Embed Size (px)
Citation preview

THE IMPACT OF INVASIVE FISH AND INVASIVE RIPARIAN PLANTS ON
THE INVERTEBRATE FAUNA OF THE RONDEGAT RIVER IN
THE CAPE FLORISTIC REGION, SOUTH AFRICA
Steven R. Lowe', Darragh J. Woodford', Dean N. Impsorr' and Jenny A. Dayl
IFreshwater Research Unit, Department of Zoology, University of Cape Town,
Rondebosch 7700, South Africa.
2Scientific Services, Cape Nature, Jonkershoek, Stellenbosch, South Africa.
Corresponding Author: S.Lowe. e-mail: [email protected]
Keywords: invasive species; bass; biotope; trophic response; invertebrateassemblages.
The current paper is presented in partial fulfilment for the degree ofM.Sc.
Percy FitzPatrick InstituteUniversity of Cape Town
Rondebosch 7700South Africa

The copyright of this thesis rests with the University of Cape Town. No
quotation from it or information derived from it is to be published
without full acknowledgement of the source. The thesis is to be used
for private study or non-commercial research purposes only.
Univers
ity of
Cap
e Tow
n

ABSTRACT
1. The freshwater biota of the Cape Floristic Region of South Africa is characterised
by high levels of endemism. Invasive plants and fish are widespread in the region and
represent major threats to indigenous communities. We investigated the impact of
invasive smallmouth bass (Micropterus dolomieu) and black wattle (Acacia mearnsii)
on macroinvertebrate assemblages in a cobble-bed foothill river within the Region.
2. Eight sites were sampled corresponding to four invasion conditions: no invasion;
invasive trees only; invasive fish only; invasive trees and invasive fish. These sites
represented two predation regimes: Sites with many indigenous fish and no bass; sites
with bass and no indigenous fish. Invertebrates were collected from 7 sampling
occasions during spring/summer seasons from September 2003 to January 2005 and
identified to family or genus, assigned to functional feeding groups and biomass
measured. Results were analysed by multivariate analyses of similarity.
3. Invertebrate assemblages clustered into three distinct biotopes: sand, cobble and
marginal vegetation. The abundance or biomass of assemblages within sand were
insensitive to invasion whereas assemblages from marginal vegetation were altered by
the presence of bass alone. Invertebrate assemblages in cobble were affected by the
presence of bass or black wattle or both. The process by which invasive trees
influence invertebrate assemblages is unclear. Cironomidae larvae were greatly
increased in the presence of bass and Simuliidae, Baetidae and most large invertebrate
predators, such as the Odonata, were moderately increased in the presence of bass.
Most other grazing and algae-consuming invertebrates, such as the Heptageniidae and
Elmidae were reduced at bass-invaded sites.
4. Analysis of fish gut contents revealed a preponderance of Chironomidae and
Simuliidae larvae in the diet of the indigenous fish, whereas a wide variety of prey,
particularly the Ephemeroptera, featured in the diet of smallmouth bass. We propose
that established populations of bass in the Rondegat River do not exert a predatory
control over their invertebrate prey whereas indigenous fish do. Bass may alter
invertebrate assemblage structure by removing the key fish predators from the
indigenous freshwater community, causing predatory release on benthic dipteran
larvae and possibly the Baetidae, with subsequent trophic effects throughout the
invertebrate assemblage.
1

INTRODUCTION
Endemism and the freshwater fauna of The Cape Floristic Region
The Cape Floristic Region (CFR) is one of the world's most diverse and endangered
bioregions (CEPF/CI 2002). The exceptional biodiversity of the CFR is under
increasing threat from habitat destruction and fragmentation by agricultural and urban
development, invasion by introduced alien species and the potential effects of global
warming. The freshwater ichthyofauna of the CFR is relatively species poor but
represents a hotspot of endemism, with 16 of the 19 currently described indigenous
species being endemic (Impson et al., 2002). Fifteen of the indigenous species are
listed as threatened, nine of these being considered endangered or critically
endangered (Baillie and Groombridge 1996).
As is the case with the freshwater fish, the freshwater invertebrate biota of the CFR is
less species rich and less intensely studied than the terrestrial floral component but
contains equivalent high levels of endemism: approximately two-thirds of the
freshwater invertebrate species of the CFR are endemic to it and represent one-third
of South Africa's freshwater invertebrate species (Wishart and Day, 2002). Many of
the species are Gondwanan relicts that have persisted in a relatively unchanged
geological and climatic landscape for more than 200 million years (Stuckenberg,
1962).
Invasive species in the CFR
One of the greatest threats to the biodiversity of the CFR is the spread of invasive
species. Invasion by introduced plants currently affects 8% of the surface area of
South Africa and 29% of the Western Cape (the province that encompasses the
majority of the CFR) is invaded by invasive plant species (Versveld et al., 1998). The
riparian zones are susceptible to invasion by dense stands of alien plants and it is
estimated that the area of invaded riparian habitat will double in the next twenty years
(Versveld et al., 1998). Rivers are effective corridors for invasion by plants due to the
transport of seeds by water, frequent disturbance (flooding and changing seasonal
water level), constant moisture and frequently a lack of a light-competing riparian
canopy (Henderson & Wells, 1986; MacDonald and Richardson, 1986). In the
Western Cape, the primary riparian invasive plant is Acacia mearnsii, with other
2

species of Acacia, Prosopis and Sesbania also invading extensive areas of riparian
habitat (Versve1d et al., 1998).
Impacts of invasive riparian plants
In addition to the well documented reduction in stream flow due to greater water
consumption by alien plants than by many indigenous plants (e.g. Versve1d et al.,
1998; Davies and Day, 1998), other dramatic changes may result from the invasion of
riparian habitats. Overshading by taller, faster-growing and more rapidly-recruiting
invasive plants may result in the loss of the indigenous riparian community. Some
invasive trees, such as A. mearnsii, lack a dense root system and may be responsible
for the loss of marginal root mats associated with low-cover indigenous riparian
vegetation, thereby increasing erosion (Rowntree, 1991; Ractliffe et al., 2003).
Invasive vegetation may increase the incidence of debris dams, features typical of
rivers with large woody plants, which restrict instream flow and typically cause
upstream channel widening (Rowntree, 1991). The nature of change will depend on
the particular biological, geophysical and hydrological characteristics of the river
(Ractliffe et al., 2003), making the impacts of invasive vegetation on a river
ecosystem difficult to predict beyond the local level.
Invasive plants and freshwater invertebrates
The replacement of indigenous vegetation with invasive plants may have diverse
effects on the freshwater invertebrate community: the loss of species due to alteration
of the marginal habitat required for specialised life cycles; a change in the
invertebrate assemblage due to hydrological changes such as altered channel shape or
reduced flow (Ractliffe et al., 2003, King & Schael 2001). Increased siltation due to
increased bank and/or bed erosion may smother the natural benthic substratum and
alter rates of leaf litter decomposition and algal growth.(Martin & Neely, 2001;
Ractliffe et al., 2003; Pusey & Arthington, 2003). Increased shading and leaf-litter
input associated with invasive trees (Le Maitre et al., 1996) may shift the river from
an autochthotrophic to an allochthotrophic system and consequently alter the
invertebrate species composition (Feminella et al., 1989). In addition to quantity, the
timing of the allochthonous input and its chemical properties differ greatly between
invasive acacia species and fynbos (King 1987) and may affect the diversity and
abundance of invertebrate taxa (King 1987, Duggan et al., 2002).
3

Smallmouth bass, Micropterus Dolomieu, in the CFR
The impacts of alien invasive fish on South African rivers are poorly documented.
Impson et al., (2002) noted the presence of 16 species of introduced fish in the CFR
with smallmouth bass (Micropterus dolomieu) being one of the most successful
invaders. M dolomieu was first introduced into the Western Cape in 1937 and to the
Olifants River in 1943 (Harrison, 1953). Since then, M dolomieu has become
established in all the major river systems in the Western Cape (Bruton & de Moor,
1988) so that indigenous fishes are now largely confined to smaller tributaries and
headwaters, often above waterfalls that prevent the spread of invasive fishes (Gaiger
et al., 1980). M dolomieu has been implicated in the decline of six endemic species in
the Olifants River (Bruton and de Moor, 1988) and several observations and studies
have noted the lack of co-occurrence between bass and indigenous fishes (Harrison,
1953; Christie, 2002; Shelton, 2003; Woodford, 2005). To date, no studies have
investigated ecosystem-wide or multi-trophic-Ievel impacts of M dolomieu in South
Africa.
The potential for ecosystem-wide impacts of invasive fish
Experimental and field studies in New Zealand (on trout) and the USA (on bass
species) have shown that invasive fish can exert a top-down impact on river
ecosystems. These impacts are often mediated by changes in the abundance,
composition or behaviour of grazing invertebrates (McIntosh & Townsend, 1994;
Peacor & Werner, 2000) and fish (Power & Matthews, 1983; Power et aI., 1985). The
effects may translate to a change in primary production by algae in the river and
altered nutrient turnover (Power 1992a,b; Flecker & Townsend 1994. The community
or ecosystem-wide impact of invasive fishes is currently difficult to predict due to the
paucity of studies undertaken and the range of systems and of fish species studied.
The context of the study
The Table Mountain Fund (TMF) was created to implement projects identified by The
Cape Action for People and the Environment (CAPE). The Rondegat River, a
tributary of the Olifants River, was identified as a priority site for rehabilitation due to
the threat posed by M dolomieu, which had invaded lower reaches of the river. This
study provides data to inform the TMF of the impacts of alien fish (M dolomieu) and
vegetation (particularly A. mearnsii) on the invertebrate fauna of the Rondegat River
4

in conjunction with a complementary report (Woodford, 2005) on the impacts of M
dolomieu on indigenous fishes.
Purpose of the study
This study investigated whether invasive fish or invasive vegetation influence the
structure of the invertebrate community within the Rondegat River. The focus was on
characterising any patterns of impacts due to invasive species and thereafter to define
some of the processes by which these may be caused.
The impact of invasive fish on river biota, particularly on indigenous fish species,
provides the context of the study. By describing responses, if any, of the invertebrate
community to invasion, information will be provided that will contribute to the
understanding of ecosystem-wide impacts of riparian invasions in the Cape Floristic
Region.
METHODS
Site description
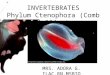
The Rondegat River (32°24'S; 19°05'E) has its source in the Cedarberg mountains at
an elevation of 1000 metres and flows in a north-westerly direction, over a
predominantly Table Mountain Sandstone substratum, for 25 km, until it joins the
Olifants River at Clanwilliam Dam (Figure 1). The catchment is approximately 111
km2 in area and receives 711 mm annual average rainfall, of which the majority falls
in winter (June-August), February being the driest month (Mannicon, 1998). The river
flows through pristine fynbos in its upper reaches, then through a catchment featuring
citrus plantations, two campsites, a forestry station, cattle pasture, areas of dense
infestation by alien trees (mainly A. mearnsii and, to a lesser extent, A. melanoxylon)
and several artificial weirs and water-abstraction points. A natural waterfall about 18
km from the source acts as a barrier to the upstream invasion of M dolomieu and
appears to be an important factor in defining the nature of the fish populations, with
large specimens of the Clanwilliam yellowfish (Labeobarbus capensis) the only
downstream representatives of the six species of indigenous fish found in the river
(Woodford, 2005).
5

Collection sites and dates
Eight sites were chosen along the river (Figure 1) as duplicates of four invasion
conditions (invasion status, IS1-4. IS1: no invasion; IS2: invasive plants only; IS3:
invasive fish only; IS4: invasive plants and fish). Some characteristics of each site are
summarised in Table 1. At sites 3-7 the small flood plain, approximately 150 m wide,
has been converted to cattle pasture. At sites 6 and 7 cattle frequently cross the river.
These sites (IS3), which were largely cleared of invasive trees by early 1990, are
characterised by a narrow margin of indigenous riparian vegetation with some small
recruits ofA. mearnsii.
Sampling was conducted over two spring/summer seasons. In total seven invertebrate
collections were made: September 2003 and October 2003 and February 2004 and
April 2004 represent year 1. October 2004 and November 2004 and January 2005
represent year 2. After the first year of sampling, site 8ii was chosen upstream of a
nearby extraction weir, which almost eliminated flow from site 8i that summer.
No flow measurements were made on the Rondegat River. However, data from a flow
meter (EIH006, DWAF hydrological services) on the Jan Dissels River, which runs
parallel to the Rondegat in an adjacent valley, and which usually receives rainfall at
the same time, was used to indicate relative flow rates on the Rondegat. Figure 2
shows the average daily flow rate for each month on the Jan Dissels River. The
average rainfall for August 2003 and 2004 was similar but the rainfall for September
in 2004 was much reduced compared to the previous year. Possibly of more
importance to the river ecosystem however, were two floods in late August and early
September 2003 which were much larger than any floods in the following year. Sitel
(lSI) and site 3 (IS2) were lacking in sand habitat during year 1, possibly as a result
of scouring by the floods. Therefore, data on sand for IS1 (pristine) and IS2 (invasive
vegetation) from year 1 are based on samples from only site 2 and site 4 respectively.
Collections from cobble, sand and marginal vegetation were made from site 6 (IS3)
starting in April 2004, therefore data for IS3 (invasive fish) for September and
October 2003 are from site 7 only.
Temperature, conductivity and pH were recorded at each site on each collection date
between 10:00 and 16:00 hours. These parameters were not measured in November
2004 or January 2005. The temperature recorded in riffles from sites 1-8 along the
river typically did not exceed a difference of 3°C during any sampling occasion with
the exception of two outstanding recordings in October 2003 and one much cooler
6

recording in September 2003. The variation in pH at different sample sites was less
than 1.0 during any sampling occasion. However, conductivity varied by an average
of 150% from site 1 to site 8 from the same sampling dates.
Shading was estimated at each site by assuming a virtual arc transversing the river
from one bank to the other. From the center of the river, the percentage of open sky
versus vegetation, river banks and rocks on the arc was estimated.
Invertebrate collection and identification
Invertebrates were collected separately from sand, marginal vegetation and cobble, if
present, at each site, using a kick-net. Each of the three habitats was sampled for two
minutes or until the entire available habitat had been sampled within a 20 m stretch of
river. All in-current cobble biotopes in at least 15 cm of water were sampled and
pooled. Habitats in pools were not sampled.
Invertebrates were preserved immediately in 5% formalin and transferred to 70%
ethanol within 48 h of collection for subsequent identification and counting using a
dissecting microscope. Organisms were identified to a taxonomic level suitable for
classification into functional feeding groups (FFG) as defined by Schael and King
(2005, in prep)(Table 2), with the exception of the Chironomidae (Diptera). Baetidae
(Ephemeroptera) from various samples were identified to genus and most of these
were classified as type 1 deposit feeders (DFl). Therefore, in the analysis of FFGs,
Baetidae are either included as DFl, or removed from the analysis as stipulated.
The following texts were used for identification: Day, Harrison and de Moor, 2002;
de Moor, Day and de Moor, 2003a,b.
Biomass
Mean biomass values were measured for all common taxa (those contributing to
>0.5% total abundance). For each taxon, between five and 300 individuals were
selected, depending on the numbers available and body size, from samples collected
in November 2003 and 2004. The samples were dried in an oven at 40°C for 48 hand
weighed to 0.1 mg accuracy.
7

Fish stomach analysis (by D. Woodford)
Fish were caught in seine or fyke nets or by electrofishing in season I (09/2003
04/2004). Smaller fish «10 em) were preserved in 10% formalin. Stomachs from
larger fish were removed immediately and injected with 10% formalin. Stomach
contents were removed and separated into major food item groups: invertebrates, fish,
detritus, filamentous algae and other plant material. These were then placed into a
glass vial containing 70% ethanol and the volume estimated according to the
following formula:
v = (x.100·I )IIr h
where: v = volume, r = radius of the vial base, x = estimated gut content cover of vial
base, h = the height of food component in the vial.
The invertebrate component was then further identified to order or family depending
on the level of digestion of the prey items.
Data analysis
Data were entered into the software package PRIMER (Clarke and Gorley, 2001) and
analysed using Multi Dimensional Scaling ordination (MDS) and CLUSTER analyses
to detect patterns of relative (dis)similarity between samples. Bray-Curtis similarity
was used for all tests and data were standardised to account for variation in the
amount of substratum sampled at each site and between habitats (cobble, sand and
marginal vegetation). ANOSIM (analysis of similarity) was performed on the
similarity matrices generated in PRIMER and differences between specific pairs of
samples compared (e.g. invertebrates in sand from invaded status I compared to those
from sand at invaded status 4). A 95% confidence interval was chosen in order to
accept (p>0.05) or reject (p<0.05) the null hypothesis (Ro) that the mean and variance
of two samples are not different. Abundance data were fourth-root transformed, in
order to give weighting to rarer taxa, and all taxa included in the analysis unless stated
otherwise. Biomass data were fourth-root transformed (unless otherwise stated), and
only those organisms contributing to >0.5% of total biomass within each habitat were
included in the analyses.
Samples that were statistically dissimilar (as determined by ANOSIM) were then
subjected to SIMPER analysis in order to determine the degree of dissimilarity and to
identify which taxa contributed to the dissimilarity.
8

The potential influence of environmental variables was investigated using the
following procedure: conductivity (IlS em"), temperature (Qq, pH, shading (% cover)
and flow (nr's") were entered into a Draftsman's Plot (PRIMER) and transformed to
increase linearity and reduce skewness. These variables were then used, in
conjunction with the similarity matrix generated for the corresponding invertebrate
sample data, in the BIO-ENV analysis procedure (PRIMER). Dissimilarities,
calculated by Normalised Euclidean distance and Spearman's Ranking, were used to
provide a correlation between combinations of environmental variables and the
invertebrate data. The association between the grouping of samples and
environmental factors was then compared and overlaid on MDS plots.
RESULTS
Invertebrate diversity in the Rondegat River
Appendix 1 lists all taxa identified from the Rondegat River. For each year, the
average abundance and standard deviation of taxa in each sample from sand, marginal
vegetation and cobble is given. Appendix 2 displays the average abundances over all
sampling occasions for each taxon at each of the eight sites during year 2
(2004/2005).
Environmental influence
The five environmental factors (conductivity, temperature, pH, shading and flow)
were not closely linked to each other, but the most closely correlated were (in order of
correlation): temperature and flow; pH and shade; conductivity and flow (correlation
= -0.599,0.483, -0.437 respectively). The environmental factors that best explained
the distribution of the collected invertebrates were a combination of conductivity, pH
and flow. However, this combination of variables appeared to be poorly correlated to
the sample data (correlation = 0.166) as confirmed by MDS ordination overlay of the
individual factors.
Similarity and dissimilarity within invertebrate abundance data
Inter-year variation
Within all the three habitat types there was significant difference between the two
sample years (2003/4 and 2004/5, p<O.Ol) for taxon abundance (Table 4a) whether
data were untransformed or fourth-root transformed. Presence/absence transformation
9

of the data showed significant differences between the means of the two years for
sand only (P<0.05). This may be accounted for by the lack of sand substratum at some
sites due to scouring by floods in August/September 2003. Due to these differences,
data were analysed separately for each year.
Habitat differences
Analysis of inter-habitat differences by MDS and cluster dendrograms showed
distinct grouping of samples from cobble, sand and marginal vegetation for both
years. Figures 3 and 4 are two-dimensional representations of the similarity of
samples from habitats representing the four different invasion conditions during year
2 (see Table 1 for details of invasion status). The invertebrate assemblages of the
three habitats were significantly different within year 1 and year 2 in pairwise analysis
of similarity (sand/marginal vegetation; sand/cobble; marginal vegetation/cobble
within each year. p < 0.01 for all tests, ANOSIM, Table 4b). Table 5 shows the
average abundance of taxa per sample and the similarity scores of the taxa that
collectively contributed to >75% of the similarity within each habitat in year 2.
The Baetidae and Chironomidae were ubiquitous and numerous and consistently
contributed the most to the similarity for each habitat. Other taxa characteristic of the
three habitats for both years were the Gomphidae in sand, the Zygoptera, Caenidae,
Veliidae and Athripsodini in marginal vegetation, and Cheumatopsyche,
Leptophlebiidae and Heptageniidae in cobble. The life histories of these taxa are
consistent with the location from which they were sampled (Day et aI, 2002; de Moor
et at., 2003a,b).
Differences between invasion conditions
The MDS ordination of invertebrate samples grouped by habitat and invasion status
indicated that within the habitat clusters there was grouping of samples for different
conditions of invasion (lSl-4) (Figure 3). Table 6a summarises the results of
significant dissimilarities of pairwise ANOSIM analysis between these groups. There
was no significant difference between the sand sites in any of the four invasive
conditions in either year. The invertebrate assemblage in cobble substrata of the
pristine, uninvaded sites (IS1) differed significantly from those in the cobble of the
other three conditions (lS2, IS3 and IS4). In addition, cobble sites invaded by alien
vegetation only (lS2) were significantly different from those with only alien fish
10

(IS3). The invertebrate communities in marginal vegetation were significantly
dissimilar only between uninvaded areas and those invaded only with alien fish (IS1
compared to IS3). All other conditions within the same habitats were not significantly
different. Figure 5 a-e graphically represents pairwise comparisons of the taxa that
cumulatively contribute to >25% of the dissimilarity between samples from year 2
grouped by habitat and invasive status. On each graph, the taxa are ranked from left to
right, with those on the left contributing most to the dissimilarity between the invasion
conditions. Consideration of the biology of each of these taxa can provide an
indication of the processes which may be operating to produce differences between
the invasion conditions.
The same analysis procedure for year 1 revealed largely similar results to those from
year 2. The MDS produced a high stress factor (0.25, 50 iterations) and less distinct
groupings than those for year 2. Samples from sand showed no visible clustering for
different invasion conditions (as for year 2) and were not statistically dissimilar from
each other (ANOSIM). Invertebrate communities in cobble from pristine sites were
significantly dissimilar to those in cobble from sites invaded by alien fish alone
(cobble IS3) and from those sites invaded by both alien fish and alien vegetation
(cobble IS4). A summary of these results is provided in table 6b. In contrast to year 2,
however, invertebrate lSI cobble samples (uninvaded sites) were not dissimilar to IS2
samples (alien plant invaded sites). Also in contrast to year 2, marginal vegetation
samples from sites invaded by alien fish alone were dissimilar to sites with both alien
fish and vegetation in year I (IS3 compared to IS4). All other sample groups from the
same habitats were not significantly dissimilar when compared between invasion
conditions.
As implied by the discrete separation of samples from sand and cobble in year 2
(Figure 4), analysis of similarity between all sample groups from sand compared to
those from cobble for the four invasion conditions are significantly dissimilar
(p<0.05). For year I however, all sand samples (sand ISl-4) are significantly
dissimilar to cobble samples from IS1 and IS2 groups, but not from IS3 and IS4
groups (with the exception of sand lSI to cobble IS3). These data imply increased
sedimentation and a more embedded cobble substratum at downstream sites.
11

Functional Feeding Groups (FFGs)
Taxa were aggregated into FFGs (see Table 2 for FFG definitions and abbreviations
and appendix 1 for the FFG of individual taxa) and analysed by the same process as
described above for unaggregated taxa. This broader-scale analysis of taxonomic
grouping was used in order to characterise particular life-history traits of taxa that
may be affected by the changing conditions along the river. The number of
individuals in each FFG from each sample was square-root transformed and revealed
clustering into the three distinct habitat types, comparable with unaggregated data.
With the exception of Grazer 2 (mostly Heptageniidae), the abundance of all FFG
categories per collection was greater in year 2 than in year 1. Notably, the number of
individuals per sample from the predator 2 (P2) group was over three times greater in
year 2 than in year 1. Clustering by invasion status was also similar to that for
unaggregated taxa: in year 2, samples from cobble at pristine sites were significantly
dissimilar (31%) from sites invaded by alien fish alone (p<0.05; r = 0.444). FFGs
contributing most to the dissimilarity were the deposit feeder 1 group (DFl), followed
by the predator 2 group (P2), followed by the filter feeders (FF). These groups vary
from the functional feeding status of the individual taxa that contributed to the
dissimilarity between pristine (lSI) and alien fish only (lS3) groups with
unaggregated data as shown in figure 5b. (The FFG categories of the individual taxa
contributing to ISl/lS3 dissimilarity for unaggregated data are (not including
chironomids): DF2, followed by P2, followed by grazer 1, followed by scraper 2).
Pristine sites were not significantly dissimilar from sites invaded by alien vegetation
(lS2, p = 0.08) and totally invaded sites (lS4, p = 0.06) despite low p-values, and
therefore warrant further investigation. FFGs from cobble IS2 samples were
significantly dissimilar from cobble IS4 samples (p = 0.04; r = 0.465; 33.1%
dissimilarity) in contrast to unaggregated taxa IS2 samples which differed from
cobble IS3 samples. As with unaggregated data, samples from marginal vegetation
differed significantly only between pristine (lSI) and alien fish only (lS3) invaded
sites (p <0.01; r = 0.464; 35% dissimilarity).
The analysis of square-root-transformed data for FFGs from year 1 shows the same
grouping of sites when compared to the analysis for individual taxon abundances.
Cobble samples from pristine (IS1) sites were dissimilar from fish-only invaded sites
(lS3) (p <0.05, r = 0.425) and fully invaded sites (p <0.01; r = 0.5). Also, samples
12

from marginal vegetation differed significantly only between fish invaded (lS3) and
fully invaded sites (lS4) ..
Biomass
The relationship of biomass and diversity between samples was analysed by grouping
samples into each habitat type and then selecting only those taxa that contribute
>0.5% to the total biomass for that habitat. A list of the taxa weighed can be seen in
Table 3. Table 7 summarises the analysis of the biomass of samples from the four
different invasion statuses. The results show related trends of dissimilarity between
groups when compared to the similar analysis performed with taxa abundance (shown
in Figure Sa and e). Samples from sand showed no significant dissimilarity between
the four invasion conditions and showed the same similarity between all sand sites
and cobble IS3 and IS4 sites in year 1 as for abundance data. Samples from marginal
vegetation were significantly dissimilar only between pristine (IS1) and alien fish
(lS3) sites and samples from cobble were dissimilar when compared between lSI and
IS2, IS1 and IS3 and IS2 and IS3 sites (not between IS1 and IS4 as for abundance).
Despite these similar trends, the taxa contributing towards the differences in biomass
were considerably different from those contributing to the dissimilarity of taxa
abundance for the same groups. Notably, the taxa contributing most to the
dissimilarity in biomass were large-bodied predators such as the Megaloptera and
Odonata.
Biomass was compared between the three habitats and subsequently within the
habitats for the four invasion conditions in year 2 using ANOYA and Tukey's tests.
These analyses were undertaken to test whether univariate analysis of untransformed
data can detect similar patterns to those observed by the multivariate Bray-Curtis
similarity method employed by PRIMER. Pairwise analysis of cobble, marginal
vegetation and sand revealed that only cobble and sand differed significantly (f =
14.7, p = 0.018, ANOYA). Subsequent analysis revealed that the only significant
differences between sites within any of the habitats were samples from cobble IS1
sites and cobble IS3 sites (p <0.05, df = 46, Tukey HSD). Figure 7 shows the average
biomass per sample (and SD) for cobble, marginal vegetation and sand from year 2 at
each site along the river.
13

Biomass of FFGs
The biomass of FFGs was assessed in order to investigate whether the changes in the
invertebrate assemblages described above are characterised by groups of taxa with
particular feeding traits. The biomass of taxa from each sample was aggregated
according to their assigned FFG (see appendix I for FFG of individual taxa). FFGs
utilising a similar food resource were aggregated: the deposit feeder 2 group and
scraper 2 group were aggregated, also, the grazer 2 and scraper 1 groups were
aggregated. Figure 8 a-d represents the average biomass and standard deviation of
each FFG per sample from each habitat. The predator 2 group is displayed separately
due to the much larger biomass of this group. The results of these analyses are
summarised in Table 8. No differences were detected between samples from different
invasion conditions from sand or marginal vegetation. Significant differences were
detected between cobble samples from pristine sites (lSI) and sites invaded by alien
vegetation alone (lS2) and sites invaded by alien fish alone (lS3). Cobble samples
from IS2 sites were also significantly different from IS3 site cobble samples. These
differences are accounted for mostly by the predator 2 and predator 1 groups. The
grazer2/scraperl group was characteristic of the dissimilarity between pristine (IS1)
and invasive vegetation-only (lS2) groups and deposit feeder 1 (DFl, primarily
Baetidae) accounted for differences between alien vegetation-only (lS2) and alien
fish-only (lS3) groups.
The removal of either the predators (PI and P2) or the herbivores (all other groups)
from the analysis did not alter the significance of the dissimilarity between pristine
sites and the IS2 and IS3 sites. However, the sites invaded by alien vegetation alone
(lS2) showed no dissimilarity to alien fish-only invaded sites (lS3) upon removal of
predators from the analysis, but retained the dissimilarity upon removal of herbivores.
Removal of herbivores from the analysis of the FFG biomass revealed a significant
dissimilarity between samples from marginal vegetation where the river was
uninvaded (lSI) compared to areas invaded by alien fish alone (lS3).
Fish diet (collected and processed by D Woodford).
An understanding of the feeding preferences of the fish that characterise different
parts of the river is important in order to appreciate how an invasive fish may exert an
impact on the invertebrate assemblages. Gut contents were analysed from M
14

dolomieu (n == 32) and four indigenous fish species: Labeobarbus capensis, n == 30;
Pseudobarbus phlegethon, n == 30; Barbus calidus, n == 30 and Austroglanis gilli ee 32
(Figure 9 a-e). Invertebrates composed a minimum of 60% of the gut content volume
of all fish species (Woodford, 2005). Identification of the invertebrate taxa from gut
contents yielded results consistent with the feeding strategies of the fish species (see
Woodford 2005 for details): the most notable differences between the indigenous fish
species and the invasive M dolomieu, was the large number of dipteran larvae
(Chironomidae and Simuliidae) in the gut contents of indigenous fish, in contrast to
the guts of M dolomieu, which contained mostly Ephemeroptera, particularly the
Baetidae. Other notable differences between the indigenous and invasive fish were
trichopteran larvae in the gut contents ofP. phlegethon and A. gilli and Odonata in the
diet ofM dolomieu.
The dipteran larvae (Chironomidae and Simuliidae) and Baetidae are the most
abundant taxa in the diet of the indigenous fish and M dolomieu respectively. The
biomass and calorific content of each prey taxa are the most important factors
determining the contribution to the diet of the fish. The numerical contribution of
Odonata compared to Baetidae in the diet of M. dolomieu is only 7.5%. However, if
those Odonata are large Gomphidae, the total biomass of Gomphidae will be 450%
greater than that of the Baetidae.
DISCUSSION
Heterogeneity is a striking characteristic of freshwater biota (Palmer and Poff, 1997;
Li et al., 2001) but the type of heterogeneity observed will depend upon the scale of
the observation, and the spatial arrangement and quantity of resources and consumers
(McIntosh et a1., 2004). The scale of the sampling technique and the level of
taxonomic identification used in this study can detect patterns of invertebrate
assemblages characteristic of the three habitat types, or biotopes, within the river:
sand, marginal vegetation and cobble. The ability to detect these trends was consistent
for similarities in abundance and biomass and for different transformations of the
data. Patterns of invertebrate species composition that coincide with different invasion
conditions can also be detected along the Rondegat River. However, the taxa
contributing to these dissimilarities vary depending on the type of data used
(unaggregated abundance of taxa, FFGs or biomass). These data imply that the causes
15

of the dissimilarity at sites invaded by alien species may have impacts at multiple
levels within the invertebrate assemblages.
Inferring causality to observed patterns is difficult even with experimental
manipulation (such as fish or invertebrate exclusion zones) or closely comparable
rivers (if, indeed, such suitable 'control' reference sites can be applied), neither of
which were included in the present study. The spatial separation of sites with different
invasion conditions along the river is problematic for formulating conclusions based
on differences between samples from these sites. However, contrary to previous
notions of longitudinal river zoning, extensive work recently undertaken on macro
invertebrate distributions within rivers in the Western Cape, has provided strong
evidence that the catchment (for the current study: the Olifants River) and the bedrock
substratum (Table Mountain sandstone) are stronger influences on invertebrate
assemblages than the longitudinal zone (King and Schae1, 2001). Upland rivers in the
Western Cape are known to exhibit high variability, but differentiation within upland
sites into mountain streams and foothill-cobble beds is not apparent (Dallas, 2002). In
the current study, comparison of temperature, pH, conductivity and coarse measures
of shade and flow with the invertebrate data set from the Rondegat River, implies that
there were factors that exerted a stronger influence in the grouping of the invertebrate
samples other than the type and method of environmental factors measured.
The potential influence of fish predation on invertebrate assemblages
By comparing the gut contents of fish with the distribution of the prey items found at
each site (see appendix 2) it seems likely that M dolomieu does not constrain the
population size of its invertebrate prey items: Baetidae and Odonata were generally
more numerous at lower sites, despite featuring proportionately higher in the diets of
Midolomieu than of the indigenous fish found upstream. In contrast, the main prey
items of the indigenous fish, the Chironomidae and Simulidae, are less numerous at
the upper sites and it is possible that the numerically superior indigenous fish exert a
top-down control of their prey items. In the absence experimental studies and of
biomass and energy assimilation measurements for fish at different sites, it is not
possible to conclude if river reaches populated by indigenous fish are subjected to
greater top-down control. It is the opinion of the author that the fish biomass in the
Rondegat River is greater in the absence of invasive fish. This is in contrast to studies
on invasive trout, which generally indicate an increased predation pressure in the
16

invaded as opposed to the native state (Flecker and Townsend, 1994; Simon and
Townsend, 2003). Trout are visual diurnal predators (as are bass) that typically
achieve a higher biomass than the indigenous fish they displace. The impacts of
increased predation pressure by trout on invertebrates have been shown to have a
number of potential mechanisms: direct control of prey by consumption, shifts from
daytime to nocturnal drift of mayflies in the water column (McIntosh et al., 2002) and
reduced total foraging time of invertebrate grazers (McIntosh and Townsend 1995).
Data on temporal changes of fish feeding preferences and experimental methods to
quantify the predatory control exerted by the fish species on invertebrate prey, will
greatly enhance our understanding of the community-level effects of introduced fish
species.
Patterns of invertebrate responses along different invasion conditions:
The cobble biotope
The invertebrate assemblage within the cobble biotope consistently possessed the
highest biomass and species diversity of the biotopes sampled, as has been shown for
other studies (Dallas 2002). Cobble samples also demonstrated the most frequent
differences between the four invasion conditions, implying that cobble is the most
sensitive of the biotopes to invasion. Cobble samples from pristine areas of the river
are significantly different from areas invaded by alien fish alone, whether analysed by
univariate statistics, or multivariate measures of similarity based on the abundance or
biomass of taxa. In the presence of M dolomieu, the total invertebrate biomass is
significantly higher. Notably, there is a greater number and biomass of predatory
invertebrates in the alien fish-only invaded areas and there are more algae-consuming
scrapers (Sl), grazers (G2) and deposit feeders (DF2) within the pristine sites (see
Figures 5b and 8d).
Significant dissimilarity between the biomass of samples from pristine sites and sites
invaded by alien vegetation or alien fish is retained whether analysed for predators or
herbivores only. This implies that the invertebrate assemblage structure of sites
invaded by alien fish or vegetation alone is altered at multiple trophic levels compared
to uninvaded sites. In contrast, invertebrate predator composition, rather than the
composition of invertebrate herbivores, is largely responsible for differences between
cobble samples from sites invaded by only alien vegetation compared to areas invaded
only by alien fish.
17

As discussed above, the indigenous fish are likely to exert a greater control of
Chironomidae larvae than M doiomieu. Most of the Chironomidae are classified
within the grazer 2 FFG, and an increase in their numbers in the invaded sites, due to
predatory release in the absence of indigenous fish, could result in competitive
exclusion of other algae-eaters. The presence of Ephemeroptera other than the
Baetidae in the diet of M doiomieu, but not in the diet of indigenous fish (Figure 8),
indicates that increased selective predation could also be responsible for the reduction
of the Heptageniidae at sites invaded by alien fish (Figure 5b and c).
The dramatic increase in both small and large invertebrate predators in the presence of
alien fish is coincidental with a large increase in the total number primary consumers.
It is possible that the increased food resource for the invertebrate predators outweighs
the increased risk of predation from M doiomieu. The greater number of Baetidae at
alien fish-invaded sites is difficult to explain considering the high propensity of
Baetidae in the diet ofMdoiomieu. It is possible that bass-invaded sites still represent
a decreased risk of predation to Baetidae, based on fish biomass at different sites.
Increased predation, however, can also result in an unchanged, or even increased
number of Baetidae, due to complex interactions between foraging behaviour and
algal abundance (Diehl et ai., 2000; McIntosh & Taylor, 2004).
The dissimilarity between cobble samples from pristine sites and those from sites
invaded by alien vegetation (IS2 and IS4 sites) is difficult to interpret. The Caenidae
contribute most to the difference between these invasion conditions and are more
numerous at both conditions invaded by alien vegetation. The Caenidae were more
numerous in marginal vegetation than in cobble samples (see appendix 1), however,
the Caenidae are known to feed in silty deposits (de Moor et al., 2003a) and their
increase in cobble samples from IS2 and IS4 conditions could represent increased
sedimentation at these sites due to the increased presence of debris dams and bank
erosion resulting from the presence of alien vegetation (with the exception of site 5,
IS4).
The sand biotope
Sand exhibited the lowest diversity and biomass of the biotopes, consistent with
SASS (South African Scoring System) scoring from other studies (Dallas, 2002). The
18

consistent lack of dissimilarity in samples from sand indicates that the organisms
inhabiting this substratum are resilient to change in the river food web. Sand may act
as a refuge from predation pressure for those organisms adapted to living in it, such as
the burrowing Gomphidae. It is also possible that sand is a lower nutrient, less
heterogenous environment, requiring specialised life histories that make the resident
organisms less vulnerable to competition. Sand was also the only biotope to show
significant differences between collection dates for year 2, for which there are many
possible explanations including emigration and reduced or increased sedimentation
from October 2004 to January 2005.
The marginal vegetation biotope
The invertebrate communities in marginal vegetation showed fewer differences
between invasion conditions than for cobble. There were significant differences only
between pristine sites and sites invaded by alien fish alone in year 2 and between
invasive fish only and both invasive fish and vegetation (lS3 and 4) in year 1. The
taxa contributing to dissimilarity between marginal vegetation samples from pristine
sites compared to those from sites invaded by alien fish alone are consistent with the
proposed predatory control of specific invertebrate taxa for cobble samples (see
Figure 5e): the Chironomidae were far more abundant in the absence of indigenous
fish (at IS3 sites). The Simulidae were also consistently more abundant in the absence
of indigenous fish with the exception of one collection date (average and standard
deviation for lSI and IS3 repectively; 23.5 ± 34.8; 23 ± 14.8). The greater number of
Athripsodes (Trichoptera) at alien fish-only invaded sites confirms the hypothesis that
indigenous fish are exerting a predatory control on their prey.
It was expected that the invertebrate assemblages most affected by the replacement of
indigenous riparian vegetation by invasive trees (as is the case for sites with invasion
status 2 and 4) would be the assemblages residing in the marginal vegetation because
of the direct loss and fragmentation of habitat. Due to the lack of instream marginal
vegetation at sites 8i and ii (lS4), samples could only be collected on two out of four
dates for year 1 (site 8i) and for two samples out of three in year 2 (site 8ii).
However, the other fully invaded site, site 5, still retained patches of indigenous in
stream riparian vegetation, possible due to the rocky nature of the banks preventing
invasion at these points. It is from this site that most of the marginal vegetation data
for invasion status 4 is derived.
19

The kick-sampling technique (described in the methods) may result in sites with a
small amount of a particular habitat being overestimated in terms of overall diversity
and abundance. This is due to sampling a higher proportion of the total amount of the
habitat. The effect will be more pronounced if the species/area relationship is
logarithmic, particularly when taxa are relatively homogenously distributed. A
species/area plot for marginal vegetation samples in year 2 (PRIMER, data not
shown) revealed that, on average, two and five collections will capture >50% and
>75% of species richness respectively. Invertebrate assemblages from marginal
vegetation show little change compared to cobble assemblages, with marginal
vegetation acting as a refuge. Based on these data, a measure of the abundance of
marginal vegetation may be a reasonableindication of the state of the populations of
resident invertebrate taxa.
Addressing potential sampling bias
In order to address the problem of uneven representation when sampling, a survey
should be conducted to map the physical characteristics of the site and then a
proportion of each habitat sampled consistently between sites. Alternatively, all of a
particular habitat should be sampled in a short stretch of river and the species
abundance adjusted for the area of habitat sampled. Normalising data for sampling
effort is necessary in heterogenous systems if different sites or streams are to be
comparable (Li et al., 2001). Analysis in the PRIMER programme partially addresses
these issues by providing the option to 'standardise' between samples.
Changes in the invertebrate community can also be masked if the level of taxonomic
identification is too coarse to capture changes at a lower taxonomic grouping.
Examples from freshwater and marine benthic systems suggest that, depending on the
questions being investigated, information may not be lost by aggregating species to
higher taxonomic groupings (Clarke and Warwick, 1994; Dallas 2002). Finer
taxonomic identification will be required, however, if the conservation of rare species
is to be addressed.
The Rondegat River appears to be a predator-dominated system with respect to the
biomass of the invertebrate assemblage. Predatory invertebrates, such as the abundant
Odonata within the Rondegat River, could exert a strong influence on primary
consumers and play an important role in structuring the invertebrate community in
20

addition to providing food for higher trophic consumers. Consistent with the findings
of the current report, dragonfly larvae may reflect the physical environment of a river
and have been shown to increase in diversity due to the presence of invasive species
(Stewart and Samways, 1998). Studies have demonstrated trophic cascades mediated
by multiple invertebrate feeding strategies (Flecker and Townsend, 1994; Huryn,
1998; Carlisle & Hawkins, 1998) and have demonstrated the potential for size-related
alteration of predatory invertebrate taxa by introduced fish species, with subsequent
consequences for trophic interactions (Huryn, 1998).
Predatory invertebrates consistently contributed to the differences between sites with
and without invasive fish, as shown in Figure 8 and summarised in Table 7 and
Appendix 2. Further investigation could reveal an important component of the river
ecosystem described in the current report and for similar rivers of the region.
Collectively, our results demonstrate that invasive plants and invasive fish may alter
invertebrate community assemblages. In the absence of experimental controls, the
mechanisms by which the exotic species exert their effects are difficult to asses. It is
also difficult to account for the influence of catchment practices on the pattern of
invertebrate assemblages in the system studied the current report. We propose,
however, that invasive fish cause multiple trophic alterations of macroinvertebrate
assemblages by removing key predators from the indigenous river ecosystem. Further
studies are required to: confirm the findings presented here, to investigate the
possibility of ecosystem-wide consequences of smallmouth bass invasions and to
determine the extent to which the pattern of invertebrate assemblage response persists
throughout the range of invasive smallmouth bass in the CFR.
Acknowledgments
We thank Donni Malherbe, Rika du Plessi and field rangers of Cape Nature for their
help and use of facilities. Also, the Nieuwouldt families at Keurbos and Grootkloof
for use of their land and discussion. Thanks to Angus McNielage and Mark Burman
for laboratory assistance. Geordie Ractliffe, Candice Hill, Sean Marr and Bruce
Paxton of the Freshwater Research Unit, UCT provided helpful discussion and
Colleen Seymore and Tammy Roberts of Zoology department for help with PRIMER.
Dennise Schae1 and Jackie King of FRU provided the FFG classifications. Funding
was provided by the TMF alien fish programme 0NWF SA).
21

REFERENCES
Baillie J. & Groombridge B. (1996) 1996 mCN Red List of Threatened Animals.Gland: rue».
Bruton M.N. & de Moor Ll. (1988) Atlas of alien and translocated indigenous aquaticanimals of South Africa. South African National Scientific Programmes Report, 144,78-133.
Carlisle D.M. & Hawkins C.P. (1998) Relationship between invertebrate assemblagestructure, 2 trout species, and habitat structure in Utah mountain lakes. Journal oftheNorth American Benthological Society,17, 286-300.
CEPF/CI Communications (2002) Critical Ecosystem Partnership Fund: AnnualReport 2002. Conservation International.
Christie D.I. (2002) The distribution and population status of the Cape whitefish,Barbus Andrewi, in the upper Hex River, Worchester and the associated impact ofsmallmouth bass, Micropterus dolomieu. BSc (Honours) project, University of CapeTown.
Clarke K R & Gorley RN. (2001) PRIMER v5: User Manual/Tutorial PRIMER-E:Plymouth U.K
Clarke K R & Warwick RM. (1994) Change in marine communities: an approach tostatistical analysis and interpretation. Plymouth: Plymouth Marine Laboratory, 144pp.
Dallas H.F. (2002) Spatial and Temporal Heterogeneity in Lotic Systems:Implications for Defining Reference Conditions for Macroinvertebrates.3rd WaterNet/Warfsa Symposium 'Water Demand Management for SustainableDevelopment', Dar es Salaam, 30-31 October 2002
Davies B. & Day J. (1998). Vanishing Waters. UCT Press.
Day lA, Harrison A.D. and de Moor Ll. (eds.) (2002) Diptera. Vol.9. WRC ReportNo. TT20l/02.
De Moor, l.J., Day J'A, and de Moore F.C. (eds.) (2003)a Insecta I: Ephemeroptera,Odonata and Plecoptera. Vol.7. WRC Report No. TT 207/03.
De Moor, Ll., Day lA and de Moore F.C. (eds.) (2003)b Insecta II: Hemiptera,Megaloptera, Neuroptera, Trichoptera and Lepidoptera. Vol. 8. WRC Report No. TT214/03.
Diehl S., Cooper S.D., Kratz KW., Nisbet RM., Roll S.K, Wiseman S.W. & JenkinsJr. T.M. (2000) Effects of Multiple, Predator-Induced Behaviors on Short-termProducer-Grazer Dynamics in Open Systems. The American Naturalist, 156,293-313.
22

Duggan I.C., Collier KJ., Champion P.D., Crocker G.F., Davies-Colley R.J., LambertP.W., Nagels J.W. & Wilcock RJ. (2002) Ecoregional differences in macrophyte andmacroinvertebrate communities between Westland and Waikato: are all New Zealandlowland streams the same? New Zealand Journal of Marine and FreshwaterResearch, 36, 831-845.
Feminella, lW., Power M.E. & Resh V.H. (1989) Periphyton responses to grazinginvertebrates and riparian canopy in three Northern California coastal streams.Freshwater Biology, 22, 445-487.
Flecker, A S. & C. R Townsend. (1994) Community-wide consequences of troutintroduction in New Zealand streams. Ecological Applications, 4, 798-807.
Gaigher C.D., Hamman K.C.D. & Thome S.D. (1980) The distribution, status andfactors affecting the survival of indigenous freshwater fishes in the Cape Province.Koedoe, 23, 57-88
Harrison AD. (1965) Geographical distribution of riverine invertebrates in SouthernAfrica. Archivfur Hydrobiologie, 61, 387-394.
Harrison AC. (1953) The acclimatization of smallmouth bass. Piscator, 27, 89-97.
Henderson, L. & Wells, M.l (1986) Alien plant invasions in the grassland andsavanna biomes. In: Macdonald, LAW., Kruger, F.l & Ferrar, AA (Eds) Theecology and management of biological invasions in southern Africa. Proceedings ofthe National Synthesis Symposium on the ecology of biological invasions. OxfordUniversity Press, Cape Town
Huryn AD. (1998) Ecosystem-level evidence for top-down and bottom-up control ofproduction in a grassland stream system. Oecologia, 115, 173-183.
Le Maitre, D.C., van WiIgen B.W., Chapmank.A. & McKelly D.H. (1996) Invasiveplants and water resources in the western Cape Province, South Africa: modelling theconsequences of a lack of management. Journal ofApplied Ecology, 33, 161-172.
Impson N.D., BiIIs R & Cambray lA (2002) A conservation plan for the unique andhighly threatened freshwater fishes of the Cape Floristic Kingdom. In Collares-PerieraM.l, Cowx LG. & Coelho M.M. (eds) Conservation offreshwater fishes: Options forthe future. Fishing News Books. Blackwell Science. 432-440.
King lM., Day lA, Henshall-Howard M.-P. & Davies B.R (1987) Leaf-packdynamics in a south-western Cape mountain stream. Freshwater Biology, 18, 325340.
King J.M. & Schae1 D.M. (2001) Assessing the ecological relevance of a spaciallynested geomorphological hierarchy for river management. WRC Report. No.754/1/01.
23

Li J., Herlihy A, Gerth W., Kaufmann P., Gregory S., Urquhart S. & Larsen D.P.Variability in stream macroinvertebrates at multiple spatial scales. FreshwaterBiology, 46,87-97.
Macdonald, LAW. & Richardson, D.M. (1986) Alien species in terrestrialecosystems of the fynbos. In: Macdonald, LAW., Kruger, F.J. & Ferrar, AA (Eds)The ecology and management of biological invasions in southern Africa. Proceedingsof the National Synthesis Symposium on the ecology of biological invasions. OxfordUniversity Press, Cape Town.
Mannicom C. (1998) Effects of Acacia meamsii on macroinvertebrates in a WesternCape stream. BSc Honours project. Zoology Department, University of Cape Town,South Africa.
Martin C.D & Neely R.K. (2001) Benthic macroinvertebrate response tosedimentation in a Typha angustifolia L. wetland. Wetlands Ecology andManagement, 9, 441-454
McIntosh A R & Townsend C.R (1995) Impacts of an introduced predatory fish onmayfly grazing periodicity in New Zealand streams. Limnology and Oceanography,40, 1508-1512.
McIntosh A R, Peckarsky RL. & Taylor B.W. (2002) The influence of predatoryfish on mayfly drift: Extrapolating from experiments to nature. Freshwater Biology,47, 1497-1513.
McIntosh AR, Peckarsky B.L. & Taylor B. W. (2004) Predator-induced resourceheterogeneity in a stream food web. Ecology, 85, 2279-2290.
Palmer M.A & Poff N.L. (1997) Heterogeneity in streams: the influence ofenvironmental heterogeneity on patterns and processes in streams. Journal of theNorth American Benthological Society,16, 169-173.
Peacor S. D. & Werner E. E. (2000) Predator Effects On An Assemblage OfConsumers Through Induced Changes In Consumer Foraging Behavior. Ecology, 81,1998.
Power, M. E. (1992a) Habitat heterogeneity and the functional significance of fish inriver food webs. Ecology, 73, 1675-1688.
Power, M. E. (1992b) Top down and bottom up forces in food webs: do plants haveprimacy? Ecology, 73, 733-746.
Power, M.E. & Matthews J. (1983) Algae-grazing minnows (Campostomaanomalum), piscivorous bass (Micropterus spp.) and the distribution of attached algaein a small prairie-margin stream. Oecologia, 60, 328-332.
Power, M.E., Matthews W.J. & Stewart AJ. (1985) Grazing minnows, piscivorousbass and stream algae: Dynamics ofa strong interaction. Ecology, 66, 1448-1456.
24

Pusey B.l & Arthington AH. (2003) Importance of the riparian zone to theconservation and management of freshwater fish: a review. Marine and FreshwaterResearch, 54, 1-16.
Ractliffe G., Ewart-Smith 1., Day E. & Grgens A (2003) External evaluation of theWorking for Water Programme. Water Research Commission Report,
Rowntree, K. (1991) An assessment of the potential impact of alien invasivevegetation on the geomorphology of river channels in South Africa. Southern AfricanJournal ofAquaticScience, 17,28-43.
Schael, D.M. and King, I.M. (2005) Western Cape river and catchment signatures.Water Research Commission, Pretoria. WRC Report No. 1303/1/05. (In Prep).
Shelton lM. (2003) The impact of the alien smallmouth bass, Micropterus dolomieu,on the indigenous fishes of a South African river. BSc (Honours) project. ZoologyDepartment, University of Cape Town.
Simon K.S. and Townsend C.R. (2003) Impacts of freshwater invaders at differentlevels of ecological organisation, with emphasis on salmonids and ecosystemconsequences. Freshwater Biology, 48, 982-994.
Skelton P. (1993) A complete guide to the freshwater fishes of South Africa. Struik,South Africa.
Stewart D.AB. & Samways M.l (1998) Conserving Dragonfly (Odonata)Assemblages Relative to River Dynamics in an African Savanna Game Reserve.Conservation Biology, 12, 683-692.
Stuckenberg B.R. (1962) The distribution of the montane palaeo genic element in theSouth African invertebrate fauna. Annals ofthe Cape Provincial Museum, 11, 119158.
Versfeld, D.B., Le Maitre, D.C. & Chapman, R.A (1998) Alien Invading Plants andWater Resources in South Africa: A preliminary assessment. WRC report No.TT99/98, Water Research Commission, Pretoria.
Wishart M'J, & Day lA (2002) Endemism in the freshwater fauna of the SouthWestern Cape, South Africa. Verhandlungen Internationale Vereinigung Limnologie,28, 1-5.
Woodford DJ. (2005) The impact of invasive smallmouth bass (Micropterusdolomieu) on the indigenous fish community of the Rondegat River, Western CapeProvince, South Africa - A quantitative Assessment with implications forrehabilitation. MSc dissertation, Zoology Department, University of Cape Town.
25

I19'00'
lIfOO'
/
N
A
f9'OO'E - "
Cedarberg
Wilderness
Area
4
3
Algeria
zkm \
5
7• Keurbos
Farm
Waterfall
Olifants River
Figure 1 Map of the Rondegat Riverand location of the study sites

9876
2004
543210 11 12 1_1 _
2003
9
3.5 ,----------------------~------3+-----------------------.-::~-----
2.5 -\-----\-------------------+--\-----
2-1-----\-----------------1----\-----
1.5 +-----\-----------------'l.........--/----\-----
1+---------\------------1------\---
0.5 -l-------~------------I-------~==~~O--l----~----,-----.--~~~;::±:~~~~~~~~~~
-";'.!flE-
month
Figure 2 Average daily discharge per month of the Jan Dissels River during the study period.(data from Department of Water Affairs and Forestry)
Key:A 0 A 0 .I. 181 • 182181 182 marginal 181 182
cobble vegetationsand
0 184 o 1830 183 e 184 • 184 • 183
Stress: 0.18
Figure 3 MD8 ordination of invertebrate samples grouped by habitat and invasion condition.Year2. 181: no invasion; 182: invasive plants only; 183: invasive fish only; 184: invasive plants and fish.

~-
~~rf
A~
~--c=t-
- -1
,--
-1 L-j
,----
e~
~~
L--j
I20
I40
I60
Similarity (%)
I80
mv4mv4mv4mv2mv2mv3mv3mv3mv3mv3mv4mv2mv1mv1mv3mv2mv3mv383mv2mv1mv1mv1mv1c2c3c4c2c3c3c3c3c1c1c1c1c1c1c4c2c4c4c4c2c2c4c2c3c184828483848282818281818484818383838182828184
I81
100
MarginalVegetationIS1-4
cobbleIS1-4
sandIS1-4
Figure 4 Dendrogram of year 2 group similarity
Hierarchical clustering of habitats across all invasion conditions Invasion condition1 (IS1) within the cobble habitat ishighlighted

a50
~ 45~ 40lii 35a.~ 30
~ 25E 20~ 15e 10"iV 5
oCaenldae 8rridae Ecnorridae SlmJlUdae L1l1l1lchldae Megaloplera
adun
b
Heplagenidae Aeshnidae O1,onorridae Gyrillidae Oigochaeta Snidee aduh T",u~dee
sllllil larvae
Caenidae f'Ilptagenidae Aeshnidae Brri<fae aduk C. afra HydrachneDae Gorrphidaesmall small
Simuliidae Baetidae Ecnomidae Tipulidae Eimidae Ubeliulidae Aeshnidaesmall adult large small
d5045403530252015105Ot-""UL....J---.-'"""--'--r-..lWWo~,----L.-L--.-"UUL--'---r----'---'--r---'lllll'--'--,
~50:;; 45~ 40~ 35~ 30~ 25~ 20
-:;: 15~ 10~ 5;:; O-+---'='---r-
c
60 214
e
50
45
i 40
~ 35
~ 30o
~ 25.a 20..& 15e~ 10
5
o
f-r-
r-r-
'-~n n Jl
SimJidae Chironorridee Athripsodes Baelidae Ubelluidae Gynnklae Hjdroptiidaesrral srral larvae
Figure 5 a - e The average number of individuals for each taxon whichcontribute to >25% cumulative dissimilarity between sample groups (rankedfrom left to right). Comparisons are between significantly dissimilar samplegroups of the same habitat with a different invasion status (181-4): 181: noinvasion; 182: invasive plants only; 183: invasive fish only; 184: invasive plantsand fish.a: cobble 181/182; b: cobble 181/183; c: cobble 181/184; d: cobble 182/183e: marginal vegetation 181/183

200
600
400
~Cobble
o Marginal \eg
lDISand
800
1400
1200
1600
~
'""C<:o'tl~
"8....~ 1000<I>OJ~
~~
..!!l
'":::l"C:;;:"C<:
~....<I>.0E:::l<:
~~~~~~~~~~~~~~~~~#~~~~~~~~~~~~~~~~~~~~~~~~~~ 'O~'O~
Figure 6 Taxa abundances of Functional Feeding Groups within each habitat for each year(see table 2 for FFG definitions)
fA.cobble
lDIm. veg
<>sand
6 7 8
181182 ~I
Invasion status (181-4) 184
Figure 7 Biomass (mean and standard deviation) at each site for cobble, marginal vegetationand sand (year 2)

a b
Cobble Marginal vegetation
0.1 0.1
0.08 0.08
" "~ 0.06 ~0.06
'" '"'"'" ~ 0.04<00.04E .S!a .c
.c 0.020.02
cobble 181 cobble 182 cobble 183 cobble IS4mveg181 mveg 182 mveg IS3 mveg IS4
hvasion Status invasion status
c
0.01
0.0081:';; 0.006
'"<0
~ 0.004:c
0.002
o
SandKey to Figure 8a-c
o Deposit Feeder 1
~ Deposit Feeder 2/scraper 2
~ Filter Feeder
II Grazer 1
• Grazer 2/scraper 1
~ Predator 1
sand 181 sand 132 sand 133 sand 134
d
invasion status
Predator 2 FFGIn each habitat
0.7
0.6
~0.5
<II 0.4<II11l
0.3E.Q.0
0.2
0.1
0
III cobble
om. veg
Dsand
151 152 153
Invasion status
154
Figure 8 a-dAverage biomass of functional feeding groups within each habitat for year 2.Note: different y-axis scales to account for large variation in biomass between FFGs.

A) Labeobarbuscapensls B)Pseudobarb/#l phlegethon
All otherInvertebrates
7.3%
Corixldae26.4%
AU otherinvertebrates
5.5% Trlchoptera6.8%
O1lrononidae80.9%
0) Barbus calidus 0) Austroglants gill/
Terrestrtalinvertebrates
20.9%
Other aquaticinvertebrates
14.2%
SinlIlidae6.7%
Baetldae9.7%
Coleoptere11.2%
Chironorridae37.3%
E)M/cropterus dolom/eu
Otheraquetlcinvertebrates
9.4%
SllllJlldae11.8%
Terrestrialinvertebrates
1.0%
O1irononidee48.1%
Trlchoptera6.7%
TerrestrialInvertebrates
8.4%
Other aquaticInvertebrates
14.4%
Odonata4.4%
OtherEphemeroptera
11.7%
Fish0.3%
Crab2.2%
Baetidae58.6%
Figure 9 a-e The proportional abundances of animal taxa from fish gutsA: Labeobarbus capensis, n =30; B: Pseudobarbus phlegethon, n =30;C:Barbus calidus, n =30; D: Austroglanis gilli =32; D: M. dolomieu n =32

Table 1. Characteristics of each sample site in the Rondegat River. Invasion statusdefinitions (lSl-4)are refered to throughout the text. Minimum and maximum figuresare derived from all sampling occasions.
Site #Invasion status (IS) pH
Average daytime Conductivity Shadeand IS temperature (0C) (IlS em") (%cover)
1 (lSI) - alien fish min: 5.2 min: 14.0 min: 20.520%
- alien vegetation max: 5.8 max: 16.3 max: 51.0
2 (lSI)- alien fish min: 5.3 min: 12.6 min: 21.4
40%- alien vegetation max: 5.9 max: 17.7 max: 52.7
3 (lS2)- alien fish min: 5.5 min: 14.5 min: 31.5
60%+ alien vegetation max: 5.8 max:J8.4 max: 102.0
4 (lS2)- alien fish min: 5.5 min: 14.6 min: 33.2
70%+ alien vegetation max: 6.0 max: 18.5 max: 103.0
5 (lS4)+ alien fish min: 5.9 min: 15.2 min: 35.5
70%+ alien vegetation max: 6.2 max: 20.6 max: 111.0
6 (lS3)+ alien fish min: 5.8 min: 15.5 min: 37.3 20%- alien vegetation max: 6.1 max: 19.0 max: 118.8
7 (lS3)+ alien fish min: 5.8 min: 16.0 min: 38.9
30%- alien vegetation max: 6.1 max: 22.0 max: 118.8
8i, ii + alien fish min: 6.1 min: 17.5 min: 34.790%
(lS4) + alien vegetation max: 6.3 max: 18.0 max: 147.7
Table 2 Functional feeding group definitions, derived from Schae1 and King (2005).
FFG (code) Feeding Mode Dominant Food TypePredator I (PI) Predator - feeds on organisms by active Micro-organisms - bacteria to
predation zooplankton, zoobenthos, etc.Predator 2 (P2) Insects to small vertebratesFilter feeder Filter by collecting small particles of Micro-organisms: bacteria - algae -(FF) food suspended in the water column zooplankton and particulate organic
matterDeposit feeder I Collecting/gathering deposited organic Detritus (VFPOM and FPOM) only(DFI) material from the substratumDeposit feeder 2 Algae and detritus in similar amounts(DF2) dependent on availabilityShredder Shredder - feeds by fragmenting leaves Vegetation. Usually allochthonous(shred) and large pieces of plant material terrestrial materialScraper I (S I) Scraper - scrapes thin film of micro- Algae (and other micro-organisms)
organisms off substrataScraper 2 (S2) Algae and detritusGrazer I (GI) Grazer - feeds on whole living plants, Living aquatic plants
leaves and stems. Algal mats can beincluded where scraping is not employedas mechanism for collection.
Grazer 2 (G2) AlgaeOmnivore (Om) Feeds on anything and everything Everything
available; mainly reserved forscavengers such as crabs.

Table 3 Taxa weighed for biomass measurements
Taxa Number·of Individuals Masslindivldual (g)weighed
Elmidae adult 150 0.00021
Elmidae larvae 150 0.00019
Gyrinldae adult 6 0.00403
Gyrinidae larvae 22 0.00052
Limnichnidae 50 0.00031
Helodldae 40 0.00018
Chlronomid 300 0.00007
Simulldae 200 0.00033
Baetidae (small) 150 0.00010
Baetldae (lame) 100 0~00052
Caenidae 100 0.00030
Heptaqenldae 50 0.00174
l.eptophlebldae 50 0.00128
Teloqonadldae 50 0.00082
Corixidae 10 0.00328
Naucoridae 20 0.00735
Meualootera 5 0.04964
Aeshnid (small) 16 0.00061
Aeshnld (large) 8 0.06156
Gomphidae (small) 50 0.00029
Gomphidae (large) 10 0.03226
Libellulldae (small) 25 0.00081
IIbeliulidae (larqe) 10 0.05179
Zygoptera (small) 50 0.00030
Zygoptera (large) 20 0.00390
Athripsodes 40 0.00064
C. afra 50 0.00190
Table 4a Inter-year variation of taxon abundance for each habitat type as indicated byANOSIM analysis. (m. veg =marginal vegetation)
Comparison r pSand yearl/year2 0.177 <0.01
m. veg yearl/year2 0.161 <0.01Cobble yearl/year2 0.171 <0.01
Table 4b Inter-habitat variation of taxon abundance for each year as indicated byANOSIM analysis.
comparison r pYear 1 sand/m. veg 0.613 <0.01Year 1 sand/cobble 0.602 <0.01Year 1 m.veg/cobb1e 0.367 <0.01Year 2 sand/m. veg 0.660 <0.01Year 2 sand/cobble 0.746 <0.01Year 2 m.veg/cobb1e 0.367 <0.01

Tab
le5.
Tax
aco
ntri
buti
ngto
>75
%cu
mul
ativ
esi
mil
arit
yfo
rth
ree
habi
tats
.Y
ear
Ian
d2.
Ha
bita
t(%
sim
ilari
tyb
etw
ee
nY
ear1
Ha
bit
at(
%si
mila
rity
Yea
r2sa
mp
les)
be
twe
en
sam
ple
s)
San
d(3
5.71
%)
Ab
un
da
nce
Sim
ilari
ty%
Sim
/SO
Co
ntr
ibu
tio
n%
San
d(4
6.56
%)
Ab
un
da
nce
Sim
ilari
ty%
Sim
/SO
Co
ntr
ibu
tio
n%
(av.
nu
mb
er/
sam
ple
)(a
vera
ge
)(a
v.n
um
be
r/sa
mp
le)(
aver
age)
Bae
tidae
(larg
e)6.
838.
591.
1424
.07
Bae
tidae
(larg
e)8.
639.
182.
4719
.72
Gom
phid
ae(s
mal
l)8.
616.
810.
9819
.07
Chi
rono
mid
ae55
.29
9.08
1.62
19.5
Chi
rono
mid
ae28
.78
6.66
1.08
18.6
6G
omph
idae
(larg
e)4.
797.
21.
9115
.46
Gom
phid
ae(la
rge)
2.17
4.63
0.8
12.9
7E
lmid
ae(la
rvae
)6.
134.
621.
199.
92E
lmid
ae(la
rvae
)2
1.72
0.54
4.83
Gom
phid
ae(s
mal
l)20
.71
4.16
0.92
8.93
Cae
nida
e7.
043.
530.
837.
59M
arg
ina
lveg
(46.
54%
)M
arg
ina
lveg
(56.
56%
)B
aetid
ae(la
rge)
50.1
10.2
15.
2321
.94
Chi
rono
mid
ae14
4.96
7.65
2.62
13.5
3C
hiro
nom
idae
28.5
57.
862.
7116
.9B
aetid
ae(la
rge)
88.8
37.
315.
4812
.92
Cae
nida
e9.
454.
011.
288.
62C
aeni
dae
23.2
24.
11.
877.
24Z
ygop
tera
(larg
e)6.
383.
61.
217.
74Z
ygop
tera
(larg
e)9.
783.
932.
716.
95B
aetid
ae(s
mal
l)4
2.79
0.92
5.99
Zyg
opte
ra(s
mal
l)24
.78
3.86
1.94
6.82
Sim
uliid
ae24
.93
2.74
0.78
5.88
Elm
idae
(larv
ae)
6.7
2.99
1.56
5.29
Ath
ripso
dini
4.03
1.9
0.68
4.08
Libe
llulid
ae(s
mal
l)8.
392.
861.
365.
05V
eliid
ae1.
411.
680.
693.
62B
aetid
ae(s
mal
l)15
.87
2.79
1.06
4.93
Zyg
opte
ra(s
mal
l)4.
831.
380.
582.
97S
imul
iidae
15.6
12.
611.
154.
62Li
bellu
lidae
(larg
e)3
2.41
1.39
4.25
Vel
iidae
4.09
1.8
0.9
3.17
Ath
ripso
dini
10.3
51.
50.
732.
65C
ob
ble
(47.
33%
)C
ob
ble
(57.
8%)
Bae
tidae
(larg
e)46
.94
10.5
13.
9822
.2B
aetid
ae(la
rge)
60.8
47.
286.
3212
.6C
hiro
nom
idae
32.8
46.
541.
8513
.82
Chi
rono
mid
ae60
.04
6.36
4.23
11.0
1S
imul
iidae
20.1
65.
651.
5711
.94
Elm
idae
(larv
ae)
29.6
5.8
4.28
10.0
3C
.afr
a7.
714
1.41
8.46
C.a
fra
35.6
5.24
2.67
9.07
Bae
tidae
(sm
all)
5.68
3.04
0.95
6.42
Sim
uliid
ae39
.44.
682.
018.
09E
lmid
ae(la
rvae
)4.
742.
650.
965.
59Le
ptop
hleb
idae
13.0
43.
811.
956.
59Le
ptop
hleb
idae
4.48
2.57
0.95
5.43
Bae
tidae
(sm
all)
12.4
43.
351.
465.
8H
epta
geni
dae
3.87
2.11
0.81
4.45
Elm
idae
(adu
lt)19
.64
3.12
1.32
5.39
Lim
nich
idae
11.8
42.
81.
414.
84H
epta
geni
dae
8.64
2.44
1.05
4.23

Table 6a Summary of the pairwise analysis of significantly dissimilar sample groupsfrom different invasion conditions within the same habitat (year 2). IS1: no invasion;IS2: invasive plants only;"IS3: invasive fish only; IS4: invasive plants and fish
Taxa, ranked in order of contribution toInvaded Status p r Dissimilarity dissimilarity between the sites (only the
ton four are listed)
Cobble ISlIIS2 0.04 0.583 59.3%Caenidae, E1midaeadult, Ecnomidae,simuliidae.
Cobble IS1/IS3 0.01 0.573 58.9%Heptageniidae, Aeshnidae small,Chironomidae, Gyrinidae larvae.
Cobble IS 1/IS4 0.02 0.417 56.0%Caenidae, Heptageniidae, Aeshnidae small,Elmidae adult.
Cobble IS211S3 0.02 0.419 55.25%Simuliidae, Baetidae small, Ecnomidae,Tipulidae.
Marginal vegatation0.03 0.386 54.0%
Simuliidae, Chironomidae, Athripsodes,IS1IlS3 Baetidae small.
Table 6b Summary of the pairwise analysis of significantly dissimilar sample groupsfrom different invasion conditions within the same habitat (year 1).
Invaded status p r DissimilarityCobble IS1IIS3 0.01 0.641 53.5%Cobble ISlIIS4 0.02 0.504 60.8%
Marginal vegetation0.02 0.736 51.6%
IS3/IS4
Table 7 Similarity analysis of taxa biomass from samples within habitats anddifferent invasion conditions (year 2).
Invaded status p r dissimilarityTaxa, ranked in order of contribution todissimilarity
Cobble IS1/IS2 0.05 0.444 30.52%Megaloptera, Elmidae adult, Simuliiidae,Aeshnidae large.
Cobble IS111S3 0.01 0.64 38.04%Libellulidae large, Megaloptera, Aeshnidae large,Heptageniidae.
Cobble IS2/IS3 0.02 0.567 36.72%Libellulidae large, Aeshnidae large, Megaloptera,Simuliidae.
M.veg IS1IIS3 0.01 0.437 41.19%Libellulidae large, Gomphidae large, Athripsodes,C. afra, Naucoridae.
Table 8 Similarity analysis of the biomass ofFFGs from samples within habitats anddifferent invasion conditions (year 2).
Invaded status p r FFG, ranked in order ofcontribution to dissimilarity
AllFFGsCobble IS1IIS2 0.05 0.437 P2, Grazer2/scraper 1, PI, DF2Cobble IS111S3 0.02 0.656 P2, PI, DF1, Grazer2/scraperlCobble IS211S3 0.02 0.622 P2,P1Herbivores onlyCobble IS1/IS2 0.01 0.561Cobble IS111S3 0.02 0.726Predators onlyCobble IS1IIS2 0.01 0.82Cobble IS111S2 0.04 0.544M. vegetation IS111S3 0.01 0.519

Appendix 1 Taxa of the Rondegat River. September 2003 - January 2005.Key: Tabanidae/Tany = Tabanidae/Tanypodidae (Diptera); C. afra = Cheumatopsyche afra, M.capense =Macrostemum capense (Trichoptera); Noto/Nepidae =notonectidae/Nepidae (Hemiptera)For key to FFGs, see Table 2.
YEAR 1 YEAR 2Taxa sand sand m.veg m.veg cobble cobble FFG sand
~~~~ ~..v~~m.veg cobble cobble
(av.) ISm lav.)- (SD) (av.) ISDl lav.l avo ISDi' (av.) ISDlChlronomldae 28.78 63.47 28.55 31.24 32.84 56.80 FF 55.29 83.33 144.96 102.50 60.04 86.54Simulildae 0.67 5.68 24.93 69.74 20.16 25.70 P1 2.04 3.47 15.61 21.40 39.40 43.35Athericldae 0.06 0.71 0.45 1.33 0.68 1.62 FF 0.08 0.41 0.26 0.62 0.20 0.65Culicidae 0.00 0.00 0.72 1.96 0.39 1.65 S1 0.58 2.86 3.22 9.83 0.16 0.80Blephariceridae 0.00 0.00 0.21 0.82 0.77 2.16 FF 0.00 0.00 0.00 0.00 0.36 1.08Dixldae 0.06 0.23 0.93 2.42 0.00 0.00 G2 0.00 0.00 0.61 1.03 0.04 0.20Ceratopogonidae 0.22 0.63 0.24 0.91 0.23 0.80 P2 0.50 0.88 0.70 1.18 0.52 1.19Empldldae 0.11 0.32 0.03 0.19 0.16 0.52 P2 0.00 0.00 0.48 1.12 0.68 1.35TabanldaelTany 0.00 0.00 0.03 0.19 0.19 0.54 P2 0.08 0.41 0.09 0.42 0.68 1.11Tlpulidae 0.00 0.00 0.07 0.37 0.16 0.45 P2 0.33 0.76 0.30 0.82 1.00 1.71Muscidae 0.06 0.23 0.03 0.19 0.06 0.36 0.08 0.41 0.13 0.46 0.16 0.47Syrphldae 0.00 0.23 0.03 0.19 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00
Baetidae (large) 6.83 8.66 50.10 42.24 46.94 32.53 DF1 8.63 5.66 88.83 72.27 60.84 34.66Caenldae 1.39 2.22 9.45 13.36 6.16 14.26 DF2 7.04 9.35 23.22 26.33 5.52 9.21
Telogonadidae 0.33 1.38 0.24 0.83 9.39 23.57 DF2 0.00 0.00 0.26 0.75 0.96 3.26
Baetidae (small) 0.33 1.02 4.00 4.86 5.68 7.38 DF1 3.96 6.02 15.87 18.11 12.44 12.58
Leptophlebidae 0.06 0.23 0.55 1.57 4.48 7.93 DF1 0.46 1.14 1.65 2.04 13.04 13.65
Hentauenldae 0.06 0.23 0.69 2.98 3.87 8.44 G2 0.04 0.20 0.00 0.00 8.64 12.64
Gomphldae (small) 8.61 13.53 0.10 0.56 2.35 9.33 P2 20.71 41.04 1.48 2.57 3.44 8.50
Gomphidae (large) 2.17 2.01 0.31 1.14 0.77 1.50 P2 4.79 3.24 0.57 1.08 0.20 0.50Zygoptera (large) 0.06 0.23 6.38 8.81 0.10 0.54 P2 0.00 0.00 9.78 8.03 0.04 0.20
Zygoptera (small) 0.11 0.46 4.83 10.29 0.16 0.73 P2 0.33 0.76 24.78 52.41 0.36 0.76
Libellulldae(small) 2.17 3.17 2.21 4.51 0.74 1.32 P2 2.42 5.11 8.39 8.41 0.84 1.95
Libellulldae(large) 0.78 2.10 1.10 1.63 0.32 0.94 P2 0.38 0.77 3.00 2.34 0.84 1.60Aeshnldae (large) 0.00 0.00 0.97 2.65 0.74 1.67 P2 0.00 0.00 0.09 0.42 0.72 1.67Aeshnldae Ismalll 0.17 0.50 0.72 1.56 0.45 1.55 P2 0.58 1.38 2.87 4.08 9.16 14.40
C. afra 0.11 1.00 1.97 3.78 7.71 8.17 P1 1.08 1.67 4.00 8.33 35.60 34.20Athripsodlni 0.89 1.63 4.03 10.46 0.48 1.00 DF2 0.75 1.19 10.35 17.59 0.44 1.26Hydroptilldae 0.94 2.85 1.69 5.66 1.32 5.39 S1 0.75 1.51 5.65 8.65 4.40 19.13Leptecho/cerus 0.50 1.17 1.48 2.08 0.29 0.53 Shr 0.50 1.06 1.91 3.42 0.32 0.69Ecnomidae 2.00 5.20 0.07 0.26 0.45 1.09 DF1 0.58 2.17 0.04 0.21 1.36 2.61Oecetis 0.06 0.23 0.55 1.57 0.45 1.09 P2 0.17 0.82 1.30 1.84 0.92 1.91Philopotamidae 0.00 0.00 0.14 0.74 0.32 1.14 FF 0.00 0.00 0.09 0.42 1.12 2.32
Parecnomlna 0.00 0.00 0.14 0.58 0.10 0.40 0.00 0.00 0.04 0.21 0.08 0.40
Sericostomatldae 0.06 0.50 0.07 0.37 0.13 0.34 Sh 0.42 1.06 0.22 0.60 0.32 0.75
Macrostemum 0.00 0.00 0.00 0.00 0.03 0.18 P2 0.00 0.00 0.17 0.83 1.00 1.91
Barbarochthonldae 0.00 0.00 0.00 0.00 0.03 0.18 0.00 0.00 0.00 0.00 0.00 0.00
Leptocerldae sp. 0.00 0.00 0.00 0.00 0.03 0.18 0.00 0.00 0.13 0.63 0.16 0.55
Glossosomatidae 0.00 0.00 0.00 0.00 0.00 0.00 0.08 0.41 0.52 1.24 2.64 7.65
Petrothrincidae 0.00 0.00 0.00 0.00 0.00 0.00 0.21 0.59 0.57 0.99 1.60 1.98
Polvcentropodldae 0.00 0.00 0.00 0.00 0.00 0.00 0.50 1.06 1.91 3.42 0.32 0.69
Elmidae (larvae) 2.00 3.16 1.03 1.59 4.74 5.55 S1 6.13 9.01 6.70 6.85 29.60 19.03
Elmidae (adult) 0.06 0.23 1.86 5.57 3.03 4.96 S2 0.58 1.06 4.30 12.14 19.64 27.53
Helodidae 0.44 1.07 1.07 1.89 0.97 2.02 G1 2.83 8.32 1.52 3.87 2.56 3.27
Hydrophllldae 0.00 0.00 0.21 0.56 0.45 0.93 S1 0.17 0.82 0.30 0.93 3.28 4.90
Limnlchldae 0.11 0.46 0.34 0.90 1.06 1.48 G1 0.88 2.27 3.04 4.55 11.84 18.14
Gyrlnldae (larvae) 0.22 0.54 0.34 1.01 0.71 2.10 P2 0.04 0.20 2.39 5.10 2.32 3.08
Dytlscldae (adult) 0.06 0.23 0.97 2.77 0.00 0.00 P2 0.00 0.00 0.74 1.39 0.00 0.00
Dytiscidae (larvae) 0.17 0.69 0.34 1.34 0.00 0.00 P2 0.42 1.67 2.78 4.49 0.36 0.95
Hydraenldae 0.00 0.00 0.07 0.37 0.26 0.73 S2 0.00 0.00 0.13 0.46 0.32 0.80
Gvrlnldae (adult) 0.06 0.23 0.07 0.37 0.00 0.00 P2 0.08 0.41 0.52 1.24 2.64 7.65
Corlxldae 3.22 7.58 1.00 1.87 0.48 1.15 P1 0.50 1.69 0.48 1.04 0.16 0.55
Naucorldae 1.06 2.83 1.86 5.22 0.26 0.82 P2 0.25 1.22 3.30 7.21 0.16 0.47
Vellldae 0.06 0.32 1.41 1.78 0.32 0.87 P1 0.17 0.56 4.09 4.94 0.16 0.55
Mesovellldae 0.00 0.00 0.14 0.74 0.03 0.18 P2 0.00 0.00 0.09 0.42 0.04 0.20
Gerridae/Saldidae 0.00 0.00 0.07 0.26 0.00 0.00 P2 0.00 0.00 0.04 0.21 0.00 0.00
Noto/NeDidae 0.00 0.00 0.03 0.19 0.00 0.00 P2 0.00 0.00 0.30 0.70 0.00 0.00
Oligochaeta 0.89 2.79 0.83 1.85 1.68 5.58 G2 0.17 0.48 1.57 4.41 1.08 2.56
Lepidoptera 0.00 0.00 0.48 1.02 0.03 0.18 G1 0.08 0.41 0.87 2.22 0.32 0.95
Hydrachnellae 0.00 0.00 0.14 0.52 0.29 0.64 0.21 0.59 0.57 0.99 1.60 1.98Megaloptera 0.00 0.00 0.07 0.37 0.26 0.63 P2 0.00 0.00 0.00 0.00 1.44 1.94Plecootera 0.00 0.00 0.17 0.54 0.10 2.72 Shr 0.17 0.82 0.04 0.21 0.40 0.91

Appendix 2The average number of taxa per sampling occasion at each site (year 2).lSI: no invasion; IS2: invasive plants only; IS3: invasive fish only; IS4: invasiveplants and fish.
Key: Tabanidae/Tany =Tabanidae/Tanypodidae (Diptera); C. afra =Cheumatopsyche afra,M. capense = Macrostemum capense (Trichoptera); Noto/Nepidae = notonectidae/Nepidae(Hemiptera)
Site number and invasion status (IS)Taxa 1 (181) 2 (181) 3 (182) 4 (182) 5 (184) 6 (183) 7 (183) 8 (184)Chironomidae 50.44 44.89 94.11 77.00 122.44 155.56 128.50 28.88Simuliidae 16.44 22.44 18.67 2.67 22.33 41.22 12.88 20.50Culicidae 0.33 0.33 0.00 0.00 2.22 6.89 0.50 0.00Ceratopogonidae 0.89 0.33 1.33 0.89 0.00 0.44 0.25 0.50Tipulidae 0.89 0.22 0.22 0.56 0.22 1.33 1.13 0.00Empldidae 0.67 0.00 0.89 0.00 0.56 0.67 0.00 0.38TabanidaelTany 0.22 0.22 0.11 0.67 0.00 0.22 0.50 0.50Dixldae 0.11 0.33 0.22 0.00 0.33 0.22 0.13 0.38Athericidae 0.22 0.11 0.33 0.00 0.44 0.22 0.13 0.00Blephariceridae 0.00 0.00 0.56 0.00 0.22 0.00 0.00 0.25Muscidae 0.00 0.00 0.22 0.00 0.33 0.00 0.25 0.25Baetidae (large) 44.44 30.11 51.11 39.78 51.22 51.67 119.63 47.88Caenidae 4.56 7.11 25.89 8.33 29.78 6.00 7.00 6.25Baetidae (small) 17.56 3.11 2.22 5.56 8.44 25.56 14.50 11.25Leptophlebidae 1.56 2.89 3.67 7.89 4.22 9.78 9.50 2.13Heptagenidae 4.11 11.89 1.67 1.33 3.89 0.22 1.00 0.00Teloqonadldae 1.33 0.33 0.00 0.00 0.00 0.00 0.00 0.00Gomphidae (small) 4.44 0.44 7.11 1.89 5.44 1.78 18.75 34.63Zygoptera (small) 1.33 1.11 6.89 4.00 32.00 7.78 7.25 6.38Aeshnidae (small) 0.44 0.67 1.78 3.00 3.33 8.44 6.25 12.50Libellulidae (small) 1.33 0.67 5.11 3.56 5.56 7.56 6.25 1.00Zygoptera (large) 2.56 1.78 2.11 2.22 3.44 1.56 7.88 5.00Gomphidae (large) 1.33 1.78 2.78 2.00 1.78 1.11 1.38 3.00L1bellulidae(large) 0.78 0.89 1.44 0.56 2.44 3.00 1.75 0.38Aeshnidae (iargef 0.00 0.00 0.00 0.22 0.22 0.11 1.50 0.25C. afra 11.44 15.00 7.33 6.22 6.00 30.89 11.75 27.25Athrip/Setodes 0.44 0.67 6.33 2.11 9.22 1.11 9.75 1.25Hydrophilidae 3.22 2.11 1.00 0.67 0.33 2.22 0.38 0.25leptecho/cerus 0.33 0.44 0.89 2.44 1.44 0.67 0.50 0.50Oecetis 0.67 1.22 0.00 1.22 2.22 0.00 0.50 0.50Ecnomidae 0.00 1.11 2.11 1.44 0.56 0.22 0.00 0.00Philopotamidae 0.22 0.22 0.67 0.11 1.33 0.78 0.00 0.00Macrostemum 0.00 0.11 0.22 0.67 0.00 1.78 0.50 0.00Sericostomatidae 0.33 0.56 0.00 0.22 1.00 0.00 0.50 0.00Polycentropodidae 0.00 0.00 0.00 0.44 1.22 0.00 0.00 0.00Lepta, unplaced 0.33 0.44 0.00 0.00 0.00 0.00 0.00 0.00Glossosomalidae 0.00 0.00 0.00 0.00 0.56 0.00 0.00 0.00Parecnomia 0.11 0.00 0.00 0.00 0.22 0.00 0.00 0.00Petrothrincidae 0.00 0.22 0.00 0.00 0.00 0.00 0.00 0.00Elmidae larvae 21.56 15.78 21.67 7.11 9.00 19.89 12.13 10.75Elmidae adult 33.11 10.22 1.89 0.56 1.89 10.44 3.00 7.00Limnichidae 3.11 2.44 15.89 4.89 0.44 12.00 3.50 1.25Hydroptilidae 0.78 0.00 2.89 2.89 2.44 14.22 5.00 1.13Helodidae 7.33 2.00 5.00 1.89 0.22 1.67 0.25 0.00Gyrinidae larvae 0.89 1.33 0.33 0.67 0.44 2.44 6.25 1.13Dytiscidae larvae 0.11 0.22 0.11 0.22 1.78 3.11 3.13 1.00Gyrinidae adult 0.22 0.00 0.00 0.67 0.00 0.22 3.63 5.13Dytiscidae adult 0.00 0.00 0.11 0.00 0.33 0.67 0.00 0.88Hvdraenidae 0.56 0.00 0.00 0.00 0.00 0.00 0.13 0.50Veliidae 1.33 0.22 3.22 3.56 0.11 0.00 1.25 2.00Naucoridae 1.00 1.89 4.67 0.89 0.44 0.22 0.50 0.00Corixidae 0.00 0.44 0.00 0.22 0.56 1.78 0.00 0.00Noto/Nepidae 0.00 0.22 0.00 0.22 0.11 0.00 0.00 0.25Mesoveliidae 0.00 0.11 0.00 0.00 0.22 0.00 0.00 0.00Gerrid/Saldidae 0.00 0.00 0.00 0.00 0.00 0.00 0.13 0.00Oligochaeta 0.44 0.11 0.78 0.44 0.22 1.11 4.50 0.38Hydrachnellae 0.44 0.56 1.00 0.56 0.78 1.00 0.50 1.88Megaloptera 0.44 0.78 0.22 0.00 1.11 1.22 0.00 0.00Lepidoptera 0.89 0.33 2.00 0.00 0.00 0.00 0.13 0.00Plecoptera 0.67 0.11 0.00 0.00 0.56 0.00 0.25 0.00Total 246.00 190.56 306.78 202.44 345.67 439.00 415.00 245.25