Embed Size (px)
Citation preview

Proc. Nat. Acad. Sci. USAVol. 70, No. 2, pp. 498-502, February 1973
The Labyrinthine Sense Organs of the Frog(otic labyrinth/ear/amphibia)
ERNEST GLEN WEVER
Auditory Research Laboratories, Princeton University, Princeton, New Jersey 08540
Contributed by Ernest Glen Wever, December 6, 1972
ABSTRACT A study of the macula and crista organsof the frog's labyrinth with the use of an improved methodof histological preparation has shown these endings to bemore complex than heretofore believed. The structure ly-ing over the layer of sensory and supporting cells is not asingle "gelatinous" body as commonly described, but con-sists of two distinct layers with separate functions.In all these endings-macula sacculi, macula utriculi,
macula lagenae, and the cristae of the three semicircularcanals-there is a special tectorial structure that lies overthe cellular surface and makes the connections to the cil-iary tufts of the hair cells. It has the general form of a re-ticulum, though in the saccule it is somewhat elaborated.A preliminary study of other vertebrates indicates that
this tectorial reticulum is present in all the labyrinthineendings throughout the series from fishes to mammals.Consideration is given to the possible advantages of this
special tectorial structure in the stimulation process.
Three types of sensory endings are present in the otic laby-rinth. These are the papilla acustica (in advanced forms calledthe cochlea), the macula acustica, and the crista acustica.The papilla acustica is lacking in the fishes, and makes itsfirst appearance in the amphibians, and in most species of thisgroup there are two types, the papilla basilaris and the papillaamphibiorum; in all higher animals there is one type only.Three principal forms of the macula occur, the macula sacculi,macula utriculi, and macula lagenae, though the last-namedis lacking in the higher mammals and in a few other species.A macula neglecta is present in several forms, but is of smallsize and uncertain function, and will not be considered furtherhere. A crista is present uniformly in the ampulla of each ofthe three semicircular canals from fishes to mammals.Our understanding of the general structure of these organs
arose out of the studies of the early anatomists, and perhapsreached its clearest formulations in the descriptions of Retziusin 1881 and 1884 (1). Extensive investigations performedsince, with the aid of electron microscopy, have added a wealthof fine details but yet have brought no important change inthe basic picture.These endings continue to be described essentially in the
language of Retzius as consisting of two layers of tissue, onean array of hair cells held in a framework of supporting cellsand the other a covering layer usually characterized as gelat-inous and varying greatly in form in the three types of end-ings.The object of the present report is, first, to show that in all
these endings the covering of the cellular elements consists oftwo distinct layers of tissue and not a single one, and then toconsider the particular functions of these separate structures.The observations are an offshoot of a study of the auditory
papillae of the frog, a study that necessitated the develop-
ment of improved methods for the histological treatment ofamphibian ears. These observations, described in detail else-where*, showed that the auditory papillae always include aspecial tectorial structure lying close over the ciliated ends ofthe hair cells, making connections with their ciliary tufts, andclearly involved in the stimulation process. This situation ledto the question whether some corresponding structures mightbe present in the nonauditory endings as well. These struc-tqres were looked for, and found to be present.
In all four types of labyrinthine endings, in the utricularmacula, lagenar macula, saccular macula, and the cristae ofthe semicircular canals, there is a tectorial structure in theform of a fiber network, or for the saccular macula a form thatseems to be an elaboration of such a network. This structurealways lies above the surface membrane that represents theexposed ends of the hair cells and supporting cells, and hassuperimposed on it a structure that for the macular endingsis an otolithic mass and for the cristae is a further tectorialbody known as the cupula. The descriptions below referspecifically to Rana pipiens.
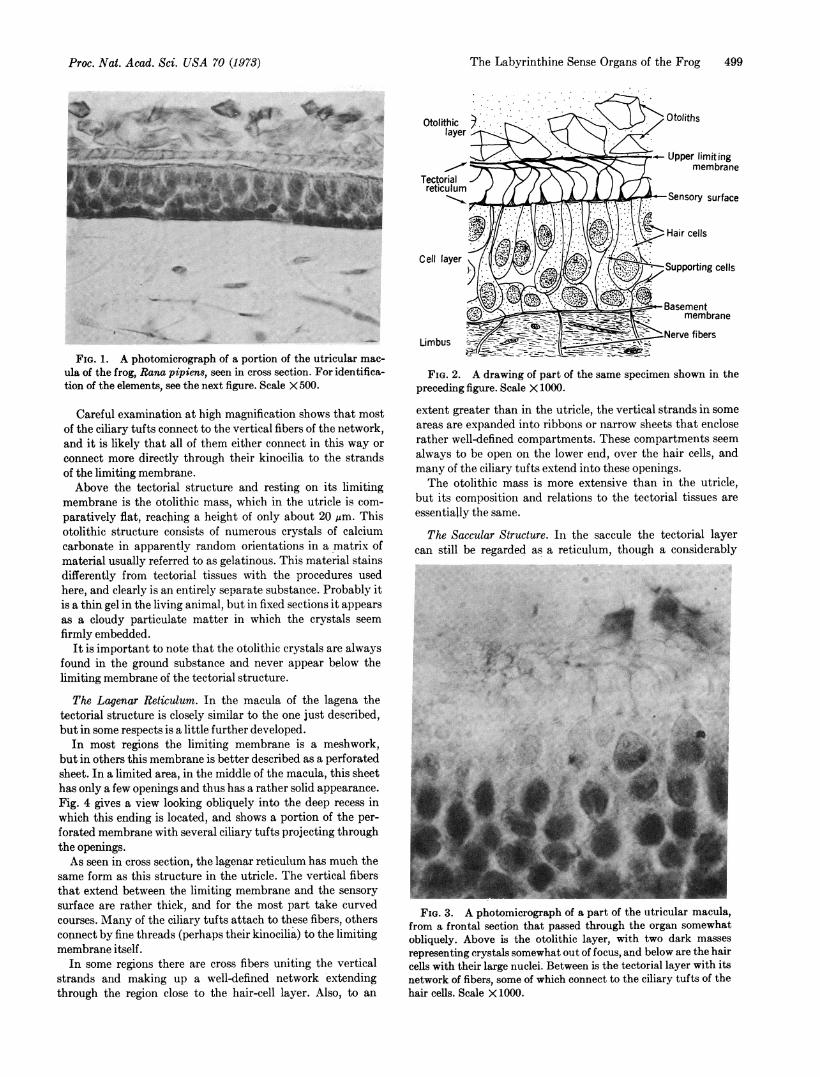
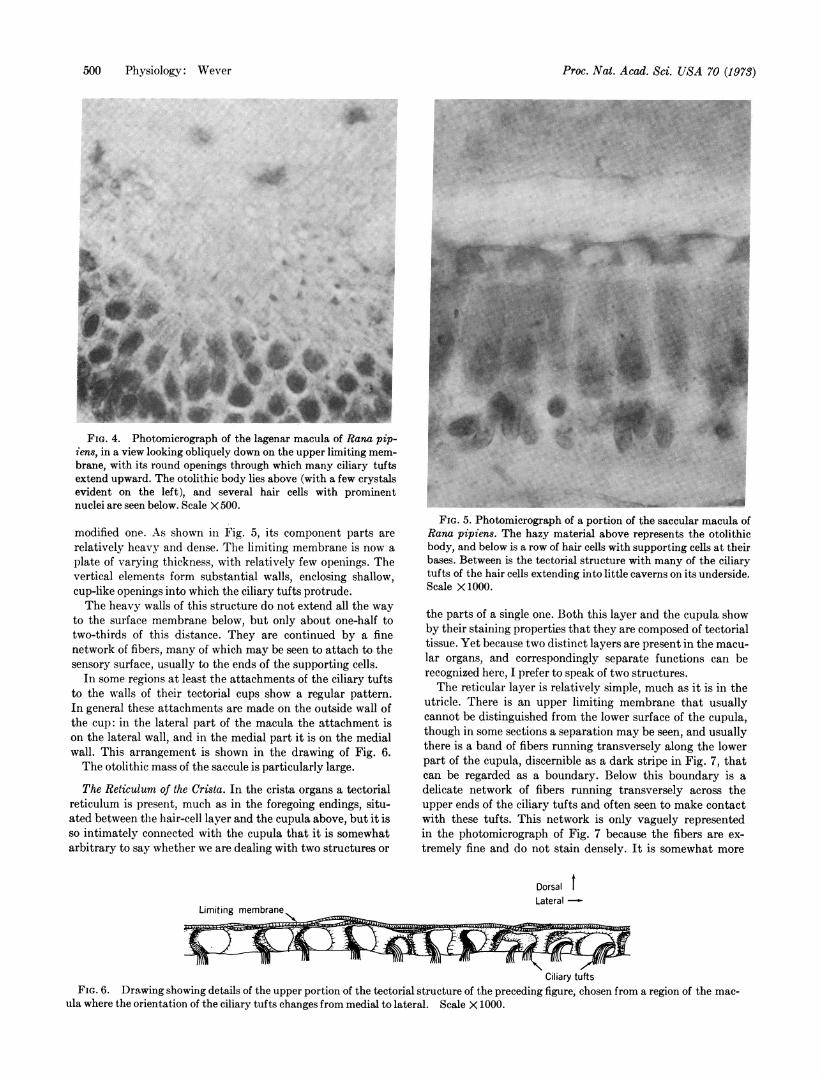
The Utricular Reticulum. The tectorial structure lying overthe utricular hair cells varies somewhat in its different re-gions. In some places, especially toward the edges of the mac-ula, there is only a simple network, as shown in the photomi-crograph of Fig. 1 and in the drawing of Fig. 2.
This network consists of an upper limiting membrane andan array of more or less vertical fibers that extend to thesensory surface below and sometimes at least attach to thissurface membrane. The limiting membrane is a loose networkwhose round, oval, and sometimes polygonal openings canbest be seen when the view is from one side. Fig. 3 shows thismembrane obliquely, and both the upper limiting network andthe vertical fibers can be seen. As will be noted, some of thesevertical fibers make connections with the tips of the ciliarytufts of the hair cells.
In some regions, and especially in the middle of the macula,the structure is more complex. The vertical fibers are oftenconnected to one another by short and usually fine transversestrands, which sometimes branch and interconnect so that thewhole constitutes an open spongework of spaces in a three-dimensional lattice.
In some places also the vertical fibers are expanded as nar-
row ribbons or even as small, irregular sheets, and the spacesof the lattice then take on the appearance of shallow recesses
or canals with imperfect walls.
* Wever, E. G., "The ear and hearing in the frog, Rana pipiens,"to appear.
498
The Labyrinthine Sense Organs of the Frog 499
. ~A.
FIG. 1. A photomicrograph of a portion of the utricular mac-ula of the frog, Rana pipiens, seen in cross section. For identifica-tion of the elements, see the next figure. Scale X 500.
Careful examination at high magnification shows that mostof the ciliary tufts connect to the vertical fibers of the network,and it is likely that all of them either connect in this way orconnect more directly through their kinocilia to the strandsof the limiting membrane.Above the tectorial structure and resting on its limiting
membrane is the otolithic mass, which in the utricle is com-paratively flat, reaching a height of only about 20 Mm. Thisotolithic structure consists of numerous crystals of calciumcarbonate in apparently random orientations in a matrix ofmaterial usually referred to as gelatinous. This material stainsdifferently from tectorial tissues with the procedures usedhere, and clearly is an entirely separate substance. Probably itis a thin gel in the living animal, but in fixed sections it appearsas a cloudy particulate matter in which the crystals seemfirmly embedded.
It is important to note that the otolithic crystals are alwaysfound in the ground substance and never appear below thelimiting membrane of the tectorial structure.
The Lagenar Reticulum. In the macula of the lagena thetectorial structure is closely similar to the one just described,but in some respects is a little further developed.
In most regions the limiting membrane is a meshwork,but in others this membrane is better described as a perforatedsheet. In a limited area, in the middle of the macula, this sheethas only a few openings and thus has a rather solid appearance.Fig. 4 gives a view looking obliquely into the deep recess inwhich this ending is located, and shows a portion of the per-forated membrane with several ciliary tufts projecting throughthe openings.As seen in cross section, the lagenar reticulum has much the
same form as this structure in the utricle. The vertical fibersthat extend between the limiting membrane and the sensorysurface are rather thick, and for the most part take curvedcourses. Many of the ciliary tufts attach to these fibers, othersconnect by fine threads (perhaps their kinocilia) to the limitingmembrane itself.
In some regions there are cross fibers uniting the verticalstrands and making up a well-defined network extendingthrough the region close to the hair-cell layer. Also, to an
FIG. 2. A drawing of part of the same specimen shown in thepreceding figure. Scale X 1000.
extent greater than in the utricle, the vertical strands in someareas are expanded into ribbons or narrow sheets that encloserather well-defined compartments. These compartments seemalways to be open on the lower end, over the hair cells, andmany of the ciliary tufts extend into these openings.The otolithic mass is more extensive than in the utricle,
but its composition and relations to the tectorial tissues areessentially the same.
The Saccular Structure. In the saccule the tectorial layercan still be regarded as a reticulum, though a considerably
.. :0 _'Y
S. 'eme Of i ifl wijP}s . F g ;F!-Woo> 4s adz 'Up OF Y Ft1* *-* e } * I* _., .i .:
.. .. ; *:# 9 * :9 ,r
.0t
:' l.
FIG. 3. A photomicrograph of a part of the utricular macula,from a frontal section that passed through the organ somewhatobliquely. Above is the otolithic layer, with two dark masses
representing crystals somewhat out of focus, and below are the haircells with their large nuclei. Between is the tectorial layer with itsnetwork of fibers, some of which connect to the ciliary tufts of thehair cells. Scale X 1000.
Proc. Nat. Acad. Sci. USA 70 (1973)
Proc. Nat. Acad. Sci. USA 70 (1973)
.~~
l en
I'w w9*__MFIG. 4. Photomicrograph of the lagenar macula of Rana pip-
iens, in a view looking obliquely down on the upper limiting mem-brane, with its round openings through which many ciliary tuftsextend upward. The otolithic body lies above (with a few crystalsevident on the left), and several hair cells with prominentnuclei are seen below. Scale X 500.
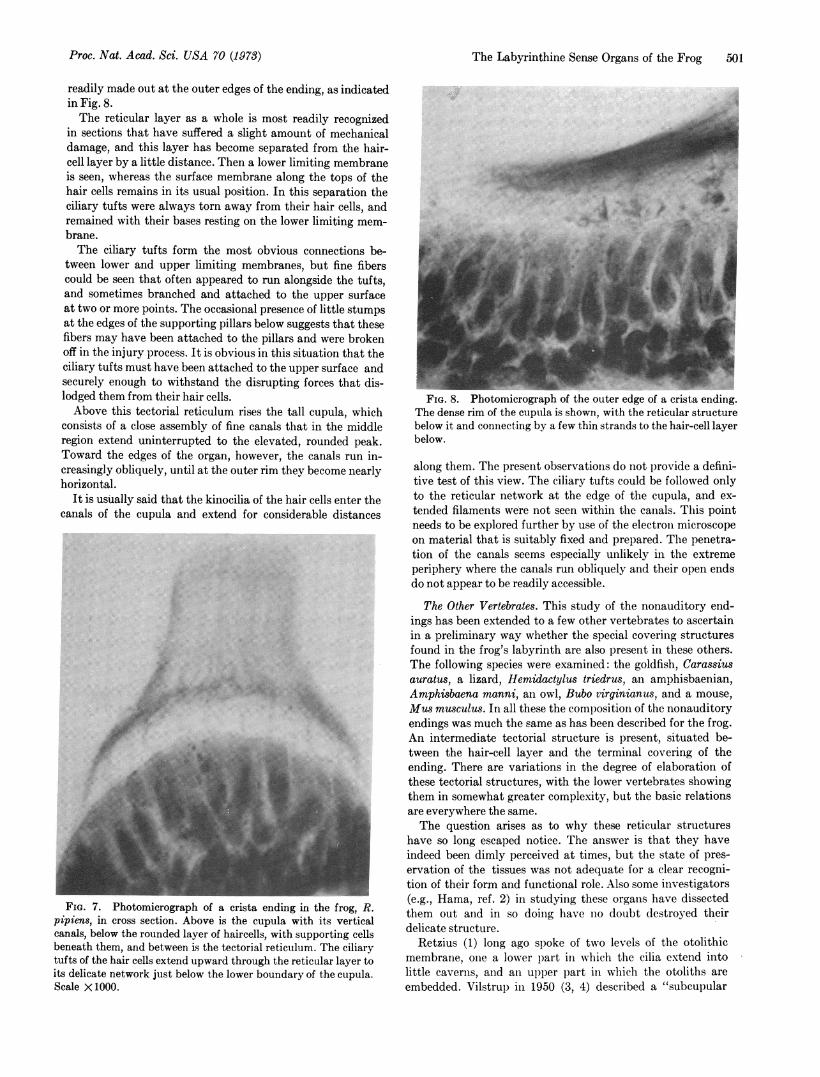
modified one. As shown in Fig. 5, its component parts arerelatively heavy and dense. The limiting membrane is now aplate of varying thickness, with relatively few openings. Thevertical elements form substantial walls, enclosing shallow,cup-like openings into which the ciliary tufts protrude.The heavy walls of this structure do not extend all the way
to the surface membrane below, but only about one-half totwo-thirds of this distance. They are continued by a finenetwork of fibers, many of which may be seen to attach to thesensory surface, usually to the ends of the supporting cells.
In some regions at least the attachments of the ciliary tuftsto the walls of their tectorial cups show a regular pattern.In general these attachments are made on the outside wall ofthe cup: in the lateral part of the macula the attachment ison the lateral wall, and in the medial part it is on the medialwall. This arrangement is shown in the drawing of Fig. 6.The otolithic mass of the saccule is particularly large.
The Reticulum of the Crista. In the crista organs a tectorialreticulum is present, much as in the foregoing endings, situ-ated between the hair-cell layer and the cupula above, but it isso intimately connected with the cupula that it is somewhatarbitrary to say whether we are dealing with two structures or
FIG. 5. Photomicrograph of a portion of the saccular macula ofRana pipiens. The hazy material above represents the otolithicbody, and below is a row of hair cells with supporting cells at theirbases. Between is the tectorial structure with many of the ciliarytufts of the hair cells extending into little caverns on its underside.Scale X1000.
the parts of a single one. Both this layer and the cupula showby their staining properties that they are composed of tectorialtissue. Yet because two distinct layers are present in the macu-lar organs, and correspondingly separate functions can berecognized here, I prefer to speak of two structures.The reticular layer is relatively simple, much as it is in the
utricle. There is an upper limiting membrane that usuallycannot be distinguished from the lower surface of the cupula,though in some sections a separation may be seen, and usuallythere is a band of fibers running transversely along the lowerpart of the cupula, discernible as a dark stripe in Fig. 7, thatcan be regarded as a boundary. Below this boundary is adelicate network of fibers running transversely across theupper ends of the ciliary tufts and often seen to make contactwith these tufts. This network is only vaguely representedin the photomicrograph of Fig. 7 because the fibers are ex-tremely fine and do not stain densely. It is somewhat more
Dorsal tLateral
Limiting membrane
Ciliary tuftsFIG. 6. Drawing showing details of the upper portion of the tectorial structure of the preceding figure, chosen from a region of the mac-
ula where the orientation of the ciliary tufts changes from medial to lateral. Scale X 1000.
500 Physiology: Wever
..:I,'i..
The Labyrinthine Sense Organs of the Frog 501
readily made out at the outer edges of the ending, as indicatedin Fig. 8.The reticular layer as a whole is most readily recognized
in sections that have suffered a slight amount of mechanicaldamage, and this layer has become separated from the hair-cell layer by a little distance. Then a lower limiting membraneis seen, whereas the surface membrane along the tops of thehair cells remains in its usual position. In this separation theciliary tufts were always torn away from their hair cells, andremained with their bases resting on the lower limiting mem-brane.The ciliary tufts form the most obvious connections be-
tween lower and upper limiting membranes, but fine fiberscould be seen that often appeared to run alongside the tufts,and sometimes branched and attached to the upper surfaceat two or more points. The occasional presence of little stumpsat the edges of the supporting pillars below suggests that thesefibers may have been attached to the pillars and were brokenoff in the injury process. It is obvious in this situation that theciliary tufts must have been attached to the upper surface andsecurely enough to withstand the disrupting forces that dis-lodged them from their hair cells.Above this tectorial reticulum rises the tall cupula, which
consists of a close assembly of fine canals that in the middleregion extend uninterrupted to the elevated, rounded peak.Toward the edges of the organ, however, the canals run in-creasingly obliquely, until at the outer rim they become nearlyhorizontal.
It is usually said that the kinocilia of the hair cells enter thecanals of the cupula and extend for considerable distances
..
FIG. 7. Photomicrograph of a crista ending in the frog, R.pipiens, in cross section. Above is the cupula with its verticalcanals, below the rounded layer of haircells, with supporting cellsbeneath them, and between is the tectorial reticulum. The ciliarytufts of the hair cells extend upward through the reticular layer toits delicate network just below the lower boundary of the cupula.Scale X1000.
FIG. 8. Photomicrograph of the outer edge of a crista ending.The dense rim of the cupula is shown, with the reticular structurebelow it and connecting by a few thin strands to the hair-cell layerbelow.
along them. The present observations do not provide a defini-tive test of this view. The ciliary tufts could be followed onlyto the reticular network at the edge of the cupula, and ex-tended filaments were not seen within the canals. This pointneeds to be explored further by use of the electron microscopeon material that is suitably fixed and prepared. The penetra-tion of the canals seems especially unlikely in the extremeperiphery where the canals run obliquely and their open endsdo not appear to be readily accessible.
The Other Vertebrates. This study of the nonauditory end-ings has been extended to a few other vertebrates to ascertainin a preliminary way whether the special covering structuresfound in the frog's labyrinth are also present in these others.The following species were examined: the goldfish, Carassiusauratus, a lizard, Hemidactylus triedrus, an amphisbaenian,Amphisbaena manni, an owl, Bubo virginianus, and a mouse,Mus musculus. In all these the composition of the nonauditoryendings was much the same as has been described for the frog.An intermediate tectorial structure is present, situated be-tween the hair-cell layer and the terminal covering of theending. There are variations in the degree of elaboration ofthese tectorial structures, with the lower vertebrates showingthem in somewhat greater complexity, but the basic relationsare everywhere the same.The question arises as to why these reticular structures
have so long escaped notice. The answer is that they haveindeed been dimly perceived at times, but the state of pres-ervation of the tissues was not adequate for a clear recogni-tion of their form and functional role. Also some investigators(e.g., Hama, ref. 2) in studying these organs have dissectedthem out and in so doing have no doubt destroyed theirdelicate structure.
Retzius (1) long ago spoke of two levels of the otolithicmembrane, one a lower part in which the cilia extend intolittle caverns, and an upper part in which the otoliths areembedded. Vilstrup in 1950 (3, 4) described a "subcupular
Proc. Nat. Acad. Sci. USA 70 (1973)
li
Al

Proc. Nat. Acad. Sci. USA 70 (1973)
zone" in the crista of the codfish, with a reticulum that heconsidered to be a part of the sensory epithelium and pro-duced in embryo by an extrusion of cytoplasmic materialsfrom this epithelium. Jurato and de Petris (5), however, wereunable to verify these observations, and regarded the sub-cupular structure "at least partly" as an artifact. The presentresults show, on the contrary, that Vilstrup's "zone," thoughpoorly preserved and somewhat vague in appearance in hisphotomicrographs, is a reality and unquestionably serves auseful purpose in stimulation as he maintained. Finally thepresence of the reticulum is at least suggested in photomicro-graphs of the saccule by Lewis and Nemanic (6) and by adrawing presented by Hillman and Lewis (7), though thisstructure was not recognized by them for what it is.
DISCUSSIONThe evidence just presented, in which the frog is treated inparticular but a cursory sampling is made of all the vertebrateclasses, indicates that the tectorial structure arose early in thephylogenetic development of the labyrinth and continuedthrough all its evolutionary stages. This structure clearlyplays an important part in the functioning of all the labyrin-thine endings.The basic function of this structure is apparent: it encloses
the ciliary tufts of the hair cells and makes the connectionswith them. Thus, it is the means by which a relative motion isset up between the bodies of the hair cells and their ciliarytufts, whereby these cells are stimulated. How this relativemotion is established varies greatly with the type of endingand its receptive function.For the auditory papillae, as brought out in another report,
the tectorial reticulum lying over the hair cells receives vibra-tory movements, sometimes directly but much more oftenfrom other tectorial tissues that lie in a path of vibratory fluidflow through the ear, and then transmits these movements tothe ciliary tufts of the auditory hair cells. Because these haircells are borne on a solid, stationary base, there is relativemotion between them and their cilia, and thus these cells arestimulated.For the macular endings the primary stimulus is head posi-
tion or, more properly, a change in this position. Because ofthe high density of the crystals in the otolithic body, gravityexerts a greater force on this body than on the remainingparts of the ending, and when the head is moved a differentialmotion is transmitted through the tectorial reticulum to theciliary tufts, thereby stimulating their cells. Sudden shocksand slow vibrations will also be effective in some degree instimulating this type of mechanoreceptor. It is important tonote that there is never a direct contact between cilia andotoliths, as has sometimes been claimed. The evidence forsuch contact as presented by Iurato and de Petris (see theirFig. 124, ref. 5) is an artifact due to mechanical or fixationdamage and a mixing of tissue materials.For the crista organs the primary stimulus is the accelera-
tive force of rotatory motion. A change of rotation in theplane of a semicircular canal, or one with a component in thisplane, because of the inertia of the fluid will exert a force onthe cupula in a direction contrary to this rotation. The re-sulting bending of the cupula communicates the motion tothe ciliary tufts of the hair cells, thereby stimulating thesecells.
It has usually been supposed that this stimulating effectarises directly from the bending of the cilia (properly, thekinocilia) or by tension exerted on these elements, whichhave been described as running through the cupular canals.If this were so, the presence of the tectorial reticulum wouldseem superfluous. The presence of the large group of stereo-cilia in the tuft would likewise be without effect. It seems morelikely that the stimulation process in the crista organs cor-responds closely to that in the other labyrinthine endings,and that the motion of the cupula is transferred first to thetectorial reticulum and then to the ciliary tuft as a whole, justas in the others.
In all these labyrinthine endings the action of the tectorialreticulum in executing the final step of transferring the rela-tive motion to the hair cell has the obvious advantage ofmaking this transfer to a considerable number of these ele-ments at the same time. There is, consequently, a degree ofspatial summation and synchronization in the response thatmakes for improved sensitivity.There are other possible advantages of the reticular struc-
ture in this stimulation process. It may have a protectivefunction. If the cilia were directly connected to the otolithicmass, an excessive stimulus could easily be disruptive, tearingthese cilia away from their hair cells and producing a seriousloss of sensitivity. As the observations show, the connectionis first through the reticular framework and finally through aslender, pliable strand that will yield somewhat to excessiveforces and thus afford a degree of protection for the sensoryelements.There is the further possibility that the framework of the
tectorial structure provides elastic restorative forces, so thatafter displacements caused by a given stimulus the system willbe brought back to a normal equilibrium condition.
Acknowledgment is made of support of the National Institute ofNeurological Diseases and Stroke and of the Office of Naval Re-search.
1. Retzius, G. (1881) Das Gehororgan der Wirbelthiere (Stock-holm), Vol. I; (1884) Vol. II.
2. Hama, K. (1969) Zeits. f. Zellforsch. 94, 155-171.3. Vilstrup. T. (1950) Ann. Otol. Rhinol. Laryngol. 59, 46-71.4. Vilstrup, T. (19.50) Ann. Otol. Rhinol. Laryngol.59, 19-54.5. lurato, S. & de Petris, S. (1969) in Submicroscopic Structures
of the Inner Ear, ed. Iurato et al. (Pergamon Press, Oxford),pp. 210-218.
6. Lewis, E. R. & Nemanic, P. (1972) Zellforsch. Mikrosk.Anat. 123, 441-457.
7. Hillman, D. E. & Lewis, E. R. (1971) Science 174, 416-419.
502 Physiology: Wever





























