Embed Size (px)
Citation preview

Crop Protection 23 (2004) 229–234
ARTICLE IN PRESS
*Correspondin
622, Uppsala S-7
17-14-94.
E-mail addres
0261-2194/$ - see
doi:10.1016/j.cro
Temporal spread of wheat dwarf virus and mature plant resistancein winter wheat
Mats Lindblad*, Roland Sigvald
Department of Ecology and Crop Production Science, Swedish University of Agricultural Sciences, P.O. Box 7043, Uppsala, S-750 07, Sweden
Received 7 May 2003; received in revised form 11 July 2003; accepted 29 August 2003
Abstract
Wheat dwarf is a potentially severe disease of winter wheat. The causal agent is wheat dwarf virus (WDV), a geminivirus
transmitted by the leafhopper Psammotettix alienus. Field surveys of WDV incidence and vector occurrence were conducted for 4
years in winter wheat fields in central Sweden. The objective was to estimate the extent of primary spread of WDV in autumn, by
adult leafhoppers migrating into the fields, and the secondary spread in the following early summer, by the progeny of the
immigrants. We also related the rate of virus spread in early summer to the phenological development of the vector and the host
plant. Furthermore, the development of mature plant resistance was assessed by inoculating winter wheat plants in various growth
stages with infectious leafhoppers. Field survey data indicate that the rate of primary infections did not exceed 5%, and that
infection rates increased to up to 50% during the summer. Infection rates ceased to increase as the crops reached the end of stem
elongation, and virus transmission experiments showed that plants became resistant to infection at growth stage DC 31 (first node
detectable).
r 2003 Elsevier Ltd. All rights reserved.
Keywords: Primary and secondary infection; Resistance; Psammotettix alienus; WDV
1. Introduction
Understanding the process of virus transmission byinsect vectors into a crop, and the secondary spreadfrom primary infected to healthy plants, is fundamentalin plant-virus disease epidemiology. Most plants aresusceptible to virus infection only during certain growthstages, and the extent of virus spread depends on theoccurrence and activity of infectious vectors during thisperiod. Hence, it is essential to estimate host plantsusceptibility in relation to the phenological develop-ment of the plant. Moreover, to develop appropriatecontrol strategies and forecasting methods, the relativeimportance of primary and secondary infections needsto be ascertained.Wheat dwarf is a serious virus disease of winter
wheat. Typical symptoms are yellowing and severedwarfing. Early sown crops are most exposed to damage
g author. National Food Administration, PO Box
51 26, Sweden. Tel.: +46-18-17-56-95; fax: +46-18-
s: [email protected] (M. Lindblad).
front matter r 2003 Elsevier Ltd. All rights reserved.
pro.2003.08.011
(Felix et al., 1992; Praslicka, 1996; Lindsten andLindsten, 1999). Diseased plants often occur in patchesin the fields but infections may, in the worst case, resultin crop failure. The disease is relatively uncommon, butduring the last decade it has attracted much attention inseveral European countries (Bisztray et al., 1989; Vacheret al., 1992; Huth, 2000). In Sweden, severe outbreaksoccurred early in the last century and in the 1940s. Afterthese reports, the disease was rare up to 1997 whenmany winter wheat fields in central Sweden sufferedconsiderable damage (Lindblad and Waern, 2002).Wheat dwarf virus (WDV) is a geminivirus, trans-
mitted by the leafhopper Psammotettix alienus (Dahl-bom) (Homoptera: Cicadellidae) (Vacke, 1961; Lindstenet al., 1980). Apart from winter wheat, different cereal,ley and weed grass species may serve as alternative hostplants (Vacke, 1972; Lindsten and Vacke, 1991;Lindsten and Lindsten, 1993; Vacke and Cibulka,1999). In common with other geminiviruses (Conti,1985), WDV is transmitted in a persistent, non-propagative manner. It does not multiply within theinsect and it is not transmitted to the eggs. The vector,P. alienus, is a Holarctic species that commonly occurs

ARTICLE IN PRESSM. Lindblad, R. Sigvald / Crop Protection 23 (2004) 229–234230
in grasslands and arable fields (Schiemenz, 1969;Raatikainen and Vasarainen, 1973; Lindblad and Aren-.o, 2002). In northern Europe, it has two generations peryear and overwinters in the egg stage (Schiemenz, 1969).The nymphs hatch in spring and pass five developmentalstages before the adult stage is reached in early summer(Giustina et al., 1991; Guglielmino and Virla, 1997).Adults of the second generation start to appear in latesummer and are present until late autumn (Lindbladand Aren .o, 2002).The primary spread of WDV to winter wheat fields
takes place in autumn, when adult leafhoppers migrateinto newly sown fields. Incidence may range from a fewpercent up to about 75% (Fohrer et al., 1992). It has notbeen established to what extent a secondary spread ofvirus occurs in spring, when leafhopper nymphs hatchand may acquire virus from plants infected in theprevious autumn. Studies concerning the relationbetween the phenological development of winter wheatand changes in susceptibility to WDV are also scarce,although Vacke (1972) reported that plants are moresusceptible at the one leaf stage than in later stages. Inthe present study, we estimated the relative extent ofprimary and secondary WDV infections in winter wheatfields in Sweden. We also related the rate of virus spreadduring early summer to the phenological development ofthe vector and the host plant. Estimates of mature plantresistance obtained from field surveys were validated ingreenhouse experiments, in which WDV was trans-mitted by leafhoppers to winter wheat plants at variousgrowth stages.
2. Material and methods
2.1. Field surveys
Field surveys of WDV incidence and vector occur-rence were carried out in winter wheat fields in theprovince of Uppland in central Sweden from 1998 to2000, and in the provinces of Uppland and .Osterg .otlandin 2001. Early sown fields, i.e. fields sown at thebeginning of September, were selected. The predominat-ing cultivar was ‘Kosack’, which is susceptible to WDV(Lindblad and Waern, 2002). None of the fields weretreated with insecticides. Eight fields (sown in theprevious year) were surveyed in 1998, five in 1999, sixin 2000 and 11 in 2001. In 2000, samples were alsocollected from an experimental plot (100� 200m2) sownat the end of August in the previous year. Virusincidences were estimated by sampling one leaf fromeach of five plants taken at 20 places along a diagonal ineach field (total 100 plants/field).The top leaf of the main shoot was collected from
plants in early growth stages, but after heading nodistinction was made between main shoots and tillers.
The first sample in each field was assumed to representthe primary infection rate, and was either collected latein November (fields surveyed in 1999–2001), after theactivity of the leafhoppers in the fields had ceased, orearly in May (fields surveyed in 1998), before anynymphs had started to hatch. In 1998–2000 the finalinfection rates were estimated by sampling the fields atthe end of June or in the middle of July. In 2001, finalsamples were only collected from three fields in whichWDV was detected in the first sample. In addition, mostfields surveyed in 1998 were sampled every second weekfrom the beginning of May to the middle of July, inorder to estimate the rate of increase of virus incidenceduring the growing season. In 2000, samples werecollected from the experimental plot two times a weekfrom 10 May to 3 July.Each sampled leaf was tested serologically for the
presence of WDV by double antibody sandwich enzyme-linked immunosorbent assay (das-ELISA), according tothe method described by Clarke and Adams (1977) withonly minor changes. Each extract was tested once (onewell/extract) using an antiserum against a Swedishisolate of WDV (see Lindsten and Lindsten, 1999). Toavoid non-specific reactions, 2 g polyvinylpyrrolidonewas added to 100ml extraction buffer. Absorbancevalues at 405 nm were read 2 h after addition ofsubstrate. Readings for negative controls (non-infectedwheat sap) ranged from 0.06 to 0.11, with a mean valueof 0.08. Leaf samples with absorbance values higherthan 0.3 were classified as positive, but readings forpositive samples were most often significantly higher(mean 2.1, maximum 3.5).The occurrence of adult and nymphal P. alienus was
monitored by using water traps, consisting of yellow,round metal trays (diameter 24 cm, depth 7 cm) filledwith water containing detergent. At the beginning ofMay, one trap was placed on the soil about 30m fromthe field edge in each surveyed field (see Lindblad andAren .o, 2002, for details concerning the placement andchoice of number of traps per field). Traps were emptiedone or two times a week and monitoring was continuedfor at least 4 weeks from the time of first occurrence ofnymphs.
2.2. Greenhouse experiments
Two virus transmission experiments were performedin the winter 2000–2001. In the first experiment, winterwheat plants at growth stages DC 11 (one leaf unfolded,Tottman and Broad, 1987), DC 30 (pseudostem erect),DC 32 (second node detectable), and DC 41 (flag leafsheath extending) were exposed to viruliferous P.
alienus. The experiment was repeated with plants ingrowth stages DC 11, DC 21 (first side shoot emerged),DC 30, DC 31 (first node detectable), and DC 32.

ARTICLE IN PRESS
Table 1
Initial and final incidences of WDV and vector occurrence in winter
wheat fields surveyed in central Sweden 1998–2001
Year Field Infected plants (%) No. of P. alienus
Initial Final Nymphs Adults
1998 A 2 0 14 0
B 3 4 17 0
C 5 12 59 6
D 5 29 13 0
E 5 47 32 0
M. Lindblad, R. Sigvald / Crop Protection 23 (2004) 229–234 231
A cultivar (Ebi) known to be very susceptible to WDV(Vacke and Cibulka, 2000) was used.
Psammotettix alienus leafhoppers were collected fromwinter wheat fields in August 2000 and reared in cages ina greenhouse at 20�C and 16 h light/8 h darkness (L:16/D:8). Young winter wheat plants were used as foodplants. Serological tests showed that all food plants hadbecome infected by WDV about 6 weeks after thecollection of leafhoppers. Food plants were replacedcontinuously as plants died, but repeated ELISA-testsshowed that new plants also became infected. Adultleafhoppers from the progeny of the field-collectedinsects, raised on the WDV infected plants, were usedfor virus transmission in November and December 2000(first and second experiment, respectively).Winter wheat kernels were laid on moist filter paper
and placed in a climate chamber for vernalization at atemperature of 4�C and L:6/D:18. After 2–3 weeks,seedlings were planted separately in pots. After 3–4weeks the first batch of four plants was brought into agreenhouse and reared at 15�C and L:8/D:16. There-after, batches of plants were brought into the green-house every week until plants in growth stages DC 11 toDC 41 were obtained. Four plants of each growth stageincluded in the experiments were then caged, and twoleafhoppers were placed on each plant. The insects wereinfused to plants of all growth stages at the same day ineach experiment. All tillers were removed to ensurefeeding on the main shoots. The plants were kept at20�C and L:16/D:8 until the leafhoppers were removedafter 6 days. At that time, new tillers had in some casesstarted to develop. The plants were sprayed with aninsecticide and placed at 18�C and L:16/D:8.One leaf was sampled from the main shoot of each
plant every second week for 6–8 weeks after the timewhen leafhoppers were placed on the plants. At eachsampling, most of the top leaf was collected from shootsin early growth stages. To allow for repeated sampling,only part of the top leaf was cut on shoots where the flagleaf had emerged. In the second experiment, the threefirst tillers of each plant were sampled, except for plantsinoculated at DC 11 which did not produce tillers. Eachsampled leaf was tested by ELISA as described above.Symptoms of disease were noted 8 weeks after inocula-tion. Shoots on which the ear failed to emergecompletely, or did not develop at all, were classified asdiseased.
2000 F� 1 12 23 164
2001 G 1 0 13 0
H 1 30 68 27
I 2 0 20 1
�Experimental plot.Note: Data are from nine fields where primary infected plants were
found. In all, 100 plants were tested by ELISA at each sampling
occasion. Numbers of P. alienus are total catches in water traps during
4 weeks from the time of first occurrence of nymphs.
3. Results
3.1. Field surveys
Primary infected plants were found in five of the eightfields surveyed in 1998 and in five out of 11 fields in2001. No infected plants were found in 1999, and in
2000 infected plants were found in the experimental plotbut not in any other winter wheat field. The rates ofinfections at the initial sampling occasions were low (0–5%), whereas high incidences of WDV (up to about50%) occurred in some fields at the final samplingoccasions (Table 1).The final rates of infected plants were, to some extent,
related to the first estimates of WDV incidence.Infection rates of at least 10% were noted in five outof nine fields in which WDV was detected with the firstELISA-test (Table 1). The final virus incidences in fieldswhere no infected plants were found at the first samplingoccasion did not exceed 1% (data from 14 fields in1998–2000). There was no clear relation betweenleafhopper occurrence and virus incidence, but the fieldin which most leafhoppers were caught in 2001 alsobecame most infected (Table 1).The increase in WDV incidence mainly occurred
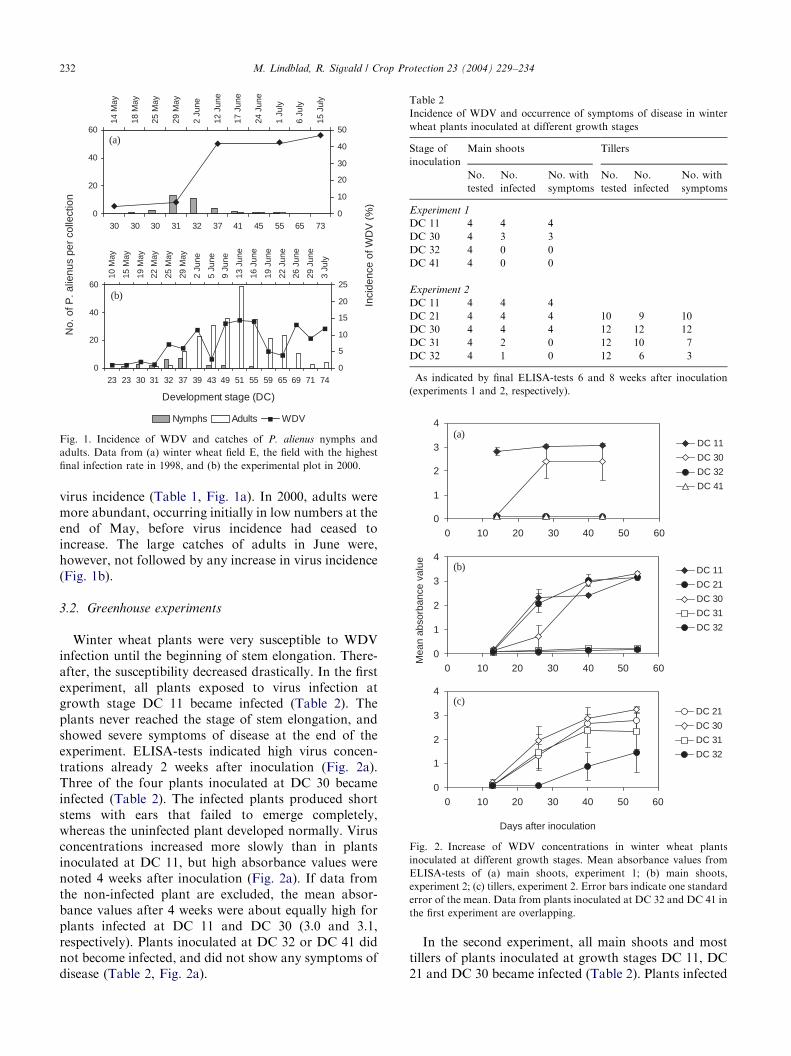
during the period of stem elongation. In 1998, virusincidence increased rapidly at the beginning of June.Thereafter, the percentage of infected plants increasedvery little (Fig. 1a). Similarly, in 2000, WDV incidenceincreased during a limited period at the end of May.Virus incidences in later samples varied, but did notsignificantly exceed the level estimated at the beginningof June (Fig. 1b). Thus, in both years the percentage ofwinter wheat plants testing positively for WDV ceasedto increase in samples taken later than at the end of stemelongation (DC 37–39).Water trap catches suggest that mainly nymphs were
present during the periods of virus spread, whereasadults occurred later in the season. In 1998, nymphswere caught in all surveyed fields whereas adults wererare. Except for in one field, no adults were caught untilthe middle of June, i.e. after the period of increase in
ARTICLE IN PRESS
0
20
40
60
30 30 30 31 32 37 41 45 55 65 73
0
10
20
30
40
50
14 M
ay
18 M
ay
25 M
ay
29 M
ay
2 Ju
ne
12 J
une
17 J
une
24 J
une
1 Ju
ly
6 Ju
ly
15 J
uly
(a)
0
20
40
60
23 23 30 31 32 37 39 43 49 51 55 59 65 69 71 74
Development stage (DC)
0
5
10
15
20
25
10 M
ay
15 M
ay
19 M
ay
22 M
ay
25 M
ay
29 M
ay
2 Ju
ne
5 Ju
ne
9 Ju
ne
13 J
une
16 J
une
19 J
une
22 J
une
26 J
une
29 J
une
3 Ju
ly
Nymphs Adults WDV
(b)
No.
ofP
. alie
nus
per
colle
ctio
n
Inci
denc
e of
WD
V (
%)
Fig. 1. Incidence of WDV and catches of P. alienus nymphs and
adults. Data from (a) winter wheat field E, the field with the highest
final infection rate in 1998, and (b) the experimental plot in 2000.
Table 2
Incidence of WDV and occurrence of symptoms of disease in winter
wheat plants inoculated at different growth stages
Stage of
inoculation
Main shoots Tillers
No.
tested
No.
infected�No. with
symptoms
No.
tested
No.
infected�No. with
symptoms
Experiment 1
DC 11 4 4 4
DC 30 4 3 3
DC 32 4 0 0
DC 41 4 0 0
Experiment 2
DC 11 4 4 4
DC 21 4 4 4 10 9 10
DC 30 4 4 4 12 12 12
DC 31 4 2 0 12 10 7
DC 32 4 1 0 12 6 3
�As indicated by final ELISA-tests 6 and 8 weeks after inoculation
(experiments 1 and 2, respectively).
Mea
n ab
sorb
ance
valu
e
0
1
2
3
4
0 10 20 30 40 50 60
DC 11
DC 21
DC 30
DC 31
DC 32
(b)
0
1
2
3
4
0 10 20 30 40 50 60
DC 21
DC 30
DC 31
DC 32
(c)
Days after inoculation
0
1
2
3
4
0 10 20 30 40 50 60
DC 11
DC 30
DC 32
DC 41
(a)
Fig. 2. Increase of WDV concentrations in winter wheat plants
inoculated at different growth stages. Mean absorbance values from
ELISA-tests of (a) main shoots, experiment 1; (b) main shoots,
experiment 2; (c) tillers, experiment 2. Error bars indicate one standard
error of the mean. Data from plants inoculated at DC 32 and DC 41 in
the first experiment are overlapping.
M. Lindblad, R. Sigvald / Crop Protection 23 (2004) 229–234232
virus incidence (Table 1, Fig. 1a). In 2000, adults weremore abundant, occurring initially in low numbers at theend of May, before virus incidence had ceased toincrease. The large catches of adults in June were,however, not followed by any increase in virus incidence(Fig. 1b).
3.2. Greenhouse experiments
Winter wheat plants were very susceptible to WDVinfection until the beginning of stem elongation. There-after, the susceptibility decreased drastically. In the firstexperiment, all plants exposed to virus infection atgrowth stage DC 11 became infected (Table 2). Theplants never reached the stage of stem elongation, andshowed severe symptoms of disease at the end of theexperiment. ELISA-tests indicated high virus concen-trations already 2 weeks after inoculation (Fig. 2a).Three of the four plants inoculated at DC 30 becameinfected (Table 2). The infected plants produced shortstems with ears that failed to emerge completely,whereas the uninfected plant developed normally. Virusconcentrations increased more slowly than in plantsinoculated at DC 11, but high absorbance values werenoted 4 weeks after inoculation (Fig. 2a). If data fromthe non-infected plant are excluded, the mean absor-bance values after 4 weeks were about equally high forplants infected at DC 11 and DC 30 (3.0 and 3.1,respectively). Plants inoculated at DC 32 or DC 41 didnot become infected, and did not show any symptoms ofdisease (Table 2, Fig. 2a).
In the second experiment, all main shoots and mosttillers of plants inoculated at growth stages DC 11, DC21 and DC 30 became infected (Table 2). Plants infected

ARTICLE IN PRESSM. Lindblad, R. Sigvald / Crop Protection 23 (2004) 229–234 233
at DC 11 showed the same severe symptoms as thosefrom the first experiment. Absorbance readings werevery low 2 weeks after inoculation, but after 4 weekshigh absorbance values were noted (Fig. 2b). Absor-bance values for main shoots of plants exposed to virustransmission at DC 30 increased slower than for thoseexposed in earlier stages, but after 6 weeks there were nosignificant differences (Fig. 2b). Virus concentrations intillers increased in a similar way, but the difference in therate of increase between plants inoculated at DC 21 andDC 30 was small (Fig. 2c). Absorbance values for mainshoots of plants inoculated at DC 31 and DC 32increased only slightly (Fig. 2b), although the finalELISA-test indicated that some of them became infected(Table 2). Infected shoots did, however, not show anysymptoms and the final absorbance values wererelatively low (between 0.3 and 0.5). Virus concentra-tions in tillers increased more rapidly, and becamehigher, than in main shoots (Fig. 2c). Most infectedtillers failed to produce ears, but there were also tillersthat developed normally in spite of high virus concen-trations (absorbance values between 1 and 3) in the finalELISA-test.WDV concentrations in the first tillers, which in some
cases were exposed to virus transmission by leafhoppers,did not exceed the levels in later developed tillers.Absorbance values actually tended to be highest in thethird tillers. Eight weeks after inoculation, the meanabsorbance values of the first, second and third tillerswere 2.2, 2.5 and 2.8, respectively. The differenceswere not statistically significant (ANOVA, F2;43 ¼0:7;P > 0:05).
4. Discussion
Our study indicates that the primary spread of WDVin autumn, by adult leafhoppers migrating into winterwheat fields, resulted in relatively low infection rates.The low extent of primary infections is in contrast withprevious studies from France (Fohrer et al., 1992),indicating that the epidemiology of wheat dwarf maydiffer between Sweden and other countries in Europe.The window for possible spread of WDV is narrow,which may be one reason for the sporadic occurrence ofwheat dwarf outbreaks. In autumn, the time whenleafhoppers are active and can transmit virus to newlysown winter wheat fields is limited by decreasingtemperatures. High numbers of P. alienus can be caughtin water traps as long as daily maximum temperaturesexceed 15�C, whereas almost none occur at tempera-tures below 10�C (Lindblad and Aren .o, 2002). Hence,wheat dwarf mainly occurs in early sown winter wheatcrops, which emerge when daily temperatures still arerelatively high. In spring and early summer, spread of
virus can only occur from the time when leafhopper eggsstart to hatch until the crops become resistant.Our results suggest that nymphal P. alienus accounted
for most of the secondary spread, since the increase invirus incidences occurred before adults started tobecome abundant. Although the nymphs are wingless,they are very mobile already in early developmentstages. In fields with dense populations, nymphs caneasily be observed jumping from plant to plant and intowater traps (Lindblad and Aren .o, 2002). An alternativeexplanation of the increase of WDV incidence duringsummer is that plants became infected in autumn butvirus infections could not be detected until new leavesdeveloped during early summer. Difficulties in detectingvirus antigen in inoculated leaves have been reported formaize streak virus, another geminivirus (Peterschmittet al., 1992). Inoculation trials have, however, shownthat WDV can be detected by ELISA in an inoculatedleaf 1 week after virus transmission, even if the winterwheat plants are kept at a low temperature (8�C)(Sandgren, M., unpublished).The results of our field surveys indicate that winter
wheat plants became resistant to WDV infection at thestem elongation. Similarly, greenhouse experimentsshowed that plants were susceptible until growth stageDC 30, and that resistance of the main shoot increasedmarkedly in later stages. The results indicate that tillersof older plants were still susceptible, but this may be anartifact due to the experimental design. All tillers of theexperimental plants were removed as leafhoppers wereintroduced into the cages, which resulted in a regenera-tion of small, and presumably more susceptible, tillerson all plants. Consequently, it is likely that earlydeveloping cultivars, which reach the resistant growthstage earlier in summer than late cultivars, are less likelyto become infected. This is corroborated by field trialdata, which indicate lower disease incidence in two earlydeveloping cultivars (Tarso and Pagode) than in thecultivar ‘Kosack’ which develops more slowly (Lindbladand Waern, 2002).Treating winter wheat seeds with an insecticide can
control wheat dwarf, whereas the effect of insecticidesprays in autumn varies (Vacher et al., 1992). Adisadvantage of seed treatments is, however, thatthis control measure has to be performed in apreventative way and is difficult to adjust to the actualneed. We suggest that control of newly hatched nymphsin early summer may be an alternative way to preventwheat dwarf damage, at least in regions such as centralSweden where high infection rates do not come up untilafter the secondary spread of WDV. Control of nymphsprovides better possibilities to forecast outbreaks ofwheat dwarf, by, for example, monitoring leafhopperimmigration to winter wheat fields in autumnand estimating the extent of primary infections byserological tests.

ARTICLE IN PRESSM. Lindblad, R. Sigvald / Crop Protection 23 (2004) 229–234234
Acknowledgements
We thank Jonathan Yuen and anonymous reviewersfor their constructive comments on earlier drafts of themanuscript. This study was supported by grants fromthe Swedish Farmer’s Foundation for AgriculturalResearch and the Swedish Board of Agriculture.
References
Bisztray, G., Gaborjanyi, R., Vacke, J., 1989. Isolation and
characterisation of wheat dwarf virus found for the first time in
Hungary. Z. Pflanzenkr. Pflanzenschutz 96, 449–454.
Clarke, M.F., Adams, A.N., 1977. Characteristics of the microplate
method of enzymelinked immunosorbent assay for the detection of
plant virus. J. Gen. Virol. 34, 475–483.
Conti, M., 1985. Transmission of plant viruses by leafhoppers and
planthoppers. In: Nault, L.R., Rodriguez, J.G. (Eds.), The
Leafhoppers and Planthoppers. Wiley, New York, pp. 289–307.
Felix, I., Larcher, J.M., Maraby, J., Philippeau, G., Vinatier, K., 1992.
Risques d’attaques de cicadelles et conditions d’efficacit!e des
insecticides. Perspect. Agric. 173, 98–106.
Fohrer, F., Lebrun, I., Lapierre, H., 1992. Acquisitions rec!ente sur le
virus du nanisme du bl!e. Phytoma 443, 18–20.
Giustina, W.D., Lebrun, I., Lapierre, H., Lochon, S., 1991. Distribu-
tion g!eographique du vecteur et du virus. Phytoma 432, 30–34.
Guglielmino, A., Virla, E.G., 1997. Postembryonic development and
biology of Psammotettix alienus (Dahlbom) (Homoptera, Cicadel-
lidae) under laboratory conditions. Boll. Zool. Agr. Bachic. 29,
65–80.
Huth, W., 2000. Viruses of Graminae—a short overview. J. Plant Dis.
Prot. 107, 406–414.
Lindblad, M., Aren .o, P., 2002. Temporal and spatial population
dynamics of Psammotettix alienus, a vector of wheat dwarf virus.
Int. J. Pest Man. 48, 233–238.
Lindblad, M., Waern, P., 2002. Correlation of wheat dwarf incidence
to winter wheat cultivation practices. Agric. Ecosys. Environ. 92,
115–122.
Lindsten, K., Lindsten, B., 1993. Occurrence and transmission of
wheat dwarf virus (WDV) in France. 3 Conference Internationale
sur les ravageurs en agriculture, Montpellier, pp. 41–48.
Lindsten, K., Lindsten, B., 1999. Wheat dwarf—an old disease with
new outbreaks in Sweden. J. Plant Dis. Prot. 106, 325–332.
Lindsten, K., Vacke, J., 1991. A possible barley adapted strain of
wheat dwarf virus (WDV). Acta Phytopatol. Entomol. Hung. 26,
175–180.
Lindsten, K., Lindsten, B., Abdelmoeti, M., Junti, N., 1980.
Purification and some properties of wheat dwarf virus. Third
Conference on Virus Diseases of Graminae in Europe, Rotham-
stead, pp. 27–30.
Praslicka, J., 1996. Vplyv niektorych faktorov na napadnutie psenice
ozimnej virusovou zakrpatenostou psenice (WDV). Ochr. Rostlin
UZPI 32, 181–186.
Peterschmitt, M., Quiot, J.B., Reynaud, B., Baudin, P., 1992.
Detection of maize streak virus antigens over time in different
parts of maize plants of a sensitive and a so-called tolerant cultivar
by ELISA. Ann. Appl. Biol. 121, 641–653.
Raatikainen, M., Vasarainen, A., 1973. Early- and high-summer flight
periods of leafhoppers. Ann. Agric. Fenn. 12, 77–94.
Schiemenz, H., 1969. Die Zikadenfauna mitteleurop.aischer Trock-
enrasen (Homoptera, Auchenorrhyncha). Entomol. Abh. Mus.
Tierk. Dresden 36, 201–280.
Tottman, D.R., Broad, H., 1987. The decimal code for the
growth stages of cereals, with illustrations. Ann. Appl. Biol. 110,
441–454.
Vacher, C., Felix, I., Larcher, J.M., Lochon, S., Pillon, M., Stoop, P.,
1992. Lutte contre le nanisme du bl!e: le point apr"es deux ann!ees
d’exp"erimentation. Perspect. Agric. 173, 89–97.
Vacke, J., 1961. Wheat dwarf virus disease. Biol. Plant. 3,
228–233.
Vacke, J., 1972. Host plants range and symptoms of wheat dwarf
virus. Ved. Pr. Vyzk. Ustavu Rostl. Vyroby Praha-Ruzyne 17,
151–162.
Vacke, J., Cibulka, R., 1999. Silky bent grass (Apera spica-venti [L.]
Beauv.)—a new host and reservoir of wheat dwarf virus.
Chundelka Metlice 35, 47–50.
Vacke, J., Cibulka, R., 2000. Response of selected winter wheat
varieties to wheat dwarf virus infection at an early growth stage.
Czech J. Gen. Plant Breeding 36, 1–4.





























