Embed Size (px)
Citation preview

ECOGRAPHY 21: 573-580. Copenhagen 1998
The breeding productivity of dark-bellied brent geese and curlewsandpipers in relation to changes in the numbers of arctic foxesand lemmings on the Taimyr Peninsula, Siberia
Ron W. Summers, Les G. Undcrhill and Evgeny E. Syroechkovski Jr.
Summers, R, W,. Undcrhill. L. G. iind Syroechkovski, E, E, Jr, 1998, The breedingprodiictiviiy ol" dark-bellied breni geese and curlew sandpipers in relation lo changesin the numbers of arctic loxes and lemmings on ihe Taimyr Peninsula. Siberia. -Ecography 21: 57? 580,
There were aboul ihree-year cycles in ihe populations of arctic foxes, and thebreeding productivities of brenl geese and curleu sandpipers on the Taimyr Penin-sula, Russia, The popukuions of arctic foxes and lemmings changed in synchrony.The breeding productivities of the birds tended to be good when the arctic foxes wereincreasing in numbers and poor when the arctic foxes were decreasing. There was anegati\e relationship belween arctic fox numbers (or occupied lairs) and the breedingproducti\ity of brent geese in the following year. Although there was evidence ofwide-spread synchrony In the lemming cycle across the Taimyr Peninsula, somelocalities showed differenees, Howe\er, such sites would still have been influenced bythe general pattern of fox abundance in the typical tundra zone of the TaimyrPeninsula, where most of the arctic foxes breed and from vvhicii extensive movementsof fo.xes occur after a decline in lemming numbers. The results support a prey-switch-ing hypothesis (also known as the alternative prey hypothesis) whereby aretie foxes,and other predators, feed largely on lemmings when these are abundant or increasing,but switch to birds when the lemming population is small or declining. Therelation,ships between arctic foxes, lemmings and brent geese may be further influ-enced by snowny owls which create fox-exclusion zones around their nests, thusproviding safe nesting areas for the geese.
R. W. Summers. Royal Sociery for ihc I'niledioti of Birds, Eiire House. BeechwoiulPark. Imerness, Svoiiaiul ll'2 3Bii'. - L. G. Viidei-hili Aiiaii Demography Unit. Deploj Sunisiii-al Sciences. Uiiir. of Cape Toivii. Roiideho.sch. South Africa. 770L - E. E.Syroechkov.ski Jr.. Insl. of Ecology eiml Erolunon. Russian Academy of Sciences, 33Leninsky Prospcci. Moscow. Russia.
Summers and Utiderhill (1987) showed thai Ihere is aca three-year cycle in the breeding productiviiy of dark-bellied brent geese Brantu b. bernicla correlated with theabundance, or change in abundance, oflcmmings Lcui-nnis sihiricus and Dicro.slonyx iorqualu.s on the TaimyrPeninsula where this subspecies of brent goose breeds.Similar patterns in breeding productivity and correla-tions with the lemming cycle have also been found forwaders breeding in this part of Siberia: curleu' sand-pipers Calidris ferrughu'ci. sanderlings C. alha. knots C.caniitus and turnstones Araniria ititcvprcs (Cramp and
Simmons 1983, Underbill 1987. Summers and Undcrhill1987. Summers et al, 1989. Underhili et al. 1989). Inaddition. Prokosch (1988) has shown a correlation be-tween the breeding productivity of brent geese and greyplovers Phnicilis squalawla.
Supporting the earlier idea of Roselaar (1979), Sum-mcrs (1986) and Summers and Underhili (1987) pro-posed that prey-switching by arctic foxes Alopexhii^opu.w and perhaps other predators, was the cause ofthese correlations. They suggested that predators feedprimarily on lemmings when these are abundant, and
Accepted 25 May 1998
Copyright © ECOGRAPHY 1998ISSN 0906-7590Primed in Ireland - all rights reserved
2l;(i (1998) 573

the birds" nests and young receive little predation,resulting in good breeding productivity by the birds.However, when lemmings arc scarce, the predators preymore heavily on the birds, causing poor breeding pro-ductivity. There are now direct observations at onelocality on the Taimyr Peninsula showing thai thisoccurs (Underhill et al. 1993).
When the above hypothesis was formulaied, dala onthe distribution and annual changes in the numbers ofarctic foxes were not considered and fox predationlevels were inferred from the changes in the size of thelemming population. Fox population data are nowavailable, allowing a re-assessment. Further, the dataon lemmings which Summers and Underhill (1987)originally used may not have been representative of theTaimyr Peninsula as a whole. The lemming data re-ferred largely to the typical tundra zone, south of lhearctic tundra zone where the above birds mainly breed(Fig. 1). If lemming cycles are asynchronous within theTaimyr Peninsula, then ii h unlikely thai prey-switchingby predators would affect simultaneously entire popula-tions of birds spread across the peninsula. We re-exam-ined farther data from different parts of the peninsulato test this. Finally, we discuss the changes in thebiomass of the potential prey as the lemming popula-tion changes, and the role of snowy owls Nyetea scundi-aca in the prey-swilching hypothesis, which is alsoknown as the alternative prey hypothesis (Hagen 1952.Lack 1954).
Material and methods
The breeding productivity of the dark-bellied brentgeese was based on estimates of the percentage offirst-year birds in samples of Hocks during winter inBritain where a large proportion of the populationspends the winter. The data were collated by TheWildfowl and Wetlands Trust and Wetlands Interna-tional (original references in Summers 1986). Thebreeding productivity of curlew sandpipers was basedon the percentage of first-year birds in trapped samplesin South Africa where they over-winler (Cramp and
Simmons 1983). Breeding productivity of geese andsandpipers has been either poor (<\JVu firsl-yearbirds) or good ( > 20'/*. first-year birds) (Summers andUnderhill 1987). There were no values between 13 and
The qualitative data on the abundance of lemmings,as used in our earlier paper (Summers and Underhill1987). were obtained from Dorogov (1983) andTomkovich (unpub!.). Dorogov's study was wide-rang-ing, encompassing the valleys of the rivers Agapa, Pura.Bystraya. Pyasina, Yangoda, Taimyr. BoFshaia Balak-nia, Boganida and Khatanga (Fig. 1). Additional dataarc now available from Krylov (1983) who collateddata on lemming numbers from the arctic fox 'Catch'Service (Dept of Biology of Commercially ImportantAnimals). Krylov (1983) did not specify the regions oflhe Taimyr Peninsula where the lemming data werecollected, but most of the game biologists from whomhe received data worked in the typical tundra zone ofthe Taimyr Peninsula (Fig, I). Krylov (1983) also pro-vided the data on arctic foxes: an estimate of their totalnumbers and the frequency of occupation of lairs onthe Taimyr Peninsula in summer.
Neither Dorogov (1983) nor K17I0V (1983) obtaineddata on lemmings from the arctic tundra zone of theTaimyr Peninsula where brent geese and curlew sand-pipers largely breed (Fig. I). Sueh dala were obtainedbetween 1989 and 1993 when expeditions were mountedby the Inst, of Evolutionary Morphology and AnimalEcology (Prys-Jones 1991). Qualitative data were ob-tained from the lower Taimyr River. Sterligova Point.Pyasina Delta and Basin and Sibirakov Island, whilstlemming densities were estimated at Lake Pron-chishcheva (Yemelenova and kybkin, in Underhill etal. 1993) (Fig. 1).
We constructed a contingency table with each yearclassified on three dimensions: good or poor breedingproductivity of brent geese, increase or decrease innumber of foxes from the previous year, and increaseor decrease in abundance of lemmings. In order to testwhether the interactions between these variables weresignificant, we fitted a log-linear model to the three-waycontingency table (Bishop et al. 1975). Because of the
ARCTIC TUNDRA
TYPICAL TUNDRA
SHRUB TUNDRA
Lakeronchishcfieva
720 N
Fig. I. The Taimyr Peninsulashowing localities where lhelemming numbers wereestimated. The dashed linesshow the boundaries of thearetie. typical and shrubtundra zones (after Chernov1985).
574 F.CXKJRAPIIY 21:6 |1998)
likelihood that successive observations in the quantita-tive variables (percentages of occupied fox Uiirs, num-bers of arctic tbxes. and percentages of first-year brentgeese and lirst-year eurlew sandpipers) are intcrcorre-lated, they cannot be treated as independent observa-tions, and it is possible that significant crosscorrelations between these time series are spurious (Boxand Jenkins 1976). We therefore fitted an autoregressivemodel to each series (or a logarithmic transformationof the series) in order to determine the structure of theseries using the computer programme TSP (Hall andLilien 1986), This structure (the component of the timeseries) was subtracted from the time series to obtain theso-called "pre-whitened" residuals. These white noiseresiduals (tested as such by the portmanteau test; Boxand Pierce 1970) then represent the shocks to the timeseries: i.e, that part that cannot be predicted from theprevious observations. If the residuals of two time serieshave shocks that tend to coincide, or coincide if one ofthe series is lagged by one or more years, then theexistence of a relationship between the two time seriesmay be inferred with more certainty than if only crosscorrelations between the original series were large.
Results
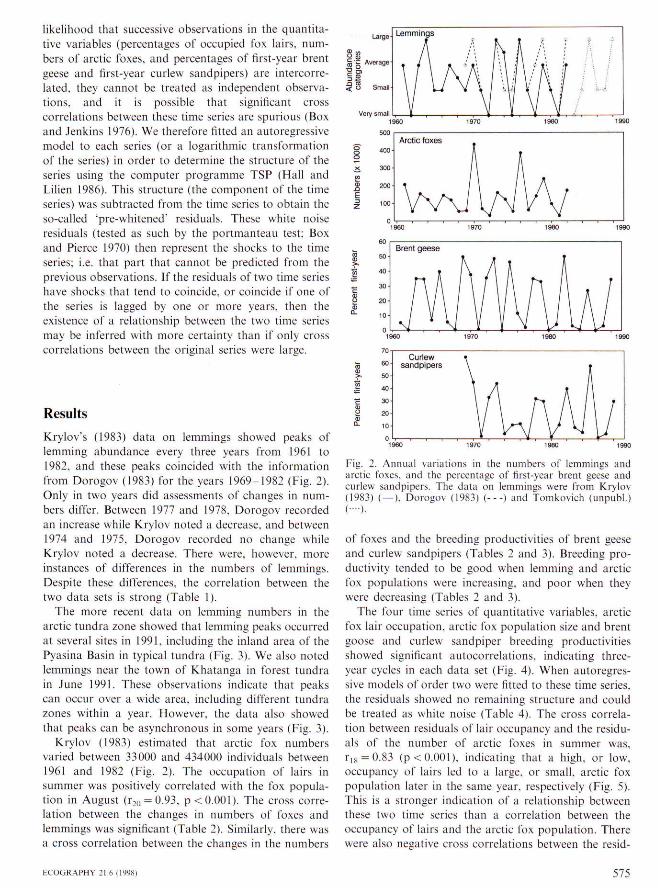
Krylov's (1983) data on lemmings showed peaks oflemming abundance every three years from 1961 to1982. and these peaks coincided with the intbrmationfrom Dorogov (1983) for the years 1969 1982 (Fig. 2).Only in two years did assessments of changes in num-bers differ. Between 1977 and 1978, Dorogov recordedan increase while Krylov noted a decrease, and between1974 and 1975. Dorogov recorded no change whileKrylov noted a decrease. There were, however, moreinstances of differenees in the numbers of lemmings.Despite these differenees. the correlation between thetwo data sets is strong (Table I).
The more recent data on lemming numbers in thearctic tundra zone showed that lemming peaks oeeurredat several sites in 1991. including the inland area of thePyasina Basin in typical tundra (Fig, 3), We also notedlemmings near the town of Khatanga in forest tundrain .Tune 1991. These observations indicate that peakscan occur over a wide area, including different tundrazones within a year. However, the data also showedthat peaks can be asynchronous in some years (Fig. 3).
Krylov (1983) estimated that arctic fox numbersvaried between 33000 and 434000 individuals between1961 and 1982 (Fig. 2). The occupation of lairs insummer was positively correlated with the fox popula-tion in August (1-20 = 0.93, p<O.OOI). The cross corre-lation between the changes in numbers of foxes andlemmings was significant {Table 2). Similarly, there wasa cross eorrelation between the changes in the numbers
Fig, 2. Annual variations in the numbers of lemmings andarctic foxes, and the percentage of first-year brent geese andcurlew sandpipers. The data on lemmings were from Krylov(1983) ( —), Dorogov (1983) (---) and Tomkovich (unpubl.)( • • • • ) ,
of foxes and the breeding productivities of brent geeseand curlew sandpipers (Tables 2 and 3). Breeding pro-ductivity tended to be good when lemming and arcticfox populations were increasing, and poor when theywere decreasing (Tables 2 and 3),
The four time series of quantitative variables, arcticfox lair occupation, arctic fox population size and brentgoose and curlew sandpiper breeding productivitiesshowed significant autocorrelations, indicating three-year cycles in each data set (Fig, 4), When autoregres-sive models of order two were fitted to these time series,the residuals showed no remaining strueture and couldbe treated as white noise (Table 4), The cross correla-tion between residuals of lair occupancy and the residu-als of the number of arctic foxes in summer was,rif< = 0.83 (p< 0.001), indicating that a high, or low.occupancy of lairs led to a large, or small, arctic foxpopulation later in the same year, respectively (Fig, 5).This is a stronger indieation of a relationship betweenthese two time series than a correlation between theoccupancy of lairs and the arctic fox population. Therewere also negative cross correlations between the resid-
ECOGRAPHY 21:6 (I9<)S) 575
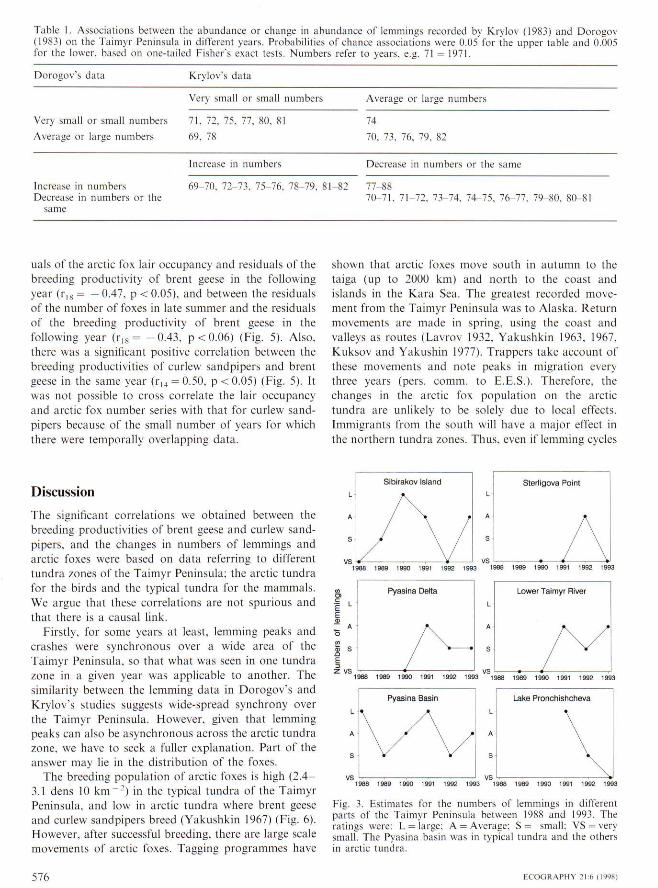
Table 1, Associations between the abundance or change in abundance of lemmings recorded by Krylov (1983) and Dorogov(1983) on the Taimyr Peninsula in different years. Probabilities of chance associations were 0,05 for the upper table and 0,005for the lower, based on one-tailed Fisher's exact tests. Numbers refer to years, e.g, 71 = 1971,
Dorogov's dala Krylov's data
Very small or small numbers Average or large numbers
Very small or small numbers 71, 72, 75. 77, 80.
Average or large numbers 69. 78
74
70. 73, 76, 79. 82
Increase in numbers Decrease in numbers or the same
increase in numbersDecrease in numbers or the
same
3-70, 72-73, 75-76, 78-79, 81-82 77-8870-71. 71-72, 7.V74, 74-75. 76-77. 79-80, 80-81
uals of the arctic fox lair occupancy and residuals of thebreeding productivity of brent geese in the followingyear (ri^ = — 0.47, p < 0,05). and between the residualsof the number of foxes in late summer and the residualsof the breeding productivity of brent geese in thefollowing year ( r |g^ - 0 . 4 3 , p < 0.06) (Fig, 5). Also,there was a significant positive correlation between thebreeding productivities of curlew sandpipers and brentgeese in the same year (ri4 = 0.50. p < 0.05) (Fig. 5). Itwas not possible to cross correlate the lair occupancyand arctic fox number series with that for curlew sand-pipers because of the small number of years for whiehthere were temporally overlapping data.
shown that arctic foxes move south in autumn to thetaiga (up to 2000 km) and north to the coast andislands in the Kara Sea. The greatest recorded move-ment from the Taimyr Peninsula was to Alaska. Returnmovements are made in spring, using the coast andvalleys as routes (Lavrov 1932, Yakushkin 1963, 1967,Kuksov and Yakushin 1977), Trappers take aeeount ofthese movements and note peaks in migration everythree years (pers. comm. to E.E.S.). Therefore, thechanges in the aretie fox population on the arctictundra are unlikely to be solely due to local effects.Immigrants from the south will have a major effect inthe northern tundra zones. Thus, even if lemming cycles
Discussion
The significant correlations we obtained between thebreeding productivities of brent geese and curlew sand-pipers, and the changes in numbers of lemmings andarctic foxes were based on data referring to differenttundra zones of the Taimyr Peninsula; the arctic tundrafor the birds and the typieal tundra for the mammals.We argue that these correlations are not spurious andthat there is a causal link.
Firstly, for some years at least, lemming peaks andcrashes were synchronous over a wide area of theTaimyr Peninsula, so that what was seen in one tundrazone in a given year was applicable to another. Thesimilarity between the lemming data in Dorogov's andKrylov's studies suggests wide-spread synchrony overthe Taimyr Peninsula, However, given that lemmingpeaks can also be asynchronous across the arctic tundrazone, we have to seek a fuller explanation. Part of theanswer may lie in the distribution of the foxes.
The breeding population of arctic foxes is high (2.4-3.1 dens 10 km " -) in the typieal tundra of the TaimyrPeninsula, and low in arctic tundra where brent geeseand curlew sandpipers breed (Yakushkin 1967) (Fig. 6).However, after successful breeding, there are large scalemovements of arctic foxes. Tagging programmes have
A
s
vs
Sibirakov Island
1989 1990 1991 1992 1993 1988 1989 1990 1991 1992 1993
19S9 1990 1991 (992 1993 19B8 19a9 1990 1991 1992 1993
Fig. 3, Estimates for the numbers of lemmings in differentparts of the Taimyr Peninsula between 1988 and 1993. Theratings were; L = large; A = Average: S = small: VS = verysmall. The Pyasina basin was in typical tundra and the othersin arctic tundra.
576 ECOGRAPHY 21:6

Table 2, Associations between the breeding success of brent geese and changes in the abundance of lemmings and arctic foxesin dilTcrent years. Numbers refer to years, e.g. 71 = 1971, There were significaiil interaclions between foxes and geese xf = 9.3,p<().01) and between foxes and lemmings x] = 5.6. p<0,02).
Change in arctic foxpopulalion
Change in lemming populalion
Good breeding for brent geese
Deciease
Increase
Decrease or same Increase
71-72, 74-75
65-66, 68-69, 77-78
63-64
62-63, 69-70. 72-73. 78-79.81-82
Poor breedina for brent ceese
DecreaseIncrease
Decrease or same Increase
61-62. 64-65. 67 68 70 7i. 73-74. 76-77. 79-80. KO-81 66 6775-76
Table .1 Associations between the breeding productivity of curlew sandpipers and the changes in the size of the arctic foxpopulalion in different years. The probability of chance association was 0.025. based on one-lLiilcd Fisher's exact test. Numbersrefer to year. e.g. 71 = 1971.
Change in
Decrease
Increase
The abundance of arctic foxes Breeding productivity
Poor
70-71, 73-74. 74-75,
75-76
of curlew
7(^77, 79
sandpipers
80. 80-81
Good
71-72
69-70, 72-73. 77-78 78 79, 81-82
are asynchronous in differeni areas or different tundrazones, the impact of arctic fox predation may be fehacross all tundra zones because mosl arctic foxes breedin one zone and emigration occurs atter successfulbreeding.
There have now been direct observations in the arctictundra supporting the Roselaar and Summers hypothe-sis (Underhill et al, 1993). In 1991 at Lake Pron-ehishcheva (Fig. I), the lemming population was at apeak. All the geese nested successfully and the dailyprobability of nest predalion for waders was 0.0022. In1992. when the lemming population was declining, onlytwo geese nested but the wader density was similar.However, large numbers ol" foxes were present and thedaily probability of nest predation for waders was 100times greater (0.20). The main differences in the foxes'behaviour in the two years were that, in 1991, they wererarely seen and only close to their dens, whereas in1992, they were regularly seen searching for food and13 dead foxes were found (Underhill et al, 1993).
The prey-switching hypothesis assumes that theehange in the diet of foxes is due to the lemming foodbase being very much greater than that of the birdswhen lemmings are at their peak, and lower relative tothe birds when the lemming population is low. Therehave been few estimates of lemming densities on theTaimyr Peninsula (reviewed by Batzli 1981), One studyreported densities of < 1 ha • ' in 1968, 130 Di-crostony.x torqualtis and 84 Lemimis sihiricus ha " ' in1969. and a peak of 725 ha ' (ca 54 kg h a " ' ) (both
species combined) in 1970 (Vinokurov et al. 1972).More recently, in 1991. Yelemenova and Rybkin (inUnderhill et al, 1993) estimated 100-392 lemmingsha" ' at Lake Pronchisheheva (7-29 kg ha" '). Countsof birds at the same time gave density estimates of 7,1
Fox numbers
Lag (years) Lag (years)
Fig. 4. Correlograms showing autocorrelations of ihe percentoccupancy of arctic fox lairs, arctic fox numbers, and thepercentage of first-year brent geese and curlew sandpipers.Peaks and troughs above and below the doited lines indicatelags that are statistically signitieant.
ECOGRAPHV 21:6 577
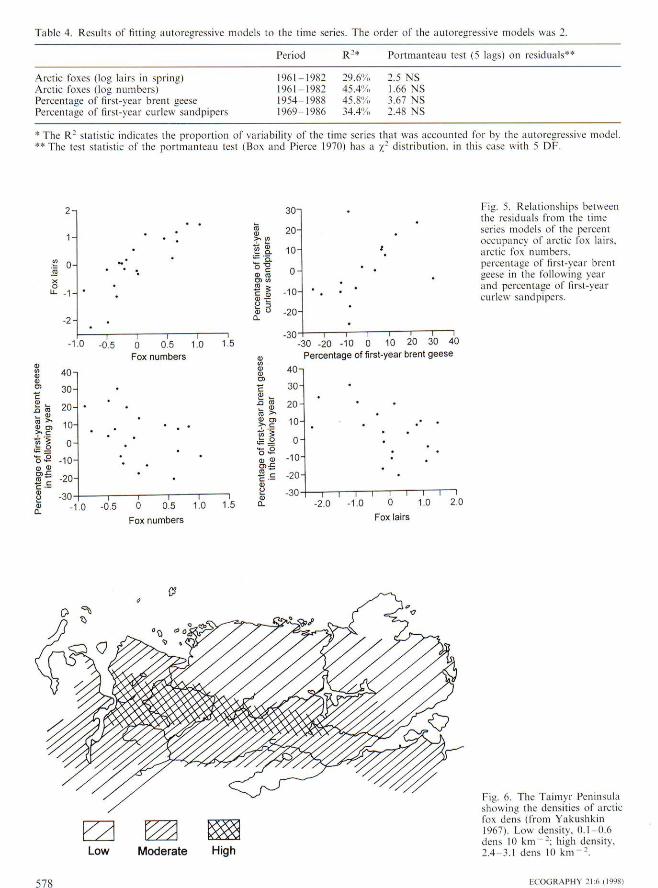
Table 4, Results of Htling aiiioregressivc models to the time series. The order of the autoregressive models was 2,
Period R-* Portmanteau test (5 lags) on
Arctic foxes (log lairs in spring)Arctic foxes (log numbtrrs)Percentage of first-year brerU geesePercentage of first-year curlew sandpipers
1961-1982 29,6% 2,5 NS1961 1982 45,4'>1. 1,66 NS1954-1988 45,8'!';, 3,67 NS1969 1986 34,47,, 2,48 NS
*The R- slalistic indicates the proportion of variability of the lime series that was accounted for by the autoregressive model.** The test statistic of the portmanleau lest (Box and Pierce 1970) has a / - distribution, in ihis case with 5 OF.
1-
0-
- 1 -
-2-
-1,0
40-1
30-
20-
10
0-
-10-
-20-
-30-
-0,5 0 0,5 1.0Fox numbers
1,5
-1,0 -0,5 0 0.5
Fox numbers
1.0 1.5
l i
20-
10-
0 -
-10-
-20-
-p to - 1 1
•
—• 1 1 1-30 -20 -10 0 10 20 30 40
Percentage of first-year brent geese
40-
30-
•° S 20-
10-
o 0--10-
~,£ -20-
-30
Fig, 5, Relationships betweenthe residuals from the timeseries models of the percentoccupancy of arctic fox lairs,arctic fox numbers,percentage of (irsl-year brentgeese in the following yearand percentage oT lirst-yearcurlew sandpipers.
-2.0 -1,0 0
Fox lairs
1 0 2.0
Low Moderate High
Fig, 6, The Taimyr Peninsulashowing the densities of arcticfox dens (from Yakushkin1967), Low density. 0,1 0,6dens 10 km -; hiuh density.2.4 .1,1 dens 10 km" ^
578 LCOGR.APHY 21

pairs of waders. 2,4 gulls and skuas, 1.4 passerines, ca0.7 ptarmigan Lagopus mutus and 0.6 pairs of brenlgeese km - (Underbill el al. 1993). Tbis was equivalentto ca 0,056 kg of birds ba~ ' and 0,011 k g e g g s b a " ' .During 1992, tbe lemming population declined from 64to ca 0 ba ~ ' during tbe summer. Tbe food supply o\'birds and eggs was similar to 1991, Tbus, tbe lemmingfood source during peaks is very mucb greater than tbatof birds, and less tban the birds wben tbe lemmings arevirtually absent.
An additional factor, wbicb affects tbe vulnerabilityof geese to predation, is important to tbe prey-switcb-ing bypotbesis. Brent geese at Lake Pronebisbcbevawere found nesting in association wilb breeding snowyowls (Summers et al, 1994), similar to the bebaviour ofblack brants B. b. nigricans on Wrangel Island (Litviiiet al, 1985, Dorogoi 1990), Tbe aggressive owls excludearciic foxes from around tbeir nests and tbus provide asafe nesting area for geese, Tbe black brants arc sodependent on tbe snowy owls tbat tbey do not attemptnesting if tbe owls do not breed (Dorogoi 1990). Giventhat snowy owls breed mainly when tbe lemming foodsupply is good (Litvin and Ovsyanikov 1990). tbe blackbrant's breeding productiviiy is tberefore indirectlylinked to lemming numbers. At present, it is unknov '̂nwbat proportion of dark-bellied brent geese nest inassociation witb snowy owls but it bas been observedelsewbcre on tbe Taimyr Peninsula, at BoFsbaia Balak-nia (Yesoi! 1994), at Medusa Bay. western Taimyr{Tulp et al. 1997) and on tbe Mamonta Peninsula westof tbe Taimyr Peninsula (Zbukov and Ivanov. inRyabitsev 1993).
Acknowledgements - We thank alt participants of the Interna-tional Arctic Expedition who kindly supplied information onlemmings and foxes, LGU is snpported by the Univ, of CapeTown and Ihe Foundation for Research and Development.Pretoria. The draft was commented upon by C, J, Feare,
ReferencesBatzli, G, O, 1981, Populalions and energetics of small mam-
mals in the tundra ecosystem. - In: Bliss. L, C . Heal. O,W, and Moore, .1, .1, (eds). Tundra ecosystems: a compara-tive analysis. Cambridge Univ, Press, pp, 377-396,
Bishop. Y, M,, Fienber. S. E, and Holland. P. W, 1975,Discrete mnltivariate analysis: theory and practice, MITPress, Cambridge. Massachusetts,
Box. G, E, P. and Pierce. D, A. 1970, Distribution of residualantocorrelations in a ut ore gressive-integrated moving aver-age time series models. - J, Am, Stat. Ass. 65: 1509 1526,
Box. G. E. P. and Jenkins. D, K. 1976, Time series analysis:forecasting and control, revised edition, - Holden Bay.San Francisco,
Chernov. Y. 1, 1985, The living tundra, Cambridge Univ,Press.
Cramp, S, and Simmons. K, E, L, (edsl 1983, The birds of thewestern Palearetic. vol, .3. - Oxford Univ, Press,
Dorogoi. I, V, 1990, Factors of communal breeding of thesnowy owls {Nyvieu scandiaca) and anseriformes birds atthe Vrangel Island, - Ornithologia 24: 26-33,
Dorogov, V. F. 1983. Nesting of the rough-legged buzzard(Buieo lagopus) on Taimyr, - Ecology and rational use ofterrestrial vertebrates of north of central Siberia. Novosi-birsk, pp. 118-126.
Hagcn. Y, 1952, Rovfuglene og viltpleien. - Gyldendal Norskforlag. Oslo,
Hall. R,^E, and Lilien, D, M. 1986, Micro TSP users manual,- McGraw-Hill. New York,
Krylov. M, K, 1983, The elTects of ecological faetors on thenumbers and hunting of arctic foxes in the Taimyr.-Ecology and rational use of terrestrial vertebrates of thenorth of central Siberia. Novosibirsk, pp, 86 93,
Kuksov, V. A, and Yakushin G. D, 1977, Arctic fox. Fauna ofgame mammals and birds. Game management of theYenisey North. Krasnoyiarsk publ. house, pp. 29-37.
Lack. D, 1954, Hie natural regulation of animal numbers, -Clarendon. Oxford,
Lavrov, P, N, 1932, Arctic fox, - Vheshtoruizdat. Moscow,Eitvin, K. E,. Pulyaev. A. I. and Syroechkovsky. E. V, 1985,
Colonies of the snow goose (Anser caerulescens). brentgoose (Branlii hcruicia) and eider (Somateria moilissimu)near the snowy owl (Nyclea seundiacu) nests on theWrangel Island,'- Zool, Zhurnal 64: 1012 1023.
Eitvin, K, Y, and Ovsyanikov. N. G, 1990, Relationshipbetween the reproduction and numbers of snowy owls andarciic foxes and the nnmbers of true lemmings on theVrangei Island. - Zool, Zhurnal 69: 52-64,
Prokosch. P, 19K8, Das Schleswig-Holsteinische Waticnmcerals Kriinhjahrs-Aufenthaltsgebiet arktischer Waivogel -Populationen am Beispiel von Kicbitzregenpfiefer {Piuvialissquaiarola. L, 1758). Knutt (Calhiis caiunus. L, 1758) undPfuhlsehnepfe (Uiiiosa iapponica. L. 1758), - Corax 12:273 442,
Prv's-Jones. R, P, 1991, Ornithological cooperation in Siberia,- Nature 354: 102.
Roselaar, C. S, 1979, Fluetuaties in aantallen krombekstrand-lopers Calidri.s ferritginea. - Watcrvogels 4: 202 210.
Ryahitsev, V. K, (ed,) (translated by jT Gromadzka) 1993.Breeding conditions for waders in the tundras of Russia in1991, - Wader Study Group Bull, 71: 20 25,
Summers. R. W, 1986, Breeding production of dark-belliedbrent geese Brania hernicUi hcruicia in relation to lemminaeycles. Bird Study 33: 105-108,
Summers. R, W, and Underhili. L, G, 1987, Factors related EObreeding production of brent geese Branta /;, in-riUcIa andwaders (Charadrii) on the Taimyr Peninsula, - Bird Study34: 161-171,
Summers. R, W, et al, 1989, Populations, migrations, biomet-rics and moult of the turnstone Arenaria i. inlerpre.s on theeast Atlantie coastline, with speeial reference to theSiberian population. - Ardea 77: 145 -168,
Summers. R, W. et al, 1994, The breeding biology of dark-bel-lied brent geese Brania b. heriiicia and king eiders Somale-ria speclabilis on the northeastern Taimyr Peninsula,especially in relation to snowy owl Nyciea .scandiaea nests,- Wildfowl 45: 110-118,
Tulp. I, et al, 1997, Breeding waders at Medusa Bay, westernTaimyr, m 1996. WtWO report 57. Zeist,
Underhili. L, G, 1987, Changes in the age structure of curlew-sandpiper populations at Eangebaan Eagoon, SouthAfrica, in relation to lemming cycles in Siberia, - Trans,Roy, Soc, South Africa 46: 209 214,
Underhili. L, G,. Waltner. M, and Summers. R, W, 1989.Three-year cycles in breeding productivity of knots Calidri.scanuius wintering in southern Africa suggest Taimyr Penin-sula provenance. Bird Study 36: 83-87,
Underhili. E, G, et al, 1993, Breeding of waders (Charadrii)and brent geese Brania herniela fjernicia at PronchishehevaEake, northeastern Taimyr, Russia, in a peak and a de-creasing lemming year. - Ibis 135: 277-292,
Vinokurov. A, A., Orlov. V, A. and Okhotsky. Y, V, 1972,Popniation and faunal dynamics of vertebrates in tundrabiocenoses (Taimyr), In: Wielgolaski. F, E. and Ross-wall. T, (eds), Proc, IV Int, Meet, Biol, Prod, Tundra.
ErOGR,'\PHY 21:6 579

Leningrad. USSR. Tundra Biome Steering Comm.. Stock- Yakushkin. G. D. 1967, Distribution of the arctic fox dens inholm, pp, 187-189, Taimyr, Problems of agriculture and game managetnenT of
Yakushkin. G. D. 1963, The preliminary results of marking of the Far North. V, 14. - Krasnoiask Publ, House, pp.arctic foxes in Taimyr with the aim of studies of seasonal 23- 32,migrations. - Proc. Inst. Agrieult, Far North. Norilsk, pp. Yesou. P. 1994, Contribution a I'etude avifaunistique dc la175-ISO, Peninsule du Taimyr. - Alauda 62: 247-252.
5g{) ECOGRAPHY


![Predatorprey relationships] arctic foxes and lemmings · 2010-04-23 · Predatorprey relationships] arctic foxes and lemmings ANDERS ANGERBJO RN˜\ MAGNUS TANNERFELDT˜ and SAM ERLINGE](https://img.dokumen.tips/doc/110x75/5ed3db9673d3d2457570060c/predatorprey-relationships-arctic-foxes-and-2010-04-23-predatorprey-relationships.jpg)



























