Embed Size (px)
Citation preview

PHYSIOLOGY
© 2006 Elsevier Ltd179ANAESTHESIA AND INTENSIVE CARE MEDICINE 7:6
The mammalian immune system has evolved to provide protec-tion against infectious agents (pathogens). Although the immune system may provide protection against tumours, the main evolu-tionary driving force has been the battle with pathogens.
Innate and specific immunityThe immune system has historically been divided into the innate and specific systems. The innate immune system includes:• physical and chemical barriers to infection (e.g. skin, hydro-
chloric acid in the stomach)• cells (e.g. macrophages, neutrophils, mast cells)• soluble factors (e.g. the complement proteins, acute-phase
proteins). The cellular components of the specific immune system are the lymphocytes. Lymphocyte precursors originate from the bone marrow and some differentiate in the thymus into T lymphocytes which are responsible for cellular immunity. Other lymphocyte precursors differentiate into B lymphocytes in the fetal liver and after birth in the bone marrow. B lymphocytes are responsible for humoral immunity. Most of the production of lymphocytes occurs during fetal and neonatal life, but there is a slow continu-ous production of new lymphocytes from stem cells in adults. After differentiation, the lymphocytes migrate to the lymph nodes and other lymphoid tissue. The two main types of T cells are CD4 and CD8 T cells. Although morphologically indistinguishable, the three types of lymphocyte perform different functions. One of the main features distinguishing specific from innate immunity is that specific immunity is not present before infection. Following initial infection with a pathogen it takes about 4 days to produce antibody against it. The innate immune system used to be regarded as a primitive defence system of limited effectiveness, found in vertebrates and invertebrates. The specific immune system is found in vertebrates only and was thought to be the main defence mechanism. It is now known that the innate immune system does more than provide the first line of defence and plays an important role in determin-ing the type of specific immune response that is mounted against infectious agents.
Self/non-self discriminationBoth the innate and specific immune systems can distinguish between foreign entities (e.g. pathogens) and the body’s cells
and tissues. Recognition is achieved by molecules of the immune system that bind only to foreign molecules associated with patho-gens and not to self-molecules.
Innate immune system: cells and proteins of the innate immune system can recognize certain molecules on the surface of patho-gens (pathogen-associated molecular patterns – PAMPs) as foreign. PAMPs are large molecules with a significant non-protein component and examples include lipopolysaccharides, peptido-glycans, lipotechoic acids and mannans. They are found only on microbes and are essential for microbial survival, therefore they cannot stop manufacturing them as a way of avoiding recognition by the innate immune system. The cell receptors that recognize PAMPs are called pattern-recognition receptors and include toll-like receptors and lectins such as the mannose receptor (Figure 1). Soluble factors recognizing PAMPs include mannose-binding lectin and C-reactive protein. There is a limit to the number of receptors that can evolve to recognize foreign PAMPs and, given the quicker replication of pathogens compared with mammals, some pathogens will evolve ways of escaping recognition by the innate immune system. For example, avirulent streptococci are recognized and eliminated by the innate immune system and therefore do not cause disease. However, virulent streptococci synthesize a waxy carbohydrate outer coat that is not recognized by the innate immune system and therefore to eliminate them a specific immune response is required.
Specific immune system B lymphocytes and antibody – B lymphocytes have antibody molecules on their cell surface. Antibody molecules consist of two identical heavy chains and two identical light chains forming a Y-shaped structure (Figure 1). Each heavy chain molecule consists of a variable (V) region at the N-terminal amino acid end and a constant region at the C-terminal end consisting of three domains. Light chains have a variable region and a constant region consisting of one domain. The variable regions of the heavy and light chain fold together to form a structure that is responsible for recognizing and binding to molecules on a microbial cell surface or soluble protein made by a microbe. The molecules to which antibody binds are called antigens. Special molecular mechanisms cause part of the gene coding for the V region of the antibody to be randomly altered. Therefore, each antibody molecule has a different amino acid sequence in the part of the molecule that recognizes antigen, and thus each antibody can bind to some antigens but not others. Enzyme digestion can split antibodies into an antigen-binding frag-ment (Fab) and a crystallizable fragment (Fc). There are a number of different classes of antibody (IgM, IgG, IgA, IgE, IgD). The class is determined by the Fc region of the heavy chain. There are also two types of light chain called κ and λ. Each class of heavy chain can associate with either κ or λ light chains. The class of heavy chain determines the function of the antibody, the light chain type has no effect on function. T lymphocytes have receptors for antigen on their surface. Although related to antibody, the T-cell receptor for antigen (TcR) is coded for by different genes. The TcR consists of two transmem-brane glycoprotein chains (α and β), linked by di-sulphide bonds. The chains have a constant region proximal to the cell surface and a V region distal to the cell surface. Different V regions are generated
Peter J Wood is Lecturer in Immunology in the Faculty of Life Sciences,
University of Manchester, UK. He obtained a BSc in Pharmacology from
Liverpool University and a PhD in Immunology from London University.
His research interests focus on immunological events in insulin-
dependent diabetes mellitus and in graft rejection.
The immune system: recognition of infectious agentsPeter J Wood
PHYSIOLOGY
© 2006 Elsevier Ltd180ANAESTHESIA AND INTENSIVE CARE MEDICINE 7:6
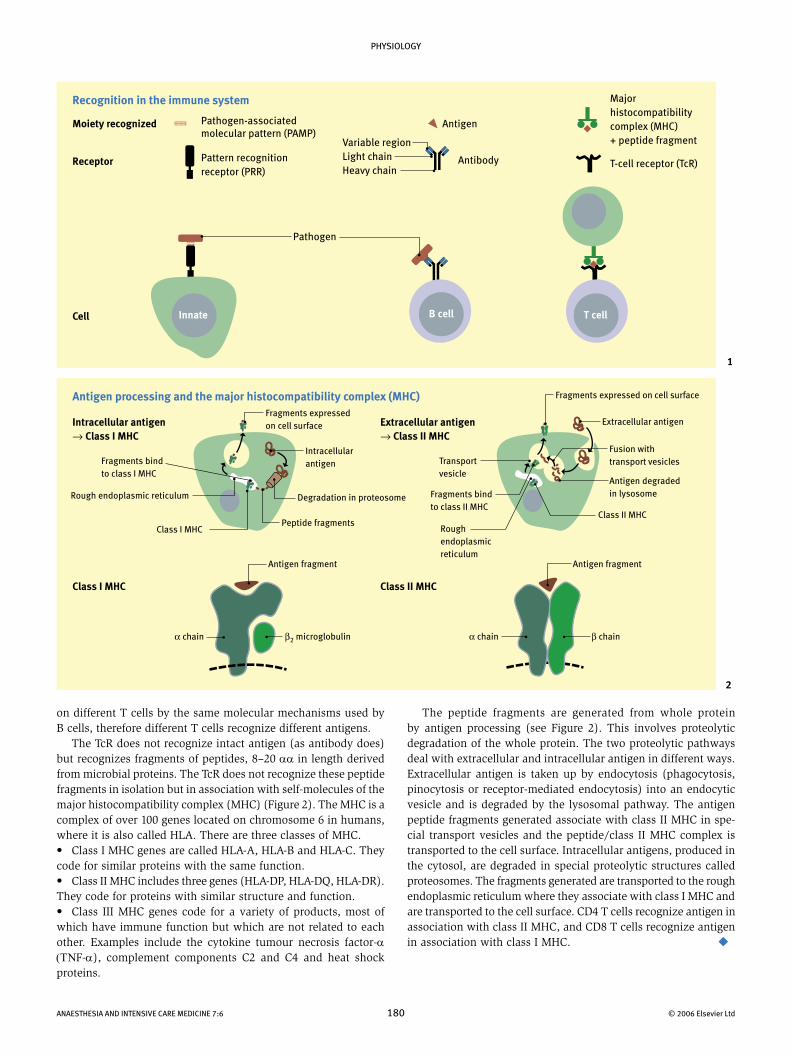
on different T cells by the same molecular mechanisms used by B cells, therefore different T cells recognize different antigens. The TcR does not recognize intact antigen (as antibody does) but recognizes fragments of peptides, 8–20 αα in length derived from microbial proteins. The TcR does not recognize these peptide fragments in isolation but in association with self-molecules of the major histocompatibility complex (MHC) (Figure 2). The MHC is a complex of over 100 genes located on chromosome 6 in humans, where it is also called HLA. There are three classes of MHC.• Class I MHC genes are called HLA-A, HLA-B and HLA-C. They code for similar proteins with the same function. • Class II MHC includes three genes (HLA-DP, HLA-DQ, HLA-DR). They code for proteins with similar structure and function.• Class III MHC genes code for a variety of products, most of which have immune function but which are not related to each other. Examples include the cytokine tumour necrosis factor-α (ΤΝF-α), complement components C2 and C4 and heat shock proteins.
1
Recognition in the immune system
Moiety recognized Antigen
Antibody
Pathogen-associated molecular pattern (PAMP)
Pattern recognition receptor (PRR)
Receptor
Cell
Pathogen
Innate B cell T cell
Variable regionLight chainHeavy chain
Major histocompatibility complex (MHC) + peptide fragment
T-cell receptor (TcR)
2
Antigen processing and the major histocompatibility complex (MHC)
Intracellular antigen → Class I MHC
Extracellular antigen → Class II MHC
Class I MHC Class II MHC
Fragments expressedon cell surface
Fragments expressed on cell surface
Fusion with transport vesiclesFragments bind
to class I MHC
Rough endoplasmic reticulum
Class I MHCPeptide fragments
Degradation in proteosome
Intracellular antigen
Antigen fragment
β2 microglobulinα chain
Antigen fragment
α chain β chain
Transport vesicle
Fragments bind to class II MHC
Rough endoplasmic reticulum
Extracellular antigen
Antigen degraded in lysosome
Class II MHC
The peptide fragments are generated from whole protein by antigen processing (see Figure 2). This involves proteolytic degradation of the whole protein. The two proteolytic pathways deal with extracellular and intracellular antigen in different ways. Extracellular antigen is taken up by endocytosis (phagocytosis, pinocytosis or receptor-mediated endocytosis) into an endocytic vesicle and is degraded by the lysosomal pathway. The antigen peptide fragments generated associate with class II MHC in spe-cial transport vesicles and the peptide/class II MHC complex is transported to the cell surface. Intracellular antigens, produced in the cytosol, are degraded in special proteolytic structures called proteosomes. The fragments generated are transported to the rough endoplasmic reticulum where they associate with class I MHC and are transported to the cell surface. CD4 T cells recognize antigen in association with class II MHC, and CD8 T cells recognize antigen in association with class I MHC.




![Mathematical modelling of immune response in tissues · infectious agents [73]. PRRs are found on cells throughout the immune system but are best described on cells of the innate](https://img.dokumen.tips/doc/110x75/5f1fa59ca635df45c76f0187/mathematical-modelling-of-immune-response-in-tissues-infectious-agents-73-prrs.jpg)
























