Dickson PW. Aldred AR. Marley PD, Guo-Fen T, Howlett GJ, Schreiber G. High prealbumin and transferrin mRNA levels in the choroid plexus of rat brain. Biochem Biophys Res Corniiiun 1985: 127: 890-5
15 Dickson PW. Aldred AR. Menting JGT, Marley PD. Sawyer WH. Schreiber G. Thyroxine transport in choroid plexus. ] Biol Chem 1987: 262: 13907-15
16 Dwork AJ, Cavallaro T. Martone RL, Goodman DS. Schon EA. Herbert J. Distribution of transthyretin in the rat eye. Invest Ophthnlmol Vis Sci 1990: 31: 489-96
17 Friedman HS. Burger PC. Bigner SH et al. Establishment and characterization of the human medulloblastoma cell line and transplantable xenograft D283 Med. 1 Neuropathol E x p NeuroI 1985: 44: 592-605
18 Gray HDA. Gray ES. Home CHW. Sites of prealbumin production in the human fetus using the indirect immuno- peroxidase technique. Virchows Arch [A] 1985: 406:
19 Herbert J. Cavallaro T, Dwork AJ. A marker for primary choroid plexus neoplasms. A m ] Pathol 1 9 9 0 136:
20 Herbert J, Wilcox JN, Pham KTC et al. Transthyretin: a choroid plexus-specific transport protein in human brain. Neurology 1986: 36: 900-11
21 Jacobsen PF, Jenkyn J, Papadimitriou JM. Establishment of a human medulloblastoma cell line and its heterotrans- plantation into nude mice. J NeuropathoI Exp Neurol 1985:
22 Jacobson B, Carlstrom A, Collins VP, Grimelius L. Trans- thyre t i in endocrine pancreatic turnours. A m J Pnthol
23 Jacobson B. Collins VP. Grimelius L. Pettersson T. Sandstedt B. Carlstrom A. Transthyretin immunoreactivity in human and porcine liver, choroid plexus, and pancreatic islets. 1 Histochem Cytochem 1989: 37: 31-7
24 Jacobson B. Pettersson T, Sandstedt B. Carlstrom A. Pre- albumin in the islets of Langerhans. IRCS Med Sci 19 79; 7 590
25 Kameko M. Ichikawa M. Katsuyama T. Kanai M, Kato M. Akamatsu T. Immunohistochemical localization of plasma retiol-binding protein and prealbumin in human pancre- atic islets. Histochem ] 1986: 18: 164-8
26 Kato M. Kato K, Goodman DS. Immunocytochemical studies on the localization of plasma and cellular retinol- binding proteins and of transthyreti (prealbumin) in rat liver and kidney. CeIl Biol 1984; 98: 1696-704
27 Kato M. Soprano DR, Makover A. Kato K, Herbert J. Goodman DS. Localization of immunoreactive transthyretin (prealbumin) and of transthyreti mRNA in fetal and adult rat brain. Diflerentiation 1986: 31: 228-35
629-36
463-73
13 1 7-25
4 4 472-85
1989: 134: 465-71
28 Eileine TO, Weber L. Zofel P. Changes with age in leucocyte counts, protein and IgG content of lumbar cerebrospinal fluid (CSF) of patients with inflammatory, ischemic. and haemorrhagic diseases or tumours of the central nervous system (CNS). Zeitschr Gerontol 1988: 21: 102-5
29 Korf HW. Korf B, Schachenmayr W, Chader GJ, Wiggert B. Irnmunocytochemical demonstration of interphotoreceptor retinoid-binding protein in cerebellar rnedulloblastoma. Acta Neuropathol. 1992: 83: 482-7
30 Kramm CM. Korf Kw, Czenvionka M. Schachenmayr W. de Grip WJ. Photoreceptor differentiation in cerebellar medulloblastoma: evidence for a functional photopigment and authentic S-antigen (arrestin). Actu Nruropntliol. 199 1: 81: 296-302
31 Liddle CN. Reid WA. Kennedy JS. Immunolocalization of prealbumin: distribution in normal human tissue. ] Pnthol
32 Maraziotis T. Perentes E. Karamitopoulou E et at Neuron- associated class III P-tubulin isotype. retinal S-antigen. synaptophysin. and glial fibrillary acidic protein in human medulloblastomas: a clinicopathological analysis of 3 6 cases. Actu Neuropathol 1992: 84: 355-63
33 Matsushma T, Jnoue T, Takeshita I. Fukui M, Iwaki T, Kitamoto T. Choroid plexus papillomas: an immuno- histochemical study with particular reference to the co- expression of prealbumin. Neurosurgery 1988: 23: 384-9
34 Miller ID. Reid WA. Liddle CN, Home CHW. Immuno- localization of prealbumin as a marker for carcinoid tumours. 1 Pathol 1984: 143: 199-204
35 Mita S. Maeda S, Shimada K, Araki S. Analyses of pre- albumin mRNAs in individuals with familial amyloidotic polyneuropathy. 1 Biochem 1986: 100: 1215-22
36 M ~ l l g k d K. Jacobsen M, Jacobsen GK, Clausen PP. Saunders NR. Immunohistochemical evidence for an intracellular localization of plasma proteins in human foetal choroid plexus and brain. Neurosci Lett 1979: 14:
3 7 Paulus W. JSnisch W. Clinicopathologic correlations in epithelial choroid plexus neoplasms: a study of 52 cases. Acta Neuropathol 1990: 80: 6 3 5 4 1
38 Pietsch T, Gottert E. Meese E et al. Characterization of a continuous cell l i e (MHH-NB-11) derived from advanced neuroblastoma. Anticancer Res 1988: 8: 1329-34
39 Pietsch T. Scharmann T. Fonatsch C et al. Characterization of five new cell lines derived from human primitive neuro- ectodermal tumours of the central nervous system. Cancer. Res 1994: 54: 3278-87
40 Russell DS, Rubinstein LJ. Medulloblastoma. In Pathology of Tumours of the Nervous System. Baltimore: Williams &
41 Seitz RJ, Wechsler W. Immunohistochemical demonstration of serum proteins in human cerebral gliomas. .Acts Neuropathol 1987: 73: 145-52
1985; 146: 107-13
85-90
Willrin~, 1989: 251-74
0 1995 Blackwell Science Ltd. Neuropathology and Applied Neurobiology. 21. 399409

Transthyretin in meditlbblostornn 409
42 Soprano DR. Herbert J, Soprano KJ, Schon EA. Goodman DS. Demonstration of transthyretin mRNA in the brain and other extrahepatic tissues in the rat. ] B i d Cliern 1955; 260: 11793-8
43 Soprano DR. Soprano KJ, Goodman DS. Retinol-binding protein and transthyretin mRNA levels in visceral yolk sac and liver during fetal development in the rat. Proc NntZ Acad Sri USA 1986; 83: 7 7 3 0 4
44 Thomas T. Power B. Hudson P. Schreiber G. Dziadek M. The expression of transthyretin mRNA in the developing rat brain. Dev Biol 1988: 128: 415-27
4 5 Tsuzuki T. klita S. Maeda S . Araki S. Shimada K. Structure of the human prealbumin gene. J BioI Chern 1985: 160: 1 2 2 2 4 7
Received 5 Jarizmry 1995 Accepted after revision 10 J Z U W 1 9 9 5
0 1995 Blackwell Science Ltd. Neuroputhology and Applied Neurobiofogy. 21, 399409



![Medulloblastoma: [Print] - eMedicine Neurology · emedicine.medscape.com eMedicine Specialties > Neurology > Pediatric Neurology Medulloblastoma George I Jallo, MD, Associate Professor](https://img.dokumen.tips/doc/110x75/5d472c3c88c993527c8b60e5/medulloblastoma-print-emedicine-neurology-emedicinemedscapecom-emedicine.jpg)

![Medulloblastoma: [Print] - eMedicine Neurology · accounts for approximately 7-8% of all intracranial tumors and 30% of ... Incidence of medulloblastoma is 1.5-2 cases per ... Medulloblastoma:](https://img.dokumen.tips/doc/110x75/5b7fc2317f8b9ae6088caa0e/medulloblastoma-print-emedicine-accounts-for-approximately-7-8-of-all.jpg)