Embed Size (px)
Citation preview

Entomol. exp. appI. 49:195-201 (1988) �9 Kluwer Academic Publishers, Dordrecht - Printed in the Netherlands 195
The adaptation process of Ceratitis capitata to the laboratory analysis of life-history traits
Hebe Myrina Laghi de Souza, Sergio Russo Matioli 1 & Wilma Nascimento de Souza Departamento de Gen~tica e Evolur Instituto de Biologia, Universidade Estadual de Campinas, Caixa Postal 6109, 13100 Campinas - SP,, Brasil - 1Present address: Departarnento de Biologia, Instituto de Bioci~ncias, Universidade de Sao Paulo, Caixa Postal H461, 05421 Sgo Paulo - SP, Brasil
Accepted: June 6, 1988
Key words: Ceratitis capitata, Diptera, Tephritidae, life-history, adaptation, colonization
Abstract
Colonizing species are believed to have genetic plasticity that permits their adaptation to new habitats. The Mediterranean fruit fly, Ceratitis capitata, is known for this ability, reflected in its present world-wide ge- ographical distribution. To investigate the genetic components of the colonizing capacity of this species, we studied the life-table parameters in (a) a recently-collected wild population, (b) a laboratory strain maintained for 6 years, and (c) a hybrid population resulting from crosses between the wild females and long-term laborato- ry maintained males. All the strains were collected in the same host and place. Within a few generations, both the hybrid and the wild strains showed life table parameters (egg, larval, pupal and adult mortalities, net reproductive rate, intrinsic rate of population growth, and generation time) similar to those of the laboratory strain. A difference in the speed of adaptation to the laboratory conditions was remarkable, with the hybrid population reaching the values of the long-term laboratory strain much faster than the wild population. An incipient reproductive isolation between the laboratory and the wild population was also observed.
The genetic plasticity that allows fast adaptation to new conditions such as those in the laboratory is high in C. capitata, although the genetical variability measured by enzyme electrophoresis in Brazilian populations is low. This suggests that the isozyme variation observed is not related to this kind of adaptation.
Introduction
The Mediterranean fruit fly, Ceratitis capitata, is one of the major fruit crop pests in Brazil. The fly originated in Africa but today presents a world-wide geographic distribution, showing great colonizing potential.
The mass-rearing procedures used under artificial conditions will inevitably produce flies that are different from the wild ones, so that problems may arise when these flies are used in control programs, such as those based on the sterile insect technique.
Laboratory-reared flies can also be useful for the study of problems concerning adaptation and colonization processes, in a more general biological approach.
The existence of adaptation processes in laboratory-reared insect populations is a general phenomenon, (Souza et al., 1970 and Boiler, 1972). In mass-reared populations of C. capitata, Rossler (1975) showed differences between collected flies reared for 12 generations and populations main- tained for more than 12 years. The adaptation processes that occurred when C. capitata was in-
196
troduced to the laboratory were fast, taking only a few generations (Mu~iz, 1986).
Using life-table analysis, we studied the genetic ef- fects on the fitness of laboratory-reared, field- collected and hybrid populations of Ceratitis capita- ta under laboratory conditions.
Material and methods
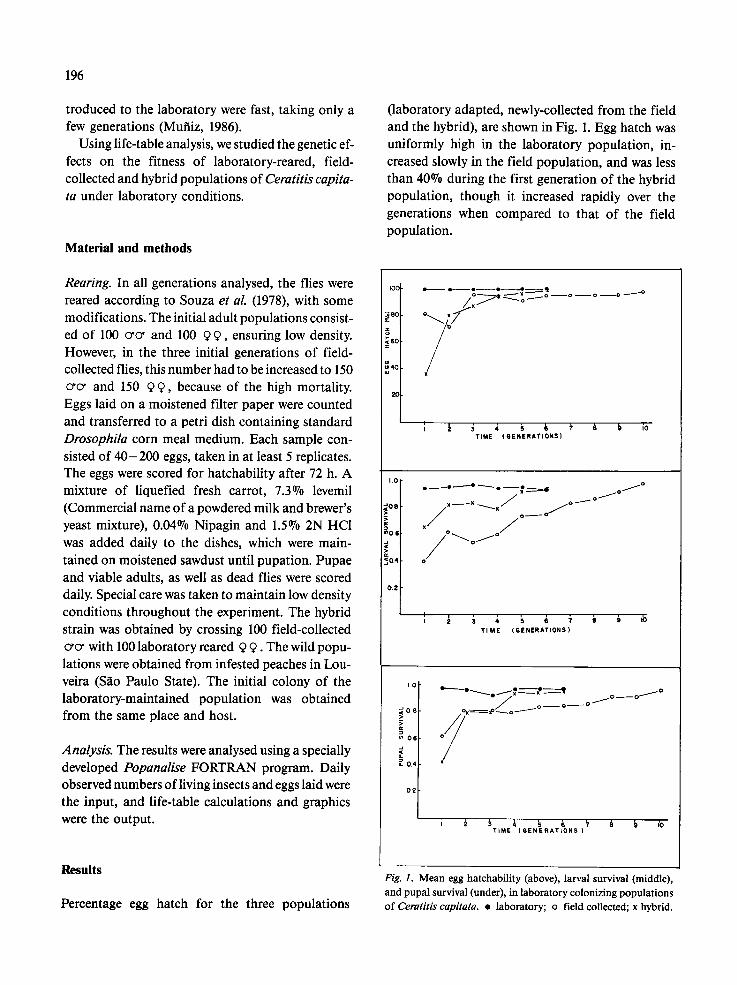
(laboratory adapted, newly-collected from the field and the hybrid), are shown in Fig. 1. Egg hatch was uniformly high in the laboratory population, in- creased slowly in the field population, and was less than 40~ during the first generation of the hybrid population, though it increased rapidly over the generations when compared to that of the field population.
Rearing. In all generations analysed, the flies were reared according to Souza et al. (1978), with some modifications. The initial adult populations consist- ed of 100 o'c~ and 100 9 9, ensuring low density. However, in the three initial generations of field- collected flies, this number had to be increased to 150 o'ce and 150 9 9, because of the high mortality. Eggs laid on a moistened filter paper were counted and transferred to a petri dish containing standard Drosophila corn meal medium. Each sample con- sisted of 40- 200 eggs, taken in at least 5 replicates. The eggs were scored for hatchability after 72 h. A mixture of liquefied fresh carrot, 7.3% levemil (Commercial name of a powdered milk and brewer's yeast mixture), 0.04~ Nipagin and 1.5~ 2N HCI was added daily to the dishes, which were main- tained on moistened sawdust until pupation. Pupae and viable adults, as well as dead flies were scored daily. Special care was taken to maintain low density conditions throughout the experiment. The hybrid strain was obtained by crossing 100 field-collected o'o' with I00 laboratory reared 9 9. The wild popu- lations were obtained from infested peaches in Lou- veira (Sao Paulo State). The initial colony of the laboratory-maintained population was obtained from the same place and host.
Analysis. The results were analysed using a specially developed Popanalise FORTRAN program. Daily observed numbers of living insects and eggs laid were the input, and life-table calculations and graphics were the output.
Results
Percentage egg hatch for the three populations
TIME (GENERATIONS}
1.0
06
Oq
0.4
0.2
I , ~ ~ ; ~ ~ § ~ ~ TIME (GENERATIONS)
TiME (GENERATIONS)
Fig. 1. Mean egg hatchability (above), larval survival (middle), and pupal survival (under), in laboratory colonizing populations of Ceratitis capitata. �9 laboratory; o field collected; x hybrid.
197
�9 5 o e o _ 7 o
' ' ' ' - 5
2 0 -
E 6 6 S l O -
P E R 5 . O - F E M A L E
PER i.O- OAr
0 . 5 -
3
T I M E (GENERATIONS)
~" 0 .I ;o ' ~o ' ; o ' 4o ' s'o ' 6o ' ; o TiME AFTER EMERGENCE(DAYS)
70 80
2 0 - E 6 6 S l O -
P E R FEMALE 5 . 0 -
PER OAr
- - 5 TI ME
i {GENERATIONS)
>o.I , ' o ' ~ o ' ~o
TIME AFTER ' ~ o ' ~o ' ~ o ' ~ o '
EMERGENCE {DAYS)
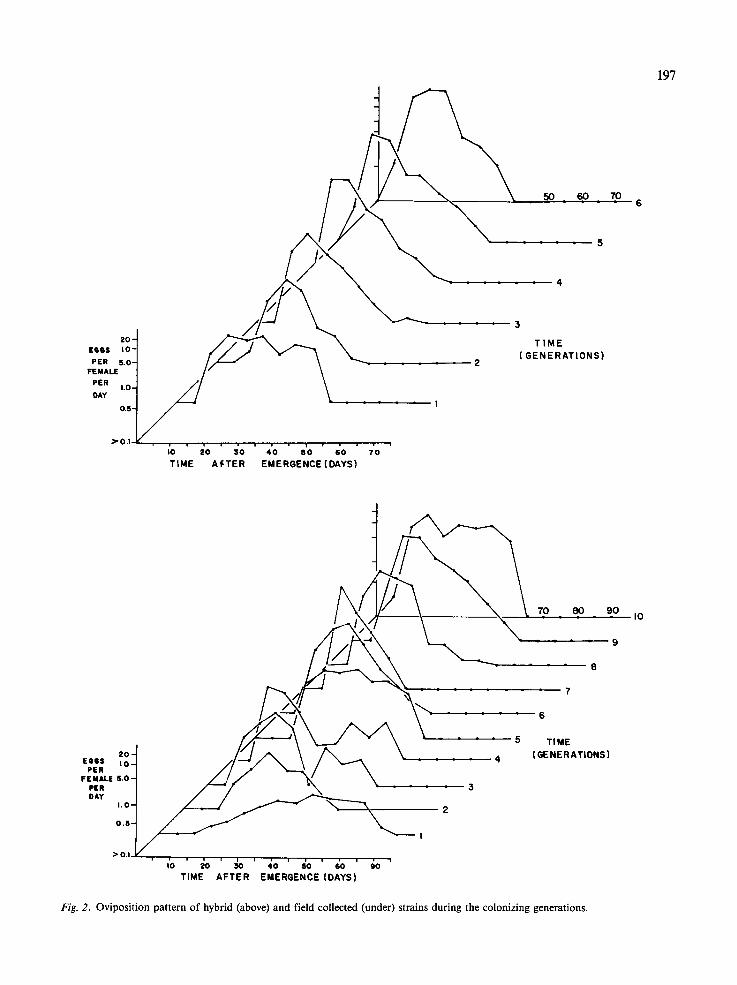
Fig. 2. Oviposition pattern of hybrid (above) and field collected (under) strains during the colonizing generations.
90
9
8
I0
198
A uniform and higher level survival (Fig. 1) was again observed in the laboratory population. Although the hybrid and the field populations showed a variable increase in larval survival, their curves were almost parallel, with a lower starting val- ue for the field population than for the hybrid popu- lation.
The pattern of pupal viability (Fig. 1) showed a behaviour of this trait similar to that observed for egg hatch, i.e., uniformity for the laboratory popula- tion, a slow increase for the field population, and a fast increase for the hybrid population, which also showed the lowest initial value.
The changing oviposition patterns of the hybrid and field populations are shown in Fig. 2. The aver- age oviposition pattern of the laboratory popula- tions is shown in Fig. 3. These graphs show three clear trends: both field and hybrid populations showed a) a progressive increase in average egg num- ber; b) a shorter duration of the pre-oviposition peri- od; and c) an earlier occurrence of the post- oviposition period. However, the improved adapta- tion to laboratory conditions was more pronounced in the hybrid than in the field population. In the first generations of the field population the oviposition pattern is very different from both hybrid and laboratory populations. In this case, the flies oviposit almost until they die, practically without post-oviposition period. The oviposition pattern of the field population remained different from the laboratory population even after 10 generations.
From these population data, the following life-
FtUA~ I0 P[fl DAY 5.0
O5
< 0 1 . . . . . 5 I 0 15 20 25 30 xm, 4 0 45
DAYS A F T E R E M E R G E N C E
Fig. 3. Mean oviposition rate (+ S.D.) of laboratory strain.
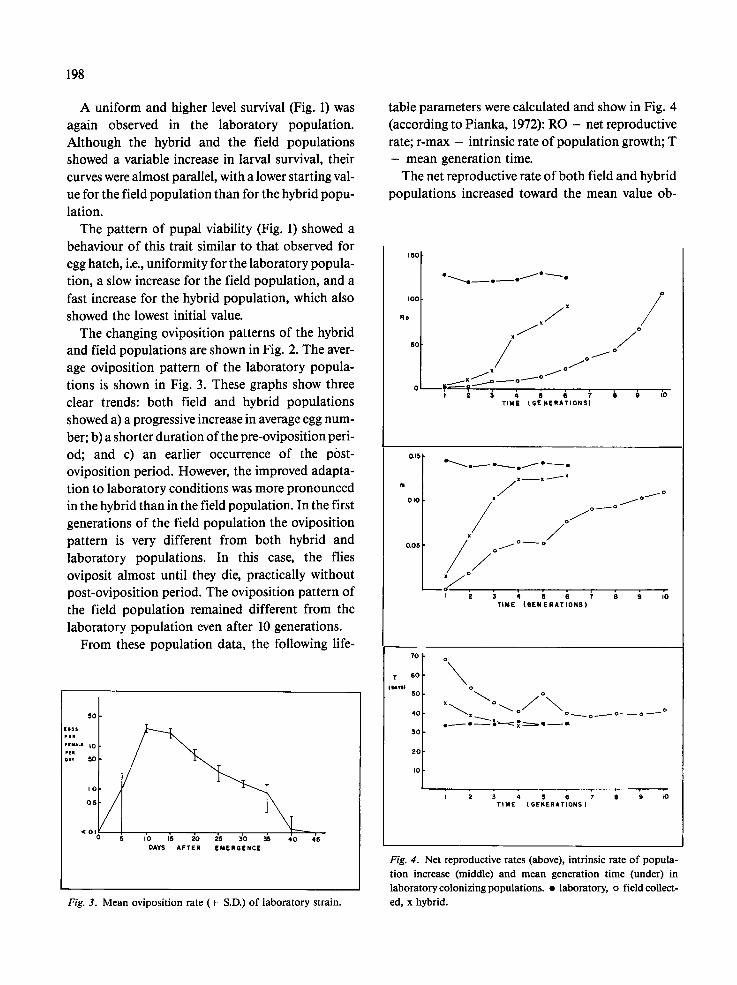
table parameters were calculated and show in Fig. 4 (according to Pianka, 1972): RO - net reproductive rate; r-max - intrinsic rate of population growth; T
- mean generation time. The net reproductive rate of both field and hybrid
populations increased toward the mean value ob-
~ f / o ~ o ~ o ~ o / " e - ' - - - - - o
i ~176 R, j x
50
0 I ~ 1 ,
T I M E LGE N E R A T I O N S )
<}.IS
m
010
0 .05
~ o ~ o...,...~ o ~ �9 ~ o
J x - x ~ x .......~. o
~ 1 7 6 o
T i M E ( G E N E R A T I O W S )
T
10#,111
70
GO
5r
4C
5C
2 (
IC
O\o x ~ o / o \
T I M E ( G E N E R A T I O N S )
Fig. 4. Net reproductive rates (above), intrinsic rate of popula- tion increase (middle) and mean generation time (under) in laboratory colonizing populations. �9 laboratory, o field collect- ed, x hybrid.

tained for the laboratory populations and the proc- ess was faster in the hybrid population. The intrinsic rate of population growth showed a pattern similar to that obtained for the RO coefficient, although the slopes of the curves were almost linear. Mean genera- tion time decreased in both field and hybrid popula- tions, and progress was also faster in the hybrid population.
Discussion
The adaptation of flies to laboratory conditions may be regarded as a kind of colonizing phenomenon. The intrinsic rate of population growth may be relat- ed to the average fitness of a population. The results presented here show changes in the traits that in- dividually lead to a colonization event. With the ex- ception of the unusual behaviour occurring in the first generation with respect to egg hatch and pupal survival, the major trend was as follows: the hybrid strain tended to approach the behaviour of the laboratory strain faster than the field-collected population, in all studied traits. This observation strongly suggests that all traits were under the con- trol of selective forces leading to a laboratory colonizing process.
This finding contrasts with some theoretical con- cepts. A colonizing species is believed to be ecologi- cally a generalist, exhibiting a fine-grained be- haviour, with phenotypes having the same fitness in various habitats (Parsons, 1983; Rozenzweig, 1981). The degree of genetic variability, as measured by en- zyme electrophoresis is lower in C. capitata, when compared to other tephritids (e.g. the genus Rhago- letis) and to other Diptera (Morgante et al., 1981), but is close to the variability of the genusAnastrepha (Morgante et al., 1980). Genetic variability did not seem to be absent in the traits anlaysed in the present study. The hypothesis of the loss of genetic variabili- ty by genetic drift caused by fluctuations in popula- tion size must therefore be revised, because this kind of evolutionary constraint would be responsible for a general decrease in genetic variability.
The exact nature of the variability shown in the colonization process remains unclear, but some hy- potheses may be advanced. Some atypical and natu-
199
rally rare behaviour could be selected for in unusual conditions such as maintenance in the laboratory. Observations of this kind were reported in Drosophi- la willistoni, which showed increased frequency of unusual behaviour in populations cages (Souza et al., 1970). This behaviour could lead to the observed incipient reproductive isolation between laboratory strain and field population represented by the low hatchability of hybrid eggs and lower pupal survival observed in the present study. These phenomena may be due to hybrid disgenesis, or, in the case of eggs, to unsuccessful mating. Rossler (1975) also ob- served this kind of reproductive isolation, but it should be pointed out that his 'field' strain was kept under laboratory conditions for twelve generations, when most of the modifications occur, as shown here. With respect to the low pupal survival in the hybrid population during the first generation, it may be the result of a certain incompatibility between the selected genotypes and the 'wild' ones, a case of hybrid inviability, also suggesting some degree of post-mating reproductive isolation.
The changes in the oviposition pattern show that during the laboratory colonization a typical r- selection occurs, whereby the strategy of long-term oviposition is fully replaced with a more opportunis- tic strategy. This may be caused by the rearing proce- dure, whereby the first eggs to hatch produce a new generation and the late ones are lost. This r-selection is characteristic of laboratory populations of Cerati- tis capitata, (Vargas et al., 1984, Mufiiz & Gil, 1986), with a remarkable similarity of independently recorded oviposition patterns. The situation in na- ture is quite different, as inferred by the first genera- tion of the wild strain. Disruptive selection could be occurring, with fast developing flies being selected as the colonizing ones, while the late developing flies may be advantageous at the end of the specific fruit season by being able to exploit another host which matures later. Evidence that this species is subject to disruptive selection with respect to larval survival comes from the work of Krainacker et al. (1987), showing that different hosts can cause different responses regarding life-history parameters, even in a genetically uniform laboratory population.
Life-history parameters were shown to be affected by selection in experiments carried out with

200
Drosophila melanogaster (Rose & Charlesworth, 1981a and 1981b), although this took place in laboratory-maintained strains which could already have been selected for many parameters.
The present study shows the dynamics of the proc- ess of initial adpatation of artificially reared Cerati- tis capitata. The practice of complete re- colonization as used in mass-rearing procedures must be revised, since the time for adaptation is very short, covering about 10-15 generations. Therefore, continuous input of natural population samples into the mass-reared population will provide a sufficient source of naturally adapted genotypes or genes, although it certainly will reduce the efficience of production proportionally. This input should be made at the shortest possible rate, in order to prevent oscillations of production. The ideal procedure to yield flies which are competitive in nature is the one that causes the minimum selective constraint, as measured by the rate of increase of the rm parameter during the early colonizing generations. It may be achieved by designing a rearing system that resem- bles natural conditions, especially those related to behaviour stimuli, as courtship places, and feeding sites.
am selben Wirt und Ort gesammelt. Nach wenigen Generationen zeigten sowohl die Kreuzungs- wie auch die wilden St~imme Lebensdatenparameter (Eier, Sterblichkeit der Larven, Puppen and Adulten, Nettofortpflanzungsrate, spezifische Populationsvermehrungsrate, Generationszeit), die denen des Laboratoriumsstammes ~ihnlich waren.
Dennoch zeigte sich ein merkwiirdiger Unter- schied in der Geschwindigkeit der Anpassung an die Laboratoriumsbedingungen: Die Fliegen der Kreu- zungspopulation erreichten die Werte des Laboratoriumsstammes viel schneller als die Fliegen der Wildpopulation. Ferner wurde eine anf/ingliche Fortpflanzungsisolierung zwischen der Laboratoriums- und der Wildpopulation beo- bachtet.
Daraus wird geschlossen, dass die genetische Plastizit~it, die eine schnelle Anpassung an neue Be- dingungen (z.B. des Laboratoriums) zul~isst, in dies- er Fliegenart hoch ist, obwohl die mit Enzymelek- trophorese gemessene Ver~inderlichkeit bei brasilianischen Populationen von C. capitata niedrig ist. Dies l~isst vermuten, dass die beobachtete Isozymver/inderlichkeit zu dieser Art Anpassung nicht in Beziehung steht.
Zusammenfassung
Der A npassungsprozess von Ceratitis capitata an das Laboratorium: Analyse der Lebensdatenparameter
Wahrscheinlich besitzen kolonisierende Insektenar- ten eine genetische Plastizit~it, die es ihnen er- m6glicht, sich an neue Habitats anzupassen. Die Mittlemeerfruchtfliege, Ceratitis capitata, ist wegen dieser F~ihigkeit allgemein bekannt, sie spiegelt sich in ihrer gegenw~irtigen geographischen Verbreitung in aller Welt wieder. Um die genetischen Anteile an der Kolonisationsf~ihigkeit dieser Fliegenart zu un- tersuchen, sch~itzten wir die Lebensdaten-parameter bei folgenden Populationen: (a) bei einem frisch gesammelten Muster der Wildpopulation; (b) bei ei- nem 6 Jahre lang geziichteten Laboratoriums- stamm; (c) bei einer Kreuzungspopulation, die aus der Kombination wilde Weibchen und Laboratori- umsm~innchen erhalten wurde. Alle St/imme wurden
Acknowledgements
The authors thank Dr. A. B. da Cunha and Dr. J. S. Morgante for criticism, Mrs. Irani M. Fernandes for drawing.
Research supported by grants from Conselho Na- cional de Desenvolvimento Cientifico e Tecnol6gico, Programa Integrado de Gen6tica, proc. #4003185.
References
Boiler, E., 1972. Behavioral aspects of mass-rearing of insects. Entomophaga 17(1): 9 - 2 5 .
Kralnacker, D. A., J. R. Carey & R. I. Vargas, 1987. Effect of larval host on life history traits of the mediterranean fruit fly, Cerati- tis capitata. Oecologia 73: 5483-590.
Morgante, J. S., A. Malavasi & G. L. Bush, 1980. Biochemical Sys- tematics and evolutionary relationships of neotropical Anas- trepha. Ann. Entomol. Soc. Am. 73." 622-630.
Morgante, J. S., H. M. L. Souza, E. De Conti & M. Cytrynowicz,

1981. Allozymic variability in an introduced fruit fly pest Cera- titis capitata (Wiedeman, 1824), (Diptera-Tephritidae). Rev. Brasil. Genet. 4: 183-191.
Mu~iz, M., 1986. Studies on a rapid adaptation of the Mediterra- nean fruit fly. In: Proceedings of the CEC/IOBC 'ad-hoc meet- ing', Hamburg on fruit flies of economic importance. (Caval- loro, R., eds). A. A. Balkema, Rotterdam pp. 121-124.
Muiliz, M. & A. Gil, 1986. Laboratory studies on isolated pairs of Ceratitis capitata (Wied.): Results obtained during the last years in Spain. In: Proceedings of the CEC/IOBC 'ad-hoc meeting', Hamburg, 1984, on fruit flies of economic impor- tance (Cavalloro, R., eds). A. A. Balkema, Rotterdam pp. 125 - 127.
Parsons, P. A., 1983. The evolutionary biology of colonizing spe- cies. Cambridge University Press, Cambridge 262 pp.
Pianka, E. R., 1972. Evolutionary Ecology. Harper & Row, New York.
Rose, M. R. & B. Charlesworth, 1981a. Genetics of life history in Drosophila melanogaster. I. Sib analysis of adult females. Genetics 97: 173-186.
201
Rose, M. R. & B. Charlesworth, 1981b. Genetics of life-history in Drosophila melanogaster. II. Exploratory selection experi- ments. Genetics 97: 187-196.
Rozensweig, M. L., 1981. A theory of habitat selection. Ecology 62: 327- 335.
Rossler, Y., 1975. Reproductive differences between laboratory- reared and field-collected populations of the mediterranean fruitfly, Ceratitis capitata. Ann. Entomol. Soc. Am. 68: 987-991.
Souza, H. M. L. de, A. B. Da Cunha & E. P. dos Santos, 1970. Adaptive polymorphism of behaviour evolved in laboratory populations of Drosophila willistonia. Am. Nat. 104: 175-179.
Souza, H. M. L. de, A. E. Piedrabuena & O. H. O. Pavan, 1978. Biologia de Ceratitis capitata (Wiedmann) (Diptera - Tephrit- idae). Um novo meio artificial de criacao para producao em massa. Papeis Avulsos Zool. Sao Paulo 31(14): 213-220.
Vargas, R. I., D. Miyashita & T. Nishida, 1984. Life history and demographic parameters of three laboratory-reared tephritids (Diptera: Tephritidae). Ann. EntomoL Soc. Am. 77:651-656.





























