Embed Size (px)
Citation preview

364
Biochimica et Biophysica Acta, 467 (1977) 364--375 © Elsevier/North-Holland Biomedical Press
BBA 77718
STIMULATORY AND INHIBITORY EFFECTS OF SULFHYDRYL REAGENT ON p-AMINOHIPPURIC ACID TRANSPORT BY ISOLATED RENAL TUBULES
E.F. BOUMENDIL-PODEVIN and R.A. PODEVIN
Unitd de Recherches de Ndphrologie Normale et Pathologique U64 de I'I.N.S.E.R.M., H~pital Tenon, 75020 -Paris (France)
(Received December 22nd, 1976)
Summary
Isolated tubules from rabbit kidney cortex were treated with several differ- ent sulfhydryl reagents in an at tempt to determine whether sulfhydryl groups are involved in organic acid transport. Disulfide reagents such as sodium tetra- thionate and 6,6'-dithionicotinic acid were found to exert a biphasic effect on p-aminohippuric acid transport, i.e. transient stimulation followed by inhibi- tion. In contrast, t reatment of tubules with the mercaptide-forming reagent, p- chloromercuribenzoate, caused only inhibition of organic acid transport. Treat- ment of tubules with reductants such as dithiothreitol or mercaptoethanol blocked the stimulatory effect of tetrathionate without affecting the inhibitory effect of this oxidant. The inhibition caused by p-chloromercuribenzoate, how- ever, was largely reversible when tubules were treated with reductants. The results suggest that the renal organic acid transport system contains sulfhydryl groups and that its activity is increased when some of these groups are oxidized.
Introduct ion
The importance of SH groups in the transport of sugars and amino acids across biological membranes has been well established [1--6]. The use of differ- ent sulfhydryl reagents has helped to identify and characterize these groups. With regard to the effects of sulfhydryl reagents on organic acid transport, the only well documented studies were concerned with diuretic and non-diuretic mercurials. In these observations the depression of renal organic acid transport [7--9] was attributed to an effect on tissue metabolism rather than a direct
Abbreviation: PCMB, p-chloromercuribenzoate.

365
action on organic acid transport. In this report we have compared the effects of a mercurial reagent to those of disulfide reagents on the accumulation of p- aminohippuric acid by isolated rabbit renal tubules.
The results indicated that, in function of time, the action of the disulfide reagents, sodium tetrathionate and 6,6'-dithionicotinic acid was biphasic i.e. a transient stimulation followed by an inhibition of p-aminohippuric acid uptake, whereas with the mercurial, p-chloromercuribenzoate (PCMB), only the latter action could be detected. Our experiments also demonstrated that the activa- tion of p-aminohippuric acid transport by tetrathionate could be blocked by the reductants dithiothreitol and mercaptoethanol. This report presents these results and other experiments describing the different effects of PCMB and tetrathionate on electrolyte and organic acid transport in isolated renal tubules.
Materials and Methods
Chemicals Bacterial collagenase was purchased from Worthington, bovine serum albu-
min and dithiothreitol from Calbiochem, sodium tetrathionate was from Fluka and was recrystallized as described by Gilman [10]; PCMB was from Aldrich. 6,6.-dithi0nicotinic acid was from Newcell Biochemicals, p-amino-[1-14C]hip- puric acid, 20--40 Ci/M, and [Me-3H]inulin 5 Ci/mM, were from New England Nuclear Corporation. All other chemicals were reagent grade and were obtained from Merck-Darmstadt.
Methods Suspensions of rabbit renal cortical tubules were prepared according to Burg
and Orloff [11]. The acetate Ringer solution used for the preparation and incu- bation of the isolated tubules contained: 135 mM NaC1; 10 mM KC1, 10 mM sodium acetate, 1 mM MgSO4, 1 mM CaC12, 0.5 mM Call4 (PO4)2, 20 mM Tris • HC1, (pH 7.4 at 25°C), 5 mM glucose, with 1.5% wt./vol bovine serum albumin.
Treatment of the tubules with the different sulfhydryl reagents (tetrathion- ate, 6,6'-dithionicotinic acid, PCMB) was performed in conical centrifuge tubes as follows: the investigated chemical was incubated in acetate Ringer solution containing 3% bovine serum albumin for 30 min to allow time for an equilibri- um of the reagent with the albumin to be reached. The reaction was started by adding an equal volume (usually 5 ml) of bovine serum albumin-free suspension of tubules. Incubation was carried out for the stated time with a gas phase con- taining 100% 02. The reaction was terminated by dilution and centrifugation at 50 × g for 2 min. Control tubules were treated as above but omitting the sulf- hydryl reagent. In experiments designed to test the reversibility of the effects of sulfhydryl reagents on p-aminohippuric acid and electrolyte transport, treated and control tubules were incubated with and without 84 mM mercapto- ethanol or 20 mM dithiothreitol for 5 min in an atmosphere of 100% N2. The reaction was ended as described above. All treatments were carried out at 25 ° C. The subsequent incubations with p-aminohippuric acid were performed in ves- sels described by Burg and Orloff [11] at 25°C with a gas phase containing 100% 02. The final suspension contained from 2 to 5% of tissue (wet wt./vol.). The final concentration of p-amino [ 1-14C] hippuric acid in the bathing medium

366
was 0.15 mM. [Me-3H]inulin was also added giving a final radioactivity of about 1 pCi/100 ml. After completion of incubation, the suspension was trans- ferred to a special centrifuge tube maintained at 0°C [12]. Centrifugation was performed for 5 min at 5000 × g in a refrigerated centrifuge. The supernate was decanted, the superficial layer of cells removed by suction, and the remain- ing tissue plugs were weighed before and after dessication at 70 ° C.
In experiments designed to examine the effects of sulfhydryl reagents on p- aminohippuric acid uptake in the absence of active electrolyte transport slices of rabbit renal cortex were used in place of isolated tubules as attempts to pre- pare (Na*, K÷)-depleted and ouabain-poisoned separated tubules resulted in fragmentation of most of the cells. Slices of cortex approximately 0.4 mm thick were prepared with a Stadie-Riggs microtome. They were leached for 80 min at 25°C in the following leaching solution: 145 mM choline chloride, 20 mM Tris. HC1 (pH 7.3) and 2 mM ouabain, as recently reported [13]. Treat- ment of these ouabain-poisoned slices with sulfhydryl reagents was performed for the times indicated at 25°C in the leaching solution. The reaction was ter- minated by transferring the suspension of slices to a beaker covered with surgi- gal gauze. The slices were washed free of reagents by pouring leaching solution over the pieces of tissue. Control ouabain-poisoned slices were treated as above but omitting the sulfhydryl reagent. Tissues were rapidly blotted and 4--6 slices (30--60 mg wet weight) were transferred for 20 min at 25°C to 5 ml of p- amino-[1-14C]hippuric acid (0.14 r aM)and [Me-3H] inulin containing solution identical in composition to that used for leaching or modified by replacing the choline chloride with an equivalent amount of NaC1. The gas phase was 100% N2. Preparation of the samples for analysis was similar for the separated tubules and slices [14].
The radioactivities of ~4C and 3H of the tissue extracts and of the incubation media were measured simultaneously, each in duplicate, in a Mark 1 Nuclear Chicago liquid scintillation counter, as previously described [15]. The p-amino- hippuric acid in the incubation media was measured using the method of Smith et al. [16]. Na* and K ÷ analyses were performed using a flame photometer (Netheler and Hinz, Gmbh, Hamburg, G.F.R.). Tissue contents of p-aminohip- puric acid, water, and electrolytes were corrected for extracellular contamina- t ion by using the inulin space measured for each sample [14].
The accumulation of p-aminohippuric acid was usually expressed as the dis- tribution ratio, which is the ratio of organic acid concentration in cellular water to that in the medium.
Results were expressed as means -+ S.D. Statistical analyses were performed by the Student 's t test.
Results
Effect o f S H reagent concentration on p-aminohippuric acid and electrolyte transport
The results in Figs. 1 and 2 compare the effects of two sulfhydryl reagents, PCMB and tetrathionate, on p-aminohippuric acid and electrolyte transport by isolated renal tubules. With a reagent exposure time of 20 min followed by a 30 min incubation period with p-aminohippuric acid alone, both the mercaptide-

367
0
43 2
O~
300
2oo
100
~ 0
8
6
c~
Q
300~
~ 2 0 0 E
100
,e
i i i i J I
{] 0'.1 13.2 0*..5 i~ ~] 131 0'7_ 0'.~ 0~. 0'5 P C M B ('m M} Tef;rathionet~ [raM)
Fig. 1. I n h i b i t i o n of p - a m i n o h i p p u r i c ac id a n d e l ec t ro ly t e t r a n s p o r t as a f u n c t i o n of PCMB c o n c e n t r a t i o n . Tubu l e s were t r e a t ed wi th the s p e c i f i e d c o n c e n t r a t i o n s of PCMB fo r 20 m i n as i n d i c a t e d u n d e r Mater ia l s
and M e t h o d s . I n c u b a t i o n wi th labe led p - a m i n o h i p p u r i c acid was fo r 30 m i n . Each p o i n t is the m e a n of
th ree e x p e r i m e n t s . Ver t i ca l l ines d e n o t e +- 1 S.D. Absence of the ve r t i ca l l ine ind ica te s t h a t S.D. was negli-
gible.
Fig. 2. I n h i b i t i o n of p - a m i n o h i p p u r i c ac id and e l ec t ro ly t e t r a n s p o r t as a f u n c t i o n of t e t r a t h i o n a t e concen- t r a t ion . E x p e r i m e n t a l c o n d i t i o n s w e r e iden t ica l w i t h those in Fig. 1 e x c e p t t h a t t e t r a t b i o n a t e r ep laced
PCMB.
forming reagent, PCMB, and the oxidizing reagent, tetrathionate, caused inhibi- t ion of p-aminohippuric acid and electrolyte transport. Figs. I and 2 show that whereas the shape of the curve relating organic acid transport to sulfhydryl rea- gent concentration was comparable whether PCMB or tetrathionate was used, a different pattern emerged for the effects of these two reagents on electrolyte content. As can be seen in Fig. 1, the net loss of K ÷ induced by PCMB was almost exactly compensated by the increase in tissue Na ÷, whereas tetrathion- ate caused a net gain of Na ÷, 2--5-times higher than the tissue K + loss. At least in part, this difference may be accounted for by the enlargement of the intra- cellular space which was significantly higher with tetrathionate than with PCMB (results not shown).
In these experiments, the suLfhydryl reagents were added to the Ringer solu- tion containing bovine serum albumin 30 min prior to the addition of tubules. Since these reagents react with albumin, the concentration of reagent to which
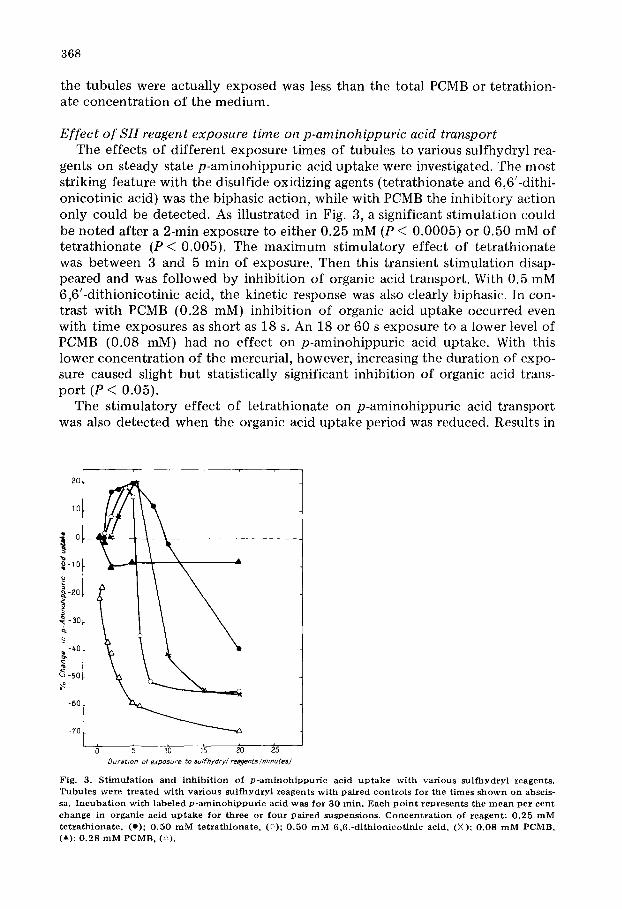
368
the tubules were actually exposed was less than the total PCMB or tetrathion- ate concentration of the medium.
Effect o f SH reagent exposure time on p-aminohippuric acid transport The effects of different exposure times of tubules to various sulfhydryl rea-
gents on steady state p-aminohippuric acid uptake were investigated. The most striking feature with the disulfide oxidizing agents (tetrathionate and 6,6'-dithi- onicotinic acid) was the biphasic action, while with PCMB the inhibitory action only could be detected. As illustrated in Fig. 3, a significant stimulation could be noted after a 2-min exposure to either 0.25 mM (P < 0.0005) or 0.50 mM of tetrathionate ( P < 0.005). The maximum stimulatory effect of tetrathionate was between 3 and 5 min of exposure. Then this transient stimulation disap- peared and was followed by inhibition of organic acid transport. With 0.5 mM 6,6'-dithionicotinic acid, the kinetic response was also clearly biphasic. In con- trast with PCMB (0.28 mM) inhibition of organic acid uptake occurred even with time exposures as short as 18 s. An 18 or 60 s exposure to a lower level of PCMB (0.08 raM) had no effect on p-aminohippuric acid uptake. With this lower concentration of the mercurial, however, increasing the duration of expo- sure caused slight but statistically significant inhibition of organic acid trans- port (P < 0.05).
The stimulatory effect of tetrathionate on p-aminohippuric acid transport was also detected when the organic acid uptake period was reduced. Results in
2O
10
-~0~ 2
~ -30
-~0
-50
-70
Duratlon of exposure to ~ulfhydry/ r~n~ /minutes}
Fig, 3. S t i m u l a t i on and inh ib i t ion of p - a m i n o h i p p u r i c acid u p t a k e wi th var ious su l fhyd ry l reagents . Tubu les were t r ea t ed wi th var ious su l fhydry l reagents wi th pa i red con t ro l s for the t imes s h o w n on abscis- sa. I n c u b a t i o n wi th labeled p - a m i n o h i p p u r i c acid was for 30 rain. Each p o in t r ep resen t s the m e a n p e r cent change in organic acid u p t a k e for three or fou r pa i red suspensions, C o n c e n t r a t i o n of reagent : 0 .25 m M t e t r a t h i o n a t e , (e) ; 0 ,50 m M t e t r a t h i o n a t e , (;~); 0 .50 m M 6,6 . -d i th ionico t in ic acid, (X); 0 .08 m M PCMB, (~,); 0 .28 m M PCMB, (A).

369
25
12
"~ 0.5
.,y,
'~ 0 300
~ 200
1 ~.~oo
¢e ~. o
___¢? i I
" 6o 7~a 1;o ~ , "ne , s e c
Fig. 4. S t imu la t ion of p - a m i n o h i p p u r i c acid and Na + t r a n s po r t by t e t r a th iona t e . Tubu les were t r ea ted w i th or w i t h o u t 0 .25 rnM t e t r a t h i o n a t e for 5 m in at 25°C. Cont ro l and t r e a t ed tubules were then i n cu b a t ed wi th labeled p - a m i n o b i p p u r i e acid for the t imes s h o w n on abscissa. Organic acid u p t ak e : con t ro l , (o); t e t r a t h i o n a t e , (e) . Tissue K+: cont ro l , (o); t e t r a t h i o n a t e , ( - ) . Tissue Na+: con t ro l , (A); t e t r a th iona t e , (1). Each s y m b o l r ep resen t s the m e a n of th ree e p x e r i m e n t s and ver t ica l lines d e n o t e -+ 1 S.D. When S.D. is n o t s h o w n it did no t e x t e n d b e y o n d the symbols .
Fig. 4 demonstrate that tetrathionate caused statistically significant stimulation of p-aminohippuric acid accumulation (P < 0.01) at the earliest time period that can be reasonably investigated with the method used, i.e., 65 s. Of interest is the observation that the stimulatory effect of tetrathionate on p-aminohip- puric acid uptake was associated with reduction in tissue Na ÷ content. For all the incubation times investigated tetrathionate treated tubules contained signi- ficantly less Na ÷ {0.0005 < P < 0.0025) than paired control tubules. The data of Fig. 4 also show that K ÷ tissue content is higher in tetrathionate-treated tubules compared to controls. This increase, however, was not large enough to demonstrate statistically. These effects occurred without significant changes in the cell water content. Mean tissue water content was 2.02 +- 0.067 and 2.04 +- 0.051 kg/kg dry wt. for control and treated tubules, respectively. Each value represents the mean +- S.D. of 12 tubule suspensions.
Reversibility of the effects of SH reagents on p-aminohippuric acid transport Thiol compounds reversed the stimulation of p-aminohippuric acid uptake

370
T A B L E I
R E V E R S I B I L I T Y O F T H E S T I M U L A T O R Y E F F E C T O F T E T R A T H I O N A T E O N p - A M I N O H I P P U R I C A C I D T R A N S P O R T BY I S O L A T E D R E N A L T U B U L E S
T r e a t m e n t s were as indicated under Materials and Methods . Incubat ions w e r e carried o u t w i t h 0 . 1 5 m M
labeled p - a m i n o h i p p u r i c acid for the spec i f i ed t imes . Each value represents the m e a n + S.D. of e i ther 6
( E x p t . 1) or 3 ( E x p t s . 2 a n d 3) e x p e r i m e n t s .
Expt . First t r e a t m e n t S e c o n d t r e a t m e n t I n c u b a t i o n Dis tr ibut ion Percent No. (5 r a in ) (5 r a in ) t i m e ( m i n ) ratio change
1 N o n e N o n e 2 2 . 8 8 ± 0 . 2 8
N o n e Di th io thre i to l , 2 0 m M 2 2 .71 ~: 0 . 1 3 - -6
Tetra th ionate , 0 . 2 5 m M N o n e 2 3 . 8 9 + 0 . 1 8 * 35
T e t r a t h i o n a t e , 0 . 2 5 m M D i t h i o t h r e i t o l , 2 0 m M 2 3 .13 + 0 . 1 5 9
2 N o n e N o n e 3 0 4 . 8 2 ~- 0 . 1 2
N o n e Di tb io thre i to l , 2 0 m M 3 0 4 . 4 8 + 0 . 1 6 ** - -7
N o n e M e r c e p t o e t h a n o l , 8 4 m M 30 4 . 2 5 -+- 0 .21 * * - - 1 1 . 8
Tetra th ionate , 0 .5 m M N o n e 30 6 . 0 5 ± 0 .16 * 2 5 . 5
Tetra th ionate , 0 .5 m M D i t h i o t h r e i t o l , 2 0 m M 3 0 4 . 7 9 +- 0 . 0 9 - 4 ) . 6
Tetra th ionate , 0 .5 m M M e r c a p t o e t h a n o l , 8 4 m M 3 0 4 . 8 7 +- 0 . 0 7 1
3 N o n e N o n e 3 0 4 . 9 7 -2 0 . 3 4
N o n e Di th io thre i to l , 2 0 m M 30 5 . 0 8 +- 0 . 2 4 2 .3
N o n e M e r c a p t o e t h a n o l , 8 4 m M 30 5 .19 ± 0 . 4 0 4 .4
Tetra th ionate , 0 . 2 5 m M N o n e 3 0 5 .93 +- 0 . 4 3 ** 1 9 . 2
Tetra th ionate , 0 . 2 5 m M D i t h i o t h r e i t o l , 2 0 m M 30 5 .07 -+ 0 .21 2
Tetra th ionate , 0 . 2 5 m M M e r c a p t o e t h a n o l , 8 4 m M 3 0 4 . 7 5 -+ 0 . 0 9 - - 4 . 4
• P ~ 0 . 0 0 0 5 .
• * P <: 0 . 0 5 vs. respect ive c on tr o l values.
by tetrathionate. Dithiothreitol and mercaptoethanol almost completely aboli- shed the tetrathionate effect (Table I). This reversal could not be attributed on an inhibition caused by the thiol compounds, since these reagents exerted either no statistically significant effect on p-aminohippuric acid transport (Expts. 1 and 3) or an inhibitory effect that was small compared to the stimula- tory effect of tetrathionate (Expt. 2).
Thiol compounds substantially antagonized the depressive effect of PCMB on organic acid and electrolyte transport. As summarized in Table II, both mer- captoethanol and dithiothreitol reversed by approximately 70% the inhibition of organic acid uptake by PCMB. The reversal of the effect on electrolyte trans- port was more marked, 93% for K ÷ and 86% for Na ~. In contrast, thiol com- pounds did not antagonize substantially the depression of p-aminohippuric acid uptake by tetrathionate (Table II). The only apparent effect of thiol treatment was the fall in tissue Na ÷ content.
Effects of SH reagents on p-aminohippuric acid uptake by (Na ÷, K÷)-depleted and ouabain-poisoned slices
As recently reported [13], the imposition of an extracellular to intracellular Na ~ gradient in the absence of active electrolyte transport induces a stimulation of p-aminohippuric acid uptake. The data in Table III show that the Na÷-depen- dent anaerobic entrance of p-aminohippuric acid is inhibited significantly by PCMB. In Expt. 1 a 20 min exposure to a 0.4 and 2.2 mM level of PCMB
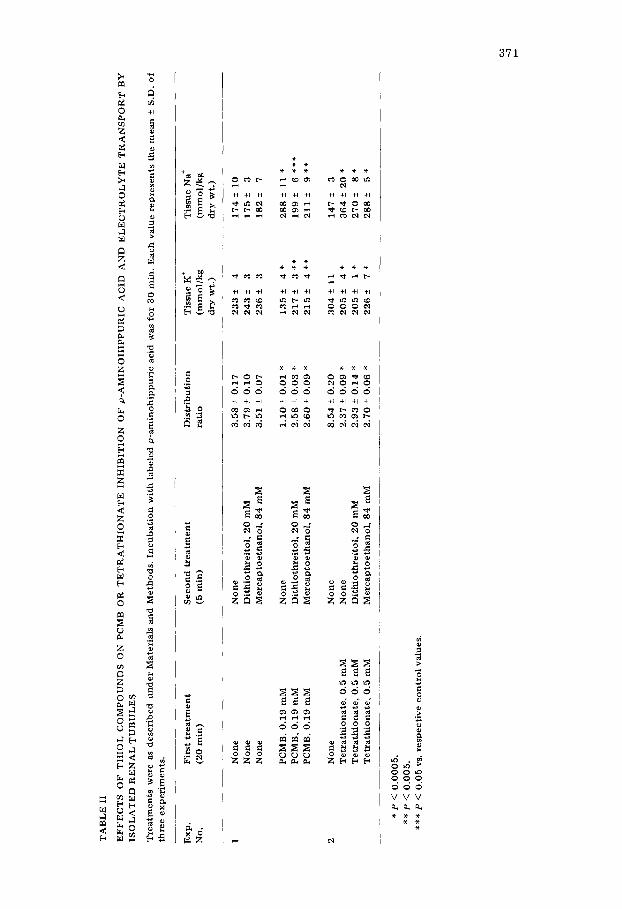
TA
BL
E
II
EF
FE
CT
S
OF
T
HIO
L
CO
MP
OU
ND
S
ON
P
CM
B
OR
T
ET
RA
TH
ION
AT
E
INH
IBIT
ION
O
F
p-A
MIN
OH
IPP
UR
IC
AC
ID
AN
D
EL
EC
TR
OL
YT
E
TR
AN
SP
OR
T
BY
" IS
OL
AT
ED
R
EN
AL
T
UB
UL
ES
Tre
atm
en
ts
wer
e as
de
scri
be
d
un
de
r M
ate
ria
ls a
nd
M
eth
od
s.
Inc
ub
ati
on
w
ith
la
be
led
p-a
min
oh
ipp
uri
c
ac
id
wa
s fo
r 3
0 r
ain
. E
ac
h v
alu
e r
ep
rese
nts
th
e m
ea
n
± S
.D.
of
thre
e e
xp
eri
me
nts
.
Ex
p.
Fir
st t
rea
tme
nt
Se
co
nd
tr
ea
tme
nt
Dis
trib
uti
on
T
issu
e K
+
Tis
sue
Na
+
No
. (2
0 r
ain
) (5
ra
in)
rati
o
(mm
ol/
kg
(m
mo
l/k
g
dry
w
t.)
dry
wt.
)
1 N
on
e
No
ne
3
.58
± 0
.17
No
ne
D
ith
ioth
reit
ol,
2
0 m
M
3.7
9 ±
0.1
0
No
ne
M
erc
ap
toe
tha
no
l,
84
ra
m
3.5
1
-+ 0
.07
PC
MB
, 0
.19
m
M
No
ne
1
.10
±
0.0
1
PC
MB
, 0
.19
m
M
Dit
hio
thre
ito
l,
20
mM
2
.58
+-
0.0
3
PC
MB
, 0
.19
m
M
Me
rca
pto
eth
an
ol,
8
4 m
M
2.6
0
+ 0
.09
2 N
on
e
No
ne
8
.54
±
0.2
0
Te
tra
thio
na
te,
0.5
mM
N
on
e
2.3
7
± 0
.09
Te
tra
thio
na
te,
0.5
mM
D
ith
ioth
reit
ol,
2
0 m
M
2.9
3
-+ 0
.14
Te
txa
thio
na
te,
0.5
mM
M
erc
ap
toe
tha
no
l,
84
m
M
2.7
0
± 0
.06
* P
<
0.0
00
5.
**
P
<
0.0
05
. *
**
P
<
0.0
5 v
s. r
esp
ec
tiv
e c
on
tro
l v
alu
es.
23
3
+
4 1
74
+
1
0
24
3
+-
3 1
75
±
3
23
6
+
3 1
82
±
7
*
13
5±
4
"
28
8±
1
1 *
*
21
7 +
- 3
"*
1
99
+-
6 *
**
*
21
5±
4 '*
*
21
1+
- 9 *
*
30
4
± 1
1
14
7
-+
3
* 2
05
+
4"
36
4±
2
0"
* 2
05
±
1
* 2
70
+-
8"
* 2
26
±
7 *
28
8±
5
"

372
T A B L E I I I
E F F E C T S O F PCMB A N D T E T R A T H I O N A T E ON p - A M I N O H I P P U R I C A C I D U P T A K E BY (Na +, K+) -
D E P L E T E D A N D O U A B A I N - P O I S O N E D S L I C E S
S o d i u m - and p o t a s s i u m - d e p l e t e d and o u a b a i n - p o i s o n e d slices w e r e t r e a t e d f o r 20 m i n in the l each ing solu-
t i on a lone ( con t ro l ) o r fo r the spec i f i ed t imes in the l each ing s o l u t i o n c o n t a i n i n g PCMB (0 .4 - -2 .2 raM), or t e t r a t h i o n a t e (0 .5 - -2 m M ) . When t r e a t m e n t wi th t e t r a t h i o n a t e was less t h a n 20 min , slices were ma in -
t a ined fo r an a p p r o p r i a t e t i m e in the l e ach ing so lu t i on a lone b e f o r e a d d i n g the d i su l f ide r e a g e n t so tha t
the overal l t r e a t m e n t p e r i o d was exac t l y 20 rain. I n c u b a t i o n w i t h labeled p - a m i n o h i p p u r i c acid was for 20 m i n ; the gas phase was 100% N 2. Deta i l s are g iven u n d e r Mater ia ls and Methods . Each value r e p r e se n t s the
m e a n -+ S.D. of 4 - - 1 0 e x p e r i m e n t s .
Exp t . T r e a t m e n t I n c u b a t i o n c o n d i t i o n s D i s t r i b u t i o n ra t io
No.
1 None , con t ro l 145 m M cho l ine ch lo r ide
PCMB 0 .4 mM, 20 ra in 145 m M chol ine ch lo r ide
2.2 mM, 20 ra in 145 m M chol ine ch lor ide
None , c o n t r o l 145 m M NaC1
PCMB 0.4 mM, 20 ra in 145 m M NaC1
2.2 mM, 20 m i n 145 m M NaCl
2 None , c o n t r o l 145 m M cho l ine ch lor ide
T e t r a t h i o n a t e 0.5 mM, 3 m i n 145 m M cho l ine ch lo r ide
0.5 mM, 15 m i n 145 m M chol ine ch lo r ide
None , c o n t r o l 145 m M NaC1
T e t r a t h i o n a t e 0 .5 mM, 3 m i n 145 m M NaCl 0.5 mM, 5 m i n 145 m M NaCl
0 .5 mM, 15 m i n 145 m M NaC1
3 None , con t ro l 145 m M cho l ine ch lo r ide
T e t r a t h i o n a t c 0 .5 mM, 5 m i n 145 m M cho l ine ch lor ide 0.5 mM, 15 ra in 145 m M chol ine ch lo r ide
0.5 mM, 20 m i n 145 m M chol ine ch lo r ide
4 None , con t ro l 145 m M cho l ine ch lo r ide
T e t r a t h i o n a t e 2 mM, 20 m i n 145 m M cho l ine ch lor ide
None , c o n t r o l 145 m M NaCl T e t r a t h i o n a t e
2 raM, 10 m i n 145 m M NaCl 2 mM, 20 rain 145 m M NaC1
* P < 0 ,02.
** P < 0 .002 . *** P < 0 .05 vs. r e spec t ive cont ro ls .
0 .64 + 0 .02
0 .54 ± 0 .05 *
0.48 ± 0.01 * 1.79 ± 0.18
1.07 ± 0.07 **
0 .55 + 0 .04 **
0.57 ± 0 .04
0 . 6 0 ± 0 .05
0 .67 ± 0 .07 *
1.57 ± 0 .13
1,41 ± 0 .14
1.46 ± 0 .06 1 .48 ± 0 .07
0 .52 -+ 0 .03
0 .54 -+ 0 .04
0 .57 ± 0 .06 *** 0.48 -+ 0 .04 **
0.54 ± 0.01
0 ,48 +- 0 .02 * 1.32 -+ 0 .08
1 .20 ± 0 .05 *** 1 .15 ± 0.03 **
decreased the uptake by 40 and 70%, respectively in the presence of NaC1, and only by 16 and 25%, respectively in the presence of choline chloride. This table (Expts. 2 and 3) also shows that t rea tment of ouabain-poisoned slices with 0.5 mM tet ra thionate caused significant stimulation of p-aminohippuric acid uptake in a choline-medium at 15 min. This stimulation ranged from 10 to 17% and was followed by inhibition when the length of exposure to the agent was increased. The lack of te t ra thionate stimulation in the Na ÷ medium could be explained by the fact that the Na ÷ gradient increased p-aminohippuric acid

3 7 3
transport by about 3-fold, thus rendering any further small effect undetectable. With a higher level of tetrathionate (Expt. 4), a modest but significant inhibition of organic acid transport was detected in both the presence or absence of exter- nal Na +.
Discussion
The present work demonstrates that p-aminohippuric acid transport in iso- lated renal tubules can be either stimulated or inhibited by sulfhydryl reagents depending on their nature and on the experimental conditions. With 6,6'-di- thionicotinic acid and tetrathionate, which are known to oxidize specifically -SH groups [17--19], we observed a stimulation of organic acid uptake fol- lowed by an inhibition whereas with the mercaptide-forming reagent, PCMB, only the latter effect could be detected. The tetrathionate stimulation and the PCMB inhibition could be reversed with dithiothreitol or mercaptoethanol but the inhibitory effect of tetrathionate could not be reversed. Several facts suggest that the stimulatory action of 6,6'-dithionicotinic acid and tetrathionate is due to effects of these reagents at the level of the tubular surface membrane. First- ly, stimulation of uptake occurs for very short time exposures of the tubules to these reagents. Secondly, 6,6'-dithionicotinic acid [20], and probably also tetrathionate [21], does not readily enter cell membranes, and thirdly, this effect is detected at the earliest time period that can be investigated with the method used (65 s) suggesting that this oxidant increases the initial rate of p- aminohippuric acid entry.
These results support the view that the p-aminohippuric acid transport sys- tem or part of it might exist in two states: a normal -SH form and an activated disulfide-sulfhydryl form. This view, which can account for the various observed effects, is presented schematically in Fig. 5. In this scheme we con-
~O~-
ACTIVATED STATE INHIBITED 3TATE M~,n~t~b~ I Tetrathtot~te
/9 s l I ,S s s
\ ] -~ 'Mercaptoet2~nol 2 Di~h,o%rhre/to/
NORMAL ,.STATE INHIBITED ,..3TATE
Fig. 5. H y p o t h e t i c a l s cheme to expla in the e f fec t s of va r ious su l fhyd ry l reagen ts on t r a n s p o r t of p - amino - h ippur ic acid by i so la ted renal tubules . F o r details, see Discussion.

374
ceive that the basal state of the organic acid transport system in isolated tubu- les is in a reduced form since its activity is inhibited by the mercaptide-forming agent PCMB. The f inding that the inhibitory process is largely reversed as a result of t reatment with reductants suggests that the main action of PCMB is not secondary to irreversible membrane changes or to reactions of this agent with groups other than sulfhydryl. Tetrathionate [18,19] and possibly 6,6'- dithionicotinic acid [17] oxidize protein -SH groups through a two-step reac- tion. Initially, tetrathionate reacts with superficial -SH groups to form sulfenyl thiosulfates which subsequently react with less accessible -SH groups to form symmetrical disulfides. The first step is a reversible process. The second step, in contrast, has been found to be either hardly reversible [22] or not at all [19]. Thus, the reversible conversion of the basal form of the organic acid transport system to the activated form is most likely to be due to formation of mixed disulfides. Hence, activation could be attributed to limited oxidation of the basal state of the organic acid transport system. The irreversible inhibition of p- aminohippuric acid transport caused by prolonged tetrathionate exposure, on the other hand, could be accounted for by permanent membrane structural alterations secondary to extensive formation of disulfide bonds.
Czech et al. [4] reported that oxidants stimulated the glucose transport sys- tem in brown fat cells whereas reductants inhibited this transport. In the pre- sent study reductants do not affect control p-aminohippuric acid transport rates. However, all the reagents used, including the reductants, were not present during the organic acid uptake period in order to avoid possible intereference between these reagents and p-aminohippuric acid uptake. The possibility exists, therefore, that -SH groups formed as a consequence of t reatment with reduc- tants are reoxidized by 02 and traces of heavy metal ions [23] during the organic acid uptake period, thus masking a possible inhibitory effect.
The data obtained with reductants, oxidants and PCMB are consistent with the notion that a sulfhydryl-disulfide interchange might play a role in the regu- lation of organic acid transport comparable to the mechanism first proposed by Czech et al. [4] for hexose transport by brown fat cells. Although the avail- able data are apparently consistent with the above concept they do not exclude other hypotheses. The present experiments show that the effects of the sulfhy- dryl reagents on p-aminohippuric acid uptake are associated with parallel effects on electrolyte tissue content. Since Na + and K ÷ ions are required for organic acid transport in mammalian kidney cortex [24--29], it is possible that the observed effects on p-aminohippuric acid uptake may be indirect and medi- ated by changes in active electrolyte transport. Against this are the findings of the experiments using ouabain-poisoned slices: PCMB and a high concentration of tetrathionate were found to diminish, and a short exposure to a low concen- tration of tetrathionate was found to stimulate organic acid uptake (Table III). Nevertheless the magnitude of the responses to the sulfhydryl reagents was diminished in comparison to the results obtained using metabolically active iso- lated tubules. Hence part of the effects of these agents could be mediated by an alteration in {Na ÷, K+)-ATPase but it is also known that in preparations using slices rather than tubules [11,30,31] the rate of p-aminohippuric acid uptake and its sensitivity to various agents is decreased.

3 7 5
Acknowledgments
The authors wish to thank Misses C. Priol and M.N. Manguy for skilful tech- nical assistance and Dr. Nitza Pauleau for critical reading of the manuscript. We are indebted to Miss M. Bizien for preparing the manuscript. The studies have been supported by Institut National de Sant4 et de la Recherche M4dicale (I.N.S.E.R.M.) and by Commissariat ~ l'Energie Atomique (C.E.A.), France.
References
1 Morgan, H.E. and Whitfield, C.F. (1973) in Cur ren t Topics in Membranes and T ranspo r t (Bronner , F. and KleinzeUer, A., eds.), Vol. IV, pp. 2 5 5 - - 3 0 3 , Academic Press Inc., New York
2 Roths te in , A. (1973) in Mercury, Mercurials and Mercaptans (Miller, W.W. and Clarkson, J.W., eds.), pp. 68- -05 , Charles C. Thomas , Springfield, Ill.
3 Bloch, R. (1974) J. Biol. Chem. 249, 1 8 1 4 - - 1 8 2 2 4 Czech, M.P., Lawrence, J.C. and Lynn , W.S. (1974) Proc. Natl. Acad. Sci. U.S. 71, 4 1 7 3 - - 4 1 7 7 5 Feng, H.W. and Riggs, T.R. (1973) Biochim. Biophys. Acta 318, 4 2 6 - - 4 3 8 6 Kwock , L., WaUach, D.F.H. and Hefter , K. (1976) Biochim. Biophys. Acta 419 , 93 - -103 7 Berliner, R.W., Kennedy , T.J. and Hil ton, J.G. (1948) Am. J. Physiol. 154, 537 - -541 8 Farah , A. and Rennick , B.R. (1956) J. Pharmacol . Exptl . Therap. 117, 4 7 8 - - 4 8 7 9 Hook , J.B. and Hirsch, G.H. (1973) in Mercury , Mercurials and Marcap tans (Millet, W.W., and Clark-
son, J.W., eds.), pp. 124 - -138 , Charles C. Thomas , Springfield, I11. 10 Gi lman, A., Philips, F.S., Koelle, E.S., Allen, R.P. and St. John , E. (1946) Am. J. Physiol. 147,
1 1 5 - - 1 2 6 11 Burg, M.B. and Orloff , J. (1962) Am. J. Physiol. 203, 3 2 7 - - 3 3 0 12 Podevin, R.A. and Boumendi l -Podevin , E.F. (1972) Biochim. Biophys. Acta 282, 2 3 4 - - 2 4 9 13 Podevin, R.A. and Boumendi l -Podevin , E.F. (1977) Am. J. Physiol. , in the press 14 Boumendi l -Podevin , E.F. , Podevin, R.A. and Richer , G. (1975) J. Clin. Invest. 55, 1 1 4 2 - - 1 1 5 2 15 Podevin, R.A. and Boumendi l -Podevin , E.F. (1975) Biochim. Biophys. Ac ta 375, 1 0 5 - - 1 1 4 16 Smith , H.W., Finkelstein, N., Aliminosa, L., Crawford , B. and Graber , M. (1945) J. Clin. Invest. 24,
3 8 8 - - 4 0 4 17 Grassett i , D.R. and Murray , J .F. (1970) Biochem. Pharmacol . 19, 1 8 3 6 - - 1 8 3 8 18 Pihl, A. and Lange, R. (1962) J. Biol. Chem. 237 , 1 3 5 6 - - 1 3 6 2 19 Parker , D.J. and Allison, W.S. (1969) J. Biol. Chem. 244, 1 8 0 - - 1 8 9 20 Grassett i , D.R., Murray , J .F. and Ruan , H.T. (1969) Biochem. Pharmacol . 18, 6 0 3 - - 6 1 1 21 Haest , C.W.M. and Deut icke, B. (1975) Biochim. Biophys. Ac ta 401 , 4 6 8 - - 4 8 0 22 Chung, S.I. and Folk , J.E. (1970) J. Biol. Chem. 245, 6 8 1 - - 6 8 9 23 Webb, J .L. (1966) in Enzyme and Metabolic Inhibi tors (Webb, J .L. , ed.), Vol. II, pp. 6 5 8 - - 6 5 9 ,
Academic Press, Inc., New York 24 Platts, M.M. and Mudge, G.M. (1961) Am. J. Physiol. 200, 3 8 7 - - 3 9 1 25 Taggart , J .V., Silverman, L. and Trayner , E.M. (1953) Am. J. Physiol. 173, 3 4 5 - - 3 5 0 26 Foulkes , E.C. and Miller, B.F. (1961) in Membrane Transpor t and Metabolism (Kleinzeller, A. and
K o t y k , A., eds.), pp. 559- -565 , Academic Press, New York 27 Burg, M,B. and Orloff, J. (1962) Am. J. Physiol. 202, 565- -571 28 Bemdt , W.O. and Beechwood , E.C. (1965) Am. J. Physiol. 208, 6 4 2 - - 6 4 8 29 Chung, S.R., Park, Y.S. and Hong, S.K. (1970) Am. J. Physiol. 219, 30 - -33 30 Huang , K.C. and Lin, D.S.T. (1965) Am. J. Physiol. 2 0 8 , 3 9 1 - - 3 9 6 31 Mur thy , L. and Foulkes, E.C. (1967) Nature 213, 180 - -181





























