Embed Size (px)
Citation preview

1877Journal of Cell Science 111, 1877-1888 (1998)Printed in Great Britain © The Company of Biologists Limited 1998JCS9764
Three distinct steps in transport of vesicular stomatitis virus glycoprotein
from the ER to the cell surface in vivo with differential sensitivities to GTP γS
Rainer Pepperkok 1,*, Martin Lowe 1,**, Brian Burke 2 and Thomas E. Kreis 1,§
1Department of Cell Biology, Sciences III, University of Geneva, 30 quai Ernest-Ansermet, CH-1211 Geneva 4, Switzerland2Department of Anatomy, University of Calgary, 3330 Hospital Drive, Calgary, AB T2N 4N1, Canada*Present address: Light Microscopy Laboratory, Imperial Cancer Research Fund, 44 Lincoln’s Inn Fields, London WC2A 3PX, UK**Present address: Cell Biology Laboratory, Imperial Cancer Research Fund, 44 Lincoln’s Inn Fields, London WC2A 3PX, UK§Author for correspondence (e-mail: [email protected])
Accepted 20 April; published on WWW 15 June 1998
Microinjected GTPγS revealed three distinct steps in theexocytic transport of the temperature sensitive glycoproteinof vesicular stomatitis virus (ts-O45-G) from the ER to thecell surface in intact Vero cells. While COPII dependentexport of ts-O45-G from the ER is blocked in cells injectedwith recombinant protein of a dominant mutant of SAR1a(SAR1a[H79G]) inhibited in GTP hydrolysis, neitherinjected GTPγS nor antibodies against β-COP (anti-EAGE)interfere with this transport step significantly. In contrast,transport to the Golgi complex is blocked by 50 µM GTPγS,a dominant mutant of ARF1 (ARF1[Q71L]) inhibited inGTP hydrolysis, or microinjected anti-EAGE, but injectedSar1a[H79G]p has no effect. Microinjection of GTPγS orexpression of ARF[Q71L] rapidly induces accumulation ofCOPI coated vesicular structures lacking ts-O45-G. Finally,transport of ts-O45-G from the trans-Golgi network (TGN)
to the cell surface is inhibited only by high concentrationsof GTPγS (500 µM). Interestingly, this step is only partiallybrefeldin A sensitive, and injected antibodies against β-COPand p200/myosin II, a TGN membrane associated protein,have no effect. These data provide first strong in vivoevidence for at least three distinct steps in the exocyticpathway of mammalian cells regulated by different sets ofGTPases and coat proteins. COPII, but not COPI, isrequired for ER export of ts-O45-G. COPI plays a role insubsequent transport to the Golgi complex, and a so farunidentified GTPγS sensitive coat appears to be involved intransport from the TGN to the cell surface.
Key words: Exocytic membrane traffic, Coat protein, GTP bindingprotein, GTP analog, Microinjection
SUMMARY
r-teyndortndr to
TCuentPIlate6),toin
t al.,l.,een
es, beslls.n to
INTRODUCTION
Transport between adjacent compartments in the endocyticexocytic pathways is thought to be mediated by vesicular carr(Palade, 1975). All so far identified transport vesicles are coawith distinct coat protein complexes (Rothman and Orci, 19Kreis et al., 1995; Schekman and Orci, 1996). Clathrin aadaptor proteins are associated with vesicles involvedendocytosis and transport from the TGN to endosom(Robinson, 1994). COPII coated vesicles mediate ER to Gtransport in yeast (Barlowe et al., 1993, 1994), and these prothave also been identified in mammalian cells (Kuge et al., 19Shaywitz et al., 1995; Paccaud et al., 1996; Tang et al., 19COPI has been shown to be involved in transport between theand the Golgi complex (Hosobuchi et al., 1992; Pepperkok et1993; Peter et al., 1993; Griffiths et al., 1995; Aridor et al., 19Bednarek et al., 1995; Letourneur et al., 1994; Cosson et1996), and it is also involved in intra Golgi membrane traf(Orci et al., 1997). Most recently, membrane transport betwthe ER and the Golgi complex has been visualized directlyliving cells, using a chimera of ts-O45-G with the greefluorescent protein (ts-G-GFP; Scales et al., 1997; Presley e
andiersted
92;nd
ines
olgieins94;97). ER
al.,95; al.,ficeen inn
t al.,
1997). COPII exports ts-G-GFP from the ER into vesiculatubular clusters (VTCs) which may build up an ‘intermediacompartment’ (IC) when membrane traffic is arrested bincubation of cells at 15°C (for a review see also Bannykh aBalch, 1997). These structures then transform into transpcomplexes (TCs), probably as COPII is replaced by COPI, amove as entire structures in a microtubule dependent mannethe Golgi complex. It has been suggested that recycling of associated factors to the ER by COPI is essential for subseqdelivery of cargo to the Golgi (Scales et al., 1997). A COrelated coat has also been proposed to regulate early to endosome transport (Whitney et al., 1995; Aniento et al., 199and a new complex of coat proteins (AP3) with homologies clathrin adaptors (AP1 and AP2) has been identified mammalian cells and yeast (Pevsner et al., 1994; Newman e1995; Simpson et al., 1996; Dell’Angelica, 1997; Cowles et a1997). Assembly of these coat proteins onto membranes has bproposed to be the driving force for budding of transport vesicland it has been speculated that related coat proteins mayinvolved in the formation of most, if not all, vesicular carriermediating distinct membrane transport steps in eukaryotic ce
Some of these coat protein complexes have been show

1878
alticitivets-ive1),0°Cly,nctntdR
s
nsII.
eassm
wasiesal.,al.,-
astehe1pe
eiss a
h
R. Pepperkok and others
directly interact with vesicular cargo. AP1 and AP2, foexample, can interact with mannose-6-phosphate receptothe TGN and LDL receptor on the plasma membranrespectively (Pearse and Robinson, 1990). COPI interacts wproteins that have an ER retrieval sequence at their carbotermini (Cosson and Letourneur, 1994; Letourneur et al., 19Lowe and Kreis, 1995; Harter et al., 1996; Sohn et al., 19or proteins of the p24 family of membrane proteins with anmotif suggested to mediate anterograde transport (Fiedler e1996), and COPII in yeast has been shown to play a roleselective packaging of cargo into ER derived transport vesic(Rexach et al., 1994; Campbell and Schekman, 199Therefore, it can be assumed that these coat proteins mayplay an important function in sorting of cargo into the differevesicular carriers mediating membrane transport.
Membrane transport is, in addition to coat proteins, aregulated by small ras-like GTPases (Zerial and Huber, 199Members of the Arf protein family, as well as Sar1p, plaessential roles in assembly of coat proteins onto membra(Oka et al., 1991; Donaldson et al., 1991; Palmer et al., 19Robinson and Kreis, 1992; Stamnes and Rothman, 1993; Tret al., 1993; Barlowe et al., 1993; Chen and Shields, 1996; Wet al., 1997), and rab proteins have been shown to play a in targeting and docking of transport vesicles to and wacceptor membranes (Zerial and Stenmark, 1993; Novick Brennwald, 1993; Nuoffer and Balch, 1994). Trimeric proteins have also been implicated in some step of formatiotransport intermediates in the regulated secretory pathw(Leyte et al., 1992; Pimplikar and Simons, 1993). Again, relaprotein families with GTPase activity appear to be ubiquitoand universal regulators of vesicular membrane traffic.
To further characterize the regulation of vesicular membratraffic in the exocytic pathway in living cells, we usemicroinjection of non-hydrolysable nucleotide analogs. Nohydrolysable GTP analogs have been shown to interfere wuncoating of coated vesicles in vitro, a prerequisite fsubsequent fusion of these transport intermediates with tacceptor membranes (Melancon et al., 1987; Beckers Balch, 1989; Barlowe et al., 1994; Oka and Nakano, 199Furthermore, mutants of Arfs and Sar1p affecting their GTPactivities lead to severe disorganization of exocytic membratransport (Dascher and Balch, 1994; Teal et al., 1994; Zh
50040030020010000
20
60
100
39.5->31
15->31
20->31
GTPγS (µM)
39.5->310
20
60
100
A B
ts-O
45-G
at t
he c
ell s
urfa
ce (
%)
rr ine,ithxy-
94;96) F-t al., inles7). alsont
lso5).ynes93;aubestroleithandGn of
aytedus
nedn-ith
orheirand4).asene
ang
et al., 1994; Aridor et al., 1995). To visualize eventuinhibitory effects of injected nucleotide analogs in the exocytransport we used as a marker the temperature senstransmembrane glycoprotein of vesicular stomatitis virus, O45-G. This protein accumulates at the nonpermisstemperature (39.5°C) in the ER (Bergmann et al., 198accumulates at 15°C (Saraste and Kuismanen, 1984) or 2(Matlin and Simons, 1983) in the IC or the TGN, respectiveand is transported to the cell surface at 31°C. Three distitransport steps could be identified in vivo by their differesensitivities to GTPγS, corroborating previous results obtaineby in vitro assays or with semi-intact cells. Export from the Eis not affected by GTPγS and requires COPII; transport in TCto the Golgi complex is sensitive to GTPγS and inhibition ofCOPI function; finally, delivery of ts-O45-G from the TGN tothe plasma membrane is inhibited only by high concentratioof GTPγS, but insensitive to perturbation of COPI and COP
MATERIALS AND METHODS
Cell culture, microinjection and immunofluorescencelabelingVero cells (African green monkey kidney cells, ATCC CCL81) wermaintained and infected with ts-O45 VSV (Indiana serotype) described earlier (Kreis and Lodish, 1986). Microinjection waperformed on a computer automated microinjection system (AIS froZeiss) as described (Pepperkok et al., 1993). Immunofluorescenceperformed as described (Pepperkok et al., 1993) with antibodagainst calnexin (Hammond and Helenius, 1994), PDI (Vaux et 1990), cd8 (Ponnambalam et al., 1994), ERGIC53 (Schweizer et 1988), γ-adaptin (Robinson and Kreis, 1992), mammalian KDELreceptor (Griffiths et al., 1994), p58 (Bloom and Brashear, 1989),β-COP (Duden et al., 1991; Pepperkok et al., 1993), β′-COP (Lowe andKreis, 1995), mammalian Sec13p (Tang et al., 1997), rab1a (Saret al., 1995), p200 (Narula et al., 1992) and VSV-G (Kreis, 1986). TGolgi complex in Vero cells stably expressing myc-tagged NAGTwas visualized using a monoclonal antibody against a myc-epito(Evan et al., 1985) as described elsewhere (Pierre et al., 1994).
Fluorescence microscopy and image analysesImages of immunofluorescence-labeled cells were recorded on a Zinverted fluorescence microscope (Axiovert TV135) equipped withcooled, slow scan CCD camera (Photometrics CH250, 1,317×1,035pixels, Tucson, AZ, USA), controlled by a Power Macintos
15->31 20->31 ( C)°
500µM ATP S g
500µM GMPPNP
Fig. 1.Transport of ts-O45-G to the plasmamembrane in GTPγS injected cells. Virusinfected Vero cells, where ts-O45-G had beenaccumulated in the ER, IC or TGN at therespective temperatures, were injected withdifferent concentrations of GTPγS (A), or with500µM GMPPNP or ATPγS (B) and 5 mg/mlcoumarin BSA. After injection, the temperaturewas shifted to 31°C for 1 hour and the amountof ts-O45-G on the cell surface quantified byimmunofluorescence and image analysis. Thedata are normalized to the value obtained inneighboring non-injected cells (100%).Transport from the ER and IC to the plasmamembrane is efficiently blocked by 100µMGTPγS, while transport from the TGN is onlyinhibited at 500µM GTPγS.

1879Three steps in exocytic membrane traffic
celloH) as
ted
PIIrane
tsing
lseN.
touragece
e
x.d
theate).r
8100/100. Images were further processed with the software pacIPLab spectrum V3.0 (Signals Analytics Corp., Vienna, VA, USAbefore printing on either Tmax100 film using a slidewriter IS200(Focus Graphics, Forster City, CA, USA), or on paper using a Xalaser press 1800 (Conware Informatic AG, Baar, Switzerland).
ts-O45-G cell surface labeling was quantified as previously descr(Pepperkok et al., 1993). Co-localization of ts-O45-G with differemarker proteins at 15°C was determined as already described (Grifet al., 1995). For each cell analyzed, a square area (200×200 imagepixels, equivalent to about 15-30% of the total projected cell area) randomly chosen, digitally zoomed using the IPLab software, andvesicular structures positive for ts-O45-G marked by circular overl(with a diameter equivalent to 250 nm in microscope coordinates). different membrane compartment marker proteins investigated windicated in the same way. Two structures were scored as co-localwhen their respective overlays overlapped by more than 50%.
Metabolic labeling and biochemical analyses ofmaturation of ts-O45-G~500 cells, grown on glass coverslips (Pepperkok et al., 1993), winfected with ts-O45 VSV and incubated for 2 hours in normal cultumedium, followed by 10 minutes incubation in labeling mediu(culture medium lacking methionine and cysteine). Cells wemetabolically labeled for 20 minutes with 2.8 mC[35S]methionine/cysteine (Amersham) per ml labeling medium. Cewere then washed and placed into low carbonate culture medcontaining 10µg/ml cycloheximide for microinjection (all incubationsup to this step and the microinjection were done at 39.5°C). Cells wfurther chased (in normal cell culture medium containing 100µg/mlcycloheximide) as indicated in the figure legends. Preparation of lysates, immunoprecipitation of ts-O45-G, endoglycosidase H (enddigestion, gel electrophoresis, and fluorography were performeddescribed elsewhere (Pepperkok et al., 1993).
RESULTS
The small GTPases of the Arf family and Sar1p, when treawith non-hydrolysable GTP analogs (e.g. GTPγS) are shiftedto their activated forms. As a consequence, COPI or COcoat proteins are locked onto membranes, and membtraffic is arrested. We have microinjected GTPγS, recombinantSar1p or Arf1 cDNA into living cells and studied their effecon membrane transport from the ER to the cell surface usts-O45-G as a marker protein.
Table 1. Co-localization of ts-O45-G with differentmembrane markers at 15°C
Antigen GTPγS Co-localization
β-COP − 88±6%+ 11±13%
Sec13p − 72±14%+ 79±11%
ERGIC 53 − 92±4%+ 84±12%
hKDEL-R − 86±4%+ 78±11%
γ-Adaptin − 13±6%+ 16±9%
Colocalization of the distinct punctate ts-O45-G positive structures (typifor the IC; marked by arrows in Fig. 3C and Fig. 4C,E,G) and specificmembrane marker proteins were determined as described in Materials anMethods. More than 500 structures in a total of at least 25 cells wereanalyzed. Shown are the means ± standard mean error.
kage)
nte
ibedntfiths
was allaysTheere
izing
erere
mreillsium
ere
GTPγS blocks transport of ts-O45-G from transportcomplexes to the cell surface in vivoGTPγS was microinjected at different concentrations into celinfected with ts-O45 VSV and kept at the appropriattemperatures to keep ts-O45-G arrested in the ER, IC or TGAfter a brief incubation to let cells recover from the injectionprocedure, they were shifted to the permissive temperatureallow transport of the viral glycoprotein to resume and, one holater, arrival of ts-O45-G was quantified on the plasmmembrane by immunofluorescence and automated imaanalysis. Transport from the ER, ICs and TGN to the cell surfawas inhibited by GTPγS. As shown in Fig. 1A, the concentrationof GTPγS required to inhibit 50% of transport to the cell surfacwas significantly lower from the ER or ICs (50µM) than fromthe TGN (>200µM). Similar results were obtained whenGMPPNP was microinjected, but only little transport inhibitionwas observed with ATPγS (Fig. 1B). The effects of GTPγS ontransport of ts-O45-G from the ER or ICs to the Golgi complewere also quantified by cellular micro-biochemistry (Fig. 2)About 500 ts-O45 VSV infected cells were metabolically labelewith 35S-met/cys at 39.5°C, injected with differentconcentrations of GTPγS at 39.5°C, subsequently incubated a31°C for 2 hours and acquisition of endoH resistance of tcarbohydrate side chains of ts-O45-G determined to evalutransport through the medial Golgi cisternae (Fig. 2ASignificant inhibition of transport of ts-O45-G from the ER o
cal
d
Fig. 2.Biochemical analysis of transport from the ER or IC to theGolgi complex in cells injected with GTPγS. About 500 virus infectedcells with ts-O45-G accumulated in the ER or IC, respectively, werelabeled with 35S-met/cys and microinjected with differentconcentrations of GTPγS at 39.5 or 15°C. Immediately after injectionthe temperature was shifted to 31°C. Cells were lysed after 2 hours,ts-O45-G immunoprecipitated and analyzed for the amount which hadacquired endoH resistance. (A) A representative autoradiographanalyzing endoH acquisition, and (B) the correspondingquantification. The mean ± standard deviation of at least twoindependent experiments is shown. n.s., not shifted; n.i., not injected.
GTPγS (µM)

1880
lns).o
t
r
e
ftd
r
r
.
R. Pepperkok and others
Fig. 3.Co-localization of ts-O45-G and mammalian Sec13pduring ER to IC transport in GTPγS injected cells. Infectedcells with ts-O45-G accumulated in the ER were injected at39.5°C with 500µM GTPγS and 5 mg/ml coumarin BSA.Cells were incubated after injection for a further 30 minutesat 39.5°C (A,B), or for 3 hours at 15°C (C,D). They werethen fixed and double stained for the viral glycoprotein(A,C) and mammalian Sec13p (B,D). GTPγS injected cells(indicated by asterisks) were identified by the co-injectedcoumarin BSA (not shown). Arrows indicate co-localizingstructures. GTPγS does not interfere with transport of ts-O45-G from the ER to the IC, and the viral glycoprotein co-localises with Sec13p in the IC. Bar, 15µm.
IC through the Golgi complex occurred already with 50µMGTPγS microinjected (Fig. 2B), corroborating that GTPγSinhibits transport from the ER through the Golgi compleInterestingly, no difference could be seen whether transpbegan from the ER or the IC.
To characterize early exocytic membrane transport mprecisely, Vero cells were injected with 500µM GTPγS at39.5°C and incubated for an additional 30 minutes at 39.5°under these conditions ts-O45-G remained arrested in theof injected and neighboring control non-injected cells (Fi3A, 4A). Injection of GTPγS had no visible effect on thedistribution of ER resident proteins calnexin and PDI (nshown) or Sec13p, a component of the COPII complex (F3B). When the temperature was subsequently shifted to 1for 3 hours, ts-O45-G accumulated as in the non-injeccontrol cells in distinct patchy structures characteristic of tIC (Figs 3C, 4C,E,G) and co-localised with Sec13p (Fig. 3CTable 1), as well as KDEL-receptor and ERGIC 53 (not showTable 1), but not with β-COP (Fig. 4C-F), γ-adaptin (notshown; Table 1) or PDI and calnexin (not shown). Thuinjected GTPγS at concentrations up to 500µM (higherconcentrations of GTPγS lead to cell damage) does nointerfere with export of ts-O45-G from the ER. Similar resuwere obtained when GMPPNP was injected (not showMicroinjection of antibodies against β-COP (anti-EAGE),which inhibit ER to Golgi transport in vivo (Pepperkok et a1993), also failed to interfere with export from ER into TC(Fig. 4G,H). A notable feature of cells injected with antibodiehowever, was the complete overlap of ts-O45-G and β-COP(visualized by the microinjected antibodies; Fig. 4G,H, arroindicate co-localizing structures). In summary the results shthat GTPγS does not inhibit accumulation of ts-O45-G in thIC, a process which is independent of COPI.
For analyzing membrane transport in TCs to the Gocomplex, Vero cells with ts-O45-G accumulated at 15°C weinjected with 500µM GTPγS and subsequently incubated a
x.ort
ore
C; ERgs
otig.
5°Ctedhe,D;n;
s,
tltsn).
l.,ss,
wsowe
lgiret
31°C for 1 hour. While ts-O45-G was transported normally tothe plasma membrane (Fig. 5A,C,E,G) in non-injected controcells or cells injected with cascade blue BSA (not shown), iGTPγS-injected cells it was found arrested in extended tubuleas well as in a few distinct punctate structures (Fig. 5A,C,E,GAgain, these ts-O45-G-containing structures exhibited nsignificant overlap with the smaller β-COP buds or vesiclesthat accumulate under these conditions (Fig. 5A,B,A*,B*), bucoincide precisely with the distribution of ERGIC53 (Fig.5C,D) and KDEL receptor (not shown). Although there wasclearly some overlap in a juxtanuclear region of ts-O45-G withthe Golgi resident enzyme NAGT1 (arrows in Fig. 5E,F), it isnot clear (because of the limited resolution of lightmicroscopy), whether this fraction of viral glycoprotein hadtruly entered the Golgi complex or just accumulated onmembranes neighboring the Golgi complex. In fact, oubiochemical analyses indicate that GTPγS inhibits ts-O45-G’sacquisition of endoH resistance by only ~50% (Fig. 2).
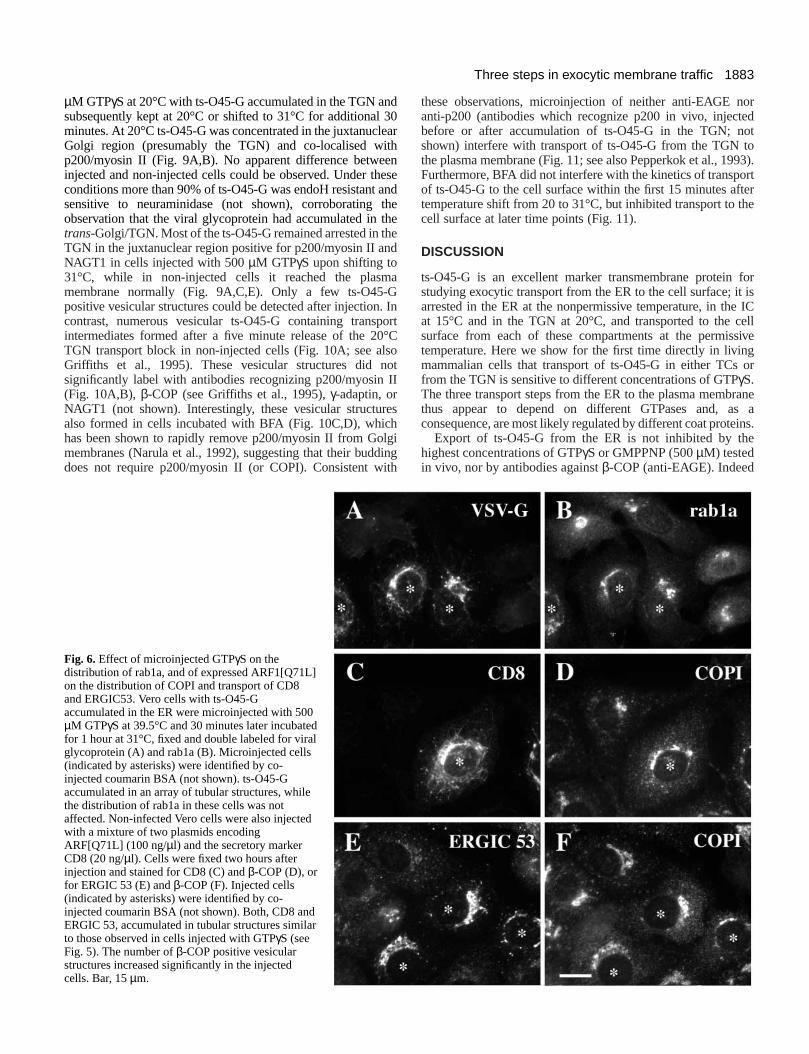
Injection of GTPγS had no effect on the distribution ofSec13p (Figs 3, 5G,H), and when cells with ts-O45-Gaccumulated in the ER or in the IC had been shifted to thpermissive temperature, little of the viral glycoproteinremained co-localised with this COPII subunit (Fig. 5G,H).Furthermore, we observed no effect on the distribution orab1a, a small GTPase involved in ER to Golgi transpor(Tisdale et al., 1992; Satoh et al., 1997) in cells microinjectewith 500 µM GTPγS (Fig. 6A,B). In contrast, microinjectedGTPγS induced a dramatic redistribution of β-COP, a subunitof COPI. Compared to non-injected cells, the juxtanuclealabeling of β-COP in the Golgi region was significantlydecreased and a large number of β-COP positive, apparentlyvesicular structures accumulated (Figs 4B, 5B). Similaeffects, albeit initially less dramatic, were also observed incells which were injected with cDNA of the Arf1[Q71L]mutant (Zhang et al., 1994) deficient in GTP hydrolysis (Fig6D,F).

1881Three steps in exocytic membrane traffic
s4;ed
utd
R
s-.atned
ad
Treatment of cells with brefeldin A (BFA), a drug whichinhibits binding of COPI to membranes (Donaldson et a1990), did not change this GTPγS induced altered distributionof β-COP (not shown; see also Donaldson et al., 199Therefore, the GTPγS induced β-COP positive structures aremost likely vesicles or buds which have accumulated as Arlocked in its activated state with bound nonhydrolyzabGTPγS (Tanigawa et al., 1993). When cells with injecteGTPγS and ts-O45-G accumulated at 15°C where shifted to permissive temperature, the viral glycoprotein moved intubular structures containing ERGIG53 and KDEL receptbut little or no β-COP (Table 1). Furthermore, under thesconditions, COPI positive vesicular structures contained litts-O45-G (Fig. 5A,B; Table 1). Similar results were obtainewhen cells were injected with ARF1[Q71L] cDNA instead owith 500µM GTPγS; in these cells transport of ts-O45-G (noshown), CD8 or ERGIC53 is also arrested in tubular arra(Fig. 6C-F). Thus we conclude that GTPγS affects most likelyArf1 and dramatically shifts the normal steady staequilibrium of cytosolic and membrane bound coatomtowards the membrane bound form and induces a bursbudding vesicles which do not appear to contain significaamounts of cargo.
l.,
1).
f isledtheto
or,etledftys
teert ofnt
A dominant negative SAR1a mutant inhibited in GTPhydrolysis interferes with export of ts-O45-G fromthe ER but not from TCs to the plasma membraneBudding of COPII coated vesicles from ER membranedepends on Sar1p GTPase activity (Barlowe et al., 199Bednarek et al., 1995), and fusion incompetent COPII coatvesicles form in the presence of GTPγS in vitro (Oka andNakano, 1994; Barlowe et al., 1994). Therefore, GTPγS shouldinhibit ER to Golgi transport by preventing COPII coatedvesicles from fusing with their target membranes. Transport oof the ER appeared not to be significantly altered or inhibitein the presence of either GTPγS or GMPPNP (Fig. 3, and datanot shown). To address more directly the role of Sar1p in Eto Golgi transport in vivo we purified recombinant protein froma dominant negative SAR1a mutant, inhibited in GTPhydrolysis (SAR1a [H79G]; Aridor et al., 1995). The purifiedprotein was microinjected at 39.5, 15 or 20°C into cells with tO45-G arrested in the ER, IC or TGN, respectivelyMicroinjected cells were subsequently incubated for 1 hour 31°C and effects on transport of the viral glycoproteivisualized by immunofluorescence as described for infectcells injected with GTPγS. While Sar1a[H79G]p completelyblocked transport from the ER to the plasma membrane, it h
Fig. 4.Co-localization of ts-O45-G and β-COP during ERto IC transport in GTPγS injected cells. Infected Vero cellswith ts-O45-G accumulated in the ER were injected at39.5°C with 500µM GTPγS and 5 mg/ml coumarin BSA(A-F) or anti-EAGE (2 mg/ml; G,H). Cells were incubatedfor a further 30 minutes at 39.5°C after the microinjectionand then either fixed and stained for ts-O45-G (A) and β-COP (B), or temperature shifted for 3 hours to 15°C beforefixation and staining for ts-O45-G (C,E,G) and β-COP(D,F) or microinjected anti-EAGE (H). GTPγS injectedcells (indicated by asterisks) were identified by the co-injected coumarin BSA (not shown). Arrows indicate co-localizing structures. Arrowheads in B indicate coatomernear the Golgi complex; microinjection of GTPγSdisperses these structures. Arrowheads in C and E indicatets-O45-G in structures devoid of β-COP. E and F representcorresponding two fold enlarged areas of the microinjectedcell shown in C and D. Neither microinjected GTPγS noranti-EAGE interfere with transport of ts-O45-G from theER to the IC. Bar, 15µm.

1882
tost
eofI5;r
R. Pepperkok and others
Fig. 5.Morphological analysis of transport of ts-O45-Gfrom the IC to the Golgi complex in cells injected withGTPγS. Infected cells with ts-O45-G accumulated inthe IC were injected with 500µM GTPγS and 5 mg/mlcoumarin BSA at 15°C. After injection, the temperaturewas shifted to 31°C for 1 hour and cells were then fixedand double stained for ts-O45-G (A,C,E,G) and β-COP(B), ERGIC53 (D), myc-tagged NAGT1 (F) or Sec13p(H). (A*) Enlargement of arrowed area in A; (B*)corresponding enlarged area in B. The Vero cells usedin E,F were stably transfected with myc-taggedNAGT1. Injected cells (indicated by asterisks) wereidentified by the co-injected coumarin BSA (notshown). Arrows in A and A* indicate ts-O45-G positivetubular structures negative for β-COP. Arrows and smallarrowheads in C and D indicate co-localizing tubularand punctate structures, respectively. The largerarrowheads in C indicate ts-O45-G at the plasmamembrane. (E-H) Arrows and arrowheads indicate co-localizing and non-co-localizing structures,respectively. ts-O45-G is arrested in punctate andtubular structures which overlap with ERGIC 53, andpartially with Sec13p or myc-tagged NAGT1, but notwith β-COP, in GTPγS injected cells. Bar, 15µm.
only a small effect on transport from the IC or TGN to thplasma membrane, respectively (Fig. 7). Microinjection Sar1a[H79G]p into cells kept at the nonpermissive temperatarrested the majority of ts-O45-G in the ER when cells weshifted to the permissive temperature for 1 hour, with onlyminor fraction of the viral glycoprotein congregating intpatches with which Sec13p co-localised (Fig. 8A,BInterestingly, when compared to neighboring control cells, number of dispersed cytoplasmic Sec13p positive structuwas significantly decreased in the injected cells, while COPII signal accumulated near the nucleus (see also Bonaal., 1989). This observation may indicate that the COPII coatransport intermediates containing Sar1a[H79G]p have motowards the Golgi region without uncoating and preventing
eofure,re ao).
theres
thetti ettedvedthe
generation of new ER derived COPII coated vesicles due depletion of the cytosolic pool of COPII. Overall these resultsuggest that COPII is required only for exit from the ER, bunot for any subsequent transport steps.
Inhibition of transport of ts-O45-G from the TGN tothe cell surface requires higher concentrations ofGTPγS than from TCs to the Golgi complexTo characterize the inhibition of transport of ts-O45-G from thTGN to the plasma membrane more precisely, the effects microinjected GTPγS and antibodies against p200, a myosin Iprotein found on buds on the TGN (Narula and Stow, 199Ikonen et al., 1996; Müsch et al., 1997), were followed in furthedetail. Cells infected with ts-O45 VSV were injected with 500
1883Three steps in exocytic membrane traffic
ordt
o93).rtere
foris ICellivegor
ane a
ins.
µM GTPγS at 20°C with ts-O45-G accumulated in the TGN asubsequently kept at 20°C or shifted to 31°C for additional minutes. At 20°C ts-O45-G was concentrated in the juxtanuclGolgi region (presumably the TGN) and co-localised wip200/myosin II (Fig. 9A,B). No apparent difference betweeinjected and non-injected cells could be observed. Under thconditions more than 90% of ts-O45-G was endoH resistant sensitive to neuraminidase (not shown), corroborating observation that the viral glycoprotein had accumulated in trans-Golgi/TGN. Most of the ts-O45-G remained arrested in tTGN in the juxtanuclear region positive for p200/myosin II anNAGT1 in cells injected with 500µM GTPγS upon shifting to31°C, while in non-injected cells it reached the plasmmembrane normally (Fig. 9A,C,E). Only a few ts-O45-positive vesicular structures could be detected after injectioncontrast, numerous vesicular ts-O45-G containing transpintermediates formed after a five minute release of the 20TGN transport block in non-injected cells (Fig. 10A; see alGriffiths et al., 1995). These vesicular structures did nsignificantly label with antibodies recognizing p200/myosin (Fig. 10A,B), β-COP (see Griffiths et al., 1995), γ-adaptin, orNAGT1 (not shown). Interestingly, these vesicular structuralso formed in cells incubated with BFA (Fig. 10C,D), whichas been shown to rapidly remove p200/myosin II from Gomembranes (Narula et al., 1992), suggesting that their budddoes not require p200/myosin II (or COPI). Consistent w
Fig. 6.Effect of microinjected GTPγS on thedistribution of rab1a, and of expressed ARF1[Q71L]on the distribution of COPI and transport of CD8and ERGIC53. Vero cells with ts-O45-Gaccumulated in the ER were microinjected with 500µM GTPγS at 39.5°C and 30 minutes later incubatedfor 1 hour at 31°C, fixed and double labeled for viralglycoprotein (A) and rab1a (B). Microinjected cells(indicated by asterisks) were identified by co-injected coumarin BSA (not shown). ts-O45-Gaccumulated in an array of tubular structures, whilethe distribution of rab1a in these cells was notaffected. Non-infected Vero cells were also injectedwith a mixture of two plasmids encodingARF[Q71L] (100 ng/µl) and the secretory markerCD8 (20 ng/µl). Cells were fixed two hours afterinjection and stained for CD8 (C) and β-COP (D), orfor ERGIC 53 (E) and β-COP (F). Injected cells(indicated by asterisks) were identified by co-injected coumarin BSA (not shown). Both, CD8 andERGIC 53, accumulated in tubular structures similarto those observed in cells injected with GTPγS (seeFig. 5). The number of β-COP positive vesicularstructures increased significantly in the injectedcells. Bar, 15µm.
nd30earthneseandthethehed
aG. Inort°C
sootII
eshlgiing
ith
these observations, microinjection of neither anti-EAGE nanti-p200 (antibodies which recognize p200 in vivo, injectebefore or after accumulation of ts-O45-G in the TGN; noshown) interfere with transport of ts-O45-G from the TGN tthe plasma membrane (Fig. 11; see also Pepperkok et al., 19Furthermore, BFA did not interfere with the kinetics of transpoof ts-O45-G to the cell surface within the first 15 minutes afttemperature shift from 20 to 31°C, but inhibited transport to thcell surface at later time points (Fig. 11).
DISCUSSION
ts-O45-G is an excellent marker transmembrane protein studying exocytic transport from the ER to the cell surface; it arrested in the ER at the nonpermissive temperature, in theat 15°C and in the TGN at 20°C, and transported to the csurface from each of these compartments at the permisstemperature. Here we show for the first time directly in livinmammalian cells that transport of ts-O45-G in either TCs from the TGN is sensitive to different concentrations of GTPγS.The three transport steps from the ER to the plasma membrthus appear to depend on different GTPases and, asconsequence, are most likely regulated by different coat prote
Export of ts-O45-G from the ER is not inhibited by thehighest concentrations of GTPγS or GMPPNP (500 µM) testedin vivo, nor by antibodies against β-COP (anti-EAGE). Indeed

1884
-disi
lgiileese
oftsER).
o),yalfalith5;s a
n,ddnre,-gPIlgin,
ficeoflgiforinis
l
edor
R. Pepperkok and others
0
25
50
75
100
BSASar1a (H79G)p
ts-
O45
-G a
t th
e ce
ll su
rfac
e (%
)
39.5->31 °C 15->31 °C 20->31 °C
Fig. 7.Transport of ts-O45-G to the plasma membrane in Vero celinjected with Sar1a(H79G)p. Coumarin BSA (5 mg/ml) was injectewith or without purified Sar1a(H79G)p (1 mg/ml) at 39.5°C, 15°C 20°C with ts-O45-G arrested in the ER, IC or TGN, respectively.After injection, the temperature was shifted to 31°C for 1 hour andthe amount of ts-O45-G on the cell surface quantified byimmunofluorescence and image analysis. The data are normalizethe value obtained with neighboring non-injected cells (100%).Sar1a(H79G)p blocks transport from the ER, but not from the IC oTGN to the plasma membrane. The mean ± standard deviation foindependent experiments with at least 50 injected cells is shown.
it has been shown that GTPγS is an at least 50-fold less goodsubstrate for Sar1p than it is for Arf1 (Aridor et al., 1995); thuwhile export of viral glycoprotein from the ER is blocked icells microinjected with GTP restricted Sar1a[H79G]pmicroinjected GTPγS had no effect on this first exocyticmembrane transport step. Compelling in vitro data (Oka et 1991; Oka and Nakano, 1994; Barlowe et al., 1994; Bednaet al., 1995) and experiments in vivo in mammalian cells wSar1p and mutants of that GTPase (Kuge et al., 1994; Ariet al., 1995) strongly suggest that COPII is essential for expof membrane proteins from the ER in yeast and mammalcells. Our results are consistent with these data and conthat in vivo GTPγS does not inhibit the formation of COPIIcoated vesicles transporting ts-O45-G out of the ER towaVTCs, where they accumulate but are probably inhibited fuse because Sar1p, deficient in GTP hydrolysis, cannot releCOPII (Barlowe et al., 1994; Oka and Nakano, 1994).
Fig. 8.Morphological analysis of ts-O45-G transport inSar1a(H79G)p injected cells. Infected cells with ts-O45-Gaccumulated in the ER were injected with Sar1a(H79G)p(1 mg/ml) at 39.5°C and subsequently shifted for 1 hour to31°C, fixed and stained for ts-O45-G (A) and Sec13p (B).Injected cells (marked by asterisks) were identified by co-injected coumarin BSA (not shown). Arrows in A and Bindicate overlapping structures. Arrowheads in A indicatets-O45-G in the nuclear envelope. Sar1a(H79G)p inhibitsexport of ts-O45-G from the ER and blocks transport tothe Golgi complex. Bar, 10µm.
s,n,
al.,rekithdorortianfirm
rdstoase
While export from the ER was not affected by either GTPanalogues or anti-EAGE, both reagents efficiently inhibitetransport in TCs to the Golgi complex, suggesting that COPI involved in membrane traffic between VTCs and the Golgcomplex but not in export from the ER. MicroinjectedSar1a[H79G]p on the other hand had no effect on VTC to Gotransport. The simplest explanation for these results is that, whCOPII regulates ER export, COPI is involved in regulating thsubsequent delivery of cargo in TCs to the Golgi complex. Theresults also support previous in vitro results with digitoninpermeabilized cells suggesting that COPII directs export cargo (ts-O45-G) from the ER and that COPII and COPI coaare sequentially coupled to regulate transport between the and Golgi complex (Aridor et al., 1995; Bannykh et al., 1996Consistent with that conclusion we have recently shown that β-COP co-localizes predominantly with TCs carrying ts-O45-G tthe Golgi complex (Griffiths et al., 1995; Scales et al., 1997providing further evidence for an important role of COPI in earlanterograde membrane traffic. Interestingly, however, about hof the coatomer positive vesicular structures did not contain vircargo, suggesting that COPI may also be associated wrecycling or retrograde transport vehicles (Griffiths et al., 199Kreis et al., 1995). Genetic evidence indeed strongly suggestrole for COPI in retrieval of material from the Golgi complex tothe ER (Letourneur et al., 1994; Cosson et al., 1996). In additiomicroinjected anti-EAGE inhibit transport between the ER anGolgi complex (Pepperkok et al., 1993) and block BFA inducerelocation of Golgi enzymes to the ER by inhibiting dissociatioof coatomer from membranes (Scheel et al., 1997). FurthermoGolgi derived COPI coated vesicles carry predominantly KDELreceptor (Sönnichsen et al., 1996), involved in retrievinmissorted ER proteins (Lewis and Pelham, 1992). While COappears to mediate transport in both directions through the Gocomplex (Orci et al., 1997; see also Schekman and Mellma1997), we propose that COPI regulates membrane trafbetween the ER and Golgi complex in a different way. Thsimplest model explaining the available data on the function COPI suggests that this coat moves with TCs to the Gocomplex (Scales et al., 1997) and on its way is responsible the retrieving, for example, of SNAREs and factors required COPII dependent vesicle budding to the ER. Inhibition of thfunction by microinjection of specific antibodies or GTPγSinterferes with retrieval of these factors and inhibits normadelivery of cargo to the Golgi complex.
Injection of GTPγS had also a profound effect on thedistribution of coatomer. β-COP no longer accumulated in theperinuclear region of these cells, but instead was scatterthroughout the cytoplasm in numerous vesicular structures
lsd
or
d to
rr 2

1885Three steps in exocytic membrane traffic
it in.
s
emrs,n
re, ityds
are;th;
n
allyl
Fig. 9.Morphological analysis of transport of ts-O45-G from the TGN to the plasma membrane inGTPγS injected cells. Infected cells with ts-O45-Gaccumulated in the TGN were injected with 500µMGTPγS and 5 mg/ml coumarin BSA at 20°C. Afterinjection, cells were incubated at 20°C (A,B) or31°C (C-F) for 30 minutes, before fixation anddouble staining for ts-O45-G (A,C,E), p200 (B,D),or myc-tagged NAGT1 (F). Injected cells (indicatedby asterisks) were identified by co-injectedcoumarin BSA (not shown). The Vero cells used inE,F were stably transfected with myc-taggedNAGT1. GTPγS inhibits ts-O45-G in the TGN. Bar,15µm.
buds. While in these injected cells, ts-O45-G colocalized asthe control cells with the proteins normally accumulating in tIC at 15°C (e.g. ERGIC53, Sec13p, KDEL receptor), little β-COP could be found colocalizing with any of these proteins.addition, shifting cells injected with GTPγS from 15°C to thepermissive temperature lead to accumulation of ts-O45-Gtubular structures (similar to cells that had been injected wanti-EAGE; see Pepperkok et al., 1993 and Scheel et al., 19but little overlap between β-COP and cargo could be seen. Wconclude from these results that while injected anti-EAGenhances binding of COPI to membranes and inhibits bformation (Scheel et al., 1997), GTPγS in vivo also enhancesthe binding of coatomer onto membranes (as has bpreviously shown in vitro by Malhotra et al., 1989 anDonaldson et al., 1991) but subsequently induces the formaof buds or vesicles. To our surprise, however, most of thstructures appeared devoid of cargo, suggesting that Ghydrolysis plays an important role in the coordinated coupliof sorting and budding of transport vesicles. While the cructarget(s) in this process remains to be definitively establishArf is most likely one, since similar results where obtained wthe GTP-restricted ARF[Q71L] mutant. Furthermore, rabappeared not to be significantly affected by the injection GTPγS or expression of ARF[Q71L]. We are currently furtheinvestigating this observation, since it may lead to importa
inhe
In
inith97),eEud
eendtioneseTP
ngialed,ith1ofrnt
new insights into cargo sorting mechanisms. In addition, suggests that caution should be exercised when interpretingvitro experiments performed in the presence of GTP analogs
Inhibition of transport of ts-O45-G from the TGN to theplasma membrane required microinjection of at least five timehigher GTPγS concentrations than those inhibiting transport inTCs to the Golgi complex. This result suggests that thGTPase(s) involved in this transport step must be distinct frothose regulating early exocytic membrane traffic (see also Milleand Moore, 1991; Xu and Shields, 1993; Pimplikar and Simon1993; Barr and Huttner, 1996; Chen and Shields, 1996; Simoet al., 1996). Since all transport vesicles identified so far acoated with specific proteins and regulated by small GTPasesis likely that TGN to cell surface transport is also mediated bspecific coated vesicles. Indeed, TGN associated coated bucontaining ts-O45-G have already been described and they negative for clathrin and COPI (Griffiths et al., 1985, 1995Duden et al., 1991). A candidate for a protein associated wicoats on TGN buds is p200/myosin II (Narula and Stow, 1995Ikonen et al., 1997); its binding to Golgi membranes is BFAsensitive p200 (Narula et al., 1992). This protein has beeimplicated in the formation of transport containing VSV-Gvesicles from the TGN (Müsch et al., 1997). Some experimentdata indicate, however, that p200/myosin II may not be directinvolved in the transport of ts-O45-G to the cell surface. Cytoso

1886
-eiat
R. Pepperkok and others
Fig. 10.Co-visualisation of ts-O45-G and p200during transport of the viral glycoprotein from theTGN to the plasma membrane. ts-O45-G wasaccumulated in the TGN at 20°C before thetemperature was shifted for 5 minutes to 31°C in theabsence (A,B) or presence of brefeldin A (C,D).Cells were then fixed and double-stained for ts-O45-G (A,C) and p200 (B,D). Most ts-O45-Gpositive vesicular transport intermediates that hadformed under these conditions (arrows) appear notto contain p200. Arrowheads in A and B indicateoverlapping structures. Bar, 15µm.
immunodepleted of p200 still supports basolateral transporthis viral glycoprotein (Ikonen et al., 1996). Furthermore, TGderived carrier vesicles containing ts-O45-G appear not to lawith antibodies against p200 and microinjected anti-p200 has
eatgr
us
d,ot
itinot
tic.ic
,ierlly
lestotheed
504030201000
25
50
75
100
time (min)
ts-O
45
-G a
t th
e c
ell
surf
ace
(%
)
BSA
anti-p200
BFA
GTP Sγ
Fig. 11.Time-course of transport of ts-O45-G from the TGN to theplasma membrane in the presence of BFA, or microinjected anti-p200 or GTPγS. Cells were injected at 20°C with GTPγS (500µM)and 5 mg/ml coumarin BSA (GTPγS), anti-p200 (anti-p200), orcoumarin BSA alone (BSA) after accumulation of ts-O45-G in theTGN at 20°C. Temperature was then shifted to 31°C for differenttime-periods and ts-O45-G quantified at the plasma membrane byimmunofluorescence and quantitative image analysis. In parallelexperiments non-injected cells were shifted from 20 to 31°C in thepresence of brefeldin A (BFA). Each data point represents the meof two experiments with at least 50 cells analyzed. The mean ±standard deviation of the mean is shown.
t ofNbel no
inhibitory effect on TGN-plasma membrane transport of ts-O45G. Moreover, vesicles containing ts-O45-G can still form in thpresence of BFA (which dissociates p200/myosin II from Golgmembranes), when cells with the glycoprotein accumulated 20°C in the TGN are shifted briefly to the permissivetemperature; about 50% of the viral glycoprotein will reach thcell surface in the presence of this drug. While we assume thp200/myosin II may not be essential for the final vesicle buddinstep from the TGN, it is possible that a BFA sensitive coat oscaffold (including p200/myosin II) may be involved in primingthe budding reaction during the 20°C incubation, and that thtransport to the cell surface is no longer fully BFA andp200/myosin II sensitive. While BFA was previously shown toinhibit constitutive secretion of heparan sulfate proteoglycan ants-O45-G from the TGN in vitro (Rosa et al., 1992; Miller et al.1992), it has recently also been reported that BFA does ninhibit transport of prohormone in GH3 cells from the TGN tothe cell surface in vitro (Xu and Shields, 1993). In contrast, has been shown by the same group that Arf1 is involved prohormone transport (Chen and Shields, 1996). We cannconclude whether or not an Arf-like protein is involved in TGNto cell surface membrane traffic, nor do we know the coaproteins that mediate this last step in exocytic membrane traffBut the observation that TGN to cell surface membrane traffof ts-O45-G is sensitive to GTPγS in vivo should facilitate theidentification of the relevant new coat proteins.
We thank Drs Bill Balch for Sar1p DNAs; Richard Kahn for theARF[Q71L] cDNA; David Shima for the plasmid encoding CD8;Brian Storrie for NAGT1-myc transformed Vero cells; George BloomStephen Fuller, Hans-Peter Hauri, Ari Helenius, Wanjin Hong, ScottRobinson, David Shima, Kai Simons and Bor Luen Tang foantibodies. The technical assistance of Jeanne Kroese is gratefuacknowledged. Special thanks go to Andy Whitney and Suzie Scafor helpful discussions. This research was supported by grants T.E.K. from the Fonds National Suisse, the Canton de Genève and International Human Frontier Science Program. R.P. was support
an

1887Three steps in exocytic membrane traffic
ct.
s.
lex
gi
ry
s.
fof
ast
ed
inll
ot
ic
y
g
rinal
by an EMBO long term postdoctoral fellowship and M.L. by Traveling Research Fellowship from the Welcome Trust.
REFERENCES
Aniento, F., Gu, F., Parton, R. G. and Gruenberg, J. (1996). An endosomalbeta COP is involved in the pH-dependent formation of transport vesicdestined for late endosomes. J. Cell Biol. 133, 29-41.
Aridor, M., Bannykh, S. I., Rowe, T. and Balch, W. E. (1995). Sequentialcoupling between COPII and COPI vesicle coats in endoplasmic reticuto Golgi transport. J. Cell Biol. 131, 875-893.
Bannykh, S. I., Rowe, T. and Balch, W. E. (1996). The organization ofendoplasmic reticulum export complexes. J. Cell Biol. 135, 19-35.
Bannykh, S. I. and Balch, W. E. (1997). Membrane dynamics at theendoplasmic reticulum-Golgi interface. J. Cell Biol. 138, 1-4.
Barlowe, C., d’Enfert, C. and Schekman, R. (1993). Purification andcharacterization of SAR1p, a small GTP-binding protein required ftransport vesicle formation from the endoplasmic reticulum. J. Biol. Chem.268, 873-879.
Barlowe, C., Orci, L., Yeung, T., Hosobuchi, M., Hamamoto, S., Salama,N., Rexach, M. F., Ravazzola, M., Amherdt, M. and Schekman, R.(1994). COPII: A membrane coat formed by sec proteins that drive vesbudding from the endoplasmic reticulum. Cell 77, 895-907.
Barr, F. A. and Huttner, W. B. (1996). A role for ADP-ribosylation factor 1,but not COP I, in secretory vesicle biogenesis from the trans-Golgi netwoFEBS Lett. 384, 65-70.
Beckers, C. J. and Balch, W. E. (1989). Calcium and GTP: essentiacomponents in vesicular trafficking between the endoplasmic reticulum Golgi apparatus. J. Cell Biol. 108, 1245-1256.
Bednarek, S. Y., Ravazzola, M., Hosobuchi, M., Amherdt, M., Perrelet, A.Schekman, R. and Orci, L. (1995). COPI- and COPII-coated vesicles budirectly from the endoplasmic reticulum in yeast. Cell 83, 1183-1196.
Bergmann, J. E., Tokuyasu, K. T. and Singer, S. J. (1981). Passage of anintegral membrane protein, the vesicular stomatitis virus glycoprotethrough the Golgi apparatus en route to the plasma membrane. Proc. Nat.Acad. Sci. USA78, 1746-1750.
Bloom, G. S. and Brashear, T. A. (1989). A novel 58-kDa protein associatewith the Golgi apparatus and microtubules. J. Biol. Chem. 264, 16083-16092.
Bonatti, S., Migliaccio, G. and Simons, K. (1989). Palmitosylation of viralmembrane glycoprotein takes place after exit from the endoplasretiulum. J. Biol. Chem. 264, 12590-12595.
Campbell, J. L. and Schekman, R. (1997). Selective packaging of cargomolecules into endoplasmic reticulum-derived COPII vesicles. Proc. Nat.Acad. Sci. USA94, 837-842.
Chen, Y. G. and Shields, D. (1996). ADP-ribosylation factor-1 stimulatesformation of nascent secretory vesicles from the trans-Golgi networkendocrine cells. J. Biol. Chem. 271, 5297-5300.
Cosson, P. and Letourneur, F. (1994). Coatomer interaction with di-lysineendoplasmic reticulum retention motifs. Science263, 1629-1631.
Cosson, P., Démollière, C., Hennecke, S., Duden, R. and Letourneur, F(1996). Delta- and zeta-COP, two coatomer subunits homologous to clathassociated proteins, are involved in ER retrieval. EMBO J. 15, 1792-1798.
Cowles, C. R., Odorizzi, G., Payne, G. S. and Emr, S. D. (1997). The AP-3 adaptor complex is essential for cargo-selective transport to the yvacuole. Cell 91, 109-118.
Dascher, C. and Balch, W. E. (1994). Dominant inhibitory mutants of ARF1block endoplasmic reticulum to Golgi transport and trigger disassemblythe Golgi apparatus. J. Biol. Chem. 269, 1437-1448.
Dell’Angelica, E. D., Ohno, H., Ooi, C. E., Rabinovich, E., Roche, W. K.and Bonifacino, J. S. (1997). AP-3: an adaptor like protein complex withubiquitous expression. EMBO J. 16, 917-928.
Donaldson, J. G., Lippincott-Schwartz, J., Bloom, G. S., Kreis, T. E. andKlausner, R. D. (1990). Dissociation of a 110-kD peripheral membranprotein from the Golgi apparatus is an early event in brefeldin A actionJ.Cell Biol. 111, 2295-2306.
Donaldson, J. G., Lippincott-Schwartz, J., Bloom, G. S., Kreis, T. E. andKlausner, R. D. (1991). Binding of ARF and β-COP to Golgi membranes:possible regulation by trimeric G protein. Science254, 1197-1199.
Duden, R., Griffiths, G., Frank, R., Argos, P. and Kreis, T. E. (1991). β-COP, a 110kd protein associated with non-clathrin-coated vesicles andGolgi complex, shows homology to β-adaptin. Cell 64, 649-665
Evan, G. I., Lewis, G. K., Ramsay, G. and Bishop, J. M. (1985). Isolation
a
les
lum
or
icle
rk.
land
d
in,
s
mic
of
.rin-
east
of
e.
the
of monoclonal antibodies specific for human c-myc proto-oncogene produMol. Cell. Biol. 5, 3610-3616.
Fiedler, K., Veit, M., Stamnes, M. A. and Rothman, J. E. (1996). Bimodalinteraction of coatomer with the p24 family of putative cargo receptorScience273, 1396-1399.
Griffiths, G., Pfeiffer, S., Simons, K. and Matlin, K. (1985). Exit of newlysynthesized membrane proteins from the trans cisterna of the Golgi compto the plasma membrane. J. Cell Biol. 101, 949-964.
Griffiths, G., Ericsson, M., Krijnse-Locker, J., Nilsson, T., Goud, B.,Soling, H. D., Tang, B. L. , Wong, S. H., Hong, W., et al. (1994).Localization of the Lys, Asp, Glu, Leu tetrepeptide receptor to the Golcomplex and the intermediate compartment in mammalian cells. J. Cell Biol.127, 1557-1574.
Griffiths, G., Pepperkok, R., Locker, J. K. and Kreis, T. E. (1995).Immunocytochemical localization of beta-COP to the ER-Golgi boundaand the TGN. J. Cell Sci. 108, 2839-2856.
Hammond, C. and Helenius, A. (1994). Quality control in the secretorypathway: retention of a misfolded viral membrane glycoprotein involvecycling between the ER, intermediate compartment and Golgi apparatusJ.Cell Biol. 126, 41-52.
Harter, C., Pavel, J., Coccia, F., Draken, E., Wegehingel, S., Tschochner,H. and Wieland, F. (1996). Nonclathrin coat protein gamma, a subunit ocoatomer, binds to the cytoplasmic dilysine motif of membrane proteins the early secretory pathway. Proc. Nat. Acad. Sci. USA93, 1902-1906.
Hosobuchi, M., Kreis, T. E. and Schekman, R. (1992). Sec21 is a generequired for ER to Golgi transport that encodes a subunit of a yecoatomer. Nature360, 603-605.
Ikonen, E., Parton, R. G., Lafont, F. and Simons, K. (1996). Analysis of therole of p200-containing vesicles in post-Golgi traffic. Mol. Biol. Cell7, 961-974.
Ikonen, E., de Almeid, J. B., Fath, K. F., Burgess, D. R., Ashman, K.,Simons, K. and Stow, J. L. (1997). Myosin II is associated with Golgimembranes: identification of p200 as nonmuscle myosin II on Golgi-derivvesicles. J. Cell Sci. 110, 2155-2164.
Kreis, T. E. (1986). Microinjected antibodies against the cytoplasmic domaof vesicular stomatitis virus glycoprotein block its transport to the cesurface. EMBO J. 5, 931-941.
Kreis, T. E. and Lodish, H. F. (1986). Oligomerization is essential fortransport of vesicular stomatitis viral glycoprotein to the cell surface. Cell46, 929-937.
Kreis, T. E., Lowe, M. and Pepperkok, R. (1995). COPs regulatingmembrane traffic. Annu. Rev. Cell Dev. Biol. 11, 677-706.
Kuge, O., Dascher, C., Orci, L., Rowe, T., Amherdt, M., Plutner, H.,Ravazzola, M., Tanigawa, G., Rothman, J. E. and Balch, W. E. (1994).Sar1 promotes vesicle budding from the endoplasmic reticulum but nGolgi compartments. J. Cell Biol. 125, 51-65.
Letourneur, F., Gaynor, E. C., Hennecke, S., Demolliere, C., Duden, R.,Emr, S. D., Riezman, H. and Cosson, P. (1994). Coatomer is essential forretrieval of dilysine-tagged proteins to the endoplasmic reticulum. Cell 79,1199-1207.
Lewis, M. J. and Pelham, H. R. (1992). Ligand-induced redistribution of ahuman KDEL receptor from the Golgi complex to the endoplasmreticulum. Cell 68, 353-364.
Leyte, A., Barr, F. A., Kehlenbach, R. H. and Huttner, W. B. (1992).Multiple trimeric G-proteins on the trans-Golgi network exert stimulatorand inhibitory effects on secretory vesicle formation. EMBO J. 11, ??-??.
Lowe, M. and Kreis, T. E. (1995). In vitro assembly and disassembly ofcoatomer. J. Biol. Chem. 270, 31364-31371.
Malhotra, V., Serafini, T., Orci, L., Shepherd, J. C. and Rothman, J. E.(1989). Purification of a novel class of coated vesicles mediatinbiosynthetic protein transport through the Golgi stack. Cell 58, 329-336.
Matlin, K. S. and Simons, K. (1983). Reduced temperature prevents transfeof a membrane glycoprotein to the cell surface but does not prevent termglycosylation. Cell 34, 233-243.
Melançon, P., Glick, B. S., Malhotra, V., Weidmann, P. J., Serafini, T.,Gleason, M. L., Orci, L. and Rothman, J. E. (1987). Involvement of GTP-binding ‘G’ proteins in transport through the Golgi stack. Cell 51, 1053-1062.
Miller, S. G., Carnell, L. and Moore, H. H. (1992). Post-Golgi membranetraffic: brefeldin A inhibits export from distal Golgi compartments to thecell surface but not recycling. J. Cell Biol. 118, 267-283.
Miller, S. G. and Moore, H. P. (1991). Reconstitution of constitutive secretionusing semi-intact cells: regulation by GTP but not calcium. J. Cell Biol. 112,39-54.
Müsch, A., Cohen, D. and Rodriguez-Boulan, E. (1997). Myosin II is

1888
t
II
-
ort
s
in
a
inhe
f.
a
f
R. Pepperkok and others
involved in the production of constitutive transport vesicles from the TGJ. Cell Biol. 138, 291-306.
Narula, N., McMorrow, I., Plopper, G., Doherty, J., Matlin, K. S., Burke,B. and Stow, J. L. (1992). Identification of a 200-kD, brefeldin-sensitiveprotein on Golgi membranes. J. Cell Biol. 117, 27-38.
Narula, N. and Stow, J. L. (1995). Distinct coated vesicles labeled for p20bud from trans-Golgi network membranes. Proc. Nat. Acad. Sci. USA92,2874-2878.
Newman, L. S., McKeever, M. O., Okano, H. J. and Darnell, R. B. (1995).b-NAP, a cebrellar degeneration antigen, is a neuron-specific vesicle protein. Cell 82, 773-783.
Novick, P. and Brennwald, P. (1993). Friends and family: the role of the RabGTPases in vesicular traffic. Cell 75, 597-601.
Nuoffer, C. and Balch, W. E. (1994). GTPases: multifunctional molecularswitches regulating vesicular traffic. Annu. Rev. Biochem. 63, 949-990.
Oka, T. and Nakano, A. (1994). Inhibition of GTP hydrolysis by Sar1p causeaccumulation of vesicles that are a functional intermediate of the ERGolgi transport in yeast. J. Cell Biol. 124, 425-434.
Oka, T., Nishikawa, S. and Nakano, A. (1991). Reconstitution of GTP-binding Sar1 protein function in ER to Golgi transport. J. Cell Biol. 114,671-679.
Orci, L., Stamnes, M., Ravazzola, M., Amherdt, M., Perrelet, A., Söllner,T. H. and Rothman, J. E. (1997). Bidirectional transport by distinctpopulations of COPI-coated vesicles. Cell 90, 335-349.
Paccaud, J. P., Reith, W., Carpentier, J.-L., Ravazzola, M., Amherdt, M.,Schekman, R. and Orci, L. (1996). Cloning and characterization ofmammalian homologues of the COPII component Sec23. Mol. Biol. Cell7,1535-1546.
Palade, G. (1975). Intracellular aspects of the process of protein syntheScience 187, 347-358.
Palmer, D. J., Helms, J. B., Beckers, C. J. L. Orci and J. E. Rothman.(1993). Binding of coatomer to Golgi membranes requires ADP-ribosylatifactor. J. Biol. Chem. 268, 12083-12089.
Pearse, B. M. F. and Robinson, M. S. (1990). Clathrin, adaptors and sortingAnnu. Rev. Cell Biol. 1, 151-171.
Pepperkok, R., Scheel, J., Horstmann, H., Hauri, H. P., Griffiths, G. andKreis, T. E. (1993). Beta-COP is essential for biosynthetic membrane transpfrom the endoplasmic reticulum to the Golgi complex in vivo. Cell 74, 71-82.
Peter, F., Plutner, H., Zhu, H., Kreis, T. E. and Balch, W. E. (1993). Beta-COP is essential for transport of protein from the endoplasmic reticulumthe Golgi in vitro. J. Cell Biol. 122, 1155-1167.
Pierre, P., Pepperkok, R. and Kreis, T. E. (1994). Molecular characterizationof two functional domains of CLIP-170 in vivo. J. Cell Sci. 107, 1909-1920.
Pevsner, J., Volknandt, W., Wong, B. R. and Scheller, R. H. (1994). Tworat homologs of clathrin-associated adaptor proteins. Gene146, 279-283.
Pimplikar, S. W. and Simons, K. (1993). Regulation of apical transport inepithelial cells by a Gs class of heterotrimeric G protein. Nature362, 456-458.
Ponnambalam, S., Rabouille, C., Luzio, J. P., Nilsson, T. and Warren, G.(1994). The TGN38 glycoprotein contains two non-overlapping signals tmediate localisation to the trans-Golgi network. J. Cell Biol. 125, 253-268.
Presley, J. F., Cole, N. B., Schroer, T. A., Hirschberg, K., Zaal, K. J. andLippincott-Schwartz, J. (1997). ER-to-Golgi transport visualized in livingcells. Nature389, 81-85.
Rexach, M. F., Latterich, M. and Schekman, R. W. (1994). Characteristicsof endoplasmic reticulum-derived transport vesicles. J. Cell Biol. 126, 1133-1148.
Robinson, M. S. (1994). The role of clathrin, adaptors and dynamin iendocytosis. Curr. Opin. Cell Biol. 6, 538-544.
Robinson, M. S. and Kreis, T. E. (1992). Recruitment of coat proteins ontoGolgi membranes in intact and permeabilized cells: effects of brefeldinand G protein activators. Cell 69, 129-138.
Rosa, P., Barr, F. A., Stinchcombe, J. C., Binacchi, C. and Huttner, W. B.(1992). Brefeldin A inhibits the formation of constitutive secretory vesicland immature secretory granules from the trans-Golgi network. Eur. J. CellBiol. 59, 265-274.
Rothman, J. E. and Orci, L. (1992). Molecular dissection of the secretorypathway. Nature355, 409-415.
Saraste, J. and Kuismanen, E. (1984). Pre- and post-Golgi vacuoles operatin the transport of Semliki Forest virus membrane glycoproteins to the csurface. Cell 38, 535-549.
Saraste, J., Lahtinen, U. and Goud, B. (1995). Localization of the smallGTP-binding protein rab1p to early compartments of the secretory pathwJ. Cell Sci. 108, 1541-1552.
Satoh, A. K., Tokunaga, F., Kawamura, S. and Ozaki, K. (1997). In situ
N.
0
coat
s to
sis.
on
.
ort
to
hat
n
A
es
eell
ay.
inhibition of vesicle transport and protein processing in the dominannegative Rab1 mutant of Drosophila. J. Cell Sci. 110, 2943-2953.
Scales, S. J., Pepperkok, R. and Kreis T. E. (1997). Visualization of ER-to-Golgi transport in living cells reveals a sequential mode of action for COPand COPI. Cell 90, 1137-1148.
Scheel, J., Pepperkok, R., Lowe, M., Griffiths, G. and Kreis, T. (1997).Dissociation of coatomer from membranes is required for brefeldin Ainduced transfer of Golgi enzymes to the endoplasmic reticulum. J. CellBiol. 137, 319-333.
Schekman, R. and Orci, L. (1996). Coat proteins and vesicle budding.Science271, 1526-1533.
Schekman, R. and Mellman, I.(1997). Does COPI go both ways? Cell 90,197-200.
Schweizer, A., Fransen, J. A. M., Bächi, T., Ginsel, L. and Hauri, H.-P.(1988). Identification, by a monoclonal antibody, of a 53-kD proteinassociated with a tubulo-vesicular compartment at the cis-side of the Golgicomplex. J. Cell Biol. 107, 1643-1653.
Shaywitz, D. A., Orci, L., Ravazzola, M., Swaroop, A. and Kaiser, C. A.(1995). Human SEC13Rp functions in yeast and is located on transpvesicles budding from the endoplasmic reticulum. J. Cell Biol. 128, 769-777.
Simon, J. P., Ivanov, I. E., Shopsin, B., Hersh, D., Adesnik, M. andSabatini, D. D. (1996). The in vitro generation of post-Golgi vesiclescarrying viral envelope glycoproteins requires an ARF-like GTP-bindingprotein and a protein kinase C associated with the Golgi apparatus. J. Biol.Chem. 271, 16952-16961.
Simpson, F., Bright, N. A., West, M. A., Newman, L. S., Darnell, R. B. andRobinson, M. S. (1996). A novel adaptor-related protein complex. J. CellBiol. 133, 749-760.
Sönnichsen, B., Watson, R., Clausen, H., Misteli, T. and Warren, G. (1996).Sorting by COP I-coated vesicles under interphase and mitotic condition.J. Cell Biol. 134, 1411-1425.
Sohn, K., Orci, L., Ravazzola, M., Amherdt, M., Bremser, M., Lottspeich,F., Fiedler, K., Helms, J. B. and Wieland, F. T. (1996). A majortransmembrane protein of Golgi-derived COPI-coated vesicles involved coatomer binding. J. Cell Biol. 135, 1239-1248.
Stamnes, M. A. and Rothman, J. E. (1993). The binding of AP-1 clathrinadaptor particles to Golgi membranes requires ADP-ribosylation factor,small GTP-binding protein. Cell 73, 999-1005.
Tanigawa, G., Orci, L., Amherdt, M., Ravazzola, M., Helms, J. B. andRothman, J. E. (1993). Hydrolysis of bound GTP by ARF protein triggersuncoating of Golgi-derived COP-coated vesicles. J. Cell Biol. 123, 1365-1371.
Tang, B. L., Peter, F., Krijnse-Locker, J., Low, S. H., Griffiths, G. andHong, W. (1997). The mammalian homolog of yeast Sec13p is enriched the intermediate compartment and is essential for protein transport from tendoplasmic reticulum to the Golgi apparatus. Mol. Cell Biol. 17, 256-266.
Teal, S. B., Hsu, V. W., Peters, P. J., Klausner, R. D. and Donaldson, J. G.(1994). An activating mutation in ARF1 stabilizes coatomer binding toGolgi membranes. J. Biol. Chem. 269, 3135-3138.
Tisdale, E. J., Bourne, J. R., Khosravi-Far, R., Der, C. J. and Balch, W. E.(1992). GTP-binding mutants of rab1 and rab2 are potent inhibitors ovesicular transport from the endoplasmic reticulum to the Golgi complexJ. Cell Biol. 119, 749-761
Traub, L. M., Ostrom, J. A. and Kornfeld, S. (1993). Biochemical dissectionof AP-1 recruitment onto Golgi membranes. J. Cell Biol. 123, 561-573.
Vaux, D., Tooze, J. and Fuller, S. (1990). Identification by anti-idiotypeantibodies of an intracellular membrane protein that recognizes mammalian endoplasmic reticulum retention signal. Nature345, 495-502.
West, M. A., Bright, N. A. and Robinson, M. S. (1997). The role of ADP-ribosylation factor and phospholipase D in adaptor recruitment. J. Cell Biol.138, 1239-1254.
Whitney, J. A., Gomez, M., Sheff, D., Kreis, T. E. and Mellman, I. (1995).Cytoplasmic coat proteins involved in endosome function. Cell 83, 703-713.
Xu, H. and Shields, D. (1993). Prohormone processing in the trans-Golginetwork: endoproteolytic cleavage of prosomatostatin and formation onascent secretory vesicles in permeabilized cells. J. Cell Biol. 122, 1169-1184.
Zerial, M. and Stenmark, H. (1993). Rab GTPases in vesicular transport.Curr. Opin. Cell Biol. 5, 613-620.
Zerial, M. and Huber, L. A. (editors) (1995). Guidebook to the SmallGTPases. Sambrook and Tooze Publication at Oxford University Press.
Zhang, C. J., Rosenwald, A. G., Willingham, M. C., Skuntz, S., Clark, J.and Kahn, R. A. (1994). Expression of a dominant allele of human ARF1inhibits membrane traffic in vivo. J. Cell Biol. 124, 289-300.





























