Embed Size (px)
Citation preview

AQUATIC CONSERVATION: MARINE AND FRESHWATER ECOSYSTEMS, VOL. 2, 309-324 (1992)
The effects of r@arian management and physicochemistry on macroinvertebrate feeding guilds and community
structure in upland British streams
S. D. RUNDLE, E. CLARE LLOYD and S. J . ORMEROD
Catchment Research Group, School of Pure and Applied Biology, University of Wales College of CardvJ CardvJ UK
ABSTRACT
1. Patterns in the community structure and feeding guild composition among macroinvertebrates in the margins and riffles of 66 streams in upland Wales and Scotland were appraised and related to site chemistry, physiography and riparian management. We aimed to assess the effectiveness of riparian strips of broadleaves and moorland/grassland in protecting stream communities and their trophic function during commercial forestry.
2. The composition of macroinvertebrate communities in margins and riffles most strongly reflected stream p H and aluminium concentrations; acidic riffles contained fewest species of all major macroinvertebrate groups. However, the character of marginal habitats had some influence on communities in stream margins through the presence of broadleaf trees or moorland/grassland vegetation in the riparian zone. Even at low pH, streams with moorland riparian zones had different macroinvertebrates from those with pure conifer.
3. Stream chemistry was important in explaining patterns among feeding guilds. Species richness and abundances of grazers and filterers were reduced at acidic sites with high aluminium concentrations. The number of shredder species was significantly lower in riffles a t low p H than at high pH.
4. In contrast, marginal habitat structure was of overriding importance in explaining patterns among collectors and predators. These functional groups were most abundant at sites with riparian zones of moorland and grassland, but scarce in streams where the bankside vegetation consisted of coniferous or broadleaved trees. Total grazer abundances were lowest a t sites with riparian zones of pure conifer.
5 . We conclude that riparian management can modify effects of forestry on the trophic dynamics and structure of the macroinvertebrate communities of upland streams, but the effect operates within a strong gradient due to acidity. Thus, the array of macroinvertebrates which may benefit from riparian management in forest streams is liable to be restricted by low pH.
6. We discuss the wider ramifications of riparian management for the conservation of streams during commercial forestry. In particular we assess if management proposals set out by the forestry industry are supported. We also discuss the problem of scale when considering measures implemented for stream conservation, emphasizing that the control of problems such as acidification may only be possible at a regional level.
INTRODUCTION
Recent interest has centred on the role of physical factors, such as flow and habitat structure, in explaining the organization of macroinvertebrate communities in streams (Statzner et al., 1988; Rutt et al., 1989; Hildrew
1052-7613/92/040309-16$13.00 01992 by John Wiley & Sons, Ltd.
Received 29 May 1992 Accepted 2 October 1992

310 S . D. RUNDLE. E. C. LLOYD AND S . J. ORMEROD
et al., in press). Chemical features such as acid-base status are also important (Townsend et al., 1983; Wade et al., 1989). Catchment land use may modify all these factors and it is important that any resulting changes do not adversely affect the structure and function of stream ecosystems in ways that jeopardize their conservation value.
In Britain, much interest has focused on the consequences of extensive afforestation for upland streams. Plantations of exotic conifers can accentuate the acidification of fresh waters in catchments with base-poor rocks and soils (Ormerod et al., 1989). Changes in the physical and energetic characteristics of streams, such as temperature (Weatherley and Ormerod, 1990), flow rate and erosion (Robinson, 1980) may also occur. Consequences for stream habitat structure have also been described, especially along stream margins (Rutt et al. , 1989).
In an attempt to counteract some of the effects of afforestation on fresh waters, management practices- such as the use of riparian ‘buffer’ strips-have been implemented in Britain on a national scale (Forestry Commission, 1991). We have now appraised the efficacy of these techniques in upland Wales and Scotland, showing that riparian management influenced stream habitat structure, but that effects on macroinvertebrate assemblages were subsumed by the impact of acidity (Ormerod et al. , in press). These previous assessments included only a preliminary investigation of effects at the community level and ignored the potential effects of land use on trophic structure (Hawkins et al., 1982; Townsend el al., 1983; Bunn, 1986). Such trophic effects may be important: conifer litter is a poor source of allochthonous energy (Sedell et al., 1975; Murphy et al., 1981; Feminella et al., 1989). Also, acidity and increased shading in conifer streams are likely to affect food quality for grazers and shredders (e.g. Winterbourn et al., 1985; Groom and Hildrew, 1989). The conservation ramifications of such functional changes are poorly known in streams generally.
Here, we extend our previous study (Ormerod et al., in press) by assessing the influences of chemistry, physical variables and riparian management on the taxonomic and trophic structure of macroinvertebrate communities in 66 streams in upland Wales and Scotland.
STUDY SITES
The streams were situated throughout the uplands of Scotland and Wales (Ormerod et al., in press). They were small (width < 2.5 m), 1-3 order tributaries representing a wide range of chemical conditions typical of upland Britain generally (pH 4.4-6.9, calcium 0.5-20 mg I - I ) . Six types of riparian management included: conifer forest (CON, 12 sites); conifer forest with a cleared buffer strip (CLR, 11 sites); conifer forest with a moorland/grassland buffer strip from the planting stage (BUF, 12 sites); conifer forest with a deciduous buffer strip left at the planting stage (BRD, 8 sites); semi-natural broadleaf woodland (SNW, 9 sites); semi-natural moorland/grassland (=moorland, in text) (MOR, 14 sites). In all cases, conifers were beyond the stage of canopy closure, crops being at least 20 years old. A more detailed description of the sites is given by Ormerod et al. (in press).
METHODS
Macroinvertebrate sampling and trophic classification
Animals were sampled during October and November 1990 from the riffle and margin habitat at each site; one 2-minute kick sample (net mesh 900pm) was taken from each habitat, and animals were preserved in the field. Previous studies, carried out as part of the Llyn Brianne project in mid Wales, showed that kick samples faithfully represented densities of animals in cylinder samples over the range typically encountered in small upland streams; moreover, kick samples of 2 minutes duration returned, on average, 81% of the taxa recovered in samples of 6 minutes duration (Weatherley et al., 1985).
In the laboratory, animals were sorted and, where practicable, identified to species. Taxa were then assigned
EFFECTS OF RIPARIAN MANAGEMENT ON STREAM INVERTEBRATES 31 1
to one of five feeding guilds as defined by Cummins (1973) (shredders, grazers, collecter/gatherers = collectors, collector/filter feeders = filterers, or predators) using information from Hynes (1 970), Edington and Hildrew (1981), Elliott et at. (1988), and Wallace eta/. (1990). The abundance and number of species in each guild in the margin and riffle at each site was then calculated. Owing to taxonomic difficulties and ambiguity over feeding behaviour, meiofaunal groups such as micro-crustaceans, Simuliidae, Chironomidae and all beetles other than the Dytiscidae were omitted from analyses involving feeding guilds. The importance of these groups to conservation and biodiversity in streams is still frequently neglected, but worthy of further research effort. The omission of these taxa from analyses, however, does not make our comparisons between sites any less valid.
Physicochemical sampling
A quantitative assessment of the substratum and marginal structure of each stream (Tables 1 and 2) was made at the same time as biological sampling, following a modified version of the procedure used by Rutt et a/. (1990). A 0.5 x 0.5 m quadrat, divided into 25 sub-quadrats of 0.1 x 0.1 m, was placed randomly on
Table I . Physicochemical variables used to describe four major attributes (margin = marginal habitat structure, substratum = substratum composition, chemistry and location) at 66 stream sites in upland Wales and Scotland. Each group of variables was reduced to one
principal component (PC) describing each attribute (see text). Variables are in order of their scores along the PC.
Margin Substratum Chemistry Location
Tree roots Percentage cover Rock Stones Leaves GravelISand MossesILiverworts Wood Macroph ytes Earth Grass roots Sphagnum Grasses
Boulders (> 256 mm) PH Linkage Bedrock Aluminium Width Tree roots Colour Altitude Leaves Calcium Slope Silt/Mud (<0.06 mm) Conductivity Moss Iron PebblesICobbles (16-256 mm) Magnesium SandIgravel (0.06-16 mm)
Table 2. The mean scores (k SD) at sites in each TWINSPAN group at level 2 (derived separately for riffle and margin macroinvertebrate data) on each principal component from different sets of habitat variables measured at 66 sites in upland Wales and Scotland (see
Table 1). F values for ANOVAs between sites are also given (significance levels *p<0.05, **p<O.OI, ***p<O.Ool).
TWINSPAN group
3 4 5 6 F? 63 ~ ~~
Riffle fauna Margin PC Substratum PC Chemistry PC Location PC
Margin fauna Margin PC Substratum PC Chemistry PC Location PC
-0.5 (2.0) -0.2 (2.0) - 1.2 (1.2)
0.5 (1.6)
1.0 (2.1) 0.2 (1.5)
- 1.2 (0.6) 0.7 (2.0)
0.7 (1.6) 0.2 (1.2)
- 0.9 (0.8) 0.5 (1.8)
0.4 (1.7) 0.4 (1.3)
-0.5 (1.2) -0.5 (1.0)
0.0 (2.0) 0.0 (1.4)
-0.1 (1.2) -0.4 (1.2)
- 1.8 (0.8) -0.8 (2.1) -0.4 (1.3)
1.3 (1.9)
-0.2 (1.6) -0.1 (1.2)
1.2 (1.2) -0.1 (1.0)
-0.3 (1.7) -0.3 (1.1)
1.1 (1.2) 0.0 (1.1)
1 .o 0.2
1.6 14.5***
4.6* 2.5
11.2*** 4.8**

312 S. D. RUNDLE, E . C. LLOYD AND S. J . ORMEROD
the stream bed at 10 points located randomly over lOOm of each stream. The dominant substratum type (see Table 1) in each submerged sub-quadrat was then recorded. Adjacent to each of the 10 positions where substratum features were recorded, the dominant marginal habitat (see Table 1) just below the water surface was recorded in contiguous 0.1 m lengths on a random choice of either the right or left bank. Stream width and the proportion of stream shaded by overhanging trees (percentage cover) were also estimated on site, and altitude, slope and stream link magnitude were derived from Ordnance Survey 1:50000 scale maps (Ormerod and Edwards, 1987). Stream chemistry was analysed in the laboratory from spot water samples (see Table 1 for a list of chemical determinants). Metal determinations were made after filtration on site (0.45 pm), and therefore reflect concentrations usually reported as ‘dissolved’ (see Ormerod et a]. (in press) for more detailed methodology). Although only one chemical sample was taken from each site, this proved sufficient to detect strong correlations between acid-base status and the stream fauna (Ormerod et al. , in press).
Statistical analyses
Physicochemistry Four groups of physicochemical variables (Table 1) describing characteristics of the substratum, marginal
habitat, chemistry and location of the streams were summarized by principal components analysis (PCA- Ormerod et al. , in press). We used the principal component (PC) for each group shown to be best related to biological parameters as variables in our analyses. These PCs were as follows:
i. Substratum-describing a trend from sites with a high proportion of sand and gravel in their substratum, to those with more bedrock and boulders.
ii. Marginal-describing a gradient from sites where the marginal habitat was dominated by ‘soft’ features, such as grasses and Sphagnum, to those with a high proportion of ‘hard’ features, such as tree roots and stones. Soft and hard marginal features were shown previously to be associated with moorland riparian zones and banksides covered with broadleaf or coniferous trees respectively (Ormerod et al., in press).
iii. Chemistry-describing a trend from streams with high pH and low aluminium concentrations, to acidic sites with high aluminium concentrations.
iv. Location-describing a gradient from small to large streams.
Invertebrates All biological and physicochemical data were log or log(N+ 1) transformed to normalize distributions.
Macroinvertebrate data were classified using the FORTRAN program TWINSPAN (Hill, 1979). This technique arranges site groups into a hierarchy based on their taxonomic composition, and species are classified simultaneously on the basis of their occurrence in site groups. Indicator species are also identified, these showing the strongest preference for any given group according to their frequency of occurrence. Abundances were taken into account through the TWINSPAN pseudospecies facility, with cut levels of 1, 10, 100 and 1000 employed. In all cases analysis was halted at level 2, beyond which groupings ceased to be easily interpreted in ecological terms. Margin and riffle data were analysed separately.
Taxon richnesses for macroinvertebrate groups (Plecoptera, Ephemeroptera and Trichoptera) were compared between TWINSPAN groups using ANOVA. Differences in physicochemical variables between TWINSPAN groups were assessed using ANOVA. The occurrence of sites from different management groups in TWINSPAN groups was assessed by x2 of association.
The species richness of each functional feeding group for separate margin and riffle data, and the species richness and abundances for feeding group data from both habitats combined, were related to physicochemical variables (including the four PCs) using product-moment correlation. Feeding guild taxon richness and abundances were compared between riparian management groups using ANOVA.

EFFECTS OF RIPARIAN MANAGEMENT ON STREAM INVERTEBRATES
RESULTS
313
Community classification
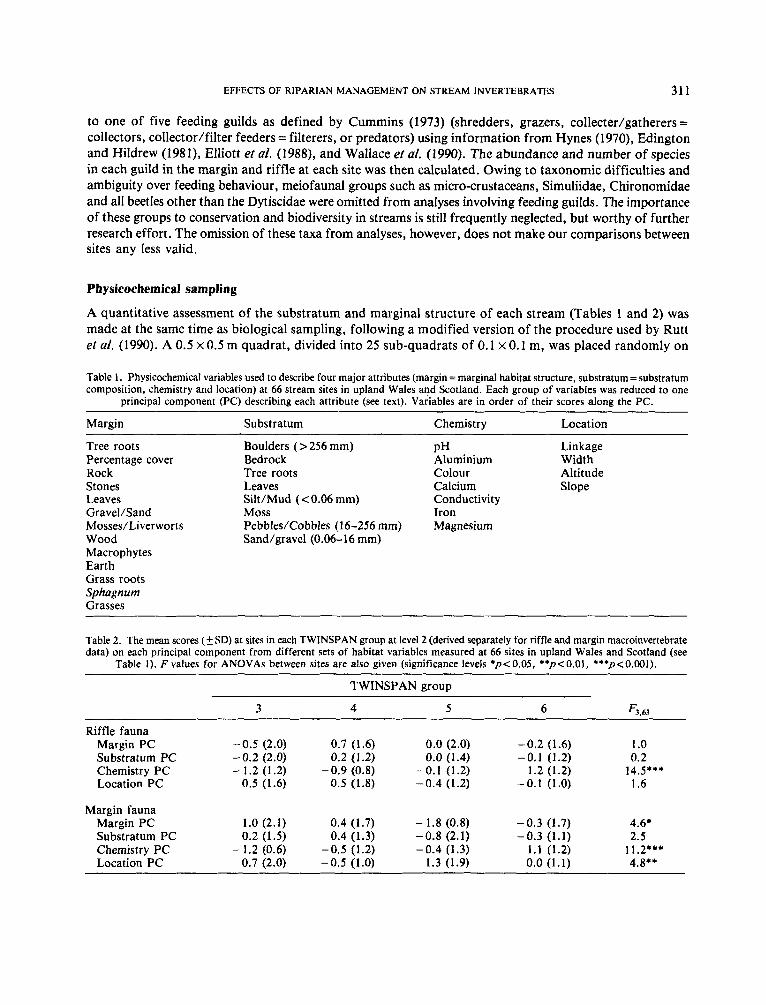
RVJe data TWINSPAN at level 1 divided riffles strongly on the basis of chemistry (Figure 1). Groups 1 and 2 differed
in their pH (6.4 f 0.4 SD and 5.6f 0.7 SD respectively, t-test pc0.001) and aluminium concentrations (0.063mgI-'x or +0.32SDand0.136mgl-lx or sO.34, t-testpc0.001). There was alsoasignificant,
RIFFLE
rl Sites
I ' 2 l Eelodidae
Bydraenidael
Eliinthidae Bydrophilidae P r o t o n e l u r a w
Tipulidae xsuExhorauatlca
HARGIN
0 Sites
E&ifiw Nydraenidae] W e l u r e l l P w Eydrophilidae
Figure 1. TWINSPAN classifications of 66 sites in upland Wales and Scotland based on macroinvertebrate samples from riffles and margins. Indicator species for each division are shown.
314 S. D . RUNDLE, E. C. LLOYD AND S. J . ORMEROD
Poljrcentropus flavomaculatus Paraleptophlebia submarqinata sialis fuliginosa Cordulegaster boltoni Chloroperla torrentium
Chloroperla tortentium Turbellaria silo pallipes 0 . . Wormaldia sp. . . . . Diplectrona felix . . Polycentropus kinqi Polycentropus flavomaculatus . ECdyonUruS dispar Taeniopteryx nebulosa
Rhrthroqena semicolorata Perla bipunctata
Asellus aquaticus Heptaqenia sulphurea Ecdyonurus venosus . . . Dinocras cephalotes . . Gammarus pulex . . Heptaqenia lateralis 0 . Perlodes microcephala 0 . . Hydropsyche siltali . . . . Chloroperla tripunctata . . . . Planorbis sp. Protonemura meyeri L@pidostoma hirtum Elminthidae
Baetis rhodani ; . . .
Isoperla qrammatica 0 0 . . Protonemura praecox . . * * Protonemura montana . . . Plectrocnemia geniculata 0 . Philopotamus montanus . . . Helodidae . . . . Odontocerum albicorne . . . . . ; . . . . . . .
0 . 0 . . * . . Tipul idae 0 . 0 . Dliqochaeta . . . . srmuliidae . . . . Limnephilidae . . . . Leuctra hippopus 0 .
Plectrocnemia conspersa . . . . Amphinemura sulcicollis
Leuctra nlqra . 0 . Diur.3 bicaudata Nemoura cambrica . . . . Dytiscidae . . . . ve1.a capra1 . . Nemoura cinerea 0 .
Lype phaeopa . Nemurella plctetl . . . Brachyptera risi . .
3 4 5 6
TWINSPAN group
. 0 . . . . . . . . .
Pisidium sp. Sericostoma personatun Polycentropus kinqi Ecdvonurus disvar
Hydropsyche siltalai Rhyacophila dorsalis Protonemura meyeri Tipulidae Dytisicidae Namoura cambrica Plectrocnemia conspersa Leuctra niqra Nemoura cinerea
Odontocerum albicorne Leuctra hipppus
Simuliidae Dicranota Limnephilidae Chironomidae Valia caprai
Elminthidae
Plectrocnemia conspersa Agapetus sp. Leptophlebia vespertina
oligochaeta
Iil
. . . . .
. . . . . .
. . . . . . . - . . . . . . . . - ~ . . -
Turbellaria Leuctra inermie Amphinemura sulcicollis Hydraenidae/Hydrophilidae Diura bicaudata
/ i I I I I
Gammarus pulex Chloroperla tripunctata Rhithrogena semicolorata Wormaldia SD. ~~ ~~ ~
Diplectrona' felix Heptaqenia lateralis Perlodes microcephsla Baetis rhodani Isoperla gramatica Helodidae
Asellus aquaticus Philoptamus montanus Protonemura praecox Protonemura montana Sericostoma SP. ~ y p e phaeopa Nemurella picteti
1 (T 3
. . . - . . . .
4
. . . . . . . - . . . . . . . . __ -
TWINSPAN
. . . . . . . - . . . . . . . -
-
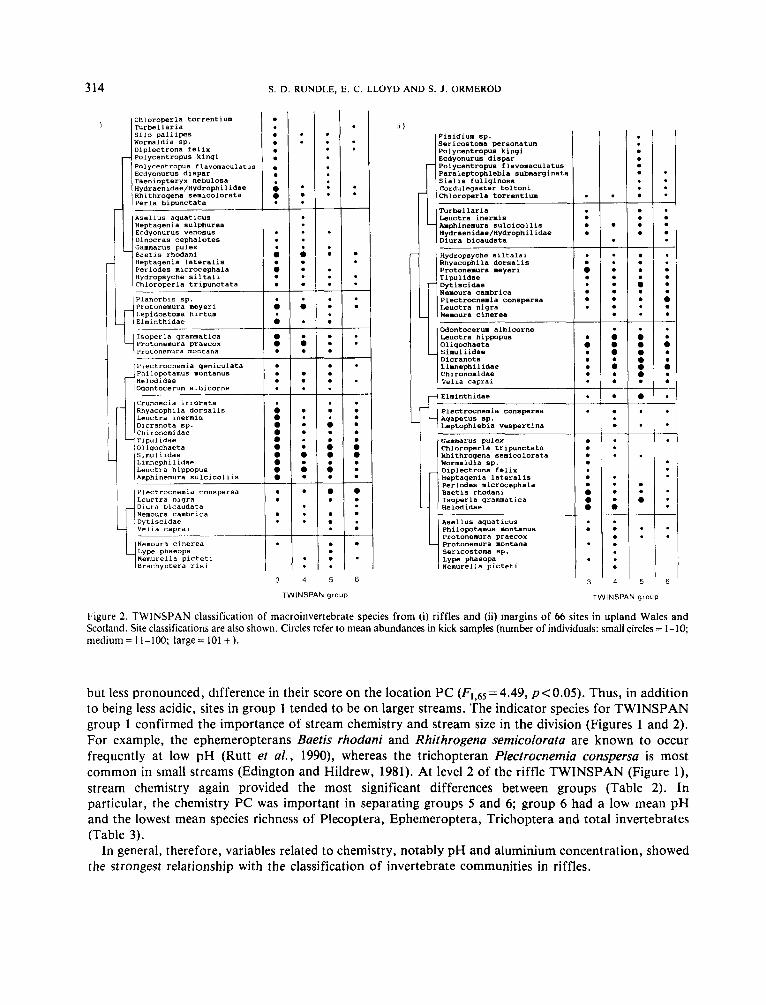
Figure 2. TWINSPAN classification of macroinvertebrate species from (i) riffles and (ii) margins of 66 sites in upland Wales and Scotland. Site classifications are also shown. Circles refer to mean abundances in kick samples (number of individuals: small circles = 1-10; medium: 1 1 - 1 0 0 ; large= 101 +).
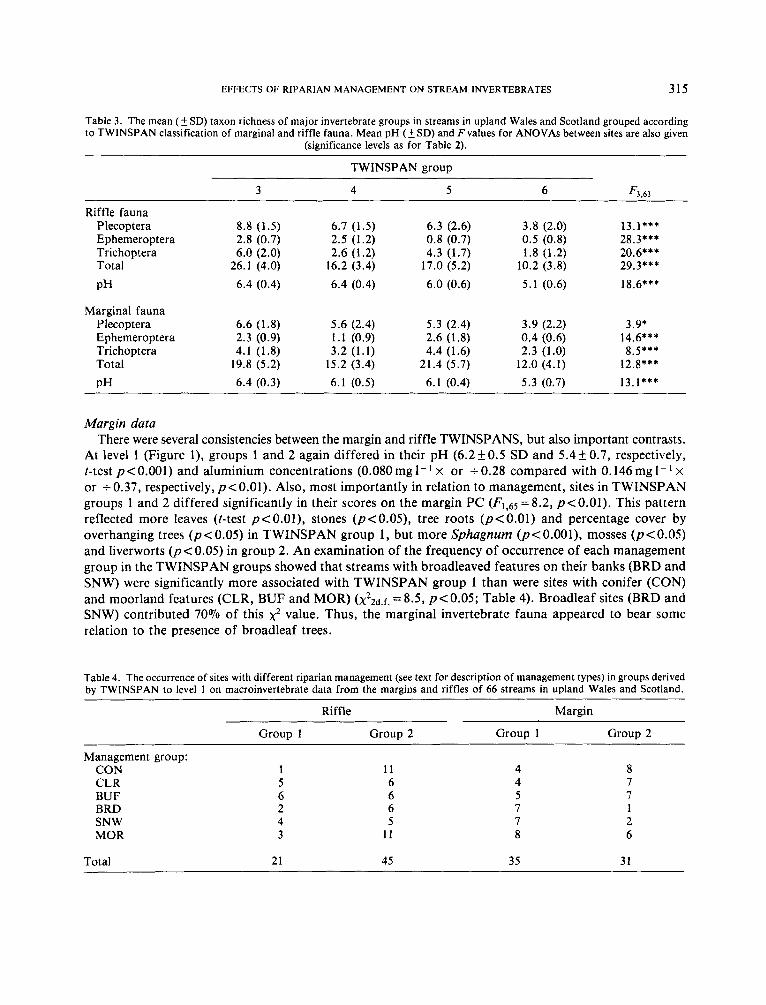
but less pronounced, difference in their score on the location PC (Fi,65=4.49, pe0 .05) . Thus, in addition to being less acidic, sites in group 1 tended to be on larger streams. The indicator species for TWINSPAN group 1 confirmed the importance of stream chemistry and stream size in the division (Figures 1 and 2). For example, the ephemeropterans Baetis rhodani and Rhithrogena semicolorata are known to occur frequently at low pH (Rutt et al., 1990), whereas the trichopteran Plectrocnemia conspersa is most common in small streams (Edington and Hildrew, 1981). At level 2 of the riffle TWINSPAN (Figure l), stream chemistry again provided the most significant differences between groups (Table 2). In particular, the chemistry PC was important in separating groups 5 and 6; group 6 had a low mean pH and the lowest mean species richness of Plecoptera, Ephemeroptera, Trichoptera and total invertebrates (Table 3).
In general, therefore, variables related to chemistry, notably pH and aluminium concentration, showed the strongest relationship with the classification of invertebrate communities in riffles.
EFFECTS OF RIPARIAN MANAGEMENT ON STREAM INVERTEBRATES 315
Table 3. The mean ( 5 SD) taxon richness of major invertebrate groups in streams in upland Wales and Scotland grouped according to TWINSPAN classification of marginal and riffle fauna. Mean pH (5 SD) and F values for ANOVAs between sites are also given
(significance levels as for Table 2).
TWINSPAN group
3 4 5 6 F161 ~ ~~
Riffle fauna Plecoptera 8.8 (1.5) 6.7 (1.5) 6.3 (2.6) 3.8 (2.0) 13.1*** Ephemeroptera 2.8 (0.7) 2.5 (1.2) 0.8 (0.7) 0.5 (0.8) 28.3*** Trichoptera 6.0 (2.0) 2.6 (1.2) 4.3 (1.7) 1.8 (1.2) 20.6* * * Total 26.1 (4.0) 16.2 (3.4) 17.0 (5.2) 10.2 (3.8) 29.3* * * PH 6.4 (0.4) 6.4 (0.4) 6.0 (0.6) 5.1 (0.6) 18.6***
Marginal fauna Plecoptera 6.6 (1.8) 5.6 (2.4) 5.3 (2.4) 3.9 (2.2) 3.9* Ephemeroptera 2.3 (0.9) 1.1 (0.9) 2.6 (1.8) 0.4 (0.6) 14.6*** Trichoptera 4.1 (1.8) 3.2 (1.1) 4.4 (1.6) 2.3 (1.0) 8.5*** Total 19.8 (5.2) 15.2 (3.4) 21.4 (5.7) 12.0 (4.1) 12.8*** PH 6.4 (0.3) 6.1 (0.5) 6.1 (0.4) 5.3 (0.7) 13.1***
Margin data There were several consistencies between the margin and riffle TWINSPANS, but also important contrasts.
At level 1 (Figure l), groups 1 and 2 again differed in their pH (6.2k0.5 SD and 5.420.7, respectively, t-test p<O.OOl) and aluminium concentrations (0.080 mg 1- x or +- 0.28 compared with 0.146 mg 1- x or + 0.37, respectively, p <0.01). Also, most importantly in relation to management, sites in TWINSPAN groups 1 and 2 differed significantly in their scores on the margin PC (F1,65 = 8.2, ~ ~ 0 . 0 1 ) . This pattern reflected more leaves (t-test p< 0.01), stones (p<0.05), tree roots (p<O.Ol) and percentage cover by overhanging trees (p < 0.05) in TWINSPAN group 1, but more Sphagnum (p < 0.001), mosses ( p < 0.05) and liverworts @<0.05) in group 2. An examination of the frequency of occurrence of each management group in the TWINSPAN groups showed that streams with broadleaved features on their banks (BRD and SNW) were significantly more associated with TWINSPAN group 1 than were sites with conifer (CON) and moorland features (CLR, BUF and MOR) (x22d.f. = 8 .5 , p<0.05; Table 4). Broadleaf sites (BRD and SNW) contributed 70% of this x2 value. Thus, the marginal invertebrate fauna appeared to bear some relation to the presence of broadleaf trees.
Table 4. The occurrence of sites with different riparian management (see text for description of management types) in groups derived by TWINSPAN to level 1 on macroinvertebrate data from the margins and riffles of 66 streams in upland Wales and Scotland.
Riffle Margin
Group I Group 2 Group 1 Group 2 ~ ~~
Management group: CON CLR BUF BRD SNW MOR
Total 21
11 6 6 6 5
11
45 35 31

316 S. D. RUNDLE, E. C. LLOYD A N D S. J . ORMEROD
Species that were important indicators in the marginal TWINSPAN at level 1 confirmed the importance of stream chemistry and habitat structure in influencing the division. For example, the mayfly Baetis rhodani and the trichopteran Philopotamus montanus both occur predominantly in less acid streams (Rutt et al. , 1990). Also, the two leaf-shredding plecopterans Protonemura praecox and Nemurella picteti might also be expected to favour wooded streams (e.g. Groom and Hildrew, 1989).
At TWINSPAN level 2 for margin data, stream chemistry remained important, separating acidic sites in group 6 from those in group 5 (Table 2). Scores on the location PC also differed significantly between groups, due to larger streams forming groups 3 and 5 (Table 2). Of most importance in relation to riparian management, scores on the margin PC were significantly reduced in group 5 relative to group 6 (Table 2 ) . In keeping with this division, sites in group 5 had more Sphagnum and grassy vegetation than group 6 , but less cover by trees and fewer tree roots. Indicators for group 5 included taxa such as the Limnephilidae, Centroptilum luteolum, Leptophlebia marginata, Paraleptophlebia submarginata, and Cordulegaster bolfoni (Figure 2 ) , species all associated with marginal habitats (e.g. Rutt et al., 1989).
Thus, while chemistry was again a major correlate with the TWINSPAN of margin data, some influence by riparian management could not be excluded. In particular, the presence of broadleaf trees, either naturally occurring or due to management in plantation forests, was involved in separating sites at level 1 . At level 2 , sites were again divided partly on the basis of the presence of ‘soft’ features, associated with moorland riparian zones (Ormerod et al., in press), in the stream margins.
Response of feeding groups to physicochemistry
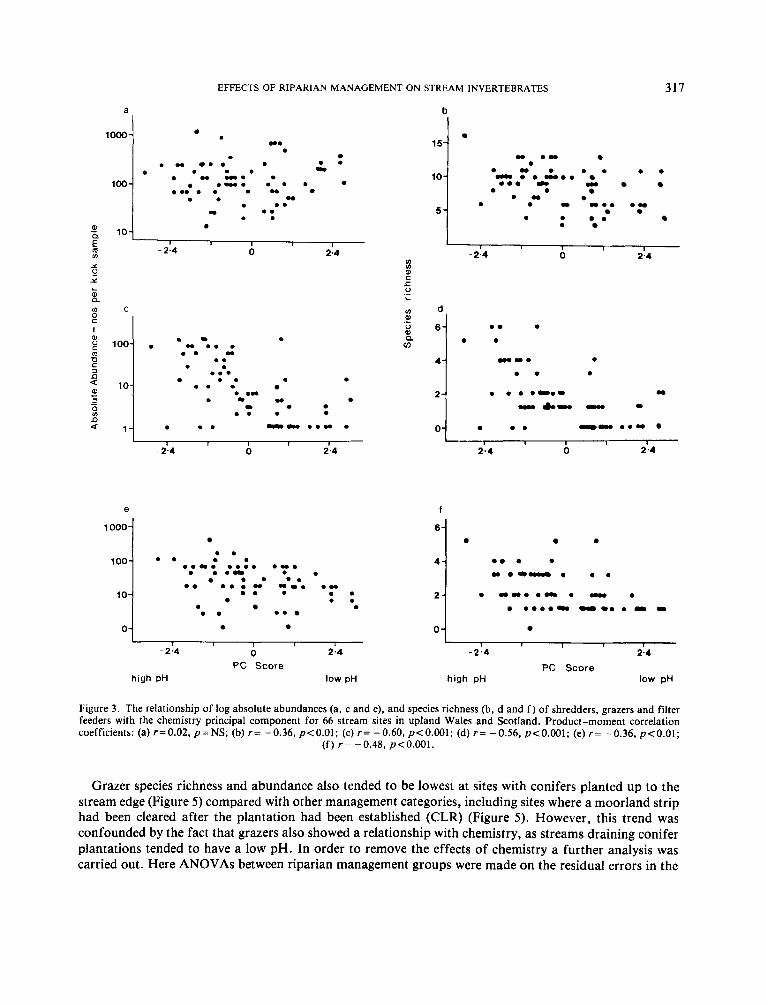
The total species richness and total abundances of grazers and filterers decreased significantly along the chemistry PC (Figure 3 ) . Thus, there were fewer species and individuals of both of these feeding groups at acidic sites with high aluminium concentrations. This was reflected in the low numbers of filterers and grazers in the TWINSPAN groups with low mean pH (i.e. groups 5 and 6 for riffles and group 6 for margins; Table 5 ) . The total number of shredder species in riffles also decreased along the chemistry PC (Figure 3 ) with fewer shredder taxa at low pH.
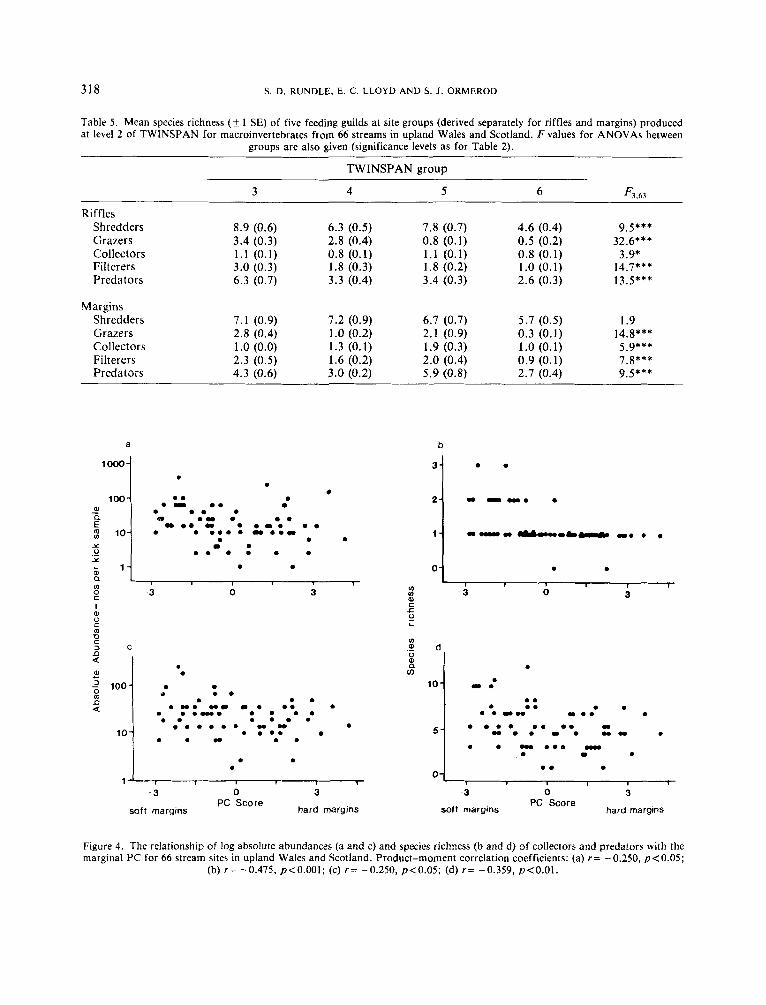
In contrast to grazers and filterers, marginal habitat structure was of greatest importance in correlations with predators and collectors. The total species richness and total abundance of both guilds decreased along the marginal principal component (Figure 4) . This was reflected in the separate margin samples where species richness correlated strongly with variables important on the marginal P C such as percentage canopy cover (collectors r= -0.47, p<O.OOl; predators r=0.33, p<O.Ol) tree roots (collectors r= -0.30, ~ ~ 0 . 0 5 ; predators r= -0.36,p<0.01) and grasses (collectors r=0.43,p<0.001; predators r=0.36,p<0.01) as well as with the PC itself (collectors r = -0.415, p<O.OOl; predators r = -0.37, p < O . O l ) . Thus, the number of species and individuals of collectors and predators in stream margins declined with an increase in hard bankside features associated with conifer forest and broadleaf woodland. This was reflected in the high species richness of both collectors and predators in TWINSPAN group 5 for margin data (i.e. the group with the highest proportion of soft features in their margins).
Feeding groups and riparian management
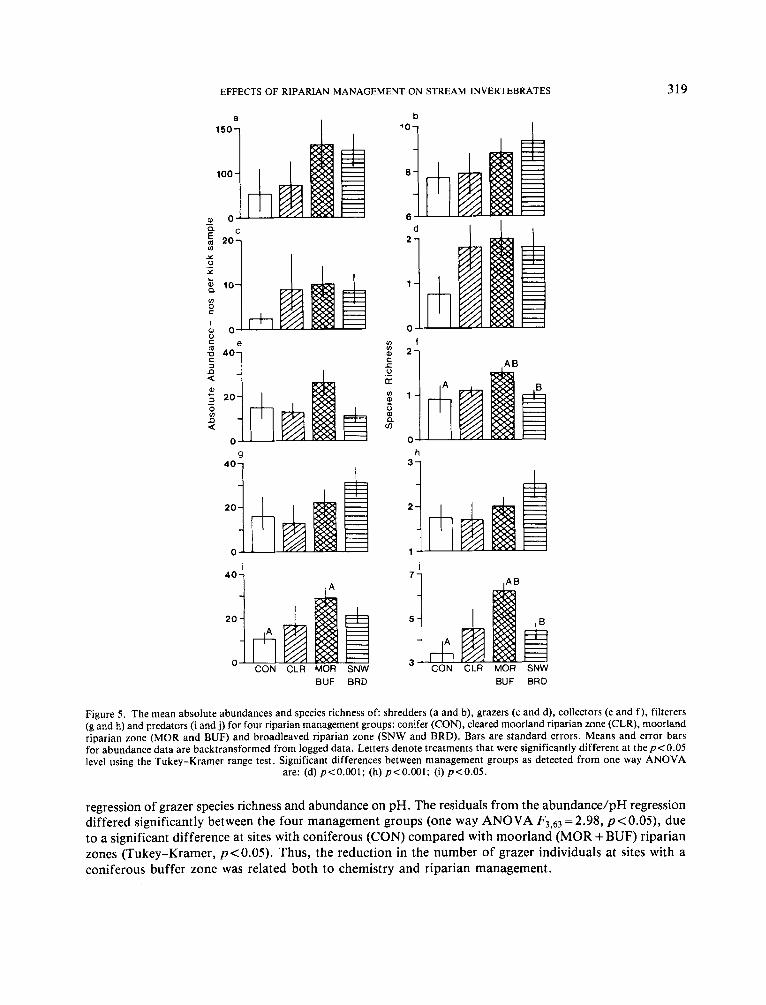
The high species richness of collectors and predators at sites with ‘soft’ marginal features was reflected in their distribution with riparian management. In this case the number of management categories was reduced to four, based on the dominant riparian component at the stream’s edge (Figure 5). A Tukey-Kramer range test showed that there were significant differences ( p < 0.05) between the number of species of collectors and predators at sites with moorland riparian zones (MOR + BUF) compared with those with riparian zones of either coniferous (CON) or broadleaved (SNW + BRD) trees (Figure 5). Moreover, predators were less abundant at sites with coniferous compared with moorland riparian zones (Tukey-Kramer, p < 0.05).
EFFECTS OF RIPARIAN MANAGEMENT ON STREAM INVERTEBRATES
10-
5 -
317
6-
4-
2-
b
I
1000-
100-
10-
a
. ... 151
0 . n . 0 .
m u
a*. 0 .
0
0 .
.., 10
l a I I I - 2.4 0 2.4
1 I I I
- 2.4 0 2.4 u)
a, A= 0 .- L
u) m 0 .-
a v)
Y
Y .- 0
L
m Q
C UI
2 I 9)
2
s 0 3
100
0 . . . . . 10 .. . m
3 0 ln
c - n a
*a 0 0 ..
0 .. . .. woo....
1 1 I 1 I
1
- 2.4 0 2.4
.. 2.4 0 2.4
0 I I I I
e f
0 . . 0 . ..
0 0 I I 1 I 1 I I
- 2.4 0 2.4 - 2 . 4 2.4
PC Score PC Score high PH low pH high pH low pH
Figure 3 . The relationship of log absolute abundances (a, c and e), and species richness (b, d and f ) of shredders, grazers and filter feeders with the chemistry principal component for 66 stream sites in upland Wales and Scotland. Product-moment correlation coefficients: (a) r=0.02, p=NS; (b) r = -0.36, p<O.OI; (c) r = -0.60, p<O.OOl; (d) r = -0.56, p<O.OOI; (e) r = -0.36, p<O.OI;
( f ) r= -0.48, p<O.OOI.
Grazer species richness and abundance also tended to be lowest at sites with conifers planted up to the stream edge (Figure 5 ) compared with other management categories, including sites where a moorland strip had been cleared after the plantation had been established (CLR) (Figure 5 ) . However, this trend was confounded by the fact that grazers also showed a relationship with chemistry, as streams draining conifer plantations tended to have a low pH. In order to remove the effects of chemistry a further analysis was carried out. Here ANOVAs between riparian management groups were made on the residual errors in the
318 S. D. RUNDLE, E. C. LLOYD AND S . J . ORMEROD
Table 5 . Mean species richness ( 5 I SE) of five feeding guilds at site groups (derived separately for riffles and margins) produced at level 2 of TWINSPAN for macroinvertebrates from 66 streams in upland Wales and Scotland. F values for ANOVAs between
groups are also given (significance levels as for Table 2).
TWINSPAN group
3 4 5 6 F3,63
Riffles Shredders 8.9 (0.6) 6.3 (0.5) 7.8 (0.7) 4.6 (0.4) 9.5***
Collectors 1 . 1 (0.1) 0.8 (0.1) 1.1 (0.1) 0.8 (0.1) 3.9* Filterers 3.0 (0.3) 1.8 (0.3) 1.8 (0.2) 1 .o (0.1) 14.7***
Grazers 3.4 (0.3) 2.8 (0.4) 0.8 (0.1) 0.5 (0.2) 32.6***
Predators 6.3 (0.7) 3.3 (0.4) 3.4 (0.3) 2.6 (0.3) 13.5***
Margins Shredders 7.1 (0.9) 7.2 (0.9) 6.7 (0.7) 5.7 (0.5) 1.9 Grazers 2.8 (0.4) 1.0 (0.2) 2.1 (0.9) 0.3 (0.1) 14.8***
Filterers 2.3 (0.5) 1.6 (0.2) 2.0 (0.4) 0.9 (0.1) 7.8*** Collectors 1.0 (0.0) 1.3 (0.1) 1.9 (0.3) 1 .o (0.1) 5.9***
Predators 4.3 (0.6) 3.0 (0.2) 5.9 (0.8) 2.7 (0.4) 9.5***
a
'"j . .
L
%! '1 . . u L
P I a, 0
m
3 c n d I
F
n 4
1°1
-3 0 3
. . . -3 0 3
PC Score soft margins hard margins
L
u)
._ i l 10 0 .
0 . . 0
1 I 1 I
-3 0 3 PC Score
soft margins hard margins
Figure 4. The relationship of log absolute abundances (a and c) and species richness (b and d) of collectors and predators with the marginal PC for 66 stream sites in upland Wales and Scotland. Product-moment correlation coefficients: (a) r = -0.250, ~<0.05;
(b) r = -0.475, p<O.OOl; (c) r = -0.250, p<0.05; (d) r = -0.359, p<O.Ol.
EFFECTS OF RIPARIAN MANAGEMENT ON STREAM INVERTEBRATES 319
a 1 5 0 1
I I
b
'01 I
100 8
m o 6
2
Y 0 Y .-
10 1
m C
I 0 0 0
A B
40i 20
0 i
40 1
!a SNW
BUF BRD
h 31
i 2hk 1
I
5
CON CLR MOR SNW BUF BRD
Figure 5 . The mean absolute abundances and species richness of: shredders (a and b), grazers (c and d), collectors ( e and f), filterers (g and h) and predators (i and j) for four riparian management groups: conifer (CON), cleared moorland riparian zone (CLR), moorland riparian zone (MOR and BUF) and broadleaved riparian zone (SNW and BRD). Bars are standard errors. Means and error bars for abundance data are backtransformed from logged data. Letters denote treatments that were significantly different at the p < 0.05 level using the Tukey-Kramer range test. Significant differences between management groups as detected from one way ANOVA
are: (d) p<O.OOl; (h) p<O.OOl; (i) p<O.OS.
regression of grazer species richness and abundance on pH. The residuals from the abundance/pH regression differed significantly between the four management groups (one way ANOVA F3,63 = 2.98, p<O.O5), due to a significant difference at sites with coniferous (CON) compared with moorland (MOR + BUF) riparian zones (Tukey-Kramer, p<O.OS). Thus, the reduction in the number of grazer individuals at sites with a coniferous buffer zone was related both to chemistry and riparian management.

320 S. D. RUNDLE, E. C. LLOYD AND S. J . ORMEROD
DISCUSSION
In summary, our results demonstrate the importance of chemistry, in particular pH, in influencing the taxonomic and trophic structure of macroinvertebrate communities in streams of upland Britain. TWINSPAN classifications of both riffle and marginal assemblages were closely related to stream acidity and aluminium concentrations; total taxon richness and the number of species of major macroinvertebrate groups were significantly lower in riffle communities at low pH. The species richness and abundances of grazers and filter feeders also decreased with increasing acidity, and there were fewer shredder species at low pH.
Despite the dominant influence of chemistry, however, habitat characteristics in stream margins had an important effect on the taxonomic and trophic structure of macroinvertebrate assemblages. Marginal habitat structure was, in turn, influenced by riparian management. For example, collectors and predators were most abundant and species-rich at sites with moorland/grassland riparian zones, and grazer abundances were lowest where conifers were planted to the stream’s edge.
Chemical influences
The importance of stream acidity to macroinvertebrate community structure has been shown repeatedly in Britain (Townsend et al., 1983; Wade et al., 1989; Rutt et al., 1990). Assemblages are consistently less diverse at acidic sites with high aluminium concentrations, with some feeding guilds absent at acid sites (cf. Sutcliffe and Hildrew, 1989). These findings were clearly confirmed by our survey.
The exact mechanisms by which invertebrates are affected by acidity are still unclear. Some species may have a physiological intolerance of the chemical conditions associated with low pH (Herrmann and Anderson, 1986; Ormerod et al., 1987). Another view, important in the context of this study, is that reduced pH may have an indirect influence by affecting food availability. This may have consequences for the trophic dynamics and food web processes in stream systems (Sutcliffe and Hildrew, 1989).
The reduction of macroinvertebrate data into functional feeding guilds can provide a useful insight into the effects on trophic structure. Previous studies suggest a lower diversity and abundance of grazers and filter feeders at acid sites (e.g. Otto and Svensson, 1983; Townsend et al., 1983), patterns consistent with our observations. Both of these feeding groups may be responding to a reduction in food quality or quantity at low pH. For instance, Winterbourn et al. (1985) found that the structure of the epilithon, on which grazers feed, was altered in acidic streams. Rosemond et al. (1992), however, have recently provided a contrasting view because food for grazers at acid sites remained abundant even though this group was scarce. Reduced decomposition of leaf litter at low pH (Chamier, 1987) may also reduce the availability of fine particulate detritus available to filter feeders.
Influence of habitat structure and riparian management
Ormerod et al. (in press) showed that the marginal habitats at sites in this study were affected by riparian management. Streams whose banks were covered by either coniferous or broadleaf trees had a higher proportion of hard features such as rocks, stones and tree roots in their margins. Conversely, streams with moorland/grassland riparian zones had a higher proportion of soft marginal features, such as grasses and Sphagnum. Our study suggests that this relationship, in turn, influenced the macroinvertebrate community structure in the stream margins. Vegetated stream margins associated with moorland tend to be more structurally complex than the harder banksides in forested streams (Rutt et al., 1989), perhaps providing habitat to certain species (e.g. Jenkins et a/. , 1984; Ormerod, 1987).
There was evidence from our study that trophic conditions were influenced by riparian features. For example, a high number of predators and collectors occurred at sites with moorland/grassland riparian zones. One possibility is that vegetation traps or retains fine particulate matter used by collectors, while

EFFECTS OF RIPARIAN MANAGEMENT ON STREAM INVERTEBRATES 321
predators may be showing a direct response to increased prey availability (as enhanced macroinvertebrate abundances) in softer margins.
Riparian management is likely to influence the trophic dynamics in streams directly, energy inputs varying with different forms of bankside vegetation. Thus, shading is liable to limit autochthonous production, while the nature of the canopy will affect the quality of allochthonous material. On this basis, Vannote et al. (1980) predicted that grazer densities and species richness would be higher at sites where riparian canopy cover was minimal and the productivity of periphytic algae high (Power, 1984; Feminella et at., 1989). In keeping with this prediction, we found that grazers were less abundant in streams where conifers were planted up to the stream edge, a result evident even after accounting for possible effects on grazer densities by low pH. Of particular relevance to management, grazer densities were not reduced at afforested sites with moorland riparian strips, even where a buffer zone had been cleared after the conifer plantation had been established.
In contrast to grazers, shredders are likely to dominate invertebrate communities in woodland streams, which receive large inputs of leaf litter (Vannote et al., 1980). However, shredders in our study showed no relationship with riparian management and were not associated with the presence of broadleaved trees. Moreover, neither their distribution nor abundance were correlated with variables associated with broadleaf litter inputs. This suggests other sources of CPOM, such as grasses, may be available in upland streams, or that some shredder species may utilize food items associated with other feeding guilds (Hawkins et al., 1982). However, one other explanation is that broadleaf streams in our study did not retain litter inputs in suitable concentrations, either because of their harder margins or steeper slope (Ormerod et al., in press). It has recently been shown that shredders responded directly to an experimental input of coarse particulate organic matter (CPOM), but only where the retentiveness of CPOM was artificially increased (Dobson and Hildrew, 1991). Finally, many chironomids are detritivores and we cannot dismiss the possibility that their inclusion in trophic analyses would have given a different view of shredder distribution and abundance.
Conservation and management-ramifications
Boon (1992) has suggested recently that five management options form the basis for the conservation of streams and rivers. They are respectively: preservation (of pristine systems), limitation or mitigation (of damaging activities), restoration (following damage) and dereliction (of totally degraded systems). Riparian management, as in our study, can be viewed as an option for both limiting and mitigating the effects of forestry, implemented either at the planting stage (limitation) or subsequently through tree clearance (mitigation). Here, we have posed questions over whether these techniques are effective in protecting the structure and function of the stream system. We have demonstrated that, with some limitations, both are options for stream conservation. For example, the character of riparian zones had a direct effect on habitat diversity through an influence on the complexity of stream margins. This habitat effect, in turn, influenced the structure and diversity of macroinvertebrate communities (Ormerod et al., in press). Moreover, the character of bankside vegetation also had an apparent effect on trophic function, probably by controlling energy inputs to the stream as light and tree products. Together, these results support the management proposals set out by the forestry industry in the Forests and Water Guidelines (Forestry Commission, 1991), hitherto mostly developed in the absence of research data. While our study did not address the effects of buffer strips in reducing effects by forestry on discharge regimes and temperature (Maitland et al., 1990), the evidence from the data on habitat structure and macroinvertebrate communities suggested these problems were not affecting the biology of our sites.
Nevertheless, it was clear from this, and our previous study (Ormerod et al. , in press) that riparian management may have been less effective in overcoming the problem of acidification: forest sites had elevated aluminium concentrations irrespective of riparian management. This contrasts with a view proposed in the Forests and Water Guidelines (Forestry Commission, 1991). This feature remains important in view of the

322 S. D. RUNDLE, E. C. LLOYD AND S. J . ORMEROD
marked and well recognized effect of acidity on stream communities and their trophic functioning. While it may be possible to try to mitigate the impact of such problems through restorative practices such as liming, these techniques may themselves have major ramifications for conservation. Moreover, their benefits to the stream community remain equivocal (Weatherley and Ormerod, 1992), while implementation at the catchment scale is likely to result in considerable financial cost. The whole problem of acidification highlights the problem of scale when considering measures implemented for the conservation of rivers: acidification through atmospheric deposition (accentuated by conifer forestry) represents an environmental impact that has an effect at the supra-catchment level, that is at a regional rather than a catchment scale (Boon, 1992).
The fact that catchment-scale and supra-catchment scale effects on rivers through afforestation and acidification can partly or wholly subsume those at the bankside level has major ramifications for the current concepts of river conservation. Boon (1991) has suggested that, when British rivers are protected through designation as SSSIs (Sites of Special Scientific Interest), control of damaging activities is often only possible at the riparian level. The lessons from our study would be that such control might allow moderate benefits to the stream habitat structure, and to the community and feeding guilds represented. However, there is a risk that chemical signals from the catchment scale could have stronger effects on both stream communities and trophic function. In such instances, one useful way forward might be the recognition or designation of whole river catchments as river ecosystem protection zones. Current changes to agricultural practice and conservation management at the landscape scale in Britain mean that such designations are a real possibility. As Petersen et al. (1987) have suggested, riparian management should represent only the first step in the protection of whole catchments.
ACKNOWLEDGEMENTS
We are grateful to Gordon Patterson of the Forestry Commission who helped in the selection of sites, and to the various Forest District Officers who provided further assistance. The National Rivers Authority (Wales) performed chemical analyses, and Ann Douglas sorted and identified invertebrates. Rob Soutar was instrumental in the initiation of this project, which was funded by the Nature Conservancy Council.
REFERENCES
Boon, P. J . 1991. ‘The role of Sites of Special Scientific Interest (SSSIs) in the conservation of British rivers’, Freshwater
Boon, P. J . 1992. ‘Essential elements in the case for river conservation’, in Boon P. J., Calow P. and Petts G. E.
Bunn, S. E. 1986. ‘Spatial and temporal variation in the macroinvertebrate fauna of streams of the northern Jarrah
Chamier, A. C. 1987. ‘The effect of pH on the microbial degradation of leaf litter in seven streams in the English
Cummins, K. W. 1973. ‘Trophic relations of aquatic insects’, Annual Review of Entomology, 18, 183-206. Dobson, M. and Hildrew, A. G. 1991. ‘A test of food limitation in stream detritivores’, Journal of Animal Ecology,
Edington, J. M. and Hildrew, A. G. (1981). A Key to the Caseless Caddis Larvae of the British Isles with Notes on
Elliott, J. M., Humpesch, U. H. and Macan, T. T. 1988. Larvae of the British Ephemeroptera: A Key with Ecological
Feminella, J. W., Power, M. E. and Resh, V. H. 1989. ‘Periphyton responses to invertebrate grazing and riparian
Forestry Commission 1991. Forests and Water Guidelines, 2nd edn. HMSO Publications, London. Groom, A. P. and Hildrew, A. G. 1989. ‘Food quality for detritivores in streams of contrasting pH’, Journal of Animal
Hawkins, C. P., Murphy, M. L. and Anderson, N. H. 1982. ‘Effects of canopy, substrate composition and gradient on the structure of macroinvertebrate communities in Cascade Range streams of Oregon’, Ecology, 63, 1840-1856.
Forum, 1, 95-108.
(Eds), River Conservation and Management, John Wiley, Chichester, 11-33.
forest, Western Australia: functional organization’, Freshwater Biology, 16, 621-632.
Lake District’, Oecologia, 71, 491-500.
61, 69-78.
their Ecology, Scientific Publication No. 43, Freshwater Biological Association, Windermere.
Notes, Scientific Publication No. 49, Freshwater Biological Association, Windermere.
canopy in three northern California coastal streams’, Freshwater Biology, 22, 445-457.
Ecology, 58, 863-881.

EFFECTS OF RIPARIAN MANAGEMENT ON STREAM INVERTEBRATES 323
Herrmann, J . and Anderson, K. J. 1986. ‘Aluminium impact on respiration of lotic mayflies at low pH’, Water, Air and Soil Pollution, 30, 703-709.
Hildrew, A. G., Dobson, M. K., Groom, A. P.. Ibbotson, A., Lancaster, J. and Rundle, S. D. in press. ‘Flow and retention in the ecology of stream invertebrates’, Verhandlungen der Internationalen Vereinigung fur theoritische und angewandte Limnologie.
Hill, M. 0. 1979. TWINSPAN-A FORTRAN Program for Arranging Multivariate Data in an Ordered Two-way Table by Classifcation of the Individuals and Attributes, Cornell University, Ithaca, New York.
Hynes, H. B. N. 1970. The Ecology of Running Waters, University of Liverpool Press, Liverpool. Jenkins, R. A., Wade, K. R. and Pugh, E. 1984. ‘Macroinvertebrate-habitat relationships in the River Teifi catchment
and the significance to conservation’, Freshwater Biology, 14, 23-42. Maitland, P. S., Newson, M. D. and Best, G. A. 1990. The Impact of Afforestation andForestry Practice on Freshwater
Habitats, Focus on Nature Conservation No. 23, Nature Conservancy Council, Peterborough. Murphy, M. L., Hawkins, C. P. and Anderson, N. H. 1981. ‘Effects of canopy modification and accumulated sediment
on stream communities’, Transactions of the American Fisheries Society, 110, 469-478. Ormerod, S. J. 1987. ‘The influences of habitat and seasonal sampling regimes on the ordination and classification
of macroinvertebrate assemblages in the catchment of the River Wye, Wales’, Hydrobiologia, 150, 143-151. Ormerod, S. J . , Boole, P., McCahon, C. P., Weatherley, N. S., Pascoe, D. and Edwards, R. W. 1987. ‘Short-term
experimental acidification of a Welsh stream: comparing the biological effects of hydrogen ions and aluminium’, Freshwater Biology, 17, 341-356.
Ormerod, S. J . , Donald, A. P. and Brown, S. J. 1989. ‘The influence of plantation forestry on the pH and aluminium concentration of upland Welsh streams: a re-examination’, Environmental Pollution, 62, 47-62.
Ormerod, S. J. and Edwards, R. W. 1987. ‘The ordination and classification of macroinvertebrate assemblages in the catchment of the River Wye in relation to environmental factors’, Freshwater Biology, 17, 533-546.
Ormerod, S. J . , Rundle, S. D., Lloyd, E. C. and Douglas, A. A. in press. ‘The influence of riparian management on the habitat structure and macroinvertebrate communities of upland streams draining plantation forests’, Journal of Applied Biology.
Otto, C. and Svensson, B. S. 1983. ‘Properties of acid, brown water streams in South Sweden’, Archiv fur Hydrobiologie,
Petersen, R. C., Madsen, B. L., Wilzbach, M. A., Magadza, C. H. D., Paarlberg, A., Kullberg, A. and Cummins, K. W. 1987. ‘Stream management: emerging global similarities’, Ambio, 16, 166-179.
Power, M. E. 1984. ‘Habitat quality and the distribution of algae-grazing catfish in a Panamanian stream’, Journal of Animal Ecology, 53, 357-374.
Robinson, M. 1980. ‘The Effects of Pre-afforestation Drainage on the Streamflow and Water Quality of a Small Upland Catchment’, Report No. 73. Institute of Hydrology, Wallingford.
Rosemond, A. D., Reice, S. R., Elwood, J . W. and Mullholland, P. J. 1992. ‘The effects of stream acidity on benthic macroinvertebrate communities in the south eastern United States’, Freshwater Biology, 27, 193-210.
Rutt, G. P., Weatherley, N. S. and Ormerod, S. J . 1989. ‘Microhabitat availability in Welsh moorland and forest streams as a determinant of macroinvertebrate distribution’, Freshwater Biology, 22, 247-261.
Rutt, G. P., Weatherley, N. S. and Ormerod, S. J . 1990. ‘Relationships between the physicochemistry and macro- invertebrates of British upland streams: the development of a modelling and indicator system for predicting fauna and detecting acidity’, Freshwater Biology, 24, 463-480.
Sedell, J . R., Triska, F. J . and Triska, N. S. 1975. ‘The processing of conifer and hardwood leaves in two coniferous streams: (i) weight loss and associated invertebrates’, Verhandlungen der Internationalen Vereinigung fur theoretische und angewandte Limnologie, 19, 1617-1627.
Statzner, B., Gore, J. A. and Resh, V. H. 1988. ‘Hydraulic stream ecology: observed patterns and potential applications’, Journal of the North American Benthological Society, 7, 307-360.
Sutcliffe, D. W. and Hildrew, A. G. 1989. ‘Invertebrate communities in acid streams’, in Morris, R., Taylor, E. W., Brown, D. J. A. and Brown, J. A. (Eds), Acid Toxicity and Aquatic Animals, Cambridge University Press, Cambridge,
Townsend, C. R., Hildrew, A. G. and Francis, J . 1983. ‘Community structure in some southern English streams: the
Vannote, R. L., Minshall, G. W., Cummins, K. W., Sedell, J. R. and Cushing, C. E. 1980. ‘The river continuum
Wade, K. R., Ormerod, S. J. and Gee, A. S. 1989. ‘Classification and ordination of macroinvertebrate assemblages
Wallace, I. D., Wallace, B. and Phillipson, G. N. 1990. A Key to the Case-bearing Caddis Larvae of Britain and Ireland,
99, 15-36.
13-29.
influence of physicochemical factors’, Freshwater Biology, 13, 521 -544.
concept’, Canadian Journal of Fisheries and Aquatic Sciences, 37, 130-137.
to predict stream acidity in upland Wales’, Hydrobiologia, 171, 59-78.
Scientific Publication No. 51, Freshwater Biological Association, Windermere.

324 s. n. RUNDLE, E. c. LLOYD AND s. J . ORMEROD
Weatherley, N. S., Ormerod, S. J . and Boole, P. 1985. A Comparison of Two Sampling Methods for Macroinvertebrates
Weatherley, N. S. and Ormerod, S. J . 1990. ‘Forests and the temperature of upland streams in Wales: a modelling
Weatherley, N. S . and Ormerod, S. J. 1992. ‘The biological response of acidic streams to catchment liming compared
Winterbourn, M. J . , Hildrew, A. G. and Box, A. 1985. ‘Structure and grazing of stone surface organic layers in some
in Small Streams, UWIST Acid Waters Report No. 1. UWIST, Cardiff.
exploration of the biological effects’, Freshwater Biology, 24, 109-122.
to changes predicted from stream chemistry’, Journal of Environmental Management, 34, 105- 1 15.
acid streams of southern England’, Freshwater Biology, 15, 363-374.





























