Embed Size (px)
Citation preview

Trans. Br, mycol. Soc. 71 (3) 419-423 (1978)
[ 419 ]
Printed in Great Britain
SURVIVAL OF AERO-AQUATIC HYPHOMYCETES ON LAND
By P. J. FISHER
Department of Biological Sciences, University of Exeter
Eight species of aero-aquatic hyphomycetes were tested for survival on garden soil underfield conditions, prolonged drying in the laboratory and desiccation over silica gel. In the fieldthree species survived after 11 months, one for 8 months, two for 6 months and two for2 months. Under laboratory conditions one species survived for 10 months, one for 8 months,two for 3 months, two for 1 month and two for 7 days. In a desiccator one species survived for9 months, one for 3 months, four for 1 month and two for 7 days.
Pure mycelia of 12 species were dried under laboratory conditions and tested for survival.Three species survived for 5 months, one for 4 months, two for 2 months, two for 1 month andfour for less than 1 month.
Spores of eight species were subjected to desiccation over silica gel. Six species survived for10 days but were dead after 20 days. The remaining two species survived for less than10 days. The results are discussed in relation to the ecology of the organisms.
Aero-aquatic hyphomycetes are characterized byformation of a vegetative mycelium under water,sporulating only when the substrate in which thefungus is growing is exposed to a moist atmosphere(van Beverwijk, 1953; Fisher, 1977).
The inability to produce spores on a submergedmycelium is an important characteristic of theaero-aquatic hyphomycetes which distinguishesthem from the aquatic hyphomycetes. Existingrecords indicate that the group is adapted for lifeunder water and in localities of high humidities(Linder, 1929; Glen-Bott, 1951, 1955; vanBeverwijk, 1951a, b, 1953, 1954; Gonczol, 1976).Our own records indicate that the preferredhabitat of the group is static and often stagnantwater (Fisher, 1977; Fisher & Webster, 1978).A few species have, however, occasionally beenreported from non-aquatic environments and fromvery small water sources (Waid, 1954; Bandoni,1972; Park, 1974b; Gonczol, 1976).
During studies undertaken in South Devon thefollowing species were recovered from moist leaflitter, in an oak and pine forest adjacent to BystockReservoir: Helicoon pluriseptatum, Candelabrumspinulosum and Spirosphaera floriformis. Thefollowing species were recovered from surface soiladjacent to a pine plantation near BurratorReservoir on Dartmoor ; Aegerita sp., Clathrosphaerina zalewskii, Helicodendron giganteum, Hd.triglitziense, Ho. pluriseptatum and Helicosporiumsp.
This paper reports the survival of a selectednumber of species in leaf mash and leaf litter ongarden soil, in air-dried leaf mash in the laboratoryand in a desiccator. Pure mycelia and spores werealso investigated for resistance to drying.
MATERIALS AND METHODS
Survival in field, laboratory and desiccatorexperiments
The following eight species were investigated :H elicodendron conglomeratum Glen-Bott, Hd. fuscum (Berk. & Curt.) Linder, Hd. giganteumGlen-Bott, Hd. hyalinum Linder, Hd. tubulosum(Riess) Linder, Hd. triglitziense (j aap) Linder,Helicoon ellipticum (Pk) Morgan, and Aegerita sp.The fungi were isolated from decaying leaf litterrecovered from static water sources in woodlandin South Devon (Fisher & Webster, 1978) andcultured for the experiments as follows.
Freshly abscissed deep frozen beech leaves weremacerated in a vegetable blender and stra inedthrough a cheese cloth. Approximately 100 g of theleaf mash was introduced into each of thirty-two1 I conical flasks together with 500 cm 3 distilledwater and autoclaved for 1 h at 120 °C.
A set of four flasks was then inoculated with twoagar disks taken from the margin of a 4-week-oldculture of the respective species. After an incubation period of 7 weeks the contents of each setof four flasks were thoroughly mixed by hand inan enamel dish and spread out to dry for 7 daysat 25-30°. The air dried mash of each species wasnow divided into three portions to be used in thefield, laboratory and desiccator experiments. Afterthe 7 day drying period small samples of leaf mashwere taken from each Jot and tested for the presenceof living mycelium by placing them into McCartneybottles with 15 ern" of sterile distilled water, andincubating for 7 days at 20°. The bottles wereshaken vigorously for about 30 s daily to ensure
15 ' 2

42 0 Aero-aquatic hyphomycetes on land
aeration. Subsequent sporulation was determinedby moist chamber incubation in Petri dishes.
The eight portions of the inoculated dried mashset aside for the field experiment were each spreadseparately in a cage 20 x 20 em" with wooden sidesand nylon mesh (1 mm-) as a base and top andplaced outside on garden soil. Before the tops weresecured ten non-sterile beech leaves taken from thefrozen stock, were loosely placed on top of eachlot of mash.
At chosen intervals 3-5 g samples were removedfor the presence of spores of the inoculated species.The samples were tested for the presence of livingmycelium and the moisture content of the leafmash determined (Table 1). The presence ofliving mycelium in the leaf mash was detected byincubating in 250 em" conical flasks with 100 em"sterile distilled water at 20° for 7 days. Sporulationon the mash was noted after incubation in Petridish moist chambers.
For the desiccator experiment eight Petri dishes,5 ern diam, were each filled with dried colonizedleaf mash, and placed over self-indicating silica gelin a desiccator. The relative humidity within thedesiccator was determined with a Relative Percentage Humidity Indicator (Negretti & Zambra).It remained at 4 ± 0' 5 % at 20° throughout theexperiment. At chosen intervals the leaf mash wassampled to test for living mycelium and themoisture content determined.
Survival of mycelia after air drying
The following species were investigated: Aegeritasp., Clathrosphaerina zalewskii Beverwijk, Helicosporium sp., Ho. fuscosporum Linder, Hd. conglomeratum Glen-Bott, Hd. giganteum Glen-Bott,Hd. hyalinum Linder, Hd. luteo-album Glen-Bott,Hd. triglitziense (Jaap) Linder, Hd. tubulosum(Riess) Linder, Hd. westerdijkiae Beverwijk,Helicodendron sp.nov.
The fungi were isolated from decaying leaf litterrecovered from static water sources in woodland inSouth Devon (Fisher & Webster, 1978).
All species were grown in beech leaf decoctionand prepared as described by Fisher & Webster(1978). After 4 weeks incubation at 20 ° themycelia were harvested on a 0'5 mm plastic sieveand washed with sterile distilled water. Eachmycelium was placed on dry filter paper in 9 erndiam Petri dishes and air dried at 20°. At the timeof harvest samples of the mycelia were incubatedin moist chambers to confirm viability at the beginning of the experiment.
At intervals sections of the dried mycelia werecut out, submerged under sterile distilled water inPetri dishes and incubated for 2 weeks at 20°,
after which the mycelia were recovered andincubated in moist chambers as previously described. Mycelia that showed active growth without sporulation were transferred on to 0'1 % maltextract agar for a further incubation period of10 days. In all cases sporulation was taken as thecriterion for survival.
The moisture content at the time of samplingwas determined by drying portions of the myceliato constant weight at 95°.
Survival of conidia after desiccation
The following species were investigated: Aegeritasp., C. zalewskii Beverwijk, Ha.fuscosporum Linder,Hd. conglomeratum Glen-Bott, Hd. giganteumGlen-Bott, Hd. hyalinum, Linder, Hd. tubulosum(Riess) Linder and Hd. westerdijkiae Beverwijk.Isolations were made as previously described.
The fungi were grown on sterile beech leafdisks as described by Fisher & Webster (1978).After a period of 4 weeks growth all disks wereincubated in moist chambers for sporulation. Tendisks from each species bearing spores were thenplaced over self-indicating silica gel in a desiccatorat 20° and after 10 days desiccation approximately50 spores from each species were streaked on to0'1 % malt extract agar and incubated at 20°.
Mycelial growth followed by sporulation was takenas criterion for spore survival.
The moisture contents of spores of Hd. giganteum before and after desiccation were determinedin the following manner: spore deposits 1-2 mmthick were gently removed with the blunt side ofa scalpel from two r-year-old cultures, distributedinto four aluminium foil boats, weighed and placedin a desiccator.
After 10 days three boats were weighed todetermine weight after desiccation and then driedto constant weight at 95° to determine the watercontent. The spores in the remaining boat wereused to test viability after desiccation.
RESULTS
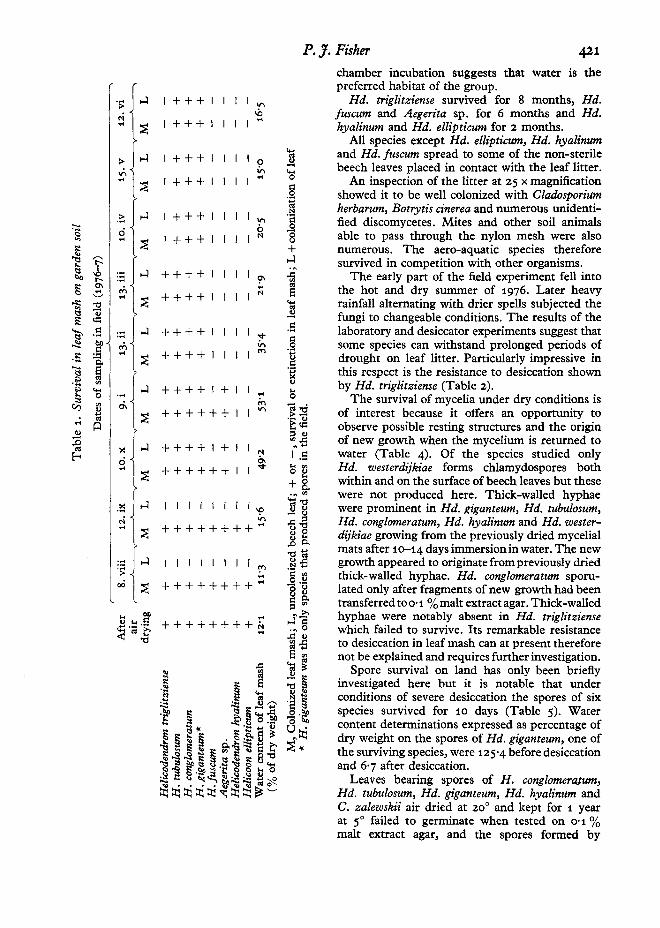
All species grown in leaf mash survived an initialdrying period of 7 days at prevailing laboratorytemperatures (Tables 1, 2, 3). Helicodendrontubulosum, Hd. conglomeratum and Hd. giganteumsurvived in mash after 11 months contact withgarden soil. Only Hd. giganteum produced sporeson the litter surface during this period but noneof these germinated when transferred on to 0'1 %malt extract agar, and were probably dead. Lack ofsporulation by the remaining species after prolonged periods on land, and renewed sporulationafter a period under water and subsequent moist
P.J. Fisher 421
++++++++~...
I I I I I I I I ~...
::E ++++++++ ...
chamber incubation suggests that water is thepreferred habitat of the group.
Rd. triglitziense survived for 8 months, Rd.fus cum and Aegerita sp. for 6 months and Rd.hyalinum and Rd. ellipticum for 2 months.
All species except Rd. ellipticum, Rd. hyalinumand Rd. fuscum spread to some of the non-sterilebeech leaves placed in contact with the leaf litter.
An inspection of the litter at 25 x magnificationshowed it to be well colonized with Cladosporiumherbarum, Botrytis cinerea and numerous unidentified discomycetes. Mites and other soil animalsable to pass through the nylon mesh were alsonumerous. The aero-aquatic species thereforesurvived in competition with other organisms .
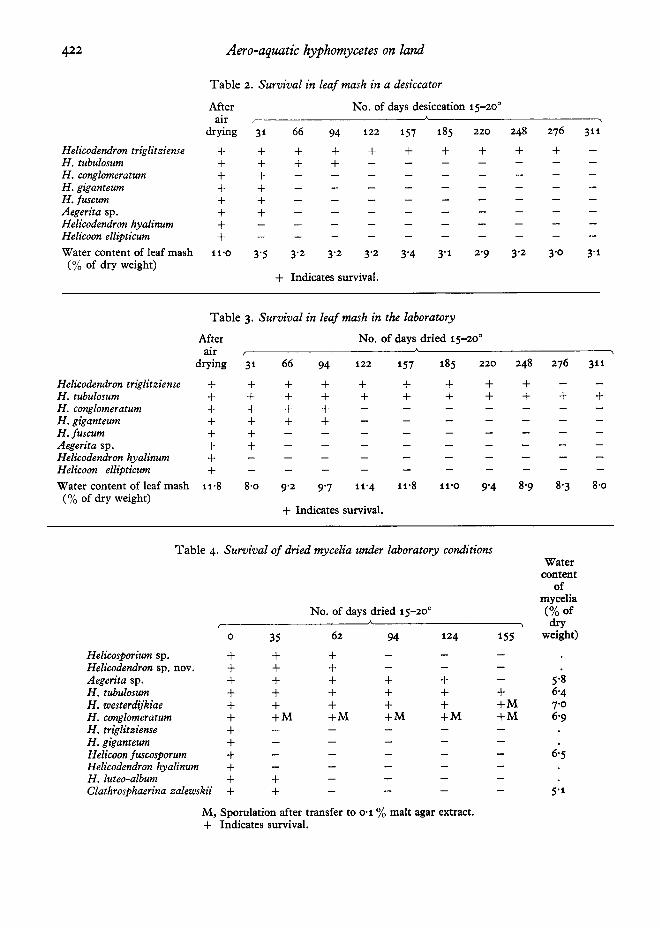
The early part of the field experiment fell intothe hot and dry summer of 1976. Later heavyrainfall alternating with drier spells subjected thefungi to changeable conditions. The results of thelaboratory and desiccator experiments suggest thatsome species can withstand prolonged periods ofdrought on leaf litter. Particularly impressive inthis respect is the resistance to desiccation shownby Rd. triglitziense (Table 2).
The survival of mycelia under dry conditions isof interest because it offers an opportunity toobserve possible resting structures and the originof new growth when the mycelium is returned towater (Table 4). Of the species studied onlyRd. westerdijkiae forms chlamydospores bothwithin and on the surface of beech leaves but thesewere not produced here. Thick-walled hyphaewere prominent in Rd. giganteum, Rd. tubulosum,Rd. conglomeratum, Rd. hyalinum and Rd. westerdijkiae growing from the previously dried mycelialmats after 10-14 days immersion in water. The newgrowth appeared to originate from previously driedthick-walled hyphae. Rd. conglomeratum sporulated only after fragments of new growth had beentransferred to 0'1 %malt extract agar.Thick-walledhyphae were notably absent in Rd. triglitziensewhich failed to survive. Its remarkable resistanceto desiccation in leaf mash can at present thereforenot be explained and requires further investigation.
Spore survival on land has only been brieflyinvestigated here but it is notable that underconditions of severe desiccation the spores of sixspecies survived for 10 days (Table 5). Watercontent determinations expressed as percentage ofdry weight on the spores of Rd. giganteum, one ofthe surviving species, were 125'4 before desiccationand 6'7 after desiccation.
Leaves bearing spores of R . conglomeratum,Rd. tubulasum, Rd. giganteum, Rd. hyalinum andC. zalewskii air dried at 20° and kept for 1 yearat 5° failed to germinate when tested on 0'1 %malt extract agar, and the spores formed by
V'>bN
...;..,V'>
o
'"...
IV'>IQ
I'"++ +
1++ + 1
1+++
1++ +1
1+++
1+++1
++++
...:l ++++1+
...:l ++ + +
...:l ++ + +
...:l ++++1+
...:l r I I I I I I I 'flV'>
::E ++++++++'"
::E ++++
::E ++++++
::E ++++++
l<
o...
m...
m...
>V'>...
'~,~
';:00
.:Sri...
.:::o...
422 Aero-aquatic hyphomycetes on land
T able 2 , S urvival in leaf mash in a desiccator
After No. of days desiccation 15-20°air
-----drying 31 66 94 122 157 185 220 248 276 311
H elicodendron triglitziense + + + + + + + + + +H, tubulosum + + + +H . conglomeratum + +H, giganteum + +H i fus cum + +A egerita sp. + +H elicodendron hy alinum +H elicoon ellipticum +Water content of leaf mash 11'0 3'5 3'2 3'2 3'2 3'4 3'1 2'9 3'2 3'0 3'1(% of dry weight)
+ Indicates survival.
Table 3, Survival in leaf mash in the laboratory
After No, of days dried 15-20°air
drying 31 66 94 122 157 185 220 248 276 311
H elicodendron triglit ziense + + + + + + + + +H , tubulosum + + + + + + + + + + +H . conglomeratum + + + +H. giganteum + + + +Hi fuscum + +A egerita sp , + +H elicodendron hyalinum +H elicoon ellipticum +Water content of leaf mash 11·8 8'0 9'2 9'7 11'4 11'8 11'0 9'4 8'9 8'3 8'0
(% of dry weight )+ Indicates survival.
Table 4, Survival of dried mycelia under labo ratory conditionsWater
contentof
myceliaNo. of days dried 15-20 ° (% of
r-- dry0 35 62 94 124 155 weight)
Helicosporium sp, + + +H elicodendron sp, nov, + + +A egerita sp. + + + + + 5'8H . tubulosum + + + + + + 6'4H. westerdij k iae + + + + + +M 7'0H , conglomeratum + +M + M +M +M +M 6'9H . triglitziense +H . giganteum +H elicoon fuscosporum + 6'5H elicodendton hy alinum +H , luteo-album + +Clathrosphaerina z alewski i + + 5'1
M, Sporulation after transfer to 0'1 % malt agar extract,+ Indicates survival.

P.J.Fisher
Table 5. Survival of spores in a desiccator
Period of desiccationdays at 15-20 0
A
10 20
Helicodendrongiganteum +Hd, westerdijkiae +Hd. conglomeratum +Helicoon[uscosporum +Clathrosphaerina zalewskiiHelicodendrum tubulosumAegerita sp, +Helicodendron hyalinum +
+ Indicates survival.
Hd. giganteum sometime during the field experiment were, as previously mentioned, unable togerminate.
This evidence suggests that spore survival onland may be very limited and that the floatingpropagules of many of the aero-aquatic fungi maybe more important in spreading species from onestatic water habitat to another.
The survival of many of these fungi in leaf litteron land and their considerable resistance to dryingwould enable them to survive during periods ofdrought and assist their spread via wind blownleaffragments. This method of spread could explaintheir widespread distribution in many isolatedstatic waters (Gonczol, 1976).
The records of land colonization cited in theintroduction of this paper suggest that certainmembers of this group commonly grow on land.Their presence on land may be explained by thesemi-aquatic nature of many terrestrial habitats intemperate zones in areas of high rainfall.
Fisher &Webster (1978) observed that the groupwas particularly abundant in shallow water-filledchannels where flooding may be intermittent andwhere the leaf litter layer is only a few centimetresbelow the water surface. During dry spells thelitter layer may become exposed to the atmosphere.
The experimental results reported here suggestthat mycelia present in infected wind blown leavesfrom such sources when resubmerged in only a fewmillimetres of water could readily regenerate. Suchfoci of infection could spread under a thin layer ofwater to surrounding leaves and soil far removedfrom any permanent water source. Thus the morepermanent aquatic habitats of this group may beextended during periods of high rainfall allowingsome species to colonize leaf litter and soil surfaces
on land. Land may thus represent a marginalhabitat for the majority of the group.
I wish to thank Mr S. K. Abdullah for therecords of aero-aquatic hyphomycetes recoveredfrom soil on Dartmoor.
REFERENCES
BANDONI, R. J. (1972). Terrestrial occurrence of someaquatic hyphomycetes. Canadian Journal of Botany50, 2283-2288.
DESCALS, E. & WEBSTER, J. (1976). Hyaloscypha:Perfect State of Clathrosphaerina zalewskii. Transactions of the British Mycological Society 67,525-528.
FISHER, P. J. (1977). New methods of detecting andstudying saprophytic behaviour of aero-aquatichyphomycetes from stagnant water. Transactions ofthe British Mycological Society 68, 407-411.
FISHER, P. J. & WEBSTER, J. (1978a). The effect ofoxygen and carbon dioxide on the growth of fouraero-aquatic hyphomycetes. Transactions of theBritish Mycological Society. (In the Press.)
FISHER, P. J. & WEBSTER, J. (1978b). Sporulation ofaero-aquatic fungi under different gas regimes inlight and darkness. Transactions of the BritishMycological Society 71, 465-468.
GLEN-BoTT, J. I. (1951). Helicodendron giganteumsp, nov. and other aerial sporing hyphomycetes ofsubmerged dead leaves. Transactions of the BritishMycological Society 34, 275-279.
GLEN-BoTT, J. I. (1955). On Helicodendron tubulosumand some similar species. Transactions of the BritishMycological Society 38, 17-30.
GONCZOL, J. (1976). Ecological observations on theaquatic hyphomycetesof Hungary, II. Acta BotanicaAcademiae Scientiarum Hungaricae 22, 51-60.
LINDER, D. H. (1929). A monograph of the helicosporous fungi imperfecti. Annals of the MissouriBotanical Gardens 16, 227-388.
PARK, D. (1974b). Aquatic Hyphomycetes in nonaquatic habitats. Transactions of the British Mycological Society 63, 183-187.
VAN BEVERWIJK, A. L. (1951a). Zalewski's Clathrosphaera spirifera. Transactions of the British Mycological Society 34, 280-29°.
VAN BEVERWIJK, A. L. (1951b). Candelabrum spinulosum, a new fungus species. Antonie van Leeuwenhoek 17,278-284.
VAN BEVERWIJK, A. L. (1953). Helicosporous Hyphomycetes. I. Transactions of the British MycologicalSociety 36, 111-124.
VAN BEVERWIJK, A. L. (1954). Three new fungi:Helicoonpluriseptatum n.sp., Papulaspora pulmonarian.sp. and Tricellula inaequalis n.gen. n.sp. Antonievan Leeuwenhoek 20, 1-16.
WAID, J. S. (1954). Occurrence of aquatic hyphomycetesupon root surfacesof beech grown in woodland soils. Transactions of the British MycologicalSociety 37, 420-421.
(Accepted for publication 10 May 1978)

![Review Article Gammarus-MicrobialInteractions:AReviewhyphomycetes was negligible [ 17]. Aquatic hyphomycetes produce secondary metabolites that function in microbe-microbe interactions](https://img.dokumen.tips/doc/110x75/60c9d3a89f80e602464161fa/review-article-gammarus-microbialinteractionsareview-hyphomycetes-was-negligible.jpg)



























