Embed Size (px)
Citation preview

J . Sci. Food Agric. 1980, 31, 877-888
The Distribution of Acyl Lipids in the Germ, Aleurone, Starch and Non-starch Endosperm of F Q U ~ Wheat Varieties
Kevin D. Hargin and William R. Morrison
Departnient of Food Science orid N~itritioii, Universit,v of Strdic lyde, 131 Albion Street, Glasgow GI I S D
(Manuscript received 9 Januarj. 1980)
The quantitative distribution of 23 classes of acyl lipids was determined in the germ, aleurone, starch and endosperm non-starch fractions of Atou (Soft English), Flinor (Hard English), Waldron (US Hard Red Spring) and Edmore (US Amber Durum) wheats. All four wheats had similar proportions (dry basis) of pericarp (6.8-8.6 %), germ (2.5-3.0 %), starchy endosperm (78.7-84.5 %) and starch (59.3-67.5 %), and similar quantities of acyl lipids in the whole kernels (2.8-3.2 :{), germ (25.7-30.5 %). starch (0.8-1.2%) and endosperm non-starch fractions (0.8-1.1 %). Flinor, Waldron and Edmore had 7.3-10% aleurone containing 8.7-10.6 %lipids, but Atou appeared to have a n abnormally low aleurone weight (4.0%) and a correspondingly high lipid content (19.4%). Pericarp acyl lipids were studied only in Atou, where they com- prised 1.3 o,{ of the dry weight and 3.8 7; of the total acyl lipids in the whole kernel. Lipids in the germ and aleurone consisted of triglycerides (60.3-79.3 %), other non- polar lipids (5.6-12.0%) and phospholipids (13.6-17.9%). Starch lipids were almost exclusively lysophospholipids (89.4-94.4 %). Greater variation was found in the endosperm non-starch lipids which consisted of triglycerides (1 3.7-34.1 $.',), other non- polar lipids (33.248.5 :Q, glycolipids (18.6-38.3 %) and phospholipids (21.9-35.3 7;) . Edmore had the highest levels of triglycerides and non-polar lipids, and the lowest levels of glycolipids (as expected in a tetraploid wheat). Atouresembled Edmore in its low levels of steryl esters and glycolipids, but it also had least phospholipids.
1. Introduction
Lipids have important functions in the developing and germinating wheat kernel, they affect many aspects of wheat and flour technology and can provide useful amounts of dietary essential fatty acids.'-3 Data on the quantitative distribution of lipids within the wheat kernel would be of greater value in relation to any of these topics, but the quality of the published data is poor, especially when compared with the information available on flour 1ipids.l In fact, until recently there was no complete analysis of the lipid classes in the individual tissues of any cereal grain.
We have now quantified the principal acyl lipids in the pericarp, germ, starch and endosperm non-starch fractions of three types of maize,4 and have studied changes in the lipids of developing maize kernel^.^ Maize was chosen because it has the largest kernel of any cereal, and i t is thus easier to dissect and to adapt analytical methods to suit each type of tissue.
In this paper the quantitative distribution of 23 classes of acyl lipids in the principal parts of the wheat kernel is described. Four varieties were studied, representing Soft English (Atou), Hard English (Flinor), U S Hard Red Spring (Waldron), and US Amber Duruni (Edmore) wheats.
2. Experimental 2.1. Materials Samples of Atou and Flinor wheats (1976 harvest) were provided by Mr K. Kingswood, R H M Research Lld. High Wycombe, and Waldron and Edmore wheats were provided by Dr B. J.
0022-5142/80/0900-0877 $02.00 (I', 1980 Society of Chemical Industry
877

878 K. D. Hargin and W. R. Morrison
Donnelly, North Dakota State University, Fargo, ND, USA. The wheats were stored as received in sealed jars at 4°C.
2.1.1. Isolation of starch
Batches of wheat were milled on a Buhler mill to give flour (72 % extraction rate) which was mixed to a stiff dough with 21 % glycerol solution ( d = 1.26), then thinned down to a slurry and centrifuged at 108 000 g for 60 min. The grey-yellow proteinaceous layer was removed and the starch recovered for further mixing and centrifuging until no more protein separated out. The recovered starch was then washed in a similar manner with 0.01 % sodium metabisulphite solution (three times) and finally with water. The starch was air-dried and gently ground to pass a 75 pm aperture sieve. The total nitrogen contents of the four starches were 0.063 % (Atou), 0.047 % (Flinor), 0.036 % (Waldron), and 0.085 % (Edmore) (calculated 0.020-0.029 :< for lipid N, assuming N: P molar ratio is 1 : l ) .
The starch content of the kernels was determined from dissected aleurone plus starchy endosperm (see section 2.1.2) which had been ground to pass a 75 pm aperture sieve. This material was boiled with water to gelatinise the starch, and the glucose produced by hydrolysis with amyloglucosidase (from Asperigilliu niger) was then measured colorimetrically by a glucose oxidase method (Karkalas, J., private communication).
2.1.2. Dissection of wheat kernels
Whole kernels were softened for dissection by immersing in boiling water; treatment times of 8-10 min were necessary to prevent formation of enzymic artefacts in germ and aleurone lipids. The kernels were cooled in cold water and stored in a moist atmosphere for dissection within the next hour. It is recognised that some loss of water solubles may have occurred, thereby affecting the accuracy of gravimetric determinations, but the percentage losses were thought to be small, except in Atou aleurone. Comparisons of lipids in germ and in starchy endosperm prepared by wet and dry dissection methods showed no evidence of lipid losses or of transfer of lipids between tissues in kernels treated with boiling water.
Individual kernels were dissected under a microscope ( x 25 or x 50 magnification). The outer pericarp was readily removed and the intact germ (embryonic axis plus scutellum) was then prised out. The remaining pericarp (down to the testa and nucellar layer) was then removed by working a scalpel between the testa and the aleurone layer until ea-iigh had been lifted for the rest to be peeled away using forceps. In this way the aleurone layer WAS t ~ j ~ e d over the entire grain, except for a small region inside the crease, and the fractions obtained cmstituted the total pericarp and the aleurone plus starchy endosperm.
It was not possible to remove the whole aleurone layer from the underlying starchy endosperm, so methods were devised to find the weight of aleurone tissue by difference, The outer pericarp was peeled off kernels which were then bisected longitudinally so that residual outer pericarp could be removed from the crease. The kernel halves were placed in boiling water for a further 10 min to soften the starchy endosperm, and the testa-aleurone sides of the half kernels were then stuck down on to gummed paper. Careful scraping with the back of a scalpel blade gave shells of testa plus aleurone and a mass of scraped starchy endosperm. All fractions were dried and weighed and the weight of the aleurone calculated from (i) outer pericarp plus testa-alerrrone minus total pericarp, and from (ii) aleurone plus starchy endosperm minus scraped starchy endosperm. Results by both methods are combined in Table 1.
After remvoing aleurone lipids (see section 2.2), aleurone plus starchy endosperm material was used for quantification of non-starch and starch lipids. Since triglycerides were found, it was thought that aleurone lipids were still contaminating the starchy endosperm and an alternative approach was therefore used to prepare aleurone-free starchy endosperm. Whole kernels were pearled to remove most of the pericarp, the kernels were split along the crease, and the remaining pericarp, germ and aleurone were cut away with a scalpel. The endosperm halves were then washed in chloroform-methanol [1:1 (v/v)] to remove any aleurone or germ lipid adhering to their surfaces.
Distribution of acyl lipids in wheat 879
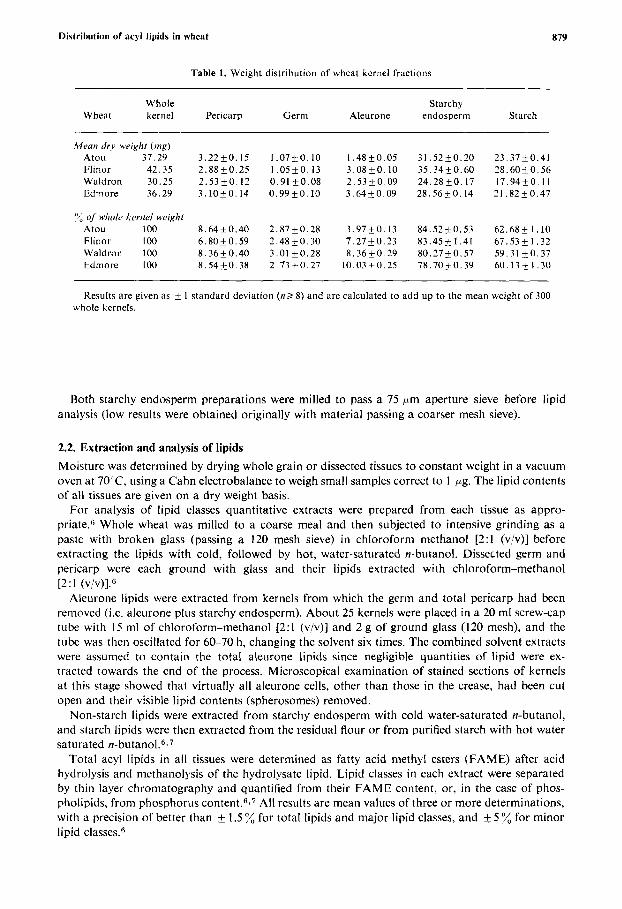
Table 1. Weight distribution of wheat kernel fractions
Whole Starchy Wheat kernel Pericarp Germ Aleurone endosperm Starch
,Wean dr.v weight ( m g ) Atou 37.29 3 .22k0 .15 1.07+0.10 1.48kO.05 31.52k0.20 23 .37k0 .41 Flinor 42.35 2.88k0.25 1.05&0.13 3 . 0 8 k 0 . 1 0 35.34k0.60 28 .60k0 .56 Waldron 30.25 2 .53k0 .12 0.91+0.08 2 . 5 3 k 0 . 0 9 24 .28k0 .17 17.94+0.11 Edmore 36.29 3 . 1 0 k 0 . 1 4 0.99kO.10 3 . 6 4 k 0 . 0 9 2 8 . 5 6 i 0 . 1 4 2 1 . 8 2 k 0 . 4 7
"3 of wholc~ kernel weight Atou 100 8 .64 f0 .40 2 . 8 7 k 0 . 2 8 3.97+0.13 84 .52k0 .53 62.68+1.10 Flinor 100 6 . 8 0 k 0 . 5 9 2.48k0.30 7 .27k0 .23 83 .45k1 .41 67.53k1.32 Waldron 100 8 .3650 .40 3.01 k 0 . 2 8 8.36kO.29 80.27k0.57 59.31 k 0 . 3 7 Edmore 100 8 . 5 4 k 0 . 3 8 2 . 7 3 k 0 . 2 7 10.03kO.25 78 .7020 .39 60.13+1.30
Results are given as k I standard deviation ( n > 8) and are calculated to add up to the mean weight of 300 whole kernels.
Both starchy endosperm preparations were milled to pass a 75 pm aperture sieve before lipid analysis (low results were obtained originally with material passing a coarser mesh sieve).
2.2. Extraction and analysis of lipids Moisture was determined by drying whole grain or dissected tissues to constant weight in a vacuum oven at 70 C. using a Cahn electrobalance to weigh small samples correct to 1 pg. The lipid contents of all tissues are given on a dry weight basis.
For analysis of lipid classes quantitative extracts were prepared from each tissue as appro- priate.6 Whole wheat was milled to a coarse meal and then subjected to intensive grinding as a paste with broken glass (passing a 120 mesh sieve) in chloroform-methanol [2:1 (v/v)] before extracting the lipids with cold, followed by hot, water-saturated n-butanol. Dissected germ and pericarp were each ground with glass and their lipids extracted with chloroform-methanol [Z: 1 (v/v)].6
Aleurone lipids were extracted from kernels from which the germ and total pericarp had been removed ( i s . aleurone plus starchy endosperm). About 25 kernels were placed in a 20 ml screw-cap tube with 15 ml of chloroform-methanol [2:1 (v/v)] and 2 g of ground glass (120 mesh), and the tube was then oscillated for 60-70 h, changing the solvent six times. The combined solvent extracts were assumed to contain the total aleurone lipids since negligible quantities of lipid were ex- tracted towards the end of the process. Microscopical examination of stained sections of kernels a t this stage showed that virtually all aleurone cells, other than those in the crease, had been cut open and their visible lipid contents (spherosomes) removed.
Non-starch lipids were extracted from starchy endosperm with cold water-saturated n-butanol, and starch lipids were then extracted from the residual flour or from purified starch with hot water saturated n-butanol.6s
Total acyl lipids in all tissues were determined as fatty acid methyl esters (FAME) after acid hydrolysis and methanolysis of the hydrolysate lipid. Lipid classes in each extract were separated by thin layer chromatography and quantified from their FAME content, or, in the case of phos- pholipids, from phosphorus All results are mean values of three or more determinations, with a precision of better than 1.5 % for total lipids and major lipid classes, and f 5 % for minor lipid classes.6
880 K. D. Hargin and W. R. Morrison
3. Results and discussion
3.1. Dissection of kernels The dry weights of the whole kernels and their dissected fractions are given in Table 1 . Atou and Flinor had characteristic large plump kernels compared with the smaller kernels of Waldron and the long, thin kernels of Edmore. Germ and pericarp weights were remarkably constant, most of the variation in kernel weight being in the starchy endosperm. The results expressed as percentages are in good agreement with published v a l u e ~ ~ - ~ 3 for pericarp, germ, and starchy endosperm. The proportions of aleurone in Flinor and Waldron were n ~ r m a l , ~ - ~ ~ but were slightly high in Edmore and exceptionally low in Atou.
The low aleurone weight in Atou was investigated using dry kernels fractured transversely, comparing aleurone cell dimensions by scanning electron microscopy. In all four wheats the aleurone layer at mid-section was 27-35 pm thick, and the cells appeared to be almost identical. Calculations showed that the large starchy endosperm and plump shape of the Atou kernels would reduce the weight percentage of a normal aleurone layer by 1-2% compared with the other wheats, but not by the margin shown in Table 1 . Faced with these conflicting observations i t has to be concluded that there is an inexplicable discrepancy in the aleurone dry weight determinations. If this is the case, the Atou aleurone lipid results listed in Table 2 must be artificially high since they were calculated from the satisfactory data in Table 3 and the erroneously low aleurone dry weight.
Table 2. Percentage composition of lipids in wheat kernels and their principal fractions
:d of total acyl lipid Kernel fraction % lipid in ~~
Wheat variety dry fraction Non-polar lipid Glycolipid Phospholipid
Whole kernela Atou Flinor Waldron Edmore
Atou Flinor Waldron Edmore
Aleitrone Atou Flinor Waldron Edmore
Atou Flinor Waldron Edmore
Atou Flinor Waldron Edmore
Germ
Non-starch endosperni
Starch
2.85 2.82 2.93 3 .21
25 .7 30.5 29 .7 29.6
19.4c 8 . 7 8 . 7
10.6
0.85 1.12 1.07 0 .88
0 . 9 8 0 . 8 2 0 .77 1.17
56.9 44 .4 4 9 . 6 56.7
8 4 . 8 83.7 79.3 84 .5
79.5 74 .9 72.3 83.0
42 .8 35.8 33 .2 4 7 . 4
4 . 4 5 . 9 4 . 5 3 . 9
12.5 14.1 14 .4 8 . 0
0 2.0" 3.50 I .9b
6.76 8.06 9 .gb 2.26
35.3 28 .9 38.3 20 .4
5 . 5 1 . 8 I . 2 6 . 7
30.6 41.5 36 .0 35.3
15.2 14.3 17.2 13.6
13.8 17.1 17.9 14.8
21.9 35.3 28.5 32.3
90.1 92.3 94 .3 89.4
a Calculated using summation of lipids in kernel parts instead of whole kernel experimental results
* Tentative identification. CThis value is probably too high-see section 3.1.
(column 1 in Tables 3-6) which were slightly low.

Distribution of acyl lipids in wheat 881
Table 3. Composition o f lipids in Atou (English Soft Winter) wheat kernels (pg lipid/kernel)
Lipid class
SE TG D (2 FFA MGC ASG MGDG M G M G DGDG D G M G NAPE NALPE D E'G PCi PC PE PI LPG LPE L PC LPI, LPSd PAd
Total FAME Total NL Total GL Total PL Total acyl lipid
~
Whole kernel
23.5 446.9 26.8 28.2 13.8 11.7 8.4 4.7 51.2 42.7 9.0 6.8
} 8.3 21.2
6.7 23. I 206.4
7.9
167.7 539.2 118.7 289.4 941.3
~.
Pericarp
5.6 6.6 14.8 9.1 0 . 5 1 .6d
-~
* * * * * * * * * * * * * * * *
37.6 36.6 2.9d 3.0 42.5
Germ Aleurone
1.5 218.3
5.6 2.2
-
- - -
-
- - -
1.7 1.1 5.7 23.4 7.9
1 .o 1 .o
266.0 233.6
41.8 275.4
-
10.0 206.4 3.7 5.9 2.4 6.5 3.5d 3.3d 1.6" 4.3d - -
1 . 5
} 3.1 25.9 5.5
2.5 1 . 1
261.9 228.4 19.2 39.6 287.2
Starchy endosperm
Non-starch" Non-starchb Starchn
1.2 69.3 17.7 15.8 6.9 13.2 8.5 5 . 0 45.7 24.0 8.2 6.6
4.1 70.0 14.6 19.4 13.4 9.2 8.8 5.1 54.5 4.6 22.4 15.7
1.3 1.5
5.0 1.9
1.4 1.2 0.9 5.4 1.2 2.8 1.4 2.6 1.6 4.2
29.9 12.4 174.6 I .2
3.5 1.7
199.5 201.7 222.5 116.9 122.1 10.1 96.4 82.2 12.6 59.9 63.2 206.4 273.2 267.5 229.1
~~~~~
Peeled, aleurone-defatted kernels.
May include some 6-0-acylmonogdlactosyldiglyceride. '> Pearled kernels.
" Tentative identification. 'l Not measured.
The percentage lipid composition of the kernels and their principal fractions are shown in Table 2. Published v a l u e s l ~ ~ ~ for the lipids in whole wheat, which would include little or no starch lipids, range from 2.1 to 3.8 %. The four wheats used in the present study were in the middle of this range and had more polar lipids than previously recorded14 because starch lysophospholipids were also quantified. Detailed analyses of lipid classes in each part of the wheat kernel are given in Tables 3-6 and are discussed in the appropriate sections below. The fatty acid compositions of the various lipid classes showed no significant differences among the four wheats (except in the non-starch steryl esters), and are therefore not discussed.
3.2. Pericarp lipids The bulk of the acyl lipids in Atou pericarp were non-polar (Table 3), as in maize.4 No attempt was made to measure unsaponifiable matter which probably formed the bulk of the lipids (cf. maize4), or to quantify galactosylglycerides and phospholipids. In view of the low levels of acyl lipids in Atou pericarp, and the fact that they are of little significance in wheat technology, it was decided not to study lipids in the pericarp of the other wheats. The high levels of diglycerides and free fatty acids in .4tou pericarp (Table 3) suggest that lipid degradation occurred during pericarp senescence and kernel ripening, as was demonstrated in maize.5
882 K. D. Hargin and W. R. Morrison
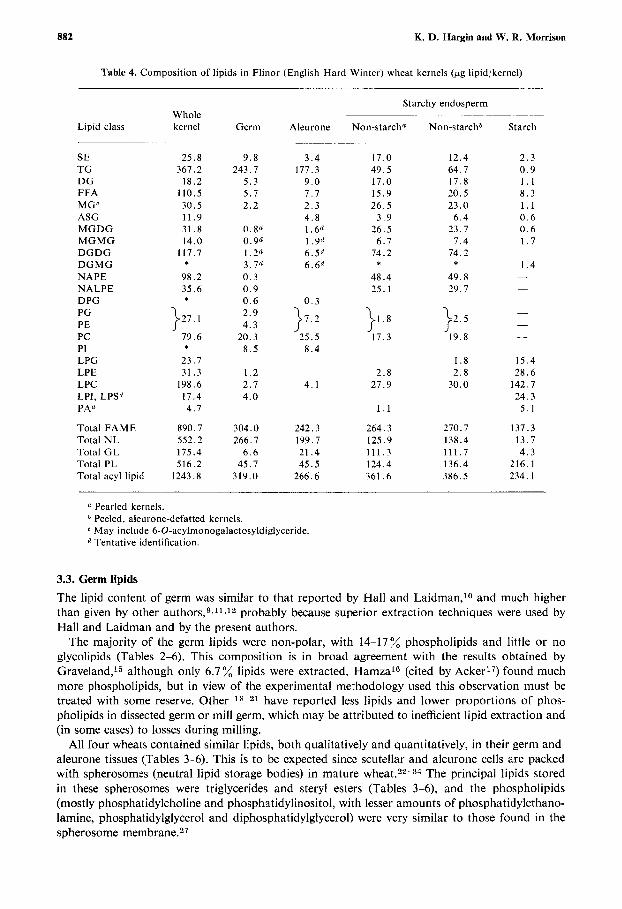
Table 4. Composition of lipids in Flinor (English Hard Winter) wheat kernels (pg lipidikernel)
Lipid class
SE TG DG FFA MG' ASG MGDG MGMG DGDG DGMG NAPE NALPE DPG PG PE PC PI LPG LPE LPC LPI, LPS" PA6
Total FAME Total NL Total G L Total PL Total acyl lipid
Whole kernel
~
25 .8 367,2
18.2 110.5 30.5 11.9 31.8 14.0
117.7
98.2 35.6
j 2 7 . 1
79 .6
23.7 31.3
198.6 17.4 4 . 7
890.7 552.2 175.4 516.2
1243.8
*
*
*
Germ Aleurone
9.8 243.7
5.3 5.7 2.2
0.8" 0.9d I .2d 3.7'' 0 . 3 0 . 9 0 .6 2 .9 4 .3
20.3 8 . 5
1 .2 2.7 4 .0
304.0 266.7
6 .6 45.7
319.0
3 . 4 177.3
9 . 0 7 .7 2 .3 4.8 1.6d 1.9" 6 .5d 6.6d
0 . 3
}7.2
25.5 8 . 4
4.1
242.3 199.7 21.4 45.5
266.6
Starchy endosperm
Non-starchn Non-starchu Starch ~~~ -~ _ _ ~~~
- ~
17 .0 49 .5 17 .0 15.9 26.5
3 .9 26.5
6 . 7 74.2
48.4 25.1
*
12.4 64.7 17.8 20.5 23.0
6 . 4 23.7
7 . 4 74.2
49.8 29.7
*
2 .3 0 . 9 1 . 1 8 . 3 1 .1 0 . 6 0 . 6 1 . 7
1 . 4 -
-
-
j I . 8 )2.5 - 17 .3 19.8 -
1 .8 15.4 2 .8 2 . 8 28.6
27.9 30.0 142.7 24.3
1 . 1 5 . I
264.3 270.7 137.3 125.9 138.4 13.7 111.3 111.7 4 . 3 124.4 136.4 216.1 361.6 386.5 234.1
Pearled kernels. Peeled, aleurone-defatted kernels. May include 6-0-acylmonogalactosyldiglyceride.
l7 Tentative identification.
3.3. Germ lipids The lipid content of germ was similar to that reported by Hall and Laidman,lo and much higher than given by other a u t h o r ~ , ~ , 1 ' ~ 1 ~ probably because superior extraction techniques were used by Hall and Laidman and by the present authors.
The majority of the germ lipids were non-polar, with 14-17% phospholipids and little or no glycolipids (Tables 2-6). This composition is in broad agreement with the results obtained by Graveland,15 although only 6.7 % lipids were extracted. HamzalG (cited by Acker17) found much more phospholipids, but in view of the experimental methodology used this observation must be treated with some reserve. Other 18-21 have reported less lipids and lower proportions of phos- pholipids in dissected germ or mill germ, which may be attributed to inefficient lipid extraction and (in some cases) to losses during milling.
All four wheats contained similar lipids, both qualitatively and quantitatively, in their germ and aleurone tissues (Tables 3-6). This is to be expected since scutellar and aleurone cells are packed with spherosomzs (neutral lipid storage bodies) in mature ~ h e a t . ~ 2 - 3 ~ The principal lipids stored in these spherosomes were triglycerides and steryl esters (Tables 3-6), and the phospholipids (mostly phosphatidylcholine and phosphatidylinositol, with lesser amounts of phosphatidylethano- lamine, phosphatidylglycerol and diphosphatidylglycerol) were very similar to those found in the spherosome membrane.27
Distribution of acyl lipids in wheat 683
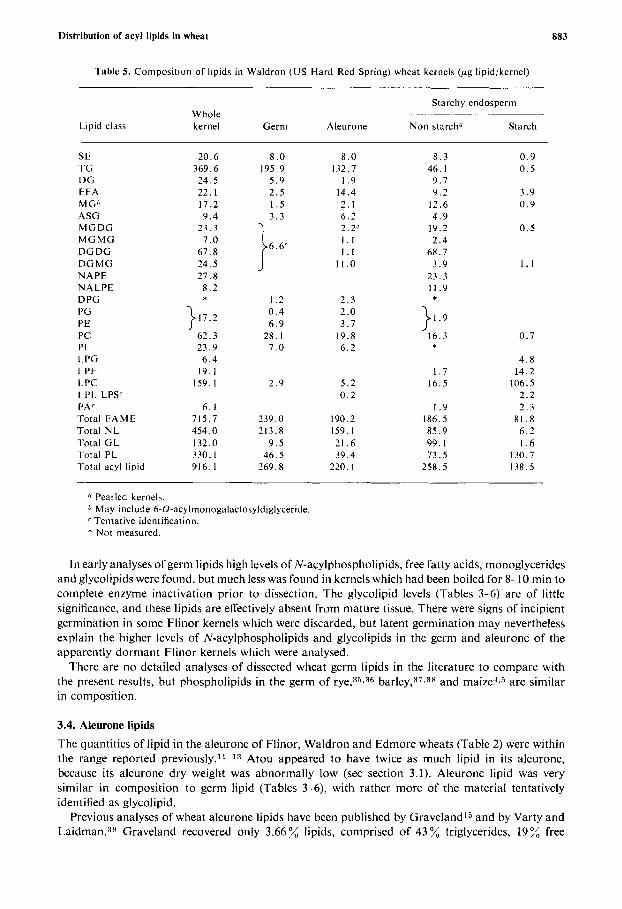
l'able 5. Composition of lipids in Waldron ( U S Hard Red Spring) wheat kernels (pg lipidikernel)
Lipid class
Starchy endosperm ~ ~-~ Whole
kernel Germ Aleurone Non-starch" Starch
SE TG DG F FA M G 1' ASG MGDG MGMG DG DG DGMG NAPE NALPE D PG I'G PE PC PI L.PG L.PE LPC LPI, LPSC PA' Total FAME Total N L Total GL Total PL
24.5 22.1 17.2 9.4 23.3 7.0 67.8 24.5 27.8 8.2
117.2
62.3 23.9 6.4 19.1 159. I
6. I 715.7 454.0 132.0 330. I
*
5.9 2.5 I .5 3.3
]6.6?
I .2 0.4 6.9 28.1 7.0
2.9
239 .0 213.8 9.5 46.5
20.6 8.0 8.0 369.6 195.9 132.7
I .9 14.4 2.1 6.2 2.2c 1 . 1 1 .1
11.0
2.3 2.0 3 . 7 19.8 6.2
5.2 0 . 2
90.2 59.1 21.6 39.4
Totol acyl lipid 916.1 269.8 220. I
8.3 46. I 9.7 9.2 12.6 4.9 19.2 2.4 68.7 3.9 23.3 11.9
}l.9
16.3
*
*
1.7 16.5
1.9 86.5 85.9 99. I 73.5 258.5
0.9 0.5
3.9 0.9
0.5
1.1
0.7
4.8 14.2 106.5 2.2 2.3 81.8 6.2 1.6
130.7 138.5
' I Pearled kernels. May include 6-0-acylmonogalactosyldiglyceride Tentative identification.
* Not measured.
1 n early analyses of germ lipids high levels of N-acylphospholipids, free fatty acids, monoglycerides and glycolipids were found, but much less was found in kernels which had been boiled for 8--10 min to complete enzyme inactivation prior to dissection. The glycolipid levels (Tables 3-6) are of little significance, and these lipids are effectively absent from mature tissue. There were signs of incipient germination in some Flinor kernels which were discarded, but latent germination may nevertheless explain the higher levels of N-acylphospholipids and glycolipids in the germ and aleurone of the apparently dormant Flinor kernels which were analysed.
There are no detailed analyses of dissected wheat germ lipids in the literature to compare with the present results, but phospholipids in the germ of rye,35,36 bar le~,3~,38 and maize4q5 are similar in composition.
3.4. Aleurone lipids The quantities of lipid in the aleurone of Flinor, Waldron and Edmore wheats (Table 2) were within the range reported p r e v i ~ u s l y . ~ ~ - ~ ~ Atou appeared to have twice as much lipid in its aleurone, because its aleurone dry weight was abnormally low (see section 3.1). Aleurone lipid was very similar in composition to germ lipid (Tables 3-6), with rather more of the material tentatively identified as glycolipid.
Previous analyses of wheat aleurone lipids have been published by GravelandI5 and by Varty and L a i d n ~ a n . ~ ~ Graveland recovered only 3.66% lipids, comprised of 43 % triglycerides, 19% free
884 K. D. Hargin and W. R. Morrison
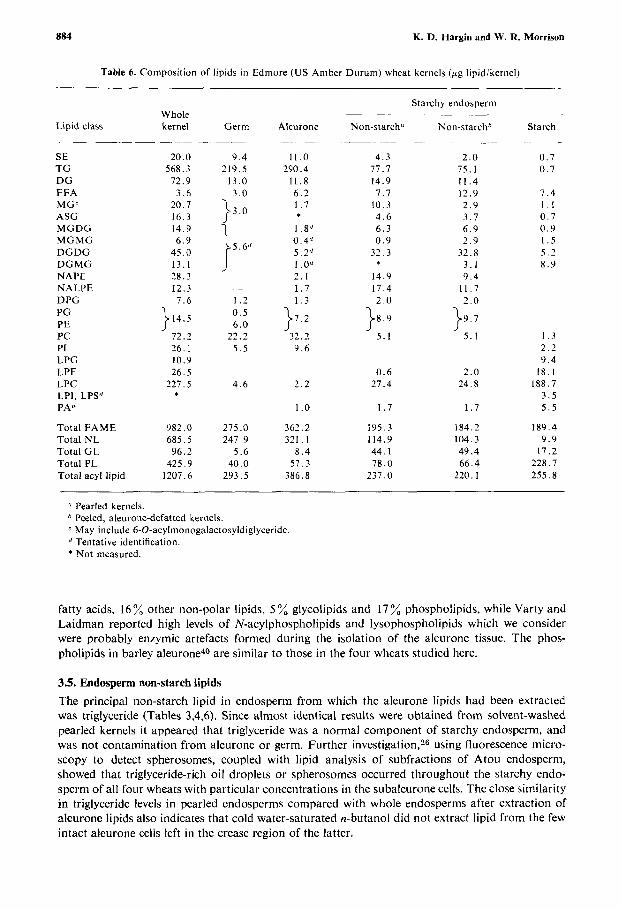
Table 6 . Composition of lipids in Edmore (US Amber Durum) wheat kernels (pg lipidikernel)
Lipid class
SE TG DG FFA MGc ASG MGDG MGMG DGDG DGMG NAPE NALPE DPG PG PE PC PI LPG LPE LPC LPI, LPS" PAd
Total FAME Total N L Total G L Total PL Total acyl lipid
Whole kernel
~ ~~
20 .0 568.3
72.9 3.6
20.7 16.3 14.9 6.9
45.0 13.1 28.3 12.3 7.6
> 1 4 s
72.2 26.1 10.9 26.5
227.5 *
982.0 685.5
96.2 425.9
1207.6
Germ kleurone
Starchy endosperm -_ Non-starcha Non-starch"
9 . 4 219.5
13.0 3 .0 i 5.6''
-
-
1.2 0 .5 6 .0
22.2 5.5
4.6
275.0 247.9
5.6 40.0
293.5
11.0 290.4
11 .8 6 . 2 I . 7
1.8" 0. 411 5.2'1 1 .Of/ 2 . I 1.7 1.3
*
}7.2
32.2 9.6
2.2
1 .o 362.2 321.1
8.4 57.3
386.8
4 .3 77.7 14.9 7.7
10.3 4 . 6 6.3 0.9
32.3
14.9 17.4 2 . 0
}8.9
5.1
*
2.0 75.1 11.4 12.9 2 .9 3.7 6 .9 2 .9
32.8 3.1 9.4
11.7 2 . 0
>9 .7
5.1
0 .6 2.0 21.4 24.8
I .7 1.7
195.3 184.2 114.9 104.3 44.1 49.4 78.0 66.4
237.0 220.1
Starch ~
0 .7 0 .7
7.4 1 1 0 . 7 0 .9 I . 5 5.2 8.9
I . 3 2 .2 9 . 4
18.1 188.7
3.5 5 .5
189.4 9.9
17.2 228.7 255.8
Pearled kernels. Peeled, aleurone-defatted kernels. May include 6-0-acylmonogalactosyldiglyceride. Tentative identification.
* Not measured.
fatty acids, 16% other non-polar lipids, 5 % glycolipids and 17% phospholipids, while Varty and Laidman reported high levels of N-acylphospholipids and lysophospholipids which we consider were probably enzymic artefacts formed during the isolation of the aleurone tissue. The phos- pholipids in barley a l e ~ r o n e ~ ~ are similar to those in the four wheats studied here.
3.5. Endosperm non-starch lipids The principal non-starch lipid in endosperm from which the aleurone lipids had been extracted was triglyceride (Tables 3,4,6). Since almost identical results were obtained from solvent-washed pearled kernels it appeared that triglyceride was a normal component of starchy endosperm, and was not contamination from aleurone or germ. Further investigation,26 using fluorescence micro- scopy to detect spherosomes, coupled with lipid analysis of subfractions of Atou endosperm, showed that triglyceride-rich oil droplets or spherosomes occurred throughout the starchy endo- sperm of all four wheats with particular concentrations in the subaleurone cells. The close similarity in triglyceride levels in pearled endosperms compared with whole endosperms after extraction of aleurone lipids also indicates that cold water-saturated n-butanol did not extract lipid from the few intact aleurone cells left in the crease region of the latter.

Distribution of acyl lipids in wheat 885
Greatest variation between the wheats was seen in the galactosylglycerides and diacylphos- pholipids. Since there is no means of determining whether the lysophospholipids were non-starch lipids or starch lipids which had leaked prematurely from the starch granules, they are excluded from this discussion.
The ratio of digalactosyldiglyceride : monogalactosyldiglyceride in Flinor and Waldron was about 3:1 (Tables 4,5), similar to the ratio in most flours from hexaploid wheats.2,7.J1-J5 Edmore had much less total glycolipids (Table 2) and the ratio of galactoslyglycerides was 5:1 (Table 6). These features, and the low level of steryl ester, are associated with the absence of the D-genome in durum ~ h e a t . 1 . ~ 6 , ~ ~ Atou had a comparatively high percentage of glycolipids in its total lipids (Table 2), but it had the lowest levels of all lipids, including glycolipids, when expressed as mg lipid 100 g-' dry weight. The ratio of galactosylglycerides in Atou (6:l) was even higher than in Edmore, and there was a low level of steryl ester (mostly linoleate), which suggests that Atou may have recessive lipid characters on the D-genome.
'The principal non-starch phospholipids in all four wheats were the N-acylphosphatidylethano- lamines and phosphatidylcholine, Atou and Edmore having much less than Flinor and Waldron. These are also the major non-starch phospholipids in white flour.7J6,41-45 N-Acylphosphatidyl- ethanolamines may be chemical or enzymic artefact^,^"^' and indeed, in the present study small quantities were formed in the germ if the kernels had received insufficient treatment in boiling water. However the N-acylphosphatidylethanolamines are not regarded as artefacts in f l o ~ r , ~ ~ , ~ s and since they were found in pearled kernels which had received no wetting in the laboratory, it seems most unlikely that they were experimental artefacts. This leaves the intriguing possibility that they were formed during the later stages of kernel maturation and should be recognised as natural lipids in wheat kernels.
3.6. Starch lipids It is now well established that wheat starch lipids are almost exclusively lysophospholipids.l~~~7~45~5~ The starch preparations used in this study had low levels of non-lipid nitrogen, but nevertheless appeared to contain traces of typical non-starch lipids such as triglyceride, diglyceride and galacto- syldiglycerides. If these are discounted, the starch lipids were almost exclusively lysophospholipids, with small amounts of free fatty acids, monoglycerides and galactosylmonoglycerides. Atou and Edrnore starches had more Iysophosphatidylcholine than Flinor and Waldron starches; this may be a genetically-determined character similar to the non-starch features discussed previously, but there is no precedent for this suggestion.
3.7. Whole kernel lipids Experimental values for most lipid classes in the whole kernel were in acceptable agreement with totals for germ, aleurone, starch and non-starch lipids (Tables 3-6). The principal lipid was tri- glyceride, mostly located in the germ and aleurone, but with significant amounts in the starchy endosperm. Steryl ester, diglyceride and monoglyceride were fairly evenly distributed in these three tissues, together with variable amounts of free fatty acids. The high level of free fatty acids in Flinor whole kernels was probably caused by an unintended delay between milling and extraction of the lipids, and indicates the potential for lipolysis in whole wheatmeal.
Glycolipids were the smallest group in the whole kernel (Table 2) and were almost exclusively in the endosperm non-starch fraction (Tables 3-6). Small quantities tentatively identified in germ and aleurone may have been wholly or partially artefacts.
The whole kernels contained appreciable quantities of phospholipids which were mostly in the starch. De la Roche et aL50 extracted similar diacylphospholipids, but obtained very poor recoveries of slarch 1 ysophospholipids.
Diphosphatidylglycerol and phosphatidylinositol were found only in germ and aleurone, the N-acylphosphatidylethanolamines only in the endosperm non-starch fraction, and lysophospho- lipids almost entirely in starch. Unfortunately, these lipids are not suitable markers for the various tissues because they are either minor components (diphosphatidylgycerol, phosphatidylinositol),

886 K. D. Hargin and W. R. Morrison
or are liable to be formed in other tissues as artefacts (N-acylphosphatidylethanolamines) or as products of lipolysis (lysophospholipids).
4. Conclusions
The results presented here give for the first time a comprehensive picture of the distribution of lipids in the prinicpal parts of the wheat kernel, and the pattern may be representative (with minor modifications) of the distribution of lipids throughout the Pooideae (wheat, barley, rye, oats and triticale). The findings complement previous studies of lipids in three types of maize kernels,4~5 and show many common features as well as some important differences. The germ (and probably also the aleurone) lipids are very similar in both cereals, although maize has a much larger germ with a higher lipid content. l ~ 5 Wheat and maize starch lipids are exclusively monoacyl and are therefore capable of forming amylose inclusion complexes, but maize starch has much more free fatty acids and less lysophospholipids4 than wheat starch. The greatest differences are seen in the endosperm non-starch lipids which are triglycerides, glycolipids and phospholipids in wheat, whereas in maize they are mostly free fatty acids formed by degradation of these lipids in later stages of kernel development.5
The results in this study are also consistent with light and electron microscopic evidence on the distribution of lipids. Triglycerides are typical storage lipids in the spherosomes which are seen in large numbers in the mature scutellum and a l e u r ~ n e , ~ ~ - ~ ~ and in lesser quantities in the embryo (Angold, R . E., private communication) and subaleurone and inner starchy endosperm cells.2fi Glycolipids are probably membrane lipids in some unidentified organelle($ present only in the starchy endosperm (the amyloplast membrane?), and diacylphospholipids are known to be ubiqui- tous in all biomembranes. The high levels of lysophospholipids in wheat and other cereal starches are unique, and to the best of our knowledge they have not been recorded as the major class of lipid in any other healthy tissue of plant, animal or microbial origin.
Acknowledgements K. D. Hargin received a CASE award from the Science Research Council in collaboration with RHM Research Ltd; Mr K . Kingswood, Dr T. Galliard and Dr R. E. Angold gave much help and Dr Angold provided all microscopy services.
References I .
2.
3. 4.
5.
6.
7.
8.
9.
10.
11.
Morrison, W. R. Cereal lipids. Advances in Cereal Scierrce and Technology (Pomeranz, Y., Ed.), Vol. 2, Am. Ass. Cereal Chem., St. Paul, Minn., 1977, pp. 221-348. Morrison W. R. Lipids in wheat and their importance in wheat products. Recerrf Advances in the Biochemistry of Cerea1.v (Laidman, D. L.; Wynn Jones, R . G., Eds), Academic Press, London, New York, San Francisco, 1979, pp. 3 13-335. Morrison, W. R. Cereal lipids. Proc. Nufr. SOC. 1977, 36, 143-148. Tan, S. L. ; Morrison, W. R. The distribution of lipids in the germ, endosperm, pericarp and tip cap of amylo- maize, LG-I 1 hybrid maize and waxy maize. J . Am. Oil Chem. SOC. 1979, 56, 531-535. Tan, S . L.; Morrison, W. R. Lipids in the germ, endosperm and pericarp of the developing maize kernel. J . Am. Oil Cliem. SOC. 1979, 56, 759-764. Morrison, W. R.; Tan, S . L.; Hargin, K. D. Methods for the quantitative analysis of lipids in cereals and similar tissues. J . Sci. Food Agric. 1980, 31, 329-340. Morrison, W. R.; Mann, D. L.; Wong, S . ; Coventry, A. M . Selective extraction and quantitative analysis of non-starch and starch lipids from wheat flour. J. Sci. Food Agric. 1975, 26, 507-521. Heathcoate, J. G.; Hinton, J. J. C.; Shaw, B. The distribution of nicotinic acid in wheat and maize. Proc. R . SOC., B 1952, 139, 216-287. Geoffroy, R. Character of specific constituents of wheat grain, germ and hemicelluloses. Tech. Ind. CPriali2res 1974, 145, 3-6. Hall, G. S.; Laidman, D. L. The determination of tocopherols and isoprenoid quinones in the grain and seedlings of wheat (Triticum vdgare). Biochem. J . 1968, 108, 465-473. Inglett, G . E. Kernel structure and composition. Wheat Production ond U/ilization AVI, Westport, Conn., 1974, pp. 108-118.

Distrihution of acyl lipids in wheat 887
12.
13.
14.
15. 16. 17. 18.
19.
20.
21.
23.
24.
2 5 .
26.
’7.
28.
29.
30.
31.
32.
13.
14.
35. 36. 37.
38.
79.
40.
41.
42.
43. 44.
45.
MacMasters. M. M.; Hinton. J. J. C.; Bradley, D. Microscopic structure and composition o f the wheat kernel. Il.%~ar: Chf,rni\trJ and Teer~hrlolugy (Pomeranz, Y.. Ed.), Am. Ass. Cereal Chem, St. Paul. Minn., 1971, pp. 51-113. Shetlar, M. R.: Rankin, G. T.; Lyman, J. F . ; France, W. G. Investigation o f the proximate chemical coni- position o f the separate bran layers o f wheat. Cereal Chern. 1947, 24, 1 1 1-122. Pomeranz. Y.; Chung, 0.; Robinson, R. J. Lipids in wheat from various classes and varieties. J. Am. Oil C l r ~ t ? ~ . Soc. 1966, 43, 51 1-514. Graveland, A. A micromethod for research on wheat lipids. TNO Nierrivr 1969, 24, 472-477. Hamza, Y . PhD Thesis, University o f Munster, Westf., 1968. Acker, L. Cereal lipids: Their composition and significance. Getreidr Mehl Brot 1974. 28, 181-187. Bolling, H. ; El Baya, A . E. ; Zwingelberg, H. Germ fat and baking quality o f wheat. Getreirk Mehl Brot 1973, 27, ‘)2-94. Moruzzi. G . : Viviani, R.; Sechi, A. M . ; Lenaz. G. Studies ofconipound and individual lipids o f wheat germ. J . Food Sc i. 1969, 34, 58 1-584. Nelson, J. H.; Glass, R. L.; Geddes, W. F. Silicic acid column chromatography o f wheat lipids. Ccreal Chem. 1963. 40. 337-343. Pomeranz, Y. ; Chung, 0. The lipid composition o f a single wheat kernel and its structural parts. J. Chroiirarogr. 1965, 19. 540-550. Ayre, L.: Angold, R. E. The occurrence o f transfer cell structures in relation to the developing aleurone layer o f the wheat caryopsis. Rerenf Adwnrrs in rhe Biochemisfry of Cvreals (Laidman, D. L.: Wyn Jones, R. G., Izds), Academic Press, London, New York. San Francisco, 1979, pp. 359-366. I3rodbury. D . : MacMasters, M. M.; Cull, I. M . Structure o f the niature wheat kernel. 111. Microscopic struc- ture o f the endosperm o f hard red winter wheat. Cereal Chern. 1956, 33, 361-373. Bradbury. D.; MacMasters. M. M.; Cull, 1. M. Structure o f the mature wheat kernel. IV. Microscopic struc- ture o f the germ o f hard red winter wheat. Crrtwl Cherri. 1956. 33, 373-391. Colborne. A. J . ; Morris, G.; Laidman, D. L. The formation o f endoplasmic reticulum in the aleurone cells of germinating wheat: an ultrastructural study. J . E.vper. But. 1976, 27, 759-767. Hargin. K. D.: Morrison, W. R.; Fulcher, R. G. Triglyceride deposits i n the starchy endosperm of wheat. Cerral C h ~ r i ~ . (in press). Jelsenia, C. L.; Morre, D. J . ; Ruddat, M.; Turner, C. Isolation and characterisation o f the l ipid reserve bodies, spherosomes, from aleurone layers o f wheat. Bor. Go.-. 1977, 138, 138-149. Morrison, I . N.: Kuo, J . : O’Brien, T. P. Histochemistry and tine structure o f developing wheat aleurone cells. P/rrrro 1975. 123, 105-1 16. Morrison, I . N . : O’Brien. T. P.; Kuo, J. Ini t ial cellularisation und difierentiation in the aleurone cells i n the ventral region o f the developing wheat grain. Plcmta 1978, 140, 19-30. Nougarede. A, ; Pilet, P.-E. Infrastructure of the scutellar cells o f Triticrrm vulgare Vill. during the first stages or germination. C. R . Acod. Sci. Paris. 1964, 258. 264-2644. Simmonds, D. H. Structure, composition and biochemistry o f cereal grains Cereals ’78: Better Nutrition for. rhe IVor/t/’.s MiNioris (Pomeranz, Y., Ed.), Am. Ass. Cereal Chem. St. Paul. Minn., 1978, pp. 105-137. Swift, J . G . ; Buttrose, M . S. Freeze-etch studies o f protein bodies in wheat scutellum. J . Ulfrasrrrrcf. Rm. 1972, 40, 378-390. Swift, J. G.; O’Brien, T. P. The fine structure o f wheat scutellum before germination. Airst. J . B i d . Sci. 1972, 25. 9-22. Swift, J. G. ; O’Brien, T. P. The fine structure o f wheat scutellum during germination. Arrst. J . B i d . Sci. 1972, 25, 469--486. Thomson. L. W.: Zalik. S. Lipids i n rye seedlings i n relation to vernalization. Platit Phyriul. 1973. 52, 268-273. Acker, L.. Schmitz. H. J . ; Hamza, Y . The lipids o f wheat. Getrricle Mehl Brot 1968, 18, 45-50. Holmer. G . ; Ory, R. H.; Hoy. C. E. Changes in l ipid composition of germinating barley embryo. lipid.^ 1973, 8. 277-283. Price, P. B.; Parson, J. Distribution o f lipids in embryonic axis, bran-endosperm, and hul l fractions of hulless barley and hulless oat grain. J . Agric. Food Chent. 1979. 27, 8 13-8 15. Varty, K . : Laidman, D. L. The pattern and control o f phospholipid metabolism in wheat aleurone tissue. J . E.uprr. Bot. 1976, 27. 748-758. Firn, R. D.; Kende, 14. Some effects of applied gibberelic acid on the synthesis and degradation of lipids in iaolated barley aleurone layers. Plairt Phy.\iol. 1974, 54, 91 1-915. Clayton, T. A,; Morrison, W. R. Changes in flour lipids during the storage o f wheat flour. J . Sci. Food Agric. 1072, 23, 721-736. Lin, M. J . Y.; Youngs, V. L.; D’Appolonia. B. L. Hard red spring and durum wheat polar lipids. 1. Isolation and quantitative determination. Cereal Cherrr. 1974. 51, 17-33. MacMurray, T. A.: Morrison, W. R. Composition o f wheat flour lipids. J . Sci. Food Agric. 1970, 21, 520-528. Mann, D. L,.; Morrison, W. R. Changes in wheat lipids during mixing and resting o f flour-water doughs. J , Sci. Food Agric . 1974, 25, 1109-1 119. Morrison. W. R. The stability o f wheat starch lipids i n untreated and chlorine-treated cake flours. J . Sci. h o d Agric. 1978, 29, 365-371.

888 K. D. Hargin and W. R. Morrison
46.
47.
48.
49.
50.
51.
52.
Hernandez-Lucas, C. ; Fernandez de Caleya, R.; Carbonero, P. : Garcia-Olmedo, F. Reconstitution of petro- leum ether soluble wheat lipopurothionin by binding of digalactosyldiglyceride to the chloroform-soluble form. J . Agric. Food Chein. 1977, 25, 1287-1289. Hernandez-Lucas, C. : Fernandez de Caleya, R. ; Carbonero, P.; Garcia-Olmedo, F. Control of galactosyl- diglycerides in wheat endosperm by group 5 chromosomes. Genetics 1977, 85, 521-527. Bomstein, R. A. A new class of phosphatides isolated from soft wheat flour. Biochem. Biophys. Res. Comnrurr. 1965, 21, 49-54. Colborne, A. J. ; Laidman, D. L. The extraction and analysis of wheat phospholipids. Phytocheniistry 1975, 14, 2639-2645. De la Roche, I . A , ; Andrews, C. J.; Kates, M . Changes in phospholipid composition of a winter wheat cultivar during germination at 2 deg. and at 24 deg. Plorir Physiol. 1973, 51, 468-473. Wren, J. J.; Merryfield, D. S. Precursors of N-acylethanolamines in hen’s egg-yolk lipid. Biochim. Biophys. Acta 1965, 98. 589-597. Becker, G.; Acker, L. Die Lipide der Getreidestroerken Monogr. V1-5 (Ullniann, M. ; Ed.), Paul Parey, Hamburg, 1976.




![Endosperm and Imprinting, Inextricably Linked1[OPEN] · In maize, endosperm is de-fined asESR(embryosurroundingregion, analogousto micropylar), starchy, aleurone, or BETL (basal](https://img.dokumen.tips/doc/110x75/60981ac179887c077266e563/endosperm-and-imprinting-inextricably-linked1open-in-maize-endosperm-is-de-ined.jpg)




![Endosperm and Imprinting, Inextricably Linked1[OPEN] · Endosperm pro-liferation affects final seed size—a greater number of endosperm cells is generally correlated with bigger](https://img.dokumen.tips/doc/110x75/5fcbefad1c6189578942e363/endosperm-and-imprinting-inextricably-linked1open-endosperm-pro-liferation-affects.jpg)



















