Embed Size (px)
Citation preview

J. Biomechanics Vol. 27, No. 1. pp. 77-87, 1994.
Printed in Great Britam
0021-9290194 WOO+ 00
i(?, 1993 Pergamon Press Ltd
THREE-DIMENSIONAL KINEMATICS OF THE INTACT AND CRANIAL CRUCIATE LIGAMENT-DEFICIENT STIFLE OF
DOGS
D. L. KORVICK, G. .I. PIJANOWSKI* and D. J. SCHAEFFER
Department of Veterinary Biosciences, University of Illinois, Urbana, IL 61801, U.S.A.
Abstract-Joint motion was measured for the intact and cranial cruciate ligament (CCL)-deficient stifle in five large dogs using an instrumental spatial linkage and radiophotogrammetry. The linkage was directly attached to the bones using modified bone plates. Motion data were presented in terms of a clinical coordinate system. Stifle kinematics were highly reproducible for a given dog, For intact stifle kinematics. the swing phase included flexion+zxtension, internal-external rotation and abduction-adduction, while stance-phase kinematics included only flexion-extension. Loss of the CCL altered joint kinematics over the entire gait cycle. At the swing-stance transition, the CCL-deficient stifle demonstrated an abrupt cranial subluxation of the tibia which was sustained throughout stance. At the beginning of the swing phase the CCL-deficient joint returned to the cranial+audal alignment seen in the intact joint. The dog’s stifle was shown to be CCL-dependent during stance. The dog compensated for CCL loss by reducing the external limb load and by carrying the limb in greater flexion throughout the gait cycle, but was unable to prevent joint subluxation during stance. Stifle kinematics are greatly altered following CCL loss. It is the repetitive, mechanical, dynamic subluxation in a CCL-deficient joint which may be the cause of joint degeneration.
INTRODUCTION
Rupture of the cranial cruciate ligament (CCL)? is a common cause of hindlimb lameness in dogs; CCL loss causes stiflet joint laxity and subsequent joint degeneration (Arnoczky, 1985). Experimental trans- ection of the CCL consistently produces joint degener- ation and has been defined as a mechanically induced osteoarthritis model (Pond and Nuki, 1973). It is commonly believed that joint laxity is a factor in the development of joint degeneration.
In the canine stifle, CCL function has been defined solely on quasi-static tests. In vitro, ligament sec- tioning studies have shown that the CCL limits cranial displacement of the tibia on the femur and internal tibia1 rotation (Arnoczky and Marshall, 1977). The dynamic function of the CCL during ambulation, however, remains unknown. In the dynamic state, joint compression and muscle forces contribute to joint stability. These dynamic components may be able to compensate for CCL loss and maintain intact joint kinematics, i.e. prevent cranial tibia1 subluxation. In man, EMG studies have shown that patients com- pensated for anterior cruciate ligament (ACL) loss by increasing their hamstrings activity while diminishing the quadriceps activity (Limbird et al., 1988). Berchuck et al. (1990) also found that patients with ACL-deficient knees had a reduced maximum flexion moment about the knee, which he equated with a reduced quadriceps force. To date, there is no convin-
Received in jinalform 13 January 1993. *Author to whom correspondence should be addressed. t The equivalent human structure is the anterior cruciate
ligament (ACL). $ The equivalent human structure is the knee.
cing evidence that the laxity which is present in the passive joint also occurs during dynamic walking
activity. Our objective was to study the function of the CCL
in the dynamic state. We measured the three-dimen-
sional kinematics of the intact and CCL-deficient stifle during walking and trotting. We hypothesized that dogs may be able to compensate for CCL loss and thus maintain intact stifle joint kinematics. A comparison between the CCL intact and CCL-deficient kinematics will be used to determine under what conditions and flexion angles the CCL is essential in maintaining intact joint kinematics and hence function.
MATERIALS AND METHODS
Five dogs weighing 33&375 N were used in this study. Joint motion was measured before and after CCL transection using an instrumented spatial link-
age (ISL) and radiophotogrammetry (Korvick, 1991). This work required performing multiple surgical pro- cedures (Table 1). The surgical protocol was reviewed and approved by the Laboratory Animal Care and Advisory Committee of the University of Illinois.
During the initial surgery, modified bone plates were surgically attached to the dog’s left femur and tibia (Fig. 1). These implants were used as a means to repeatably and acutely attach the linkage directly to the bone. Also during the initial surgery, three spher- ical tungsten markers (0.8 mm) were embedded in the femur and tibia to establish radiographically visible surgical coordinate systems. The dogs were given 7 weeks to recover from surgery before kinematic data collection.
We chose bone plates over threaded bone pins as a means to attach the spatial linkage. In a pilot study

78 D. L. KORVICK et al.
using three dogs, we found that acute insertion of bone pins caused a non-weight-bearing lameness despite the use of systemic and local analgesics. Additionally, 6-8 weeks following their insertion, all bone pins were loose and the dogs were still lame.
Prior to kinematic data collection, the linkage was attached to the limb through the plates. The dogs were anesthetized with isoflurane and their limbs were clipped, scrubbed and draped for aseptic surgery. Each implant post was approached through a stab incision and a pin (3.2mm diameter) was threaded into the post. A rod was attached across each pair of femoral or tibia1 pins with Kirschner clamps. The connecting rods carried mounting blocks to which the linkage arms were attached (Fig. 2). The linkage and blocks weighed 2.24 N.
The dogs also received a morphine epidural (0.011 mg kg- ’ diluted in 1 ml of normal saline per 4.54 kg of body mass) to alleviate pain associated with the four stab incisions made to access the plates. Morphine blocks the p-receptors in the spinal cord which are responsible for pain perception. Pain relief from epidural morphine persists for 10 h in all dogs (Bonath and Saleh, 1985). Preliminary trials demon- strated that epidural morphine given to normal dogs did not alter their proprioceptive ability or gait.
The dogs’ recovery from isoflurane was rapid. They were allowed to stand, walk and become accustomed
Table I. Surgical protocol
Implant bone plates (7 weeks)
1 Percutaneous attachment of bone pins
and data collection: intact joint (1 week)
I CCL trtnsection
(7 weeks) 1
Percutaneous attachment of bone pins and data collection: CCL deficient joint
1 Euthanasia
I Post
.5 mm Bone
Pla
1.6 cm
to the linkage prior to collecting kinematic data. The linkage was removed following data collection.
One week following intact stifle kinematic data collection, the CCL was transected at its midsection through a medial arthrotomy. No attempt was made to excise the transected ligament. The joint was lav- aged with lactated Ringers solution. The joint capsule was closed with 3-O prolene sutures and the incision was closed in three layers. The limb was placed in a soft padded bandage for 1 week. The dogs received 5 grains of aspirin twice daily for the remainder of the experiment. We allowed the dogs 7 weeks to recover from CCL transection before kinematic data collec- tion. At the conclusion of data collection the dogs were euthanized.
Limb function was evaluated both by visual obser- vation of the gaiting dogs and by forceplate analysis. Forceplate data were collected by repeatedly walking and trotting the dogs on a leash across a forceplate (AMTI model OR6-3A) until five samples per limb were collected for each gait. The vertical peak force (VPF) was used to assess limb function. The VPF was determined for dogs prior to surgery (presurgical condition), following the dog’s recovery from bone plate implantation and 7 weeks after CCL transection. Limb function in the plated and CCL-deficient condi- tion was reported as the VPF as a percentage of the presurgical VPF. In all cases, forceplate data were collected the day prior to kinematic trials.
A multistep protocol was used to determine stifle kinematics (Table 2). A six-degree-of-freedom spatial linkage similar to Kinzel’s design (Kinzel et al., 1972) was attached to the limb. To signal foot contact with the ground, a pressure-sensitive switch was attached to the fourth digit of the instrumented limb. The dogs were walked and trotted over a level surface covered with rubber mats. Data were collected at 200 Hz for l-5 s using a 1Zbit A/D converter in a 286 computer system. Following data collection, the dogs were anesthetized and stifle kinematics were collected for manual cranial drawer tests at flexion angles approx- imating 40 and 90”. No attempt was made to measure the applied force.
Side View
11 0.12 cm
Top View
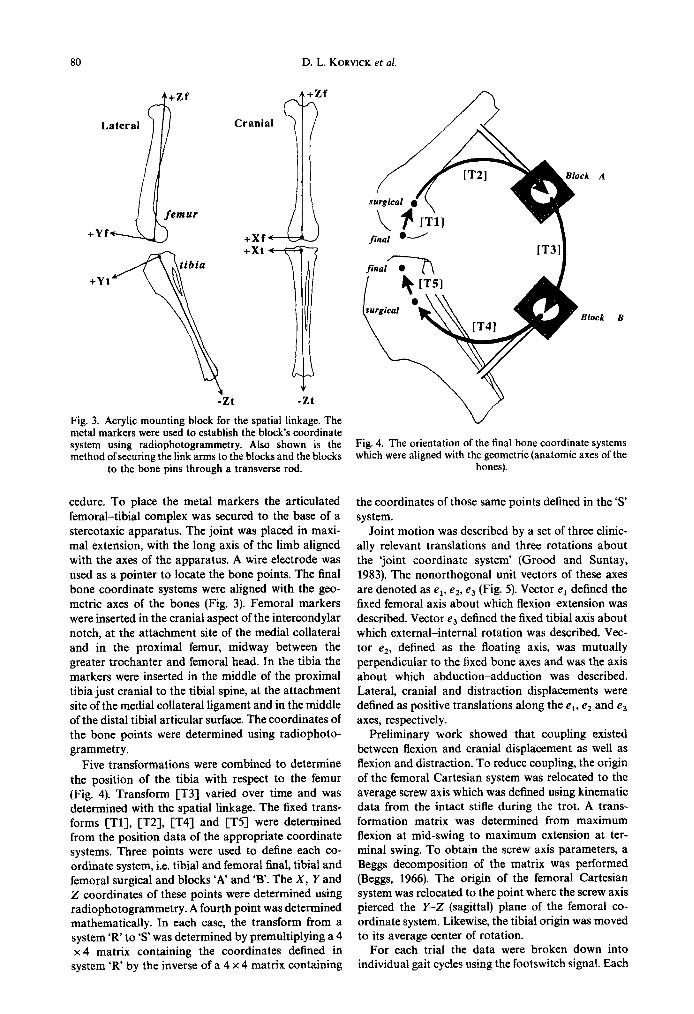
Fig. 1. The five transformations used to determine the final femoral-to-tibia1 transformations. The origins of the surgical and final coordinate systems are shown. Surgical coordinates were established prior to conducting kinematic trials; the final coordinate system established in the cadaver bones. [T3] was determined by using the ISL and varied over time. [Tl], [T2], fl4] and CT53 were Iixed over time and were determined using radiophotogrammetry to determine the coordinates of the points defining each system.

Intact and CCL-deficient stifle of dogs 79
metal
BLOCK 1.6 cm
Fig. 2. The components of the bone implant. Two implants were fixed to the femur and tibia and were attached parallel to the long axis of the bone. The post provided a method to locate the plates under the skin
and as an attachment site for the threaded pins.
Table 2. Data collection protocol
(1) Attach the linkage across the stifle (2) Collect linkage data for the walk and trot (3) Perform cranial drawer tests while collecting linkage
data (dogs are anesthetized) (4) Radiograph limb to determine the bone to linkage
transforms (5) Calibrate the linkage (6) Repeat steps l-5 for CCL-deficient stifle (7) Euthanize dogs (8) Establish final bone coordinate systems (9) Radiograph bones. Determine surgical to final trans-
forms (10) Combine transformations to determine joint motion
To relate the linkage data with the surgical bone based coordinate systems, radiophotogrammetry was conducted. The linkage was removed from the leg, leaving the mounting blocks in place. The dog’s leg was positioned in a radiographic reference object. The reference object was a four-sided PlexigIas box (16.5 cm wide by 30.4 cm long by 12.7 cm deep) with 15 steel balls (1.6 mm reference markers) embedded in each side. Two radiographic exposures were taken using a 102 cm focal-film distance, with the X-ray tube positioned + 15-20” off the vertical axis.
The reference object’s points, bone and mounting block points on both radiographs were digitized. The
digitized and known three-dimensional coordinates of the reference points were used to determine the three- dimensional coordinates of bone and block points using the direct linear transformation (DLT) al- gorithm (Adbel-Aziz and Karara, 1971). Each pair of radiographs was digitized three times and the average DLT derived bone and block coordinates calculated. From these data the bone to block transforms were obtained.
The linkage output voltages were filtered using a fourth-order low-pass Butterworth digital filter with a 10 Hz cutoff frequency. This was appropriate for gait data which have a frequency component of 15 Hz or less (Antonsson and Mann, 1985). The linkage was calibrated using the method of Sommer and Miller (1981). The calibration parameter estimates and the filtered voltages were used to determine the linkage transformations.
The experimental protocol was repeated for the CCL-deficient condition. After taking radiographs of the CCL-deficient joint, the dogs were euthanized with an intravenous overdose of barbiturate.
The final coordinate systems were established in the cadaver bones. Establishing a final coordinate system was necessary to allow comparisons between animals. Placing markers in cadaver bones was more repro- ducible than placing markers during a surgical pro-
80 D. L. KORVICK et al.
c Lateral
il
Cranial
:I
+Yf ? -..
I II
femur
+Xf +Xt
tibia
+Yt !
T 1
1
-zt -zt
Fig. 3. Acrylic mounting block for the spatial linkage. The metal markers were used to establish the block’s coordinate system using radiophotogrammetry. Also shown is the method of securing the link arms to the blocks and the blocks
to the bone pins through a transverse rod.
cedure. To place the metal markers the articulated femoral-tibia1 complex was secured to the base of a stereotaxic apparatus. The joint was placed in maxi- mal extension, with the long axis of the limb aligned with the axes of the apparatus. A wire electrode was used as a pointer to locate the bone points. The final bone coordinate systems were aligned with the geo- metric axes of the bones (Fig. 3). Femoral markers were inserted in the cranial aspect of the intercondylar notch, at the attachment site of the medial collateral and in the proximal femur, midway between the greater trochanter and femoral head. In the tibia the markers were inserted in the middle of the proximal tibia just cranial to the tibia1 spine, at the attachment site of the medial collateral ligament and in the middle of the distal tibia1 articular surface. The coordinates of the bone points were determined using radiophoto- grammetry.
Five transformations were combined to determine the position of the tibia with respect to the femur (Fig. 4). Transform [T3] varied over time and was determined with the spatial linkage. The fixed trans- forms [Tl], [T2], [T4] and [T5] were determined from the position data of the appropriate coordinate systems. Three points were used to define each co- ordinate system, i.e. tibia1 and femoral final, tibia1 and femoral surgical and blocks ‘A’ and ‘B’. The X, Y and Z coordinates of these points were determined using radiophotogrammetry. A fourth point was determined mathematically. In each case, the transform from a system ‘R’ to ‘s’ was determined by premultiplying a 4 x 4 matrix containing the coordinates defined in
system ‘R’ by the inverse of a 4 x 4 matrix containing
fina; i-
Block B
Fig. 4. The orientation of the final bone coordinate systems which were aligned with the geometric (anatomic axes of the
bones).
the coordinates of those same points defined in the ‘S system.
Joint motion was described by a set of three clinic- ally relevant translations and three rotations about the ‘joint coordinate system’ (Good and Suntay, 1983). The nonorthogonal unit vectors of these axes are denoted as e,, e2. e3 (Fig. 5). Vector e, defined the fixed femoral axis about which flexionextension was described. Vector e3 defined the fixed tibia1 axis about which external-internal rotation was described. Vec- tor e2, defined as the floating axis, was mutually perpendicular to the fixed bone axes and was the axis about which abduction-adduction was described. Lateral, cranial and distraction displacements were defined as positive translations along the e,, e, and e3 axes, respectively.
Preliminary work showed that coupling existed between flexion and cranial displacement as well as flexion and distraction. To reduce coupling, the origin of the femoral Cartesian system was relocated to the average screw axis which was defined using kinematic data from the intact stifle during the trot. A trans- formation matrix was determined from maximum flexion at mid-swing to maximum extension at ter- minal swing. To obtain the screw axis parameters, a Beggs decomposition of the matrix was performed @eggs, 1966). The origin of the femoral Cartesian system was relocated to the point where the screw axis pierced the Y-Z (sagittal) plane of the femoral co- ordinate system. Likewise, the tibia1 origin was moved to its average center of rotation.
For each trial the data were broken down into individual gait cycles using the footswitch signal. Each

Intact and CCL-deficient stifle of dogs 81
External rotation
GIL Fixed Tibia1 Axis
Fig. 5. Stifle kinematics were defined using the clinical joint coordinate system. The drawing shows axes of rotation and translation. The six parameters of motion were flexion-extension, abduction-adduction, internal-external rotations and craniakaudal, medial-lateral and compression-distraction translations.
cycle was divided into its swing and stance compon- ents and the components interpolated at 2% intervals using a cubic spline to normalize the data from 0 to 50%. Normalization of the data permitted averaging across multiple cycles despite variations in the speed of gait. Normalization did not affect the shape of the curves or relative timing during the gait cycle. The average kinematic curves for a given dog, gait and condition were obtained from averaging 25 gait cycles. The average kinematic curves across all dogs were determined using distance-weighted least-squares smoothing.
To determine whether there was coupling between select kinematic parameters, pooled Pearson correla- tion matrices were determined for swing and stance across dog, gait and condition. A matrix of Bonferroni probabilities was calculated. The gait parameter pairs were said to be coupled when P<O.OS.
The difference between intact and deficient kin- ematics was determined for each dog at the walk and trot. An average difference across all dogs was calcu- lated.
To determine experimentally the error of the com- bined system, a plastic model was built which contain-
ed multiple markers to simulate the process of data acquisition used in the dog trials. The model contain- ed the four transformations which were acquired using the radiophotogrammetry and the fifth using the linkage. Error was defined as the difference between the actual and experimental measurements.
RESULTS
At the time of the intact stifle kinematic trials, three levels of recovery from implant surgery were identi- fied. Two dogs recovered fully with no reduction in VPF compared to the presurgical VPF. One dog was classified as mildly impaired because it demonstrated a 0% reduction in walk VPF and a 20% reduction in trot VPF compared to the presurgical VPF; the dog showed no visible signs of lameness. Two dogs were moderately impaired by the implants; they experi- enced a 25% reduction in trot VPF and were slightly lame. These two dogs would shift their weight to the opposite hindlimb during standing. In evaluating these results it became clear that the two dogs with the worst limb function had longer implant posts. The
82 D. L. KORVICK et al.
improved function of subsequent dogs was attributed to using shorter posts. Subjectively, dogs were not bothered by having the linkage attached to the limb and were able to walk and trot with the linkage attached to the leg.
The dogs were given 7 weeks to regain partial weight bearing on the limb following CCL trans- ection. At the time of CCL-deficient kinematic data collection, all dogs were visibly lame. The trot VPF was reduced by 60% compared to the presurgical, intact stifle VPF data. The lameness was accentuated at the faster gaits. One dog could use its left hindlimb only at the slower walking gait. All dogs had notice- able atrophy of quadriceps and biceps muscles and a positive cranial drawer sign at the time of the CCL- deficient trials.
No changes were observed in the appearance of the articular cartilage or synovial tissue at the time of surgical transection of the CCL 7 weeks following the attachment of the implants to the bone.
Post mortem examination of the stifle revealed changes consistent with early-stage osteoarthritis. The trochlear ridges were covered by a few flat raised osteophytes. Synovial tissue had proliferated to cover the joint capsule and remnant of the CCL. Cartilage fibrillation was present on the tibia1 plateau in the
I .!
90 ;‘ . .
IO ::. : 70 .!.
90
30
~\ .:
..!!
40 ’ *c.,! e,
*-_----a--_ _.dL___-_-~-,+ 30 . :
z;m :::___:,.::::::_::;:: 9
I :;., ,,::..I:.. _,.*.” .‘(I:,,,,.........I‘:::!:: _,o . ..., :
-so’
30
20 1
. . ...’ .’
-30 . . . ( + . . ’ . - 0 10 20 30 40 50 0 IO 20 30 40 so
SWING STANCE
areas not covered by the menisci. However, the menisci were intact in all dogs.
The average linkage error and standard deviation for the 10 experiments was X =0.33 kO.03 mm, Y=O.37+0.03 mm, 2=0.57+0.04mm. For the Z-Y-X Euler angles, the average error, in degrees, was Z=OS+O.l, Y=1.3+0.5, X=1.0+0.2. This error was similar to the linkage error reported by Scherrer et al. (1979), Sommer and Miller (1981) and Suntay et al. (1983).
The radiophotogrammetry average error and standard deviation for the 10 experiments was X = 0.23 f 0.02 mm, Y=O.12+0.01 mm, Z=O.27 f 0.02 mm.
The error in combining four radiophotogrammetric transformations and a fifth linkage transformation was defined in terms of the clinical coordinate system. The average error and standard deviation for four experiments was 0.7 + 0.4” for flexion*xtension, 3.0 f2.2”C for abduction-adduction and 1.9-t 1.1” for external-internal rotation. The average error for iat- era1 displacement was 1.8 f 1.6 mm, cranial displace- ment 3.1+ 1.9 mm and distraction 1.5+ 1.9 mm.
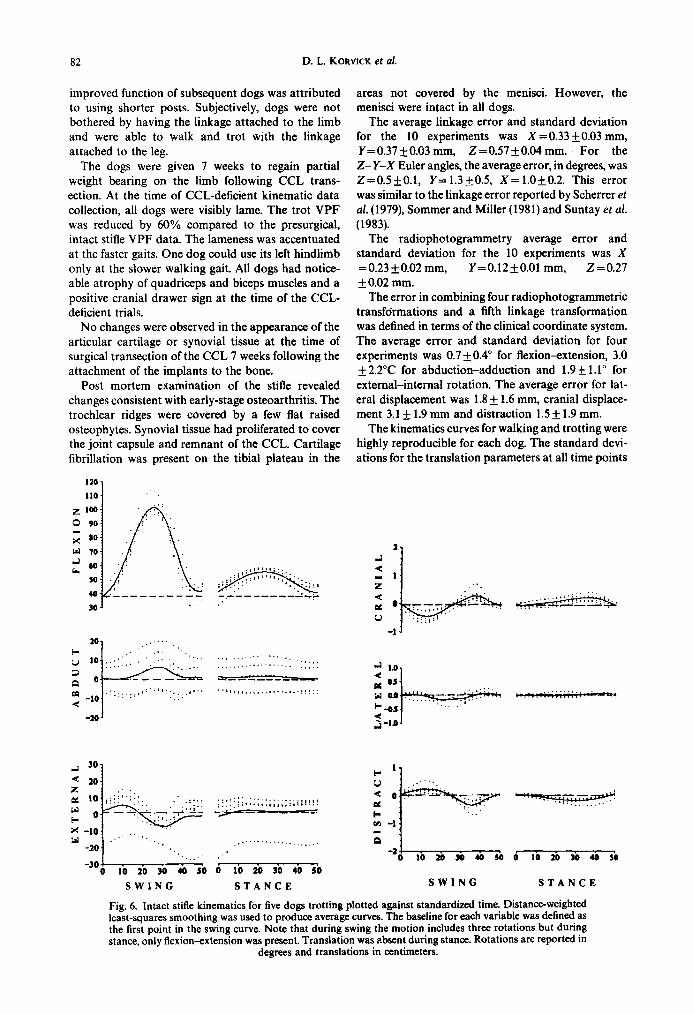
The kinematics curves for walking and trotting were highly reproducible for each dog. The standard devi- ations for the translation parameters at all time points
I- l v .‘..,
q o ::‘,._:, ’ i-t----- 7.7 :, ,, . ,. .’ ..’ 1.. --
m -:
c .:
“-21. . . . . ? 0 10 29 30 40 50 0 IO 20 30 40 50
SWING STANCE
Fig. 6. Intact stifle kinematics for five dogs trotting plotted against standardized time. Distance-weighted least-squares smoothing was used to produce average curves. The baseline for each variable was defined as the first point in the swing curve. Note that during swing the motion includes three rotations but during stance, only flexion-extension was present. Translation was absent during stance. Rotations are reported in
degrees and translations in centimeters.

Intact and CCL-deficient stifle of dogs 83
across the 25 gait cycles averaged 0.2 mm for all dogs. ard deviations averaged 0.2 mm for lateral, 0.5 mm for The average standard deviation for flexion was 1.2”, cranial and 0.5 mm for distraction displacements. abduction 0.4” and external rotation 0.8”. Figure 6 Figure 7 shows the average kinematic variables plot- shows the intact stifle kinematic variables for all dogs ted against standardized time for the CCL-deficient plotted against standardized time for the trot. Dis- trials for all dogs at the trot. Distance-weighted least- tance-weighted least-squares smoothing was used to squares smoothing was used to produce the average fit a line through the points to produce average kinematic curves across dogs. A horizontal line was kinematic curves across dogs. The baseline for each drawn across the graphs to represent the baseline for parameter was defined using the first point in the the intact stifle kinematic parameters. In the CCL- swing curve. Kinematic curves for walk were very deficient stifle, walking and trotting parameters were similar to the trotting data. Differences included larger generally similar. Trotting produced larger rotation flexion, internal rotation and abduction peaks at mid- maxima for flexion, internal rotation and abduction swing for the trotting gait. compared to walking.
Pearson correlations for abduction and external rotation were significant (P<O.O5) for both walking and trotting during both swing and stance. For the intact stifle, an increase in abduction was accom- panied by an increase in internal rotation. Likewise, Pearson correlations for cranial displacement and distraction were significant (P < 0.05) for the walk and trot. For the intact stifle, cranial displacement was inversely correlated with distraction.
Pearson correlations were significant (P < 0.05) for abduction and external rotation for the walk during swing and stance. Correlations were significant be- tween cranial displacement and distraction during both swing and stance phases of the walk and trot.
Stifle kinematics for the CCL-deficient joint were likewise very reproducible for each dog and gait. The rotation standard deviations for the 25 cycles aver- aged 1.3, 0.5 and 0.9” for flexion, abduction and external rotation, respectively. The translation stand-
The dogs carried the CCL-deficient joint in approx- imately 5-14” more flexion during walking (Fig. 8). These differences were less pronounced during trot- ting. Internal rotation increased following CCL trans- ection, most noticeably during the mid-stance phase of gait.
The changes in cranial translation following CCL transection were consistent across all dogs. Most notable was the marked cranial displacement at the
30
20 ,.
IO .., ,,,.:”
0
-10 -_ .’ ..,..,
7.7 - - - - -.-7 ..:.
. . . .
. . . -20 ‘.
-3oA I . 1 * 0 10 20 30 40 so 0 IO 20 30 40 50
SWING STANCE SWING STANCE
_A---_----_
-g--&J . . _:.’
0 IO 20 30 6a so
Fig. 7. CCL-deficient stifle kinematics for five dogs trotting plotted against standardized time. Distance- weighted least-squares smoothing was used to produce average curves. The baseline for each variable was that used for the intact kinematics. Note that during swing and stance the motion includes three rotations. Translation is present during stance.. Rotations are reported in degrees and translations in centimeters.
27:1-F
84 D. L. KORVICK et al.
16 - flex
10 -
5-
0
+_&$zA+
Stance
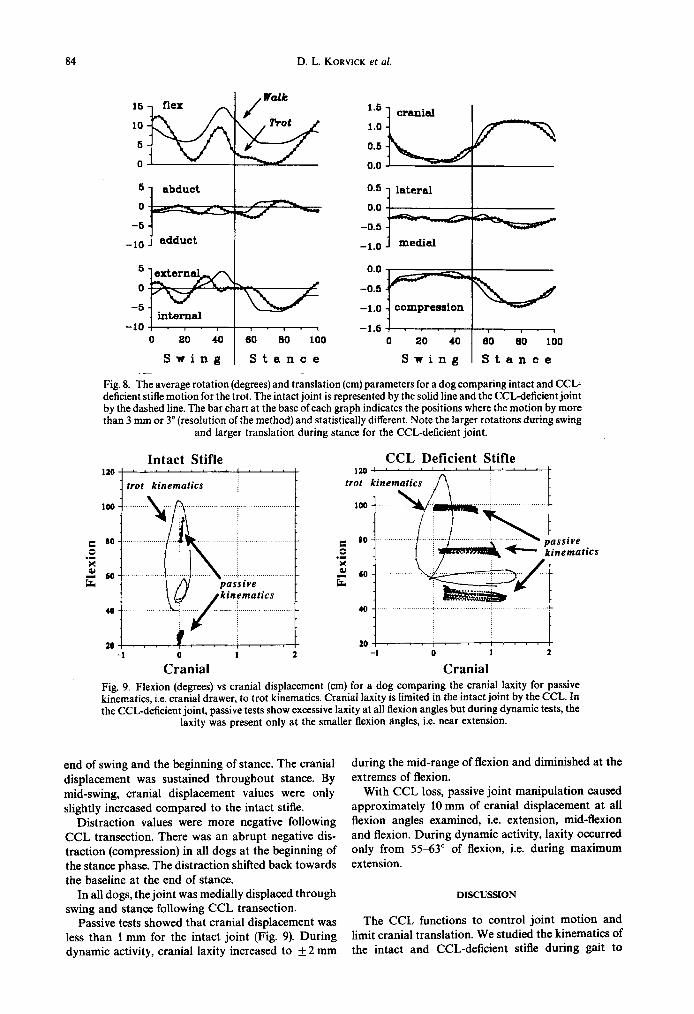
Fig. 8. The average rotation (degrees) and translation (cm) parameters for a dog comparing intact and CCL- deficient stifle motion for the trot. The intact joint is represented by the solid line and the CCL-deficient joint by the dashed line. The bar chart at the base of each graph indicates the positions where the motion by more than 3 mm or 3” (resolution of the method) and statistically different. Note the larger rotations during swing
and larger translation during stance for the CCL-deficient joint.
Intact Stifle 110""' : ',"I""'
trot kinematics i
CCL Deficient Stifle 120""' I
trot kinematics
-1
CrOanial 1 1 0 1 2
Cranial Fig. 9. Flexion (degrees) vs cranial displacement (cm) for a dog comparing the cranial laxity for passive kinematics, i.e. cranial drawer, to trot kinematics. Cranial laxity is limited in the intact joint by the CCL. In the CCL-deficient joint, passive tests show excessive laxity at all flexion angles but during dynamic tests, the
laxity was present only at the smaller flexion angles, i.e. near extension.
end of swing and the beginning of stance. The cranial displacement was sustained throughout stance. By mid-swing, cranial displacement values were only slightly increased compared to the intact stifle.
Distraction values were more negative following CCL transection. There was an abrupt negative dis- traction (compression) in all dogs at the beginning of the stance phase. The distraction shifted back towards the baseline at the end of stance.
In all dogs, the joint was medially displaced through swing and stance following CCL transection.
Passive tests showed that cranial displacement was less than 1 mm for the intact joint (Fig. 9). During dynamic activity, crania1 laxity increased to f2 mm
during the mid-range of flexion and diminished at the extremes of flexion.
With CCL loss, passive joint manipulation caused approximately 10 mm of crania1 displacement at all flexion angles examined, i.e. extension, mid-flexion and flexion. During dynamic activity, laxity occurred only from 5563” of flexion, i.e. during maximum extension.
DISCUSSION
The CCL functions to control joint motion and limit crania1 translation. We studied the kinematics of the intact and CCL-deficient stifle during gait to

Intact and CCL-deficient stifle of dogs 85
determine whether dogs could compensate for CCL loss and maintain intact joint kinematics. In addition, we were interested in identifying the periods during the gait cycle where the CCL is essential in preventing excessive cranial joint subluxation.
An invasive method, similar to that described by Scherrer et al. (1979), was used to measure stifle kinematics. All dogs gaited without difficulty. In three of five dogs limb function was normal based on forceplate data. In two of five animals, the external ground reaction force was reduced by 25%. This is an unavoidable limitation in using an invasive method of measuring stifle motion. The tradeoff for accepting these changes in limb function was that we could accurately track joint motion before and after cutting the CCL. High accuracy was essential to this study because the cranial translations we were measuring were small. Additionally, the effect of the implant on alteration of stifle kinematics should be similar across
intact and CCL-deficient joints. Differences in the kinematics between the two conditions should be due to removal of the CCL alone.
The kinematic data were reproducible within a dog for a given gait, as indicated by the small standard deviations seen across the 25 gait cycles. The shapes of the intact stifle kinematic curves between dogs were similar, but their starting point (baseline) varied. This between-dog variation was caused by the inability to locate reproducibly the bone coordinate systems from dog to dog. The between-dog differences were espe- cially pronounced for the external rotation and abduc- tion parameters which have the smallest range of motion. In contrast, the between-dog translation cur- ves for the intact stifle trials were nearly coincident. This resulted from relocating the femoral and tibia1 origins to an average screw axis and forcing the origins to be coincident at the beginning of the swing phase. This final step was important in unifying the trans- lation descriptions of motion between dogs.
The between-dog kinematic curves for the CCL- deficient stifle showed more variability even for the previously coherent translation data. This suggests that there was variability in the way each dog modified its gait to adapt to the absence of the CCL. It may also reflect the effect of a shift of the instant center of rotation caused by CCL transection. As noted pre- viously, the translation parameters are especially sensitive to having the bones’ coordinate origins posi- tioned away from the joint’s center of rotation.
There is no reported three-dimensional kinematic data in the literature for the canine stifle. Lafortune (1983) and Lafortune and Cavanagh (1985) measured human knee kinematics using an invasive technique and described knee motion in terms of the clinical coordinate system. Comparisons between the dog stifle and human knee kinematics are of interest because the dog is frequently used to model the human knee. Additionally, the anatomic features of the dog and human joints are generally quite comparable. The shapes of the kinematic curves for flexion and abduc-
tion were similar between the canine stifle and human knee. Both the canine and human joints demonstrated internal rotation during swing but the temporal com- ponents differed. In the human knee maximum inter- nal rotation occurred before peak flexion, while in the dog’s stifle maximum internal rotation occurred at peak flexion. For both man and dog, the motion during the swing phase of gait is dominated by flexion-extension and also include internal-external and abduction-adduction rotations. By contrast, the motion during stance included only small flexion-extension rotations for both dog and man.
The CCL force was shown to be essential in balan- cing the sum of forces across the joint during stance. A
comparison between intact and CCL-deficient stifle cranial displacement curves showed that the CCL functions to resist cranial displacement during the stance phase of gait. Our assumption was that the increased cranial translation in the CCL-deficient joint was due to an unopposed cranial force.
All dogs adapted to the CCL insufficiency by dra- matically reducing the load on the injured limb. The
dogs also carried their CCL-deficient limbs in more flexion while walking. This may represent a conscious effort on the part of the dog to avoid maximum extension, a position where the joint is likely to subluxate cranially under load. Alternatively, the in- crease in flexion angle may represent reflex inhibition of quadriceps contraction related to excessive joint fluid. Experimental evidence demonstrated a 3&50% inhibition of reflex-evoked quadriceps contraction caused by injection of 60 ml of saline into the human knee (Kennedy et al., 1982). Dogs experience joint effusion following CCL transection (Person, 1982) and presumably will experience an inhibition of reflex- evoked quadriceps as in man.
Dynamic and passive cranial laxity patterns differ- ed. This difference was due to the addition of external limb and muscle forces in the dynamic state. In the intact joint a manual test demonstrated less than a millimeter of cranial laxity, while in the dynamic tests the laxity increased to f2 mm. This difference was due to the larger forces imposed on the joint during walking. In the CCL-deficient joint manual tests demonstrated up to 11 mm of cranial laxity at all flexion angles, whereas during the dynamic tests large cranial displacements only occurred while the joint was maximally extended. During walking, the dynam- ic components of joint compression and muscle forces were presumably able to mask the excessive cranial joint laxity during the swing phase of gait. By contrast, the dynamic components could not prevent excessive cranial joint laxity during the stance phase of gait.
Quadriceps contraction may be one component driving the cranial tibia1 subluxation in the dogs lacking a CCL. Quadriceps contraction generates strain in the CCL at specific flexion angles (Arms et al., 1984; Daniel et al., 1987). Kain et al. (1988) showed that quadriceps contraction in rhesus monkeys produced CCL strain at 0 and 45” flexion and CCL shortening

86 D. L. KORVICK et al.
(laxity) at 90” flexion. Presumably in the absence of the CCL, quadriceps contraction will cause cranial tibia1 subluxation at flexion angles from 0 to 45”. In the dog, the quadriceps muscle contracts during the stance phase of gait (Wentink, 1976), at which time the joint angle varies from 4&60”. This is within the range of flexion where quadriceps contraction can produce CCL strain (Kain et al. 1988) or subluxate a CCL- deficient joint. During the swing phase, however, the stifle is flexed past 60” and quadriceps forces cannot load the CCL or cause tibia1 subluxation in a CCL- deficient joint. Thus, during swing there is no driving force to alter the cranial position of the joint in the absence of the CCL. The swing phase is thus CCL- independent, while the stance phase is CCL-depend- ent.
It is interesting to note that in one dog cranial tibia1 subluxation occurred during the swing phase follow- ing CCL loss. The joint components never fully re- aligned themselves during the swing phase as seen in the other dogs and the joint would drift from min- imum to maximum drawer through the complete cycle. This pattern probably represents a more ad- vanced case of the injury where secondary restraints, i.e. joint capsule and collateral ligaments, have been stretched by the chronic instability. Serial kinematic studies would be necessary to substantiate this idea.
Cranial displacement and distraction were signific- antly correlated for both intact and CCL-deficient stifle kinematics. The coupling was most likely due to the geometry of the tibia1 plateau which slopes in a caudal-distal direction on an average of 30” (Slocum and Devine, 1983). As the tibia slides forward the femur must, by design, move caudally and distally. At some point it is possible for the femoral origin to drop below the tibia1 origin, which results in negative distraction values. Both the cranial displacement and negative distraction increased following CCL trans- ection, most notably when the dog was weight-bear- ing. This observation suggests that the geometry of the joint together with load across the joint may provide a mechanism to either load the CCL or subluxate the joint in the CCL-deficient stifle.
The sequential pattern of joint changes which occur following CCL loss proceeds gradually over the course of several months. Initially, there is synovitis and cartilage fibrillation followed by osteophyte formation, meniscal tearing and cartilage loss. This pattern of injury suggests that a repetitive factor is causing the damage. We suggest that it is the cranial tibia1 subluxation which occurs with each step that slowly degrades the CCL-deficient joint. Additional studies are needed to substantiate this idea.
In summary, a combined method using a linkage with radiophotogrammetry provided a sensitive, accurate method of measuring the three-dimensional motion of the canine stifle joint over time. It was possible to attach the linkage to the bones directly without severely impairing the dog’s gait. Kinematic data were hiehlv renroducible for a &en dog for both
the lame (CCL-deficient stifle) and sound dog (intact stifle). Intact stifle kinematics were characterized by a combination of three rotations during the swing phase and a pure flexionextension motion during the stance phase of gait. Translation motion was absent during stance for the intact stifle. CCL loss altered the joint kinematics over the entire cycle. Changes in trans- lation were most severe for the stance phase of gait. In the CCL-deficient stifle, there was an abrupt cranial subluxation of the tibia beginning at heel strike and ending at foot-off. The dogs compensated for loss of the CCL by reducing the external load on the limb and carrying the limb in greater flexion. Despite these adaptations, the dogs could not prevent the cranial joint subluxation during the stance phase of gait. The dog’s stifle was thus shown to be CCL-dependent during the stance phase.
REFERENCES
Abdel-Aziz, Y. I. and Karara, H. M. (1971) Direct linear transformation from comparator coordinates into object space coordinates in close-range photogrammetry. In Proc. ASP/U1 Symp. on Close-Range Photogrammetry, pp. 1-18, Urbana, IL. American Society of Photogram- metry, Falls Church.
Antonsson, E. K. and Mann, R. W. (1985) The frequency content of gait. J. Biomechanics 18, 3947.
Arms, S. W., Pope, M. H., Johnson, R. J., Fisher, R. A., Arvidsson, I. and Eriksson, E. (1984) The biomechanics of anterior cruciate rehabilitation and reconstruction. Am. 1. Sports Med. 12, 8-l 1.
Arnoczky, S. P. (1985) Crania1 cruciate ligament rupture and associated injuries. In Textbook of Small Animal Ortho- pedics (edited by Newton, C. and Nunamaker, D.), pp. 932-940. Lippincott, Philadelphia.
Arnoczky, S. P. and Marshall, J. L. (1977) The cruciate ligaments of the canine stifle: an anatomical and functional analvsis. Am. J. Vet. Res. 38. 1807-1814.
Beggs,J. (1966) Advanced Mechbnics. MacMillan, New York. Berchuck, M., Andriacchi, T. P., Bach, B. R. and Reider, B.
(1990) Gait adaptations by patients who have a deficient cruciate ligament. J. Bone Jt Surg. 72A, 871-877.
Bonath, K. H. and Saleh, A. S. (1985) Long term pain treatment in the dog by peridural morphine. Proc. 2nd Int. Cong. Vet. Anesth. p. 161.
Daniel, D. M., Malcom, L. L. and Losse, G. (1987) The synergistic action on the anterior cruciate ligament and thigh muscles in maintaining joint stability. Am. J. Sports Med. 15,207-231.
Grood, E. S. and Suntay, W. J. (1983) A joint coordinate system for the clinical description of three-dimensional motions: application to the knee. J. biomech. Engng 105, 136144.
Kain, C. C., McCarthy, J. A., Arms, S., Pope, M. H., Steadman.1. R.. Manske, P. R. and Shively, R. A. (1988) An in viuo analysis of the effect of transcutaneous electrical stimulation of the quadriceps and hamstrings on anterior cruciate ligament deformation. Am. J. Sports Med. 16, 147-152.
Kennedy, J. C., Alexander, I. J. and Hayes, K. C. (1982) Nerve supply of the human knee and its functional importance. Am. J. Sports Med. 10, 329-335.
Kinzel. G. L., Hall, A. S. and Hillberry, B. M. (19721 Measurement of total motion between two body segments. analytical development. J. Biomechanics 5, 93-105.
Korvick, D. L. (1991) Joint motion studies of the normal and

Intact and CCL-deficient stifle of dogs 81
cranial cruciate deficient stifle in large breed dogs. Ph.D. osteoarthritis in the dog. Ann. Rheum. Dis. 32, 387-388. thesis, University of Illinois. Scherrer, P. K., Hillberry, B. M. and VanSickle, D. C. (1979)
Lafortune, M. A. (1983) A three dimensional investigation of Determining the in oivo areas of contact in the canine the knee joint during walking. Ph.D. thesis, The Pennsyl- shoulder. .I. biomech. Engnq 101, 271-278. vania State University.
Lafortune, M. A. and Cavanagh, P. R. (1985) The measure- ment of normal knee joint motion during walking using intracortical pins. In Biomechanical Measurement in Ortho- pedic Practice (edited by Wittle, M. and Harris, D.), pp. 234-243. Clarendon Press, Oxford.
Limbird, T. J., Shiavi, R., Frazer, M. and Borra, H. (1988) EMG profiles of knee joint musculature during walking: change induced by anterior cruciate ligament deficiency.
Slocum, B. and Devine, T. (1983) Cranial tibia1 thrust: a primary force in the canine stifle. J. Am. Vet. Med. Assoc. 1e3,45&459.
Sommer, H. J. and Miller, N. R. (1981) A technique for the calibration of instrumented spatial linkages used for bio- mechanical kinematic measurements. J. Biomechanics 14, 91-98.
Suntay, W. J., Grood, E. S., Hefty, M. S., Butler, D. L. and Noyes, F. R. (1983) Error analysis of a system for
J. orihop. Res. 6, 63&638. measuring three-dimensional joint motion. _J: biomech. Person, M. W. (1982) An evaluation of hyaluronic acid on Engng 105, 127-135.
experimentally induced osteoarthritis in dogs. M.S. thesis. Wentink, G. H. (1976) The action of hind limb musculature of Washington State University. the dog in walking. Acta Anat. 96, 7&80.
Pond, M. J. and Nuki, G. (1973) Experimentally induced