Embed Size (px)
Citation preview

1062-3590/02/2906- $27.00 © 2002
MAIK “Nauka
/Interperiodica”0618
Biology Bulletin, Vol. 29, No. 6, 2002, pp. 618–627. Translated from Izvestiya Akademii Nauk, Seriya Biologicheskaya, No. 6, 2002, pp. 752–762.Original Russian Text Copyright © 2002 by Rakhleeva.
INTRODUCTION
At present spatial distribution of soil testaceans byvarious natural zones receives considerable attention.This group of nannofauna actively colonizes the upperorganogenic and organo-mineral soil horizons of theforest and meadow habitats yielding high population,biomass, and species diversity in many of them (Bon-net, 1953; Thomas, 1954; Couteaux, 1976; Gel’tser
et al.
, 1980; etc.). The data on specific structure oftestacean population in a number of biocenoses inCiscaucasian Russia are available for southern taigasoils of Moscow Region (Korganova, 1979), middletaiga soils of Tver and Arkhangelsk Regions (Alekseev,1984), southern taiga soils of Ryazan Region (Rakhle-eva, 1998), zonal soil types of the Russian Plain(Bobrov, 1999), etc. Testaceans prefer wetland habitatswith low rates of organic matter decomposition.
There is very little evidence concerning testaceanpopulations at the ample territories of Western Siberiaand no data at all is available for Surgut Polesye. At thesame time, this slumpy and waterlogged region pre-sents favorable conditions for development of testaceancomplex making their investigation quite attractive. Inaddition, investigation of pedobionts is substantiated byactive exploitation of oil and gas fields in the regionfrom the mid-1980s. This raises the problem of conser-vation of Surgut Polesye and neighboring territorieswith soil invertebrates as an integral component.
The goal of this work was to evaluate specific struc-ture of testacean complexes (Testacea, Protozoa) oftaiga soils in Surgut Polesye.
REGION OF INVESTIGATION
The investigation was carried out in the territory ofSurgut Polesye (Tyumen Region) that belongs to theLower Ob pedogeographical province of boggy andgley–weakly podzolic soils of northern taiga (at theboundary between the middle and northern taiga)(Dobrovol’skii and Urusevskaya, 1984). By terrain thisis a flat low plain (neighboring Siberian Uvaly at thenorth) with 50–150 m true altitude. The bedrocks arecovered with powerful layer of quaternary sedimentswith fluvial and glaciofluvial deposits as the most com-mon soil-forming rocks. These sediments are mostlypresented by small- and fine-grained sand (Dobro-vol’skii
et al.
, 1981). By landscape the low plains of theglacial zone are quite similar to the regions of Belorus-sian Polesye; which substantiated wide application ofthis purely East European term for description of theWestern Siberia lowland within recent years.
Specific climatic conditions, soil-forming rocks,and terrain features have considerable effect on soil for-mation. Considerable moistening at the background oflow evaporation and prolonged stay in frozen conditionreducing the active period of soil formation deceleratebiochemical transformations and favor active boggingand peat deposition. This also benefits from low drain-ing of the territory. These processes result in consider-able bogging of the studied region reaching 90% in Sur-gut Polesye (
Atlas…
, 1968; Liss and Berezina, 1981).
The soil cover of the sand plains features wide dis-tribution of boggy soils covering 85–90% total territoryon the average. Only 10–15% falls on sandy podzolsassociated with hill-like interamnian elevations and riv-erine regions with a better draining. Sandy podzols are
Testaceans (Testacea, Protozoa) of Taiga Soilsin Western Siberia (Surgut Polesye)
A. A. Rakhleeva
Faculty of Soil Science, Moscow State University, Vorob’evy gory, Moscow, 119899 RussiaE-mail: [email protected]
Received September 1, 2000
Abstract
—We studied population structure of testaceans of automorphic and hydromorphic soils in the middletaiga of Western Siberia. Rich testacean fauna has been revealed. The fauna representation was comparable tothe studied soils of Ciscaucasian Russia. The bryophilic group was the most typical for the region, since its rep-resentatives inhabited the predominating hydromorphic soils of bogs. The typical species included
Amphitremaflavum
,
Heleopera petricola
,
Placocista spinosa
,
Trigonopyxis arcula
,
Centropyxis laevigata
,
Nebela tincta
,
Arcella catinus
, as well as other moss forms. Flood plain proved to be the richest by testacean species variabilityamong the studied habitats. The set of species here included both representatives of watershed and typicalaquatic forms present due to the flood conditions of a given area. The highest quantitative indices of the testa-cean complexes were revealed in the forest area soils.
Quadrulella quadrigera
Deflandre, 1936 found in a floodplain area has been revealed for the first time in Russia.
ECOLOGY

BIOLOGY BULLETIN
Vol. 29
No. 6
2002
TESTACEANS (TESTACEA, PROTOZOA) OF TAIGA SOILS 619
presented by illuvial-ferruginous, illuvial-ferruginouswith tongue-shaped podzolic horizon, and illuvial-humic podzols (Dolgova and Gavrilova, 1971;Gavrilova and Dolgova, 1972).
Flora on the positive well-draining terrain elementsis presented by light lichen pine forest with poor anduniform floristic composition. There is no underwoodand pine reproduction is low. Hydrophilous smallshrubs appear in the depressions with increased moist-ening: marsh tea and bog whortleberry. Reindeer mosspredominates in the soil cover. Vast bog areas are occu-pied by bog vegetation.
MATERIALS AND METHODS
Protozoological investigation was carried out inSeptember 1999 as a part of complex investigation ofWestern Siberia soils carried out at the Department ofSoil Geography (Faculty of Soil Science, Moscow StateUniversity). Four habitats were studied.
1.
Cowberry-lichen pine forest
. This habited is asso-ciated to flat top of ridgy elevation. The tree layer iscomposed of pine. The soil cover includes cowberry,whortleberry, bog whortleberry, marsh tea, and lichen.The soil is illuvial-ferruginous podzol.
The upper organogenic horizon (A0) is fresh andconsiderably decomposed; it readily smears in thelower part; can be easily divided into two subhorizons:L (0–2 cm) with 45.3% field moisture content andF + H (2–5 cm) with 66.4% field moisture content; thelitter pH is 4.7.
1
2.
Pine-spruce sphagnum forest
neighbors a bog.The tree layer includes pine, spruce, and cedar. Under-growth includes cedar, birch, aspen, and spruce. Theshrub layer includes ashberry and wild rose. The soilcover includes whortleberry, common cow-wheat, cow-berry, marsh tea, and sphagnum. The soil is illuvial-fer-ruginous gley podzol.
The upper horizon of litter can be easily differenti-ated by layers. Subhorizon L (0–6 cm) (field moisturecontent is 83.8%, pH 5.0) includes undecomposed veg-etative green part of moss, rag, and needles; it is wet.Subhorizon F (6–12 cm) (field moisture content is76.3%, pH 4.35) includes brown decomposing mass; itis quite wet.
3.
Birch-cedar moss fen
. Flood plain of the fen river(30 m from the streamway). Ligneous vegetationincludes birch and cedar; shrubs include ashberry andmarsh tea; the grass cover includes rough blue-joint,buckbean, marsh cinquefoil, sedges, sphagnum, andgreen mosses. The soil is fluvial lowland peat-humus.Depth to watertable is 10 cm.
The upper organic horizons Ws
2
(0–10 cm) and T1(10–25 cm) are considerably wet and include vegetative(Ws) (field moisture content 100%, pH 6.2) and mori-
1
The upper organic soil horizons inhabited by testaceans werestudied in all habitats. Soil nomenclature corresponds to
Klassi-fiklatsiya…
, 1977.
bund (T1) (field moisture content 100%, pH 6.3) partsof sphagnum and green mosses.
2
4. Ridge-pool subshrub-sphagnum bog
. It is the cen-ter of flat plain inclined to river valley. Ridge-pool bogis oligotrophic and slightly mesotrophic. Arctic birch,white birch, myrtle, cloudberry, marsh tea, sheatingcottongrass, cranberry, bog whortleberry, cowberry,sphagnum and green mosses (field moisture content86.0%, pH 3.3). The vegetation of pool (field moisturecontent 100%, pH 3.7) includes mud sedge, butterbur,cranberry, sheating cottongrass, and sphagnums. Depthto watertable in pool and ridge is 3 and 25 cm, respec-tively. The soil is bog peat.
Samples for protozoological test were taken fromthe sample soil profiles. The samples were collectedfrom the middle of genetic horizons down to the depthwhere testaceans occurred (about 20 cm) and keptrefrigerated. Simultaneously the samples for fieldmoisture assay were collected.
Species composition and quantitation of testaceanswere studied in water suspension of a fresh soil sample:10 g substrate was mixed with 50 ml water and settledfor 1 h. After shaking up for 10 min it was put througha sieve (1 mm cell) to remove large masking particlesof litter and detritus. The filtrate volume was adjustedto 100 ml and the aquatic suspension was inspectedunder microscope (up to
×
600). A drop of suspensionapplied on a coverslip was examined in six repetitions.A drop of glycerol was added to a drop of suspensionfor a more convenient inspection (to make it more vis-cous). The number of testaceans was reduced per 1 goven-dry sample.
Testacean species were identified after Deflandre(1929), Bonnet and Thomas (1960), Decloitre (1962),Gel’tser
et al.
(1995), etc.
Testaceans biomass was determined in 1 ml of theobtained suspension placed in a 4 cm plastic Petri dishwith 2 ml water and 5–8 drops of carbolic erythrosinand stained for 1 day. The stained specimens wereaccounted under microscope at 4 cm
2
area outlined onthe dish bottom. The number of testaceans was reducedper 1 g open-dry soil using biomass recalculation for-mula (Korganova, 1997).
In addition we determined the indices of diversity(Shannon and Weaver, 1963), uniformity (Pielou,1969), diversity of dominant species (Simpson, 1949),and
β
-diversity (Megarran, 1992). Classes of domi-nance were distinguished (Whittaker, 1977). The struc-ture of morphotypes was studied on the basis of thestandard testacean morphotypes (Korganova, 1997).Cluster analysis was carried out by the indices of pairedfaunistic similarity (Jackard, cited from Chernov,1975). Ecological preferendums of individual specieswere determined.
2
Ws is sphagnum waste composed of undecomposed stems of sph-agnum mosses with infusion of rhizomes of subshrubs and rareligneous roots.
620
BIOLOGY BULLETIN
Vol. 29
No. 6
2002
RAKHLEEVA
RESULTS AND DISCUSSION
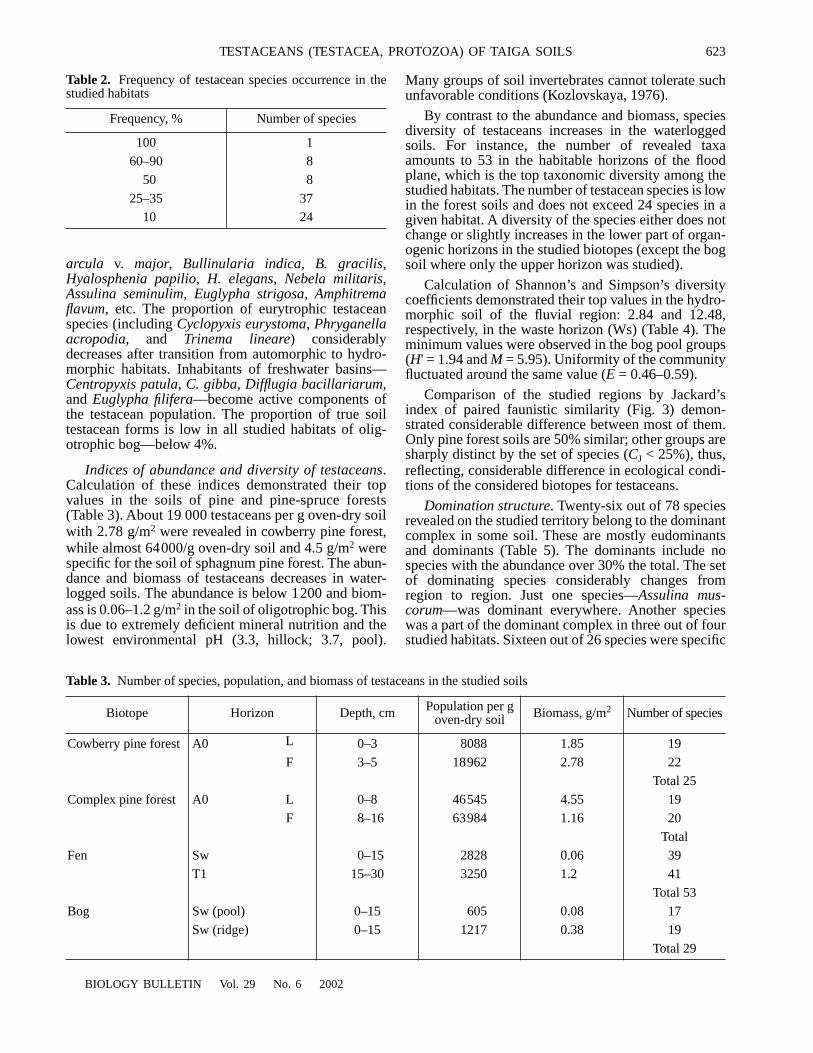
High waterlogging and low rates of organic mattertransformation favor development of testacean com-plex in the studied region. Rich fauna of testaceans hasbeen revealed in the soil of Surgut Polesye: 78 speciesand intraspecific taxa falling in 23 genera and 15 fami-lies (Table 1). Thirty found species have never beenmentioned in the publications on testacean populationsof Western Siberia (southern taiga, Tomsk Region)(Rat’kova, 1971, cited from Korganova, 1978; Bobrov
et al.
, 1995).
Almost 55% fauna of the studied habitats representCentropyxidae, Hyalospheniidae, and Euglyphidaefamilies amounting to 11–13 species each. Four fami-lies—Gromiidae, Amphitrematiidae, Cyphoderiidae,and Cryptodifflugiidae—include single species each.Other families include 2–8 testacean taxa. Representa-tives of Plagiopyxidae family typical for the Europeansoils amounts to only 6% fauna; the proportion ofCyclopyxidae and Trinematiidae is also decreased.
Note relatively low frequency of the revealed testa-ceans in the studied soil samples (Table 2). Over61 species (80% species composition) occur only in1
−2
studied habitats, which indicates highly contrasthabitats. Only one species
Assulina muscorum
wasfound in all studied samples. Its presence indicates rawhumus in the substrate and low pH (below 6.0) (Lous-ier, 1972). Only nine species are widespread. By con-trast to this region, widespread testacean species com-prised almost 30% fauna in the Oka Reserve (RyazanRegion, southern taiga in Ciscaucasian Russia)(Rakhleeva, 2000).
Structure of morphological types.
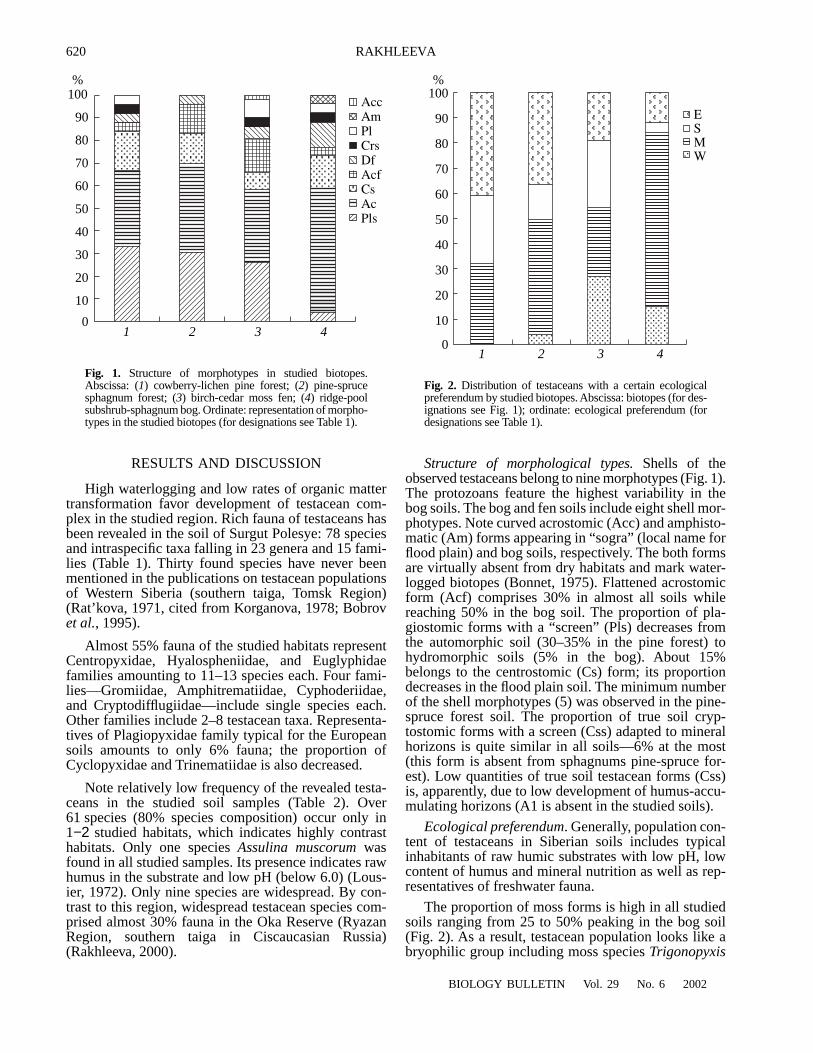
Shells of theobserved testaceans belong to nine morphotypes (Fig. 1).The protozoans feature the highest variability in thebog soils. The bog and fen soils include eight shell mor-photypes. Note curved acrostomic (Acc) and amphisto-matic (Am) forms appearing in “sogra” (local name forflood plain) and bog soils, respectively. The both formsare virtually absent from dry habitats and mark water-logged biotopes (Bonnet, 1975). Flattened acrostomicform (Acf) comprises 30% in almost all soils whilereaching 50% in the bog soil. The proportion of pla-giostomic forms with a “screen” (Pls) decreases fromthe automorphic soil (30–35% in the pine forest) tohydromorphic soils (5% in the bog). About 15%belongs to the centrostomic (Cs) form; its proportiondecreases in the flood plain soil. The minimum numberof the shell morphotypes (5) was observed in the pine-spruce forest soil. The proportion of true soil cryp-tostomic forms with a screen (Css) adapted to mineralhorizons is quite similar in all soils—6% at the most(this form is absent from sphagnums pine-spruce for-est). Low quantities of true soil testacean forms (Css)is, apparently, due to low development of humus-accu-mulating horizons (A1 is absent in the studied soils).
Ecological preferendum
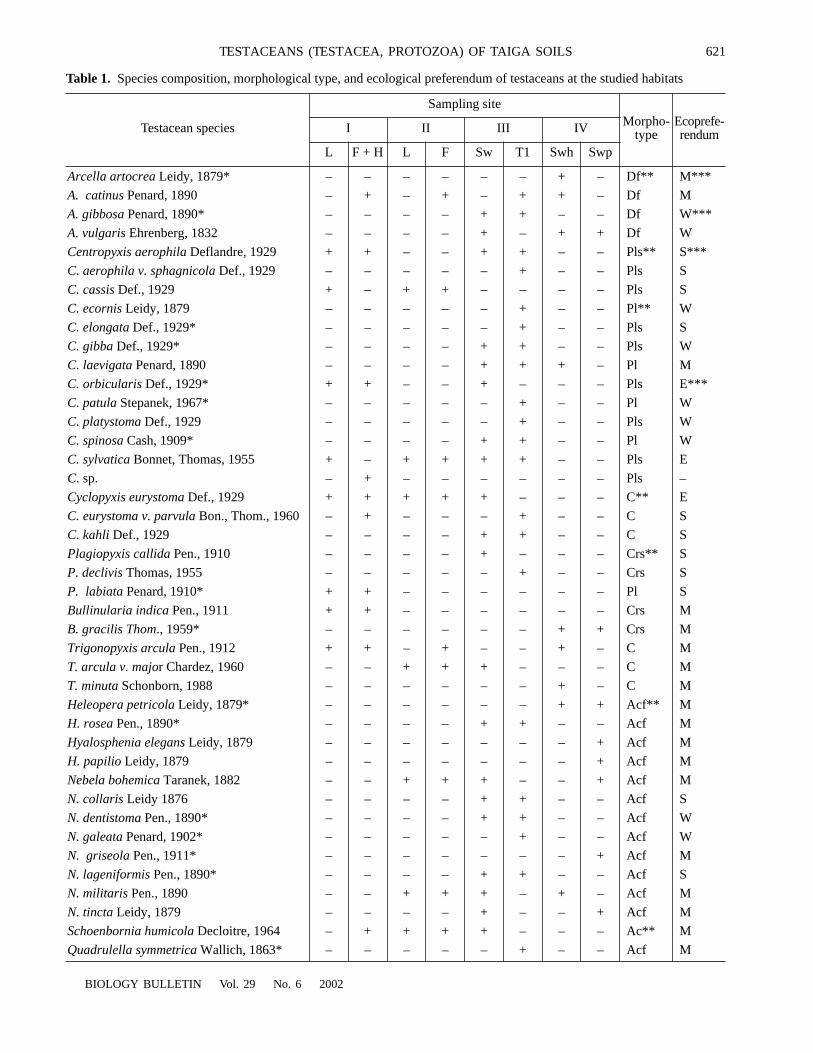
. Generally, population con-tent of testaceans in Siberian soils includes typicalinhabitants of raw humic substrates with low pH, lowcontent of humus and mineral nutrition as well as rep-resentatives of freshwater fauna.
The proportion of moss forms is high in all studiedsoils ranging from 25 to 50% peaking in the bog soil(Fig. 2). As a result, testacean population looks like abryophilic group including moss species
Trigonopyxis
1 2 3 4
0
10
20
30
40
50
60
70
80
90
100%
AccAmPlCrsDfAcfCsAcPls
Fig. 1.
Structure of morphotypes in studied biotopes.Abscissa: (
1
) cowberry-lichen pine forest; (
2
) pine-sprucesphagnum forest; (
3
) birch-cedar moss fen; (
4
) ridge-poolsubshrub-sphagnum bog. Ordinate: representation of morpho-types in the studied biotopes (for designations see Table 1).
1 2 3 4
0
10
20
30
40
50
60
70
80
90
100%
ESMW
Fig. 2.
Distribution of testaceans with a certain ecologicalpreferendum by studied biotopes. Abscissa: biotopes (for des-ignations see Fig. 1); ordinate: ecological preferendum (fordesignations see Table 1).
BIOLOGY BULLETIN
Vol. 29
No. 6
2002
TESTACEANS (TESTACEA, PROTOZOA) OF TAIGA SOILS 621
Table 1.
Species composition, morphological type, and ecological preferendum of testaceans at the studied habitats
Testacean species
Sampling siteMorpho-
typeEcoprefe-rendumI II III IV
L F + H L F Sw T1 Swh Swp
Arcella artocrea
Leidy, 1879* – – – – – – + – Df** M***
A. catinus
Penard, 1890 – + – + – + + – Df M
A. gibbosa
Penard, 1890* – – – – + + – – Df W***
A. vulgaris
Ehrenberg, 1832 – – – – + – + + Df W
Centropyxis aerophila
Deflandre, 1929 + + – – + + – – Pls** S***
C. aerophila v. sphagnicola
Def., 1929 – – – – – + – – Pls S
C. cassis
Def., 1929 + – + + – – – – Pls S
C. ecornis
Leidy, 1879 – – – – – + – – Pl** W
C. elongata
Def., 1929* – – – – – + – – Pls S
C. gibba
Def., 1929* – – – – + + – – Pls W
C. laevigata
Penard, 1890 – – – – + + + – Pl M
C. orbicularis
Def., 1929* + + – – + – – – Pls E***
C. patula
Stepanek, 1967* – – – – – + – – Pl W
C. platystoma
Def., 1929 – – – – – + – – Pls W
C. spinosa
Cash, 1909* – – – – + + – – Pl W
C. sylvatica
Bonnet, Thomas, 1955 + – + + + + – – Pls E
C
. sp. – + – – – – – – Pls –
Cyclopyxis eurystoma
Def., 1929 + + + + + – – – C** E
C. eurystoma v. parvula
Bon., Thom., 1960 – + – – – + – – C S
C. kahli
Def., 1929 – – – – + + – – C S
Plagiopyxis callida
Pen., 1910 – – – – + – – – Crs** S
P. declivis
Thomas, 1955 – – – – – + – – Crs S
P. labiata
Penard, 1910* + + – – – – – – Pl S
Bullinularia indica
Pen., 1911 + + – – – – – – Crs M
B. gracilis Thom
., 1959* – – – – – – + + Crs M
Trigonopyxis arcula
Pen., 1912 + + – + – – + – C M
T. arcula v. majo
r Chardez, 1960 – – + + + – – – C M
T. minuta
Schonborn, 1988 – – – – – – + – C M
Heleopera petricola
Leidy, 1879* – – – – – – + + Acf** M
H. rosea
Pen., 1890* – – – – + + – – Acf M
Hyalosphenia elegans
Leidy, 1879 – – – – – – – + Acf M
H. papilio
Leidy, 1879 – – – – – – – + Acf M
Nebela bohemica
Taranek, 1882 – – + + + – – + Acf M
N. collaris
Leidy 1876 – – – – + + – – Acf S
N. dentistoma
Pen., 1890* – – – – + + – – Acf W
N. galeata
Penard, 1902* – – – – – + – – Acf W
N. griseola
Pen., 1911* – – – – – – – + Acf M
N. lageniformis
Pen., 1890* – – – – + + – – Acf S
N. militaris
Pen., 1890 – – + + + – + – Acf M
N. tincta
Leidy, 1879 – – – – + – – + Acf M
Schoenbornia humicola
Decloitre, 1964 – + + + + – – – Ac** M
Quadrulella symmetrica
Wallich, 1863* – – – – – + – – Acf M

622
BIOLOGY BULLETIN
Vol. 29
No. 6
2002
RAKHLEEVA
Table 1.
(Contd.)
Testacean species
Sampling siteMorpho-
typeEcoprefe-rendumI II III IV
L F + H L F Sw T1 Swh Swp
Q. quadrigera
Def., 1936* – – – – + + – – Acf M
Difflugia bacillariarum
Perty, 1849* – – – – – + – – Ac W
D. lucida
Pen., 1890* – – – – + + – – Acf W
D
. sp – – – – + + – – Ac W
Pseudodifflugia gracilis
Schlumberger, 1845 – – + + – – – – Ac W
Phryganella acropodia Hopkinson
, 1909 + + – – – – – – C E
P. hemisphaerica Pen., 1902* – – – – – – – + C W
P. sp. – – – – – – + + C M
Euglypha acantophora Ehrenberg, 1949* – – – – – – – + Ac M
E. anadonta v. magna Schon. , 1964* + + – – – – – – Acf S
E. ciliata Leidy, 1878 + + + + + + + – Acf E
E. compressa Carter, 1964 – – – – – – – + Acf E
E. compressa f. glabra Wailes, 1915* + + – + – – – – Acf E
E. cristata Leidy, 1878 – – – – + + – – Ac W
E. filifera Pen., 1890 – – – – + + – + Acf W
E. laevis Perty, 1849 + + + + + + – + Acf E
E. strigosa Leidy, 1879 + + – – + + – – Acf M
E. tuberculata Dujardin, 1841 – – – – – + – – Ac M
Assulina muscorum Greef, 1888 + + + + + + + + Acf S
A. seminulum Leidy, 1879 – + + + – – + + Acf M
A. sp. + + + + + + + – Acf –
Placocista lens Pen., 1899* – – – + – – – + Acf M
P. spinosa Carter, 1865* – – – – – – + + Acf M
Tracheleuglypha acolla Bon., Thom., 1955 – – – – + + – – Ac S
T. acolla v. aspera Bon. , Thom. , 1955 – – – – + – – – Ac S
Trinema complanatum Pen., 1890 – – + + + + – – Pls S
T. enchelys Leidy, 1878 – – – – – + – – Pls E
T. lineare Pen., 1890 + + + + + + + – Pls E
T. lineare v. minuscula Chardez, 1971* – – – – + + + – Pls E
T. penardi Thom., Chardez, 1958* – – – – + + – – Pls S
Corythion delamarei Bon., Thom., 1960* + – + – – – – – Pls M
C. dubium Taranek, 1881 + + + + + – – – Pls E
C. orbicularis Iudina, 1996* – + + – + – – – Pls M
Amphitrema flavum Archer, 1877 – – – – – – – + Am** W
Cyphoderia ampulla Ehrenberg, 1840* – – – – – + – – Acc** W
Difflugiella oviformis v. fusca Pen. , 1890 – – + – + + – – Ac E
Note: Sampling sites: I, cowberry-lichen pine forest, subhorizons L and F + H; II, pine-spruce sphagnum forest, subhorizons L and F; III, birch-cedar moss fen, horizons Ws and T1; IV, ridge-pool subshrub-sphagnum bog, Sw horizons developed on hillock (Swh) and pool (Swp).
* Species first found in Western Siberia ** Morphotypes: Df, flattened discoid; Pl, plagiostomic simple; Pls, plagiostomic with a screen; C, centrostomic; Ac, acrostomic sim-
ple; Acf, flattened acrostomic; Am, amphistomatic; Crs, cryptostomic with a screen; Acc, curved acrostomic *** Testacean forms: W, water; M, moss; S, soil; E, eurytopic (after Chardez, 1965; Ogden and Hedley, 1980; Aescht and Foissner, 1988;
our observations).
BIOLOGY BULLETIN Vol. 29 No. 6 2002
TESTACEANS (TESTACEA, PROTOZOA) OF TAIGA SOILS 623
arcula v. major, Bullinularia indica, B. gracilis,Hyalosphenia papilio, H. elegans, Nebela militaris,Assulina seminulim, Euglypha strigosa, Amphitremaflavum, etc. The proportion of eurytrophic testaceanspecies (including Cyclopyxis eurystoma, Phryganellaacropodia, and Trinema lineare) considerablydecreases after transition from automorphic to hydro-morphic habitats. Inhabitants of freshwater basins—Centropyxis patula, C. gibba, Difflugia bacillariarum,and Euglypha filifera—become active components ofthe testacean population. The proportion of true soiltestacean forms is low in all studied habitats of olig-otrophic bog—below 4%.
Indices of abundance and diversity of testaceans.Calculation of these indices demonstrated their topvalues in the soils of pine and pine-spruce forests(Table 3). About 19 000 testaceans per g oven-dry soilwith 2.78 g/m2 were revealed in cowberry pine forest,while almost 64000/g oven-dry soil and 4.5 g/m2 werespecific for the soil of sphagnum pine forest. The abun-dance and biomass of testaceans decreases in water-logged soils. The abundance is below 1200 and biom-ass is 0.06–1.2 g/m2 in the soil of oligotrophic bog. Thisis due to extremely deficient mineral nutrition and thelowest environmental pH (3.3, hillock; 3.7, pool).
Many groups of soil invertebrates cannot tolerate suchunfavorable conditions (Kozlovskaya, 1976).
By contrast to the abundance and biomass, speciesdiversity of testaceans increases in the waterloggedsoils. For instance, the number of revealed taxaamounts to 53 in the habitable horizons of the floodplane, which is the top taxonomic diversity among thestudied habitats. The number of testacean species is lowin the forest soils and does not exceed 24 species in agiven habitat. A diversity of the species either does notchange or slightly increases in the lower part of organ-ogenic horizons in the studied biotopes (except the bogsoil where only the upper horizon was studied).
Calculation of Shannon’s and Simpson’s diversitycoefficients demonstrated their top values in the hydro-morphic soil of the fluvial region: 2.84 and 12.48,respectively, in the waste horizon (Ws) (Table 4). Theminimum values were observed in the bog pool groups(H' = 1.94 and M = 5.95). Uniformity of the communityfluctuated around the same value (E = 0.46–0.59).
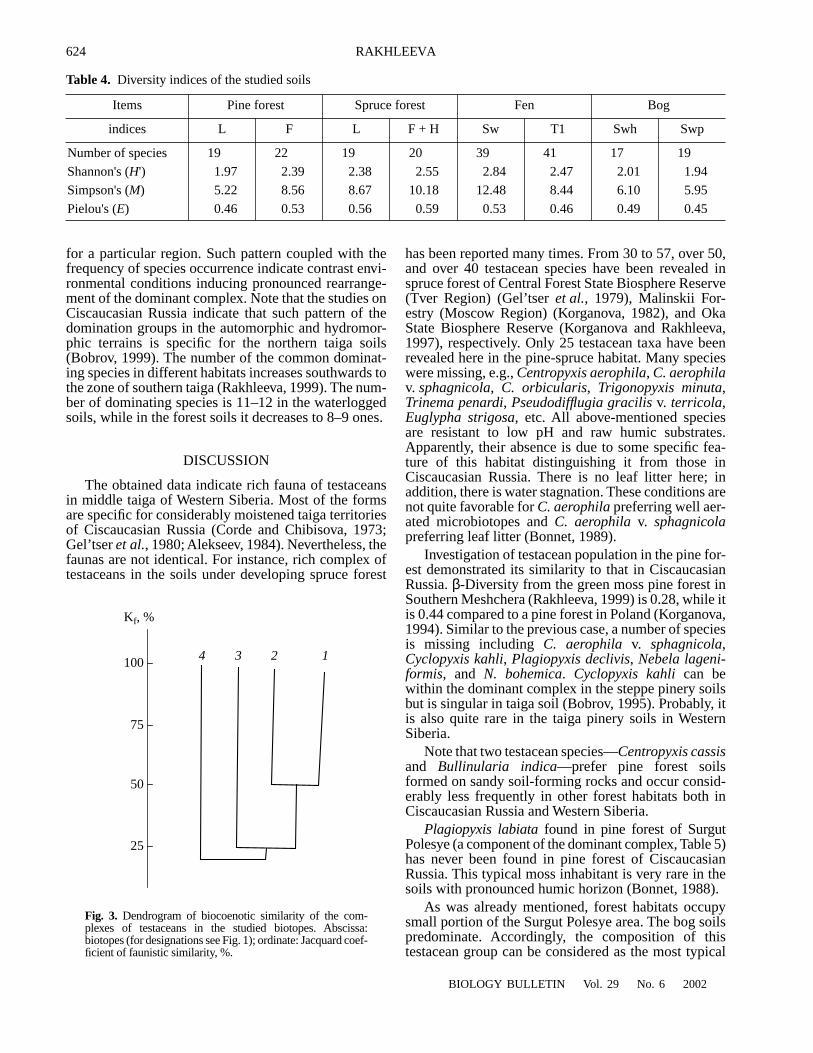
Comparison of the studied regions by Jackard’sindex of paired faunistic similarity (Fig. 3) demon-strated considerable difference between most of them.Only pine forest soils are 50% similar; other groups aresharply distinct by the set of species (CJ < 25%), thus,reflecting, considerable difference in ecological condi-tions of the considered biotopes for testaceans.
Domination structure. Twenty-six out of 78 speciesrevealed on the studied territory belong to the dominantcomplex in some soil. These are mostly eudominantsand dominants (Table 5). The dominants include nospecies with the abundance over 30% the total. The setof dominating species considerably changes fromregion to region. Just one species—Assulina mus-corum—was dominant everywhere. Another specieswas a part of the dominant complex in three out of fourstudied habitats. Sixteen out of 26 species were specific
Table 2. Frequency of testacean species occurrence in thestudied habitats
Frequency, % Number of species
100 1
60–90 8
50 8
25–35 37
10 24
Table 3. Number of species, population, and biomass of testaceans in the studied soils
Biotope Horizon Depth, cm Population per goven-dry soil Biomass, g/m2 Number of species
Cowberry pine forest A0 L 0–3 8088 1.85 19
F 3–5 18962 2.78 22
Total 25
Complex pine forest A0 L 0–8 46545 4.55 19
F 8–16 63984 1.16 20
Total
Fen Sw 0–15 2828 0.06 39
T1 15–30 3250 1.2 41
Total 53
Bog Sw (pool) 0–15 605 0.08 17
Sw (ridge) 0–15 1217 0.38 19
Total 29
624
BIOLOGY BULLETIN Vol. 29 No. 6 2002
RAKHLEEVA
for a particular region. Such pattern coupled with thefrequency of species occurrence indicate contrast envi-ronmental conditions inducing pronounced rearrange-ment of the dominant complex. Note that the studies onCiscaucasian Russia indicate that such pattern of thedomination groups in the automorphic and hydromor-phic terrains is specific for the northern taiga soils(Bobrov, 1999). The number of the common dominat-ing species in different habitats increases southwards tothe zone of southern taiga (Rakhleeva, 1999). The num-ber of dominating species is 11–12 in the waterloggedsoils, while in the forest soils it decreases to 8–9 ones.
DISCUSSION
The obtained data indicate rich fauna of testaceansin middle taiga of Western Siberia. Most of the formsare specific for considerably moistened taiga territoriesof Ciscaucasian Russia (Corde and Chibisova, 1973;Gel’tser et al., 1980; Alekseev, 1984). Nevertheless, thefaunas are not identical. For instance, rich complex oftestaceans in the soils under developing spruce forest
has been reported many times. From 30 to 57, over 50,and over 40 testacean species have been revealed inspruce forest of Central Forest State Biosphere Reserve(Tver Region) (Gel’tser et al., 1979), Malinskii For-estry (Moscow Region) (Korganova, 1982), and OkaState Biosphere Reserve (Korganova and Rakhleeva,1997), respectively. Only 25 testacean taxa have beenrevealed here in the pine-spruce habitat. Many specieswere missing, e.g., Centropyxis aerophila, C. aerophilav. sphagnicola, C. orbicularis, Trigonopyxis minuta,Trinema penardi, Pseudodifflugia gracilis v. terricola,Euglypha strigosa, etc. All above-mentioned speciesare resistant to low pH and raw humic substrates.Apparently, their absence is due to some specific fea-ture of this habitat distinguishing it from those inCiscaucasian Russia. There is no leaf litter here; inaddition, there is water stagnation. These conditions arenot quite favorable for C. aerophila preferring well aer-ated microbiotopes and C. aerophila v. sphagnicolapreferring leaf litter (Bonnet, 1989).
Investigation of testacean population in the pine for-est demonstrated its similarity to that in CiscaucasianRussia. β-Diversity from the green moss pine forest inSouthern Meshchera (Rakhleeva, 1999) is 0.28, while itis 0.44 compared to a pine forest in Poland (Korganova,1994). Similar to the previous case, a number of speciesis missing including C. aerophila v. sphagnicola,Cyclopyxis kahli, Plagiopyxis declivis, Nebela lageni-formis, and N. bohemica. Cyclopyxis kahli can bewithin the dominant complex in the steppe pinery soilsbut is singular in taiga soil (Bobrov, 1995). Probably, itis also quite rare in the taiga pinery soils in WesternSiberia.
Note that two testacean species—Centropyxis cassisand Bullinularia indica—prefer pine forest soilsformed on sandy soil-forming rocks and occur consid-erably less frequently in other forest habitats both inCiscaucasian Russia and Western Siberia.
Plagiopyxis labiata found in pine forest of SurgutPolesye (a component of the dominant complex, Table 5)has never been found in pine forest of CiscaucasianRussia. This typical moss inhabitant is very rare in thesoils with pronounced humic horizon (Bonnet, 1988).
As was already mentioned, forest habitats occupysmall portion of the Surgut Polesye area. The bog soilspredominate. Accordingly, the composition of thistestacean group can be considered as the most typical
Table 4. Diversity indices of the studied soils
Items Pine forest Spruce forest Fen Bog
indices L F L F + H Sw T1 Swh Swp
Number of species 19 22 19 20 39 41 17 19
Shannon's (H') 1.97 2.39 2.38 2.55 2.84 2.47 2.01 1.94
Simpson's (M) 5.22 8.56 8.67 10.18 12.48 8.44 6.10 5.95
Pielou's (E) 0.46 0.53 0.56 0.59 0.53 0.46 0.49 0.45
100
Kf, %
75
50
25
4 3 2 1
Fig. 3. Dendrogram of biocoenotic similarity of the com-plexes of testaceans in the studied biotopes. Abscissa:biotopes (for designations see Fig. 1); ordinate: Jacquard coef-ficient of faunistic similarity, %.

BIOLOGY BULLETIN Vol. 29 No. 6 2002
TESTACEANS (TESTACEA, PROTOZOA) OF TAIGA SOILS 625
for the studied territory. Note higher species variabilityof testaceans (29 species) in the bog soils as comparedto the forest ones (Table 3). A typical bryophilic groupis present (Bartos, 1954; Schönborn, 1966; Gel’tseret al., 1980; Alekseev, 1984). Microrelief becomes par-ticularly important here. The set of testacean species iscomposed of those inhabiting ridges and pools. Theridges permanently include Trigonopyxis arcula, T.minuta, and Nebela militaris, that are also common inthe forest biotopes with raw moss litter. Typical repre-sentatives of bogs—Hyalosphenia papilio, H. elegans,Amphitrema flavum, Nebela griseola, and Placocistaspinosa—were observed in the pool substrate andproved to be resistant to highly unfavorable conditionsof this habitat. Jackard’s index of paired faunistic simi-larity demonstrates low similarity between testaceanpopulations of the ridge and pool (24%).
The region of fen in the flood plane of a river provedto be the most variable by taxa composition of testa-ceans. Fifty-three out of 78 testacean species have beenrevealed there. Apparently, more neutral pH (6.2 and6.3 at Ws and T1, respectively), increase in the avail-able nutrition elements from river drifts, ample moist-ening and peat favor active development of testaceans.Considerable number of moss and aquatic testaceanforms has been revealed (Table 1).
Finding Quadrulella quadrigera in this habitat is ofinterest. This species was never found in Russia before.It was described by Deflandre in 1936 and has beenfound in the soils of North America, Guinea, Sey-chelles, and British Columbia (Chardez, 1967). Byecology it is an inhabitant of hyperhydromorphic mosssubstrates. It features a combination of square and rect-angular idiosomas specific for Quadrulella genus and
Table 5. Set of dominant species in the studied samples (>5% total population)
Testacean species I* II* III* IV*
Plagiopyxis labiata 10.8 – – –
Euglypha compressa f. glabra 21.6–10.8 – – –
Phryganella acropodia 6.5 – – –
Pseudodifflugia gracilis – 12.6 – –
Cyclopyxis eurystoma – – 7 –
Plagiopyxis callida – – 5.8 –
Euglypha laevis – – 14.2 –
E. tuberculata – – 5 –
Trinema complanatum – – 9.3 –
Tracheleuglypha acolla – – 19.8–20.2 –
Assulina sp. – – – 11.4
Heleopera petricola – – – 6.8
Placocista spinosa – – – 26
Nebela tincta – – – 13
Hyalosphenia papilio – – – 22
Phryganella sp. – – – 8.6
Nebela griseola – – – 13
Amphitrema flavum – – – 13
Nebela bohemica – 17.1–9.3 5.8 –
N. militaris – 15.4–13.1 7 –
Trinema lineare – 6.6 16.2 –
Centropyxis sylvatica – 11.6–9.3 14.2 –
Trinema lineare v. minuscule – – 7 11.4
Arcella catinus – 13.7 – 11.4
Trigonopyxis arcula 13.4 – – 20.4
Corythion dubium 20.4–14.7 15.4 – –
Schoenbornia humicola 19.6 14.8 – –
Assulina muscorum 29.6–12.5 12.1 8 27.2
* See Table 1 for the sampling site designations.

626
BIOLOGY BULLETIN Vol. 29 No. 6 2002
RAKHLEEVA
oval and rounded idiosomas specific for Nebela genus.Quadrulella quadrigera has been found in all horizonswithin the studied habitat. In the lower part of the pro-file it was complemented by one more Quadrulella spe-cies—Q. symmetrica. This species occurs in the north-ern and southern taiga soils of Ciscaucasian Russia(Gel’tser et al., 1980; Bobrov, 1999), particularly, inthe hydromorphic soils with mesotrophic waterloggingin Karelia, where it can be a component of the dominantcomplex.
Another hydrophilous species—Cyphoderiaampulla—has been found in the same soil. It is of inter-est that this species has been revealed only in the north-ern taiga soils of Ciscaucasian Russia (Bobrov, 1999).This species prefers wetland raw humic substrates(Lousier, 1972). It is possible that its range in the mod-erate zone is limited by the northern taiga territories.
Thus, the bryophilic group is the most typical forthis region, since its representatives inhabit hydromor-phic soils of bogs predominating by area. The typicalspecies include Amphitrema flavum, Heleopera petri-cola, Placocista spinosa, Trigonopyxis arcula, Centro-pyxis laevigata, Nebela tincta, Arcella catinus, as wellas other moss forms.
The flood plane proved to be the richest by testaceanspecies diversity among the studied habitats. The set ofits species include both representatives of watershedareas and typical aquatic forms, which is due to theflood plane conditions of this region.
Quantitative investigation of the testacean complexdemonstrates that these indices peak in the forest soils.
ACKNOWLEDGMENTS
Where grateful to the organizers of Siberian expedi-tion S.Ya. Trofimov and N.A. Avetov for introducinginto field studies, chief environmental manager of OAOSurgutneftegaz L.A. Malyshkina for the help in fieldmaterial collection, student of Faculty of Soil Science(Moscow State University) V. Perevozchikov for thedata on pH of the soil samples. This work was sup-ported by the Russian Foundation for Basic Research(project no. 00-04-48510a).
REFERENCES
Aescht, E. and Foissner, W., Catalogus Faunae Austria. TeilIa: Stamm Rhizopoda, Verlag Osterreichischen Acad. Wis-senschaften, 1989, pp. 1–81.Alekseev, D.A., Testaceans in Soils of Boggy Forests in theNorthern Subzone of European Taiga, Cand. Sc. (Biol.) Dis-sertation, Moscow: Moscow State University, 1984.Atlas torfyanykh resursov SSSR (Atlas of Peat Resources ofUSSR), GU Geodezii i Kartografii, 1968.Bartos,ˇ E., ˇKorenonozceˇ Radu Testacea, Bratislava: Vyda-vatel’stvo Slovenskej Akademie Vied., 1954.Bobrov, A.A., Species Distribution of Testaceans (Protozoa,Testacea) in Pinery Soils, Biologicheskoe raznoobrazie
lesnykh ekosistem (Biodiversity of Forest Ecosystems),1995, pp. 134–136.Bobrov, A.A., Ecogeographical Pattern of Testaceans Distri-bution and Community Structure (Protozoa: Testacea), Doc-toral (Biol.) Dissertation, Moscow: Moscow State Univer-sity, 1999.Bobrov, A.A., Yazvenco, S.B., and Warner, B.G., Taxonomicand Ecological Implications of Shell Morphology of TreeTestaceans (Protozoa: Rhizopoda) in Russian and Canada,Arch. Protistenkd., 1995, vol. 145, pp. 119–126.Bonnet, L., Thécamoebiens de Quelques Stations du Midi dela France, Bull. Soc. Hist. Natur. Toulouse, 1953, vol. 88,vols. 1–2. pp. 31–33.Bonnet, L., Types Morphologiques, Écologie et évolution dela Théque chez les Thécamoebiens, Protistologica, 1975,vol. XI, no. 3, pp. 363–378.Bonnet, L., Ecology du genre Plagiopyxis (Thecamoebiensdes sols), Bull. Soc. Hist. Nat. Toulouse, 1988, vol. 124,pp. 13–21.Bonnet, L., Donnees Ecologiques sur Quelques Centropyxi-dae (Thecamoebiens) des Sols, Bull. Soc. Hist. Nat. Tou-louse, 1989, vol. 125, pp. 7–16.Bonnet, L. and Thomas, R., Thecamoebiens du Sol, FauneTerrestre d’Eau Douce des Pyrenees-Orientales, 1960,vol. 5, pp. 1–103.Chardez, D., Écologie générale des Thecamoebiens(Rhizopoda, Testacea), Bull. Inst. Agron. Stat. Rech. Gembl,1965, vol. 33, no. 3, pp. 307–341.Chardez, D., Monographie du Genre Quadrulella Cockerell(Protozoa, Rhizopoda Testacea), Bull. Rech. Agron. Gem-blou, 1967, vol. 2, no. 2, pp. 230–241.Chernov, Yu.I., Main Synecological Characteristics of SoilInvertebrates and Methods of Their Analysis, Metody poch-venno-zoologicheskikh issledovanii (Methods of Soil-Zoo-logical Studies), Moscow: Nauka, 1975, pp. 217–241.Couteaux, ˇ M.-M., Dynamisme de l’Équilibre des Thé-camoebiens dans Quelques Sols Climaciques, Memoires duMuseum National d’Hystoire Naturelle, 1976, vol. 96,pp. 44–79.Decloitre, L., Le Genre Euglypha Dujardin, Arch. F. Protis-tenk, 1962, vol. 106, no. 1, pp. 18–33.Deflandre, G., Le Genre Centropyxis Stein, Arch. Protistenk,1929, vol. 67, pp. 322–375.Dobrovol’skii, G.V. and Urusevskaya, I.S., Geografiya pochv(The Geography of Soils), Moscow: Mosk. Gos. Univ., 1984,pp. 1–415.Dobrovol’skii, G.V., Nikitin, E.D., and Afanas’eva, T.V.,Taezhnoe pochvoobrazovanie v kontinental’nykh usloviyakh(Taiga Soil Formation in Continental Conditions), Moscow:Mosk. Gos. Univ., 1981.Dolgova, L.S. and Gavrilova, I.P., Specific Soil Properties inthe Middle and Northern Taiga of Western Siberia (within theLimits of Tyumen Region), Prirodnye usloviya ZapadnoiSibiri (Natural Conditions of Western Siberia), Moscow:Mosk. Gos. Univ., 1971, no. 1, pp. 77–90.Gavrilova, I.P. and Dolgova, L.S., Sandy Podzols in the Mid-dle Taiga Subzone of Western Siberia, Prirodnye usloviyaZapadnoi Sibiri (Natural Conditions of Western Siberia),1972, no. 2, pp. 34–50.Gel’tser, Yu.G., Korganova, G.A., and Alekseev, D.A., Testa-ceans (Testacida, Protozoa) in Soil, Genezis i ekologiya

BIOLOGY BULLETIN Vol. 29 No. 6 2002
TESTACEANS (TESTACEA, PROTOZOA) OF TAIGA SOILS 627
pochv Tsentral’no-lesnogo gosudarstvennogo zapovednika(Genesis and Ecology of Soil in the Central Forest StateReserve), Moscow: Nauka, 1979, pp. 197–229.Gel’tser, Yu.G., Korganova, G.A., Yakovlev, A.S., and Alek-seev, D.A., Soil testaceans Testacida, Pochvennye prosteishie(Soil Protozoans), Leningrad: Nauka, 1980, pp. 108–142.Gel’tser, Yu.G., Korganova, G.A., and Alekseev, D.A., Opre-delitel’ pochvoobitayushchikh rakovinnykh ameb (Key toSoil Inhabiting Testacea), Moscow: Mosk. Gos. Univ., 1995.Klassifikatsiya i diagnostika pochv SSSR (Classification andDiagnostics of Soils of the USSR), Moscow: Kolos, 1977.Korde, N.V. and Chibisova, O.I., Soil Micropopulation of theMain Forest Types in the Moscow Region—Testacida, Leso-vodstvennye issledovaniya v Serebryanoborskom opytnomlesnichestve (Forestry Studies in the Serebryanyi Bor Exper-imental Forestry), Moscow: Nauka, 1973, pp. 167–174.Korganova, G.A., Testaceans (Protozoa, Testacida) as aMarker of Soil Conditions, Zoologiya bespozvonochnykh,(Zoology of Invertebrates), 1978, pp. 153–176.Korganova, G.A., Testacea in Soils Coniferous-Broad-leavedForests as an Index of Environmental Features, Cand. Sc.(Biol.) Dissertation, Moscow, 1979.Korganova, G.A., Pochvennye bespozvonochnye Moskovskoiobl. (Testaceans (Testacida) of Forest Soils in MoscowRegion), Moscow: Nauka, 1982, pp. 25–41.Korganova, G.A., Fauna and Ecology of (Protozoa, Testacea)from Forest Soils in Eastern Europe, Issledovaniya strukturyzhivotnogo naseleniya pochv (Studies of Animal PopulationStructure of Soil), Moscow: Nauka, 1994, pp. 85–102.Korganova, G.A., Soil Testacea (Protozoa, Testacea): Fauna,Ecology, and Principles of Organization of the Communities,Doctoral (Biol.) Dissertation, Moscow, 1997.Korganova, G.A. and Rakhleeva, A.A., Testacea in Soils ofMeshchera Lowland, Zool. Zh., 1997, vol. 76, no. 3, pp. 261–268.Kozlovskaya, L.S., Rol’ bespozvonochnykh v transformatsiiorganicheskogo veshchestva bolotnykh pochv (Role of SoilInvertebrates in the Transformation of Organic Matter in BogSoils), Leningrad: Nauka, 1976.
Liss, O.L. and Berezina, N.A., Bolota Zapadno-Sibirskoiravniny (Bogs of West Siberian Plain), Moscow: Mosk. Gos.Univ., 1981.Lousier, D., Taxonomy and Ecology of Testacea (Rhizopoda)in Rocly Mountain Soils (Kananaskis Vallay, Alberta, Can-ada), M. Sc. Thesis, Univ. Calgary, Canada, 1972.Megarran, E., Ekologicheskoe raznoobrazie i ego izmerenie(Eclogical Diversity and How to Measure It), Moscow: Mir,1992.Ogden, C.G. and Hedley, R., An Atlas of Freshwater TestateAmoebae, British Museum (Natur. Hist.), 1980.Pielou, E.C., An Introduction to Mathematical Ecology, NewYork, 1969.Rakhleeva, A.A., Changes in Structure and Diversity ofComplexes of Soil Testacea (Protozoa) in Mesorelief Ele-ments of the Southern Meshchera, Izv. Ross. Akad. Nauk. Ser.Biol., 1998, no. 6, pp. 749–754.Rakhleeva, A.A., Changes of Testacean Complexes (Proto-zoa) in a Sequence of Flood Plain Soils of the Oka State Bio-sphere Reserve (Southern Meshchera), Izv. AN. Ser. Biol,1999, no. 4, pp. 478–487.Rakhleeva, A.A., Specific Spatial Distribution of Testaceans(Testacea, Protozoa) on Plain Landscape (at the Example ofSouthern Meshchera), Cand. Sc. (Biol.) Dissertation, Mos-cow, 2000.Schönborn, W., Beschalten Amoben (Testacea). Wittenberg,1966.Shannon, C.E. and Weaver, W., The Mathematical Theory ofCommunication, Urbana, 1963.Simpson, E.H., Measurement of Diversity, Nature, 1949,vol. 163, p. 688.Thomas, R., Thecamoebiens de la Region Bordelaise, Bull.Soc. Hist. Natur. Toulouse, 1954, vol. 88, nos. 3–4, pp. 245–265.Whittaker, R.H., Evolution of Species Diversity in LandCommunities, Evolutionary Biology, New York: Plenum,1977, vol. 10, pp. 1–67.





























