Embed Size (px)
Citation preview

lable at ScienceDirect
Food Control xxx (2016) 1e10
Contents lists avai
Food Control
journal homepage: www.elsevier .com/locate/ foodcont
Targeting double genes in multiplex PCR for discriminating bovine,buffalo and porcine materials in food chain
M.A. Motalib Hossain a, Md. Eaqub Ali a, b, *, Sharifah Bee Abd Hamid a, Asing a,Shuhaimi Mustafa c, Mohd Nasir Mohd Desa c, I.S.M. Zaidul d
a Nanotechnology and Catalysis Research Centre (NANOCAT), University of Malaya, Kuala Lumpur 50603, Malaysiab Centre for Research in Biotechnology for Agriculture (CEBAR), University of Malaya, Kuala Lumpur 50603, Malaysiac Institute of Halal Products Research, Universiti Putra Malaysia, 43400 UPM, Serdang, Selangor, Malaysiad Deparment of Pharmaceutical Technology, Faculty of Pharmacy, International Islamic University, Kuantan 25200, Pahang, Malaysia
a r t i c l e i n f o
Article history:Received 14 April 2016Received in revised form18 July 2016Accepted 2 August 2016Available online xxx
Keywords:Multiplex PCRDouble gene-targetsShort-length DNA targetsEconomicReligious and health concernDecomposed specimensMeatball products
* Corresponding author. Nanotechnology and CatalCAT), University of Malaya, Kuala Lumpur 50603, Ma
E-mail addresses: [email protected], eaqubali@
http://dx.doi.org/10.1016/j.foodcont.2016.08.0080956-7135/© 2016 Elsevier Ltd. All rights reserved.
Please cite this article in press as: Hossain, Mmaterials in food chain, Food Control (2016)
a b s t r a c t
Beef, buffalo and pork are the major meat of economic, religious and health concern. Current methods toauthenticate these materials in food chain are based on mainly single gene targets which are susceptibleto break down by food processing treatments. We, for the first time, described here a double gene tar-geting short-amplicon length multiplex polymerase chain reaction assay for discriminating bovine,buffalo and porcine materials in a single assay platform. The advantage of the assay is evidenced in termsof fidelity, cost and time since it is highly unlikely that two different targets would be missing even in adecomposed specimen. Detection of multiple targets in a single assay definitely saves analytical cost andtime. Mitochondrial cytochrome b (cytb) and ND5 genes were targeted and six different targets (length:90e146 bp), two for each of cow (120 and 106bp), buffalo (90 and 138bp) and pig (73 and 146bp), wereamplified from raw, boiled, autoclaved and microwaved cooked meat under pure and mixed matrices.The detection limit was 0.02 ng DNA under pure states and 0.1% meat in binary mixtures and meatballproducts. Screening of Malaysian meatball products revealed all beef products were buffalo positive inwhich 35% were totally replaced. In contrast, all pork products were found uncontaminated from beefand buffalo.
© 2016 Elsevier Ltd. All rights reserved.
1. Introduction
Authentication of animal origins of food products is a rapidlygrowing field having relevance to community health, religious andcultural issues and fair-trade policy (Bottero & Dalmasso, 2011;Haider, Nabulsi, & Al-Safadi, 2012). The recent inclusion of somealien species such as horse meat and pork in beef products(Walkera, Burnsb,& Burns, 2013; NEWS STRAITS TIMES, 2015), porkand rat meat in lamb products (Ali, Razzak, & Hamid, 2014), andmonkey, dog and cat meat in exotic menu (Amin, Hamid, & Ali,2016; Rahman et al., 2014; Rashid et al., 2015) are highly alarm-ing since most of these species are the potential carriers of variousinfecting agents and prohibited in the most religions and cultures.
Beef, buffalo and pork are the major meat of economic, health
ysis Research Centre (NANO-laysia.gmail.com (Md.E. Ali).
. A. M., et al., Targeting double, http://dx.doi.org/10.1016/j.fo
and religious interests heavily consumed all over the world. Forinstances, while there is a huge turnover of buffalo in India both fordomestic consumption and exports, the slaughtering, consumptionand export of beef is totally prohibited there because of the re-striction of Hinduism (Girish, Haunshi, Vaithiyanathan, Rajitha, &Ramakrishna, 2013). The recent killing of a man and beating ofhis familymembers in the city of Dadri in India by a group of radicalHindus based on a false rumour that beef was kept in his refriger-ator is a strong piece of evidence that meat scandal can provokesocial unrest and take innocent lives (Daily Mail Online, 2015).However, despite having religious prohibition and regulatory con-trol, the counterfeiting of buffalo meat with beef and beef withbuffalo has been rampant all over the world (Girish et al., 2013;Karabasanavar, Singh, Umapathi, Girish et al., 2011,Karabasanavar, Singh, Umapathi, Kumar et al., 2011). While theEgyptians and some Europeans prefer buffalo to beef because of thecultural preference and/or fear of bovine spongiform encephalop-athy (BSE) (Sakaridis, Ganopoulos, Argiriou, & Tsaftaris, 2013), beef
genes inmultiplex PCR for discriminating bovine, buffalo and porcineodcont.2016.08.008

Table 1Composition of model meatball products.
Ingredients Meat ball (�35 g/piece)
Beef Buffalo Pork
Minced meat 23a 23a 23a
Soy protein 3 3 3Starch/breadcrumb 5 5 5Chopped onion 1 1 1Chopped ginger 0.1 0.1 0.1Cumin powder 0.75 0.75 0.75Garlic power 0.5 0.5 0.5Black pepper 0.14 0.14 0.14Tomato paste 1.5 1.5 1.5Butter 1.5 1.5 1.5Salt SA SA SAOthersb SA SA SA
SA, suitable amounts.Note:
a 1%, 0.5% and 0.1% of beef, buffalo and pork meat were mixed with a balancedamount of respective minced meat to prepare �35 g meat ball specimen.
b Flavouring agents and enhancers.
M.A.M. Hossain et al. / Food Control xxx (2016) 1e102
has evolved as the meat of choice and consumed heavily in mostparts of the world (Karabasanavar, Singh, Umapathi, Girish et al.,2011, Karabasanavar, Singh, Umapathi, Kumar et al., 2011). On theother hand, pork is a popular meat in most of the western countriesin spite of its total unacceptability to the Muslims and Jewishconsumers (Ali et al., 2012). Thus the social, religious, health andbusiness interests in beef, buffalo and pork are huge and hencethere is a need of a trustworthy but low cost method for theirdiscrimination in food chain.
Currently, several molecular detection schemes are proposed forthe authentication of beef (Mane, Mendiratta, Tiwari, &Bhilegaokar, 2012, Mane, Mendiratta, & Tiwari, 2012; Herrero,Royo, Lago, Vieites, & Espi~neira, 2013), buffalo (Girish et al., 2013;Karabasanavar, Singh, Umapathi, Girish et al., 2011,Karabasanavar, Singh, Umapathi, Kumar et al., 2011) and pork(Erwanto, Abidin, Sugiyono, & Rohman, 2014). However, thesemethods are mostly based on a single and long DNA marker whichoften breaks down under food processing treatments as well asnatural and environmental decomposition, making them lesstrustworthy but more expensive (Ali, Asing Hamid, et al., 2015). Therecent evolution of multiplex polymerase change reaction (mPCR)assays are especially promising since they offer the opportunity ofidentifying multiple target oligos in a single assay platform, savingboth analytical cost and time (Ali, Razzak, et al., 2014). Instead oftargeting a single gene, double gene targeting short-ampliconlength mPCR assays would be more reliable and trustworthybecause of the complementation opportunities in which thedetection of an alternative target can compensate the missingtarget, in case it is lost under the states of decomposition. Recently,mPCR assays have been reported for pig, dog, cat, rat and monkeyspecies (Ali, Razzak, et al., 2015), beef, pork, horse and sheep spe-cies (K€oppel, Ruf, & Rentsch, 2011) and beef, pork, lamb, chicken,ostrich and horse meat (Kitpipit, Sittichan, & Thanakiatkrai, 2014).However, no mPCR assay has been documented for the differenti-ation of heavily consumed meat, namely, beef, buffalo and pork infood chain. Thus the objective of this study was to develop atrustworthy and cost-saving mPCR assay targeting two differentgene sites for the each of bovine, buffalo and porcine species totrace them in the food chain.
2. Methods and materials
2.1. Meat samples collection
Beef (Bos taurus), buffalo (Bubalus bubalis), goat (Capra hiscus),lamb (Ovis aries), chicken (Gallus gallus), duck (Anas platyrhychos),pigeon (Columba livia), quail (Coturnix coturnix)), cod (Gadusmorhua), salmon (Salmo salar), pangas (Pangasius pangasius), tuna(Thunnus orientalis), tilapia (Oreochromis niloticus) and rohu (Labeorohita), frog (Rana kunyuensis) and turtle (Cuora amboinensis)),were purchased in triplicate on three different dates from variouswet markets and supermarkets (Pasar Borong Selangor, Serdang,PuduWet Market, Kuala Lumpur and Tesco, Petaling Jaya, Selangor)across Malaysia. Pork (Sus scrofa) was purchased in triplicates fromthree different vendors from a Chinese wet market in Sri Kam-bangan, Selangor, Malaysia. Meat from three different euthanizeddogs (Canis lupus familiaris), cats (Felis catus) and rats (Rattus rattus)were collected from Dewan Bandaraya Kuala Lumpur (DBKL) asdescribed elsewhere (Ali, Razzak et al., 2015). Monkey (Macacafascicularis) meat was donated by the Wildlife Malaysia (Rashidet al., 2015). Commercial beef and pork meatballs of threedifferent brands were purchased from various stores outlets acrossMalaysia. All samples were collected in 2015 and transported underice-chilled (4� C) and were stored at �20� C until DNA extraction(Ali, Razzak et al., 2015).
Please cite this article in press as: Hossain, M. A. M., et al., Targeting doublematerials in food chain, Food Control (2016), http://dx.doi.org/10.1016/j.fo
2.2. Preparation of beef, buffalo and pork meatballs
Three types of ready-to-eat model meatballs of beef, buffalo andpork were prepared as per Razzak, Hamid, and Ali (2015) (Table 1).Meatball products were deliberately adulterated by spiking of 1%,0.5% and 0.1% of buffalo and pork, beef and pig, and beef and buffalomeat, respectively (Razzak et al., 2015). Thus prepared 0.1%contaminated meatballs were autoclaved at 121� C under 45-psipressure for 2.5 h (Rahman et al., 2014). All samples were storedat �20� C until DNA was extracted.
2.3. Thermal treatment of meat samples
To mimic the usual processing practices, the raw meat sampleswere subjected to three different types of heat treatments. The firstone was boiling, in which the beef, buffalo and pork meat wereboiled in water at 98� C on a hot plate for 90 min (Ali, Al Amin,Razzak, et al., 2015). To simulate a conventional canning processand steam cooking practices, the meat samples were autoclaved at121� C and 45 psi pressure for 20 min and 2.5 h (Ali, Razzak et al.,2015). Furthermore, meat samples were subjected to microwavecooking which is a quick and modern system of cooking at 500 W,600 W and 700 W for 30 min (Ali, Asing Hamid, et al., 2015). Alltreated samples were kept at �20� C until DNA extraction.
2.4. Extraction of DNA from raw meats and meatballs
DNA was extracted from meat and fish samples using YeasternGenomic DNA Mini Kit (Yeastern Biotech Co., Ltd, Taipei, Taiwan)(Rashid et al., 2015). Briefly, 20 mg of muscle tissues was groundedand homogenised with a micro pestle followed by the addition oflysis buffer and proteinase K. The mixturewas incubated at 60� C tolyse cells and proteins. A spin column was used for the attachmentof DNA to the glass fibre matrix under centrifugation. Ethanolcontaining wash buffer was used to remove potential contami-nants. The purified DNA was eluted in an elution buffer. DNA fromPlant species (wheat (Triticum aestivum), onion (Allium cepa), garlic(Allium sativum), ginger (Zingiber officinale) and pepper (Capsicumannuum)) was extracted using the DNeasy Plant Mini Kit (QIAGENGmgH, Hilden, Germany) (Ali, Razzak et al., 2015). DNA frommeatballs was extracted using NucleoSpin Food DNA kit(MACHEREY-NAGEL GmbH& Co. KG, Duren, Germany) (Hird, Lloyd,Goodier, Brown, & Reece, 2003). Concentration and purity of theextracted DNA were determined using UVevis Spectrophotometer
genes inmultiplex PCR for discriminating bovine, buffalo and porcineodcont.2016.08.008

Table 3Concentration of PCR components.
PCR dNTP (mM) MgCl2 (mM) Tag pol (unit) Primer (mM)
Duplex and Triplex 0.2 2.5 0.94 0.2e0.4Tetraplex 0.25 3.5 1.0 0.16e0.4Multiplex 0.25 4.0 1.25 0.12e0.6
M.A.M. Hossain et al. / Food Control xxx (2016) 1e10 3
(NanoPhotometer Pearl, Implen GmbH, Germany) based on theabsorbance at 260 nm and ratio at A260/A280, respectively(Napolitano et al., 2014).
2.5. Design of species-specific primers
Six sets of primers were designed targeting mitochondrial cytband ND5 genes because of their higher degree of divergence andavailability of sufficient conserved regions within the species butadequate polymorphism among the closely related species(Mohamad, El Sheikha, Mustafa, & Mokhtar, 2013; Razzak et al.,2015). The Sequences of cytb and ND5 genes of cow (V00654.1),buffalo (NC_006295.1) and pig (AF034253.1) species were retrievedfrom the National Centre of Biotechnology Information (NCBI)database andwere aligned using theMEGA5 alignment tool (http://www.megasoftware.net/) to identify the hyper-variable andconserved regions. Publicly available software Primer3Plus (http://www.bioinformatics.nl/cgi-bin/primer3plus/primer3plus.cgi) wasused to design the six sets (two sets for each species (one set fromcytb and another set from ND5)) of primers (Table 2). The theo-retical specificity of the designed primers for the similar and distantspecies was ensured using online Basic Local Alignment Tool(BLAST) in NCBI database (http://blast.ncbi.nlm.nih.gov/Blast.cgi).To determine the total mismatch between the target and non-target species, the primers were in-silico screened with 17 landanimals, namely, cow (Bos taurus), buffalo (Bubalus bubalis), pig(Sus scrofa), horse (Equus caballus), goat (Capra hircus), deer (Cervusnippon), sheep (Ovis aries), donkey (Equus africanus), dog (Canislupus), rabbit (Oryctolagus cuniculus), monkey (Macaca fascicularis),cat (Felis catus), chicken (Gallus gallus), duck (Anas platyrhynchos),pigeon (Columba livia), quail (Coturnix coturnix), rat (Rattus norve-gicus), 8 aquatic species, namely, cod (Gadus morhua), salmon(Salmo salar), tuna (Thunnus orientalis), tilapia (Oreochromis niloti-cus), rohu (Labeo rohita), Pangas (Pangasius pangasius), frog (Ranakunyuensis) and turtle (Cuora amboinensis) and four plant species,namely, wheat (Triticum aestivum), onion (Allium cepa), ginger(Zingiber officinale) and pepper (Capsicum annuum) using a Clus-talW multiple sequence alignment program (http://www.genome.jp/tools/clustalw/). The final specificity was confirmed through aPCR assay against templates of 25 alien species. The primers weresynthesized by Integrated DNA Technologies (IDT), Singapore andsupplied by First BASE Laboratories Sdn Bhd., Selangor, Malaysia.
2.6. Multiplex PCR assay
Prior to develop themPCR assay, simplex PCRwas performed foreach of the target species with individual set of primers asdescribed in our earlier report (Ali, Razzak et al., 2015). The simplexPCR was performed in a 25 ml reaction mixture comprised of 5 ml of
Table 2Primer sequences used in this study.
Name Species Target gene
Cocytb Cow (Bos taurus) Cytb
CoND5 Cow (Bos taurus) ND5
Bucytb Buffalo (Bubalus bubalis) Cytb
BuND5 Buffalo (Bubalus bubalis) ND5
Pocytb Pork (Sus scrofa) Cytb
PoND5 Pork (Sus scrofa) ND5
Please cite this article in press as: Hossain, M. A. M., et al., Targeting doublematerials in food chain, Food Control (2016), http://dx.doi.org/10.1016/j.fo
5� GoTaq Flexi Buffer, 0.2 mM each of dNTP, 2.5 mMMgCl2, 0.625 UGoTaq Flexi DNA Polymerase (Promega, Madison, USA), 0.4 mM ofeach primer and 2 ml (20 ng/ml) of DNA template. For negativecontrol, template DNA was replaced by distilled water. All PCRswere performed in an ABI 96 Well Verity Thermal Cycler (AppliedBiosystems, Foster City, CA, USA) with an initial denaturation at95� C for 3 min followed by 35 cycles of denaturation at 95� C for30 s, annealing at 60� C for 30e35 s, extension at 72� C for 40 s andthe final extension at 72� C for 5 min. PCR products were visualizedin 2% agarose gel stained with Florosafe DNA stain (1st base Lab-oratories, Selangor, Malaysia) under a gel image documentationsystem (AlphaImager HP, Alpha Innotech Corp., California, USA)(data not shown). After optimising simplex PCR assays for indi-vidual species, duplex, triplex, tetraplex and finally hexaplex PCRsystems were developed as given in Tables 3 and 4. The repro-ducibility and robustness of the assay was ensured with a change ofPCRmaster mix kit (QIAGENMultiplex PCR Plus Kit, QIAGENGmbH,Hilden, Germany) and operators. Due to the poor resolution ofagarose gel, multiplex PCR products were separated and visualizedin an automated QIAxcel Advanced Capillary Electrophoresis Sys-tem (QIAGEN Hilden, Germany) (Fig. 1).
3. Results
3.1. Quality and quantity of extracted DNA
Extracted DNA concentration was determined based on theabsorbance value at 260 nm and absorbance ratio at 260 nm and280 nm (Ali, Razzak, et al., 2015). The absorbance ratio at A260/A280was between 1.8 and 1.96 for all extracted DNA. This ensured thatgood quality DNA was extracted from all samples and it was suit-able for PCR reaction (Nejad, Tafvizi, Ebrahimi, & Hosseni, 2014).
3.2. PCR specificity
First of all, a simplex PCR was optimized for each primer pairagainst the template of each target species to ensure that eachtarget was amplified (Dalmasso et al., 2004). The primer sets of beef(Cocytb and CoND5), buffalo (Bucytb and BuND5) and pork (Pocytband PoND5) amplified 120, 106, 90, 138, 73 and 146 bp productsfrom the respective sites of cytb and ND5 genes in mitochondrial
Sequence (50 - 30) Amplicon size (bp)
Forward: CGGCACAAATTTAGTCGAATReverse: TGGACTATGGCAATTGCTATG
120
Forward: GGTTTCATTTTAGCAATAGCATGGReverse: GTCCAATCAAGGGTATGTTTGAG
106
Forward: GGGTTCTAGCCCTAGTTCTCTCTReverse: ATGGCCGGAACATCATACTT
90
Forward: TCGCCTAGCTTCTTACACAAACReverse: TGGTTTGTGACTGTGATGGAT
138
Forward: TATCCCTTATATCGGAACAGACCTCReverse: GCAGGAATAGGAGATGTACGG
146
Forward: GATTCCTAACCCACTCAAACGReverse: GGTATGTTTGGGCATTCATTG
73
Note: 5 ml of 5� GoTaq Flexi Buffer was used in all PCR experiments.
genes inmultiplex PCR for discriminating bovine, buffalo and porcineodcont.2016.08.008

Table: 4Cycling parameters for PCR reactions.
PCR reaction Initial denaturation 35 cycles (40 cycles for multiplex) Final extension
Denaturation Annealing Extension
Duplex and Triplex 95� C for 3 min 95� C for 30 s 60� C for 45 s 72� C for 45 s 72� C for 5 minTetraplex 95� C for 3 min 95� C for 40 s 60� C for 60 s 72� C for 50 s 72� C for 5 minMultiplex 95� C for 5 min 95� C for 50 s 60� C for 90 s 72� C for 50 s 72� C for 7 min
Fig. 1. The gel image (a) and electroferogram (b) of double gene-site targeting multiplex PCR for beef, buffalo and pork authentication. In the gel image: lane M represents DNAladder; lanes 1e11 PCR products from pork ND5 (lane 1); buffalo Cytb (lane 2); beef ND5 (lane 3); beef Cytb (lane 4); buffalo ND5 (lane 5); pork cytb (lane 6); duplex PCR of Cytb ofbeef and buffalo (lane 7) and ND5 of beef and buffalo (lane 8); triplex PCR of Cytb of beef and ND5 of beef and buffalo (lane 9); tetraplex PCR of Cytb and ND5 of beef and buffalo (lane10); hexaplex PCR of Cytb and ND5 of beef, buffalo and pork (lane 11); and negative control (lane N). The corresponding electropherogram of lane 11 is represented with labels.
M.A.M. Hossain et al. / Food Control xxx (2016) 1e104
genome (Fig. 1). Secondly, species specificity of the primers werecross-tested against 25 non-target species (goat, lamb, dog, cat,rabbit, monkey, donkey, chicken, duck, pigeon, quail, rat, salmon,tuna, cod, tilapia, rohu, pangas, frog and turtle, wheat, onion, garlic,ginger and pepper) (data not shown). The results demonstratedthat only the specific target products were amplified and no non-targets were co-amplified. The use of a universal eukaryoticprimer set which amplified 99 bp product from all species, reflectedthe presence of good quality DNA in all tubes, eliminating thepossibility of any false-negative detection (data not shown). Thisindicated a high fidelity of the each set of the primers for theamplification of specific targets. All tests were repeated three timeson three different days with a change of operators but the sameoutcomes were materialized.
After confirming the specificity of each set of primers, mPCRassay was developed step by step for the simultaneous detection of
Please cite this article in press as: Hossain, M. A. M., et al., Targeting doublematerials in food chain, Food Control (2016), http://dx.doi.org/10.1016/j.fo
all target species (Fig.1). The order of the developmental stagewentthrough the simplex (lanes 1e6), duplex (lanes 7 and 8), triplex(lane 9), tetraplex (lane 10) and hexaplex (lane 11) PCR systems toensure the consistency of the multiplex system (Ali, Razzak, et al.,2015). The simplex and all the multiplex systems (duplex, triplex,tetraplex and hexaplex) amplified the target gene (Cytb and ND5)sites (120 and 106 bp for beef, 90 and 138 bp for buffalo and 146 and73 for pig) respectively (Fig. 1), reflecting full conformity with thesimplex PCR system. Furthermore, consistence results were foundin hexaplex PCR when the operators were changed and a differentmultiplex PCR master mix kit (QIAGENMultiplex PCR Plus Kit (datanot shown) was used. Finally, the novel mPCR assay was screenedagainst 20 non-target animal and 5 plant species (described abovefor simplex PCR) for the analysis of cross-specificity. Every time, theassay yielded PCR products only from the beef, buffalo and pigtargets and no products from any other non-targets (data not
genes inmultiplex PCR for discriminating bovine, buffalo and porcineodcont.2016.08.008

M.A.M. Hossain et al. / Food Control xxx (2016) 1e10 5
shown). The amplified PCR products were separated by an auto-mated capillary electrophoretic system (QIAGEN GmbH, Hilden,Germany) which is capable of separating nucleic acid with 5e10 bpresolution by using an in-built gel cartridge within 12 min throughthe application of an electrical current to a gel-filled capillary car-tridge through individual electrode for each capillary. The migratedoligos were detected in an in-built-detector and displayed both asgel image (Fig. 1) and electropherogram (Inset of Fig. 1) through aphotomultiplier.
3.3. Limit of detection (LOD)
To determine the sensitivity of themPCR assay, extracted DNA ofthe target species (beef, buffalo and pig) were diluted serially fromhigher (50 ng/ml) to lower concentrations (10.0, 5.0, 1.0, 0.5, 0.2, 0.1,0.05, 0.02 and 0.01 ng/ml) since spectroscopic measurement at lowconcentration is not reproducible and trustworthy (www.biochrom.co.uk/download/72/). The QIAxcel automated capillaryelectrophoresis yielded six bands corresponding to two cytb andtwo ND5 genes of beef, buffalo and pig species from as low as0.02 ng of total DNA extracted from muscle tissues (Fig. 2, lane 8).The electroferogram (inset of Fig. 2) also clearly represented sixpeaks corresponding to the six different bands displayed in the gel-view. Thus the limit of detection (LOD) of the developed mPCRassay was concluded to be 0.02 ng DNA.
3.4. The screening of meatballs
Meatballs are popular all of the world and could be consumedeither as a separate menu or as additives for other dishes. Delib-erately contaminated model meatballs of each target species were
Fig. 2. Sensitivity analysis of multiplex PCR system. Shown are in the gel view (a), laneM is Dof beef, buffalo and pig species, respectively, and lane N is negative control (0 ng of DNA).
Please cite this article in press as: Hossain, M. A. M., et al., Targeting doublematerials in food chain, Food Control (2016), http://dx.doi.org/10.1016/j.fo
prepared in laboratory as described in section 2.2 and commercialmeatballs of five different brands for beef (described as A-E) andpork (described as A-E) were purchased from different sellingoutlets across Malaysia on three different dates. The model meat-balls of each target species were deliberately adulterated with 1%,0.5% and 0.1% raw meat of other two target species as given insection 2.2. The 0.1% spiked meatballs of three species were auto-claved at 121� C and 15-psi pressure for 2.5 h to simulate extensivecooking effect. The experimental finding is given in Fig. 3 and theanalytical data are presented in Table 5.
The model beef, buffalo and pork meatballs, adulterated with1%, 0.5% and 0.1% of buffalo and pork; beef and pork; and beef andbuffalo, amplified all the six targets (Fig. 3; lanes 1e3; 5e7 and9e11) representing the three target species. The 0.1% adulteratedautoclaved meatballs also positively amplified six targets for beef,buffalo and pork (lane 4, 8 and 12). In case of commercial meatballproducts, only pork was contained in pork meatballs but 80% of theanalysed beef meatball was found to contain both beef and buffaloand 20% was found to contain only buffalo species. Thus theabsence of pure beef meatballs in Malaysian markets reflected thatbuffalo substitution in beef products is rampant for the pursuit ofillegal economic gain.
3.5. Validation of mPCR assay in thermally processed meat
To assess the detection efficiency of the described mPCR assay,various thermally treated meat samples were analysed (Arslan,Ilhak, & Calicioglu, 2006; Karabasanavar, Singh, Umapathi, Girishet al., 2011, Karabasanavar, Singh, Umapathi, Kumar et al., 2011).As reported in methodology (section 2.3), raw meat of beef, buffaloand pork were subjected to three different heat treatment
NA ladder, lane 1e8 are the PCR products of 10, 5, 1, 0.5, 0.2, 0.1, 0.05 and 0.02 ng of DNAThe electropherogram (b) of lanes 8 is presented with labels.
genes inmultiplex PCR for discriminating bovine, buffalo and porcineodcont.2016.08.008

Fig. 3. The gel image (a) and electropherograms (bed) of multiplex PCR (m-PCR) for the detection of double gene-targeting beef, buffalo and pig in deliberately adulterated modelbeef, buffalo and pork meatball under raw and processed states. In the gel image; lane M, Ladder; lanes 1e3, m-PCR of beef meatball spiked with 1%, 0.5% and 0.1% of buffalo andpork, respectively, under raw state; lanes 5e7, m-PCR of buffalo meatball spiked with 1%, 0.5% and 0.1% of beef and pig, respectively, under raw state; lanes 9e11, m-PCR of porkmeatball spiked with 1%, 0.5% and 0.1% of beef and buffalo, respectively, under raw state; lane 4, 8 and 12, m-PCR of heat treated (autoclaved for 2.5 h) 0.1% adulterated beef, buffaloand pork meatballs respectively; lane N negative control. The corresponding electroferograms of lane 4, 8 and 12 are shown with labels in b, c and d, respectively.
Table 5Screening results of model and commercial meatball products using mPCR.
Sample Adulteration State Detected species PCR accuracy (%)
Species % Beef Buffalo Pig
Model meatballsBeef meatball Buffalo and Pig 1.0 Raw 9/9 9/9 9/9 100Beef meatball Buffalo and Pig 0.5 Raw 9/9 9/9 9/9 100Beef meatball Buffalo and Pig 0.1 Raw 9/9 9/9 9/9 100Beef meatball Buffalo and Pig 0.1 Autoclaved for 2.5 h 9/9 9/9 9/9 100Buffalo meatball Beef and Pig 1.0 Raw 9/9 9/9 9/9 100Buffalo meatball Beef and Pig 0.5 Raw 9/9 9/9 9/9 100Buffalo meatball Beef and Pig 0.1 Raw 9/9 9/9 9/9 100Buffalo meatball Beef and Pig 0.1 Autoclaved for 2.5 h 9/9 9/9 9/9 100Pork meatball Beef and Buffalo 1.0 Raw 9/9 9/9 9/9 100Pork meatball Beef and Buffalo 0.5 Raw 9/9 9/9 9/9 100Pork meatball Beef and Buffalo 0.1 Raw 9/9 9/9 9/9 100Pork meatball Beef and Buffalo 0.1 Autoclaved for 2.5 h 9/9 9/9 9/9 100Commercial meatballs*Beef meatball A e e Raw 9/9 9/9 0/9 100Beef meatball B e e Raw 9/9 9/9 0/9 100Beef meatball C e e Raw 6/9 9/9 0/9 100Beef meatball D e e Raw 7/9 9/9 0/9 100Beef meatball E e e Raw 5/9 9/9 0/9 100Pork meatball A e e Raw 0/9 0/9 9/9 100Pork meatball B e e Raw 0/9 0/9 9/9 100Pork meatball C e e Raw 0/9 0/9 9/9 100Pork meatball D e e Raw 0/9 0/9 9/9 100Pork meatball E e e Raw 0/9 0/9 9/9 100
Notes: *Five different brands (A-E) of each beef and pork meatballs were analysed to validate the developed multiplex PCR assay. The numerator and denominator of eachfraction denote the number of positive detection and total number of samples analysed using the multiplex PCR assay.
M.A.M. Hossain et al. / Food Control xxx (2016) 1e106
processes, namely boiling, autoclaving andmicrowave cooking. Theextracted DNA of the all processed meat samples were used astemplates for the mPCR assay. The system successfully detected alltarget species, beef, buffalo and pork under all thermal processingconditions, including extensive autoclaving (121� C at 15-psi for2.5 h) and extreme microwaving at 700 W for 30 min (Fig. 4).
Please cite this article in press as: Hossain, M. A. M., et al., Targeting doublematerials in food chain, Food Control (2016), http://dx.doi.org/10.1016/j.fo
4. Discussion
The motivation of adulteration comes from a company's interestin making an on growing profit by selling a cheaper item in thename of its expensive counterparts. It incurs a serious risk espe-cially when an animal material is involved. According to the USDepartment of Agriculture, about 75% of the recently emerging
genes inmultiplex PCR for discriminating bovine, buffalo and porcineodcont.2016.08.008
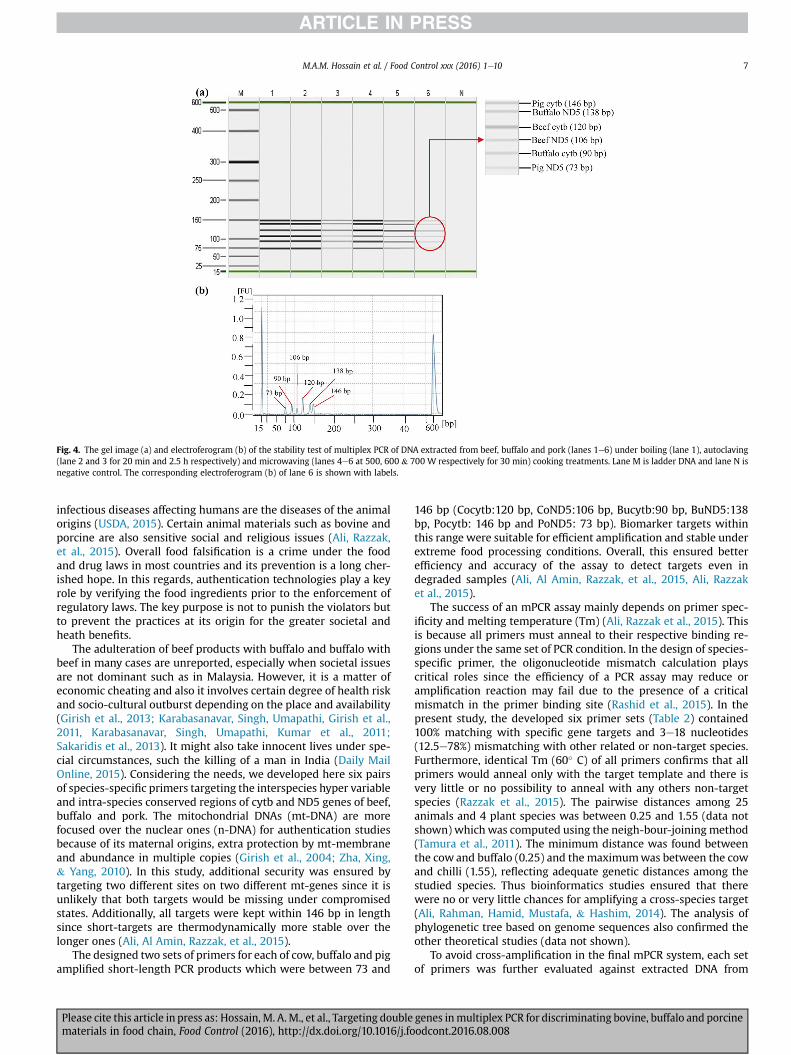
Fig. 4. The gel image (a) and electroferogram (b) of the stability test of multiplex PCR of DNA extracted from beef, buffalo and pork (lanes 1e6) under boiling (lane 1), autoclaving(lane 2 and 3 for 20 min and 2.5 h respectively) and microwaving (lanes 4e6 at 500, 600 & 700 W respectively for 30 min) cooking treatments. Lane M is ladder DNA and lane N isnegative control. The corresponding electroferogram (b) of lane 6 is shown with labels.
M.A.M. Hossain et al. / Food Control xxx (2016) 1e10 7
infectious diseases affecting humans are the diseases of the animalorigins (USDA, 2015). Certain animal materials such as bovine andporcine are also sensitive social and religious issues (Ali, Razzak,et al., 2015). Overall food falsification is a crime under the foodand drug laws in most countries and its prevention is a long cher-ished hope. In this regards, authentication technologies play a keyrole by verifying the food ingredients prior to the enforcement ofregulatory laws. The key purpose is not to punish the violators butto prevent the practices at its origin for the greater societal andheath benefits.
The adulteration of beef products with buffalo and buffalo withbeef in many cases are unreported, especially when societal issuesare not dominant such as in Malaysia. However, it is a matter ofeconomic cheating and also it involves certain degree of health riskand socio-cultural outburst depending on the place and availability(Girish et al., 2013; Karabasanavar, Singh, Umapathi, Girish et al.,2011, Karabasanavar, Singh, Umapathi, Kumar et al., 2011;Sakaridis et al., 2013). It might also take innocent lives under spe-cial circumstances, such the killing of a man in India (Daily MailOnline, 2015). Considering the needs, we developed here six pairsof species-specific primers targeting the interspecies hyper variableand intra-species conserved regions of cytb and ND5 genes of beef,buffalo and pork. The mitochondrial DNAs (mt-DNA) are morefocused over the nuclear ones (n-DNA) for authentication studiesbecause of its maternal origins, extra protection by mt-membraneand abundance in multiple copies (Girish et al., 2004; Zha, Xing,& Yang, 2010). In this study, additional security was ensured bytargeting two different sites on two different mt-genes since it isunlikely that both targets would be missing under compromisedstates. Additionally, all targets were kept within 146 bp in lengthsince short-targets are thermodynamically more stable over thelonger ones (Ali, Al Amin, Razzak, et al., 2015).
The designed two sets of primers for each of cow, buffalo and pigamplified short-length PCR products which were between 73 and
Please cite this article in press as: Hossain, M. A. M., et al., Targeting doublematerials in food chain, Food Control (2016), http://dx.doi.org/10.1016/j.fo
146 bp (Cocytb:120 bp, CoND5:106 bp, Bucytb:90 bp, BuND5:138bp, Pocytb: 146 bp and PoND5: 73 bp). Biomarker targets withinthis range were suitable for efficient amplification and stable underextreme food processing conditions. Overall, this ensured betterefficiency and accuracy of the assay to detect targets even indegraded samples (Ali, Al Amin, Razzak, et al., 2015, Ali, Razzaket al., 2015).
The success of an mPCR assay mainly depends on primer spec-ificity and melting temperature (Tm) (Ali, Razzak et al., 2015). Thisis because all primers must anneal to their respective binding re-gions under the same set of PCR condition. In the design of species-specific primer, the oligonucleotide mismatch calculation playscritical roles since the efficiency of a PCR assay may reduce oramplification reaction may fail due to the presence of a criticalmismatch in the primer binding site (Rashid et al., 2015). In thepresent study, the developed six primer sets (Table 2) contained100% matching with specific gene targets and 3e18 nucleotides(12.5e78%) mismatching with other related or non-target species.Furthermore, identical Tm (60� C) of all primers confirms that allprimers would anneal only with the target template and there isvery little or no possibility to anneal with any others non-targetspecies (Razzak et al., 2015). The pairwise distances among 25animals and 4 plant species was between 0.25 and 1.55 (data notshown) which was computed using the neigh-bour-joining method(Tamura et al., 2011). The minimum distance was found betweenthe cow and buffalo (0.25) and the maximumwas between the cowand chilli (1.55), reflecting adequate genetic distances among thestudied species. Thus bioinformatics studies ensured that therewere no or very little chances for amplifying a cross-species target(Ali, Rahman, Hamid, Mustafa, & Hashim, 2014). The analysis ofphylogenetic tree based on genome sequences also confirmed theother theoretical studies (data not shown).
To avoid cross-amplification in the final mPCR system, each setof primers was further evaluated against extracted DNA from
genes inmultiplex PCR for discriminating bovine, buffalo and porcineodcont.2016.08.008

M.A.M. Hossain et al. / Food Control xxx (2016) 1e108
muscle tissues of other two target species in a simplex PCR reaction,i.e. the beef primer sets were assayed with DNA template of buffaloand pig and buffalo primer sets were assayed with DNA template ofbeef and pig and so on. Then, each primer pair were checkedagainst 25 non-target species (lamb, goat, rabbit, cat, dog, donkey,duck, chicken, monkey, pigeon, quail, rat, salmon, tuna, cod, tilapia,rohu, pangas, turtle and frog, wheat, onion, ginger, garlic andpepper). Results demonstrated no cross-amplifications with any ofthe primer pairs. All of these indicated that the designed primerswere highly specific for the target species. Triplicate assays per-formed on three different days with a change of operators andblindly labelled samples and reproducible results were obtained.
After optimization the simplex PCR, two duplex PCR for Cocytband Bucytb and CoND5 and BuND5; one triplex of Cocytb, CoND5and BuND5; one tetraplex of Cocytb, Bucytb, CoND5 and BuND5and finally multiplex (hexaplex) PCR of Pocytb, Cocytb, Bucytb,CoND5, BuND5 and PoND5 were optimized step by step to elimi-nate the possibility of forming any primer dimers or multimers(Fig. 1) (Ali, Razzak, et al., 2015). Due to the narrow differences inthe length of the targets, agarose gel electrophoresis, which hardlyseparate nucleic acid of less than 50 bp difference in length, couldnot be used to separate the amplicons for visualization (Bottero &Dalmasso, 2011). It is also a laborious technique and requirerather longer separation time. Consequently, we used here a fullyautomated multi-capillary electrophoresis device (QIAxcelAdvanced Capillary Electrophoresis System, Germany) for theseparation and visualization of PCR products. This effectivelyenhanced sensitivity and resolution (~5 bp) and shortened analysistime, minimizing the manual handling errors and exposure tohazardous chemicals by virtue of its in-built gelmatrices in a ready-to-run gel cartridge (DOOLEY et al., 2005; Fajardo, Gonz�alez, Rojas,García, & Martín, 2010). The well separated mPCR products wereclearly visualized in the gel image along with the electrophero-grams (Fig. 1) for all of six targets.
To check the sensitivity of the mPCR assay, DNA template of alltarget species was serially diluted from higher to lower concen-tration (10 nge0.01 ng) by adding required amount of deionizedwater (Ali, Razzak, et al., 2015) (Fig. 2). The six distinct bands of thesix PCR target amplified from 0.02 ng DNA template were observedin both the gel-view (lane 8) and electroferogram (inset), con-firming the detection limit (LOD) at 0.02 ng DNA. LOD at this rangewas sufficient to detect any commercial frauding for profit makingpurposes (Razzak et al., 2015). Previously, Ali, Razzak et al., (2015)detected 0.01 ng DNA of dog, monkey and rat but 0.02 ng wasfound for pig and cat in a pentaplex PCR system of 108e172 bpproduct sizes. Zhang, (2013) documented 1 pg (0.001 ng) sensitivityfor a semi-nested mPCR for beef (263 bp), pork (387 bp), mutton(322 bp) and chicken (216 bp). However, in semi-nested mPCR acommon primer pair is amplified at first place and the amplifiedproduct is then used as a template for the multiplex PCR. Thesemake the assay more difficult, costlier time consuming and lesstrustworthy. In addition, the identical efficiency of the shortenedprimers for different templates often makes them incapable todetect an accurate species (Ali, Razzak, et al., 2015). Furthermore,Kitpipit et al. (2014) obtained 7e21 fg LOD for beef, Pork, lamb,chicken, ostrich and horse with amplicon sizes of 100e311 bp. Onthe other hand, Luo et al. (2008) detected 0.1e0.2 ng DNA in amPCRassay for cattle, pig, sheep and chicken containing 149e274 bpproduct sizes. Additionally, a duplex PCR identified beef (113 bp)and buffalo (152 bp) milk and meat from 0.001 ng DNA (REA et al.,2001; Gupta, Rank, & Joshi, 2012). Thus the variation in sensitivityis a common phenomenon in mPCR and it depends on severalfactors such as target gene, target species, amplicon size and qualityof the source material (Hou et al., 2015).
Although several assays are reported for bovine, buffalo and
Please cite this article in press as: Hossain, M. A. M., et al., Targeting doublematerials in food chain, Food Control (2016), http://dx.doi.org/10.1016/j.fo
porcine identification, the most of the reported mPCR assays havenot been optimized and tested for adulteration detection undercommercial matrices and extensive food processing conditions.This novel mPCR assay was optimized and validated for the analysisof a very popular meat product, meatball under raw and heattreated condition to evaluate the reliability and accuracy of themethod. It positively detected target species from up to 0.1%adulterated and severely autoclaved model meatballs (Fig. 3). Morerecently, Razzak et al. (2015) documented a sensitivity threshold of0.1% for adulterated meatball, burger and frankfurter productsunder raw states but up to 1% for 2.5 h autoclaved samples. Houet al. (2015) reported a sensitivity limit up to 1% for chicken, duckand goose boiled at 100 �C. In simplex PCR, Karabasanavar, Singh,Kumar, and Shebannavar (2014) identified 0.1% adulterated porkunder raw states. Mane, Mendiratta, Tiwari (2012) documented lessthan 1% autoclaved beef in mixed states. To the best of ourknowledge, no other studies have tested mPCR assay underextreme food processing conditions such as boiling (90 min),autoclaving (2.5 h) and micro-oven cooking (700 W for 30 min)(Fig. 4), although simplex PCR assays have been documented underharsh processing condition (Ali, Al Amin, Razzak et al., 2015, Ali,Asing Hamid et al., 2015; Karabasanavar, Singh, Umapathi, Girishet al., 2011, Karabasanavar, Singh, Umapathi, Kumar et al., 2011).The specialty of our assay was double gene target, short ampliconlength, exceptional stability and sufficient sensitivity under raw,admixed and processed states. A double targeted PCR assay is morereliable and trustworthy since alternative targets can compensatethe detection of a missing target under the state of decomposition.Finally, we applied the developed mPCR for the analysis of com-mercial beef and pork meatballs as buffalo meatball was notavailable in the Malaysian markets. All pork meatballs were posi-tively detected only pork but beef meatballs were detected bothbeef and buffalo for 80% samples and total replacement of beef withbuffalo in other 20% (the total number of samples were tested5 � 9 ¼ 45 for the each of beef and pork products) (Table 5).Although both beef and buffalo are permitted by the Halal Au-thority of Malaysia, beef meatball was adulterated with buffalo forgaining unjustified profit since the buffalo meat is cheaper thanbeef in Malaysia. However, as per the European guideline, fakelabelling in food is a crime and it must be prevented (Przyrembel,2004). However, because of the strict monitoring of haram spe-cies, Halal certified buffalo adulterated beef meatballs were foundfree from any non-Halal porcine materials.
5. Conclusion
Counterfeiting of animal materials in food products is anemerging concern and to combat this problem regulatory bodymust need a trustworthy technique for ingredient authentication.This novel mPCR assay for the discriminatory detection of bovine,buffalo and porcine materials is outstanding because it ensuredextra security through targeting two sites of two different geneswhich are less likely to break down even under the states ofdecomposition. Furthermore, alternative targets could complementthe detection of a missing target. The assay was sensitive enough todetect 0.02 ng pure DNA and 0.1% adulterated meat under mixedmatrices. Due to shorter length of the targets (73e146 bp), all tar-gets were stable under extensive boiling, micro-waving and auto-claving treatments. Finally, its optimization and validation for thescreening of commercial meatball products reflected the methodwas operationally feasible for the detection of trace amount bovine,buffalo and porcinematerials in food chain. Market survey revealedabout 80% of beef meatballs are adulterated with buffalo and about20% are totally replaced with buffalo. However, porcine meatballswere found to be 100% authentic and no porcine was detected in
genes inmultiplex PCR for discriminating bovine, buffalo and porcineodcont.2016.08.008

M.A.M. Hossain et al. / Food Control xxx (2016) 1e10 9
halal branded beef meatballs. The findings demonstrated sufficientmerit for using this novel mPCR assay as a practical tool for theroutine analysis of beef, buffalo and pig products even under thestate of degraded specimens.
Ethical compliance
The ethical permission was obtained from the University ofMalaya Ethical Committee and all the animals and meats used inthis study were handled following the institutional and nationalguideline for animal care.
Conflict of interest
All authors declare that there is no conflict of interest to publishthe content of this article.
Acknowledgments
This work was supported by University of Malaya ResearchGrant No. GC001-14SBS to M.E. Ali. The authors acknowledgeWildlife Malaysia and Dewan Bandaraya, Kuala Lumpur (DBKL) forproviding monkey, dog and cat meat samples.
References
Ali, M. E., Al Amin, M., Razzak, M. A., Hamid, S. B. A., Rahman, M. M., & Rashid, N. A.(2015). Short amplicon-length PCR assay targeting mitochondrial cytochrome bgene for the detection of feline meats in burger formulation. Food AnalyticalMethods, 1e11.
Ali, M. E., Asing Hamid, S. B. A., Razzak, M. A., Rashid, N. R. A., Amin, M. A., &Mustafa, S. (2015). A suitable method to detect potential fraud of bringingMalayan box turtle (Cuora amboinensis) meat into the food chain. Food Addi-tives & Contaminants: Part A, 1e11 (ahead-of-print).
Ali, M. E., Hashim, U., Mustafa, S., Man, Y. C., Dhahi, T. S., Kashif, M., et al. (2012).Analysis of pork adulteration in commercial meatballs targeting porcine-specific mitochondrial cytochrome b gene by TaqMan probe real-time poly-merase chain reaction. Meat science, 91(4), 454e459.
Ali, M. E., Rahman, M. M., Hamid, S. B. A., Mustafa, S., & Hashim, S. B. U. (2014).Canine-specific PCR assay targeting cytochrome b gene for the detection of dogmeat adulteration in commercial frankfurters. Food Anal. Methods, 7, 234e241.
Ali, M. E., Razzak, M. A., & Hamid, S. B. A. (2014). Multiplex PCR in speciesAuthentication: Probability and prospectsda review. Food Analytical Methods,7(10), 1933e1949.
Ali, M. E., Razzak, M. A., Hamid, S. B. A., Rahman, M. M., Amin, M. A., Rashid, N. R. A.,et al. (2015). Multiplex PCR assay for the detection of five meat speciesforbidden in Islamic foods. Food Chemistry, 177, 214e224.
Amin, M. A., Hamid, S. B. A., & Ali, M. E. (2016). A method for the detection ofpotential fraud of bringing feline meat in food chain. International Journal ofFood Properties, 19(7), 1645e1658.
Arslan, A., Ilhak, O. I., & Calicioglu, M. (2006). Effect of method of cooking onidentification of heat processed beef using polymerase chain reaction (PCR)technique. Meat Science, 72(2), 326e330.
Bottero, M. T., & Dalmasso, A. (2011). Animal species identification in food products:Evolution of biomolecular methods. The Veterinary Journal, 190, 34e38.
Daily Mail Online. (2015). Father is beaten to death with bricks by Indian mob and hisson is left fighting for life after rumours circulate he'd eaten beef… but it wasactually lamb. http://www.dailymail.co.uk/news/article-3254623/ Accessed03.02.16.
Dalmasso, A., Fontanella, E., Piatti, P., Civera, T., Rosati, S., & Bottero, M. T. (2004).A multiplex PCR assay for the identification of animal species in feedstuffs.Molecular and Cellular Probes, 18(2), 81e87.
Dooley, j. j., Sage, H. D., Clarke, L. M.-A., Brown, H. M., & Garrett, S. D. (2005). Fishspecies identification using pcr-RFLP analysis and lab-on-a-chip capillaryElectrophoresis: Application to detect white fish species in food products andan interlaboratory study. Journal of Agricultural and Food Chemistry, 53,3348e3357.
Erwanto, Y., Abidin, M. Z., Sugiyono, E. Y. P. M., & Rohman, A. (2014). Identification ofpork contamination in meatballs of Indonesia local market using polymerasechain reaction-restriction fragment length polymorphism (PCR-RFLP) analysis.Asian-Australasian journal of animal sciences, 27(10), 1487.
Fajardo, V., Gonz�alez, I., Rojas, M., García, T., & Martín, R. (2010). A review of currentPCR-based methodologies for the authentication of meats from game animalspecies. Trends in food science & technology, 21(8), 408e421.
Girish, P., Anjaneyulu, A., Viswas, K., Anand, M., Rajkumar, N., Shivakumar, B., et al.
Please cite this article in press as: Hossain, M. A. M., et al., Targeting doublematerials in food chain, Food Control (2016), http://dx.doi.org/10.1016/j.fo
(2004). Sequence analysis of mitochondrial 12S rRNA gene can identify meatspecies. Meat Science, 66(3), 551e556.
Girish, P. S., Haunshi, S., Vaithiyanathan, S., Rajitha, R., & Ramakrishna, C. (2013).A rapid method for authentication of Buffalo (Bubalus bubalis) meat by AlkalineLysis method of DNA extraction and species specific polymerase chain reaction.Journal of Food Science Technology, 50(1), 141e146.
Gupta, R., Rank, D., & Joshi, C. (2012). Identification and differentiation of cattle andbuffalo processed meat by duplex-pcr. Buffalo bulleitn, 31(1), 6. IBIC, KasetsartUniversity, PO Box 1084 Bangkok 10903, Thailand.
Haider, N., Nabulsi, I., & Al-Safadi, B. (2012). Identification of meat species by PCR-RFLP of the mitochondrial COI gene. Meat Sci, 90(2), 490e493.
Herrero, B., Royo, L. J., Lago, F. C., Vieites, J. M., & Espi~neira, M. (2013). Authenticationof male beef by multiplex fast real-time PCR. Food Additives & Contaminants:Part A, 30(2), 218e225.
Hird, H., Lloyd, J., Goodier, R., Brown, J., & Reece, P. (2003). Detection of peanut usingreal-time polymerase chain reaction. European Food Research and Technology,217(3), 265e268.
Hou, B., Meng, X., Zhang, L., Guo, J., Li, S., & Jin, H. (2015). Development of a sensitiveand specific multiplex PCR method for the simultaneous detection of chicken,duck and goose DNA in meat products. Meat Science, 101, 90e94.
Karabasanavar, N. S., Singh, S. P., Kumar, D., & Shebannavar, S. N. (2014). Detection ofpork adulteration by highly-specific PCR assay of mitochondrial D-loop. FoodChemistry, 145, 530e534.
Karabasanavar, N. S., Singh, S. P., Umapathi, V., Girish, P. S., Shebannavar, S. N., &Kumar, D. (2011). Authentication of carabeef (water buffalo, Bubalus bubalis)using highly specific polymerase chain reaction. European Food Research andTechnology, 233(6), 985e989.
Karabasanavar, N. S., Singh, S., Umapathi, V., Kumar, D., Patil, G., & Shebannavar, S. N.(2011). A highly specific PCR assay for identification of raw and heat treatedmutton (Ovis aries). Small Ruminant Research, 100(2), 153e158.
Kitpipit, T., Sittichan, K., & Thanakiatkrai, P. (2014). Direct-multiplex PCR assay formeat species identification in food products. Food Chemistry, 163, 77e82.
K€oppel, R., Ruf, J., & Rentsch, J. (2011). Multiplex real-time PCR for the detection andquantification of DNA from beef, pork, horse and sheep. European Food Researchand Technology, 232(1), 151e155.
Luo, J. Q., Wang, J. Q., Bu, D. P., Dan, L. I., Li, W. A. N. G., Wei, H. Y., et al. (2008).Development and application of a PCR approach for detection of bovis, sheep,pig, and chicken derived materials in feedstuff. Agricultural Sciences in China,7(10), 1260e1266.
Mane, B. G., Mendiratta, S. K., & Tiwari, A. K. (2012). Beef specific polymerase chainreaction assay for authentication of meat and meat products. Food Control,28(2), 246e249.
Mane, Mendiratta, S. K., Tiwari, A. K., & Bhilegaokar, K. N. (2012). Detection ofadulteration of meat and meat products with buffalo meat employing poly-merase chain reaction assay. Food Analytical Methods, 5, 296e300.
Mohamad, N. A., El Sheikha, A. F., Mustafa, S., & Mokhtar, N. F. K. (2013). Comparisonof gene nature used in real-time PCR for porcine identification and quantifi-cation: A review. Food Research International, 50, 330e338.
Napolitano, F., Annicchiarico, G., Catillo, G., Cris�a, A., Grandoni, F., Marchitelli, C.,et al. (2014). Identification of Ovis aries Gelsolin isoform b, a candidate gene formilk quality. Small Ruminant Research, 116(1), 21e27.
Nejad, F. P., Tafvizi, F., Ebrahimi, M. T., & Hosseni, S. E. (2014). Optimization ofmultiplex PCR for the identification of animal species using mitochondrialgenes in sausages. European Food Research and Technology, 239(3), 533e541.
News Straits Times. (2015). Russia says wrong animal DNA found in Auchan mincedmeat. http://www.nst.com.my/news/2015/09/russia-says-wrong-animal-dna-found-auchan-minced-meat/ Accessed14.01.16.
Przyrembel, H. (2004). Food labelling legislation in the EU and consumers infor-mation. Trends in Food Science & Technology, 15(7), 360e365.
Rahman, M. M., Ali, M. E., Hamid, S. B. A., Mustafa, S., Hashim, U., & Hanapi, U. K.(2014). Polymerase chain reaction assay targeting cytochrome b gene for thedetection of dog meat adulteration in meatball formulation. Meat Science, 97,404e409.
Rashid, N. R., Ali, M. E., Hamid, S. B., Rahman, M. M., Razzak, M. A., Asing, et al.(2015). A suitable method for the detection of a potential fraud of bringingmacaque monkey meat into the food chain. Food Additives & Contaminants: PartA, 32(7), 1013e1022.
Razzak, M. A., Hamid, S. B. A., & Ali, M. E. (2015). A lab-on-a-chip-based multiplexplatform to detect potential fraud of introducing pig, dog, cat, rat and monkeymeat into the food chain. Food Additives & Contaminants: Part A, 32(11),1902e1913.
Rea, S., Chikuni, K., Branciari, R., Sangamayya, R. S., Ranucci, D., & Avellini, P. (2001).Use of duplex polymerase chain reaction (duplex-PCR) technique to identifybovine and water buffalo milk used in making mozzarella cheese. Journal ofDairy Research, 68(04), 689e698.
Sakaridis, I., Ganopoulos, I., Argiriou, A., & Tsaftaris, A. (2013). A fast and accuratemethod for controlling the correct labeling of products containing buffalo meatusing High Resolution Melting (HRM) analysis. Meat Science, 94(1), 84e88.
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., & Kumar, S. (2011).MEGA5: Molecular evolutionary genetics analysis using maximum likelihood,evolutionary distance, and maximum parsimony methods. Molecular Biologyand Evolution, 28(10), 2731e2739.
United States Department of Agriculture (USDA). (2015). Zoonotic diseases of sheep
genes inmultiplex PCR for discriminating bovine, buffalo and porcineodcont.2016.08.008

M.A.M. Hossain et al. / Food Control xxx (2016) 1e1010
and goats. https://www.aphis.usda.gov/aphis/ourfocus/animalhealth/animal-disease-information/sheep-and-goat-health/national-scrapie-eradication-program/ct_zoonotic_disease/ Accessed 14.04.16.
Walkera, M. J., Burnsb, M. a., & Burns, D. T. (2013). Horse meat in beef products-species substitution 2013. Journal of the Association of Public Analysts (Online),
Please cite this article in press as: Hossain, M. A. M., et al., Targeting doublematerials in food chain, Food Control (2016), http://dx.doi.org/10.1016/j.fo
41, 67e106.Zhang, C. (2013). Semi-nested multiplex PCR enhanced method sensitivity of spe-
cies detection in further-processed meats. Food Control, 31(2), 326e330.Zha, D., Xing, X., & Yang, F. (2010). A multiplex PCR assay for fraud identification of
deer products. Food Control, 21, 1402e1407.
genes inmultiplex PCR for discriminating bovine, buffalo and porcineodcont.2016.08.008























![QIAGEN Multiplex PCR Handbook[1]](https://img.dokumen.tips/doc/110x75/577cc7781a28aba711a107f5/qiagen-multiplex-pcr-handbook1.jpg)





