Embed Size (px)
Citation preview

TMRJ
BsiMfudwR2tCm
Kt
Se(1Msts2cr1hhLimc(mrbir
F
A
R
0d
he Anxiogenic Drug Yohimbine Reinstatesethamphetamine Seeking in a Rat Model of Drug
elapseack D. Shepard, Jennifer M. Bossert, Shirley Y. Liu, and Yavin Shaham
ackground: Brain noradrenaline is involved in footshock stress-induced reinstatement of drug seeking in a rat relapse model. Wetudied whether yohimbine, an �-2 adrenoceptor antagonist that increases noradrenaline release and induces anxiety-like responsesn human and nonhuman subjects, would reinstate methamphetamine seeking in rats.
ethods: In experiment 1, the effect of yohimbine (1.25–2.5 mg/kg) on reinstatement was compared with that of intermittentootshock (5 min; .2–.6 mA) in rats that were trained to lever press for intravenous methamphetamine (9–11 days) and subsequentlynderwent 7 days of extinction training. In experiment 2, the effect of yohimbine on reinstatement of drug seeking was determineduring early (1 day) and late (21 or 51 days) withdrawal periods. On the test days, rats were first given 3-hour extinction sessions andere then tested for reinstatement induced by yohimbine.esults: In experiment 1, both yohimbine and footshock stress reinstated methamphetamine seeking after extinction. In experiment, extinction responding was higher after 21 or 51 withdrawal days than after 1 withdrawal day. In contrast, no significantime-dependent changes in yohimbine-induced reinstatement were observed.onclusions: Results indicate that yohimbine is a potent stimulus for reinstatement of methamphetamine seeking in a rat relapseodel.
ey Words: �-2 adrenoceptors, drug self-administration, extinc-ion, incubation, reinstatement, relapse, stress
tress can provoke drug craving and relapse in humans, butthe mechanisms underlying these effects are not known(Jacobsen et al 2001; Sinha 2001). In laboratory rats,
xposure to intermittent footshock stress reinstates heroinAhmed et al 2000; Shaham and Stewart 1995), cocaine (Erb et al996; Mantsch and Goeders 1999), alcohol (Le et al 1998;artin-Fardon et al 2000), and nicotine (Buczek et al 1999)
eeking. Studies using neuropharmacologic methods indicatehat brain noradrenaline (NA) is a critical mediator of footshocktress-induced reinstatement of drug seeking (Shaham et al000a). Systemic injections of the �-2 adrenoceptor agonists,lonidine and lofexidine, which decrease NA cell firing andelease (Aghajanian and VanderMaelen 1982; Mongeau et al997), attenuate footshock-induced reinstatement of heroin (Sha-am et al 2000b), cocaine (Erb et al 2000), and speedball (aeroin-cocaine combination) (Highfield et al 2001) seeking.esions of the ventral noradrenergic bundle attenuate footshock-nduced reinstatement of heroin (Shaham et al 2000b) andorphine (Wang et al 2001) seeking, and infusions of �-adreno-
eptor antagonists into the bed nucleus of the stria terminalisBNST) or the amygdala attenuate footshock-induced reinstate-ent of cocaine seeking (Leri et al 2002). Finally, Lee et al (2004)
ecently studied the effect of antagonism of the �-2 adrenoceptory yohimbine and RS-79948 on reinstatement of cocaine seekingn squirrel monkeys and reported that these drugs provokeesumption of lever pressing after extinction.
rom the Behavioral Neuroscience (JMB, SYL, YS) and Cellular NeurobiologyBranches (JDS), Intramural Research Program, National Institute on DrugAbuse, National Institutes of Health/Department of Health and HumanServices, Baltimore, Maryland.
ddress reprint requests to Dr. Yavin Shaham, Behavioral NeuroscienceBranch, IRP/NIDA/NIH, 5500 Nathan Shock Drive, Baltimore, MD 21224.
eceived December 31, 2003; revised February 25, 2004; accepted February 29,2004.
006-3223/04/$30.00oi:10.1016/j.biopsych.2004.02.032
Based on these previous findings, we studied the effect ofyohimbine on reinstatement of methamphetamine seeking inrats. Methamphetamine is abused by humans (Rawson et al 2002)and self-administered by laboratory animals (Yokel and Pickens1973), but the effect of stress on reinstatement of methamphet-amine seeking has not been investigated. Yohimbine increasesNA cell firing (Aghajanian and VanderMaelen 1982) and inducesNA release in several terminal areas, including the hypothalamus(Pacak et al 1992), hippocampus (Abercrombie et al 1988),prefrontal cortex (Becker et al 1999), BNST (Forray et al 1997),and amygdala (Khoshbouei et al 2002). Yohimbine also inducesanxiety-like responses in both human (Charney et al 1983;Holmberg and Gershon 1961) and nonhuman (Davis et al 1979;Lang and Gershon 1963) subjects. McDougle et al (1994) re-ported that in abstinent cocaine users, yohimbine induces anxi-ety-like symptoms that are more pronounced during early with-drawal. Stine et al (2001, 2002) found that in methadone-maintained patients, yohimbine induces anxiety- and opioidwithdrawal-like symptoms, as well as drug craving. These au-thors also reported that yohimbine-induced increases in cortisolsecretion, systolic blood pressure, and startle response are higherin methadone-maintained patients than in control subjects.
In this study, we first compared the effect of yohimbine onreinstatement of methamphetamine seeking to that of a standardintermittent footshock stressor, which was previously found toreinstate responding maintained by other drugs (Lu et al 2003).For this purpose, we used a “between-session” reinstatementprocedure in which self-administration training, extinction train-ing, and subsequent tests for reinstatement (under extinctionconditions) are conducted on different days (Shaham et al 2003).Based on recent demonstrations of time-dependent changes afterwithdrawal in reinstatement induced by footshock stress (Shalevet al 2001), drug priming (De Vries et al 2002; Tran-Nguyen et al1998), and drug cues (Grimm et al 2001), we also studied theimpact of the methamphetamine withdrawal period on yohim-bine-induced reinstatement. For this purpose, we used a “be-tween-within” reinstatement procedure in which rats are trainedfor drug self-administration on different days, and after differentwithdrawal periods, subjects are tested on the same day for
BIOL PSYCHIATRY 2004;55:1082–1089© 2004 Society of Biological Psychiatry

rsT
M
S
RR(wwL
I
(wis2m2afir
A
calirMpto(
Eo
tfdc
thaafMfmpalpfa
J.D. Shepard et al BIOL PSYCHIATRY 2004;55:1082–1089 1083
esistance to extinction and subsequent reinstatement of drugeeking induced by drug or nondrug stimuli (Shaham et al 2003;ran-Nguyen et al 1998).
ethods and Materials
ubjectsThe subjects were 73 male Long-Evans rats (Charles River,
aleigh, North Carolina) that weighed 350–400 g before surgery.ats were maintained on a reverse 12-hour light-dark cyclelights off at 9 or 10AM). Before and one week after surgery, ratsere kept in the animal facility with free access to food andater. Experimental procedures followed the “Principles ofaboratory Animal Care” (NIH publication no. 86–23, 1996).
ntravenous SurgeryRats were anesthetized with a xylazine � ketamine mixture
10 � 100 mg/kg, intraperitoneal [IP]) and surgically implantedith intravenous Silastic catheters (Dow Corning, Midland, Mich-
gan) into the right jugular vein. The catheter was passedubcutaneously to the top of the skull, attached to a modified2-gauge cannula (Plastics One, Roanoke, Virginia), andounted to the rat’s skull with dental cement (see Shalev et al000). Buprenorphine (.1 mg/kg, subcutaneous) was given as annalgesic after surgery, and rats were given 5–7 days to recoverrom surgery. Catheters were flushed with sterile saline contain-ng gentamicin (.08 mg/mL) every 24–48 hours during theecovery and training phases.
pparatusThe self-administration chambers, controlled by a Med Asso-
iates (Georgia, Vermont) system, had two levers located 9 cmbove the grid floor, but only one lever (an active, retractableever) activated the infusion pump. Presses on the other lever (annactive, stationary lever) were also recorded. The cannula on theat’s skull was connected to a liquid swivel (Instech, Plymoutheeting, Pennsylvania) with polyethylene-50 tubing that wasrotected by a metal spring and connected to a 20-mL syringe ofhe infusion pump. For the footshock experiment, the grid floorsf the chambers were connected to electric shock generatorsMed Associates).
xperiment 1: Effect of Intermittent Footshock or Yohimbinen Reinstatement
The experiment consisted of three phases: self-administrationraining (9–11 days), extinction training (7 days), and tests forootshock- or yohimbine-induced reinstatement (5–6 days). Theaily sessions started within 1-hour after the onset of the darkycle.
Training Phase. On the day before training, rats were broughto the self-administration room where they were chronicallyoused in the experimental chambers with free access to foodnd water. Rats were trained to self-administer methamphet-mine-HCl (supplied by the National Institute on Drug Abuse)or 6 hours/day for 9–11 days over a period of 12–14 days.ethamphetamine was dissolved in sterile saline and was in-
used at a volume of .065 mL over 2.3 sec at a dose of .1g/kg/infusion. The dose of methamphetamine is based onrevious studies (Harrod et al 2001; Stefanski et al 2002; Yokelnd Pickens 1973). During the sessions, responding on the activeever resulted in methamphetamine infusions that were accom-anied by a 20-sec light cue that was located above the lever. Aixed-ratio-1 schedule was used, with a 20-sec timeout periodfter each drug infusion. Responses on the inactive lever were
recorded but had no programmed consequences. Each sessionbegan with the insertion of the active lever and the illuminationof a houselight that remained on for the entire session. At the endof each session, the houselight was turned off and the activelever retracted. The number of infusions was limited to 100/6hours to minimize overdose. Six rats were excluded due to lossof catheter patency during training, poor health, or failure toacquire methamphetamine self-administration (less then 15 infu-sions/day).
Extinction Phase. Extinction training consisted of daily 6-hoursessions for 7 days. The experimental conditions were the sameas in training, with the exception that the responses on thepreviously active lever were not reinforced with methamphet-amine. Rats tested for yohimbine-induced reinstatement weregiven 4–5 daily injections of distilled water (.5 mL/kg, IP) duringthe extinction phase to habituate them to the injection proce-dure.
Test for Reinstatement, Experiment 1a: Intermittent Foot-shock. After extinction, one group of rats (n � 10) was tested, ina counterbalanced order, for the effects on reinstatement ofdifferent intensities of intermittent footshock (0, .2, .4, and .6mA). During the test days, rats were given five 1-hour extinctionsessions that were separated by 5 min. Rats were exposed to 5min of intermittent shock just before the fourth session. Thereinstatement tests were conducted every 48–72 hours; duringthe intervening days, rats were given five 1-hour extinctionsessions that were separated by 5 min. Constant-current, inter-mittent, inescapable, electric footshock (.5 sec ON, mean OFFperiod of 40 sec, range 10–70 sec) was delivered through ascrambler to the grid floor of the chambers. The footshockparameters are based on previous studies (see Shaham et al2000b).
Test for Reinstatement, Experiment 1b: Yohimbine. After ex-tinction, one group of rats (n � 13) was tested, in a counterbal-anced order, for the effects of different doses of yohimbine (0,1.25, and 2.5, mg/kg, IP) on reinstatement. During the test days,rats were given five 1-hour extinction sessions that were sepa-rated by 30 min. Rats were injected with the vehicle or yohimbine30 min before the fourth session. The reinstatement tests wereconducted every 48–72 hours and during the intervening days,rats were given five 1-hour extinction sessions that were sepa-rated by 30 min. Yohimbine (Research Biochemicals, Natick,Massachusetts) was dissolved in distilled water and was injectedin a volume of .5 mL/kg. The doses of yohimbine are based onprevious reports on the behavioral and physiologic effects ofyohimbine in rats (Schroeder et al 2003; Wada and Fukuda 1991;White and Birkle 2001).
Experiment 2: Time Course of Yohimbine-InducedReinstatement
Experiment 2 consisted of three phases: self-administrationtraining (10 days), withdrawal phase (1, 21, or 51 days), and testsfor yohimbine-induced reinstatement at the different withdrawaldays.
Training Phase. Three groups of rats (final n � 9–11 pergroup) were trained to self-administer methamphetamine underthe experimental conditions described for experiment 1. Thegroups tested at the different withdrawal days were matched fortheir drug intake during training. At the end of training, rats werebrought back to the animal housing facility. One rat was ex-cluded because of poor health.
Withdrawal Phase. During the withdrawal phase, rats wereindividually housed in the animal facility and were not exposed
www.elsevier.com/locate/biopsych

ttt
ttp1twrpjscqbeaay
S
awDssf(eWSYfam(l
R
ER
ndteig�osoir
urfu
1084 BIOL PSYCHIATRY 2004;55:1082–1089 J.D. Shepard et al
w
o methamphetamine or to the self-administration chambers. Ratsested after 21 or 51 days of withdrawal were handled threeimes/week.
Test for Reinstatement. The experimental conditions werehe same as in training with the exception that the responses onhe previously active lever were not reinforced with metham-hetamine. During the test days, rats were first given three-hour extinction sessions that were separated by 30 min. Afterhese extinction sessions, rats were injected IP with distilledater (.5 mL) 30 min before the fourth 1-hour session. If the rats
eached the extinction criterion of � 20 responses on thereviously active lever, then yohimbine (.625 mg/kg) was in-
ected 30 min before the subsequent (and last) 1-hour testession. If, however, the rats did not reach the extinctionriterion, then vehicle was injected between each of the subse-uent 1-hour extinction sessions up to three times before yohim-ine was administered. Because the yohimbine doses used inxperiment 1 (1.25 or 2.5 mg/kg) induced robust reinstatement,lower dose (.625 mg/kg) of the drug was used to maximize ourbility to detect time-dependent differences in responsiveness toohimbine after withdrawal.
tatistical AnalysesData from the reinstatement tests of experiment 1 were
nalyzed with repeated-measures analysis of variance (ANOVA),ith Footshock Intensity (0, .2, .4, and .6 mA) or Yohimbineose (0, 1.25, and 2.5 mg/kg) and Test Session (the two 1-hour
essions after footshock or yohimbine exposure) as the within-ubjects factors. Post hoc analyses of significant effects wereollowed by Fisher protected least significant difference teststwo-tailed). Data from the extinction and reinstatement tests inxperiment 2 were analyzed with a mixed-model ANOVA, withithdrawal Day as the between-subjects factor and Extinction
ession (the three 1-hour sessions before vehicle exposure) orohimbine Dose (0 and .625 mg/kg) as the within-subjects
actors. Data were analyzed separately for responding on thective and inactive levers; however, because the experimentalanipulations had a minimal effect on inactive lever responding
see Figures), we only describe the statistical analyses for activeever responding.
esults
xperiment 1: Effect of Intermittent Shock or Yohimbine oneinstatement
Training and Extinction. Figure 1 shows the mean � SEMumber of presses on the active lever during the last 3 trainingays and during extinction for the two groups of rats that wereested for footshock- or yohimbine-induced reinstatement afterxtinction. The mean � SEM number of methamphetaminenfusions (data collapsed from the footshock and yohimbineroups, total n � 23) during the last three training days was 50.0
3.1, 51.2 � 3.2, 55.0 � 2.9, respectively. Surprisingly, underur experimental conditions of chronic housing and long dailyessions, the rats did not demonstrate the typical extinction burstn the first extinction day (Yokel 1987), and extinction respond-ng remained stable over the 7 extinction days (Figure 1). Theeasons for this unusual extinction curve are not known.
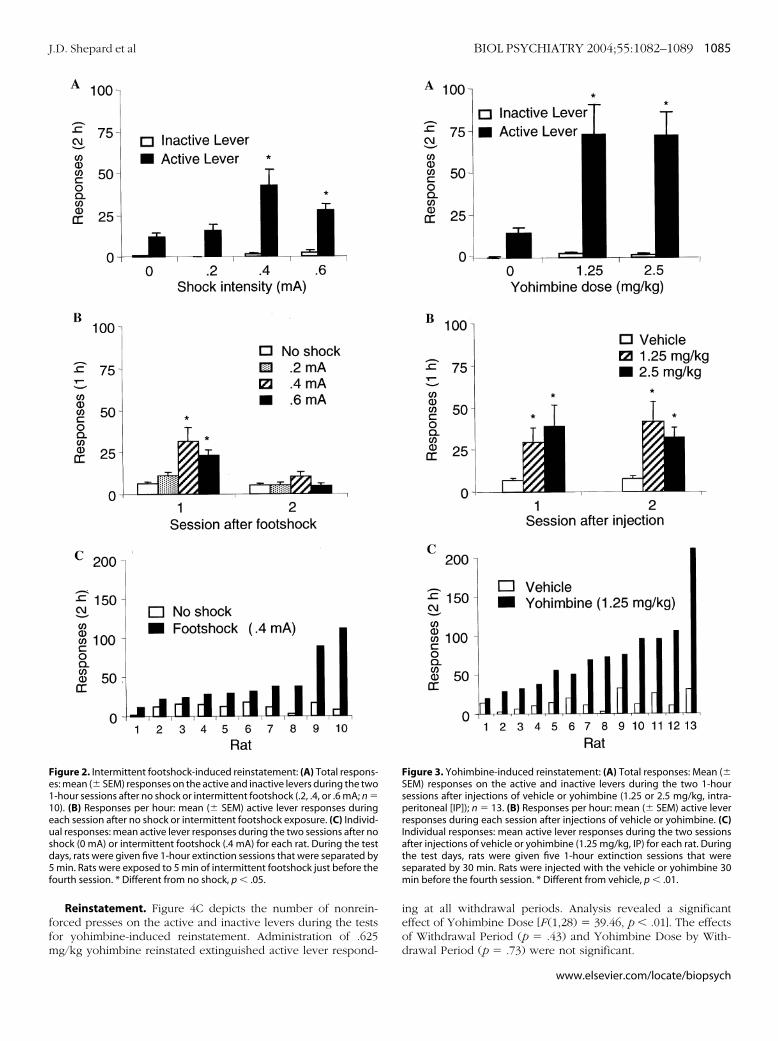
Reinstatement, Experiment 1a: Intermittent Footshock. Fig-re 2A,B depicts the mean � SEM number of nonreinforcedesponses on the active and inactive levers during the tests forootshock-induced reinstatement. Figure 3C depicts the individ-al responses to .4 mA footshock, the most effective shock
ww.elsevier.com/locate/biopsych
intensity. Footshock reinstated extinguished active lever re-sponding in an intensity dependent manner during the first, butnot the second, hour after stress exposure. Analysis of activelever responding revealed significant effects of Footshock Inten-sity [F (3,27) � 7.13, p � .01], Test Session [F (1,27) � 15.84, p �.01], and Footshock Intensity by Test Session [F (3,27) � 6.97, p �.01].
Reinstatement, Experiment 1b: Yohimbine. Figure 3A,B de-picts the mean � SEM number of nonreinforced presses on theactive and inactive levers during the tests for yohimbine-inducedreinstatement. Figure 1C depicts the individual responses to 1.25mg/kg of yohimbine. Both doses of yohimbine inducedreinstatement of extinguished active lever responding, aneffect that was observed at both hours of testing. Analysis ofactive lever responding revealed a significant effect of Yohim-bine Dose [F (2,24) � 9.39, p � .01] but not Test Session[F (1,24) � .19, ns] or Yohimbine Dose by Test Session [F (2,24)� .71, ns].
Experiment 2: Time Course of Yohimbine-InducedReinstatement
Training. Rats demonstrated reliable methamphetamine self-administration during the training phase. The mean � SEMnumber of methamphetamine infusions (n � 31) on the last threetraining days was 48.5 � 3.2, 48.3 � 3.5, 46.9 � 3.3, respectively.
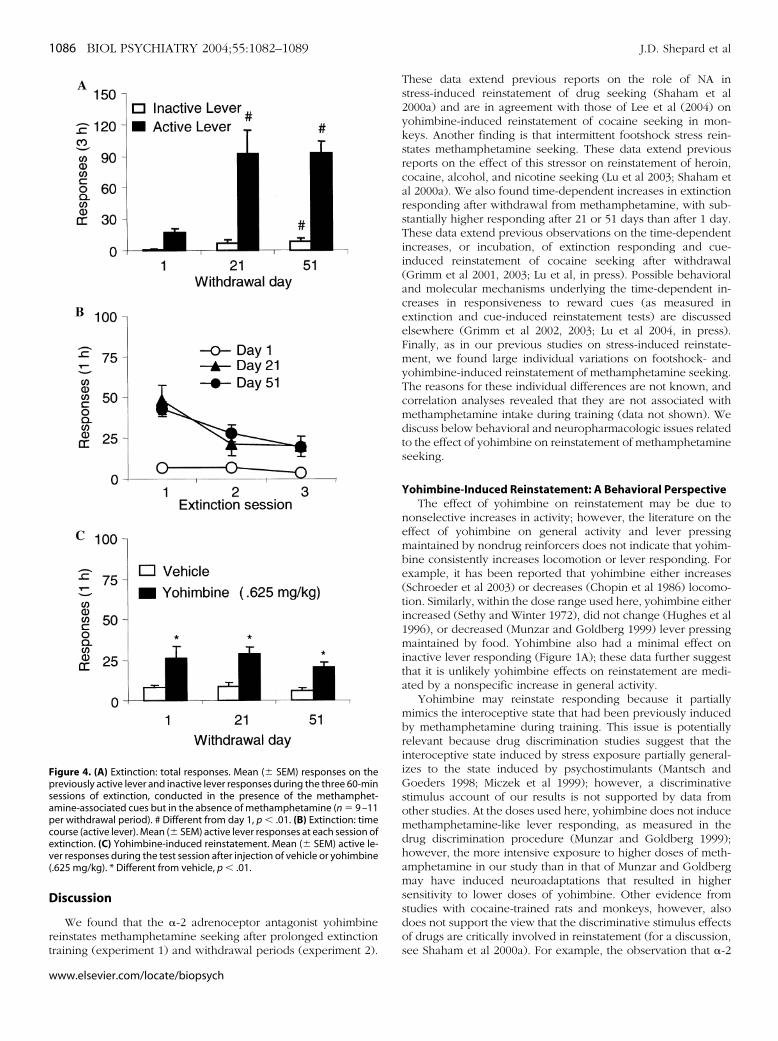
Extinction. Figure 4A,B depicts the number of nonreinforcedlever presses during the extinction tests conducted 1, 21, and 51days after withdrawal from methamphetamine. Extinction re-sponding was substantially higher after 21 and 51 days than after1 day. Analysis of responding on the previously active leverduring extinction revealed significant effects of Withdrawal Day[F (2,28) � 11.12, p � .01], Extinction Session [F (2,56) � 42.33, p� .01], and Extinction Session by Withdrawal Day [F (4,56) �9.96, p � .01].
Figure 1. Mean (� SEM) responses (6 hours/day) on the active and inactivelevers during the last 3 days of training and the 7 days of extinction ofExperiment 1. Data are from two groups of rats (n � 10 –13) that weresubsequently tested for the effect of intermittent footshock (Experiment 1a)and yohimbine (Experiment 1b) on reinstatement.
ffm
Fe11eusd5f
J.D. Shepard et al BIOL PSYCHIATRY 2004;55:1082–1089 1085
Reinstatement. Figure 4C depicts the number of nonrein-orced presses on the active and inactive levers during the testsor yohimbine-induced reinstatement. Administration of .625g/kg yohimbine reinstated extinguished active lever respond-
igure 2. Intermittent footshock-induced reinstatement: (A) Total respons-s: mean (� SEM) responses on the active and inactive levers during the two-hour sessions after no shock or intermittent footshock (.2, .4, or .6 mA; n �0). (B) Responses per hour: mean (� SEM) active lever responses duringach session after no shock or intermittent footshock exposure. (C) Individ-al responses: mean active lever responses during the two sessions after nohock (0 mA) or intermittent footshock (.4 mA) for each rat. During the testays, rats were given five 1-hour extinction sessions that were separated bymin. Rats were exposed to 5 min of intermittent footshock just before the
ourth session. * Different from no shock, p � .05.
ing at all withdrawal periods. Analysis revealed a significanteffect of Yohimbine Dose [F (1,28) � 39.46, p � .01]. The effectsof Withdrawal Period (p � .43) and Yohimbine Dose by With-drawal Period (p � .73) were not significant.
Figure 3. Yohimbine-induced reinstatement: (A) Total responses: Mean (�SEM) responses on the active and inactive levers during the two 1-hoursessions after injections of vehicle or yohimbine (1.25 or 2.5 mg/kg, intra-peritoneal [IP]); n � 13. (B) Responses per hour: mean (� SEM) active leverresponses during each session after injections of vehicle or yohimbine. (C)Individual responses: mean active lever responses during the two sessionsafter injections of vehicle or yohimbine (1.25 mg/kg, IP) for each rat. Duringthe test days, rats were given five 1-hour extinction sessions that wereseparated by 30 min. Rats were injected with the vehicle or yohimbine 30min before the fourth session. * Different from vehicle, p � .01.
www.elsevier.com/locate/biopsych
D
rt
Fpsapcev(
1086 BIOL PSYCHIATRY 2004;55:1082–1089 J.D. Shepard et al
w
iscussion
We found that the �-2 adrenoceptor antagonist yohimbineeinstates methamphetamine seeking after prolonged extinctionraining (experiment 1) and withdrawal periods (experiment 2).
igure 4. (A) Extinction: total responses. Mean (� SEM) responses on thereviously active lever and inactive lever responses during the three 60-minessions of extinction, conducted in the presence of the methamphet-mine-associated cues but in the absence of methamphetamine (n � 9 –11er withdrawal period). # Different from day 1, p � .01. (B) Extinction: timeourse (active lever). Mean (� SEM) active lever responses at each session ofxtinction. (C) Yohimbine-induced reinstatement. Mean (� SEM) active le-er responses during the test session after injection of vehicle or yohimbine.625 mg/kg). * Different from vehicle, p � .01.
ww.elsevier.com/locate/biopsych
These data extend previous reports on the role of NA instress-induced reinstatement of drug seeking (Shaham et al2000a) and are in agreement with those of Lee et al (2004) onyohimbine-induced reinstatement of cocaine seeking in mon-keys. Another finding is that intermittent footshock stress rein-states methamphetamine seeking. These data extend previousreports on the effect of this stressor on reinstatement of heroin,cocaine, alcohol, and nicotine seeking (Lu et al 2003; Shaham etal 2000a). We also found time-dependent increases in extinctionresponding after withdrawal from methamphetamine, with sub-stantially higher responding after 21 or 51 days than after 1 day.These data extend previous observations on the time-dependentincreases, or incubation, of extinction responding and cue-induced reinstatement of cocaine seeking after withdrawal(Grimm et al 2001, 2003; Lu et al, in press). Possible behavioraland molecular mechanisms underlying the time-dependent in-creases in responsiveness to reward cues (as measured inextinction and cue-induced reinstatement tests) are discussedelsewhere (Grimm et al 2002, 2003; Lu et al 2004, in press).Finally, as in our previous studies on stress-induced reinstate-ment, we found large individual variations on footshock- andyohimbine-induced reinstatement of methamphetamine seeking.The reasons for these individual differences are not known, andcorrelation analyses revealed that they are not associated withmethamphetamine intake during training (data not shown). Wediscuss below behavioral and neuropharmacologic issues relatedto the effect of yohimbine on reinstatement of methamphetamineseeking.
Yohimbine-Induced Reinstatement: A Behavioral PerspectiveThe effect of yohimbine on reinstatement may be due to
nonselective increases in activity; however, the literature on theeffect of yohimbine on general activity and lever pressingmaintained by nondrug reinforcers does not indicate that yohim-bine consistently increases locomotion or lever responding. Forexample, it has been reported that yohimbine either increases(Schroeder et al 2003) or decreases (Chopin et al 1986) locomo-tion. Similarly, within the dose range used here, yohimbine eitherincreased (Sethy and Winter 1972), did not change (Hughes et al1996), or decreased (Munzar and Goldberg 1999) lever pressingmaintained by food. Yohimbine also had a minimal effect oninactive lever responding (Figure 1A); these data further suggestthat it is unlikely yohimbine effects on reinstatement are medi-ated by a nonspecific increase in general activity.
Yohimbine may reinstate responding because it partiallymimics the interoceptive state that had been previously inducedby methamphetamine during training. This issue is potentiallyrelevant because drug discrimination studies suggest that theinteroceptive state induced by stress exposure partially general-izes to the state induced by psychostimulants (Mantsch andGoeders 1998; Miczek et al 1999); however, a discriminativestimulus account of our results is not supported by data fromother studies. At the doses used here, yohimbine does not inducemethamphetamine-like lever responding, as measured in thedrug discrimination procedure (Munzar and Goldberg 1999);however, the more intensive exposure to higher doses of meth-amphetamine in our study than in that of Munzar and Goldbergmay have induced neuroadaptations that resulted in highersensitivity to lower doses of yohimbine. Other evidence fromstudies with cocaine-trained rats and monkeys, however, alsodoes not support the view that the discriminative stimulus effectsof drugs are critically involved in reinstatement (for a discussion,see Shaham et al 2000a). For example, the observation that �-2

acartdr
ytAmsrehi
YP
ai(sby(aytat(aTyTb1isymeaea�1
ybt(rdtbsLpc
J.D. Shepard et al BIOL PSYCHIATRY 2004;55:1082–1089 1087
drenoceptor agonists attenuate footshock-induced, but not co-aine priming-induced, reinstatement of cocaine seeking (Erb etl 2000) is incompatible with a discriminative stimulus account ofeinstatement. In addition, studies with monkeys demonstratehat the ability of a drug to substitute for cocaine in the drugiscrimination procedure is not a good predictor of its effect oneinstatement (Spealman et al 1999).
A surprising finding in this report is that the effect ofohimbine on reinstatement appears to be more pronouncedhan that of footshock, especially during the second test session.
likely explanation for this finding is that yohimbine induced aore sustained increase in brain NA levels than the footshock
tressor. Studies using microdialysis reported that NA brain levelseturn to baseline values within 80–100 min after footshockxposure (Erb et al 2000), whereas yohimbine, which has aalf-life of about 7–8 hours in the rat (Hubbard et al 1988),ncreases NA levels for longer time periods (Becker et al 1999).
ohimbine-Induced Reinstatement: A Neuropharmacologicerspective
Yohimbine is widely used as a prototypical �-2 adrenoceptorntagonist in neuropharmacologic studies, and previous researchmplicated brain NA in footshock stress-induced reinstatementsee Introduction). Thus, our interpretation of the data from thistudy is that yohimbine induces reinstatement by increasing NArain levels via its actions on the �-2 adrenoceptor; however,ohimbine also acts on other receptor subtypes, including D2Scatton et al 1980), �-1 (Doxey et al 1984), and 5-HT1a (Winternd Rabin 1992) receptors. It appears unlikely, that the effect ofohimbine on reinstatement is mediated by these other recep-ors. First, yohimbine is an antagonist of D2 receptors (Scatton etl 1980). Results from reinstatement studies, however, indicatehat D2 receptor antagonists do not reinstate drug seekingShalev et al 2002; Spealman et al 1999). Second, yohimbine hasn �-2/�-1 selectivity ratio of about 40–45 (Doxey et al 1984).hus, it is unlikely that, at the doses used here, the effect ofohimbine on reinstatement is mediated by �-1 adrenoceptors.hird, yohimbine binds to 5-HT1a receptors and mimics certainehavioral effects of agonists of this receptor (Winter and Rabin992). Also, Le et al (2002) reported that intramedian raphenfusions of the 5-HT1a agonist, 8-OH-DPAT, reinstate alcoholeeking; however, it is relatively unlikely that the effect ofohimbine at the 5-HT1a receptor mediates its effect on reinstate-ent. Systemic injections of 8-OH-DPAT had a much weaker
ffect on reinstatement of alcohol seeking than yohimbine (Le etl, unpublished data). Finally, Lee et al (2004) found that theffect of yohimbine on reinstatement is blocked by the �-2drenoceptor agonist clonidine and is mimicked by the selective-2 antagonist, RS-799948–197 (Uhlen et al 1998; Virtanen et al989).
A potential neuropharmacologic interpretation of the data onohimbine-induced reinstatement is that this effect is mediatedy the drug’s action on the mesocorticolimbic dopamine sys-ems, involved in both drug- and stress-induced reinstatementKalivas and McFarland 2003; Shaham et al 2003). Particularlyelevant in this regard is the finding that yohimbine increasesopamine levels in the prefrontal cortex (Tanda et al 1996) andhat local infusions of D1 receptor antagonists into this brain sitelock footshock- or restraint-induced reinstatement of cocaineeeking (Capriles et al 2003; Sanchez et al 2003). The finding ofee et al (2004) that the dopamine receptor antagonist flu-enthixol has no effect on yohimbine-induced reinstatement ofocaine seeking in monkeys is, however, incompatible with the
notion that dopamine is critically involved in yohimbine-inducedreinstatement. Their data should be interpreted with cautionbecause only one dose of flupenthixol was used.
Based on the pharmacologic and neuroanatomic consider-ations described here and on data from previous studies on therole of brain NA in stress-induced reinstatement, it is possiblethat the effect of yohimbine on reinstatement is mediated by theventral noradrenergic projection to the BNST and amygdala thatwere previously implicated in the effect of footshock stress onreinstatement of heroin and cocaine seeking (Shaham et al2000b).
Concluding Remarks and Clinical ImplicationsThe phenomenon of stress-induced reinstatement of drug
seeking in a rat relapse model (de Wit and Stewart 1981; Stewart2000) was first demonstrated using intermittent footshock as thestressor (Shaham and Stewart 1995). This phenomenon wasgeneralized to other pharmacologic (e.g., corticotropin-releasingfactor, metyrapone; Erb et al 2001; Le et al 2002; Shaham et al1997) and environmental (e.g., acute food deprivation; Shalev etal 2000) stressors; however, several other stressors were found tobe ineffective (Shaham et al 2000a), and even in the case ofintermittent footshock, the stressor does not reinstate drugseeking when given in a nondrug environment (Shalev et al2000). These observations, along with other issues related to thevalidity of the reinstatement model, as well as the generalquestion as to whether stress manipulations in nonhumans arerelated to stress in humans, may question the relevance of thephenomenon of stress-induced reinstatement to human drugrelapse (Katz and Higgins 2003). A thorough discussion on therelevance of data from nonhuman reinstatement studies tohuman relapse is beyond the scope of this article (de Wit 1996;Epstein and Preston 2003; Katz and Higgins 2003); however, itappears that our data in rats (Le et al, unpublished data) and thedata of Lee et al (2004) in monkeys on the potent effect ofyohimbine (a drug that induces stress- and anxiety-like symp-toms in humans) on reinstatement of drug seeking, support thenotion that the reinstatement model may be useful in elucidatingthe mechanisms underlying stress-induced drug relapse. Finallyour data with yohimbine and previous data on the inhibitoryeffect of �-2 adrenoceptor agonists on footshock-induced rein-statement of drug seeking (Shaham et al 2003) may provide arationale for the use of these agonists for the prevention ofrelapse to methamphetamine use.
We thank David Chuang for expert technical assistance.
Abercrombie ED, Keller RW Jr, Zigmond MJ (1988): Characterization of hip-pocampal norepinephrine release as measured by microdialysis perfu-sion: Pharmacological and behavioral studies. Neuroscience 27:897–904.
Aghajanian GK, VanderMaelen CP (1982): Alpha 2-adrenoceptor-mediatedhyperpolarization of locus coeruleus neurons: Intracellular studies invivo. Science 215:1394 –1396.
Ahmed SH, Walker JR, Koob GF (2000): Persistent increase in the motivationto take heroin in rats with history of drug escalation. Neuropsychophar-macology 22:413–421.
Becker C, Hamon M, Benoliel JJ (1999): Prevention by 5-HT1A receptoragonists of restraint stress- and yohimbine-induced release of cholecys-tokinin in the frontal cortex of the freely moving rat. Neuropharmacology38:525–532.
Buczek Y, Le AD, Wang A, Stewart J, Shaham Y (1999): Stress reinstatesnicotine seeking but not sucrose solution seeking in rats. Psychopharma-cology 144:183–188.
Capriles N, Rodaros D, Sorge RE, Stewart J (2003): A role for the prefrontalcortex in stress- and cocaine-induced reinstatement of cocaine seekingin rats. Psychopharmacology 168:66 –74.
www.elsevier.com/locate/biopsych

C
C
D
D
d
d
D
E
E
E
E
F
G
G
G
H
H
H
H
H
J
K
K
K
1088 BIOL PSYCHIATRY 2004;55:1082–1089 J.D. Shepard et al
w
harney DS, Heninger GR, Redmond DE Jr (1983): Yohimbine induced anxi-ety and increased noradrenergic function in humans: Effects of diaze-pam and clonidine. Life Sci 33:19 –29.
hopin P, Pellow S, File SE (1986): The effects of yohimbine on explor-atory and locomotor behaviour are attributable to its effects at nor-adrenaline and not at benzodiazepine receptors. Neuropharmacol-ogy 25:53–57.
avis M, Redmond DE Jr, Baraban JM (1979): Noradrenergic agonists andantagonists: Effects on conditioned fear as measured by the potentiatedstartle paradigm. Psychopharmacology (Berl) 65:111–118.
e Vries TJ, Schoffelmeer ANM, Binnekade R, Raasø H, Vanderschuren LJMJ(2002): Relapse to cocaine- and heroin-seeking behavior mediated bydopamine D2 receptors is time-dependent and associated with behav-ioral sensitization. Neuropsychopharmacology 26:18 –26.
e Wit H (1996): Priming effects with drugs and other reinforcers. Exp ClinPsychopharmacol 4:5–10.
e Wit H, Stewart J (1981): Reinstatement of cocaine-reinforced respondingin the rat. Psychopharmacology 75:134 –143.
oxey JC, Lane AC, Roach AG, Virdee NK (1984): Comparison of the alpha-adrenoceptor antagonist profiles of idazoxan (RX 781094), yohimbine,rauwolscine and corynanthine. Naunyn Schmiedebergs Arch Pharmacol325:136 –144.
pstein DH, Preston KL (2003): The reinstatement model and relapse pre-vention: A clinical perspective. Psychopharmacology 168:31–41.
rb S, Hitchcott PK, Rajabi H, Mueller D, Shaham Y, Stewart J (2000): Alpha-2adrenergic agonists block stress-induced reinstatement of cocaine seek-ing. Neuropsychopharmacology 23:138 –150.
rb S, Salmaso N, Rodaros D, Stewart J (2001): A role for the CRF-containingpathway projecting from central nucleus of the amygdala to bed nu-cleus of the stria terminalis in the stress-induced reinstatement of co-caine seeking in rats. Psychopharmacology 158:360 –365.
rb S, Shaham Y, Stewart J (1996): Stress reinstates cocaine-seeking behaviorafter prolonged extinction and drug-free periods. Psychopharmacology128:408 –412.
orray MI, Bustos G, Gysling K (1997): Regulation of norepinephrine releasefrom the rat bed nucleus of the stria terminalis: In vivo microdialysisstudies. J Neurosci Res 50:1040 –1046.
rimm JW, Hope BT, Wise RA, Shaham Y (2001): Incubation of cocainecraving after withdrawal. Nature 412:141–142.
rimm JW, Lu L, Hayashi T, Hope BT, Su TP, Shaham Y (2003): Time-depen-dent increases in brain-derived neurotrophic factor protein levels withinthe mesolimbic dopamine system after withdrawal from cocaine: Impli-cations for incubation of cocaine craving. J Neurosci 23:742–747.
rimm JW, Shaham Y, Hope BT (2002): Effect of the cocaine and sucrosewithdrawal period on extinction behavior, cue-induced reinstatementand protein levels of the dopamine transporter and tyrosine hydroxylasein limbic and cortical areas in rats. Behav Pharmacol 13:379 –388.
arrod SB, Dwoskin LP, Crooks PA, Klebaur JE, Bardo MT (2001): Lobelineattenuates d-methamphetamine self-administration in rats. J PharmacolExp Ther 298:172–179.
ighfield D, Yap J, Grimm J, Shalev U, Shaham Y (2001): Repeated lofexidinetreatment attenuates stress-induced, but not drug cues-induced rein-statement of a heroin-cocaine mixture (speedball) seeking in rats. Neu-ropsychopharmacology 25:320 –331.
olmberg G, Gershon S (1961): Autonomic and psychic effects of yohimbinehydrochloride. Psychopharmacologia 2:93–106.
ubbard JW, Pfister SL, Biediger AM, Herzig TC, Keeton TK (1988): Thepharmacokinetic properties of yohimbine in the conscious rat. NaunynSchmiedebergs Arch Pharmacol 337:583–587.
ughes CE, Habash T, Dykstra LA, Picker MJ (1996): Discriminative-stimuluseffects of morphine in combination with alpha- and beta-noradrenergicagonists and antagonists. Pharmacol Biochem Behav 53:979 –986.
acobsen LK, Southwick SM, Kosten TR (2001): Substance use disorders inpatients with posttraumatic stress disorder: A review of the literature.Am J Psychiatry 158:1184 –1890.
alivas PW, McFarland K (2003): Brain circuitry and the reinstatement ofcocaine-seeking behavior. Psychopharmacology 168:44 –56.
atz JL, Higgins ST (2003): The validity of the reinstatement model of cravingand relapse to drug use. Psychopharmacology 168:21–30.
hoshbouei H, Cecchi M, Dove S, Javors M, Morilak DA (2002): Behavioralreactivity to stress: Amplification of stress-induced noradrenergic acti-vation elicits a galanin-mediated anxiolytic effect in central amygdala.Pharmacol Biochem Behav 71:407–417.
ww.elsevier.com/locate/biopsych
Lang WJ, Gershon S (1963): Effects of psychoactive drugs on yohimbineinduced responses in conscious dogs. A proposed screening procedurefor anti-anxiety agents. Arch Int Pharmacodyn Ther 142:457–472.
Le AD, Harding S, Juzytsch W, Fletcher PJ, Shaham Y (2002): The role ofcorticotropin-releasing factor in the median raphe nucleus in relapse toalcohol. J Neurosci 22:7844 –7849.
Le AD, Quan B, Juzystch W, Fletcher PJ, Joharchi N, Shaham Y (1998): Rein-statement of alcohol-seeking by priming injections of alcohol and expo-sure to stress in rats. Psychopharmacology 135:169 –174.
Lee B, Tiefenbacher S, Platt DM, Spealman RD (2004): Pharmacologicalblockade of alpha(2)-arenoceptors induces reinstatement of cocaine-seeking behavior in squirrel monkeys. Neuropsychopharmacology29:686 –693.
Leri F, Flores J, Rodaros D, Stewart J (2002): Blockade of stress-induced, butnot cocaine-induced reinstatement, by infusion of noradrenergic antag-onists into the bed nucleus of the stria terminalis or the central nucleusof the amygdala. J Neurosci 22:5713–5718.
Lu L, Dempsey J, Liu S, Bossert J, Shaham Y (2004): A single infusion of BDNFinto the ventral tegmental area induces long-lasting potentiation ofcocaine-seeking after withdrawal. J Neurosci 24:1604 –1611.
Lu L, Grimm JW, Dempsey J, Shaham Y (in press): Cocaine seeking overextended withdrawal periods in rats: Different time courses of respond-ing induced by cocaine cues versus cocaine priming over the first 6months. Psychopharmacology.
Lu L, Shepard JD, Scott Hall F, Shaham Y (2003): Effect of environmentalstressors on opiate and psychostimulant reinforcement, reinstatementand discrimination in rats: A review. Neurosci Biobehav Rev 27:457–491.
Mantsch JR, Goeders NE (1998): Generalization of restraint-induced discrim-inative stimulus to cocaine. Psychopharamacology 135:423–426.
Mantsch JR, Goeders NE (1999): Ketoconazole blocks the stress-inducedreinstatement of cocaine-seeking behavior in rats: Relationship to thediscriminative stimulus effects of cocaine. Psychopharmacology142:399 –407.
Martin-Fardon R, Ciccocioppo R, Massi M, Weiss F (2000): Nociceptin pre-vents stress-induced ethanol- but not cocaine-seeking behavior in rats.Neuroreport 11:1939 –1943.
McDougle CJ, Black JE, Malison RT, Zimmermann RC, Kosten TR, HeningerGR, Price LH (1994): Noradrenergic dysregulation during discontinua-tion of cocaine use in addicts. Arch Gen Psychiatry 51:713–719.
Miczek KA, Mutschler NH, van Erp AM, Blank AD, McInerney SC (1999):D-amphetamine “cue” generalizes to social defeat stress: Behavioralsensitization and attenuated accumbens dopamine. Psychopharmacol-ogy 147:190 –199.
Mongeau R, Blier P, de Montigny C (1997): The serotonergic and noradren-ergic systems of the hippocampus: Their interactions and the effects ofantidepressant treatments. Brain Res Rev 23:145–195.
Munzar P, Goldberg SR (1999): Noradrenergic modulation of the discrimina-tive-stimulus effects of methamphetamine in rats. Psychopharmacology143:293–301.
Pacak K, Armando I, Komoly S, Fukuhara K, Weise VK, Holmes C, et al (1992):Hypercortisolemia inhibits yohimbine-induced release of norepineph-rine in the posterolateral hypothalamus of conscious rats. Endocrinology131:1369 –1376.
Rawson RA, Anglin MD, Ling W (2002): Will the methamphetamine problemgo away? J Addict Dis 21:5–19.
Sanchez CJ, Bailie TM, Wu W, Li N, Sorg BA (2003): Manipulation of dopamined1-like receptor activation in the rat medial prefrontal cortex altersstress- and cocaine-induced reinstatement of conditioned place prefer-ence behavior. Neuroscience 119:497–505.
Scatton B, Zivkovic B, Dedek J (1980): Antidopaminergic properties of yo-himbine. J Pharmacol Exp Ther 215:494 –499.
Schroeder BE, Schiltz CA, Kelley AE (2003): Neural activation profile elicitedby cues associated with the anxiogenic drug yohimbine differs from thatobserved for reward-paired cues. Neuropsychopharmacology 28:14 –21.
Sethy VH, Winter JC (1972): Effects of yohimbine and mescaline on punishedbehavior in the rat. Psychopharmacologia 23:160 –166.
Shaham Y, Erb S, Stewart J (2000a): Stress-induced relapse to heroin andcocaine seeking in rats: A review. Brain Res Rev 33:13–33.
Shaham Y, Funk D, Erb S, Brown TJ, Walker CD, Stewart J (1997): Corticotrop-in-releasing factor, but not corticosterone, is involved in stress-inducedrelapse to heroin-seeking in rats. J Neurosci 17:2605–2614.
Shaham Y, Highfield D, Delfs JM, Leung S, Stewart J (2000b): Clonidine blocksstress-induced reinstatement of heroin seeking in rats: An effect inde-

S
S
S
S
S
S
S
S
S
S
S
J.D. Shepard et al BIOL PSYCHIATRY 2004;55:1082–1089 1089
pendent of the locus coeruleus noradrenergic neurons. Eur J Neurosci12:292–302.
haham Y, Shalev U, Lu L, De Wit H, Stewart J (2003): The reinstatementmodel of drug relapse: History, methodology and major findings. Psy-chopharmacology 168:3–20.
haham Y, Stewart J (1995): Stress reinstates heroin self-administration be-havior in drug-free animals: An effect mimicking heroin, not withdrawal.Psychopharmacology 119:334 –341.
halev U, Grimm JW, Shaham Y (2002): Neurobiology of relapse to heroinand cocaine seeking: A review. Pharmacol Rev 54:1–42.
halev U, Highfield D, Yap J, Shaham Y (2000): Stress and relapse to drugseeking in rats: Studies on the generality of the effect. Psychopharmacol-ogy 150:337–346.
halev U, Morales M, Hope B, Yap J, Shaham Y (2001): Time-dependentchanges in extinction behavior and stress-induced reinstatement ofdrug seeking following withdrawal from heroin in rats. Psychopharma-cology 156:98 –107.
inha R (2001): How does stress increase risk of drug abuse and relapse.Psychopharmacology 158:343–359.
pealman RD, Barrett-Larimore RL, Rowlett JK, Platt DM, Khroyan TV (1999):Pharmacological and environmental determinants of relapse to co-caine-seeking behavior. Pharmacol Biochem Behav 64:327–336.
tefanski R, Lee SH, Yasar S, Cadet JL, Goldberg SR (2002): Lack of persistentchanges in the dopaminergic system of rats withdrawn from metham-phetamine self-administration. Eur J Pharmacol 439:59 –68.
tewart J (2000): Pathways to relapse: The neurobiology of drug- and stress-induced relapse to drug-taking. J Psychiatry Neurosci 25:125–136.
tine SM, Grillon CG, Morgan CA 3rd, Kosten TR, Charney DS, Krystal JH (2001):Methadone patients exhibit increased startle and cortisol response afterintravenous yohimbine. Psychopharmacology 154:274–281.
tine SM, Southwick SM, Petrakis IL, Kosten TR, Charney DS, Krystal JH (2002):Yohimbine-induced withdrawal and anxiety symptoms in opioid-de-
pendent patients. Biol Psychiatry 51:642–651.Tanda G, Bassareo V, Di Chiara G (1996): Mianserin markedly and selectivelyincreases extracellular dopamine in the prefrontal cortex as comparedto the nucleus accumbens of the rat. Psychopharmacology 123:127–130.
Tran-Nguyen TL, Fuchs RA, Coffey GP, O’Dell LE, Baker DA, Neisewander JL(1998): Time-dependent changes in cocaine-seeking behavior and do-pamine overflow in the amygdala during cocaine withdrawal. Neuropsy-chopharmacology 19:48 –59.
Uhlen S, Dambrova M, Nasman J, Schioth HB, Gu Y, Wikberg-Matsson A,Wikberg JE (1998): [3H]RS79948 –197 binding to human, rat, guinea pigand pig alpha2A-, alpha2B- and alpha2C-adrenoceptors. Comparisonwith MK912, RX821002, rauwolscine and yohimbine. Eur J Pharmacol343:93–101.
Virtanen R, Savola JM, Saano V (1989): Highly selective and specific antago-nism of central and peripheral alpha 2-adrenoceptors by atipamezole.Arch Int Pharmacodyn Ther 297:190 –204.
Wada T, Fukuda N (1991): Effects of DN-2327, a new anxiolytic, diazepamand buspirone on exploratory activity of the rat in an elevated plus-maze. Psychopharmacology 104:444 –450.
Wang X, Cen X, Lu L (2001): Noradrenaline in the bed nucleus of the striaterminalis is critical for stress-induced reactivation of morphine-condi-tioned place preference in rats. Eur J Pharmacol 432:153–161.
White DA, Birkle DL (2001): The differential effects of prenatal stress in ratson the acoustic startle reflex under baseline conditions and in responseto anxiogenic drugs. Psychopharmacology 154:169 –176.
Winter JC, Rabin RA (1992): Yohimbine as a serotonergic agent: Evidencefrom receptor binding and drug discrimination. J Pharmacol Exp Ther263:682–689.
Yokel RA (1987): Intravenous self-administration: Response rates, the effectsof pharmacological challenges, and drug preference. In: Bozarth MA,editor. Methods of Assessing the Reinforcing properties of Abused Drug.New York: Springer-Verlag, 1–34.
Yokel RA, Pickens R (1973): Self-administration of optical isomers ofamphetamine and methylamphetamine by rats. J Pharmacol Exp Ther
187:27–33.www.elsevier.com/locate/biopsych





























