Embed Size (px)
Citation preview

E L S E V I E R Journal of the Neurological Sciences 134 (Suppl.) (1995) 69-78
~ R N A L OF m E
NEUROLOGICAL SCIENCES
The role of transition metals in the pathogenesis of Parkinson's disease
Elisabeth Kienzl ~,b, Leopold Puchinger b, Kurt Jellinger a , * , Wolfgang Linert c, Herbert Stachelberger b, Reginald F. Jameson d
Ludwig Boltzmann Insntute of Clinical Neurobiology, Lainz Hospital, Wolkersbergenstrafle 1, A-I I 30 Vienna, Austria Institute of Applied Botany, Technical Microscopy and Organic Raw Material Studies. University of Technology, Vienna, Austria
Institute of Inorganic Chemistry, University of Technology, Vienna, Austria d Department of Chemistry, University of Dundee, Dundee, Scotland, UK
Accepted 14 July 1995
Abstract
The mechanisms that lead to degeneration of melanized dopaminergic neurons in the brain stem, and particularly in the substantia nigra (SN) in patients with Parkinson's disease (PD) are still unknown. Demonstration of increased iron (Fe) in SN of PD brain has suggested that Fe-melanin interaction may contribute to oxidative neuronal damage. Energy dispersive X-ray electron microscopic analysis of the cellular distribution of trace elements revealed significant Fe peaks, similar to those of a synthetic melanin-Fe 3+ complex, in intraneuronal electron-dense neuromelanin granules of the SN zona compacta, with highest levels in a case of PD and Alzheimer's disease (AD). No Fe increase was found in Lewy bodies or in SN neurons of control specimens. The relevance of the in vitro chemical reactions of dopamine (DA), 5-hydroxydopamine (5-OHDA), and 6-hydroxydopamine (6-OHDA) with Fe 3÷ and with dioxygens for the pathogenesis of PD was investigated. An initiating mechanism for a chain reaction is suggested by which excessive Fe 3+ arises. Melanin can act as an efficient antioxidant and in the presence of Fe can promote the formation of cytotoxic hydroxyl free radicals (. OH) which, in turn, initiate lipid peroxidation and consequent cell damage. While in vitro studies indicate that DA oxidation leading to melanin formation is independent of metal ions, saturation of melanin with large amounts of Fe 3+ causes a significant generation of free • OH radicals. The relevance of the events induced by the melanin-Fe complex for the degeneration of dopaminergic neurons in PD is discussed. Free redox-active metal ions in the cytoplasm may cause site-specific peroxidation and thus exert neurotoxic effects. Excessive hydrogen peroxide in post mortem frontal cortex of a patient with PD and AD could be shown by confocal fluorescence microscopy, and this observation may be a direct indicator of oxidative stress.
Keywords: Dopamine; Hydrogen peroxide; 6-Hydroxydopamine; Iron(ll); Iron(Ill); Melanin; Oxygen radicals; Parkinson's disease
1. Introduct ion
Iron (Fe) and manganese (Mn) are transition elements because their ions have incompletely filled orbitals. Iron occurs in two principal oxidation states, ferrous Fe 2+ (d 6)
and ferric Fe 3+ (dS). The binding strength of Fe z+ and Mn 2+ to ligands, and the intermediate concenlration of free Fe ions suggest that these two metals may play a role in catalytic and regulatory processes in the cell (Crichton, 1991). Transition metals can accept or donate single elec- trons to promote redox reactions and free radical forma- tion. Transferrin and ferritin are proteins responsible for
" Corresponding author. Tel.: ( + 43- I ) 80110 3431 : Fax: ( + 43-1 ) 804 5401.
0022-510X/95/$09.50 © 1995 Elsevier Science B V All rights rese~ed SSDI 0 0 2 2 - 5 1 0 X ( 9 5 ) 0 0 2 1 0 - 3
transport and storage of Fe and are necessary for Fe homeostasis (Morris et al., 1992a).
In the central nervous system (CNS), bound Fe is usually concentrated in specialized membranes, such as in mitochondria, or in the cellular cytoplasm. Most brain iron is stored in an inactive form bound to intracellular ferritin which can be considered a special compartment of its own: a membrane-l ike protein wraps around solid Fe oxyhy- droxide. Analyses of brain ferritin suggested that about 90% of the non-heme Fe content of brain is associated with ferritin and localized in microglia and oligodendro- cytes (Koeppen and Dentinger, 1988; Kaneko et al., 1989; Connor et al., 1990). The human brain Fe content is highest in the substantia nigra, globus pallidus, red nu- cleus, caudate nucleus, and putamen (reviews in Hill and Switzer, 1984; Dwork et al., 1988; Riederer et al., 1989;

70 E. Kienct et al. /Journal ()[the Neurological Sciences 134 (Suppl.) (1995) 69-78
Riederer and Youdim, 1993). Brain transferrin receptors also exhibit a regional distribution with high densities in the cerebral cortex, hippocampus, amygdala, and certain brain stem nuclei (Morris et al., 1994). Immunocyto- chemical studies have shown that brain contains transferrin in the choroid plexus (M~llgard et al., 1979), oligodendro- cytes (Connor and Fine, 1986), some neurons (Dwork et al., 1988), and certain types of astrocytes (Connor et al.. 1990; Gocht and Liihler, 1990).
Increased Fe levels have been observed in several neu- rological disorders, e.g. Parkinson's disease (PD), Hunting- ton's disease, striatonigral degeneration, Hallervorden- Spatz disease (Good et al., 1992a, Good et al., 1992b; Dexter et ai., 1992; Jellinger and Kienzl, 1993). These data and the demonstration of increased Fe content in SN in experimentally induced lesions of the nigrostriatal dopaminergic system by 6-hydroxydopamine (6-OHDA) (Oestreicher et al., 1994) suggest that Fe accumulation is not the primary cause of the selective degeneration of dopaminergic neurons in PD. The degeneration is more likely due to a secondary process after an initial toxic insult involving heterogeneous catecholaminergic cell groups in the brain stem (Hirsch et al., 1988; Jellinger. 1991; Morris and Edwardson, 1994; Schapira, 1995). The potential pathogenicity of iron in PD is related to its ability to generate free radicals and its selective binding to new romelanin, producing Fe3+-melanin complexes. These complexes may in turn induce oxidative stress (Fornstedt et al., 1989; Ben-Shachar and Youdim, 1990, Ben-Shachar and Youdim, 1991; Youdim et al., 1993, Youdim et al., 1994; Gerlach et al., 1994). Although there is severe degeneration of dopaminergic neurons in PD, not all of them are affected in an equal manner. In the midbrain, those in the central gray substance are much less involved (5%) than those of the SN zona compacta (SNZC) (75%) (Hirsch, 1992). In PD, the variably pigmented ventrolateral part of the SNZC sustains the greatest cell loss, while the highly melanized dorsal tier is less involved but suffers the heaviest losses with aging (Fearnley and Lees, 1991). Hence, the role of neuromelanin-iron complexes in the degeneration of nigral neurons in PD is not clear. Neu romelanin, a polymer bound to lipofuscin granules is pre- sent in human and other mammalian brains. It is located in catecholaminergic neurons in the SN, locus ceruleus, and other brain stem nuclei (Bogerts, 1981; Marsden, 1983). Cytophotometric studies have shown a linear increase in the pigmentation of catecholaminergic neurons with age reaching a peak at age 60 years. Thereafter, the mean neuromelanin content of individual neurons decreases (Mann and Yates, 1974).
Recent biochemical and histochemical studies revealed a selective increase of both total Fe and Fe 3+ in the SNZC of PD brains (Dexter et al., 1989; Riederer et al., 1989: Jellinger et al., 1990; Sofic et al., 1991). Total Fe ~s elevated by 25%, and Fe 3+ is almost twice as high as in Alzheimer's disease (AD) or controls. Ferrous iron r e
mains unchanged. Total Fe in SN zona reticulata, also does not change in either PD or AD. The ratios of Fe2+/Fe 3+ in SNZC shifts from almost 2:1 in controls to 1:2 in PD brains (Sofic et al., 1988, 1991). The increase of Fe is related to the severity of degeneration of dopaminergic SNZC neurons (Riederer et al., 1989; Youdim et al., 1993). Semiquantitative histochemical studies using Perls' stain revealed a significant increase of Fe 3÷ in SNZC in both PD and PD with AD, but not in AD which was unassociated with PD. Ferric iron is mainly located in microglia, macrophages, occasional non-pigmented SN neurons, and in cells of the damaged SN that is devoid of melanized neurons (Jellinger et al., 1990; Riederer et al., 1992). No Fe 3+ was detected in melanized SN cells, probably due to its binding to a protein matrix, or in Lewy bodies (Jellinger et al., 1990). X-ray microanalysis of the midbrain in PD, but not in progressive supranuclear palsy, revealed a 3.4-fold increase of Fe in areas devoid of melanin but only a 35% increase in neuromelanin aggre- gates (Hirsch et al., 1991). In contrast, MiSssbauer spec- troscopy of SN in PD and control brains revealed equal amounts of Fe in both groups. At least 90% of this Fe resembled ferritin (Gerlach et al., 1995). Fe 2+ and/or neuromelanin-Fe constitute less than 10% of the total Fe (Bauminger et al., 1994). Low levels of neuromelanin- bound Fe were thought to represent end-stages of degener- ating dopaminergic SN neurons having lost tyrosine hy- droxylase (TH) activity (Hirsch et al., 1988) and messen- ger ribonucleic acid (mRNA) for TH (Javoy-Agid et al., 1990).
The present study addresses a series of unresolved problems concerning the role of transition metals, particu- larly Fe, in the pathogenesis and progression of PD. In view of the disparate data between histochemical studies (Jellinger et al., 1990) and X-ray microanalysis in the SN of PD brain (Hirsch et al., 1991), we performed energy dispersive X-ray microanalysis (EDX) to detect and mea- sure Fe density variations in neuromelanin, cytoplasm, neuropil, and Lewy bodies in SNZC of PD and control brains, and compared the observations to synthetic Fe- loaded melanin.
Many cytotoxic compounds that promote oxidative stress (Ben-Shachar et al., 1991b) can release Fe from its storage proteins (Thomas and Aust, 1986). The neurotoxic- ity of 6-hydroxydopamine (6-OHDA) has been linked to the release of Fe from its binding sites in ferritin (Monteiro and Winterbourne, 1989). There is experimental evidence that 6-OHDA-induced lesions of nigrostriatal DA neurons in rats are attenuated by prior intraventricular injection of desferrioxamine (Desferal), a selective Fe-chelator and free radical inhibitor. It prevents loss of dopamine (DA) and the consequent decline of its metabolites (Ben-Shachar et al., 1991a). On the other hand, direct injection of Fe into SN experimentally produces selective damage to nigrostri- atal dopamine neurons, resulting in a behavioral and bio- chemical model of PD (Ben Shachar and Youdim, 1991;

k. Kwn=i e't al .Iournal o f the Neuro ioeaat Sclence.~ t34 (Suppl.) (1995) 6 9 - 7 8 71
Sengstock et al., 1992, Sengstock et al., 1993: Arendash et al., 1994; Wesemann et al., 1994). In order to shed light on the roles of Fe and 6-OHDA in the pathogenesis and progression of PD, in vitro studies were performed to demonstrate that uncomplexed Fe 2+ is able to interact with hydrogen peroxide (H 202), promoting a Fenton-type reac- tion, and to form cytotoxic hydroxyl radicals ( -OH) (Chiueh et al., 1993; Mohanakumar et al., 1994). This oxygen radical reacts with DA to yield 5-OHDA and 6-OHDA. Chemical studies with Fe 2~, DA, 5-OHDA, or 6-OHDA using stopped-flow spectrometry (Linert et al.. 1991) revealed that 6-OHDA alone is capable of releasing Fe from strongly complexing species. Kinetic studies of the reaction between DA and dioxygen with and without the presence of Fe 3+ revealed that neither melanin alone nor Fe-containing melanin participate in this reaction, and that Fe 3+ is not a catalyst for this reaction. However, it can initiate the reaction even in acidic media (Jellinger et al., 1995; Linert et al., 1995). Since DA is in large excess. and its complex is a good indicator for Fe 3+, it can be concluded that Fe is removed from solution, obviously by inclusion into melanin. Absorption of Fe at the surface of melanin may account for only 5% of the total Fe present (Herlinger et al., 1994).
There is considerable evidence that endogenous Fe stores are also partially available for peroxidative events in tissue (Coyle and Puttfarcken, 1993). Free radicals are not the cause of such disorders, but damaged tissues react much more readily than normal tissues and thus exacerbate the primary lesion (Gutteridge, 1994; Cbiueh et al., 1994). A peroxidative injury in PD and/or AD has been demon- strated by spectrofluorometric microscopy utilizing the fluorescent reaction product when H202 reacts with cer tain dyes (Royall and Ischiropoulos, 1993).
2. Material and methods
2.1. Electron microscopy and EDA analy.sts:
Brain tissue was obtained trom 3 patients with PD aged 72-84 years, all at clinical stage V (Hoehn and Yahr. 1967), and a duration of illness between 12 and 23 years, The diagnosis was confirmed pathologically by marked loss of pigmented cells in SNZC and the presence of Lew~ bodies. One brain also showed the histological changes ol AD (Khachaturian, 1985). Two control brains from pa- tients aged 60 and 74 years were tree of CNS disease~ Autopsies were obtained within 3 -9 h alter death, and the brains were halved in the sagittal plane. One half was frozen and stored in a brain bank, the other half was fixed by immersion in 10% phosphate-buffered formalin. Tissue blocks of 2 mm 3 each were dissected from fixed brain specimens, postfixed in 4% glutaraldehyde dilution also containing 2% OsO 4, dehydrated and embedded in Spurr's epoxy resin. Semithin sections were cut with glass knive.s
(Reichert Ultracut E), and stained with a 0.05% solution of toluidine blue. The areas containing pigmented SN neurons and Lewy bodies were identified under a light microscope. Ultrathin sections (250 nm thickness) of the same tissue were contrasted with OsO 4 and mounted on carbonized copper or nylon grids.
The analyses were performed with an EDX system attached to a scanning electron microscope (EM) (Jeol 100C) which could be operated in scanning transmission (STEM) or transmission (TEM) modes, and which was equipped with a microanalysis system. Control specimens were also analysed with a Jeol 2000 FX instrument equipped with an Oxford EXL microanalysis system. The analyses were made at 80 kV acceleration voltage to minimize specimen damage. The acquisition time for spec- tra was 600-1000 sec. Probe current was 100-150 mA. A low angle detector was used to collect the spectra, and the specimen tilt angle was 37 ° . The analyses were performed with electron microprobes of 10 nm diameter on speci- mens with a thickness of 100-300 nm. Electronic control of the scanning guidance allowed exact delineation of the analyzed area to rectangular fields of 50 X 100 nm. To optimize essential peak-to-background ratios and imaging conditions, several combinations of specimen thickness were tested. It was shown that Fe radiation originating from the surrounding media could be neglected. With electron beam excitation, Fe in the specimen emitted the characteristic Fe Ka and Kb lines. A test with Fe-loaded ferritin as a standard displayed both Fe peaks and two copper peaks which arose from the grids. Spectra were collected from intracytoplasmic neuromelanin granules, the non-melanized portion of the cytoplasm, Lewy bodies in pigmented SNZC neurons, and the immediately adjacent neuropil. Identical instrument parameters were used for control cases. To check detection levels and background radiation originating from surrounding media, specimen holder or supporting grids, aggregates of synthetic melanin (Das et al., 1978), with and without Fe were analysed in a similar manner. The analysed area was documented by double exposure polaroid photography in the STEM and TEM modes (see Jellinger et al., 1992).
2.2 Fluore.~cence microscopy
Unfixed, frozen frontal cortex of a patient with PD and AD was cut in 25 /.tin-thick transverse sections with a freezing microtome (Leitz Kryostat 1720). A set of sec- tions was stained for H202 with conjugated 2',7'-dichloro- fluorescein diacetate (DCFDA) (Lambda, Fluoreszenztech- nologie, Vienna, Austria). In tissue cells, DCFDA is hy- drolyzed to 2',7'-dichlorofluorescein (DCF). Reaction of DCF with peroxides yields fluorescent 2',7'-dichlorofluo- rescein which can be detected by confocal fluorescence microscopy (CLSM) (Leica Lasertechnologie, Heidelberg, Germany). The CLSM system was equipped with an argon laser with two major emission lines at 488 nm and 514 nm.

72 E. Kienzl et al. / Journal o f the Neurological Sciences 134 (Suppl.) (1995) 69-78
2.3. Kinetic studies and chromatographic separation of 5-OHDA and 6-OHDA
Dopamine (3-hydroxytyramine) and DA hydrochloride were from Sigma Chemical Co. (St. Louis, MO, USA) and were used without further purification. The ionic strength of solution was adjusted with KC1 (Merck) to 0.100 mol dm-3 in chloride. Nitrates as background electrolytes gave identical results. 5-OHDA and 6-OHDA (as hydrochlo-
A b) a)
¢-
O o
O .,I,.,w'
100 ,, Fe K~
FeK.
Q
"q
~ee
O ~
i I ,w,,, W 3
9 11 [keY]
N,,
B b) " Si a)
~"~,~CI Ca I=~ 8 o 0 , ~ . j J"<L~" .. . K~
4 0 0 / ~o 2 0 0 I 'F' ' ,,~ 4 ,,
11 [keV] Fig. l. A: comparative presentation of x ray spectra of trace elements in Lewy body (a) and neuromelanin of nigral neuron (b) in PD brain. Energy interval examined 4800-7760 eV. Iron (Fe) is undetectable in Lewy body (a), but is present in neuromelanin (b). B: comparative presentation of x-ray spectra of synthetic dopamine-melanin (a) and synthetic Fe3+-melanin (b). The spectrum of the latter (b) strongly resembles the emission of neuromelanin in the SN of PD (Fig. 1Ab) The copper grids cause two peaks (A and B).
Fig. 2. Unfixed cryostat sections (25 p,m) of post mortem frontal cortex in PD with AD, stained for hydrogen peroxide with conjugated 2',7'-di- chlorofluorescein. A: section photographed under light microscopy; B: the same section under confocal laser fluorescence microscopy. Magnifi- cation marker = 10 ~m.
rides), were also obtained from Sigma. Synthetic melanin chloride was prepared by autoxidation according to Das et al. (1978): FeC13 w a s added to a suspension of DA (pH 7.0) under oxygen and magnetic stirring. After polymer formation for 24 h, the precipitate was centrifuged, washed with distilled water and dried. Catalase (from bovine liver) and superoxide dismutase (SOD; from bovine erythro- cytes) were supplied by Sigma.
Oxygen consumption was measured using a Clark-type electrode (EO 96, Watman Lab. Sales) connected to a corresponding processor unit (Oxi 537, from the same supplier) and a Goerz-Metrawatt x-t recorder. The pH was
E Kwnzl et al /Journal of the Neurologtcal Sciences 134 iSuppl.) (1995) 69-78 73
Table 1 Fischer's f-values of Fe density calculated from the measured x-ray spectra m human post-mortem brain tissue. PD, Parkinson's disease (2 patients of LB type, aged 76 and 82 years); CO, controls (2 cases): NM, neuromelanin: CP, cytoplasm; LB, Lewy body.
Case/structure examined PD,/NM PI)/CP. LB C O / N M C O / C P
Number of measured spectra 8 6 4 Total count rate (counts/sec) 12 42 ~ 25 16 49
Rat io ~ C Fe-Ka/ /ZC backg ......
[FeK,~+140eV] 12 31 0 9 - 1 1 0.9-1.2 Fischer's f 0 0.007 0.06 0.79 0.27-0.61 Conclusion ~ron present iron absent
5 15-19
0.9-1.0 0.11-0.57
kept constant (within _+0.003 units) with an RTS 822 titrating unit (Radiometer, Copenhagen, Denmark) coupled to a PHM 84 pH meter. The temperature was held constant within +0.1°C. The slightly acidified dopamine solution was saturated with oxygen until a constant value of the oxygen concentration was reached (usually within 10-15 min). The oxygen tube was removed, and the reaction was started by automatic addition of base (the desired pH was established in less than 2 min). The solution was stirred continuously for the correct functioning of the oxygen electrode by a circular Teflon non-vortexing magnetic
stirrer (Watman Lab-Sales). Spectra in ultraviolet (UV) and visible light were recorded by a Hitachi U-2000 spectrometer.
Additional spectra and the kinetics of dopaminochrome formation were obtained with a Tracor Northern diode array high-speed spectrophotometer, supplied by Applied Photophysics Ltd. (London, UK). These stopped-flow data v~ere recorded by mixing slightly acidified dopamine solu- tions with alkaline phosphate buffer under condition of oxygen saturation. The details of this technique have been described by Linert et al. (199t). The method allows the
AA lil fl " l
-L ,h KTNU'£ES
I •
o i t 4
O
I !
O o !
0 e~ ,,~ n .
0
I
e,l
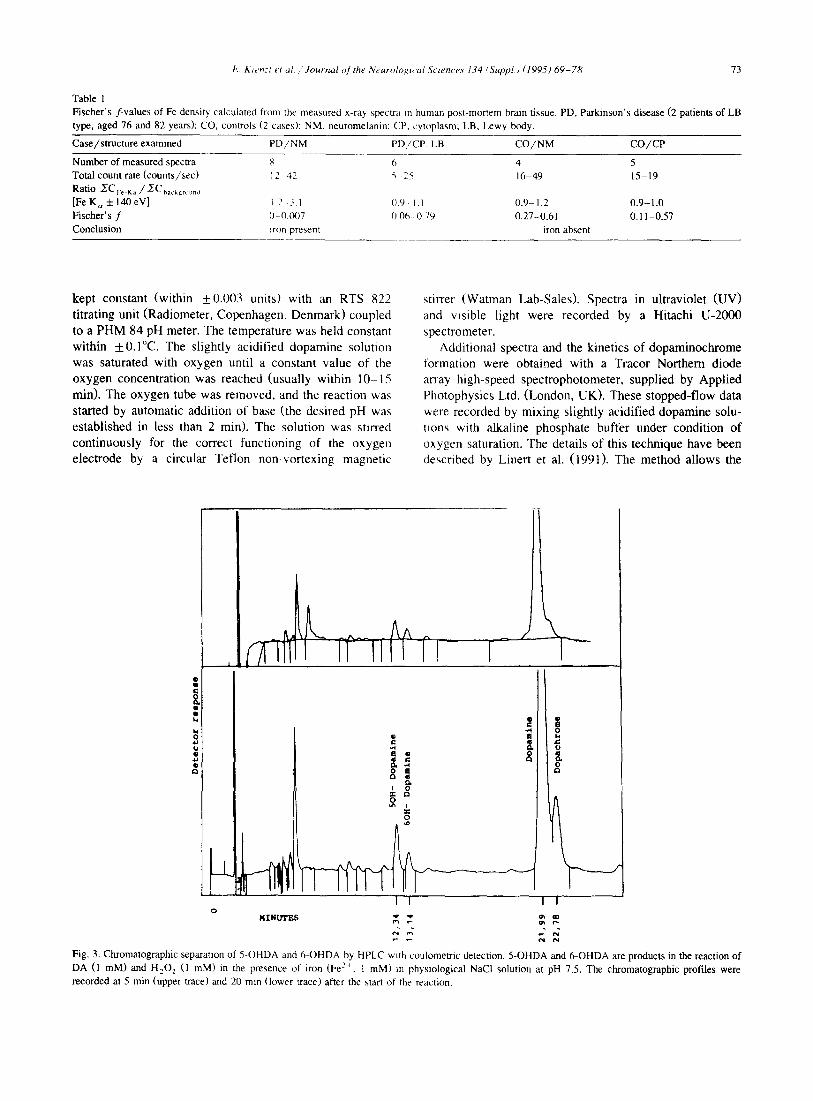
Fig. 3. Chromatographic separation of 5-OHDA and 6-OHDA by HPLC with coulometric detection. 5-OHDA and 6-OHDA are products in the reaction of DA (1 raM) and H202 (1 raM) in the presence of iron (Fe 2+, t raM) in physiological NaCI solution at pH 7.5. The chromatographic profiles were recorded at 5 rain (upper a'ace) and 20 rain (lower trace) after the start of the reaction.

74 E. Ktenzl et a l . / Journal of the Neurological Sciences 134 (Suppl.) (1995) 69-78
recording of spectra at millisecond intervals and observa tion of rapid development and decay of chemical com- pounds.
Reverse-phase high performance liquid chromatography (HPLC) was performed with a Model 5000 solvent deliv- ery module (ESA, Bedford, MA, USA) and a Coulochem 5100 A detector (ESA) with an analytical cell connected to a CI-10 recording integrator (LDC/Miiton Roy, Riviera Beach, FL, USA). Potential differences were +0.46 and -0 . 22 V, respectively. Samples were introduced through a 100-/xl injection loop. Separations were accomplished on a 250 X 4.6 mm column of Supelcosil LC-18 (5 /zm average particle size) (Supelco, Inc., College Park, PA, USA). The mobile phase contained 20 mM KH 2PO4 , 0.4raM octane- sulphonic acid and 14% methanol (v /v , pH 3). The flow- rate of the mobile phase was 1.0 ml/min. Prior to use, the mobile phase was filtered through a 0.22-/zm membrane filter (Sartorius, G6ttingen, Germany). Peak identifications were based on the elution profile of authentic standards.
Reactive iron catalyzes DA autooxidation and the Fenton reaction to produce • OH (Chiueh et al., 1993). The forma- tion of neuromelanin by oxidation of DA and polymerisa- tion of its oxidation products is a non-enzymatic process. At physiological pH, the in vitro oxidation of DA by dioxygen does not require catalytically active metal ions. The primary reaction products are dopamine-quinones which further react to dopaminochrome and H202, even in the presence of Fe ions (Linert et al., 1991).
Addition of Fe z+ and H202 to samples of DA and analysis of the resulting mixture by HPLC allowed a successful separation and identification of DA, 5-OHDA, and 6-OHDA (Fig. 3). Recent stopped-flow spectrometric studies of the electron transfer between Fe 3+ and the dopamines by anaerobic interaction of Fe 3 + with DA have demonstrated the rapid formation of the Fe-DA-complex which then, rather slowly, decomposes by means of inter- nal electron transfer, yielding Fe 2+ and dopamine-semi- quinone (Jellinger et al., 1995; Linert et al., 1995). A
3. Results
3.1. X-ray microanalysis of iron in PD brain
Energy dispersive x-ray microanalysis of neuromelanin in SN neurons in PD revealed two Fe peaks (Ka- and Kb-line) (Fig. lab) . The central core and peripheral halo of Lewy bodies (Fig. l Aa) in SNZC neurons showed extremely low Fe peaks at baseline levels. The highest amplitudes for Fe K c~ and K/3 were seen in the case of PD with AD. By contrast, analysis of neuromelanin in SN neurons of control brains revealed extremely low Fe peaks similar to those of synthetic melanin without Fe charge (Fig. 1Ba). The spectrum of Fe~+-loaded melanin resem- bled that of neuromelanin in PD (Fig. 1Bb). No Fe de- posits could be detected in the neuronal cytoplasm adja- cent to neuromelanin or Lewy bodies, and in the neuropil of SNZC in either PD or control specimens (Table 1). The statistical significance of X-ray counts in the region of the Fe lines was checked by Fisher's test (f-values), and comparisons were made with a calculated background (Statham, 1976) All regions with the strongest Fe peaks. Ka (6398 _+ 80 eV) and Kb (7657 4-80 eV) were exam- ined.
3.2. Demonstration of intrace[lular H 2 ()2
An excess of H202 was found m the post-mortem frontal cortex of a patient with PD and AD with the confocal fluorescence microscope (Fig. 2A, B).
3.3. Kinetic studies cddopamme
Dopamine is oxidized by either monoamine oxidase (MAO) or converted by auto-oxidation to generate H202.
Absorbance A
0 220
0 200
0 180
0 160
0 140
0 120
0 100
0 080
0060
0 040
0 020
0 000 Wavelength (nm)
400 480 560 640 720
AbsorDaqce
0250 i
0200 • , .
,,.,h
0 150, \ .
\, x, 0 100,
0 050
0 000,
B +Z
0 050 . . . . . . . Wavelength,(nm)
360 450 540 630 720
Fig. 4. Time dependent spectra of stopped-flow spectrometry. A: reacUon of 5-OHDA (3.0 mM) with iron (Fe ~+, 0.25 mM), pH 2.53 in 0.1 M KCI. Spectra taken after 0.625, 1.25, 2.50, 5.0, 7.50 sec (from bottom to top). Rate of electron transfer: 4.44 × 10 ~ M ~ sec- ~. B: reaction of 6-OHDA (3.0 raM) with iron (Fe 3+. 0.25 raM), pH 2.49 in 0.1 M KCI. Rate of electron transfer 600 M ~ sec- t, taken at the same time intervals as in (A). The lines indicate that no iron complex forms (compare with A).

E Ktcn. i er ,~t "Journal ¢,/the .Veur¢)ioet~ al 5ctencc~ 134 (Suppl.) (1995) 69 78 75
similar result occurs with the use 5-OHDA (Fig. 4A), although the reactions are much faster, at least in part, due to steric factors arising from the symmetry of the 5-OHDA molecule. Fig. 4B presents the results obtained when 6- OHDA is used and clearly demonstrates that no initial formation of an Fe 3 + complex is involved in the oxidation of 6-OHDA.
Metal ions such as Fe 3+ or Cu :~ have little if any influence on the rate of autooxidation. As exemplified by Fe 3+, an added metal ion becomes encapsulated by an inert, polymeric but soluble, precursor of melanin. Ulti- mately, metal ion-containing melanin is precipitated. Nei- ther melanin nor Fe-containing melanin have any effect on the rate of oxidation at neutral pH (Jellinger et al., 1995).
4. Discussion
Selective damage to neuromelanin-containing SN neu rons in PD has been related to a variety of mechanisms involving neuromelanin-Fe complexes that, under certain circumstances, may induce oxidative stress reactions and thus exert neurotoxic effects (Ben-Shachar et al., 1991b, 1995; Youdim et al., 1993). Recent biochemical and EDX studies demonstrated a significant increase of Fe ~+ m the SNZC of PD brain with significant Fe peaks only in neuromelanin granules of nigral neurons (Sofic et al.. 1991; Jellinger et al., 1992, Jellinger et al., 1993). These data were confirmed by studies using laser microscope analysis (Good et al., 1992a, b), while MiSssbauer studies of SN in PD brain showed no Fe increase compared to controls (Bauminger et al., 1994). No Fe -~ ~ was detected in Lewy bodies or in the adjacent SN neuropil of PD or control brain (Jellinger et al., 1992). In view of the antioxi dant/prooxidant properties of melanin, selective accumu- lation of Fe 3- in melanin granules of SNZC neurons in PD in the form of neuromelanin-iron complexes may be i n volved in the degeneration of dopaminergic neurons (Youdim et al., 1993, 1994).
On the other hand, melanin binds high amounts of metals and probably participates in scavenging metal-in duced free radicals (Mochizuki et al., 1993; Odh et al., 1994; Enochs et al., 1992, 1994: Korytowski et al., 1995), and it is thought to potentiate the formation of oxygen-de rived radicals under certain conditions. In vitro studies have shown that the mechanism of DA oxidation leading to polymerisation and production of melanin is indepen dent of metal ions such as Fe ~' or Cu 2' {Linert et al., 1995; Jellinger et al., 1995). At physiological pH, melanin could be considered an efficient antioxidant. Studies using electron spin resonance (ESR)-spm trapping technique confirmed the antioxidant properties of DA-melanin by lowering the one-electron reduction potential of Fe >- by complexation (Zareba el al., 1995). Once bound to melanin. Fe 3+ ions are protected against undesirable redox activa tion. Moreover, a significant generation of free hydroxyl
radicals could be detected only when melanin became saturated with large amounts of Fe ions by complexation. Synthetic DA-melanin has high-affinity (K D = 13 nm) and low-affinity binding sites (K o = 200 nM) for Fe 3+ (Ben- Shachar and Youdim, 1990). Bound Fe 3+ can be displaced only by compounds with Fe-chelating capacity.
Studies using neuronal co-culture systems, simulating the in vivo nigrostriatal system, demonstrated a neurotoxic effect of Fe on cultured neurons in a dose-dependent manner. It was more pronounced after the addition of melanin (Mochizuki et al., 1993). Neuromelanin in the SN has been shown to be a mixed melanin, possibly derived from a precursor containing sulfur (Enochs et al., 1992, Enochs et al., 1993). Moreover, oxidation products derived from both DA and cysteinyldopamine were identified con- stituents of melanin (Odh et at., 1994). Therefore, a deficit of glutathione due to exhaustion of the cysteine-glutath- lone reduction system seems to occur. Neuromelanin itself may contribute to oxidative stress when overloaded with Fe ~" by overwhelming the antioxidative defense mecha- nisms. DA significantly reduces the formation of Fe-in- duced malondialdehyde in vitro, thus suggesting that ear- lier events in cell damage are involved in DA toxicity. Recent experimental data suggest that catecholamines can cause toxicity not only by inducing oxidative stress but also possibly through direct interaction with the mitochon- drial electron transport system (Ben-Shachar et at., 1995). Hence, it is speculative to suggest that Fe may exert neurotoxic effects either by radical mechanisms or in conjunction with DA and its metabolites (Tanaka et at., 1992; Michel et al., 1992; Morris and Edwardson, 1994; Schapira, 1995). In an experimental cell culture model, Hartley et al. (1993) showed that increased intracellular Fe resulted in elevated malondialdehyde and decreased glu- tathione levels, consistent with oxidative damage, while a decrease in both complex I and IV activities contrasted with mitochondrial complex I deficiency found in the SN of human PD brain. These data and a lack of correlation between Fe levels or Fe-ferritin ratios and complex I activity, in SN in PD (Mann et al., 1994) suggest that increased Fe alone may not play a significant role in the respiratory chain dysfunction found in PD, although it may contribute to oxidative damage.
In vitro studies by stopped-flow spectrometry demon- strate (1) that the presence of Fe 2+ and H202 efficiently convert DA into the cytotoxic 6-OHDA via a Fenton-type reaction, and (12) that the oxidation of 6-OHDA by Fe 3+ proceeds without prior formation of a metal-ligand com- plex (unlike the other dopamines). In other words, the reaction of Fe ~+ with 6-OHDA is an "outer sphere" electron transfer reaction, whereas with the other catechol- amines it is an "'inner sphere" reaction (Herlinger et at., 1994). The reason for this difference is probably linked to the greater stability of the para-semiquinones and para- quinones over the ortho-species. This observation implies that 6-OHDA alone should be capable of removing Fe 3+

76 L Kienzl et al. /Journal o f the Neurological Sciences 134 (Suppl.) (1995) 69 78
(as Fe 2+) from complexes such as ferritin. It is not surpris- ing that this effect can be achieved experimentally (Monteiro and Winterbourne, 1989; Jellinger et al., 1995). Of equal importance is the observation that 6-OHDA is readily regenerated by hydroxylation of DA, thus leading to a continuous production of cytotoxic species. These hydroxylation reactions are obviously related to the occur- rence of 5-OHDA and 6-OHDA found in human urine, especially in patients on L-dopa therapy (Andrew et al., 1993). Iron mobilisation in PD could be understood as Fe release from ferritin stores by endogenously or exoge- nously derived compounds, like 6-OHDA, l-methyl-4- phenyl-l,2,3,6-tetrahydropyridine (MPTP) or paraquat, which react via an "outer sphere" electron transfer. Al- tered mechanisms for the translocation of Fe and move- ments of internalized Fe may increase Fe levels in PD brain. Following MPTP intoxication, both retrograde trans- port of Fe by transferrin receptors in brain and loss of these transferrin receptors on dopaminergic nerve termi- nals in rat striatum have been reported (Mash et al., 1993; Mochizuki et al., 1994). Free Fe, not bound to neurome- lanin, was not only found in degenerating DA cells but also in the surrounding matrix and glial cells in MPTP-in- duced hemiparkinsonian African green monkeys (Temlett et al., 1994). With a sensitive histochemical procedure, Morris and Edwardson (1994) demonstrated that non-heme Fe in PD is mainly located in glial cells and as granular deposits in the neuropil, possibly also in glia (Morris et al., 1992b). The specific neurotoxicity of 6-OHDA which induces degeneration of nigrostriatal dopaminergic neurons in animals, was studied by unilateral 6-OHDA infusion. Three weeks following infusion, a 35% increase of Fe content was seen in the ipsilateral SN (Sengstock et al., 1992, 1993; Oestreicher et al., 1994). Intranigral Fe-infu- sions into the rat SN also induced dose-dependent SN neurodegeneration. Single exposure of the SN to a modest amount of Fe induced persistent and progressive changes several months after the infusion (Sengstock et al., 1994). In post mortem SN of human PD, the levels of transferrin receptors and H-chain ferritin were significantly reduced reflecting the loss of melanized neurons (Morris et al., 1994). In contrast, L-chain ferritin was increased in SN of PD, probably associated with activated microglia that are known to respond to neuronal tissue damage (Mash et al., 1993, 1994). Increased L-chain ferritin may serve as a source of Fe for oxidative stress damage in PD.
The enzymatic and nonenzymatic cascade of DA oxida- tion, its hydroxylation to the neurotoxic 6-OHDA, and the rapid decay to semiquinone and H 2 0 2 could explain the selective degeneration of SN neurons in PD. Electron-dense reaction products of H 2 0 2 on mitochondrial membranes in damaged neurons in SN 3 days after intraventricular 6- OHDA administration in mice give evidence for alteration or oxidative inactivation of the mitochondrial electron transport chain (Ogawa et al., 1993). The relative stability of H 2 0 2 and its ability to diffuse long distances in cells
have led to the proposal that its cytotoxicity ultimately results from its decomposition to .OH, catalyzed in a Fenton-type reaction. Free redox-active metal ions in the cytoplasm cause site-specific peroxidation of lipids, pro- teins, and nucleic acids (Gutteridge, 1994; Enochs et al., 1994). Excess H 2 0 z production in degenerative nervous tissue may cause a selective decrease in complex I of the mitochondrial electron transport chain in SN and is possi- bly related to the pathogenesis of PD (Mizuno and Ohfa, 1986; Mann et al., 1994; Schapira, 1995). An analogous complex 1 defect has been observed in association with MPTP-induced parkinsonism (Mochizuki et al., 1994). The neurotoxic action of MPTP on dopaminergic neurons in primates can be blocked by N-methyl-D-aspartate (NMDA) antagonists (Bradley et al., 1993).
Disorders involving excessive superoxide radical gener- ation (and hydrogen peroxide from its dismutation), iron deposition or delocalisation of Fe complexes from Fe-con- raining proteins and metalloenzymes, limited antioxidant defences, may all be accompanied by disruption of mito- chondrial electron transport and may have severe conse- quences at excitatory postsynaptic terminals (Rice-Evans and Burdon, 1993). The demonstration of excessive H 2 0 2 in post mortem frontal cortex of a patient with PD and AD by confocal fluorescence microscopy (this report) may indicate such oxidative stress reactions. However, the ex- planation for selective vulnerability of individual neurons may come from the understanding of several factors, such as metabolic activity, excitatory amino acid input, calcium homeostasis, and the role of free-radical scavengers
The increase of a low molecular weight Fe pool is thought to be one of the first steps in the cascade creating free radical-mediated oxidative tissue damage. The search for powerful modern methodologies to understand neu- rodegenerative processes has to proceed from the molecu- lar to the cellular level, and from there to the whole CNS.
References
Andrew, R., D.G. Watson, S.A. Best, J.M. Midgley, H. Wenlong and R.K.H. Petty (1993) The determination of hydroxydopamines and other trace amines in the urine of Parkinsonian patients and normal controls. Neurochem. Res., 18:1175-1177.
Arendash, G., G. Sengstock, C. Olanow, S. Barone and A. Dunn (1994) lntranigral iron infusion as a model for Parkinson's disease. In: Woodruff, W. and Nonneman, A. (Eds.), Toxin-induced Models of Neurological Disorders, Plenum Press, New York, pp. 175-212.
Bauminger, E.R., M. Barcikowska, A. Friedman, J. Galazka-Friedman, D. Hechel and I. Nowik (1994) Does iron play a role in Parkinson's disease? Hyperfine Interact., 91: 853-857.
Ben-Shachar, D. and M.B.H. Youdim (1990) Selectivity of melanized nigra-striatal dopamine neurons to degeneration in Parkinson's dis- ease may depend on iron-melanin interaction. J. Neural Transm., 29 (Suppl.): 251-258.
Ben-Shachar, D. and M.B.H. Youdim (1991) Intranigral iron injection induces behavioral and biochemical "parkinsonism" in rats. J. Neu- rochem., 57:2133 2135.

t ; Kecncl ~'t ul Journal o f the Neuroh>g, ai gcwnces 134 ¢ Suppl.) (1995) 6 9 - 7 8 77
Ben-Shachar, D., G. Eshel, J.P.M. Finberg and M.B.H. Youdim (1991a1 The iron chelator desferrioxamine (desferal) retards 6-hydroxydopw mine-induced degeneration of nigrostriatal dopamine neurons. J. New rochem., 56: 1441-1444.
Ben-Shachar, D., P. Riederer and MB.H. Youdim (1991b) Iron-melanin interaction and lipid peroxidation: implications for Parkinson's dis ease. J. Neurochem.. 57: 1609-1614.
Ben-Shachar, D., R. Zuk and Y. Glinka (19951 Dopamme neurotoxicity inhibition of mitochondrial respiration 1. Neurochem., 64:718-~723
Bogerts, B. (19811 A brainstem atlas of catecholaminergic neurons in man using melanin as a natural marker. J. Comp. Neurol., 197: 63-80.
Bradley, M.F.B., T. Hyman and W. Koroshetz (19931 Do defects in mitochondrial energy metabolism underlie the pathology of neurode- generative diseases? Trends Neurosci., 16:125 131
Chiueh, C.C., D.L. Murphy, H. Miyake, K. Lang, PK. Tulsi and 53 Huang (19931 Hydroxal free radical ( .OH) formation reflected b~ salicylate hxdroxalation and neuromelanin: in vivo markers for oxida tive injury of nigral neurons. Ann. N. Y. Acad. Sci., 679:370 375
Chiueh, C.C., D.L. Gilbert and C.A. Cohon. eds. (19941 The neurobiol ogy of NO. and .OH. Ann. N. Y. Acad Sci., 738: 2-467.
Connor, J.R. and R.E. Fine (1986) The d+stribution of transferrin-posmve oligodendrocytes in the rat central nervous system Brain Res., 368 319-328.
Connor, J.R., S.L. Menzies, S.M. St. Martin and E.J. Mufson (19901 Cellular distribution of transferrin, ferritin and ~ron in normal and aged human brains. J. Neurosci. Res.. 27 595
Coyle, J.T. and P. Putffarcken (19931 Oxidative stress, glutamate, and neurodegenerative disorders. Science, 262: 689+ 695.
Crichton, R.R. (19911 Inorganic Biochemistry of Iron Metabolism, Ellb, Horwood Ltd.
Das, K.C., M.B. Abramson and R. Katzman (1978) Neuronal pigments: spectroscopic characterization of human brain melanin. J. Nearochem. 30: 601-605.
Dexter, D.T., P. Jenner, A.H.V. Schap~ra and C.1). Marsden (19921 Alterations in levels of iron, ferritin and other trace metals in neurode- generative disease affecting the based ganglia. Ann. Neurol, 32: 94-100.
Dwork, A.J., E.A. Schon and J. Herbert (19881 Non identical distribution of transferrin and iron in the human brain. Neuroscience, 27:333 345
Enochs, W.S., M.J. Nilges and HM. Swartz (19921 Purified human neuromelanin, synthetic dopamine melanin as a potential model pig ment, and the normal human substantia nigra: characterization b~ electron paramagnetic resonance spectroscopy 1. Neurochem.. 61 68 79.
Enochs, W.S., M.J. Nilges and tt.M. Swartz t t9931 Purified Human neuromelanin, synthetic dopamme melanin as a potential model pig ment, and the normal human substantia-mgra-characterization b~ electron paramagnetic resonance spectrometry J. Neurochem.. 61 68-79.
Enochs, W.S., T. Sarna, L. Zecca, P.A Rileyand and II..M. Swarle (1994) The roles of neuromelanin, binding of metal ions, and oxida tive cytotoxicity in the pathogenesis of Parkinson's disease: a hypoth. esis. J. Neural Transm. (P-D Sect.). 7 :83 100.
Fearnley, J. and A.J. Lees (19911 Ageing and Parkinson's disease Substantia nigra regional selectivity. Brain, 114:2289 2301.
Fornstedt, B., A. Brun, E. Rosengren and A+ Carlsson (1989) The apparent autoxidation rate of catechols m dopannne-rich regions ol human brains increases with the degree of depigmentation of substan. tia nigra. J. Neural Transm. (P-D SectL 1: 279+ 295.
Gerlach, M., D. Ben-Shachar, P. Riederer and M.B.H. Youdim (19941 Altered brain metabolism of iron as a cause of neurrxtegeneranve diseases? J. Neurochem., 63:793 806
Gerlach, M., A.X. Trautwein, L. Zecca. M.B.t t Youdin and P. Riederer (1995) Moessbauer spectroscopic studies of purified human neurome lanin isolated from the substantia nigra. J Neur(x:hem., 65:923 926.
Gocht, A. and J. LiShler (1990) Changes in glial cell markers in rccenl
and old demyelinated lesions in central pontine myelinosis. Acta Neuropathol., 80: 46-47.
Good, P.F., D.P. Pel, L.M. Bierer and J. Schmeidler (1992a) Selective accumulation of aluminium and iron in the neurofibrillary tangles of Alzheimer's disease: a laser microprobe (LAMMA) study. Ann. Neurol., 31: 286-292.
Good, P., C. Olanow and D. Perl (1992b) Neuromelanin-containing neurons of the substantia nigra accumulate iron and aluminium in Parkinson's disease: a LAMMA study. Brain Res., 593: 343-346.
Gutteridge, J.M.C. (1994) Hydroxyl radicals, iron, oxidative stress, and neurodegeneration. Ann. N. Y. Acad. Sci., 738: 201-213.
ttartley, A., J.J. Cooper and A.H.V. Schapira (1993) Iron induced oxidative stress and mitochondrial dysfunction: relevance to Parkin- son's disea~. Brain Res., 627: 349-353.
tterlinger, E., R.F. Jameson and W. Linert (1994) The spontaneous autoxidation of dopamine. J. Chem. Soc. Perkin Trans., It: 259-263.
Itill, J.M+ and R.C. Switzer (1984) The regional distribution and cellular Iocalisation of iron in the rat brain. Neuroscience, 11: 595-603.
Hirsch, E.C. (1992) Why are nigral catecholaminergic neurons more vulnerable than other cells in Parkinson's disease? Ann. Neurol., 32:
88 93. tlirsch, E.C., A..M. Graybiel and Y. Agid (1988) Melanized dopaminer-
gic neurons are differently susceptible to degeneration in Parkinson's disease. Nature, 334: 345-348.
ttirsch. E.C., J.P. Brandel, P. Galle, F. Javoy-Agid and Y. Agid (1991) Iron and aluminium increase in the substantia nigra of patients with Parkinson's disease: an x-ray microanalysis. J. Neurochem., 56: 446- 451.
li(yehn. M.M., and M. Yahr (1967) Parkinsonism. Onset, progression, and mortality. Neurology, 17: 427-442.
Ja,,oy -Agid, F.. E.C. Hirsch, S. Dumas et al. (1990) Decreased tyrosine hydroxylase messenger RNA in the surviving dopamine neurons of the substantia nigra in Parkinson's disease: an in situ hybridization study. Neuroscience, 38: 245-253.
lcllinger, K. ( 1991 ) Pathology of Parkinson's disease. Changes other than the nigrostriatal pathway. Mol. Chem. Neuropathol., 14: 153-197.
Jcllinger, K. and E. Kienzl (1993) Iron deposits in brain disorders. In: Riederer, P. and M.B.H. Youdim (Eds.), Iron in Central Nervous System Disorders, Springer-Verlag, Vienna-New York, pp. 19-36.
Jellinger, K.. W. Paulus, J. Grundke-lqbal, P. Riederer and M.B.H. Youdim (1990) Brain iron and ferritin in Parkinson's and Alzheimer's diseases. J. Neural Transm. (PD-D Sect.), 2: 327-340.
Jellinger, K.. E. Kienzl, G. Rumplmair, W. Paulus, P. Riederer, H. Stachelberger, D. Ben-Shachar and M.B.H. Youdim (1992) Iron- melanin complexes in substantia nigra of parkinsonian brains: an X-ray microanalysis. J. Neurochem., 59:1168-1172.
Jellinger, K., E. Kienzl, G. Rumplmair, W. Paulus, P. Riederer, E. Sofic, D. Ben-Shachar and M.B.H. Youdim (1993) Iron and ferritin in substantia nigra in Parkinson's disease. Adv. Neurol., 60: 267-272.
Jellinger, K., W. Linert, E. Kienz], E. Herlinger and M.B.H. Youdim t19951 Chemical evidence for 6-hydroxydopamine to be an endoge- nous toxic factor in the pathogenesis of Parkinson's disease. J. Neural. Transm. (Suppl.)+ in press
Kaneko. Y.. -I'. Kimmoto, I. Tateishi and K. Yamaguchi (1989) Ferritin immunohistochemistry as a marker of microglia. Acta Neuropathol., ~9: 129-136.
Khachaturian. Z.S. (1985) Diagnosis of Alzheimer's disease. Arch. Neu- rol. 42:1097-1105.
Koeppen, A.tt. and M.P. Dentinger (1988) Brain haemosiderin and superficial siderosis of the central nervous system. J. Neuropathol. Exp. Neurol.. 47:249-251.
Koryto~ski. W., T. Sama and M. Zareba (1995) Antioxidant action of neuromelanin - The mechanism of inhibitory effect on lipid peroxida- lion. Arch. Biochem. Biophys., 319: 142-148.
l~mert, W., RF. Jameson and E. Herlinger (1991) Complex formation followed by internal electron transfer: the reaction between L-dopa and ~ron (liD. lnorg. Chim. Acta, 187:239 247.

78 E Kienzl et al /Journal of the Neurological Sciences 1.74 (Suppl.) (1995) 69 78
Linert, W., E. Herlinger, R.F. Jameson, E. Kienzl and K. Jellinger (1995) Transition-metal catalysed oxidation processes of catecholamines: a key to the understanding or Parkinson's disease'? In: G. Ondrajovlc and A. Sirota (Eds.), Current Trends in Coordination Chemistry, Slovak Technical Univ. Press, Bratislava, pp. 313-318.
Mann, D.M.A. and P.O. Yates (1974) Lipoprotein pigments. Their rela tionship to aging in the human nervous system, lI. The melanin content of pigmented nerve cells. Brain, 97:489 498.
Mann, V.M., J.M. Cooper, S.S. Daniel, P. Jenner, C.D. Marsden and A.H.V. Schapira (1994) Complex l, iron. and ferritin in Parkinson's disease substantia nigra. Ann. Neurol., 36:876 881.
Marsden, C.D. (1983) Neuromelanin and Parkinson's disease. J. Neural Transm., 19:121 141.
Mash, D.C., J. Singer, J. Pablo, M. Basile. J. Bruce and W.J. Wemer (1993) Iron storage and transport marker~ in Parkinson's disease and MPTP-treated mice In: Riederer, P., and M.B.H. Youdim (Eds.), Iron in Central Nervous System Disorders, Springer-Verlag, Vienna New York, pp. 103-116.
Mash, D.C., J. Singer, J. Pablo. J. Bruce and W.J. Weiner (1994). Differential regulation of iron storage and transport markers in Parkin son's disease. Neurology, 44 (Suppl. 2): 269.
Michel, P.D., S. Vyas and Y. Agid (1992) Toxic effects of iron fol cultured mesencephalic dopaminergic neurones derived fi'om rat em bryonic brains. J. Neurochem., 59 :118- 127.
Mizuno, Y. and K. Ohfa (1986) Regional distribution of thiobarbituric acid-reactive products, activities of enzymes regulating the metabolism of oxygen free radicals, and some of the related enzymes in aduh and aged rat brains. J. Neurochem.. '46: 1344-1352.
Mochizuki, H., K. Nishi and Y. Mizuno (1993) Iron Melanin complex is toxic to dopaminergic neurons in a nigrostriatal co-culture. Neurode generation, 2:1 7.
Mochizuki, H., H. Imai, K. Endo et al.. (1994) Iron accumulation in the substantia nigra of 1 methyl 4 phenyl 1,2,3,6-tetrahydropyridmc (MPTP) induced hemiparkinsonism in monkeys. Neurosci. Lett., 168 251-253.
Mohanakumar, K.P., A. De Barlolomeis. R-M. Wu, K.J. Yeh, L.M. Sternberger, S.-Y. Peng, D.L. Murphy and C.C. Chiueh (1994) Fer rous-citrate complex and nigral degeneration; evidence for fi-ee-radi- cal formation and lipid peroxidation. Ann. N. Y. Acad. Sci.. 7~g~ 392-399.
MNlgard, K., M. Jacobsen, G. Krag-Jacobsen, P. Praetorius-Clausen and N.R. Saunders. (1979) Immunohistochemical evidence for an intra cellular localisation of plasma proteins in human foetal choroid plexus and brain. Neurosci. Lett. 14: 85~ 87.
Monteiro, H.P. and C.C. Winterbourne (1989) 6-Hydroxydopamine re leases iron from ferritin and promotes ferritin-dependent lipid per oxidation. Biochem. Pharmacol., 38 :4144. 4182.
Morris, C.M. and J.A. Edwardson (1994) Iron histochemistry o! the substantia nigra in Parkinson's disease. Neurodegeration, 3:277 282.
Morris, C.M., J.M. Candy, A. Keith, R.G.L. Pullen and J.A. Edwardson (1992a) Brain iron homeostasis. J. lnorg. Biochem., 47:257 265.
Morris, C.M, J.M. Candy, A.E. Oakley, C.A. Bloxham and J.A. Edward- son (1992b) Histochemical distribution of non-haem iron in the human brain. Acta Anat. Basel, 144:235-257
Morris, C.M., J.M. Candy, S. Omas. C.A. Bloxham and J.A. Edwardson (1994) Transferrin receptors in the parkinsonian midbrain. Neu ropathol. Appl. Neurobiol., 20:468 472.
Odh, G., R. Carstam, J. Paulson, A. Wittbjer, E. Rosengren and H Rorsman (1994) Neuromelanin of the human substantia mgra: a mixed-type melanin. J. Neurochem., 62:2030 2036.
Oestreicher, E., G.J. Sengstock, P. Riederer. C.W. Olanow, A.J. l)uan G.W. and Arendash (1994) I)egeneration of nigrostriatal dopammer gic neurons increases iron within the substantia nigra: a histochem~¢al and neurochemical study. Brain Res , 660: 8- 18.
Ogawa, N., R. Edarnatsu, K Mizukawa, M. Asanuma, M. Kohno and -\
Mori (1993) Degeneration and free radicals. Possible participation of levodopa. Adv. Neurol., 60: 242-250.
Rice-Evans, C. and R. Burdon (1993) Free radical- and lipid interactions and their pathological consequences. Prog. Lipid Res., 32:71-110.
Riederer, P., E. Sofic, W.D. Rausch, B. Schmidt, G.P. Reynolds, K. Jellinger and M.B.H. Youdim (1989) Transition metals, ferritin, glu- tathione, and ascorbic acid in parkinsonian brains. J. Neurochem., 52: 515-520.
Riederer, P., A. Dirt, M. Goetz, E. Sofic, K. Jellinger and M.B.H. Youdim (1992) Distribution of iron in different brain regions and subcellular compartments in Parkinson's disease. Ann. Neurol., 32: 101- 104.
Riederer, P. and M.B.H. Youdim (Eds.) (1993) Iron in Central Nervous System Disorders, Springer-Verlag, Vienna-New York.
Royall, J.A. and H. Ischiropoulos (1993) Evaluation of 2',7'-dichlorofluo- rescein and dihydrorhodamine 123 as fluorescent probes for intra- cellular H :02 in cultured endothelial cells. Arch. Biochem. Biophys., 302: 348-355.
Schapira, A.H.V. (1995) Oxidative stress in Parkinson's disease. Neu- ropathol. Appl. Neurobiol. 21: 3-9.
Sengstock, G.J., C.W. Olanow, A.J. Dunn and G.W. Arendash (1992) Iron induces degeneration of nigrastriatal neurons. Brain Res. Bull., 645- 649.
Sengstock, G., C. Olanow, R. Menzies, A. Dunn and G. Arendash (1993) Infusion of iron into the rat substantia nigra; nigral pathology and dose-dependent loss of striatal dopaminergic markers. J. Neurosci. Res., 35: 67-82.
Sengstock, G.J., C.W. Olanow, A.J. Dunn, S. Barone and G.W. Arendash (1994) Striatal dopaminergic markers, nigral volume, and rotational behavior following iron infusion into the rat substantia nigra. Exp. Neurol., 130: 82-94.
Sofic, E.. P. Riederer, H. Heinsen, H. Beckmann, G.P. Reynolds, G. Hebenstreit and M.B.M. Youdim (1988) Increased iron (11I) and total iron content in post mortem substantia nigra of parkinsonian brain, J. Neural Transm., 74: 199-205.
Sofic, E., W. Paulus, K. Jellinger, P. Riederer and M.B.M. Youdim ( 1991 ) Selective increase of iron in substantia nigra zona compacta of parkinsonian brain. J. Neurochem., 56: 978-982.
Statham, P.H. (1976) A comparative study of techniques for quantitative analysis of the x-ray spectra obtained with a Si (Li) detector. X-ray Spectrom., 5: 16-28.
Tanaka, M., A. Sotomatsu, M. Kanai and S. Hirai (1992) Combined histochemical and biochemical demonstration of nigral vulnerability to lipid peroxidation induced by DOPA and iron. Neurosci. Lett., 140: 42- 46.
Temlett, J.A., J.P. Landsberg, F. Watt and G.W. Grimme (1994) In- creased iron in the substantia compacta of the MPTP-lesioned hemi- parkinsonian African green monkey: evidence from proton micro- probe elemental microanatysis. J. Neurochem., 62: 134-146.
Thomas, C.E. and D. Aust (1986) Reductive release of iron from ferritin by cation free radicals or paraquat and other bipyridyls. J. Biol. Chem., 261: 13064-13070.
Wesemann, W., S. Blaschka, M. Salbach, C. Grote, H.W. Clement and P. Riederer (1994) lntranigral injected iron progressively reduces striatal dopamine metabolism. J. Neural Transm. (PD-D-Sect.), 8: 209-214.
Youdim, M.B.H., D. Ben-Shachar and P. Riederer (1993) The possible role of iron in the etiopathology of Parkinson's disease. Movement Disord., 8: 1-12.
Youdim, M.B.H., D. Ben-Shachar and P. Riederer (1994) The enigma of neuromelanin in Parkinson's disease substantia nigra. J. Neural Transm., 43 (Suppl.): 113-122.
Zareba, M., A. Bober, W. Korytowski, L. Zeccha and T. Sarna (1995) The effect of synthetic neuromelanin on yield of free hydroxyl radical generated in model systems. Biochim. Biophys. Acta, 1271: 343-348.





























